STUDIBREA $I BVALUAREA VIRUSURILOR GRIPALE iN ...
165
MINISTERUL SANATATII AL REPUBLICI MOLDOVA CENTRUL NATIONAL DE SANATATE PUBLICA Cu titlu de manuscris C.Z.U:578.821 .83(043.2) EDER VERONICA STUDIBREA $I BVALUAREA VIRUSURILOR GRIPALE iN PERIOADELB PANDEMICA $I INTERBPIDEMICA 163.04 - MICROBIOLOGIE Tezade doctorin gtiinfebiologice Conducitor qtiinfific : Autor: ftk l_-z Spinu Constantin - a l't v dr. hab. in medicini, profesor universitar, Om Emerito Laureat al Premiului Na{ional, Centrul Nafional de Slnitate Publici Eder Veronica CHI$INAU,2or5
Transcript of STUDIBREA $I BVALUAREA VIRUSURILOR GRIPALE iN ...
-
MINISTERUL SANATATII AL REPUBLICI MOLDOVACENTRUL NATIONAL DE SANATATE PUBLICA
Cu titlu de manuscrisC.Z.U: 578.821 .83 (043.2)
EDER VERONICA
STUDIBREA $I BVALUAREA VIRUSURILOR GRIPALE iNPERIOADELB PANDEMICA $I INTERBPIDEMICA
163.04 - MICROBIOLOGIE
Teza de doctor in gtiinfe biologice
Conducitor qtiinfific :
Autor:
ftk l_-z Spinu Constantin- a
l ' tv dr. hab. in medicini, profesor universitar,
Om Emerito Laureat al Premiului Na{ional,Centrul Nafional de Slnitate Publici
Eder Veronica
CHI$INAU,2or5
-
2
Eder Veronica, 2015
-
3
CUPRINS
ADNOTARE (n romn, rus i englez).......................................................................... 5
LISTA ABREVIERILOR.................................................................................................... 8
INTRODUCERE................................................................................................................... 9
1. EVOLUIA CAPACITILOR DE DIAGNOSTIC PRIVIND STUDIEREA I
EVALUAREA PARTICULARITILOR ETIOLOGICE ALE INFECIILOR
GRIPALE .
18
1.1 Aspecte etiologice ale infeciilor gripale i caracteristicile morfostructurale ale
virusurilor gripale ...................................................................................................
18
1.2 Caracteristica fenotipic i genotipic a virusurilor gripale........................................ 25
1.3 Diagnosticul de laborator: tehnici clasice i de biologie molecular de ultim
generaie......................................................................................
1.4 Concluzii la capitolul 1...............................................................................................
35
42
2. MATERIALE I METODE DE CERCETARE............................................................ 44
2.1 Liniaritatea cercetrilor i obiectul de studiu ..................................................... 44
2.2 Metodele de cercetare aplicate i volumul investigaiilor.......................................
2.3 Concluzii la capitolul 2...............................................................................................
44
54
3. STUDIEREA I EVALUAREA TULPINILOR DE VIRUSURI GRIPALE
IZOLATE I IDENTIFICATE N PERIOADA PANDEMIC ...............................
55
3.1 Particularitile virusurilor gripale n perioada pre-pandemic...................................
3.2 Rezultatele determinrii caracteristicilor tulpinilor de virusuri gripale n
perioada pandemic...................................................................................................
3.3 Studierea particularitilor tulpinilor de virusuri gripale n perioada
post-pandemic..........................................................................................................
3.4 Concluzii la capitolul 3...............................................................................................
55
62
70
80
4. EVALUAREA PARTICULARITILOR VIRUSURILOR GRIPALE N
PERIOADA INTEREPIDEMIC NTRU OPTIMIZAREA SUPRAVEGHERII
EPIDEMIOLOGICE I VIRUSOLOGICE A GRIPEI ...................................................
83
4.1 Rezultatele evalurii particularitilor virusurilor gripale n sezonul 2011-
2012.............................................................................................................................
83
4.2 Rezultatele evalurii particularitilor virusurilor gripale n sezonul 2012-
2013............................................................................................................................
4.3 Studierea i evaluarea particularitilor virusurilor gripale n sezonul 2013-2014....
88
101
-
4
4.4 Valorificarea rezultatelor obinute ntru optimizarea supravegherii epidemiologice
i virusologice a gripei................................................................................................
4.5 Concluzii la capitolul 4...............................................................................................
109
114
CONCLUZII GENERALE I RECOMANDRI........................................................... 116
BIBLIOGRAFIE................................................................................................................... 120
ANEXE...................................................................................................................................
ANEXA 1 Acte de implementare ..
ANEXA 2 Acreditarea laboratorului CNSP de ctre OMS ..
ANEXA 3 Chestionarul privind acreditarea de ctre OMS ..
ANEXA 4 Certificate de realizare a Programelor de Control Extern de Calitate .
ANEXA 5 Brevet de invenie ....
ANEXA 6 Diploma Cupa Marele Premiu ..
ANEXA 7 Diploma Medalia de aur ...
ANEXA 8 Diploma de decernare ..
ANEXA 9 Diploma de Excelen i Medalia de aur ......
ANEXA 10 Diploma Gala Premiilor n Sntate ..
140
141
145
146
147
154
156
157
158
159
160
DECLARAIA PRIVIND ASUMAREA RSPUNDERII .............................................. 161
CV-ul AUTORULUI ............................................................................................................ 162
-
5
ADNOTARE
Eder Veronica: Studierea i evaluarea virusurilor gripale n perioadele pandemic i
interepidemic, teza de doctor n tiine biologice, Chiinu, 2015. Structura tezei: introducere,
4 capitole, concluzii i recomandri, bibliografie (191 surse), 118 pagini de text de baz, 22
tabele, 47 figuri i 10 anexe. Rezultatele sunt publicate n 28 lucrri tiinifice. Cuvinte cheie:
virusuri gripale, HA, NA, grup genetic, sensibilitate la antivirale. Domeniu de studiu: 163.04 -
Microbiologie. Scopul lucrrii: Studierea i evaluarea particularitilor antigenice, fenotipice i
genotipice ale tulpinilor de virusuri gripale izolate i identificate n perioadele pandemic i
interepidemic n Republica Moldova ntru argumentarea msurilor de sntate cu optimizarea
sistemului naional de supraveghere epidemiologic i virusologic a gripei. Obiectivele
studiului: Determinarea i evaluarea tipurilor i subtipurilor de virusuri gripale evideniate n
perioada de studiu; studierea i evaluarea particularitilor antigenice ale tulpinilor de virusuri
gripale n perioadele pandemic i interepidemic; studierea i evaluarea particularitilor
genetice ale tulpinilor de virusuri gripale A i B n baza genelor HA i NA n perioadele
precutate n studiu; analiza caracteristicilor fenotipice cu evaluarea sensibilitii tulpinilor de
virusuri gripale la remediile antivirale de ultim generaie (oseltamivir, zanamivir) n perioadele
evideniate; valorificarea rezultatelor obinute i optimizarea supravegherii epidemiologice i
virusologice a gripei. Noutatea i originalitatea tiinific: n premier au fost obinute
rezultate originale privind particularitile antigenice, genotipice i fenotipice ale virusurilor
gripale circulante n perioadele pandemic i interepidemic. Problema tiinific: Evidenierea
i evaluarea particularitilor antigenice, genotipice i fenotipice ale virusurilor gripale circulante
n Republica Moldova, care au servit ca argumente i suport metodic ntru perfecionarea
continu a sistemului de supraveghere epidemiologic, clinic i virusologic la grip, IACRS i
SARI implementat, racordat la exigenele OMS, ECDC i CDC. Semnificaia teoretic. Acest
studiu a evideniat particularitile virusurilor gripale circulante n Republica Moldova, ceea ce a
permis de a identifica i evalua locul tulpinilor izolate n arborele filogenetic global, circumstane
extrem de importante pentru argumentarea formulei vaccinului gripal. Valoarea aplicativ a
lucrrii. Rezultatele au permis de a evidenia particularitile virusologice evolutive ale
procesului epidemic prin grip n perioadele nominalizate elemente semnificative utilizate n
perfecionarea sistemului naional de supraveghere la grip, IACRS i SARI ajustat la exigenele
OMS, ECDC, CDC. Implementarea rezultatelor tiinifice: Rezultatele obinute au servit la
valorificarea de noi metode standardizate de diagnostic al gripei, optimizarea monitorizrii
infeciilor gripale, IACRS i SARI pentru realizarea n timp real a msurilor adecvate de control
i rspuns n contextul integrrii n reelele de supraveghere regionale (EuroFlu/TESSy) i
globale (FluNet) ale OMS.
-
6
:
.
, , 2015. : , 4 ,
, (191 ), 118 , 22 ,
47 10 . 28 .
: , , , ,
. : 163.04 - .
: ,
. :
;
;
;
;
. :
,
. :
,
, ,
, ECDC CDC. :
,
. :
,
,
(EuroFlu/TESSy) (FluNet)
.
-
7
SUMMARY
Eder Veronica: The study and evaluation of influenza viruses in the pandemic and
interepidemic periods, PhD thesis in biological sciences, Chisinau, Republic of Moldova,
2015. Thesis structure: introduction, 4 chapters, conclusions and recommendations, bibliography
(191 sources), 118 pages of basic text, 22 tables, 47 figures, and 10 annexes. The results have
been published in 28 scientific papers. Keywords: influenza viruses, HA, NA, genetic group,
antiviral susceptibility. Field of research: 163.04 - Microbiology. The purpose of study: Study
and evaluation of antigenic, phenotypic and genotypic characteristics of influenza strains isolated
and identified in pandemic and inter-epidemic periods in Moldova. Research objectives: The
determination and evaluation of the types/subtypes of influenza viruses in the period of the
study; study and evaluation of antigenic specificities of influenza viruses strains in pandemic and
interepidemic periods; study and evaluation of genetic peculiarities of A and B influenza
viruses strains based on HA and NA genes in the mentioned periods; sensitivity analysis
evaluating phenotypic characteristics of influenza viruses strains to antiviral remedies of latest
generation (oseltamivir, zanamivir) in the highlighted periods; results achieved and optimize
epidemiological and virological influenza surveillance system. The scientific novelty and
originality: For the first time were obtained the original results on antigenic, genotypic and
phenotypic characteristics of circulating influenza viruses during pandemic and inter-epidemic
periods. Scientific problem: Highlighting and evaluation of features of influenza viruses
circulating in Republic of Moldova served as arguments and methodical support to continuous
improvement of implemented influenza, ARI and SARI epidemiological, clinical and virological
surveillance system, connected to the requirements of WHO, ECDC and CDC. The scientific
and practical value: The results allowed to highlight the virological peculiarities of influenza
epidemic evolution in mentioned periods significant items used in improving the national
influenza, ARI and SARI surveillance adjusted to the WHO, ECDC, CDC requirements.
Obtained results allowed to identify and evaluate the place of isolated strains in the phylogenetic
tree globally extremely important circumstances to argue influenza vaccine formulation. The
implementation of scientific results: The results of this research are used for clinical,
epidemiological and virological influenza, ARI and SARI surveillance system improving
through the implementation of new standardized methods of influenza diagnostics by approving
a series of documents in order to strengthen epidemiological surveillance, systematic monitoring
of influenza, ARI and SARI for achieving in real time the appropriate preventive and control
measures in the context of integration in regional (EuroFlu/TESSy) and global (FluNet)
surveillance networks of the WHO.
