Membrana celulara
10
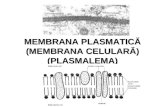
Membrana celulara Celula este separate de mediul extracellular prin formatiuni inalt differentiate care au rolul essential de a mentine in limite constante structura sa complexa. Structurile de la periferia celulei asigura in mod selective si controlat schimbul de substante dintre mediul intra si extracellular. Termenul de system de membrane celulare include in principal doua categorii de formatiuni membranare, si anume: membrane celulara externa sau plasmalema, dispusa la periferia celulei, si endomembranele celulare, ce intra in constitutia majoritatii organitelor citoplasmatice. Invelisul cellular Este alcatuit din formatiuni complexe intre care se stabilesc relatii structurale si functioanale. Componentele invelisului sunt in numar de 3. Plasmalema are o compozitie moleculara lipidica si proteica complexa. Glicocalixul se prezinta ca un invelis de natura glucidica, glicoproteica si glicolipidica, la suprafata externa a plasmalemei. Corticala celulara e reprezentata de o zona cu grosimea de 1-2mm din citoplasma celulara din imediata vecinatate a fetei citoplasmatice a plasmalemei. Invelisul cellular prezinta la suprafata celulei o serie de diferentieri cu rol important in realizarea adeziunii intercelulare- jonctiuni celulare, in marirea suprafetei celulare- microvili si in motilitatea celulara cili, flageli, pseudopode.
-
Upload
veronica99 -
Category
Documents
-
view
45 -
download
3
description
membrana celulara biochimie
Transcript of Membrana celulara
Membrana celulara
Membrana celulara
Celula este separate de mediul extracellular prin formatiuni inalt differentiate care au rolul essential de a mentine in limite constante structura sa complexa. Structurile de la periferia celulei asigura in mod selective si controlat schimbul de substante dintre mediul intra si extracellular.
Termenul de system de membrane celulare include in principal doua categorii de formatiuni membranare, si anume: membrane celulara externa sau plasmalema, dispusa la periferia celulei, si endomembranele celulare, ce intra in constitutia majoritatii organitelor citoplasmatice.
Invelisul cellular
Este alcatuit din formatiuni complexe intre care se stabilesc relatii structurale si functioanale.
Componentele invelisului sunt in numar de 3.
Plasmalema are o compozitie moleculara lipidica si proteica complexa.
Glicocalixul se prezinta ca un invelis de natura glucidica, glicoproteica si glicolipidica, la suprafata externa a plasmalemei.
Corticala celulara e reprezentata de o zona cu grosimea de 1-2mm din citoplasma celulara din imediata vecinatate a fetei citoplasmatice a plasmalemei.
Invelisul cellular prezinta la suprafata celulei o serie de diferentieri cu rol important in realizarea adeziunii intercelulare- jonctiuni celulare, in marirea suprafetei celulare- microvili si in motilitatea celulara cili, flageli, pseudopode.
Functiile invelisului cellular sunt in functie de structura moleculara si proprietatile sale.
Functia de bariera fizica. Prin aceasta se mentine structurarea mediului intracellular, la un inalt nivel de organizare, stare care se pastreaza in anumite limite. Membranele intracelulare realizeaza compartimentarea celulei asigurand desfasurarea anumitor functii.
Functia de transport. Plasmalema asigura desfasurarea schimburilor de energie si materie inter cele doua medii pe care le separa.
Functia de a receptiona informatii. Plasmalema contine receptori capabili de a capta foarte specific mesageri extracelulari.
Functia de a asigura adeziunea intercelulara, prin care invelisul participa la realizarea conexiunilor dintre celulele aflate in tesut.
Functia de a asigura apararea celulei. Se realizeaza impotriva unor factori care interactioneaza agresiv cu celula, plasmalema fiind implicate in imunitatea celulara a organismului.
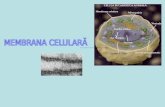
Structura moleculara a plasmalemei
Componentele chimice care intra in structura plasmalemei sunt lipidele si proteinele.
Tipuri de lipide membranare Principalele tipuri de lipide care intra in constitutia membranei sunt: fosfolipidele,colesterolul, glicolipide. Toate acestea au moleculele amfipatice sau amfifile, adica au un capat hidrofil si un capat hidrofob. Se formeaza un bistrat lipidic.
Fosfolipide membranare
1.Fosfolipide fosfogliceridele si sfingolipidele.
Fosfogliceridele au la baza molecula de glycerol. Lanturile de acizi grasi utilizate ptr esterificarea a doua grupari hidroxil alcatuiesc portiunea hodrofoba a moleculei.COmponenta polara, hidrofila reprezentata de a treia grupare hidroxil a glicerolului.
Sfingolipidele au aceeasi structura spatiala cu a fosfogliceridelor, avand o grupare polara hidrofila si doua lanturi hidrocarbonate hidrofobe. Cel mai raspandit sfingolipid este sfingomielina
2. Glicolipide membranare- au o structura moleculara asemanatoare cu a sfingolipidelor deosebindu-se de acestea prin faptul ca in locul gruparii polare fosforilcolinice au radicali glucidici, care sunt orientate inspre fata externa a plasmalemei.
