Anatomie
104
OBIECTIVELE DISCIPLINEI Disciplina contribuie la formarea şi unor competenţe generale şi specifice ale absolventului domeniului de kinetoterapie şi motricitate specială. Prin parcurgerea programului educaţional prevăzut la Disciplina Anatomie II, studentul va fi capabil să: 1. identifice elementele structurale ale componentelor anatomice studiate prin observaţie şi palpare la subiectul de analizat; 2. utilizeze corect terminologia pentru descrierea componentelor sistemului nervos central şi perifric, precum şi ale organelor interne studiate; 3. localizeze corect principalele zone topografice ale corpului; 4. localizeze corect principalele organe interne corespunzătoare zonelor topografice; 5. diferenţieze componentele viscerale ale organismului, rolul şi importanţa lor în cadrul conceptului de mişcare biologică. 6.diferenţieze conformaţia anatomică normală, de conformaţia anatomică patologică, precizând elementele anatomice afectate; COMPETENŢE ASIGURATE PRIN PARCURGEREA DISCIPLINEI Disciplina contribuie la formarea competenţelor praxiologice în legătură cu structura sistemului nervos, aparatelor şi a competenţelor specifice în legătură cu: observarea conformaţiei anatomice şi a semnelor diverselor categorii de indivizi şi sesizarea abnormalităţilor. FOND DE TIMP ALOCAT, FORME DE ACTIVITATE, FORME DE VERIFICARE, CREDITE Forma de activitate Număr ore semestru Număr credite Seminar 24 Studiu individual 41 Verificare finală Examen Validare total credite: 5
-
Upload
marius-ilisei -
Category
Documents
-
view
117 -
download
11
Transcript of Anatomie

OBIECTIVELE DISCIPLINEI
Disciplina contribuie la formarea şi unor competenţe generale şi specifice ale
absolventului domeniului de kinetoterapie şi motricitate specială. Prin parcurgerea programului
educaţional prevăzut la Disciplina Anatomie II, studentul va fi capabil să:
1. identifice elementele structurale ale componentelor anatomice studiate prin
observaţie şi palpare la subiectul de analizat;
2. utilizeze corect terminologia pentru descrierea componentelor sistemului nervos
central şi perifric, precum şi ale organelor interne studiate;
3. localizeze corect principalele zone topografice ale corpului;
4. localizeze corect principalele organe interne corespunzătoare zonelor topografice;
5. diferenţieze componentele viscerale ale organismului, rolul şi importanţa lor în
cadrul conceptului de mişcare biologică.
6.diferenţieze conformaţia anatomică normală, de conformaţia anatomică patologică,
precizând elementele anatomice afectate;
COMPETENŢE ASIGURATE PRIN PARCURGEREA DISCIPLINEI
Disciplina contribuie la formarea competenţelor praxiologice în legătură cu structura
sistemului nervos, aparatelor şi a competenţelor specifice în legătură cu: observarea
conformaţiei anatomice şi a semnelor diverselor categorii de indivizi şi sesizarea
abnormalităţilor.
FOND DE TIMP ALOCAT, FORME DE ACTIVITATE,
FORME DE VERIFICARE, CREDITE
Forma de activitate Număr ore semestru Număr credite Seminar 24
Studiu individual 41
Verificare finală Examen Validare total credite: 5

STABILIREA NOTEI FINALE
Forma de verificare (Examen, Colocviu, Verificare pe parcurs)
Modalitatea de susţinere (Scris şi Oral, Oral, Test grilă, etc.) Puncte sau procentaj
Răspunsuri la examen / colocviu 5 (50%)
Verificări practice / Evaluarea activităţii la seminar 4 (40%)
Verificări pe parcursul semestrului 0,5 (10%)
Teme de control 0,5 (10%)
Alte cerinte NO
TA
RE
TOTAL PUNCTE SAU PROCENTE 10 (100%)
TIMP MEDIU NECESAR PENTRU ASIMILAREA FIECĂRUI MODUL
Timp mediu
necesar Nr.
Crt. Denumire modul
SI S Total
1 Sistemul nervos de relaţie - generalităţi. Anatomia măduvei
spinării. Configuraţie externă. 2 -
2 Structura măduvei spinării. Substanţa cenuşie. Substanţa albă.
Substanţa reticulată. 2 -
3. Anatomia bulbului rahidian. 2 -
4. Anatomia punţii. 2 -
5. Anatomia cerebelului. 2 -
6. Anatomia pedunculilor cerebrali. Anatomia diencefalului. 2 -
7. Anatomia emisferelor cerebrale. 2 -
8. Analizatorii. 4 -
9. Aparatul respirator. 2
10. Aparatul cardiovascular. Inima (configuraţie externă şi structură). 2
11. Principalele vase de sânge şi limfatice. 2
12. Aparatul digestiv. 2

13. Glandele anexe ale tubului digestiv. 2
Timp total necesar 28 28
INSTRUCŢIUNI PENTRU PARCURGEREA RESURSEI DE
INVĂTĂMÂNT
Prezenta resursă de învăţare conţine toate informaţiile necesare însuşirii cunoştinţelor teoretice referitoare
la anatomia umana.
Conţinutul este structurat în module, în cadrul fiecărui modul regăsindu-se unul sau mai multe unităţi de
studiu, în aşa fel încât să eşaloneze şi să faciliteze parcurgerea materialului şi însuşirea sa.
Pentru parcurgerea resursei de învăţământ se recomandă următoarea succesiune :
1. Citirea, cu atenţie, a unităţii de studiu şi, consultarea recomandărilor bibliografice în legătură cu aceasta.
2. Parcurgerea rezumatului unităţii de studiu.
3. Rezolvarea testului de autocontrol.
4. Pregătirea schemei de discuţie pentru seminar.

ANATOMIE
Lector. univ. drd. Marinela Raţă
Asist. Univ. drd. Şlicaru Adina

Cuprins
Unitatea de curs 1 Sistemul nervos de relaţie
Unitatea de curs 2 Sistemul nervos central. Anatomia măduvei spinării
Unitatea de curs 3 Encefalul. Anatomia trunchiului cerebral
Unitatea de curs 4 Anatomia cerebelului
Unitatea de curs 5 Anatomia diencefalului.
Unitatea de curs 6 Anatomia emisferelor cerebrale.
Unitatea de curs 7 Sistemul nervos periferic.
Unitatea de curs 8 Sistemul nervos vegetativ.
Unitatea de curs 9 Analizatorii
Unitatea de curs 10 Aparatul respirator
Unitatea de curs 11 Aparatul cardiovascular. Inima �i principalele vase de sânge şi limfatice.
Unitatea de curs 12 Aparatul digestiv. Glandele anexe ale tubului digestiv.
Unitatea de curs 13 Sistemul endocrin

Unitatea de curs 1 Sistemul nervos de relaţie
Conţinutul unităţii de studiu
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului sistemului nervos al organismului uman.
Obiectivele operaţionale
1. Prezentarea clasificării sistemului nervos
2. Prezentarea caracteristicilor dezvoltării sistemului nervos
3. Prezentarea rolului sistemului nervos
4. Prezentarea caracteristicilor nervilor şi al grupări lor
Sistemul nervos
Sistemul nervos reprezintă totalitatea organelor formate predominant din ţesut nervos
ce recepţionează, transmite şi integrează informaţiile primite din mediul extern sau intern şi
permite elaborarea unui răspuns adecvat mesajului primit. Recepţionarea mesajelor se
realizează prin intermediul receptorilor, specializaţi în transformarea stimulilot - ce poartă
diferite forme de energie - în impuls nervos, ce reprezintă energia internă specifică
organismului.
De la receptori, influxul nervos este transmis pe căile nervoase aferente (care aduc) spre
centrii nervoşi de integrare de la nivelul diferitelor etaje de la nivelul sistemului nervos central.
Integrarea nervoasă reprezintă prelucrarea de către centrii nervoşi, a mesajelor primite
şi elaborarea comenzilor pentru organele efectoare sau, aceste informaţii sunt stocate ca "acte
de memorie" (formarea de engrame) şi reactualizate ulterior. Răspunsurile care pleacă de la
centrii sunt conduse pe căile nervoase eferente (care duc) spre efectori, cărora le va determina
intrarea în activitate (contracţia muşchilor scheletici, contracţia musculaturii viscerale,
activitatea gleandelor cu secreţie internă şi externă).
Sistemul nervos este împărţit în:
- sistemul nervos al vieţii de relaţie sau somatic ce stabileşte legătura între
organism şi mediul extern;

- sistemul nervos al vieţii vegetative ce coordonează activitatea organelor
interne, în strânsă legătură cu sistemul nervos somatic şi cu sistemul endocrin (hormonii
amplifică, generalizează şi prelungesc în timp, reacţiile iniţiate de sistemul nervos vegetativ).
Sistemul nervos somatic este format din:
- sistemul nervos central (S.N.C.) sau axul cerebro-spinal sau nevrax,
reprezentat de encefal şi măduva spinării;
- sistemul nervos periferic (S.N.P.) ce cuprinde ganglionii nervoşi, nervii spinali
şi nervii cranieni.
Sistemul nervos vegetativ este format din:
- sistemul nervos central cu centrii nervoşi vegetativi situaţi în axul
cerebrospinal;
- sistemul nervos periferic cu ganglionii nervoşi vegetativi şi fibrelel nervoase
vegetative preganglionare şi postganglionare care intră în componenţa nervilor spinali şi
cranieni.
Dezvoltarea sistemului nervos
Dezvoltarea filogenetică a sistemului nervos
Sistemul nervos apare la celenterate ca o reţea nervoasă în care celule sunt cu
prelungirile în continuitate. Sinapsele apar la artropode la care sistemul nervos are aspectul
unor mase gelatinoase ce pot realiza reflexe simple. La vertebrate schema de organizare devine
mai complexă. Măduvei spinării care este centrul reflexelor simple segmentare, i se suprapun
centrii superiori de integrare. Treptat, apare cerebelul, creierul tectal (tuberculii cvadrigemeni),
creierul bazal (nucleii bazali), iar la mamifere şi om se dezvoltă preponderent creierul cortical,
care se suprapune la rândul său prin funcţii complexe şi coordonează în ultimă instanţă toată
activitatea lui. Scoarţa cerebrală împreună cu centrii subcorticali transformă omul într-un
animal sociabil şi constituie substratul material al personalităţii umane.
Dezvoltarea ontogenetică a sistemului nervos
Sistemul nervos (SN) se dezvoltă din ectodermul feţei dorsale (placa neurală) a discului
embrionar, care se invaginează şi formează şanţul neural, mărginit de două buze, denumite
creste neurale. Acesta se adânceşte şi se separă de ectoderm devenind tub neural. În pereţii
tubului neural prin multiplicarea celulelor se formează trei straturi: intern, este zona

germinativă cu celule în mitoză. Acestea vor genera neuroblaşti şi spongioblaşti care formează
zona de manta. Neuroblaştii devin neuroni, iar spongioblaşti celule gliale. În stratul extern sau
zona marginală, prelungirile neuroblaştilor dau naştere substanţei albe a SNC. Din lumenul
tubului neural la nivelul măduvei se formează canalul central (ependimar), iar la nivelul
encefalului apar ventriculii cerebrali. Prin îngroşarea pereţilor tubului neural, apar: lama
bazală, anterior; lama dorsală, posterior; lamele fundamentale, antero-lateral şi lamele aleare,
postero-lateral.
Măduva spinării ia naştere din partea mijlocie şi terminală a tubului neural. Fibrele
nervoase ale substanţei albe a măduvei sunt iniţial fără teacă de mielină. Aceasta apare în luna
a IV-a intrauterină la nivelul cordoanelor posterioare senzitive. Fibrele corticospinale încep să
se mielinizeze după naştere, fapt ce explică posibilitatea apariţiei mersului mai târziu după
naştere. Encefalul se dezvoltă din trei vezicule cerecbrale denumite prozencefal, mezencefal şi
rombencefal. Prima şi ultima se mai segmentează ulterior şi astfel se ajunge la stadiu de 5
vezicule cerebrale. Din prozencefal iau naştere telencefalul şi diencefalul. Mezencefalul
rămâne nesegmentat, iar prin segmentarea rombencefalului iau naştere metencefalul şi
mielencefalul.
Din mielencefal se dezvoltă bulbul; din metencefal puntea şi cerebelul; din mezencefal
pedunculii cerebrali şi tuberculii cvadrigemeni; din diencefal, talamusul, hipotalamusul cu
lobul posterior al hipofizei, subtalamusul, metatalamusul şi epitalamusul; din telencefal
emisferele cerebrale şi ganglionii bazali.
Datorită creşterii mai mari în lungime a veziculelor cerebrale decât a cutiei craniene
apar o serie de curburi (a vertexului, pentină şi nucală), iar emisferele ajung să acopere
celelelalte formaţiuni şi suprafaţa lor se cutează, adaptându-se la volumul cutiei craniene.
Sistemul nervos periferic şi cel vegetativ iau naştere din crestele ganglionare ce se
dezvoltă din crestele neurale. După formarea tubului neural crestele ganglionare se separă de
el, aşezându-se între acesta şi somite. Din crestele ganglionare se dezvoltă ganglionii spinali şi
ai nervilor cranieni; tot din celule migrate de aici se vor forma ganglionii vagetativi ai lanţului
simpatic, ai plexurilor prevertebrale şi ai plexurilor nervoase intramurale ale organelor cavitare.

Gruparea neuronilor în sistemul nervos
Pentru o mai bună înţelegere a unor termenilor specifici ai sistemului nervos este
necesară precizarea unor noţiuni ca: nuclei nervoşi, substanţă cenuşie, centrii nervoşi, gangioni
nervoşi, substanţa albă, nervi, plexul nervos.
Nuclei nervoşi sunt grupări de corpi neuronali, localizaţi în sistemul nervos central,
unde formează (împreună cu fibrele amielinice) substanţa cenuşie; noţiunea de centrii nervoşi
se referă la aspectele funcţionale ale nucleilor nervoşi.
Ganglionii nervoşi sunt grupările de corpuri neuronale situate în sistemul nervos
periferic; ei se împart în ganglioni senzitivi (spinali şi cranieni) şi ganglioni vegetativi.
Fasciculele sau căile nervoase sunt grupări de fibre nervoase dispuse în interiorul
sistemului nervos central (intranervraxial) unde formează substanţa albă.; în masa lor nu există
ţesut conjunctiv.
Nervii reprezintă gruparea fibrelor nervoase în sistemul nervos periferic
(extranevraxial) şi sunt înveliţi în teci conjunctive. Unii nervi conţin şi fibre mielinice şi
amielinice. În unele regiuni ale corpului ramificaţiile nervului se împletesc între ele şi
formează plexuri nervoase, ce conţin şi celule nervoase.
Din punct de vedere fiziologic nervii pot fi: senzitivi, motori sau micşti. Nervii senzitivi
sunt formaţi din fibre nervoase senzitive, reprezentate de dendritele neuronilor senzitivi din
ganglionii spinali sau cranieni; ei primesc excitaţiile de la receptori şi le transmit la sistemul
nervos central. Nervii motori conţin fibre nervoase motoare reprezentate de axonii neuronilor
motori din nucleii de substanţă cenuşie ai axului cerebrospinal; terminaţiile lor se distribuie la
diverse organe efectoare. Nervii micşti conţin fibre senzitive şi fibre motoare.