-
8
LISTA ABREVIERILOR
ADN acid dezoxiribonucleic
Ag antigen
ARI Acute Respiratory Infections
ARN acid ribonucleic
CDC Centers for Disease Control and
Prevention
CNSP Centrul Naional de Sntate Public
CSP Centru de Sntate Public
ECDC European Centre for Disease
Prevention and Control
FITC izotiocianat de fluorescein
GIHSN Global Influenza Hospital
Surveillance Network
GISN Global Influenza Surveillance
Network
GISRS Global Influenza Surveillance and
Response System
HA hemaglutinin
HAU uniti de hemaglutinare
IACRS infecii acute ale cilor respiratorii
superioare
IMSP Instituie Medico - Sanitar Public
IRVA infecii respiratorii virale acute
M, M1, M2 proteine matrice
NA neuraminidaz
NAI inhibitor al neuraminidazei
NEP nuclear export protein
NP nucleoprotein
NS1, NS2 proteine nestructurale
OMS Organizaia Mondial a Sntii
ORF open reading frame
PA proteaza acid
PB1, PB2 proteaze bazice
PBS soluie-tampon fosfat salin
PCR reacie de polimerizare n lan
(Polymerase Chain Reaction)
RHA reacie de hemaglutinare
RHAI reacie de inhibare a hemaglutinrii
rRT-PCR Real Time Revers Transcription
Polimerase Chain Reaction
SARI severe acute respiratory infections
SUA Statele Unite ale Americii
WHO World Health Organization
-
9
INTRODUCERE
Gripa i IACRS sunt maladii infecioase cu impact global, ce evolueaz n epidemii sau/i
pandemii soldate cu mortalitate crescut.
Virusurile gripale reprezint o problem permanent de sntate public pentru toate
rile, iar eforturile de a preveni mbolnvirile i rspndirea acestora sunt constante, dar i
considerabile.
Larg rspndite n ntreaga lume, virusurile gripale infecteaz nu numai omul, ci i
numeroase specii de animale domestice (cai, porci) i slbatice (nurc, foc, balen etc.) sau
psri (rae, pescrui, curcani, gini etc.) [1-4].
Variaia antigenic i circulaia virusurilor gripale ntre specii sunt cauza izbucnirilor
epidemice care au loc anual, mbolnvirile fiind favorizate de rspunsul imun neadecvat, chiar n
acele segmente de populaie care au fost anterior expuse infeciei gripale [1-4].
Epidemiile de grip se nregistreaz n fiecare an datorit variaiei antigenice minore
(antigenic drift). Acestea sunt mutaii nesemnificative n genele care codific HA i NA. Ele
constau n apariia lent de mutaii punctiforme n HA i NA n rezultatul circulaiei continue a
virusurilor gripale n populaiile parial imunizate. Structurile antigenice existente sufer
modificri de conformaie spaial i/sau modificri n compoziia unor aminoacizi. Protecia
imun fa de contactele anterioare cu virusul gripal se menine, dar este insuficient. Astfel, fa
de subtipurile existente ale virusurilor gripale, la om se reinstaleaz o susceptibilitate la
reinfectri multiple. De regul, astfel de succesiuni se produc n fiecare an [4-7].
De obicei, epidemiile de grip apar n perioada rece a anului: n Emisfera de Nord n
lunile noiembrie martie, n Emisfera de Sud lunile mai august. n regiunile tropice gripa nu
se manifest sezonier, ci se nregistreaz pe tot parcursul anului, fiind dependent, totui, de
schimbrile climei [3-5].
n Republica Moldova cea mai nalt morbiditate prin grip se nregistreaz, de obicei, n
luna februarie, ns au fost cazuri cnd cel mai nalt nivel de morbiditate prin grip s-a nregistrat
n lunile decembrie, ianuarie i martie [2, 3, 6, 8].
Durata epidemiei este de regul 1-2 luni (4-8 sptmni), dup care virusurile dispar. Una
din enigmele principale ale virusului gripal este faptul c n cea mai mare parte a anului ele
lipsesc n circulaia populaiei umane i nu se cunoate unde circul n perioada interepidemic,
cum i unde se produc modificrile antigenice (drift antigen). Se presupune c virusurile ar
circula i s-ar modifica n regiunile ecuatoriale, pe motiv c mbolnvirile prin grip aici se
nregistreaz pe toat perioada anului [1, 3-5].
-
10
Totodat, pe lng variaia antigenic minor, exist i variaie antigenic major
(antigenic shift), ce se ntlnete doar la virusul gripal de tip A. Mutaiile de tip shift reprezint o
form mult mai brutal de schimbri n virusul gripal A, ceea ce duce la apariia unui nou virus
gripal cu o nou combinaie a proteinelor HA i NA, care anterior nu a existat mult timp sau
niciodat n populaia uman. Dac acest virus apare n populaia uman, unde majoritatea este
lipsit de protecie sau exist o protecie nesemnificativ fa de aceast variant, virusul
obinnd abilitatea de a se transmite de la om la om, atunci aceste circumstane pot conduce la
apariia de pandemii de proporii. Important este faptul, c modificrile n structura antigenic nu
pot fi previzibile. Virusurile cu deosebiri genotipice i fenotipice semnificative fa de cele
antecedente apar la intervale neregulate de timp (10-40 de ani). Aceste modificri sunt
responsabile de apariia marilor pandemii de grip [1, 3-5].
Pe parcursul a ultimii 300 de ani s-au nregistrat 10 pandemii de grip, din care 4 au avut
loc n secolul XX [3-5, 9, 10]. Pandemia spaniol din 1918 a fost una din cea mai sever
dintre pandemiile cunoscute i descrise vreodat. Gripa spaniol a afectat peste 20,0% din
populaia globului avnd o evoluie foarte rapid. Timp de 10 luni gripa s-a rspndit n ntreaga
lume. n aceast perioad au fost afectate peste 400 milioane de persoane. Valuri repetate de
mbolnviri prin grip s-au nregistrat i n anii 1918-1919 i 1919-1920 cu o intensitate mai
mic. Atunci s-au mbolnvit cei care nu au suferit n primul val. Pandemia spaniol a fost
cauzat de un nou virus gripal A(H1N1), genomul cruia coninea poriuni de gene ale
virusurilor gripale aviare i porcine. Cei mai afectai au fost tinerii, btrnii i persoanele
imunocompromise. ntr-un singur an gripa spaniol a fcut mai multe victime dect ciuma
bubonic n patru ani (1347-1351) i dect Primul Rzboi Mondial, numrnd de la 50 la 100
milioane de decese. Cea mai nalt mortalitate s-a nregistrat la persoanele cu vrsta cuprins
ntre 20 i 40 ani, rata mortalitii fiind de 259 ori mai mare dect n cazul unei gripe obinuite.
Referitor la locul i debutul pandemiei sunt dou ipoteze: prima pandemia a debutat n
februarie-martie 1918 n SUA cu transmiterea ulterioar de ctre trupele americane n Europa, a
doua ipotez pandemia a debutat n ianuarie 1918 n China cu rspndirea ulterioar la Apus
[3-5, 9, 10]. Gripa asiatic a avut loc n 1957-1958 i a fost cauzat de virusul gripal
A(H2N2), pandemia fiind de intensitate mai mic dect cea din 1918. Pandemia a nceput n
februarie 1957 n Orientul ndeprtat i s-a rspndit rapid n ntreaga lume nregistrnd 2
miliarde de bolnavi i 1 milion de decese [3-5, 9, 10]. Gripa Hong Kong n 1968-1969 a
aprut gripa de tip Hong Kong cauzat de virusul gripal A(H3N2). Pandemia avnd severitate
medie, a afectat, n special, persoanele cu vrsta de 65 de ani, nregistrndu-se peste 1 milion de
decese [3-5, 9, 10]. Gripa Ruseasc n 1977-1978 a avut loc o pandemie relativ uoar
-
11
cauzat de virusul gripal A(H1N1) agent etiologic implicat n epidemia din 1950, astfel au avut
de suferit n primul rnd persoanele nscute dup acest an [3, 4, 10].
Apariia virusului gripal A(H5N1) n 1997 n Hong Kong , care a cauzat o epizootie
masiv printre psrile domestice afectnd concomitent 18 persoane cu moartea a 6 dintre ele, a
servit ca un semnal de apariie a unei noi pandemii de grip. Distrugerea la acel moment a cca
1,5 milioane de psri a stopat rspndirea virusului A(H5N1) printre psrile domestice i
populaia uman. Virusul gripal aviar A(H5N1) a reaprut printre psrile domestice n anul
2003 ntr-un ir de ri asiatice: Cambodia, China, Indonezia, Japonia, Laos, Thailanda, Vietnam,
Coreea. n 2004 din nou s-au nregistrat mbolnviri printre oameni. Distrugerea a peste 160
milioane de psri a contribuit la reducerea rspndirii infeciei cu virusul gripal A(H5N1). n
pofida faptului c acest virus a trecut bariera de specie cu afectarea nu numai a psrilor, dar i a
unor specii de mamifere (tigri, cini, pisici), totui transmiterea lui de la om la om a fost
limitat. Persoanele care s-au mbolnvit de grip cauzat de virusul A(H5N1) au avut contact
direct i nemijlocit cu psrile bolnave [3-5, 9, 10].
Cu apariia virusului gripal aviar A(H5N1) n 2003 a persistat pericolul declanrii unei
noi pandemii de grip. Dac la nceputul anului 2004 se credea, c noua pandemie va fi cauzat
de virusul gripal A(H5N1), atunci evenimentele din 2009 au demonstrat, c ngrijorrile
specialitilor n domeniu nu au fost n zdar [3-5, 9, 10].
Noua pandemie de grip, prima din secolul XXI, a debutat n aprilie 2009 n SUA
(California de Sud i Texas), fiind cauzat de virusul gripal de tip nou A(H1N1) numit "triplu
reasortant", deoarece genomul su conine gene ale virusurilor gripale porcine, umane i aviare.
Virusul de tip nou a aprut brusc, fiind detectat concomitent n alte 2 ri: Mexic i Canada.
Procesul epidemic prin infecia cu virusul gripal de tip nou A(H1N1) a evoluat foarte repede,
afectnd ntr-un timp scurt un numr mare de oameni de pe toate continentele. Aceste
evenimente au impus OMS s ridice la 11 iunie 2009 nivelul de alert pandemic de la faza 5 la
faza 6, ceea ce a nsemnat nceputul primei pandemii de grip din secolul XXI. Totodat OMS a
subliniat, c severitatea pandemiei date a fost moderat. Majoritatea persoanelor infectate s-au
recuperat i s-au nsntoit fr a necesita spitalizare sau tratament medical specific, iar numrul
cazurilor cu complicaii i decese a fost similar epidemiilor gripale sezoniere [3-6, 8-12].
n Republica Moldova, gripa i IACRS se nregistreaz n fiecare an, numrul cazurilor
de mbolnviri variind de la an la an i reprezint 2/3 din numrul total de maladii infecioase
nregistrate pe parcursul anului. Apariia n anul 2009 a unui nou subtip de virus gripal A(H1N1)
a cauzat pandemia de grip la nivel global, resimit i n Republica Moldova. Acest fapt a impus
fortificarea sistemului de supraveghere epidemiologic a gripei prin crearea strategiilor de
-
12
prevenire i control permanent, inclusiv prin sisteme speciale de monitorizare a gripei, IACRS i
SARI [3, 6, 8].