Exista glicolipide neutre, care au radicali glucidici neutri, de ex. Galactocerebrozida ce este componenta majora a mielinei, ce formeaza teaca axonilor neuronali, si gangliozide care sunt cele mai complexe glicolipide. Contin pe langa radicalii glucidici neutri si acid sialic.
3.Colesterolul membranarMembrana celulara la eucariote contine cantitati insemnate de cholesterol.Molecula de cholesterol este amfifila, capatul hidrofil polar fiind reprezentat de gruparea hidroxil a inelului sterolic, iar capatul hidrofob e reprezentat de lantul hidrocarbonat atasat la acest inel.
Pentru ca in structura bistratului lipidic membranar se insera intre molelculele fosfolipidice, colesterolul influenteaza fluiditatea membranei. Colesterolul impiedica miscarea libera a portiunilor externe a lanturilor de acizi grasi din bistrat, dar In acelasi timp desparte si indeparteaza regiunile interne ale acestora ceea ce face ca zona interna a bistratului sa devina mai fluida.
Colesterolul scade permeabilitatea bistratului la molecule mici, hidrofile si creste flexibilitatea si stabilitatea fizica a acestuia.
Bistratul fosfolipidic
Deoarece moleculele lipidice membranare au proprietatea de a fi amfifile ele se dispun in structura membranei celulare intr-un dublu strat, cu capetele hidrofile la exterior, si cu cele hidrofobe la interior.Confera bistratului proprietati de autoasamblare, fluiditate, asimetria.Autoasamblarea bistratului fosfolipidic
Moleculele lipidice ( caracter amfifil ) au tendinta de a forma bistraturi in mediul apos.
Fosfolipidele si glicolipidele in mediul apos tind sa-si orinteze capatul hidrofob in asa fel incat sa nu vina in contact cu apa.
Prin agitarea acestor molecule de apa se formeaza micele si bistraturi.Daca se formeaza bistraturi au tendinta de a forma vezicule ce inchid in interior compartimente apoase, astfel se pot sintetiza pe cale artificiala doua tipuri de bistraturi lipidice: formeaza vezicule sferice liposomi, respectiv membrane negre plane, formate pe suprafata unui mic orificiu in asa fel incat sa separe 2 medii apoase.Fluiditatea bistratului fosfolipidicMiscarea moleculelor lipidice in bistrat este de trei tipuri:
- rotarea in jurul axului molecular
- difuziune in planul stratului in care se afla - trecerea dintr-un strat in altul ( miscarea flip flop )
Factorii care determina gradul de fluiditate al membranei celulare sunt gradul de saturare al acizilor grasi si lungimea lantului lor de carbon, temperatura (temperatura de tranzitie prin care structura de cristal lichid a bistratului trece intr-o structura cristalina de gel) si proprietatile particulare ale moleculelor de colesterol.Asimetria bistratului fosfolipidicEste determinata de compozitia lipidica diferita a celor 2 jumatati ale bistratului care este urmatoarea:1. in stratul extern
lipide cu grupare colinica
- toate glicolipidele
- lipidele cu acizi grasi nesaturati.
2. In stratul intern
- lipidele cu sarcina electrica ( fosfatidilserina negativa, fosfatidiletanolamina )
- lipidele cu acizi grasi nesaturati.
Ptr ca moleculele lipidice nu pot executa miscari flip-flop se considera ca asimetria bistratului este determinata din momentul biosintezei intracelulare a membranei, proces ce are loc la nivelul reticulului endoplasmic. Acest proces e asigura de translocatori fosfolipidici, care determina miscarea flip-flop a fosfolipidelor in cursul sintezei membranelor, proces ce sta la baza asimetriei.
Asimetria are o importanta functionala deosebita deoarece determina orientarea proteinelor membranare. O importanta deosebita in determinarea asimetriei bistratului o au glicolipidele.
Proteinele membranare
Majoritatea functiilor specifice ale membranei sunt indeplinite de proteine.
Tipuri de proteine: Unele proteine membranare traverseaza bistratul fosfolipidic. La fel ca moleculele fosfolipidice aceste proteine numite transmembranare sunt amfipatice, au regiuni hidrofobe care trec prin membrana.
Caracterul hidrofob al proteinelor transmembranare poate fi crescut prin atasarea la molecula a unui lant de acid gras care este inserat in monostratul lipidic de pe fata citoplasmatica.
Unele proteine membranare se asociaza cu bistratul prin intermediul unor lanturi hidrocarbonate. Aceste proteine legate de lipide sunt grupare in 3 clase:
a. Proteine ancorate la glicozil-fosfatidilinozitol sunt caracteristice pentru fata externa a membranei
b. Proteinele ancorate la miristat si proteinele ancoratte la farnezil ce se leaga intotdeauna pe fata citoplasmatica a membranei . Aceste proteine intra in categoria proteinelor transformante deoarece sunt implicate mai mult sau mai putin direct in procesele de transformare celulara si diviziune celulara.