Unitatea de curs 2 Sistemul nervos central. Măduva spinării
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului măduvei spinării.
Obiectivele operaţionale
1. Prezentarea configuraţiei externe a măduvei spinării
2. Prezentarea structurii interne a măduvei spinării
3. Prezentarea căilor de conducere medulare
Măduva spinării
Sistemul nervos central este format din măduva spinării şi encefal care
alcătuiesc la un loc sistemul nervos cerebrospinal sau nevrax.
Configuraţia externă. Măduva spinării este partea SNC adăpostită în canalul vertebral
format din suprapunerea găurilor vertebrale, completat între corpurile vertebrale de discurile
intervertebrale, iar între arcuri de ligamentele galbene.
Se întinde de la nivelul arcului anterior al atlasului unde iese primul nerv cervical până
în dreptul discului L1-L2. La embrionul de 3 luni umple tot canalul vertebral până la coccis, dar
ulterior creşte mai puţin decât coloana vertebrală, realizându-se o ascensiune aparentă. La
bărbat are o lungime de 45 cm, iar la femei de 43 cm.

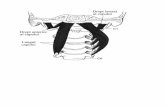
Fig. 1 Configuraţia externă a măduvei spinării
Măduva spinării este un
cordon cilindric antero-
posterior cu diametrul de 12 / 9
cm. Ea prezintă două regiuni
mai umflate: una cervicală
(intumiscentia cervicala) în
dreptul vertebrelor C3-T2 şi alta
lombară (intumiscentia
lumbalis) în dreptul vertebrelor
T9-T12.
Aceste umflături sunt
locul de unde pleacă nervii
pentru membre şi dezvoltarea
lor este condiţionată de
funcţionalitatea membrelor
respective. De exemplu,
cangurul nu are intumiscenţă
cervicală, foca pe cea lombară,
şarpele nu are nici o
intumiscenţă, iar la amputaţi
sau la cei cu lipsă congenitală a
unui membru dispare sau scade
intumiscenţa respectivă (fig. 1).
ajungă la gaura de conjugare respectivă,aşa că sub L1 vom În partea inferioară, măduva se
termină cu conul medular, care se prelungeşte până la coccis cu filum terminale din substanţa
albă. Rădăcinile nervilor spinali, datorită ascensiunii aparente a măduvei, vor coborî din ce în
ce mai oblic ca să găsi doar rădăcini ale nervilor lombari şi sacrali alături de firul terminal,
formând ceea ce se numeşte coada de cal.
Măduva spinării se împarte în mai multe segmente (miolomere) şi din fiecare iese câte
un nerv spinal. Sunt 8 perechi de nervi spinali cervicali, 12 perechi de nervi toracali, 5 perechi
de nervi lombari, 5 perechi de nervi sacrali şi 1 pereche de nervi coccigieni (fig. 19). Pe

suprafaţa măduvei se văd mai multe şanţuri longitudinale a căror denumire arată şi situaţia
lor: fisura mediană anterioară, şanţul median posterior, şanţul lateral anterior pe unde ies
rădăcinile anterioare ale nervilor spinali şi şanţul lateral posterior pe unde intră rădăcinile lor
posterioare. La nivelul măduvei cervicale se descrie şi un şanţ intermediar posterior ce
corespunde limitei între fasciculele Goll şi Burdach.
Structura internă. Pe secţiune transversală se constată că măduva spinării este alcătuită
din substanţa cenuşie situată central şi care pe secţiune are forma literei "H” şi subtanţa albă,
aşezată la periferie. Datorită fisurii anterioare şi a şanţului median posterior, care se
prelungeşte în interior cu un sept median posterior, măduva apare împărţită în două jumătăţi
legate între ele prin două comisuri: una cenuşie posterioară, formată de bara transversală a
"H”-ului şi alta albă anterioară, între fundul fisurii mediene anterioare şi comisura cenuşie.
- Substanţa cenuşie (fig. 2). Prezintă pe suprafaţa sa de secţiune două prelungiri
anterioare, numite coarne anterioare, care în lungul măduvei realizează coloanele anterioare.
Ele sunt mai umflate şi nu ajung până la suprafaţa măduvei. Prelungirile posterioare se numesc
coarne posterioare şi formează în lungul măduvei coloanele posterioare. Ele sunt mai subţiri
şi ajung până la şanţul colateral posterior de care sunt separate printr-o lamă de substanţă albă
numită zona marginală (Lissauer). Coarnele posterioare au un apex, un cap, un col şi o bază.
Coarnele laterale formează în lungul măduvei columna laterală întinsă între mielomerele C8-
L2. În grosimea comisurii posterioare este situat canalul central. Substanţa cenuşie este
formată mai ales din corpul neuronilor. Datorită legii neurobiotaxiei care afirmă că toţi
neuronii cu aceeaşi funcţie stau grupaţi la un loc, în substanţa cenuşie se vor descrie nuclei.
După funcţia neuronilor, în substanţa cenuşie se descriu patru zone:
- Zona somatomotorie, în cornul anterior cu nucleii motori pentru musculatura striată
scheletică. Se numesc şi neuroni radiculari deoarece axonul lor intră în rădăcina anterioară a
nervului spinal. Neuronii radiculari sunt de 2 tipuri: alfa care inervează fibrele contractile ale
muşchilor şi gama care sunt fusiformi (inervaţia fusurilor neuromusculare). Neuronii motori au
şi rol trofic asupra fibrelor musculare.
- Zona visceromotorie la baza conului anterior şi în cornul lateral, conţine nuclei
vegetativi. Între C8 şi L2 este nucleul intermedio-lateral în care sunt centrii simpatici pentru
tot

organismul începând cu nucleul ciliospinal (C8-T3) care produce midriaza (dilatarea pupilei) şi
terminând cu nucleul veziculo-spinal la L1 şi anorectul la L2.În măduva sacrată sunt nucleii
parasimpatici ai micţiunii, defecaţiei şi erecţiei. Axonii acestor neuroni trec prin cornul
anterior, rădăcina anterioară a nervului spinal şi din nervul spinal ajung prin ramul comunicant
alb la lanţul ganglionar simpatic laterovertebral.
- Zona viscerosenzitivă din partea posterioară a regiunii intermediare are neuronii care
recepţionează interoreceptivitatea şi care nu se grupează în nuclei. La ei vin fibrele nervoase
de la viscere, prin rădăcina posterioară a nervilor spinali.
- Zona somatoreceptivă din cornul posterior prezintă nucleul pericornual. În nucleul
pericornual, sub zona marginală (Lissauer), se găseşte stratul zonal Waldeyer, iar mai
profund, substanţa gelatinoasă Rolando. Acest nucleu are deutoneuronii pe calea
exteroceptivă. La baza cornului posterior, în partea medială se găseşte nucleul toracic (Clark-
Stilling), iar în partea laterală nucleul latero-bazilar (Bechterow). Aceşti doi nuclei
recepţionează stimulii proprioceptivi pentru coodonarea motorie fiind alcătuiţi din
deutoneuronii acestei căi. Pe lângă neuronii motori şi senzitivi, somatici şi vegetativi, se găsesc
şi numeroşi neuroni intercalari sau de asociere.
- Substanţa reticulată a măduvei este formată din grămezi de celule nervoase dispuse
“în reţea” în substanţa albă, între canalul posterior şi cel lateral, în vecinătatea imediată a
substanţei cenuşii.
-Substanţa albă este formată din 3 perechi de funicule sau cordoane. Cordonul anterior
între fisura mediană anterioară şi cornul anterior, cordonul lateral între cornul anterior şi septul
median dorsal. Cordoanele sunt alcătuite din fibre care pot fi grupate în: endogene care pleacă
de la neuronii din măduvă şi care pot fi scurte şi lungi. Cele scurte nu părăsesc măduva şi
alcătuiesc fasciculele de asociere intersegmentară a măduvei (fasciculele fundamentale). Cele
lungi sau de proiecţie ies din măduvă spre etajele superioare formând căi ascendente. Fibrele
exogene aparţin neuronilor din afara măduvei. Dacă provin de la neuronii ganglionului spinal
formează căi ascendente, pe când cele de centrii superiori, căi descendente.
A. Căile ascendente. Ele conduc sensibilitatea la etajele superioare. Primul neuron
(protoneuronul = N1) pe calea tuturor sensibilităţilor de la nivelul trunchiului şi membrelor este
neuronul pseudounipolar din ganglionul spinal de pe rădăcina posterioară a nervului spinal

aflat lângă gaura de conjugare unde cele două rădăcini se unesc spre a forma nervul. Pe căile
sensibilităţilor, deutoneuronul conduce mesajul la al III-lea neuron care cu excepţia
sensibilităţii olfactive se află în talmus, de unde stimulul ajunge la cortex în zona respectivă de
intergrare.
a) Căile sensibilităţii exteroceptive.
Dendrita protoneuronului se duce la periferie pe calea ramurilor nervului spinal de unde
culege stimulii de la exteroceptori (tactili, termici şi dureroşi), iar axonul pe calea rădăcinii
posterioare ajunge la nucleul pericornual. Axonul acestuia trece în jumătatea opusă a măduvei,
devine ascendent şi formează în cordonul anterior, tractul spinotalamic anterior, iar în cordonul
lateral tractul spinotalamic lateral şi spinorectal. Prin tractul spinotalamic anterior este condusă
sensibilitatea de presiune şi tactilă grosieră, afectivă, difuză sau protopatică pe când
sensibilitatea tactilă, fină gnostică sau epicritică este condusă prin fasciculele Goll şi Burdach.
Tractul spinotalamic lateral conduce termoalgezia. Fasciculul spinotectal conduce sensibilitatea
exteroceptivă la nivelul coliculilor optici din gama cvadrigemină.
b) Căile sensibilităţii proprioceptive.
Sunt diferite pentru stimulii ce deservesc coordonarea motorie de cei ce realizează
simţul kinestezic. Pentru sensibilitatea proprioceptivă kinestezică (aşa-zis "conştientă”, fig. 2.),
axonii protoneuronilor intră în măduvă, devin ascendenţi formând fasciculele Goll şi Burdach
din cordoanele posterioare şi merg să facă sinapsă cu al doilea neuron în nucleii omonimi din
bulb. De aici, pe calea lemnisculului medial mesajele ajung la talamus de unde sunt proiectate
pe cotex.
Pentru sensibilitatea proprioceptivă de coordonare motorie mesajele recepţionate de la
fusurile neuromusculare şi corpusculii tendinoşi Golgi, sunt trimise de protoneuron la nucelii
latero-bazilari şi toracici din măduvă. Axonii acestora fac două fascicule ascendente ce merg la
suprafaţa cordonului lateral. Tractul spinocerebelos posterior sau direct (Flechsig) pleacă din
nucleul toracic de aceeaşi parte, iar tractul spinocerebelor anterior încrucişat (Gowers) îşi are
originea în nucleul latero-bazilar din partea opusă.
c) Căile sensibilităţii interoceptive.
Ele nu au tracturi speciale la nivelul măduvei. Informaţiile culese de la viscere sunt conduse
prin fibrele tractului spinotalamic, dar mai ales din aproape în aproape cu multiple staţii în
substanţa reticulată prin căile ascendente nespecifice.

B. Căile descendente.
Aceste căi aduc comenzi de la etajele superioare pentru mişcări voluntare, automate,
reflexe de echilibru, ca şi pentru coordonarea lor. Cele pentru mişcări voluntare se numesc căi
piramidale, iar celelalte căi extrapiramidale.
Fig. 2 Căile de conducere ale măduvei spinării
a) Căile pentru mişcările
voluntare ale corpului şi ale
membrelor.
Pleacă din girusul precentral
din celulele giganto-piramidale
Betz. La măduvă, ele formează
tractul rticospinal anterior
(Türk) situat de o parte şi de alta a
fisurii mediene anterioare
(piramidal direct) şi tractul
corticospinal lateral (piramidal
încrucişat) din fasciculul lateral.
Fibrele tractului piramidal
încrucişat, au făcut decusaţia la
nivelul bulbului. Cele ale tractului
piramidal direct se încrucişează dar
pe parcurs şi merg şi ele să se
co
termine la nivelul neuronilor motori din coarnele anterioare de partea opusă.
b) Căile pentru mişcările automate. Sunt reprezentate de fasciculul tectospinal ce pleacă
de la coliculii cvadrigemeni superiori.c) Mişcările reflexe de echilibru.
Sunt comandate prin fasciculul vestibulospinal din cordonul anterior.
d) Alte căi extrapiramidale.
Sunt reprezentate de tractul rubrospinal de la nucleul roşu din mezencefal situat în
cordonul lateral; tractul olivospinal ce pleacă din oliva bulbară şi tractul reticulospinal. În
substanţa albă a măduvei spinării, pe lângă sistemele de fibre ascendente şi descendente sunt şi
fibre intersegmentare aparţinând aparatului elementar al măduvei spinării. Alături de

fasciculele fundamentale situate în fiecare cordon lângă substanţa cenuşie, mai se descriu cu
aceeaşi funcţie şi fasciculul triunghiular, septomarginal şi semiluna

Unitatea de curs 3 Encefalul. Anatomia trunchiului cerebral
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului măduvei spinării.
Obiectivele operaţionale
4. Prezentarea configuraţiei externe al trunchiului cerebral
5. Prezentarea structurii interne al trunchiului cerebral
6. Prezentarea căilor de conducere alr trunchiului cerebral
Encefalul reprezintă partea sistemului nervos central adăpostită în cutia craniană. Este
alcătuit dintr-o parte axilară denumită trunchi cerebral, înapoia căruia se găseşte cerebelul şi
din creier sau cerebrum situat superior de acesta.
Trunchiul cerebral.
Trunchiul cerebral este aşezat în etajul inferior al endobazei, pe clivus (faţa dorsală a
apofizei bazilare). Are aspectul unei fâşii longitudinale de fibre albe peste care trece ca o bandă
transversală, un alt mănunchi de fibre care formează puntea.
Configuraţia externă.
La nivelul trunchiului cerebral se distinge o faţă antero-laterală şi alta posterioară.
- Faţa antero-laterală. Pe această faţă se observă foarte bine cele 3 componente ale
trucnhiului cerebral: bulbul, puntea şi superior pedunculii cerebrali ai mezencefalului (fig. 3).