Monitorizarea circulaiei virusurilor gripale umane face parte din sistemul complex de
supraveghere a gripei. Anual, OMS monitorizeaz circulaia i profilul antigenic al virusurilor
gripale n vederea descifrrii n timp real a variantelor cu potenial epidemic i pandemic,
precum i a sensibilitii lor la produsele medicamentoase cu aciune antiviral, n special, a celor
de ultim generaie (oseltamivir, zanamivir) i profilul imunitii antigripale a populaiei cu
scopul preparrii vaccinului antigripal potrivit sezonului curent.
Lund n consideraie potenialul epidemic (pandemic) ale anumitor tipuri/subtipuri de
virusuri gripale, inclusiv impactul socio-economic al acestora asupra sntii publice se impune
necesitatea studierii i evalurii virusologice a tulpinilor de virusuri gripale izolate i identificate,
inclusiv i n Republica Moldova.
Scopul lucrrii: Studierea i evaluarea caracteristicilor antigenice, fenotipice i
genotipice ale tulpinilor de virusuri gripale izolate i identificate n perioadele pandemic i
interepidemic n Republica Moldova ntru argumentarea msurilor de sntate cu optimizarea
sistemului naional de supraveghere epidemiologic i virusologic a gripei.
Obiectivele lucrrii:
1. Determinarea i evaluarea tipurilor i subtipurilor de virusuri gripale evideniate n
perioadele de studiu.
2. Studierea i evaluarea particularitilor antigenice ale tulpinilor de virusuri gripale n
perioadele pandemic i interepidemic.
3. Studierea i evaluarea particularitilor genetice ale tulpinilor de virusuri gripale A i B n
baza genelor HA i NA n perioadele precutate n studiu.
4. Analiza caracteristicilor fenotipice cu evaluarea sensibilitii tulpinilor de virusuri gripale la
remediile antivirale de ultim generaie (oseltamivir, zanamivir) n perioadele evideniate.
5. Valorificarea rezultatelor obinute ntru optimizarea supravegherii epidemiologice i
virusologice a gripei.
Noutatea i originalitatea tiinific: n premier pentru Republica Moldova, au fost
obinute rezultate originale privind particularitile antigenice, genotipice i fenotipice ale
virusurilor gripale circulante n perioadele pandemic i interepidemic. Utilizarea, de rnd cu
metodele clasice, a tehnicilor de performan de biologie molecular (rRT-PCR, genotipare,
secveniere) n studierea tulpinilor de virusuri gripale a permis aprecierea i evaluarea poziiei
acestor virusuri n arborii filogenetici globali pentru fiecare gen n parte circumstane extrem
de importante pentru formularea coctailului vaccinal. Analiza i evaluarea susceptibilitii
-
13
tulpinilor de virusuri gripale circulante a demonstrat sensibilitatea lor la remediile antivirale de
ultim generaie, abordare extrem de important pentru perfecionarea msurilor de control i
rspuns.
Problema tiinific soluionat important const n evidenierea i evaluarea
particularitilor antigenice, genotipice i fenotipice ale virusurilor gripale circulante n
Republica Moldova, care au servit ca argumente i suport metodic ntru perfecionarea continu a
sistemului de supraveghere epidemiologic, clinic i virusologic la grip, IACRS i SARI
implementat, racordat la exigenele OMS, ECDC i CDC.
Semnificaia teoretic: Acest studiu a evideniat particularitile antigenice, genotipice
i fenotipice ale virusurilor gripale circulante n Republica Moldova, ceea ce a permis de a
identifica i evalua locul tulpinilor identificate n arborele filogenetic global, circumstane extrem
de importante pentru argumentarea formulei vaccinului gripal.
Valoarea aplicativ a lucrrii: Rezultatele obinute pe parcursul realizrii acestui studiu
au permis de a evidenia particularitile virusologice de evoluie a procesului epidemic prin
grip n perioadele pandemic i interepidemic elemente semnificative utilizate ulterior n
perfecionarea sistemului naional de supraveghere la grip, IACRS i SARI ajustat la exigenele
OMS, CDC i ECDC. Graie perfecionrii, n corespundere cu strategiile organismelor
internaionale nominalizate, sistemul de supraveghere autohton a fost integrat n reelele de
supraveghere european EuroFlu/TESSy i global FluNet.
Sistemul de supraveghere nominalizat permite de a evalua n timp real particularitile
tulpinilor de virusuri gripale circulante, potenialul lor epidemic/pandemic, sensibilitatea la
antivirale, pronosticul situaiei epidemiologice i volumul de investigaii cu msuri specifice i
nespecifice de profilaxie n situaii de urgen.
Implementarea rezultatelor tiinifice: Rezultatele obinute la realizarea cercetrilor
tiinifice n cadrul acestui studiu au servit la perfecionarea sistemului de supraveghere clinico-
epidemiologic i virusologic la grip, IACRS i SARI prin implementarea de noi metode
standardizate de diagnostic al gripei, recunoaterea de ctre organismele internaionale
nominalizate a subdiviziunii de profil a CNSP ca Centru Naional de Grip i prin aprobarea unui
ir de acte normative, n special, a Planului-cadru intersectorial gradual pentru combaterea
efectelor pandemiei cu virusul gripal nou A(H1N1) n Republica Moldova, aprobat prin
Hotrrea Guvernului nr. 824 din 15.12.2009 i a Ordinului Ministerului Sntii nr. 824 din
31.10.2011 Cu privire la perfectarea sistemului de supraveghere la grip i infeciile acute ale
cilor respiratorii n Republica Moldova n scopul fortificrii supravegherii epidemiologice,
monitorizrii sistematice a circulaiei infeciilor gripale, IACRS i SARI pentru realizarea n
-
14
timp real a msurilor adecvate de prevenire i control i n contextul integrrii n reelele de
supraveghere regionale (EuroFlu/TESSy) i globale (FluNet) ale Organizaiei Mondiale a
Sntii.
Aprobarea rezultatelor tiinifice: Direciile studiate i rezultatele obinute la realizarea
acestui studiu au fost prezentate i discutate n cadrul forurilor tiinifice naionale i
internaionale: Conferina tiinifico-Practic cu Participare Internaional CMP Chiinu
trecut, prezent i viitor, Chiinu, 23 octombrie 2009; A doua Conferin Naional de
Microbiologie i Epidemiologie, Sinaia, Romnia, 14-16 octombrie 2010; Congresul Naional de
Microbiologie i Conferina Naional de Epidemiologie, Iai, Romnia, 10-12 noiembrie 2011;
International Conference on Emerging Infectious Diseases, Atlanta, Georgia, USA, March 11-
14, 2012; Conferina a VII-a a medicilor-infecioniti din Republica Moldova, Chiinu, 2012;
3rd International Influenza Meeting, Muenster, Germany, September 2nd
4th
, 2012; -
i II
I I, K, , 10 2012; European Scientific
Conference on Applied Infectious Disease Epidemiology (ESCAIDE), Edinburgh, UK, 24-26
October 2012; The second isirv-Antiviral Group Conference in conjunction with NIHE. Severe
Influenza: Burden, Pathogenesis and Management, Hanoi, Viet Nam, 29th 31st October 2012;
23rd
Annual Meeting of the Society for Virology, Kiel, Germany, 6-9 March 2013; Options for
the Control of Influenza, Cape Town, South Africa, 5-10 September 2013; The XIX-th Session
of the Balkan Medical Days and the Second Congress of Emergency Medicine of the Republic of
Moldova, Chisinau, Republic of Moldova, 22-24 September 2013; -
, 90-
, . .. ,
, 2013; Congresul specialitilor din domeniul Sntii Publice i Management Sanitar
din Republica Moldova, Chiinu 25-26 octombrie 2013; A XII-a Ediie a Salonului
Internaional al Cercetrii, Inovrii i Inventicii PRO INVENT 2014, Cluj-Napoca, Romnia,
19-21 martie 2014; Third ISIRV-AVG Conference Influenza and other Respiratory Virus
Infections: Advances in Clinical Management 4th-6th June 2014, Tokyo, Japan; The XVIII-th
International Exhibition of Research, Innovation and Technological Transfer INVENTICA
2014, Iasi, Romania, 2-4 July 2014; 4th International Influenza Meeting, Muenster, Germany,
September 21-23, 2014; Conferina Centrul de Sntate Public din municipiul Chiinu 70 de
ani la Straja Sntii, Chiinu, Republica Moldova, 22 octombrie 2014; A VII-a Conferin
Naional de Microbiologie i Epidemiologie, Bucureti, Romnia, 12-14 Noiembrie 2014;
-
15
Salonul Internaional de Inventic PRO INVENT, Ediia a XIII-a, Cluj-Napoca, Romnia, 25-27
martie 2015.
Rezultatele cercetrilor au fost discutate i aprobate la edina Centrului Controlul Bolilor
virale al Centrului Naional de Sntate Public (Proces verbal nr. 3 din 16.12.2014), la edina
Consiliului tiinific al CNSP (Proces verbal nr. 1 din 20.01.2015), i la edina Seminarului
tiinific de profil, specialitatea 163.04 Microbiologie, din cadrul Institutului de Microbiologie
i Biotehnologie al AM (Proces verbal nr. 2 din 02.04.2015),
Publicaii la tema tezei: Rezultatele cercetrilor sunt reflectate n 28 lucrri tiinifice (2
din ele fr coautori), inclusiv: 6 articole n reviste tiinifice naionale, 2 articole n reviste
tiinifice internaionale (cotate ISI IF = 4.659 i SCOPUS), 1 tez a comunicrii la nivel
naional, 17 teze ale comunicrilor internaionale, 1 brevet de invenie MD 782 Z publicat n
Buletinul Oficial de Proprietate Industrial (BOPI) nr. 6, 30 iunie 2014.
Volumul i structura tezei: Teza este scris n limba romn, tehnoredactat la
calculator, compartimentat tradiional i include: foaia de titlu, foaia privind dreptul de autor,
rezumate n limbile romn, englez i rus, lista abrevierilor, cuprins, introducere, cuvinte-
cheie, 4 capitole, concluzii i recomandri practice, referine bibliografice, anexe, declaraia
privind asumarea rspunderii, CV-ul autorului. Teza este expus pe 118 pagini de text cules la
calculator n editorul Word, este ilustrat cu 22 tabele, 47 figuri, 10 anexe. Teza este
fundamentat pe 191 referine bibliografice.
Sumarul compartimentelor tezei
n Introducere sunt argumentate actualitatea, importana i necesitatea cercetrii
tiinifice efectuate, este reflectat situaia actual n domeniu, sunt formulate scopul i
obiectivele studiului. Descrierea i expunerea inovaiei tiinifice i a rezultatelor obinute
confirm aspectul tiinific al studiului efectuat i importana lui pentru medicin, n special
pentru sntatea public.
Capitolul 1. EVOLUIA CAPACITILOR DE DIAGNOSTIC PRIVIND
STUDIEREA I EVALUAREA PARTICULARITILOR ETIOLOGICE ALE
INFECIILOR GRIPALE cuprinde o sintez a bazelor teoretice, inclusiv rezultatele
cercetrilor tiinifice privind etiologia, caracteristicile antigenice, fenotipice i genotipice ale
virusurilor gripale efectuate de ctre cercettorii i specialitii din acest domeniu. Sunt expuse
principalele repere existente n domeniul de cercetare la nivel naional, european i mondial.