Alte proteine membranare un patrund in interiorul hidrofob al bistratului, ci sunt legate pe o fata sau alta a membranei prin interactiuni necovalente cu proteinele transmembranare.
Proprietatile specifice ale proteinelor transmembranare
In cazul proteinelor transmembranare, zona din lantul polipeptidic scufundat in regiunea hidrofoba a bistratului, este alcatuita din aminoacizi cu radicali nepolari. Legaturile peptidice fiind insa polare si neexistand apa in acest mediu, se vor forma legaturi de hidrogen intre legaturile peptidice una cu cealalta. Formarea puntilor de hidrogen dintre legaturile peptidice este favorizata atunci cand proteina patrunde in bistrat sub forma de alfa-helix. Astfel, toate proteinele transmembranare traverseaza bistratul sub forma de lanturi alfa.
Ele au o orientare unica in membrana. Acest lucru se datoreaza mecanismului asimetric dupa care sunt sintetizate si inserate in bistrat la nivelul reticulului endoplasmatic, cat si functiile diferite din domeniul citoplasmatic respectiv extracelular.Majoritatea proteinelor transmembranare sunt glicozilate. La fel ca in cazul glicolipidelor, lanturile de oligozaharide sunt despartite pe fata extracelulara a membranei.Porinele sunt o clasa de proteine transmembranare diferite ca structura a radicalului fata de celelalte. Membrana lor externa asigura protectia impotriva agentilor daunatori, precum antibioticele, sarurile biliare sau proteazele intestinale, dar permit captarea sau eliminarea unor molecule mici hidrofile ( ex. Substante nutritive sau produsi de metabolism ).
Reconstituirea experimentala a functiilor proteinelor transmembranareProteinele pot fi solubizate doar cu agenti care distrug legaturi hidrofobe de la nivelul bistratului. Cei mai utilizati agenti => detergentii.
In contact cu membrana, capetele hidrofobe ale moleculei de detergent se asociaza cu capup hidrofob al proteinelor transmembranare, eliminand astfel moleculele lipidice din substrat.
Capatul polar al moleculelor de detrgent poate fi incarcat electric cum este cazul SDS, sau neutru tip Tritton.Detergentul SDS denatureaza moleculele proteice deoarece se poate lega de miezul hidrofob ceea ce le face inactive si neutralizabile pt cercetari functionale. Totusi proteinele pot fi purificate usor in aceasta forma denaturata si pot fi renaturate la forma lor functional activa prin indepartarea detergentului.
Mobilitatea proteinelor membranare
Proteinele pot prezenta miscari de rotatie in jurul unui ax perpendcular pe planul membranei, si miscari de difuziune laterala in planul membranei.
Difuziunea laterala a proteinelor in planul membranei este foarte importanta ptr exercitarea functiilor acestor molecule.
Tehnicile de fuziune celulara si de marcare cu anticorpi au permis evidentierea directa a mobilitatii proteinelor membranare.
In functie de tipul de celula sau de tipul proteinei se considera ca o proportie de 30-90% din totalitatea proteinelor membranare difuzeaza lateral in planul membranei.
Modelul de mozaic fluid al membranei
Acesta a fost descris detaliat de Singer si Nicolson.
Postuleaza ca:
- lipidele si proteinele transmemembranare sunt dispuse in mozaic
- membrana are o structura semifluida, atat lipidele cat si proteinele executand miscari de translatie in planul bistratului lipidic.
- Fluiditatea implica existenta intre componentele moleculare, a interactiunilor fizico-chimice slabe, necovalente
- Toate moleculele din structura de baza a membranei celulare sunt amfifite.
- Fluiditatea mozaicului este posibila doar in conditii de temperatura superioara temperaturii de tranzitie din faza de gel in faza de cristal lichid, conditie realizata la nivelul temperaturii fiziologice.
- Bistratul lipidic are o structura bidimensionala, fapt ce decurge din despozitia asimetrica a moeleculei in membrana
Astfel membrana celulara poate fi imaginata ca o mare lipidca in care plutesc niste iceberguri proteice.Structura microvililorLa structurarea microvililor participa mai multe componente ale membranei plasmatice si ale citoscheletului. Proteinele din structura lor le determina aspectul si diametrul uniform. In axul fiecarui microvil se afla un manunchi de filamente de actina care se ancoreaza la o extremitate de proteinele de pe fata citoplasmatica a membranei microvilare, iar la baza se intersecteaza cu o retea de filamente ce contin proteine de legare a actinei.
Microfilamentele asigura rigiditatea microvililor si pot determina miscarea lor inainte si inapoi.
Reteaua filamentoasa situata in celula aproape de baza microvililor se insera pe fata citoplasmatica a membranei laterale la un tip special de jonctiuni celulare.