a. Bulbul este limitat
inferior de planul convenţional
de sub desucaţia piramidelor, iar
superior de şanţul bulbopontin.
Fiind în continuarea măduvei ale
cărei formaţiuni superficale se
continuă şi la acest nivel, se mai
numeşte şi măduva prelungită.
Se observă astfel fisura mediană
anterioară care este întreruptă
inferior de decusaţia
piramidelor. Superior ea se
termină cu o depresiune,
foramen caecum, la nivelul
şanţului bulbopontin. De o parte
şi de alta a fisurii mediane sunt
două cordoane de substanţă albă Figura 3 Trunchiul cerebral - faţa anterioară
proeminete, denumite piramide bulbare, situate în continuarea cordoanelor anterioare ale
măduvei. Lateral de acestea se află şanţul lateral anterior ce continuă pe cel din măduvă.
Mai lateral este cordonul lateral al bulbului în continuarea celui medular şi care va avea
ca limită posterioară şanţul lateral posterior.
În partea superioară a cordonului lateral se găseşte o umflătură ovoidală numită oliva
bulbară, limitată anterior de un şanţ retroolivar ce coincide cu şanţul retroolivar situat ceva mai
înaintea şanţului lateral posterior. De pe suprafaţa bulbului se descriu nervii cranieni de la
perechea a VI-a la a XII-a şi anume: nervul abducens (VI) din şanţul bulbopontin în dreptul
piramidelor, nervul facial (VII) şi acusticovestibular (VIII) tot din şanţul bulbopontin în dreptul
unei depresiuni numită foseta supraretroolivară. Nervul glosofaringian (IX), nervul vag (X) şi
nervul accesor (XI) din şanţul lateral posterior, iar nervul hipoglos (XII) din şanţul preolivar
sau lateral anterior.
b. Puntea este limitată inferior de şanţul bulbopontin, iar superior de un şanţ
transversal paralel cu acesta numit şanţul pontopeduncular care o separă de pedunculii

cerebrali. Pe linia mediană se găseşte un şanţ denumit şanţul artrei bazilare. Lateral de acesta
se văd piramidele punţii, mai puţin proeminente care se continuă lateral şi posterior cu braţele
punţii sau pedunculii cerebeloşi mijlocii. Din punte se desprinde nervul trigemen (V) la limita
dintre piramide şi braţele pontine.
Pedunculii cerebali sunt limitaţi inferior prin şanţul potopeduncular, iar superior se
continuă cu diencefalul. Au aspectul a două benzi divergente denumite picioarele pedunculilor
cerebrali, între care se formează o depresiune triunghiulară cu fundul perforat de vase sanguine
numită substanţă perforată posterioară. În această fosă, de pe faţa medială a picioarelor
peduculilor cerebrali se desprinde nervul oculomotor (III). La baza acestei fose se găsesc corpii
mamilari, două proeminenţe de mărimea unui sâmbure de cireaşă, care aparţin diencefalului.
- Faţa posterioară a
trunchiului cerebral nu poate fi
observată decât după ridicarea
cerebelului prin secţionarea celor 3
perechi de pedunculi cerebeloşi
(superiori, mijlocii şi inferiori). Pe
această faţă limita dintre cele 3
componente ale trunchiului cerebral
este mai puţin evidentă. Bulbul în
jumătatea inferioară prezintă în
continuare formaţiile de la măduvă.
Se observă şanţul median posterior
ce separă între ele cordoanele
posterioare formate de fasciculele
gracilla (Goll) şi cuneatus
(Burdach). Aceasta se termină
superior cu câte o proeminenţă ce
corespunde nucleilor Goll şi Figura 4 Trunchiul cerebral - faţa posterioară
Burdach. În jumătatea superioară, cordoanele posterioare par că se continuă cu două benzi
divergente numite corpi restiformi sau pedunculi cerebeloşi inferiori. Jumătatea superioară a
feţei posterioare a bulbului participă la formarea ventriculului al IV-lea (fig. 4).

Ventriculul al IV-lea este o dilatare a canalului ependimar care apare datorită curburii
pontine.
Este situat la nivelul trunchiului cerebral între bulb şi punte care îi formează podeaua şi
cerebel care îi formează tavanul. I se vor descrie o podea, pereţi laterali şi un tavan.
Podeaua este numită fosa romboidă, deoarece are o formă rombică. La nivelul
trigonului inferior sau bulbar se disting 3 zone: trigonul hipoglosului (aripa albă internă),
fovea vegi (aripa cenuşie) ce corespunde nucleului dorsal al vagului) şi cel mai lateral, aria
vestibulară (aripa albă externă), care se întinde şi în trigonul pontin şi la nivelul căreia se
găsesc nucleii cohleari şi vestibulari. Fisura mediană posterioară împreună cu nişte filamente
superficiale de substanţă albă întinse lateral şi numite strii medulare formează calamus-ul
scriptorius.
La nivelul triunghiului superior sau pontin se observă de o parte şi de alta a fisurii
mediene, două proeminenţe mediale fiecare având în partea inferioară câte o proeminenţă
rotundă, numită coliculul facialului. În profunzime la acest nivel se găseşte nucleul
abducensului (VI). Mai lateral se găseşte o depresiune numită fovea rostralis căreia îi
corespunde în profunzime nucleul motor sau masticator al trigemenului. Pereţii laterali ai
ventriculului al IV-lea vor fi corpii restiformi în trigonul inferior, iar pentru trigonul pontin vor
fi pedunculii cerebeloşi mijlocii (braţele punţii) şi superiori (braţele conjunctive) (fig.23).
Tavanul, în trigonul pontin între breţele conjunctive este format de o lamă de substanţă
albă numită văl medular anterior. Pe diagonala mică a rombului, între braţele punţii este
cerebelul, iar în trigonul bulbar este un înveliş subţire – membrana tectoria care are 3 orificii: 2
laterale (Luschka) şi 1 median (Magendie), prin care sistemul cavitar al nevraxului comunică
cu spaţiul subarahnoidian la nivelul cisternei medulocerebeloase. La nivelul membranei
tectoria, vasele din piamater formează plexurile coroide ale ventriculului IV. Acestea sunt nişte
franjuri vasculare ce proemină în interiorul ventriculilor de lumenul cărora sunt separate prin
învelişul lor ependimar. Ele formează lichidul cerebrospinal.
c. Mezencefalul prezintă pe faţa posterioară tectum-ul mezencefalic sau lama
cvadrigemină format din 2 coliculi superior sau optici şi 2 inferiori sau acustici separaţi între ei
printr-un şanţ cruciat. Coliculul superior este legat prin braţul coliculului superior de corpul
geniculat, iar cel inferior prin braţul colicului inferior de corpul geniculat. Corpii geniculaţi
sunt nucleii diencefalici (aparţin metatalamusului). Sub coliculul inferior se desprinde nervul

trohlear (IV) singurul care îşi are originea pe faţa posterioară a trunchiului cerebral şi care se
încrucişează în nevrax.
Structura internă.
Substanţa cenuşie este la interior, iar cea albă la exterior. Substanţa cenuşie nu mai este
compactă ci datorită fribrelor care se încrucişează este fragmentată în nuclei. Datorită apariţiei
ventriculului IV zonele de substanţă cenuşie care la măduvă erau suprapuse, aici sunt juxta-
puse.
- Substanţa cenuşie. Se disting nuceli ai aparatului elementar consideraţi omologi cu
cei formaţi de substanţa cenuşie a măduvei şi fromaţiuni ale paratului supraelementar ce sunt
reprezentate de nucelii şi de centri supraelementari ce determină acte complexe, prin
intermediul aparatului elementar.
- Aparatul elementar este alcătuit din următorii nuclei din coloana somatomotorie. În
mezencefal sunt nucleul oculomotorului (III) şi al trohlearului (IV), iar în punte nucleul
abducensului (VI), toţi nervi motori pentru musculatura extrinsecă a globului ocular. În bulb
este nucleul hipoglosului (XII) motor pentru muşchii limbii.
Din coloana visceromotorie specială sau branhiomotorie, deoarece inervează
musculatura striată derivată din arcurile branhiale se descriu în punte nucleul motor al
trigemenului şi al facialului, iar în bulb nucleul ambiguu. Trigemenul inervează musculatura
masticatoare, facialul pe cea a mimicei, iar nucelul ambiguu care îşi trimite fibrele prin nervii
IX, X şi XI este motor pentru muşchii laringelui, faringelui şi ai esofagului superior.
Din coloana vegetativă motorie (viscereomotorie generală) în mezencefal este
nucleul accesor al nervului oculomotor (Eddinger Westfall) cu rol iridoconstrictor; în punte se
găsesc nucleii lacrimo-nazal şi salivar superior. Primul determină secreţia glandelor lacrimale
şi a glandelor mucoasei nazale, iar celălalt este secretor pentru glandele submandibulară şi
sublinguală. În bulb se găsesc nucleii salivar inferior secretor pentru glanda parotidă şi nucelul
dorsal al vagului numit şi centru cardiopneumoenteric deoarece inervează parasimpatic aceste
viscere. De fapt, nucleii din această coloană reprezintă şi centrii parasimpaticului cranian.
Din coloana viscerală aferentă se găsesc în bulb nucelul tractului solitar care culege
gustul prin nervii VII, IX şi X şi interoceptivitatea prin nervul IX şi X.

Din coloana somatică aferentă specială sunt nucleii cohleari şi vestibulari care culeg
stimulii de auz şi echilibru. Nucleii cohleari dorsali şi ventrali sunt în punte, iar nucleii
vestibulari: superior, medial, lateral şi inferior, atât în punte, cât şi în bulb.
Din coloana somatică aferentă generală nucleul tractului mezencefalic al trigonului
culege proprioceptivitatea kinestezică de la cap, iar nucleul principal senzitiv al trigemenului
din punte şi nucleul tractului spinal al trigemenului din bulb, pe cea exteroceptivă de la pielea
feţei şi mucoasele cavităţilor feţei.
- Aparatul supraelementar este alcătuit din nucleii proprii şi centri supraelementari
situaţi în formaţia reticulată, a trunchiului cerebral. În bulb se găsesc nucleii Goll şi Burdach
staţie pe calea proprioceptivă kinestezică “conştientă” şi oliva bulbară, staţie pe calea de
coordonare motorie. Se descriu apoi centrii unor funcţii vitale ca cel respirator, vasomotor şi
cardiomotor; centrii unot funcţii digestive ca: deglutiţia, salivaţia, suptul; centrii unor reflexe
de apărare ca: centrul vomei, tusei, strănutului, clipitului (corneean); centrii ai unor funcţii
metabolice ca. Centrul glicozuric şi centrii ai tonusului muscular.
În punte, ca nuclei proprii sunt nucelii pontini, staţie pe calea de coordonare motorie
corticopontocerebeloasă. Centrii supraelementari sunt reprezentaţi de centrul masticator,
centrul salivar superior, centrul lacrimal, centrul clipitului auditivo-palpebral, centrul reflexului
audiooculocefalogir, centrii ai tonusului muscular etc.
Figura 5. Secţiune prin mezencefal
În mezencefal se descriu ca nuclei
proprii, nucleul roşu, substanţa
neagră şi coliculii cvadrigemeni.
Lama cvadrigemină formează tectum-
ul mezencefalic şi este limitată inferior
printr-un plan convenţional ce trece
prin apeductul lui Silvius (fig. 5). Sub
tectum, este tegumentul mezencefalic
limitat inferior de substanţa neagră care îl separă de picioarele pedunculilor cerebrali situaţi sub
substanţa neagră.
Substanţa neagră are celule bogate în pigmenţi. Ea este un centru al motilităţii
extrapiramidale şi este legată şi de substanţa reticulată a trunchiului cerebral.

Nucleul roşu este situat în tegumentul mezencefalic şi are o formă ovalară. Funcţional
este un centru al motilităţii, reglând tonusul centrilor inferiori. Secţiunea sub nucleul roşu dă o
hipertonie marcată, determinată de centrii inferiori pontini şi bulbari, care nu sunt inhibaţi
(rigiditate prin decerebrare).
Coliculii cvadrigemeni constituie un organ superior de integrare la vertebratele
inferioare. La om, funcţiile lui au fost preluate de centrii superiori. Coliculii optici (superiori)
sunt legaţi de reflexele oculocefalogire, fotomotorii, reflexul retinopalpebral, ca şi de o serie de
mişcări automate pentru care trimit comanda prin fasciculele tacospinal şi tectonuclear. Au o
structură laminată şi primesc aferenţe vizuale de la retină, exteroceptive prin tractul tractul
spinotectal, proprioceptive de la cerebel şi de auz de la coliculul acustic. Coliculii optici trimit
fibre eferente către nucleii motori din trunchiul cerebral şi măduvă prin tracturile tectobulbar şi
tectospinal; către cerebel prin tracturile tectocerebelos superior şi inferior şi către cortex, de la
care primeşte şi aferenţe corticotectale.
Coliculii acustici (inferiori) au o structură ganglionară. Ei primesc aferenţe de la
lemniscul lateral şi sunt centrii pentru reflexele declanşate de stimulii acustici (reflexul
acusticooculocefalogir). Ca centrii supraelementari se descriu centru de veghe, centrul
reflexelor de redresare etc.
- Substanţa albă. Substanţa albă a trunchiului cerebral este alcătuită din căi ascendente
şi descendente care asigură funcţia de conducere a acestui segment.
Căile ascendente.
Cuprind fibrele ascendente ce duc sensibilitatea de la trunchi şi membre, fibrele ce duc
sensibilitatea de la nivelul extremităţii cefalice şi fibrele ascendente din căile de coordonare
motorie.
a) Căile senzitive de la măduvă sunt reprezentate de următoarele fascicule:
- pentru sensibilitatea exteroceptivă este tractul spinotalamic anterior şi lateral, care de
la punte formează un singur traiect spinotalamic ce se termină în nucleul ventral postero-lateral
talamic. Tractul spinotectal conduce exteroceptivitatea la coliculii optivi.
- sensibilitatea kinestezică (proprioceptivă "conştientă”) ajunge prin fasciculul Goll şi
Burdach la nucelii omonimi din bulb, unde este deutoneuronul acestei căi. Axonii