Sunt descrise studii tiinifice importante, efectuate n ultimul deceniu, privind evaluarea
particularitilor etiologice, inclusiv particularitile antigenice, fenotipice i genotipice ale
virusurilor gripale circulante la nivel de mapamond. Sunt argumentate scopul i obiectivele tezei
-
16
prin analiza profund a referinelor bibliografice recente ale savanilor contemporani cu renume
internaional. n concluzii sunt nominalizate premisele care au stat la baza iniierii acestui studiu.
Capitolul 2. Materiale i metode include reflectarea detaliat a metodologiei,
metodelor i materialelor de cercetare folosite n cadrul studiului tiinific efectuat. Acesta
include metode descriptive, virusologice, imunologice, de biologie molecular, analitice i
statistice.
n calitate de materiale de studiu au fost folosite 9799 de probe codificate cu material
biologic colectat de la pacienii cu diagnosticul prezumtiv Grip, IRVA i SARI, 127
tulpini de virusuri gripale izolate din probele pozitive la prezena ARN virusurilor gripale
circulante, 78 tulpini cercetate pentru evaluarea particularitilor genotipice ale virusurilor
gripale la nivelul genei HA i 71 tulpini evaluate genetic pentru gena NA.
Capitolul 3. Studierea i evaluarea tulpinilor de virusuri gripale izolate i
identificate n perioada pandemic elucideaz rezultatele unei cercetri moderne, cu aplicarea
n premier pentru Republica Moldova a tehnicilor de biologie molecular n diagnosticarea
gripei. Sunt prezentate rezultatele evidenierii particularitilor antigenice, genotipice i
fenotipice ale virusurilor gripale circulante n perioadele evideniate, caracterizate prin
similaritate antigenic cu tulpinile vaccinale, apartenena la grupurile genetice n baza arborilor
filogenetici globali, sensibilitatea la remediile antivirale de ultim generaie. Rezultatele obinute
au permis de a confirma procesul evolutiv al virusurilor gripale de la o perioad la alta (pre-
pandemic pandemic post-pandemic) prin variaii antigenice majore, manifestat prin
apariia n circulaie a unei tulpini noi de virus gripal A(H1N1)pdm i substituia din circulaie a
virusului gripal sezonier A(H1N1) cu virusul gripal A(H1N1)pdm.
Capitolul 4. Evaluarea particularitilor virusurilor gripale n perioada
interepidemic ntru optimizarea supravegherii epidemiologice i virusologice a gripei
include evidenierea particularitilor care influeneaz n timp evoluia virusurilor gripale, fapt
ce a permis de a extrapola concordana rezultatelor obinute n acest studiu la datele selectate din
literatura internaional, i anume prin confirmarea procesului evolutiv al virusurilor gripale de la
un sezon la altul prin variaii antigenice minore.
Rezultatele studiului au permis valorificarea sistemului de supraveghere clinico-
epidemiologic i virusologic la grip, IACRS i SARI de tip sentinel racordat la exigenele
OMS, ECDC i CDC prin implementarea i valorificarea algoritmului de investigaii ce in de
evaluarea particularitilor antigenice, genotipice i fenotipice ale virusurilor gripale, ceea ce
contribuie la studierea tulpinilor de virusuri gripale n cadrul Reelei Globale de Supraveghere i
-
17
Rspuns la Grip (GISRS) din cadrul Reelei Globale de Supraveghere la Grip al OMS (WHO
GISN).
Rezultatele lucrrii sunt utilizate n cadrul reelei WHO GISN, fapt ce contribuie la:
aprecierea formulei coctailului vaccinal antigripal pentru fiecare sezon; depistarea ct mai
precoce a noilor substituii de aminoacizi care ar putea influena patogenitatea, tropismul i
capacitatea de transmisie a virusurilor gripale de la o specie la alta; evidenierea factorilor
determinani ai reasortrii virusurilor, n special n apariia de noi virusuri cu potenial pandemic,
inclusiv rezistente la remediile antivirale aflate n uz, pentru evaluarea remediilor antivirale noi
orientate spre ajustarea unui tratament antiviral adecvat.
Rezultatele studiului efectuat n laboratorul Epidemiologia infeciilor respiratorii virale
din cadrul CNSP cu valorificarea lor n practic, au permis de a fortifica sistemele de
supraveghere clinico-epidemiologic i virusologic la grip, IACRS i SARI naional i
sentinel cu realizarea msurilor de control i rspuns. De asemenea, rezultatele expuse au fost
utilizate n procesul de acreditare a laboratorului nominalizat de ctre OMS, care a fost
recunoscut ca Centru Naional de Grip inclus n Reeaua Global de Supraveghere la Grip
(GISN) prin intermediul Sistemului Global de Supraveghere i Rspuns la Grip al OMS (WHO
GISRS).
-
18
1. EVOLUIA CAPACITILOR DE DIAGNOSTIC PRIVIND STUDIEREA I
EVALUAREA PARTICULARITILOR ETIOLOGICE
ALE INFECIILOR GRIPALE
1.1. Aspecte etiologice ale infeciilor gripale i caracteristicile morfostructurale ale
virusurilor gripale
Infeciile gripale, ce se declaneaz regulat i sezonier prin epidemii i periodic (la diferit
interval de timp: 10-40 ani) prin pandemii, au un impact negativ att asupra sntii publice,
sistemului de sntate, ct i asupra economiei naionale, i astfel, impun eforturi considerabile
de control i rspuns.
Gripa este provocat de virusurile gripale din familia Orthomyxoviridae, genul Influenza
virus, care include virusurile gripale de tip A, B i C cu genom ARN, monocatenar, segmentat,
de sens negativ.
Virusul gripal A pentru prima dat a fost izolat n 1930 de la porci, de ctre virusologul
american Richard Shope, ns primul caz de infectare a omului cu grip porcin a fost atestat cu
trei ani mai nainte [1-3]. De la oameni, virusul a fost izolat n 1933, de un grup de cercettori
englezi Wilson Smith, Cristofer Andrews i Patrick Laidlow. Virusul gripal A este unul din cele
mai cunoscute i mai nfricotoare dintre virusurile gripale, cauznd cele mai serioase epidemii
i pandemii n istoria omenirii cu o rat extrem de nalt de spitalizri i decese [6, 15-18].
Virusul gripal B, a fost descoperit n 1940 de ctre virusologul american Thomas Francis-
Junior. Dup gradul de virulen, contagiozitate i semnificaie epidemic, virusul gripal B,
mereu a cedat n faa virusului gripal A. Acest virus nu cauzeaz pandemii, ns este un agent
patogen al focarelor epidemice moderate, soldate uneori cu spitalizri i cazuri de deces [6, 15-
18].
Virusul gripal C a fost izolat pentru prima dat n 1947 de ctre virusologul american
Richard Taylor. Spre deosebire de virusurile gripale A i B, virusul gripal C cauzeaz infecii
respiratorii uoare, similare rcelilor banale, ns la copiii mici poate evolua destul de grav.
Pentru virusul gripal C este caracteristic o stabilitate major a proprietilor antigenice i
biologice comparativ cu virusurile A i B. De obicei nu cauzeaz epidemii, poate doar s
nsoeasc epidemiile de grip A i B i nu are impact sever asupra sntii publice [6, 15-18].
Genomul virusurilor gripale este de tip ARN monocatenar, de sens negativ, segmentat.
Numrul de segmente difer n funcie de tipul viral: virusurile gripale A i B au genomul
constituit din 8 segmente, iar virusul gripal C din 7 segmente. De asemenea, aranjarea anumitor
fragmente/segmente de ARN n virion pentru fiecare tulpin de virus gripal n parte, poart un
caracter individual [1, 5]. Cercetrile din mijlocul sec. XX au stabilit c ARN virusurilor gripale
-
19
este segmentat datorit proprietilor lor deosebite: rata nalt de recombinare; participarea la
reactivri multiple i ncruciate; reasortarea care are loc n natur, ceea ce sporete diversitatea
genetic a virusului; precum i inactivarea diverselor funcii ale virusului gripal cu anumii
ageni chimici i raze UV [5, 6, 16, 18].
Din cele 8 fragmente ale ARN viral 7 codific proteinele structurale i doar un singur
fragment (segmentul 8) proteinele nestructurale (NS1 i NS2), funciile crora sunt legate de
reproducerea virusului gripal. n general, genomul virusului gripal codific cel puin 12
(cunoscute n prezent) proteine virale, majoritatea dintre ele sunt necesare pentru replicarea
eficient a virusului n celula-gazd i la structurarea virionilor [5, 6, 18, 19].
Genomul viral mpreun cu proteinele virale: proteaza acid (PA), proteazele bazice (PB1
i PB2) i nucleoproteina (NP) formeaz nucleocapsida. Cea din urm fiind protejat de ctre
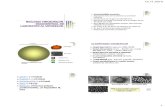
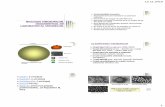
nveliul viral, derivat, de facto, din membrana celulei-gazd n timpul exocitozei (figura 1.1).
Fig. 1.1. Structura virusului gripal [19].
nveliul viral este compus predominant din proteinele de suprafa hemaglutinina (HA),
neuraminidaza (NA) i proteinele matri (M: M1 i M2). HA este responsabil de ataarea
particulei virale la receptorii celulei-gazd (acidul sialic) i de ptrunderea virusului n celul, n
special, n procesul contopirii membranei virale cu cea celular. NA nltur acidul sialic de la
HA pentru a-i facilita ptrunderea n celul i participarea n urmtoarele etape de reproducere a
virusului. Totodat, NA conduce la eliberarea particulelor virale de pe suprafaa celulei i
rspndirea virusului n tractusul respirator [5, 6, 19, 20].
Proteinele matri (M: M1 i M2) sunt componente ale stratului lipidic intern al
nveliului viral, fapt ce rezult n interaciunea proteinei M cu HA i NA. Proteina M1 este
mediatorul aranjamentului dintre nucleocapsida helicoidal i nveliul viral, astfel participnd la
structurarea virionului. Pe cnd, proteina M2 (prezent doar n virusurile gripale de tip A)
-
20
reprezint, de fapt, un canal ionic, care regleaz pH n procesul dezbrcrii virusului n
endosomi i n aparatul Golgi, unde i are loc sinteza HA. n acest caz, crearea unui pH acid este
o condiie absolut necesar n procesul de eliberare a virusului i stabilizare a conformrii HA n
timpul transportului lui intracelular, asigurnd astfel, existena mecanismului de interaciune
cooperativ a structurilor interne cu glicoproteinele externe i canalele ionice ale membranei
lipidice a virusului gripal (figura 1.1) [5, 6, 11, 19-22].
Dat fiind faptul, c principalii determinani antigenici ai virusurilor gripale A i B sunt
HA i NA glicoproteine transmembranare, ele sunt capabile s induc rspunsuri subtip-
specifice i imun, care sunt pe deplin protective n cadrul subtipului i numai parial protective
ntre diferite subtipuri. n baza antigenicitii acestor glicoproteine, virusurile gripale A, la
momentul actual, sunt grupate n 16 subtipuri de HA (H1-H16) i 9 subtipuri de NA (N1-N9).
Anume combinaia dintre diferitele subtipuri de HA cu diferitele subtipuri de NA i se numete
subtip viral: din 144 (16 x 9) subtipuri teoretic posibile, n prezent se cunosc 115, dintre care la
om au fost identificate subtipurile H1, H2, H3 de HA i N1, N2 de NA (i anume virusurile
gripale A(H1N1), A(H2N2), A(H3N2)). Aceste grupri devin considerabile cnd se efectueaz
analiza filogenetic a nucleotidelor i sunt deduse secvenele de aminoacizi ale genelor de HA i
NA respectiv [5, 6, 16-19].