deutoneuronilor se încrucişează şi formează în continuare lemniscul medial (panglica Reil) care
urcă până la nucleul ventral postero-lateral talamic unde este al III-lea neuron.
- sensibilitatea proprioceptivă de coordonare motorie ce merge prin tractul
spinocerebelos posterior, urcă la cortexul cerebelos prin pedunculii cerebeloşi inferiori, iar cea
adusă de tractul spinocerebelos anterior străbate tot trunchiul cerecbral şi prin pedunculii
cerebeloşi superiori se îndreaptă spre cortexul cerebelos.
- sensibilitatea interoceptivă de la măduvă urcă prin substanţa reticulată cu multiple
staţii şi parţial prin tractul spinotalamic.
b) Căile senzitive de la extremitatea cefalică sunt reprezentate de următoarele fascicule:
- lemniscul trigeminal (fasciculul trigeminotalamic) conduce extero şi
proprioceptivitatea extremităţii cefalice. El este format din axonii deutoneuronilor din nucleii
senzitivi ai trigemenului situaţi în tot lungul trunchiului cerebral. El se termină la nucleul
ventral postero-medial (arcuat) talamic unde este al III-lea neuron.
- calea cohleară este formată din axonii deutoneuronilor din nucleii cohleari; aceşti
axoni se încrucişează la nivelul punţii formând corpul trapezoid care separă partea tegumentală
de cea bazilară a punţii, apoi devin ascendenţi şi alcătuiesc lemniscul lateral care urcă spre
corpii geniculaţi mediali. Din lemniscul lateral merg colaterale la coliculii acustici ce mijlocesc
reflexe ale trunchiului cerebral declanşate de auz.
- sensibilitatea gustativă şi cea interoceptivă culeasă de glosofaringian şi vag merg prin
fibre solitariotalamice la al III-lea neuron din talamus.
c) Căile ascendente fac parte din căile de coordonare motorie sunt reprezentate în bulb
de fasciculul vestibulocerebelos şi fibrele clivocerebeloase, iar în mezencefal de fasciculele
dentotalamic, dentorubric şi dentotectal.
Căile descendente.
Acest căi duc comenzile de la etajele superioare şi din trunchiul cerebral, pentru
mişcările voluntare, automate, reflexe de echilibru, ca şi pentru coordonarea lor. Cele pentru
mişcări voluntare se numesc căi piramidale, iar celelalte căi extrapiramidale.
a) Căile pentru mişcări voluntare ale trunchiului şi membrelor merg prin fasciculul
corticospinal, iar cele pentru mişcările voluntare de la nivelul capului, prin fasciculul
corticonuclear (fasciculul geniculat). Ambele fascicule pleacă din girul precentral. La nivelul
diencefalului fasciculul corticospinal trece prin braţul posterior al capsulei interne, o zonă de

substanţă albă situată între talamus şi nucleul lentiform, iar fasciculul corticonuclear prin
genunchiul capsulei interne şi de aceea este numit şi fasciulul geniculat. Fasciulul geniculat
merge în trunchiul cerebral la nucleii motori ai acestuia. La nivelul bulbului, peste 80 % din
fibrele fasciculului corticospinal se încrucişează (decusaţia piramidelor) aşa că în partea
inferioară a bulbului şi în măduvă se găseşte fasciculul corticospinal anterior (piramidal direct)
şi unul lateral (piramidal încrucişat).
b) Căile pentru mişcări automate ale trunchiului şi membrelor ca şi calea pentru
mişcările automate de la nivelul capului merg prin fasciculul tectospinal şi tectonuclear. La
origine în mezencefal, tracturile tectospinale se încrucişează formând decusaţia dorsală a
tegmentului (Meynert).
c) Mişcările reflexe de echilibru sunt comandate prin fasciculul vestibulospinal, ce va
străbate bulbul şi măduva.
d) Alte căi extrapiramidale sunt reprezentate de: tractul rubrospinal şi rubronuclear,
tractul corticopontin, tractul olivospinal şi tractul reticulospinal. Tracturile rubrospinale se
încrucişează în mezencefal formând decusaţia ventrală a tegmentului (Forel). De la nucleii
pontini unde se opreşte tractul corticopontin, pleacă fibrele pontocerebeloase prin pedunculii
cerebeloşi mijlocii, spre scoarţa cerebrală.
Căile proprii.
Căile proprii ale trunchiului cerebral sunt asociative între diferiţi nuclei ai acestuia.
Printre acestea se descriu:
- fasciculul longitudinal medial care asociază nucleii vestibulari cu nucleii nervilor
motori oculari şi cu nucleul accesorului, intervin astfel în menţinerea axului de prvire prin
coordonarea mişcărilor capului şi globilor oculari, în funcţie de stimulii veniţi de la receptorii
vestibulari din urechea internă.
- fasciculul longitudinal dorsal cu fibre de natură vegetativă ce pleacă de la centrii
vegetativi diencefalici spre nucelii vegetativi ai trunchiului cerebral.
- fasciculul central al tegmentului sau palidorubroolivar este pe calea extrapiramidală
de coordonare motorie.

Unitatea de curs 4 Anatomia cerebelului
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului cerebelului.
Obiectivele operaţionale
1. Prezentarea configuraţiei externe a cerebelului
2. Prezentarea structuriiinterne a cerebelului
3. Prezentarea căilor de conducere cerebelare.
Cerebelul
Configuraţia externă.
Cerebelul sau creierul mic este un organ nervos cu rol în coordonarea motorie. Se
dezvoltă din lamele alare ale metencefalului. Cerebelul are o formă de hemiovoid cu o faţă
superioară care este separată de lobii occipitali ai emisferelor cerebrale printr-o prelungire
orizontală a dureimater denumită cortul cerebelului. Faţa inferioară a cerebelului este în raport
cu fosele cerebeloase ale scuamei occipitalului, iar faţa anterioară vine în raport pe linia
mediană cu trunchiul cerebral de care o desparte ventriculul IV, iar pe lături cu faţa posterioară
a stâncilor temporalului.
Cerebelul este legat de trunchiul cerebral prin 3 perechi de pedunculi cerebeloşi.
Pedunculii cerebeloşi inferiori sau corpii restiformi îl leagă de bulb, pedunculii cerebeloşi
mijlocii sau braţele punţii îl leagă de punte, iar pedunculii cerebeloşi superiori sau braţele
conjunctive îl leagă de mezencefal.
Clasic este împărţit prin 2 şanţuri sagitale într-o parte mediană denumită vermis şi 2
părţi laterale, lăţite denumite emisferele cerebeloase. Mai recent, corespunzând şi din punct de
vedere fiziologic şi filogenetic, se descriu şanţuri orizontale care împart atât vermisul, cât şi
emisferele în lobi. {anţurile primare separă între ei, lobul anterior (paleocerebelul) de lobul
mijlociu care este mai recent apărut (neocerebelul), iar fisura postero-laterală separă lobul
mijlociu de lobul floculonodular care este cel mai vechi filogenetic (arhicerebelul) (fig. 6).

Figura 6. Cerebelul - faţa superioară (stânga), faţa inferioară (dreapta)
Fisura orizontală separă faţa superioară de faţa inferioară a cerebelului. şanţurile mai
puţin adânci determină o lobulaţia a vermisului căreia îi corespunde o lobulaţie a emisferelor
cerebeloase. În final şanţuri şi mai puţin adânci separă suprafaţa cerebelului în folii sau lamele,
mărind astfel suprafaţa acestuia. Dinainte înapoi se disting în vermis lobuli, care au
corespondentul în emisferele cerebeloase.
În lobul anterior se descrie lingula cu frâurile lingulei, lobulul central cu aripa lobulului
central, culmen cu lobul cvadrangular anterior, posterior de care este şanţul primar.
În lobul mijlociu sunt declive cu lobul cvadrangular posterior (culmen şi declive
formează împreună monticulus), iar lui folium îi corespunde lobul semilunar superior. Faţa
superioară este separată de faţa inferioară prin fisura orizontală.
În continuare pe faţa inferioară, dar în sens postero-anterior se disting: tuber, vermis cu
lobul semilunar inferior, piramida cu lobulul biventer, uvula cu tonsila sau amigdala
cerebelului, iar după fisura postero-laterală nodulus cu floccus ce aparţin arhicerebelului. De
remarcat, că piramidele şi uvula sunt formaţiuni paleocerebeloase.
Structura cerebelului.
Cerebelul este alcătuit din substanţă cenuşie dispusă la suprafaţă formând scoarţa sau
cortexul cerebelos, iar substanţa albă formată din fibre se află la interior. În substanţa albă se
mai găsesc grămezi de neuroni care formează nucleii cerebelului.
- Substanţa cenuşie. Cortexul cerebelos este alcătuit dint 3 straturi celulare. Stratul
superificial sau stratul molecular, pentru că este format din celule mici are printre celulele sale
unele denumite "în coşuleţ”. Acestea îşi trimit axonii către stratul următor unde colateralele lor
se ramifică în jurul celulelor Purkinje, cuprinzându-le ca într-un coş.

Al doilea strat este denumit stratul ganglionar sau al celulelor lui Purkinje, cu cele mari
piriforme, bipolare care trimit dendritele în stratul molecular, iar axonii în substanţa albă spre
nucleii cerebelului. Cel mai profund este stratul granular format din celule mici, stelate al
căror axon urcă în stratul molecular unde se ramifică în "T”, făcând sinapsă cu dendritele
neuronilor Purkinje.
Nucleii cerebelului sunt reprezentaţi de nucleii fastigiali în dreptul vermisului şi lateral
de ei, spre emisferele cerebeloase sunt nucleii globoşi, emboliformi şi dinţaţi. Toţi sunt
paleocerbeloşi cu excepţia unei părţi din dinţat care este de apariţia neocerebeloasă.
- Substanţa albă. Este situată la interior şi apare pe o secţiune medio-sagitală cu un
aspect ramificat caracteristic, care a fost numit "arborele vieţii”. În ea se disting fibre
asociative intracerebeloase care leagă puncte din aceeşai emisferă cerebeloasă, fibre
comisurale care leagă zone simetrice şi fibre de proiecţie aferente şi eferente. Aferenţele ajung
la cortexul cerebelos, iar eferenţele pleacă de la nucleii cerebelului. Toate conexiunile
cerebelului se realizează prin intermediul pedunculilor cerebeloşi.
Pentru a înţelege conexiunile cerebelului trebuie să amintim că cerebelul are rol
principal în coordonarea motorie. El poate realiza această coordonare datorită faptului că
centrii care comandă mişcările voluntare, automate şi de echilibru (cortexul cerebral, coliculii
optici, nucleii vestibulari) informează în derivaţie şi cerebelul prin căile
corticopontocerebeloase, tectocerebeloase superioare şi inferioare şi respectiv, vestibulo-
cerebeloase.
În acelaşi timp prin fasciculele spinocerebeloase anterioare şi posterioare, cerebelul este
informat de ceea ce s-a executat din mişcarea comandată. El integrează aceste aferenţe şi
printr-un mecanism de feed-back (conexiune inversă) reinformează centrii care au dat
comanda, ca să şi-o modifice în funcţie de gradul ei de executare. Această informare şi
modelare a centrilor motori se face prin: fasciculul cerebelo-dento-talamo-cortical pentru
centrii corticali ce au comandat mişcări voluntare; prin fasciculul cerebelo-dento-tectal pentru
coliculii optici care au determinat mişcările automate; şi prin fasciculele cerebelo-fastigio-
vestibular pentru nucleii vestibulari care au imprimat mişcări reflexe de echilibru.
Concomitent cerebelul intervine şi direct la periferie acţionând asupra grupelor de
muşchi antagonişti pentru a bloca mişcarea la momentul necesar, fapt care se realizează prin

multiple căi, ca de exemplu cerebelo-dentro-rubro-spinală şi rubrospinală, cerebelo-fastigio-
recticulo-spinală, cerebelo-fastigi-oolivo-spinală, cerebelo-fastigio-olivo-spinală etc.
În pedunculii cerebeloşi inferiori, aferente sunt fibrele din fasciculele spinocerebelos
posterior, tectcerebelos inferior, vestibulocerebelos, olivocerebelos, iar eferente cele din
fasciculele fastigiovestibular, fastigioolivar, fastigioreticular.
În pedunculii cerebeloşi mijlocii se descriu doar fibre aferente pontocerebeloase. În
pedunculii cerebeloşi superiori fasciculul spinocerebelos anterior şi tectocerebelor superior
sunt aferente, iar fasciculele dentotalamic, dentorubric şi dentotectal sunt eferente.

Unitatea de curs 5 Anatomia diencefalului
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului diencefalului.
Obiectivele operaţionale
1. Prezentarea configuraţiei externe a diencefalului.
4. Prezentarea structuriiinterne a diencefalului.
5. Prezentarea căilor de conducere diencefalice.
Diencefalul sau creierul intermediar se dezvoltă din vezicula diencefalică ce rezultă din
segmentarea prozencefalului. Din diencefal se desprind veziculele optice care vor da naştere
tracturilor, chiasmei şi nervilor optici, ca şi unei părţi din globul ocular (retina). În vezicula
primitivă se vor evidenţia cele 3 straturi (ependimar, paleal şi marginal), care se dezvoltă mult
şi fac la nivelul diencefalului, mase mari cenuşii, cu substanţă albă redusă.
Diencefalul este aşezat în
continuarea mezencefalului sub
emisferele cerebrale care îl acoperă
lăsând vizibilă la exterior doar faţa
inferioară ce corespunde
hipotalamusului. În interiorul său se
găseşte ventriculul al III-lea.
Diencefalul este alcătuit din talamus,
metatalamus, subtalamus, epitalamus
şi hipotalamus, situate în jurul
ventriculului al III-lea (fig. 7).
Configuraţia exterioară.
Pe singura faţă vizibilă la exterior sau
faţa bazală care corespunde
hipotalamusul (HT) se observă în
partea anterioară chiasma optică de la
care pleacă spre corpii geniculaţi Figura 7. Regiunea habenulară şi faţa superioară a talamusului

laterali, tracturile optice ce limitează lateral această faţă bazală. Pe linia mediană, înapoia
chiasmei optice se găseşte o formaţiune proeminentă denumită tuber cinerum, cu o prelungire
în formă de pâlnie denumită infundibulum de care printr-o tijă este prinsă glanda hipofiză.
Posterior se găsesc 2 mici proeminenţe denumite corpi mamilari (fig. 8). Faţa
superioară a diencefalului este făcută de talamus, ce participă la formarea ventriculilor laterali.
Feţele laterale sunt separate prin capsula internă de corpii striaţi.
Ventriculul al III-lea situat median este o dilatare care provine din cavitatea primitivă a
tubului neural. Podeaua ventriculului este formată de hipotalamus, pereţii laterali sunt formaţi
în cea mai mare parte de talamus, dar o porţiune mică inferioară unui şanţ numit şanţ
hipotalamic este formată de hipotalamus.
Tavanul este format de pânza coroidiană epitelială a ventriculului III, căptuşită de
piamater. La nivelul tavanului se găsesc plexurile coroide ale acestui ventricul. Pânza
coroidiană este întinsă între 2 benzi subţiri de substanţă albă numite strii medulare
(habenulare) care se află la marginea ce separă faţa superioară de cea medială a talamusului. În
partea posterioară, ventriculului III comunică prin apenductul Syvius cu ventriculul al IV-lea,
iar anterior şi lateral prin 2 orificii denumite găurile interventriculare (Monroe) cu ventriculii
laterali (I şi II) din interiorul emisferelor cerebrale.

Figura 8. Hipotalamusul
Structura diencefalului.
Talamusul este format din 2
mase mari cenuşii, cu o formă
ovoidă cu polul anterior mai ascuţit
numit rosturm, iar cu cel posterior
mai rotunjit numit pulvinar. Lateral,
braţul posterior al capsilei interne îl
separă de nucleul lentiform. În
interior o lamă medulară internă de
substanţă albă, bifurcată anterior ca
un “Y”, îl separă în nuclei. Se
descriu următorii nuclei: rostral,
posterior, medial, lateral şi ventral.
În nucleul ventral sunt 3
subdiviziuni: nucleul ventral
anterior, nucleul ventral lateral şi
nucleul ventral posterior, care la
rândul său este împărţit într-un nucleul lateral şi altul medial sau arcuat. Tot în talamus,
majoritatea specialiştilor include şi corpul geniculat lateral şi cel medial, metatalamusul
rămânând mai mult o noţiune topografică.
Aferenţele talamusului cu excepţia căilor olfactive, toate celelalte căi ascendente către
cortex, cu al III-lra neuron în talamus. Fasciculul spinotalamic cu exteroceptivitatea şi
lemniscul medial cu proprioceptivitatea kinestezică de la corp, merg la nucleul ventral
posterolateral. Exteroceptivitatea şi proprioceptivitatea kinestezică de la extremitatea cefalică,
ca şi stimulii gustativi, ajung la nucleul ventral postero-medial sau arcuat prin lemniscul
trigeminal.
Interoceptivitatea vine din hipotalamus la nucleii rostrali. Auzul condus prin lemniscul
lateral ajunge la corpul geniculat medial, iar stimulii optici prin tractul optic ajung la corpul
geniculat lateral.

Eferenţele talamusului. Fibrele care pleacă de la talamus formează pedunculii sau
radiaţiile talamice, care merg spre aria corticală de integrare a sensibilităţii respective şi care
vor fi studiate la emisferele cerebrale. Se descriu radiaţii optice, acustice, gustative etc.
Substalamusul este situat între talamus şi mezencefal. La nivelul lui se găsesc: nucleul
subtalamic, zona incertă şi câmpul lui Forell cu 2 fascicule de substanţă albă, formaţiunile lui
sunt staţii de legătură ale sistemului extrapiramidal, strâns legate funcţional de corpii striaţi de
la baza emisferelor cerebrale.
Epitalamusul este situat în partea posterioară şi superioară a diencefalului şi este format
din:
- glanda epifiză legată printr-o tulpină de comisura habenulei;
- trigonul habenulei cu nucleul habenular;
- striile habenulei.
Prin comisura şi striile habenulare nucleul habenulei se leagă de centrii olfactivi de la
baza emisferelor cerebrale constituind un centru al reflexelor olfactivo-somatice.
Hipotalamusul este situat în partea inferioară a diencefalului unde formează podeaua şi
partea inferioară a pereţilor laterali ai ventriculului al III-lea. Este alcătuit dintr-o porţiune
supraoptică unde se individualizează NPV, din tuber cinerum unde se găsesc nucleii tuberali
regiune căreia îi aparţin şi neurohipofiza, iar posterior din corpii mamilari.
În rest nucleii din hipotalamus (HT) nu pot fi bine individualizaţi, dar sunt
parasimpatici în hipotalamusul antero-posterior şi simpatici în hipotalamusul posterior, deci
hipotalamusul poate fi considerat ca un creier vegetativ. În plus în hipotalamus se află şi
centrul somnului, centrul matabolismului glucidic, protidic, lipidic şi al apei; centrul setei,
centrul foamei, centrul saţietăţii. Tot în el sunt centrii ce intervin în funcţiile sexuale şi în
coportamentul individului.
Conexiunile hipotalamusului sunt internucleare sau asociative şi de proiecţie.
Hipotalamusul primeşte aferenţe de la centrii vegetativi inferiori, pe care la proiectează pe
cortexul cerebral direct sau prin intermediul nucleilor rostrali talamici în ariile 14, 15, 24, 28,
32 (interoceptive) de pe faţa medială a emisferelor cerebrale, în girus cinguli. Hipotalamusul
primeşte informaţii şi comenzi de la cortex, trimiţând eferenţe spre centrii vegetativi inferiori
(fasciculul dorsal Schütz).

Din punct de vedere funcţional, hipotalamusul apare ca un tablou central de comandă
pentru reglarea majorităţii funcţiuinilor din organism.
El este informat prin 2 căi: umorală şi nervoasă. Umoral prin calităţile sângelui care-l
irigă, iar nervos prin căile aferente atât ascendente, cât şi desendente de la cortex. El
prelucrează mesajele şi ia decizii pe care le trimite la organele efectoare tot prin cele 2 căi
(nervoasă şi umorală).
Calea nervoasă este realizată de căile descendente către centrii vegetativi inferiori, care
prin fibrele simpatice şi parasimpatice modifică activitatea efectorilor, dar şi prin intervenţia pe
componenta somatică (ex. frisonul termic).
Calea umorală prin care hipotalamusul intervine la periferie este reprezentat de
secreţiile hipofizei, glandă cu care are strânse legături. De la nucleul supraoptic şi
paraventricular care secretă ADH şi ocitocina prin axonii tractului hipotalamohipofizar aceste
neurosecreţii ajung în neurohipofiză unde sunt depozitate. De aici ele sunt eliminate în sânge în
funcţie de necesităţi. Rolul de depozit al neurohipofizei a fost dovedit prin ligatura tijei care a
făcut să scadă cantitatea de hormoni din glandă şi să se acumuleze în tijă deasupra ligaturii.
Nucleii tuberali reglează activitatea adenohipofizei. Ei secretă factori de eliberare (RF
= releasing factors) care pe calea adenohipofiză, determinând secreţia de hormoni tropi a
acesteia. Sistemul venos porthipofizar are reţeaua primară în HT, venele în tija hipofizei, iar
reţeaua capilară secundară în adenohipofiză.
Secreţia nucleilor tuberali este reglată printr-un mecanism umoral de conexiune inversă
de către concentraţia hormonilor specifici secretaţi de glande sub acţiunea hormonilor tropi
adenohipofizari.

Unitatea de curs 6 Anatomia emisferele cerebrale
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului emisferelor cerebrale.
Obiectivele operaţionale
1. Prezentarea configuraţiei externe a emisferelor cerebrale
2. Prezentarea structuriiinterne a emisferelor cerebrale
3. Prezentarea ariilor şi ai centrilor de comandă ai emisferelor cerebrale.
Emisferele cerebrale în număr de două sunt separate de fisura interemisferică, se
dezvoltă din vezicula telencefalică şi formează, împreună cu diencefalul, creierul sau
cerebrum. La om scoarţa cerebrală are întinderea maximă şi o structură complexă, devenind
centrul superior de integrare a tuturor funcţiilor somatice şi vegetative ale organismului.
Emisferele sunt legate prin comisuri, graţie cărora ele funcţionează simultan, iar în interior
conţin cavităţi denumite ventriculi laterali.
Configuraţia exterioară.
Emisferele cerebrale de formă ovoidă şi cu axul mare antero-posterior, au 3 feţe:
convexă, medială, bazală şi 3 poli: frontal, temporal şi occipital. Pe suprafaţa lor se văd şanţuri
mai adânci care delimitează lobii, iar la nivelul lobilor şanţuri mai puţin adânci care separă
între ei girii cerebrali. Fisuraţia şi giraţia duc la creşterea suprafeţei scoarţei cerebrale (fig. 9).
Figura 9 Faţa laterală a encefalului

Faţa convexă prezintă 2 şanţuri mai adânci: şanţul lateral şi şanţul central. {anţul lateral
(Sylvius) separă lobul temporal situat dedesubt de lobii frontal şi parietal situaţi deasupra lui.
{anţul lateral începe pe faţa bazală înconjură marginea laterală şi se îndreaptă oblic în sus şi
posterior. În fundul şanţului se vede o proeminenţă numită lobul insulei. {anţul central aproape
perpendicular pe cel precedent separă între ei lobul frontal situat anterior, de lobul parietal
situat posterior. Capul şanţului parietooccipital separă incomplet lobul parietal de cel occipital.
Lobul frontal are 2 şanţuri orizontale – şanţul frontal superior şi inferior care sunt
biturcate posterior, formând şanţul precentral. Aceste şanţuri delimitează girii frontal, superior,
mediu, inferior şi precentral.
Lobul parietal prezintă un şanţ postcentral care limitează posterior girul postcentral şi
un şanţ intraparietal care separă lobulul parietal superior de cel inferior care prezintă girul
supramarginal la capătul şanţului lateral Sylvius şi mai posterior girul angular în jurul capului
şanţului temporal superior.
Lobul temporal are la nivelul buzei inferioare a şanţului lateral Sylvius 2 şanţuri
temporale transverse, care delimitează girii temporali transverşi, iar pe faţa convexă se află
şanţurile temporal superior şi mediu, ce separă girii temporal superior, mijlociu şi inferior.
Lobul occipital are şanţul transvers şi lateral ce mărginesc girii occipitali superior şi
laterali. Faţa medială este vizibilă după secţionarea formaţiunilor comisurale interemisferice.
Pe faţa superioară a corpului calos este şanţul corpului calos. Superior şi lateral cu acest şanţ se
află şanţul cingului ce devine ascendent în partea sa posterioară. Din el se desprinde şanţul
paracentral cu traiect ascendent. În lobul parietal se observă şanţul subparietal, iar posterior
şanţul perietooccipital.
Sub aceste şanţuri delimitează următorii giri: girul cinguli, paralel cu corpul calos care
în partea posterioară printr-un istm se continuă cu girul parahipocampal ce aparţine feţei
bazale. Deasupra şanţului cingului se găseşte girul frontal superior (intern). Posterior de el este
lobulul paracentral. Înaintea şanţului parietoocipital este o parte a lobulului parietal numită
precuneus, iar acest şanţ şi fisura calcarină se află cuneusul ce aparţine lobului occipital
(fig.10).

Figura 10. Faţa medială a emisferei cerebrale
Faţa bazală a emisferelor cerebrale este formată de faţa inferioară a lobilor: frontal,
temporal şi occipital. La nivelul lobului frontal se găseşte şanţul olfactiv, cu direcţia antero-
posterioară care adăposteşte bulbul şi tractul olfactiv. Medial de acest şanţ se află girul drept
(fig. 11). Lateral de el se găsesc şanţurile orbitale, de forma literei "H”, care delimitează girii
orbitali al lobului frontal. În partea temporooccipitală, girul temporal inferior este separat prin
şanţul temporal inferior de lobul temporooccipital. Acesta este împărţit printr-un şanţ colateral
în 2 giri temporooccipitali.

Figura 11. Faţa bazală (inferioară) a emisferei cerebrale
Girul temporooccipital lateral se numeşte şi girul fusiform, iar cel medial se
subdivizează în girul lingual situat posterior, care formează buza inferioară a şanţului calcarin
şi girul parahipocampic situat anterior, care prezintă o formaţiune ca un cârlig numită uncus
(lat. uncus = cârlig). Girul parahipocampic este limitat medial de şanţul hipocampului care
proemină în interiorul ventriculului lateral unde formează cornul lui Ammon sau hipocampul.
Structura emisferelor cerebrale.
- Substanţa cenuşie ca şi la cerebel este dispusă la suprafaţă, formând scoarţa cerebrală
şi la baza emisferelor alcătuind nucleii bazali sau corpul striat.
Scoarţa cerebrală (pallim).
Scoarţa cerebrală este organul superior de integrare a funcţiilor întregului organism.
Prin giraţie şi fisuraţie suprafaţa ei a crescut ajungând la 1300-2300 cm2, cuprinzând un număr
de cca. 14 miliarde de neuroni. Cortexul cerebral are o regiune mai veche, netedă şi redusă ca
întindere denumită paleocortex, corespunzătoare riencefalului şi alta de achiziţie recentă, cu
giraţie şi figuraţie bogată şi cu întindere mare în suprafaţă, denumită neocortex.
Structura scoarţei cerebrale.

La microscop se constată că scoarţa cerebrală este alcătuită din neuroni şi prelungirile
lor, dispuşi în mai multe straturi. Există o regiune cu 2-3 straturi numită allocortex şi alta mai
întinsă, alcătuită din 6 straturi, denumită izocortex. La nivelul allocortexului care corespunde în
mare parte riencefalului, se disting 2 straturi celulare: unul superficial, stratul granular, format
mai ales din celule receptoare şi altul profund, stratul piramidal, cu celule motorii şi de
asociere.
La nivelul izocortexului se descriu începând dinspre suprafaţă următoarele straturi:
- stratul molecular, alcătuit din celule mici cu numeroase prelungiri dendritice şi axoni
care formează o reţea;
- stratul granular extern, format din celule mici poligonale sau tringhiulare, cu nucleu
mare, care îi dau aspect granular;
- stratul piramidal extern, cu celule piramidale mici, cu o dendrită care pleacă de la vârf
şi ajunge în stratul molecular. Celelelalte dendrite rămân la nivelul acestui strat. Axonii trec în
substanţa albă;
- stratul granular intern, cu celule asemănătoare stratului granular extern şi ale căror
prelungiri nu părăsesc scoarţa;
- stratul piramidal intern sau ganglionar, alcătuit din celule piramidale mari şi altele
foarte mari (giganto-piramidale Betz) ale căror prelungiri se comportă ca şi acelea ale stratului
piramidal extern;
- stratul polimorf, format din celule numeroase, cu aspecte variate. Unele dintre ele,
celulele lui Martinotti îşi trimit axonul în stratul molecular.
Celulele granulare au rol receptor, iar cele piramidale un rol efector. Izocortexul la
nivelul căruia cele 6 straturi sunt dezvoltate proporţional se numeşte homotipic, spre deosebire
de cortexul heterotipic în care predomină dezvoltarea mai mare a unor straturi. Astfel, se
descrie un izocortex heterotipic piramidal sau granular în zonele cu funcţie motorie şi un
izocortex heterotipic granular sau coniocortex, în zonele senzitive şi senzoriale.
Se descriu la nivelul scoarţei cerebrale o citoarhitectonică ce se ocupă de celulele din
cortex şi dispoziţia lor şi a mieloarhitectonică ce analizează dispoziţia fibrelor din scoarţa
cerebrală. Scoarţa cerebrală cuprinde zone sau câmpuri corticale cu structură şi funcţii
specifice, între care nu există limite nete. Brodmann, bazându-se pe citoarhitectonică, a descris
la nivelul cortexului 52 de câmpuri sau arii corticale, cu funcţii specifice.