Nomenclatura convenional a tulpinilor de virusuri gripale este folosit pentru codarea
lor i necesit conotarea tipului de virus gripal, specia gazdei (omis n cazul originii umane a
virusului), originea geografic, numrul de serie i anul izolrii, i numai pentru virusul gripal A
n paranteze se indic subtipurile de HA i NA ex. A/HongKong/156/97 (H5N1) tulpin de
virus gripal de tip A, izolat de la om, originea geografic Hong Kong, din proba nr. 156, n
anul 1997, cu subtipul H5N1 [5, 6, 15-18].
Variaia antigenic i circulaia virusurilor gripale ntre specii sunt cauza izbucnirilor
epidemice care au loc anual, mbolnvirile fiind favorizate de rspunsul imun neadecvat, chiar n
acele segmente de populaie care au fost anterior expuse infeciei gripale. Variaia antigenic este
particularitatea fundamental a virusurilor gripale A i B, care are loc la nivelul antigenelor de
suprafa HA i NA, reprezentnd astfel, un mecanism evolutiv de adaptare a virusurilor pentru
asigurarea supravieuirii lor ca specie [5, 6, 9, 15-18, 20, 22, 23].
Se cunosc dou mecanisme ale variaiei antigenice: minor (antigenic drift) i major
(antigenic shift). Variaia antigenic minor se ntlnete la toate tipurile de virusuri gripale, ns
totui, se presupune c virusul gripal C nu se supune drift-ului antigenic, deoarece mutaiile n
gena HA nu poart caracter consecvent. Totodat, att variaia antigenic minor drift-ul
antigenic, ct i cea major shift-ul antigenic se ntlnesc la virusul gripal de tip A. Acest fapt
-
21
poate fi explicat prin numeroasele epidemii, care au loc n fiecare sezon rece, ct i prin
cunoscutele pandemii de grip descrise n baza investigaiilor efectuate de ctre numeroi
cercettori virusologi [5, 6, 9, 15, 18, 24]. Drift-ul antigenic apare n rezultatul mutaiilor
punctiforme n genomul viral, ceea ce conduce, la rndul su, la modificarea proteinelor
determinanilor antigenici pn la pierderea capacitii de recunoatere de ctre sistemul imun al
gazdei. Anume mutaiile, inclusiv nlocuirile, deleiile i inseriile sunt responsabile de apariia
variantelor antigenice noi. Sub aciunea imunitii colective are loc selecia virusurilor, ce se
deosebesc dup caracteristicile antigenelor de suprafa de tulpina patern iniial. Cu toate
acestea, gene aparte ale virusurilor gripale umane acumuleaz mutaii cu o vitez aproximativ
constant, ceea ce permite de a meniona despre timpul molecular al virusului gripal. Variaiile
antigenice (drift) ale virusurilor gripale A i B apar i domin timp de 2-5 ani i numai dup
aceasta sunt nlocuite cu o alt diversitate antigenic [6, 16, 18, 20].
Pn la sfritul anilor 70 ai sec. XX era recunoscut ideea c subtipurile virusului gripal
A se schimb reciproc n mod consecutiv. n aceeai perioad savantul virusolog E. Kilbourn a
atestat faptul, c fiecare serotip al virusurilor gripale A umane imediat l nlocuiete pe
predecesorul su i singur va fi nlocuit de urmtorul. ns, cercetrile i studiile ulterioare ale
agenilor patogeni ai sezoanelor epidemice au demonstrat c lucrurile nu stau aa dup cum se
menionase. Astfel, particularitile etiologice ale gripei contemporane sunt prezentate de
cocirculaia a dou subtipuri de virus gripal A: A(H1N1) i A(H3N2), precum i a virusului
gripal de tip B, avnd, ns, fiecare din ele semnificaie epidemic diferit [15, 18, 25, 26].
n contrast, shift-ul antigenic, denot o schimbare momentan i profund n
determinanii antigenici, cu alte cuvinte, o nlocuire a ambilor sau a unuia din subtipurile de HA
i NA, ntr-un singur ciclu de replicare. Aceasta are loc n celula care este infectat simultan cu
dou sau mai multe virusuri gripale de tip A de diferite subtipuri. Odat ce distribuirea
segmentelor replicate ai genomului viral n interiorul virionului nou format are loc independent
de originea subtipului fiecrui segment, atunci poate s apar un aa virion, capabil deja de
replicare, care va purta informaia genetic a diferitor virusuri parentale (aa-numiii reasortani)
[5, 6, 9, 15-18, 23, 26].
n general, numeroasele teorii despre apariia variantelor pandemice a virusurilor gripale
pot fi divizate n ipotezele antroponozic i zooantroponozic. Se presupune, c una din
modalitile de meninere a virusului gripal A n populaia uman este persistena lui n
organismul uman. Asemenea presupunere a fost bazat pe cercetrile care au stabilit, c dup
ncheierea ciclului pandemic virusul gripal foarte repede disprea din circulaie. Conform
ipotezei date, virusul gripal n organismul uman trecea ntr-o form inactiv, care, ns, peste mai
-
22
multe luni sau chiar ani, el putea s se reactiveze. Reiese, c prezena infeciei latente i
persistente poate explica mecanismul shift-ului antigenic i includerile repetate n circulaie a
principalelor subtipuri ai virusului gripal de tip A [5, 6, 15, 26].
Urmrind istoria ciclurilor pandemice ale virusului gripal A, a putut fi confirmat ipoteza
persistenei virusurilor gripale prin reapariii repetate la diferit interval de timp, fapt ce nu
exclude posibilitatea persistenei lui n rezervorul animal. Pe de alt parte, exist date veridice, c
virusurile de origine aviar, uman i porcin clasic au un strmo comun i anume virusul
gripal de origine aviar. Unii cercettori presupun, c anume virusul gripal aviar trece bariera de
specie, infectnd la nceput porcii, apoi dup o anumit adaptare, alte mamifere, i ulterior,
nimerete n populaia uman. Acest fapt, a putut fi urmrit n 1918, cnd a aprut pandemia de
grip spaniol, determinat de virusurile gripale ale mamiferelor, HA crora era strns legat
de varianta HA aviare [4-7, 9, 15-18, 20, 22, 23, 25, 27-29].
Modalitatea cu care virusul gripal este capabil s treac periodic bariera de specie poate
explica ipoteza despre prezena n una din genele polimerazice a mutaiei care duce la sporirea
gradului de variabilitate a agentului, la apariia a unui numr considerabil de variante i la
crearea unor condiii mai bune pentru adaptarea n organismul diferitor specii de animale i
psri. ntru confirmarea ipotezei date au fost aduse dovezi, care au demonstrat c HA virusurilor
gripale A(H1N1) izolate de la porcinele bolnave din Europa de Nord a fost nrudit antigenic i
genetic cu HA aviar. Apoi, aceste virusuri porcine noi au putut din nou s treac bariera de
specie i s provoace o epizootie la curcani fr producerea de variaii genetice. Aceti ageni se
caracterizau printr-o instabilitate extrem, variabilitate i vitez de evoluie nalte. Faptul dat ar
putea explica fenomenul istoric de trecere a barierei de specie a virusului gripal aviar, astfel
fcnd posibil apariia unei linii clasice stabile de virus gripal porcin n SUA [5, 18, 24, 30].
Un alt mecanism posibil al shift-ului antigenic, n rezultatul cruia au aprut dou tulpini
pandemice de virusuri gripale este reasortarea genelor virale. n prezent sunt dovezi
convingtoare asupra faptului, c tulpinile pandemice asiatic A(H2N2) (1957) i de Hong
Kong A(H3N2) (1968) au aprut n rezultatul reasortrii virusurilor umane i aviare; genele
HA, NA i a unei proteine din complexul polimerazic avnd origine aviar. Alte gene ale acestor
tulpini pandemice sunt similare cu genele analoage ale virusurilor gripale umane anterioare
A(H1N1) i A(H3N2). Cu toate acestea, porcinele fiind sensibile att fa de virusurile gripale
umane, ct i fa de cele aviare, au putut fi gazde intermediare pentru formarea variantelor shift
(figura 1.2) [5, 18, 21, 24].
-
23
Fig. 1.2. Mecanismul shift-ului antigenic [6].
Astfel, similaritatea genofondului virusurilor gripale de tip A umane i ale altor specii de
mamifere n biosfer acord o actualitate deosebit fenomenului de participare a virusurilor
gripale animale i aviare n formarea variantelor pandemice. Totodat, trebuie de menionat, c
-
24
semnificaie epidemic considerabil pentru umanitate, pe parcursul studierii virusurilor gripale
au avut doar 3 subtipuri de virusuri gripale A: A(H1N1), A(H2N2) i A(H3N2) [18, 23].
Aa dar, pandemiile de grip au loc atunci, cnd i face apariia un nou virus gripal, la
care marea majoritate a populaiei umane de pe glob nu are protecie imun sau aceasta este
foarte slab, care are un potenial nalt de infectare a oamenilor provocnd cazuri clinice de boal
i cu o transmisibilitate nalt de la persoan la persoan. Aceste particulariti rezult ntr-un
impact de talie global a infeciei gripale ce afecteaz un procent considerabil de persoane i
cauzeaz o sporire a nivelului de mortalitate [1, 4, 6, 9, 16-18, 20, 23-36].
Practic, ntru confirmarea celor expuse mai sus poate fi adus exemplul ultimei pandemii
din irul istoric care i a avut loc n 2009-2010 prima pandemie a sec. XXI. Aceast pandemie,
de o severitate moderat, a fost provocat de un nou virus gripal reasortant A(H1N1)pdm09.
Analiza genetic a acestui virus pandemic a dezvluit o nou combinaie a genelor virusurilor
umane, porcine i aviare eurasiatice. Virusul conine genele HA, NP i NS ale virusului clasic de
origine porcin, genele PB2 i PA ale virusurilor aviare nord americane, gena PB1 a virusurilor
H3N2 de origine uman i genele NA i M ale virusurilor porcine eurasiatice similare celor
aviare. Nu se exclude probabilitatea, c acest virus provine de la virusurile porcine triplu
reasortante circulante la porcine n perioada anilor 1997-1998 coninnd genele HA, NA i PB1
similare virusurilor umane i genele interne PB2 i PA de origine aviar [4, 7, 15, 16, 20, 23, 30,
37, 38]. Cercetrile, ns, au demonstrat c aceste virusuri sunt srace n determinani moleculari
specifici pentru adaptarea la gazda uman, astfel sugernd un rol nc necunoscut/nedocumentat
a marcherilor moleculari asociat transmisiei umane [7, 11, 20, 28, 30, 38-51]. Aceste virusuri, n
consecin, nu posed marcheri asociai cu virulena nalt sau patogenitatea care au fost
observate la virusul H1N1 din 1918 sau patogenitatea nalt a virusurilor H5N1 [12, 21, 30, 42].
Totodat, analiza filogenetic a demonstrat c secvena de HA a tulpinii de virus gripal
A(H1N1)pdm 09 cel mai mult se aseamn cu cea a tulpinii din 1918 i a fost antigenic distinct
de virusurile H1N1 umane recente, precum i de componenii vaccinurilor. Antigenic, ns,
virusurile A(H1N1)pdm09 sunt omogene i printre tulpinile istorice de virusuri gripale, sunt cele
mai similare cu virusurile gripale porcine A(H1N1) clasice (triplu reasortant) [20, 21, 23, 28, 30,
42, 52].