1. Ariile senzitive şi senzoriale sunt locul de proiecţie a sensibilităţii generale şi
senzoriale. În ceea ce priveşte fiziologia la nivelul ariilor receptoare se disting 2 zone: una
primară de maximă specializare, zona receptoare propriu-zisă şi a cărei distrugere duce la
dispariţia sensibilităţii respective, iar una în jurul acesteia aşa-zisa arie senzitivă secundară sau
psihosenzorială, de asociaţie. Lezarea ei duce la pierderea posibilităţilor de asociere a senzaţiei
primare cu informaţii de la ceilalaţi analizatori, la incapacitatea de formare de reprezentări şi
noţiuni. La nivelul ariilor receptoare se găseşte coniocortex.
Câmpurile 3, 2 şi 1 din girul postcentral reprezintă aria somestezică unde sunt centrii de
integrare a sensibilităţii generale somatice. Aici ajunge sensibilitatea tactilă epicritică, termică,
dureroasă şi kinestezică (proprioceptivă "conştientă”). Unii proiectează kinestezia şi la nivelul
ariei 4 care este însă predominant motorie. Se descrie şi o somatotopie a neuronilor ce
realizează aşa numitul homunculus senzitiv.
Câmpurile 5 şi 7 din lobul parietal sunt arii de asociaţie ce deservesc somestezia.
Câmpurile 13, 14, 24, 25, 32 de pe faţa medială a emisferelor şi din girus cinguli reglează
funcţiile vegetative, fiind în strânsă legătură cu centrii vegetativi diencefali (HT). Câmpurile
17, 18, 19 reprezintă aria vizuală situată de o parte şi de alta a fisurii calcarine. Câmpul 17 se
numeşte şi arie striată pentru că la nivelul lui scoarţa are şi o strie de substanţă albă. Ariile 18 şi
19 se numesc peristriată şi respectiv parastriată (fig. 12, 13). Câmpurile 41, 42, 22 reprezintă
aria de integrare a mesajelor acustice. Câmpul 43 de la piciorul girului postcentral este centrul
gustativ.

Figura 12 Cortexul feţei externe a emisferei cerebrale stângi (Brodmann)
2. Ariile motorii sunt acelea de la care pleacă comenzi pentru mişcările voluntare ca şi
pentru coordonarea lor. Câmpul 4 din girul precentral este locul de origine a tracturilor
corticospinale şi corticonucleare. La acest nivel se găseşte izocortex heterotipic piramidal cu
numeroase celule gigantopiramidale. Excitarea la nivelul câmpului 4 produce contracţia
muşchilor scheletici, iar distrugerea sa dă paralizii musculare.
Există o somatotropie a neuronilor motori care realizează aşa-zisul homunculus motor
(Penfield) cu reprezentare răsturnată deoarece de la piciorul girului pleacă inervaţia pentru cap
(1/3), din partea mijlocie cea pentru membrul superior şi trunchi (1/3), iar cea din partea
superioară a girului (1/3) şi chiar de la nivelul feţei interemisferice, pentru membrul inferior şi
perineu.
Reprezentarea este inegală datorită raportului de inervaţie al unităţilor motorii. Un
neuron din cortex inervează numai 3-6 fibre la muşchii care execută mişcări fine cum sunt cei
ai globului ocular sau ai policelui şi 300-600 fibre de la nivelul muşchilor spatelui.
Câmpurile 6 şi 8 sunt arii motorii de asociere, coordonând mişcări complexe ca
oculocefalogiria, masticaţia, deglutiţia, etc. Distrugerea lor duce la lipsa de coordonare a

mişcărilor complexe şi nu dă paralizie. Acste câmpuri reprezintă şi segmentul cortical al căilor
extrapiramidale.
3. Ariile de asociere sau zonele psihice, care se formează în cursul veţii, se împart în
zone de asociaţie motorie ca centrii vorbirii şi ai scrisului şi arii de asociere senzitivă ca centrul
înţelegerii cuvintelor scrise sau vorbite.
Câmpurile 44 şi 45 din girul frontal inferior reprezintă centrul limbajului articulat
(câmpul 44 este centrul lui Broca). La stângaci se află în hemisfera dreaptă. Lezarea lui dă
tulburări de vorbire (disartrie). Centrul scrisului se dezvoltă lanivelul girului frontal mijlociu,
lezarea lui produce incapacitatea de a scrie sau agrafia.
Câmpul 22 este şi locul înţelegerii cuvintelor vorbite. Lezarea lui dă surditatea verbală
(deşi aude nu înţelege ce vorbeşte). Câmpurile 39 şi 40 au funcţii complexe de analiză şi de
sinteză, lezarea lor produce şi incapacitatea de a înţelege cuvintele scrise sau alexia. Câmpurile
9, 10 şi 11 de la nivelul lobului frontal, reprezintă sediul integrării sentimentelor, de stăpânire a
emoţiilor etc. Lezarea lor produce tulburări de comportament. În general lobul frontal este
sediul personalităţii umane.
Figura 13 Zonele senzoriale de pe faţa internă a emisferei cerebrale drepte.
Nucelii bazali.
Sunt mase de substanţă cenuşie, situate la baza emisferelor cerebrale, superior şi lateral
de talamus. Din cauza aspectului pe care îl au pe secţiune se numesc şi corpul striat. Sunt

formaţi din: nucleul caudat înfăşurat în jurul talamusului şi din nucleul lentiform alcătuit la
rândul său dintr-o parte laterală numită putamen şi alta medială mai decolorată numită globus
pallidus. Nucleului caudat i se descriu un cap, un corp şi o coadă, care se termină în nucleul
amigdalian. Anexat nucelilor bazali este şi claustrum care apre ca o lamă cenuşie lateral de
putamen.
Prezenţa nucleilor bazali datermină în substanţa albă de la baza emisferelor formarea
unor lame de substanţă albă. Acestea sunt reprezentate de capsula internă, externă şi extremă.
Capsula internă are un braţ anterior între capul nucleului caudat şi nucleul lentiform şi unul
posterior între talamus şi nucleul lentiform. La unirea celor 2 braţe se găseşte genunchiul
capsulei interne. Ea este străbătută de fibre de proiecţie ale emisferelor cerebrale. Capsula
externă este situată între nucleul lentiform şi claustrum, iar capsula extremă este între
claustrum şi scoarţa lobulului insulei. Nucleii bazali se leagă prin fibre nervoase între ei şi au
conexiuni şi cu scoarţa cerebrală, talamusul, hipotalamusul, subtalamusul, nucleul roşu,
substanţa neagră, nucleul olivar şi formaţiunea reticulată.
- Substanţa albă a emisferelor cerebrale.
Formează cemtrul emisferelor, masa de substanţă albă, denumită centrul semioval. Este
formată din fibre nervoase: de asociere, comisurale şi de proiecţie.
- Fibrele de asociere leagă între ele puncte din aceeaşi emisferă cerebrală. Cele scurte
se întind între 2 giri. Fibrele mijlocii fac legătura între 2 lobi sau între 2 giri ceva mai
îndepărtaţi, iar cele lungi leagă regiuni îndepărtate ale scoarţei ce de exemplu, fasciculul
longitudinal superior care uneşte scoarţa lobului frontal cu cea a lobilor parietal şi occipital.
Fornixul sau trigonul este formaţiunea care în majoritate este constituită din fibre asociative
între hipocamp sau cornul lui Ammon şi corpul mamilar de aceeaşi parte. Fornixul este aşezat
sub corpul calos şi între aceste 2 formaţiuni, anterior, pe linia mediană se găseşte o membrană
denumită septum pellucidum care desparte cei 2 ventriculi laterali. El are un corp central
triunghiular cu vârful anterior, din dreptul căruia pleacă spre corpii amilari 2 tracturi numite
columnele fornixului. Din unghiurile posterioare se desprind alte 2 tracturi numite picioarele
fornixului care se îndreaptă spre hipocamp. Prin intermediul picioarelor şi a părţii posterioare a
corpului fornixului trec şi fibre comisurale între cei 2 hipocampi (comisura interamoniană sau
comisura fornicis).

- Fibrele comisurale sunt acelea care leagă între ele cele 2 emisfere cerebrale, realizând
funcţia lor simultană. Ca formaţiuni comisurale se descriu corpul calos, comisura albă
anterioară şi comisura fornicis.
Corpul calos este un sistem de fibre care în interiorul emisferelor formează tavanul
ventriculilor laterali. Pe o secţiune medio-sagitală este ca o lamă care descrie un arc cu
concavitatea inferioară. I se descriu o prelungire anterioară sau rostrum care schimbându-şi
direcţia, formează genunchiul corpului calos, după care urmează corpul corpului calos care la
extremitatea posterioară este mai îngroşată şi se numeşte splenium. Comisura albă anterioară
este subţire şi leagă între ei bulbii olfactivi şi scoarţa lobului temporal.
În interiorul substanţei albe se găsesc ventriculii laterali I şi II, cavităţi pline cu lichid
cefalorahidian. Ventriculii laterali sunt separaţi între ei prin septum pellucidium şi comunică
prin găurile interventriculare (Monroe) cu ventriculul III. Tavanul ventriculilor laterali este
format de corpul calos. Fiecare ventricul are câte o prelungire în lobul frontal, occipital şi
temporal denumite respectiv, corn anterior, posterior şi inferior. {i la nivelul acestor ventriculi
există o pânză coroidiană ca va produce lichidul cefalorahidian din interiorul ventriculului.
Sistemul limbic (Rinecefalul).
Este partea cea mai veche ca apariţie filogenetică a emisferelor cerebrale, îndeplinind la
animalele inferioare funcţii olfactive. Este alcătuit din bulbul olfactiv aşezat în şanţul omonim
al lobului orbital. În bulbii olfactivi se termină nervii olfactivi. Bulbul olfactiv se continuă cu
tractul olfactiv care merge posterior până la trigonul şi tuberculul olfactiv. Aceste formaţiuni se
leagă cu centrii corticali ai rinencefalului: aria parolfactorie, nucelii septali, substanţa perforată
anterioară, nucleul amigdalian. Scoarţa cerebrală a acestor centrii este reprezentată de
allocortex.
Sistemul cavitar al nevraxului.
În interiorul sistemul nervos central se găsesc o serie de canale sau regiuni mai dilatate
numite ventriculi, care sunt derivate din lumenul tubului neural primitiv. În interiorul măduvei
spinării, în grosimea comisurii cenuşii se găseşte canalul central. Acesta are un diametru de
cca. 1-2 mm, putând însă să fie astupat pe alocuri. În dreptul conului medular prezintă o mică
dilataţie numită ventricul terminal. În partea superioară, canalul central se continuă cu
ventriculul IV situat între bulb şi punte care-i formează podeaua şi cerebel care-i formează
tavanul.

Formaţiunile care-l delimitează au fost descrise la trunchiul cerebral. La unghiul său
anterior, ventriculul al IV-lea se continuă cu apeductul lui Sylvius săpat în grosimea
mezencefalului la limita dintre tegment şi tectum. La capătul său anterior apeductul se deschide
într-o nouă dilataţie situată în diencefal, numită ventriculul III (fig. 14).
Figura 14. Ventriculii encefalului - vedere sagitală
Prin găurile interventriculare (Monroe) situate anterior şi lateral se face comunicarea
între ventriculul III şi ventriculii laterali I şi II din interiorul emisferelor cerebrale şi care au
fost descrişi odată cu substanţa albă a emisferelor. În sistemul cavitar al nevraxului ca şi în
spaţiul subarahnoidian se găseşte un lichid incolor, lichidul cefalorahidian (LCR). Acesta este
secretat incontinuu de pânza coroidiană a fiecărui ventricul. Plexurile coroide sunt franjuri
vasculare ale vaselor pieimater care proemină în interiorul ventriculilor, dar rămân separate de
cavitatea ventriculară printr-o lamă epitelială de natură ependimară (interventriculare şi
extraventriculare).
Din ventriculul I şi II, LCR trece în ventriculul III unde se adaugă cel secretat aici şi
prin apeductul lui Sylvius dernează mai departe în ventriculul IV. De aici prin găurile laterale

(Luschka) şi mediană (Megendie) din membrana tectria a acestui ventricul, LCR trece în
spaţiul subarahnoidian la nivelul cisternei medulocerebeloase. Din spaţiul subarahnoidian este
incontinuu resorbit şi trecut în circulaţia venoasă de către granulaţiile arahnoidale (Pachioni).
El se mai absoarbe şi la nivelul tecilor durale ale nervilor spinali. Dacă congenital sau în urma
unei meningite la copii se astupă parţial găurile din tavanul ventriculului IV, LCR se
acumulează în sistemul cavitar destinde encefalul şi apare hidrocefalia internă. Când la copii
posibilităţile de resorbţie sunt afectate poate apare o hidrocefalie externă. LCR este deci, într-o
continuă reînnoire.
Meningele.
Mengele sunt 3 membrane conjunctive de protecţie, care învelesc măduva spinării şi
encefalul. Meningele spinal se continuă la nivelul găurii occipitale cu meningele cerebrale.
Membrana exteriară denumită duramater (lat. dura = tare + mater = mamă), formată din ţesut
fibros şi elastic dens, este mai groasă. Arahnoida (gr. Arachne = păianjen + eidos = aspect)
situată sub duramater, împreună cu piamater (lat. pia = bună + mater = mamă) care înveleşte la
suprafaţă măduva spinării şi encefalul sunt mai subţiri.
Duramater spinală este despărţită de peretele osteofibros al canalului vertebral printr-un
spaţiu plin cu ţesut adipos şi numeroase vene, denumit spaţiul extradural sau epidural. Acest
spaţiu este străbătut de rădăcinile nervilor spinali în jurul cărora duramater formează teci
radiculare, care se continuă la periferie cu perinervul. În acest spaţiu se fac anesteziile
rahidiene epidurale.
Duramater cerebrală, spre deosebire de cea spinală aderă strâns de oasele cutiei
craniene, formând periostul intern al acestor oase şi se continuă la nivelul suturilor cu periostul
extern. În interiorul cutiei craniene ea trimite o prelungire orizontală care separă emisferele
cerebrale de cerebel, denumită cortul cerebelului care se inseră pe buzele şanţului transvers şi
pe marginea superioară a stâncii temporalului.
De pe faţa inferioară a acestuia pleacă o mică prelungire sagitală, denumită coasa
cerebelului care proemină între emisferele cerebeloase. O altă prelungire a membranei
duramater, cu direcţia sagitală şi perpendiculară pe faţa superioară a cortului cerebelului este
coasa creierului, care părtunde în fisura interemisferică şi ajunge în profunzime, până la faţa
superioară a corpului calos. Aceasta desparte cele 2 emisfere cerebrale. O prelungire a
membranei duramater acoperă şaua turcească, formând loja glandei hipofize. Ea poartă

denumirea de diafragma şeii şi prezintă în centru un orificiu prin care trece tija glandei
hipofize.
În grosimea membranei duramater craniene sunt cuprinse şi sinusurile venoase ale
acesteia. Între duramater şi arahnoidă se află un mic spaţiu capilar, străbătut de tracturi fibroase
şi vase sanguine, denumit spaţiul subdural.
Piamater aderă de suprafaţa măduvei şi a encefalului şi este o membrană vasculară. Prin
ea şi prelungirile pe care le trimite, pătrund în sistemul nervos central, vasele sanguine. Între
piamater şi arahnoidă se află spaţiul subarahnoidian, plin cu lichid cafalorahidian.
Măduva spinării este ancorată de duramater în părţile laterale prin prelungiri ale
membranei piamater situate în plan frontal, între rădăcinile anterioare şi posterioare ale nervilor
spinali, denumite ligamente dinţate. Piamater cerecbrală trimite prelungiri bogate în vase
sanguine, în ventriculii cerebrali, care formează plexurile coroide ale ventriculilor, cu rol în
secreţia lichidului cefalorahidian. Spaţiul subarahnoidian cerebral este mult mai neregulat decât
cel spinal. Astfel, la baza encefalului el prezintă, în dreptul depresiunilor, lacuri pline de LCR
denumite cisterne aubarahnoide, cum sunt: cisterna pontină, cisterna interpedunculară,
cisterna chiasmatică, cisterna cerebelomedulară din dreptul găurii Megendie, cisterna şanţului
lateral şi cisterna corpului calos.