Printre virusurile A(H1N1)pdm09 analizate au existat doar cteva substituii de
aminoacizi la nivel de HA, i nici una din ele nu au avut efect antigenic. Deci, variaia antigenic
printre virusurile A(H1N1)pdm09 circulante n populaia uman este la moment mai mic fa de
cea observat n timpul unui sezon gripal tipic [15, 20, 23].
-
25
Cu toate acestea, s-a atestat c virusul A(H1N1)pdm09 posed o transmisibilitate nalt i
are un avantaj biologic distinct n replicare, transmisie, tropism i patogenez n comparaie cu
ambele virusuri sezoniere reprezentative A(H1N1) i A(H3N2). Similaritatea n manifestrile
epidemiologice ale acestei tulpini de virus gripal a fost observat n populaie att n emisfera
nordic, ct i n cea sudic [15, 21, 30, 42, 48, 53-55].
Astfel, pe msur ce cunoatem mai mult despre virusurile gripale, un efort considerabil
este necesar pentru a rspunde la ntrebrile: care sunt factorii determinani n transmisia
virusului de la o specie la alta; care sunt factorii ce determin reasortarea virusurilor, - factori
critici n apariia unor noi virusuri pandemice. La nivel global, ns, exist posibilitatea de a
urmri evoluia virusurilor gripale, practic n timp real, ceea ce ne poate asigura cu informaii
inestimabile ntru stabilirea factorilor ce determin patogenitatea i/sau transmisibilitatea lor.
1.2. Caracteristica fenotipic i genotipic a virusurilor gripale
Virusurile gripale se caracterizeaz printr-o structur unical a genomului, dar i prin
instabilitate genetic, fapt datorat mutaiilor punctiforme i evenimentelor de reasortare, ceea ce
contribuie la apariia noilor variante sau tulpini de virusuri gripale cu potenial epidemic sau
pandemic. Att epidemiile, ct i pandemiile au un impact economic substanial datorit
costurilor profilaxiei i tratamentului, absenteismului, vizitelor la medic, precum i excesul de
spitalizri. Prin urmare, este necesar o nelegere detaliat a mecanismelor ce determin
patogenitatea i transmisia virusurilor gripale ntre specii, combinat cu disponibilitatea
msurilor efective de prevenie i tratament ntru realizarea msurilor de control i rspuns al
infeciilor gripale [23].
n timp ce persoanele dezvolt imunitate de lung durat la o anumit tulpin de virus
gripal, mutaiile antigenice fa de genomul virusului gripal rezult n proteine ce sunt
recunoscute ntr-o msur mai mic de ctre sistemul imun uman, lsnd persoanele susceptibile
n viitor la infectare. Evoluia fenotipului antigenic apare punctiform cu episoade de nnoire
intercalate de perioade de staz, n timp ce evoluia genetic pare mai continu, ceea ce
sugereaz c un numr relativ mic de modificri genetice sau combinaii de modificri genetice
pot conduce la apariia modificrilor la nivelul fenotipului antigenic. Populaia de virusuri
gripale evolueaz continuu n fenotip antigenic n aa-numitul proces de drift antigenic care se
realizeaz prin intermediul antigenelor de suprafa HA i NA. Aceste dou proteine sunt codate
de segmentele 4 i 6 ale genomului viral (constituit n general din 8 segmente), localizate pe
suprafaa virionului, ele sunt prima int pentru rspunsul imun al gazdei. Celelalte ase
segmente sunt codificate de ctre proteinele: PB2 (segmentul 1), PB1 (segmentul 2), PA
(segmentul 3), NP (segmentul 5), M1 i M2 (segmentul 7), NS1 i NS2/NEP (nuclear export
-
26
protein) (segmentul 8). O protein-accesoriu, suplimentar, PB1-F2 poate fi codat de segmentul
2 i ea poate conferi proprieti de virulen virusurilor care o expreseaz prin faptul, c ea se
asociaz cu proteinele mitocondriale inducnd procesul de apoptoz n celulele imune. Acest
factor de virulen a fost identificat printre virusurile gripale de tip A, stabilindu-se, totodat,
asocierea lui cu patogenitatea sporit a virusului gripal nalt patogen A(H5N1), precum i a
virusului gripal pandemic din 1918. Proteina fragmentat PB1-F2 cu fragment deschis pentru
citire (PB1-F2 ORF open reading frame) s-a dovedit a fi legat de replicarea viral continu,
inclusiv i de rspunsul proinflamator sporit. Totodat, proteina nestructural NS1 suprim
expresia genelor antivirale n celulele-gazd. Marcherii virulenei precum i factorii ce contribuie
la transmisia ntre specii au fost identificai n proteina PB2, pe cnd determinanii rezistenei
antivirale au fost depistai n proteinele NA i M [56-61].
Din momentul izolrii pentru prima dat n 1930 a virusului gripal A(H1N1) de la
porcine - antigenic foarte asemntor virusului gripal uman reconstruit A(H1N1) din 1918, ele
mprtesc, probabil, un strmo comun i pn la sfritul anilor 90 ai secolului XX acest virus
gripal clasic porcin a circulat n populaia porcinelor i a fost antigenic relativ stabil. n anul
1998, sau mai nainte, acest virus gripal clasic porcin a reasortat cu virusul gripal uman
contemporan A(H3N2) i cu un virus aviar de linie american cu subtip necunoscut, rezultnd n
apariia n populaia porcin nord-american i mai trziu n populaia porcin asiatic a virusului
gripal porcin triplu reasortant H3N2 (rH3N2). Cercetrile au demonstrat c acest virus rH3N2
posed genele HA, NA i PB1 de la virusurile gripale umane, genele PA i PB2 de la virusurile
gripale aviare i genele interne NP, M i NS de la virusurile gripale porcine [36, 42, 56, 58].
Datorit diferenelor intrinsece dintre gazde (ex. receptorii celulari) i ale aprrii selective
(sistemele umoral i celular de aprare nnscute i dobndite), diferite modificri n structura
antigenic a virusurilor gripale sunt provocate mai mult n populaia porcin dect n cea uman.
Drept consecin, drift-ul antigenic al virusurilor gripale urmeaz ci diferite la porcine
comparativ cu populaia uman. Precum drift-ul antigenic n virusurile gripale umane A(H3N2)
este, de obicei, atribuit aprrii imune stabilite n populaia uman ca urmare a infeciilor
precedente cu variantele anterioare de virusuri gripale, tot aa potenialul comun al mutaiilor de
difereniere a cluster-elor pot sugera c modificrile antigenice n virusurile gripale porcine, de
asemenea, sunt urmare ale aprrii imune. Periodic, aceste mutaii ar putea fi selectate n ambele
gazde n baza avantajelor diferite fa de evaziunea imun sau ar putea fi sporadic cuplate cu alte
mutaii cu acelai efect [36, 42, 62-65].
Totodat, virusul gripal A(H1N1) din 1918 a circulat n populaia uman pn la
declanarea n 1957 a pandemiei de grip provocat de virusul gripal A(H2N2) (gripa asiatic).
-
27
n aceast perioad s-a constatat un drift antigenic substanial al virusului gripal A(H1N1)
departe de virusul din 1918. De la nceputul anilor 1950 virusul gripal A(H1N1) a reaprut n
populaia uman n 1977, iar din 1977 pn n 2009 a fost o evoluie antigenic substanial, fapt
ce a determinat rennoirea de 8 ori a componentului H1 al vaccinului antigripal [36, 42, 56-64].
Staza antigenic relativ a virusului gripal clasic H1N1 n populaia porcin s-a estimat
pn n 1998, n acelai timp, observndu-se drift-ul antigenic substanial al virusului gripal
H1N1 n populaia uman, n cele din urm, acestea au condus la crearea unui decalaj antigenic
esenial ntre virusurile gripale clasice porcine A(H1N1) i virusurile gripale umane A(H1N1).
De asemenea, cercetrile au demonstrat, c din 2005, izolatele virusurilor gripale umane H1N1
de origine porcin au manifestat cel mai nalt grad de similaritate cu virusurile porcine H1
circulante n Asia i SUA i c proteina N1 are legtur cu virusurile porcine circulante n
Europa [41]. Se consider c porcinele joac un rol vital n transmisia ntre specii a virusurilor
gripale prin faptul c ele poart receptori att pentru tulpinile de virus gripal aviar, ct i pentru
tulpinile de virus gripal uman. Aceasta a pus n eviden porcinele drept "vas de amestec" n care
materialul genetic poate fi schimbat cu un potenial de a rezulta ntr-un nou progenitor viral la
care populaia uman este nalt susceptibil i nu posed imunitate. Astfel, populaia porcin a
devenit un rezervor pentru virusurile gripale H1N1 cu potenial de cauzare a unor epidemii
majore sau posibile pandemii n populaia uman [36, 42, 58-64].
Caracteristica unui ir de virusuri gripale A(H1N1)pdm pentru determinarea
proprietilor antigenice, n prima jumtate a anului 2009, a pus n eviden faptul c antigenic
aceste virusuri sunt omogene i printre virusurile istorice, ele sunt antigenic similare cu virusurile
clasice porcine A(H1N1), precum i virusurile triplu reasortate A(H1N1) ale liniei nord-
americane care au circulat n populaia porcin n ultimii 10 ani n SUA, i care ocazional au
infectat populaia uman n aceiai perioad. Analiza antigenic a virusurilor A(H1N1)pdm
izolate n perioada nominalizat a demonstrat prezena doar a cteva substituii ale aminoacizilor
n gena HA, ns niciuna nu s-a dovedit a avea vre-un efect antigenic. Variaia antigenic printre
aceste virusuri s-a constatat a fi, la acel moment, mai mic dect variaia antigenic observat n
populaia uman n timpul unui sezon tipic/epidemic de grip [42].
Cu toate acestea, sezonul epidemic 2008-2009 s-a caracterizat prin circulaia att a
virusurilor sezoniere clasice cunoscute A(H1N1), A(H3N2) i B, ct i a virusurilor
A(H1N1)pdm. Caracteristica antigenic a acestor virusuri gripale a demonstrat c virusurile
gripale A(H1N1) au fost asociate cu tulpina A/Brisbane/59/2007-like; virusurile A(H3N2) au
fost similare tulpinii A/Brisbane/10/2007-like, ambele tulpini fiind componente ale vaccinului
antigripal recomandat de OMS pentru sezonul epidemic 2008-2009. Virusurile gripale de tip B
-
28
circulante n sezonul respectiv s-au divizat n dou linii distincte reprezentate de tulpinile
B/Yamagata/16/88 i B/Victoria/02/87. Virusurile gripale de tip B care au corespuns liniei
B/Yamagata au fost analoage tulpinii B/Florida/04/2006, de asemenea, component al vaccinului
antigripal din sezonul nominalizat. ns, paralel cu virusurile enumerate, au fost izolate i tulpini
de virusuri gripale de tip B care aparineau liniei B/Victoria, care la acel moment nu au fost
componente ale vaccinului antigripal recomandat pentru sezonul epidemic 2008-2009. n acest
context, este necesar de interpretat cu precauie rezultatele caracterizrii antigenice, deoarece
aceste rezultate s-au bazat, de facto, pe reacia de hemaglutinoinhibare (RHAI) folosind un panel
de seruri de referin existente n acea perioad, i care puteau s nu corespund proteciei clinice
mpotriva virusurilor circulante asigurate de vaccinul antigripal [66-68].