Unitatea de curs 7 Sistemul nervos periferic.
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului sistemului nervos periferic.
Obiectivele operaţionale
1. Prezentarea structurii sistemului nervos periferic
2. Prezentarea clasificării sistemului nervos periferic.
3. Prezentarea rolului sistemului nervos periferic
Sistemul nervos periferic este alcătuit din 12 perechi de nervi cranieni şi 31 de perechi
de nervi spinali.
Nervii cranieni.
Din punct de vedere funcţional pot fi:
-receptori: olfactivul (I); opticul (II); acusticul (VIII);
-motori: oculomotor comun (III); pateticul (IV); oculomotor extern (VI); spinal (XI);
hipoglosul (XII);
-micşti: trigemenul (V); facialul (VII); pneumogastricul (IX); glosofaringianul (X).
În afară de primele 2 perechi care sunt derivate ale veziculelor cerebrale, restul ies din
trunchiul cerebral şi au o origine reală şi alta aparentă dacă au fibre motorii şi o terminare
aparentă şi alta reală dacă au fibrele senzitive. Nervii senzitivi ca şi nervii micşti pentru fibrele
lor senzitive, vor avea pe traiectul lor cel puţin un ganglion unde se vor găsi protoneuronii
receptori ai sensibilităţii culeasă de nervul respectiv. Nucleii motori din trunchiul cerebral vor
fi originea reală, iar cei senzitivi vor reprezenta terminarea reală a nervilor cranieni.
Originea sau terminarea aparentă va fi locul unde nervii se desprind din trunchiul
cerebral.
I. Nervii olfactivi. Sunt reprezentaţi de un număr de 20-30 de filete nervoase formate
din axonii neuronilor din mucoasa olfactivă, care este singurul epiteliu senzorial propriu-zis,
adică un neuroepiteliu. Axonii se termină la celulele mitrale din bulbul olfactiv.
II. Nervul optic este alcătuit din axonii neuronilor multipolari din retină. Fibrele care
vin de la retină nazală se încrucişează cu cele simetrice formând chiasma optică de la care

pleacă apoi tracturile optice, care se termină în corpii geniculaţi laterali.
III. Nervul oculomotor are fibre somatomotorii cu originea reală în nucleul
oculomotorului din tegmentul mezencefalic în dreptul coliculilor optici sub apeductul lui
Sylvius. Originea aparentă este situată pe faţa medială a picioarelor pedunculilor cerebrali.
Ei inervează cea mai mare parte a muşchilor exterinseci ai globului ocular, respectiv:
muşchiul drept superior şi muşchiul ridicător al pleoapei superioare, muşchiul drept medial,
muşchiul drept inferior şi muşchiul oblic inferior. Nervul oculomotor are şi fibrele vegetative
parasimatice cu originea în nucleul accesor al oculomotorului (Edinger-Westphal), care sunt
iridoconstrictoare. Ele fac sinapsă în ganglionul ciliar ataşat ramurei oftalmice a trigemenului,
cu neuronul postganglionar.
IV. Nervul trohlear are fibrele somatomotorii cu originea reală în nucleul trohlearului
din tegmentul mezencefalic în dreptul coliculilor acustici. Originea aparentă este dorsal de
coliculilor acustici. Originea aparentă este dorsal de coliculii acustici, fiind singurul nerv
cranian care se încrucişează în SNC şi iese de pe faţa dorsală a trunchiului cerebral. Prin fisura
orbitală superioară pătrunde în orbită şi inervează muşchiul oblic superior.
V. Nervul trigemen este nerv mixt. Pe traiectul lui se află ganglionul semilunar (Gasser)
de la care pleacă cele 3 ramuri ale lui: oftalmicul, maxilarul şi mandibularul, care se distribuie
în teritoriile feţei corespunzătoare denumirii lor.
- Fibrele motorii pleacă din nucleul branhimotor din tegumentul pontin şi merg prin
ramura mendibulară la muşchii masticatori, muşchiul tensor al vălului palatin şi la muşchiul
tensor timpanic.
- Culege sensibilitatea exteroceptivă de la pielea feţei şi de la mucoasele cavităţilor feţei
prin toate cele 3 ramuri. Protoneuronul este situat în ganglionul Gasser, iar deutoneuronul în
nucleii senzitivi din bulb şi punte.
- Culege sensibilitatea propioceptivă de la aparatul masticator prin ramura mandibulară.
Protoneuronii sunt situaţi în ganglionul Gasser, iar axonii lor merg la nucleul tractului
mezencefalic al trigemenului unde sunt deutoneuronii.
Pe ramurile trigemenului se găsesc ganglionii vegetativi parasimpatici şi fibrele
vegetative ale parasimpaticului cranian.
VI. Nervul abducens este motor pentru muşchiul drept extern al globului ocular.
Originea reală a fibrelor este în nucleul somatomotor al abducensului din dreptul coliculului

facialului (eminenţa rotundă) din fosa romboidă. Originea aparentă este din spaţiul bulbopontin
în dreptul piramidelor.
VII. Nervul facial este nerv mixt şi pe traiectul lui are ganglionul geniculat:
– fibrele motorii pleacă din nucleul branhiomotor din tegmenul pontin şi merg la
muşchii mimicii, muşchiul stilohiodian şi la muşchiul scăriţei;
- fibrele senzitive culeg gustul de la 2/3 anterioare ale limbii. Protoneuronul este situat
în ganglionul geniculat. Axonii se duc la nucleul tractului solitar (din bulb);
- fibrele visceromotorii (vegetative) pentru glanda lacrimală şi glandele mucoasei
nazale pleacă din nucleul lacrimonazal, iar cele pentru glandele submandibulară şi sublinguală
sunt situaţi în tegmentul pontin.
Nervul se desprinde la nivelul şanţului bulbopontin în dreptul fosetei supraretroolivare.
Fibrele vegetative şi senzoriale sunt descrise de unii autori ca nervul VII bis (nervul
intermediar Wriesberg).
VIII. Nervul acusticovestibular este senzorial. El are pe traiectul său 2 ganglioni: Corti
şi Scarpa.
- culege excitaţiile acustice de la organul lui Corti prin dendritele neuronilor situaţi în
ganglionul spinal Corti (protoneuronul). Axonii lor fac ramura cochleară şi merg să se termine
în nucleii cochleari din punte (deutoneuronul);
- culege excitaţiile de echilibru de la receptorii vestibulari. Protoneuronul se află în
ganglionul Scarpa, iar axonii lor fac sinapsă ramura vestibulară a perechii a VIII-a care se
termină în nucelii vestibululari din dreptul unghiului lateral al fosei romboide. Nervul se
termină în şanţul bulbopontin (originea aparentă) în foseta supraretroolivară, lateral de nervul
facial.
IX. Nervul glosofaringian este mixt şi pe traiectul lui se găsesc 2 ganglioni: intra şi
extracranian.
– fibrele motorii pentru muşchii faringelui pleacă din nucleul ambiguu din bulb
(originea reală);
- fibrele senzoriale ale protoneuronului din ganglionul extracranian culeg gustul de la
"V-ul” lingual şi de la o îngustă zonă posterioară acestuia;
- fibrele interoceptive de la sinusul carotidian îşi au protoneuronl în ganglionul
extracranian şi ca şi cele gustative se duc în bulb la nucleul tractului solitar;

- culege sensibilitatea exteroceptoare de la 1/3 posterioară a limbii şi sensibilitatea
faringelui. Protoneuronul este în ganglionul intracranian, iar axonul se duce la nucleii
exteroceptivi ai trigemenului din punte şi bulb (deutoneuronul);
- fibrele visceromotorii (vegetative) pleacă de la nucleul salivator inferior din bulb şi
determină secreţia parotidei.
Nervul glosofaringian se desprinde din dreptul şanţului lateral posterior al bulbului
(originea aparentă).
X. Nervul vag este nerv mixt şi pe traiectul lui se găsesc 2 ganglioni: jugular (superior)
şi nodos (inferior). Are cel mai mare teritoriu de inervaţie viscerală, distribuindu-se în torace şi
apoi la nivelul abdomenului unde ajunge până la unghiul splenic al colonului:
– fibrele motorii pentru o parte din muşchii faringelui şi pentru cei ai laringelui şi ai
esofagului superior îşi au originea în nucleul ambiguu;
- fibrele senzoriale ale neuronilor din ganglionul nodos culege gustul din regiunea
posterioară a limbei şi de la valecule (cele 2 fosete gloso-epiglotice);
- fibrele interoceptive de la zonele reflexogene cardiovasculare de la aparatul respirator,
de la tubul digestiv până la unghiul splenic, şi de rinichi şi căile urinare până la ureter inclusiv,
îşi au originea în protoneuronul din ganglionul nodos. Ca şi cele de gust ele se duc în bulb la
nucleul tractului solitar;
- culege sensibilitatea exteroceptivă de la pielea conductului auditiv extern.
Protoneuronul este în ganglionul jugular, iar axonii se duc la nucleii exteroceptivi ai
trigemenului din punte şi bulb;
- fibrele visceromotorii pleacă de la nucleul dorsal al vagului din bulb (coloana visceral
eferentă generală). Acest nucleul, datorită teritoriului său de distrtibuţie este denumit şi centrul
cardio-pneumo-enteric şi asigură inervaţia parasimpatică a organelor respective.
Nervul vag se desprinde din dreptul şanţului lateral posterior al bulbului (originea
aparentă), sub glosofaringian şi deasupra accesorului.
XI. Nervul accesor este motor. El are o dubllă origine: bulbară şi medulară. După
unirea celor 2 rădăcini, el se împarte într-un ram intern şi altul extern:
- fibrele care provin din nucleul ambiguu vor părăsi nervul prin ramul intern care se
alătură nervului vag şi ajung la laringe prin nervul laringean inferior;

- fibrele din nucleul spinal (medular) merg prin ramul extern la muşchii trapez şi
sternocleidomastoidian.
XII. Nervul hipoglos este motor pentru muşchii limbii. Originea reală este în nucleul
somatomotor din fosa romboidă în dreptul aripii albe interne (trigonul hipoglosului). Originea
aparentă este în şanţul preolivar.
Nervii spinali.
Nervii spinali sunt 31 de perechi metamerice care se subîmpart în: 8 perechi cervivali,
12 perechi toracali, 5 perechi lombari, 5 perechi sacrali şi 1 nerv coccigian pereche.
Fiecare nerv spinal este un nerv mixt, ce se formează prin unirea în apropierea găurii de
conjugare a rădăcinii ventrale cu rădăcinea dorsală. Pe traiectul ădăcinii posterioare se află
ganglionul spinal unde sunt situaţi protoneuronii receptori pe calea extero, proprio şi
interoceptivă. Axonii acestor neuroni pseudounipolari merg prin rădăcina posterioară şi intră
prin şanţul lateral posterior la nucleii senzitivi din nevrax (fig.15).
Funcţional rădăcina posterioară este senzitivă, aferentă, iar cea anterioară eferentă sau
motorie, deşi s-au descris fibre, care fac excepţie de la regulă. Rădăcina anterioară este formată
din axonii neuronilor somato şi visceromotorii din măduvă, care ies prin şanţul lateral anterior
şi merg la organele efectoare. Trunchiul comun, format prin unirea celor 2 rădăcini este foarte
scurt şi imediat ce iese prin gaura intervertebrală se împarte în 4 ramuri periferice.
- Ramura meningeală care reintră în canalul vertebral prin gaura intervertebrală
asigurând sensibilitatea meningelor. Tot pe ea merg şi fibrele vasomotorii.
- Ramura comunicantă conduce fibrele vegetative. Poate fi albă şi cenuşie. Cele albe
conţin fibre preganglionare simpatice şi fibre aferente vegetative. Ramura comunicantă cenuşie
are fibre postganglionare ce vin la nervul spinal din lanţul simpatic.

Figura 15. Schema structurii nervului spinal (rahidian)
- Ramura posterioară. Aceste ramuri nu se anastomozează între ele păstrându-si
dispoziţia metamerică, segmentară. Ramurile posterioare prin fibrele lor senzitive culeg
exteroceptivitatea de la pielea cefei şi a spatelui în interiorul ariei delimitată de linia parieto-
acromio-trochantero-coccigiană. Prin fibrele lor motorii inervează musculatura autohtonă a
spatelui. Ceilalţi muşchi ai spatelui care deservesc membrele sunt inervaţi de ramurile
anterioare ale nervului spinal. Primul nerv cervical nu are fibre senzitive, iar fibrele celui de-al
doilea nerv, alcătuiesc marele nerv occipital.
- Rădăcina anterioară. Ramurile ventrale se distribuie cutanat şi motor la peretele
antero-lateral al trunchiului şi la membre. Dintre ele, îşi păstrează dispoziţia segmentară numai
cele ale nervilor toracici, care formează nervii intercostali. Celelalte ramuri se anastomozează
între ele şi formează 5 plexuri nervoase: cervical, brahial, lombar, sacral şi coccigian.
Plexul cervical.
Este format de anastomozele ramurilor anterioare ale primilor 4 nervi cervicali spinali
(C1, 2, 3, 4.). De la el pronesc anastomoze pentru nervul hipoglos (XII) şi nervul accesor (XI).