Vaccinarea antigripal anual presupune asigurarea unei protecii maxime mpotriva
acelor tulpini de virusuri gripale care coincid cu virusurile vaccinale, ns o protecie limitat
sau, n general, lipsa ei poate fi observat atunci, cnd tulpinile de virus gripal vaccinal i cele
circulante sunt aa de diferite, nct s fie din linii diferite aa cum a putut fi observat cu liniile
de virus gripal de tip B (linia B/Yamagata i linia B/Victoria - una fiind component al vaccinului
nu poate asigura protecie mpotriva celeilalte). Aadar, caracteristica antigenic a tulpinilor de
virusuri gripale A(H1N1)pdm a demonstrat c aceste virusuri au fost i sunt antigenic i genetic
diferite de tulpinile A(H1N1), fapt ce a sugerat lipsa proteciei n urma vaccinrii cu vaccinul
trivalent recomandat pentru sezonul epidemic 2008-2009 fa de virusul gripal nou
A(H1N1)pdm [66].
Apariia recent a virusului pandemic (H1N1) 2009, cunoscut anterior ca virus gripal de
tip A de origine porcin, a condus la infectarea pn la mijlocul anului 2009 peste 296 mii de
persoane pe ntreg globul pmntesc cauznd n perioada menionat cca 3486 cazuri de deces.
Analiza mutaiilor de adaptare a virusului gripal de tip nou A(H1N1) 2009 a devenit o prioritate
pentru cercettori care au putut evalua probabilitatea c virusurile de la alte specii non-umane se
vor adapta la populaia uman. Virusul gripal pandemic const din mai multe gene virale
reasortate de diferit origine. Dou gene polimerazice din cele 8 ale ARN genomic segmentat, i
anume PB2 i PA, au fost de origine aviar aparinnd liniei nord americane i au fost introduse
n populaia porcin aproximativ n anul 1998. Alt gen polimerazic, PB1, de asemenea, a
evoluat recent din virusul gripal uman sezonier A(H3N2) practic n acelai an. n particular,
aceast gen PB1 a virusului A(H3N2), este cunoscut c, ar fi provenit de la virusul gripal aviar
care a intrat n populaia uman n anul 1968. Totui, genele proteice HA, NP i NS ale virusului
gripal pandemic (H1N1) 2009 se trag direct de la virusul gripal A clasic porcin al liniei nord
americane care poate fi urmrit pornind de la virusul gripal din anul 1918. Alte dou gene NA i
-
29
M, avnd originea de la virusul gripal porcin eurasiatic, au fost introduse de la psri
aproximativ n anul 1979. Cu toate c a fost determinat originea segmentelor genelor virusului
gripal pandemic (H1N1) 2009, nu este clar mecanismul de transformare a semnelor
aminoacizilor gazd-specifice, deoarece genele virusului gripal de tip nou au evoluat dup
introducerea lor n circulaie n populaia porcin civa ani n urm [30, 31]. Totodat, interesant
este faptul, c n populaia porcin au fost observate aceleai substituii care au avut loc la nivelul
genelor HA1 (substituia S203T) i NA (substituiile V106I i N248D) ale virusurilor gripale
A(H1N1)pdm circulante n populaia uman. Sistemul imun de aprare al gazdei este considerat
a fi principala for motrice selectiv a substituiilor de aminoacizi, care pot conduce la apariia
drift-ului antigenic, iar HA este inta principal a anticorpilor neutralizani. Diversitatea genetic
a HA este mult mai nalt fa de gena NA att n populaia porcin, ct i n populaia uman.
Genele HA i NA ale izolatelor virusurilor gripale A(H1N1)pdm din pri distincte ale lumii sunt
legate unele de altele cu o distan relativ mic i au o singur origine comun, cum era de
ateptat pentru un focar pandemic. Remarcarea faptului c majoritatea izolatelor derivate din
populaiile uman i porcin par s fi provenit din izolatele derivate din populaia uman, i este,
de asemenea, n concordan cu o frecven mai mare de transmisie de la om la om i de la om la
porcine n comparaie cu transmisia zoonotic de la porcine la om. ntr-adevr, rapoarte cu
privire la izbucnirea gripei sugereaz c virusul a evoluat n tcere la porcine pn la
introducerea sa n populaia uman, dup care s-a rspndit rapid printre oameni i frecvent s-a
retransmis de la oameni la porci. S-a demonstrat c virusul gripal A(H1N1)pdm a evoluat i s-a
transferat (shift antigenic) de la o clad prototip iniial amestecat la clada prototip 7
predominant. Selecia i evoluia ulterioar a cladei 7 a rezultat n apariia n circulaie a
variantelor cu mutaiile genetice D222G/N sau E. Mutaia D222G n proteina genei HA ce
conduce la lrgirea specificitii receptorilor s-a demonstrat a corela cu debutul clinic al bolii i
frecvent detectat n cazurile severe/fatale ale gripei pandemice la oameni [32-34]. De asemenea,
s-a atestat c substituia D222N este mai frecvent n cazurile fatale ale bolii la oameni, pe cnd
aceleai substituii observate n izolatele virusurilor gripale A(H1N1)pdm de la porcine nu
provoac semne clinice severe de boal [58-64, 69-73].
Caracteristica genotipic a virusurilor gripale A(H1N1)pdm a atestat c n cadrul fiecrui
segment de gene exist o congruen nalt printre virusurile gripale pandemice secveniate pan
la mijlocul anului 2009, fapt ce a sugerat c introducerea cross-specific n populaia uman a
fost un singur eveniment sau mai multe evenimente ale virusurilor genetic similare. Analiza
genoamelor virusurilor A(H1N1)pdm din Mexic i SUA, la acel moment, a evideniat 5 variante
genomice mici: secvena consensus; mutaia T373I n gena NP pereche cu mutaia M582L n
-
30
gena PA; substituiile aminoacizilor V106I i N247D n gena NA pereche cu substituia V100I n
gena NP; substituiile aminoacizilor S206T n gena HA1 grupndu-se cu ambele substituii
V106I i N247D din gena NA, substituiile V100I din gena NP i cu substituia I123V din gena
NS1; substituiile aminoacizilor S91P, V323I mpreun cu substituia S224P din gena PA [42-
44]. Includerea izolatelor din Mexic sau alte state vecine printre aceste 5 variante genomice
reflect probabilitatea c aceste variante genomice timpurii au reprezentat introducerea iniial
independent n SUA din Mexic. Datorit intervalului scurt de timp de la detecia pentru prima
dat a virusului gripal A(H1N1)pdm, nu era clar ce efect, dac era n general, au putut avea
aceste variaii genomice asupra caracteristicilor virale, precum transmisibilitate sau patogeneza.
Analiza de secveniere, ns, nu a identificat caracteristici moleculare deosebite anterior, n
cercetri ai altor virusuri gripale de tip A, presupuse c ar conferi transmisibilitate sporit sau
virulen [38, 42, 45-46]. Cunoscutul receptor al locusului de legare a proteinei hemaglutinina
H1 s-a dovedit a fi tipic multor altor virusuri gripale clasice porcine H1N1 izolate n SUA n
perioada nominalizat. Totui, au fost atestate unele mutaii detectate n gena HA a virusurilor
gripale A(H1N1) 2009 ce se deosebeau de secvena consensus a virusurilor clasice porcine,
niciuna ns, nu a fost identificat n locusul funcional semnificativ al receptorului de legare
cunoscut. Dup cum a fost de ateptat, multe din virusurile A(H1N1) 2009 conineau substituii
de aminoacizi la locusul antigenic presupus n comparaie cu gena HA a virusurilor gripale H1
sezoniere [38-42, 58, 61].
Analiza de secveniere, de asemenea, a pus n eviden prezena substituiilor de
aminoacizi n gena HA a virusurilor gripale A(H1N1)pdm n poziiile S220T, D239G/N/S,
Y247H, E252K, M247V, Q310H i E391K. S-a observat o predominan a mutaiei D239G n
unele ri n cazurile soldate cu deces, similar cu substituia D222G, dar care, probabil a fost n
concordan cu vrsta pacienilor, factorii de risc, manifestarea i progresarea bolii [73, 77, 78].
Un interes aparte prezint particularitile specifice, individuale ale structurii locusului de
legare a HA virusului gripal A(H1N1)pdm, i anume, mutaiile D94N, N125D i V250A.
Substituiile perechi n poziiile 94 i 250 (D94N i V250A) sunt similare n baza structurii
situsului de legare cu acizii sialici cu tipul receptorului caracteristic pentru esuturile umane. Cu
toate acestea, substituia V30A, identificat n structura virusului gripal A(H1N1)pdm, poate
compensa aciunea substituiei N125D, fapt care poate fi urmrit la prezena concomitent a
substituiilor D94N i V250A, cnd schimbul Valinei n poziia 250 cu Alanina influeneaz
semnificativ manifestarea substituiei D94N. n special, este stabilit, c substituiile 30A i 125N
sporesc afinitatea HA fa de receptorii sialici de tip uman. Mai mult ca att, combinaia 94D,
125N i 250V este caracteristic pentru izolatele de virus A(H1N1) din 1918, iar combinaia
-
31
94N, 125D i 250A conduce la creterea afinitii HA ctre receptorii virusurilor gripale aviare.
Pentru virusul gripal A(H1N1)pdm este caracteristic combinaia 94D, 125N i 250V care
completamente corespunde cu reziduurile aminoacidice n poziiile respective ale virusului gripal
care a cauzat pandemia spaniol din 1918. Este necesar de subliniat, c combinaia de
substituii E190D, Q226L i G228S n hemaglutininele H1, H2 i H3 conduce la trecerea
ambigu a HA de pe receptorii de tip aviar spre receptorii celulelor umane, fapt ce se realizeaz
parial [50, 73, 77, 78].
Secvenierea unor tulpini de virusuri gripale A(H3N2) a atestat c n toate secvenele
genei M1 a fost detectat substituia K174R, la nivel de gena HA a fost caracteristic substituia
K29R, iar pentru gena NA au fost caracteristice dou substituii sinonimice 351 A>G (n codonul
T117) i 408 G>A (n codonul Q136). Acestea, posibil, au indicat la faptul c tulpinile de
virusuri gripale A(H3N2) din sezonul 2008-2009 au prezentat o continuitate evoluionist a
tulpinilor de virusuri gripale A(H3N2) din sezonul 2007-2008 [74-76]. Este necesar de
menionat, c n secvenele genelor NA ale virusurilor gripale A(H3N2) din sezonul 2008-2009 a
fost detectat substituia D147N n locusul de glicozilare. Se cunoate, ns, c glicozilarea n
locusul 130 N1 (ce corespunde locusului146 n N2) a tulpinii A/Wisconsin/33 (H1N1)
deregleaz interaciunea dintre NA i plasminogenul, ceea ce conduce la diminuarea patogenitii
virusului [73, 74-76].
Marea majoritate din tulpinile gripale A(H3N2) cercetate n perioada nominalizat au
purtat n secvena domenului cu terminaia C a proteinei M1 substituia K174R, care sporete
tendina sectorului dat al lanului proteic spre formarea -spiralei. Proteina M2 s-a dovedit a
conine substituia S31N care este caracteristic rezistenei la remantadin. Cea mai important
consecin a substituiilor de aminoacizi ce rezult n descreterea afinitii pentru receptori este
efectul lor asupra eficienei infeciei i transmisiei [73, 79]. A fost remarcat faptul, c n sezonul
2009-2010 virusurile gripale A(H3N2) practic nu au participat n procesul epidemic, iar n
sezonul 2010-2011 aceste virusuri nu au avut semnificaie epidemic [73, 79, 80].
Comparaia hrilor genetice i antigenice ale virusului gripal A(H3N2) a atestat c
impactul antigenic a modificrilor genetice variaz n dependen de natura substituiilor de
aminoacizi, poziionarea lor structural i interaciunea epistatic cu alte situsuri [75].
Sezoanele epidemice 2008-2009, 2009-2010 i 2010-2011 s-au caracterizat prin reapariia
n circulaie a virusurilor gripale de tip B apartenente liniei B/Victoria n care au avut loc
schimbri semnificative ai proprietilor antigenice izolatele au fost similare unei tulpini etalon
noi, i anume B/Brisbane/60/2008. Analiza genetic a tulpinilor de virusuri gripale de tip B,
izolate n perioada anilor 2009-2011, a demonstrat c aceste tulpini au aparinut liniei B/Victoria,
-
32
n special cladei 111-ii, sau tulpinii similare B/Brisbane/60/2008 cu substituii caracteristice de
aminoacizi (V146I, N165K) n gena HA, afectnd regiunile antigenice (buclele 150 i 160).
Indiferent de faptul c timp de dou sezoane epidemice (2009-2010 i 2010-2011) n lume a
circulat i a dominat virusul gripal pandemic A/H1N1/pdm09, virusurile gripale de tip B nu au
fost eliminate din circulaie, ns rata lor n sezonul 2009-2010 a fost foarte joas, iar n sezonul
2010-2011 moderat [81-83].
n acest context, spre deosebire de sezonul 2008-2009, n sezonul epidemic 2009-2010 s-
a intensificat circulaia tulpinilor de virusuri gripale de tip B apartenente liniei B/Victoria, iar
circulaia virusurilor gripale B/Yamagata a diminuat. Astfel, componenta ce ine de tulpina
virusului gripal B din vaccinul antigripal pentru sezonul nominalizat s-a schimbat, tulpina
B/Florida/04/2006-like linia B/Yamagata a fost nlocuit cu tulpina B/Brisbane/60/2008-like
linia B/Victoria [66, 67, 81-84].
Comparaia filogenetic a tulpinilor de virusuri gripale de tip B linia B/Victoria a
demonstrat c substituiile de aminoacizi N75K, N165K, i S172P n regiunea de codificare HA1
a genei HA definesc clada genetic B/Brisbane/60/2008. Toate virusurile gripale de tip B
apartenente liniei B/Victoria circulante n sezonul 2010-2011 au fcut parte din clada
menionat. De asemenea, s-a atestat c majoritatea acestor virusuri au purtat substituia de
aminoacizi I146V n gena HA comparativ cu virusul vaccinal B/Brisbane/60/2008 i multe
virusuri au purtat i substituia L58P, ns nici-una din ele nu s-a dovedit a avea efect antigenic
[84].
Arborele filogenetic construit n baza regiunii de codificare HA1 a genei HA a tulpinilor
de virusuri gripale de tip B apartenente liniei B/Yamagata, care a reaprut n circulaie n sezonul
2012-2013, a pus n evidena prezena substituiilor de aminoacizi S150I, N165Y, G229D care
definesc grupul genetic B/Bangladesh/3333/2007. ns o parte din virusuri (grupul de virusuri
din Suedia, Finlanda i Estonia) s-au plasat n grupul genetic distinct reprezentat de tulpina
B/Brisbane/3/2007 virus prototip al acestui grup. Gena HA a tulpinii de virus gripal
B/Athens/9784/2011, ce a prezentat un model distinct n reacia HAI, s-a plasat n grupul genetic
B/Bangladesh/3333/2007 codificnd, ns, substituiile V29A, L172Q i M251V comparativ cu
virusurile de referin [84].
Este cunoscut faptul c reasortarea rearanjarea segmentelor genelor virale n celulele-
gazd infectate cu dou sau mai multe virusuri gripale reprezint un mecanism important n
evoluia virusurilor gripale. Infeciile mixte cu multiple tipuri/subtipuri de virusuri gripale pot
conduce la apariia de reasortri. Un aspect substanial al reasortrii este generarea unor noi
tulpini de virusuri gripale cu potenial pandemic, unde virusurile gripale umane obin un subtip
-
33
nou de HA i/sau NA de la virusurile gripale aviare i/sau porcine. Reasortarea, de asemenea,
faciliteaz apariia de fenotipuri mai virulente prin permiterea virusurilor de a cpta segmente
cu marcheri pentru virulen. Prin reasortare virusurile gripale au obinut gene cu mutaii ce in
de rezistena la remediile antivirale i aceste virusuri rezistente se pot replica tot aa de eficient
ca i cele sensibile i, de asemenea, pot avea o capacitate de transmisie nalt [85].
Vaccinurile curente contra gripei sezoniere sunt destinate pentru asigurarea proteciei
specifice n dependen de tulpin contra dou subtipuri ai virusului gripal de tip A circulante
(H1N1 i H3N2) i contra unui virus gripal de tip B. inta principal a acestor vaccinuri este HA
care deine rolul de mediere a ptrunderii n celula-gazd. Astfel, anticorpii neutralizani indui
de vaccin blocheaz ptrunderea virusului n celul sau prin prevenirea atarii virusului de
receptorii ce conin acid sialic de pe suprafaa celulei-gazd sau prin interferena cu fuziunea
viral mediat de HA [85].
Substituiile de aminoacizi S31N, V27A, A30V, G34E i L26F reprezint mutaiile ce
induc rezistena la adamantane (amantadina i remantadina) n virusurile gripale [73, 85, 86].
inta adamatanelor este proteina M2 a virusurilor gripale, care funcioneaz drept canal ionic
activat de acizi i este necesar pentru eliberarea nucleoproteinei dup fuziunea cu membrana
endosomal. Adamatanele inhib replicarea viral prin prevenirea deschiderii canalului ionic
M2, interfernd astfel cu dezbrcarea virusului n timpul endocitozei [87-90]. Cea mai
frecvent substituie ce caracterizeaz rezistena fa de adamatane este substituia S31N, care
predomin n 98-100% din tulpinile transmisibile rezistente la amantadin H1N1, H5N1 i
H3N2 izolate de la oameni, psri i porcine n ultimii zece ani, pe cnd mutaiile V27A i L26F
sunt mai puin frecvente [87-89]. Fiecare mutaie rezult sau n legarea redus a remediului
antiviral de ligandul M2 sau n extensia canalului ionic M2 i ambele permit canalului s-i
exercite funcia n prezena antiviralelor [87-89, 91-93]. Cercetarea mutaiilor punctiforme la
nivelul reziduurilor cptuelii porilor canalului M2 a estimat numrul mic de variante naturale
[87, 90, 91]. Astfel, numeroi mutani n captul N-terminal apos al porului rein abilitatea de a
conduce selectiv protonii asupra altor ioni, dei dependena de magnitudine i pH ai
conductibilitii lor variaz. Totui, doar cteva mutaii V27A, S31N i L26F au proprieti
similare cu canalul ionic M2 la cele mai distale locusuri [87-89, 91-93].
Rezistena la amantadin a evoluat rapid in vivo din momentul administrrii ei pacienilor
cu infecie gripal. Baza genetic a rezistenei fa de amantadin este asociat cu substituiile de
aminoacizi, dup cum s-a menionat, n regiunea transmembranar a genei M2. Cercettorii au
atestat o inciden nalt de izolate de virus gripal A(H3N2) rezistente la amantadin, care n
gena M2 posed mutaia Ser-31-Asn i o schimbare dubl n gena HA la reziduurile din poziiile
-
34
193 i 225 (clada liniei N). Totui, puin se cunoate despre faptul, dac modificrile la nivelul
genei HA au fost sinergice cu modificrile care au avut loc n gena M2 ca rspuns la presiunea
selectiv a medicamentului sau au avut loc separat i s-au asociat randomizat cu mutaiile de
mbuntire a afinitii [87-89, 91-93].
O alt clas de remedii antivirale este reprezentat de inhibitorii neuraminidazei (NAI):
oseltamivir (Tamiflu) i zanamivir (Relenza) [94]. inta NAI este proteina NA a virusurilor
gripale care este responsabil de clivajul reziduurilor acidului sialic al celulelor-gazd permind,
prin aceasta, eliberarea virionilor aflai n maturizare. NA, de asemenea, are importan n
stabilirea infeciilor respiratorii la nivelul cilor respiratorii superioare, deoarece clivarea
acidului sialic pe suprafaa mucoasei expune celulele epiteliale la aciunea virusului. NAI previn
eliberarea virionilor din celulele infectate i, astfel, reduce att infeciile cilor respiratorii
superioare, ct i durata simptoamelor [87, 95, 96]. La nivelul genei NA au fost observate
substituiile de aminoacizi H274Y, E119V, N294S i R292K, substituii care s-a dovedit a
conferi virusurilor gripale rezisten la NAI. Mutaia H274Y s-a estimat ca cea mai frecvent i
pare a fi exclusiv limitat la gena N1 att n virusurile gripale sezoniere, ct i n cele pandemice
n populaia uman i pot avea loc spontan fr presiunea aparent a antiviralelor sau reasortare
[87, 95-100]. Totui, un numr foarte mic de mutaii N294S au fost detectate n virusurile
A(H1N1)pdm, mai general, ns, toate cele patru mutaii s-a stabilit c exist n gena N1. n plus
la aceasta, dei virusurile gripale A(H3N2) sunt dominate de substituiile E119V i R292K,
substituia H274Y nu a fost niciodat identificat n gena N2 [87, 97]. S-a stabilit c o nou
substituie I223R cauzeaz o rezisten modest la NAI, ns, n combinaie cu substituia H274Y
rezistena la oseltamivir crete semnificativ, pe cnd rezistena la zanamivir rmne la nivel
sczut. O alt combinaie a dou mutaii n gena NA, Q313R i I427T, de asemenea cauzeaz
rezisten la ambele remedii NAI: oseltamivir i zanamivir [101, 102-104].
Evoluia rezistenei la oseltamivir n virusurile gripale A(H1N1) pandemice se poate
datora mutaiilor punctiforme n orice regiune a genelor NA sau evenimentului de reasortare.
Rezistena la oseltamivir n tulpinile de virusuri gripale A(H1N1) pandemice pot aprea in
diverse forme: o evoluie sporadic la un pacient infectat drept rspuns la tratament; evoluia
rezistenei la oseltamivir la un pacient infectat i transmisia tulpinii date contacilor; meninerea
genotipului ce confer rezisten la oseltamivir ntr-o linie viral datorit presiunii de selecie
i/sau evenimentului de reasortare dintre tulpinile A(H1N1) sezoniere rezistente la oseltamivir i
tulpinile A(H1N1) pandemice. Acest eveniment poate oferi un se









![GENETICA, BIOLOGIA MOLECULARĂ ŞI AMELIORAREA · neuro-vegetative, insomnii, afecţiuni renale, boli reumatice, dischinezii biliare, stări gripale, răceli [4, 24]. Cercetările](https://static.fdocumente.com/doc/165x107/5e61e6dc1ad28b461e332f25/genetica-biologia-molecular-i-ameliorarea-neuro-vegetative-insomnii-afeciuni.jpg)