Acest plex dă 2 feluri de ramuri: ramuri profunde care formează plexul cervical profund;
ramuri cutanate (superficiale) care formează plexul cervical superficial.
- Ramurile profunde sunt ramuri motorii. Ele alcătuiesc ramurile musculare şi
inervează muşchii gâtului (muşchiul lung al capului, muşchiul lung al gâtului, muşchiul scalen
mediu, muşchiul sternocleidomatsoidian) şi unii muşchi ai spatelui (trapezul, ridicătorul
scapulei).
Ansa hipoglosului este formată dintr-o ramură descendentă laterală (ramura nervului
hipoglos, provenită din C1) şi o ramură descendetă medială, provenită din C2 şi C3. Ea
inervează muşchii subhioidieni (omohiodianul, sternohioidianul şi sternotiroidianul).
Nervul frenic este format din fibre provenite din C3,C 4, şi C5 şi coboară prin gât, apoi
prin torace şi traversează diafragmul, prin orificiul venei cave inferioare şi orificiul esofagian.
Nervul frenic dă ramuri pentru diafragm şi alte ramuri frenico-abdominale traversează
diafragmul prin orificiile amintite, se anastomozează cu ramuri ale nervilor intercostali şi
împreună cu fibre simpatice alcătuiesc plexul diafragmatic, situat pe faţa inferioară a
diafragmului.
- Ramurile cutanate sunt ramuri senzitive şi se împart în 2 grupuri: un grup de 3 nervi
şi alt grup format care formează un trunchi.
Primul grup se compune din:
- nervul occipital mic, care inervează pielea din regiunea parietală şi occipitală a
capului;
- nervul auricular mare, care inervează pielea de pe feţele anterioară şi posterioară ale
pavilionului urechii;
- nervul cutanat al gâtului, care inervează pielea din regiunile suprahioidiană,
subhioidiană şi cervicală.
Al doilea grup formează un trunchi ce se divide în 3 ramuri: ramura suprasterală,
ramura supraclaviculară şi ramura supraacromială care inervează pielea din regiunea antero-
superioară a toracelui şi antero-laterală a umărului.
Plexul brahial.
Este format prin anastomoza ramurilor anterioare ale ultimilor 4 perechi de nervi
cervicali (C5 ,C 6, ,C7, ,C8 ) şi ale primei perechi de nervi toracali (T1). El prezintă o porţiune

supraclaviculară şi o porţiune infraclaviculară (axilară) şi este format din 3 trunchiuri primare:
superior (C5 -C 6, ), mijlociu (C7 ) şi inferior (C8 ,T1,), care dau fiecare ramuri ventrale şi dorsale.
Ramurile dorsale ale celor 3 trunchiuri formează fasciculul posterior al plexului
brahial.
Ramurile ventrale ale trunchiului primar, superior şi mijlociu, formează fasciculul
lateral, iar ramura ventrală a trunchiului inferior, fasciculul medial al plexului brahial.
Ramurile colaterale ale plexului brahial sunt: nervul dorsal al scapulei, nervul toracic
lung, nervul subscapular, nervul subclavicular, nervul pectoral medial şi lateral, nervul
suprascapular şi nervul toracodorsal.
Ramurile terminale ale plexului brahial sunt: nervul median, care ia naştere prin 2
rădăcini (una din fasciculul medial şi alta din fasciculul lateral), nervul musculo-cutanat (din
fasciculul lateral), nervul ulnar, nervul cutanat brahial medial, nervul cutanat antibrahial medial
(din fasciculul medial), nervul radial şi axilar (din fasciculul posterior).
Plexul brahial inervează, prin ramurile colaterale şi terminale tegumentul umărului şi al
membrului superior, muşchii centrurii scapulare şi ai extremităţii libere al membrului superior.
Nervii intercostali.
În regiunea toracală, ramurile anterioare ale nervilor spinali nu formează plexuri. Sunt
12 perechi de nervi toracali, dintre care numai 10 (T2-T11) alcătuiesc nervii intercostali,
deoarece T1 participă la formarea plexului brahial, iar T12 la formarea plexului lombar. Nervii
intercostali formează ramuri intercostale şi ramuri cutanate. Ei străbat spaţiile intercostale
mergând împreună cu vasele omonime în şanţul costal. Inervează muşchii peretelui toracic şi al
jumătăţii superioare a peretelui abdominal, ca şi pielea aceleiaşi regiuni prin ramurile lor
cutanate. Pentru abdomen merg ultimii 6 nervi intercostali.
Plexul lombar.
Este format din anastomozele ramurilor anterioare ale T12 şi ale primilor 4 nervi
lombari (L1, L2, L3, şi parţial L4). Ramurile plexului lombar sunt ramuri colaterale (nervul
patratul lombelor şi nervul psoas) şi terminale.
Ramurile colaterale formează:
- nervul iliohipogastric prin ramura muscuară inervează muşchii laţi ai abdomenului şi
drepţi abdominali; o altă ramură este cutanată şi inervează pielea din regiunea şoldurilor, din
regiunea pubiană şi a scrotului la bărbat şi a labiilor mari la femeie;

- nervul ilioinghinal inervează pielea scortului la bărbat sau a labiilor mari la femei;
- nervul femurocutanat lateral inervează pielea regiunii fesiere şi pielea regiunii laterale
a coapsei;
- nervul genitofemural inervează pielea feţei mediale a coapsei şi pielea scrotului sau a
labiilor mari.
Ramurile terminale formează:
- nervul obturator care are o ramură anterioară şi alta posterioară; ramura anterioară
inervează muşchiul obturator extern, muşchiul pectineu, muşchii adductor lung şi scurt,
precum şi pielea din regiunea genunchiului şi a feţei mediale a coapsei; ramura posterioară
inervează muşchiul adductor mare şi articulaţia coxofemurală.
- nervul femural este cel mai voluminos nerv al plexului lombar; în cavitatea pelviană
dă ramuri pentru muşchiul psoas iliac, muşchiul pectineu şi formează ramuri pentru muşchii
din loja antero-medială a coapsei, ca şi pielea acestei regiuni.
Plexul sacral.
Rezultă din anastomozele ramurilor anterioare ale trunchiului lombosacral (anastomoză
L4-parţial L5) şi ale primilor 3 nervi sacraţi (S1, S2, S3). Este cel mai voluminos plex şi se află
situat în bazin. Are forma unui triunghi a cărui baze corespunde găurilor sacrale anterioare, iar
vârful situat în scobitura sciatică (incizura ischiatică) a osului coxal.
Ramurile plexului sacral sunt: nervul fesier superior, nervul fesier inferior, nervul
cutanat femural posterior (micul sciatic), nervul ridicătorului anal, nervul obturatorului,
gemenului superior şi inferior, piramidalului şi nervul ischiatic (marele sciatic), cel mai
voluminos nerv din organism.
Ramurile terminale ale marelui sciatic sunt: nervul peronier comun şi nervul tibial.
Nervul peronier comun dă nervul cutanat sural lateral (ca ramură colaterală) şi ca
ramuri terminale nervul peronier superficial (nervul musculo-cutanat) şi nervul peronier
profund (nervul tibial anterior).
Nervul tibial dă nervul cutanat sural medial (nervul safen extern) ca ramuri colaterale
musculare şi ca ramuri terminale nervul plantar medial şi nervul plantar lateral.
Nervul marele sciatic provine deci din toate componentele plexului sacral şi părăseşte
bazinul prin gaura ischiatică, inervând tegumentul regiunii fesiere (neinervat de plexul lombar)

şi muşchii din grupul posterior al coapsei, precum şi muşchii gambei şi ai piciorului (muşchiul
biceps femural, muşchiul semitendinos, semimembranos şi aductorul mare).
Plexul ruşinos
Rezultă din anastomozele ramurilor anterioare ale celui de-al patrulea nerv sacral (S4);
el este o anexă a plexului sacral cu care se leagă printr-o ramură a nervului S3, iar cu plexul
următor, coccigian prin unirea unei ramuri a nervului S4, cu o ramură a nervului S5.
Acest plex conţine întreg parasimpaticul pelvian, având ramuri musculare şi viscerale
(motorii şi senzitive).
Prin ramurile musculare inervează muşchii ridicători anali şi ischiococcigianul.
Prin ramurile viscerale formează nervii rectali inferiori (nervi hemoroidali inferiori),
nervii perineali, nervii scrotali posteriori, nervii labiali posteriori, nervul dorsal al penisului şi
nervul dorsal al clitorisului.
Plexul coccgian.
Ia naştere din anastomozele ramurilor anterioare ale lui S5 cu nervul coccigia. Ramurile
cutanate inervează muşchiul ischiococcigian şi pielea cuprinsă între vârful coccisului şi
orificiul anal. Ramurile viscerale intră în alcătuirea plexului hipogastric.

Unitatea de curs 8 Sistemul nervos vegetativ
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului sistemului nervos vegetativ.
Obiectivele operaţionale
4. Prezentarea structurii sistemului nervos vegetativ
5. Prezentarea clasificării sistemului nervos vegetativ.
6. Prezentarea rolului şi căilor de conducere ale sistemului nervos vegetativ
Sistemul nervos vegetativ (SNV) integrează şi coordonează, în strânsă legătură cu
SNC, funcţiile viscerale. El este alcătuit din centrii nervoşi vegetativi cu rol de integrare, din
căi nervoase aferente care conduc mesajele spre centrii nervoşi şi din căile eferente care
transmit organelor efectoare, comenzile centrilor.
În cadrul SNV se disting o parte simpatică şi alta parasimpatică cu acţiune antagonistă
la nivelul organelor efectoare, în sensul că acolo unde simpaticul este excitat, parasimaticul
este inhibitor şi invers. Baza structurală şi funcţională a SNV o constituie arcurile reflexe
vegetative.
A. Calea aferentă.
Pentru viscerele pelvine colonul stâng o parte din organele retro-peritoneale şi din
mediastinul posterior, ca şi pentru interceptorii din muşchi, protoneuronul receptor este
reprezentat de neuronul pseudounipolar situat lanivelul genglionului spinal. Se mai găsesc însă
neuroni receptori la nivelul lanţului simpatic, a plexurilor prevertebrale şi chiar intramurali.
Cea mai mare parte a viscerelor toracice, a celor abdominale incluzând tubul digestiv
până la unghiul splenic, ca şi a zonelor reflexogene vasculare au sensibilitatea recepţionată de
nervul vag. Protoneuronul este situat în ganglionul nodos, iar axonul lui se duce nucleul
tractului solitar. De la sinusul carotidian, excitaţiile sunt culese de neuroni din ganglionul
extracranian al glosofaringianului şi conduse tot la nucleul tractului solitar.
B. Centrii vegetativi.
În ordinea apariţiei lor filogenetice se disting centrii nervoşi inferiori în trunchi şi
măduvă, centrii supraetajaţi subcorticali în hipotalamus şi centrii vegetativi corticali.

- Centrii vegetativi inferiori se numesc şi centrii preganglionari. De la ei pleacă spre
periferie calea eferentă. Se disting centrii parasimpatici în trunchiul cerebral şi în măduva
sacrată şi centrii simpatici în măduva toracolombară.
- Centrii vegetativi subcorticali sunt situaţi în hipotalamus. Cei parasimpatici la nivelul
regiunii lui antero-laterale, iar cei simpatici în partea posterioară. Hipotalamusul este
principalul centru subcortical de reglare a activităţii simpaticului şi parasimpaticului. El este
informat de la centrii inferiori prin căi ascendente nespecifice, multineuronale, iar influenţa lui
se exercită prin căile descendente ca fasciculul dorsal Schütz şi prin substanţa reticulată.
Proiectează informaţiile directe pe scoarţa cerebrală sau prin intermediul nucleilor rostrali
talamici şi este controlat de cortexul cerebral prin căi descendente corticohipotalamice.
- Centrii vegetativi corticali au fost menţionaţi mai sus. Se descriu cu funcţii viscerale
câmpurile corticale 13, 14, 24, 25, 32, de pe feţele orbitală şi medială ale lobului frontal ca şi
cornul lui Ammon. Astăzi se crede că în scoarţa cerebrală există centrii în raport cu inervaţia
simpatică şi parasimpatică.
C. Calea eferentă.
Calea eferentă a reflexului vegetativ spre deosebire de cea a reflexului somatic este
formată din 2 neuroni: unul situat în centrul vegetativ preganglionar din măduvă sau trunghiul
cerebral, a cărui prelungire formează fibra preganglionară care face sinapsă cu al doilea neuron
numit neuron postganglionar, situat într-un ganglion periferic.
Fibra postganglionară merge la organul efector pe calea nervilor somatici, prin nervii
vegetativi proprii sau pe calea vaselor sanguine cu plexuri perivasculare. Neuronii
postganglionari simpatici sunt aşezaţi în apropierea nervraxului, în lanţul latero-vertebral sau
într-un plex prevertebral, pe când cei parasimatici se află cel mai frecvent în imediata apropiere
sau chiar în peretele viscerului respectiv (intramurali).
Sistemul nervos simpatic.
Centrii preganglionari simpatici se întind între mielomerele C8-L2, la nivelul cornului
lateral, unde formează nucelul intermedio-lateral. Începând cu centrul ciliospinal de la C8-L2,
care produce iriodilataţie se înşiră toţi centrii simpatici în succesiunea lor firească, terminându-
se cu centrul vezicospinal la L1 şi ano-raectal la L2.

Fibra preganglionară merge prin cornul anterior, rădăcina anterioară a nervului spinal,
nervul spinal, ramura comunicantă albă şi ajunge la lanţul ganglionar latero-vertebral. Dacă
fibra se distribuie la viscerele supradiafragmatice sau la partea somatică, face sinapsă în lanţul
simpatic. Dacă este pentru viscerele subdiafragmatice trece fără oprire în nervii splahnici
abdominali şi pelvini către plexurile prevertebrale.
Nervul postganglionar este situat aşa cum se poate deduce după comportarea fibrei
preganglionare fie în lanţul latero-vertebral simpatic, fie în plexurile prevertebrale.
Fibra postganglionară se comportă diferit după organul efector la care se distribuie.
Cele pentru partea somatică merg prin ramura comunicantă cenuşie înapoi la nervul spinal şi
cu ramificaţiile acestuia se distribuie la periferie.
Pentru viscerele supradiafragmatice părăsesc lanţul simpatic ca nervii vegetativi prorpii
(ex. nervii cardiaci) sau ca plexuri perivasculare (plexul subclavicular, plex vertebral,
carotidian etc.). Desigur că la viscerele corpului va ajunge ca plexuri perivasculare cu ramurile
carotidelor. Pentru viscerele subdiafragmatice pleacă de la plexurile prevertebrale celiac şi
hipogastric ca plexuri perivasculare şi cu arterele care irigă viscerul respectiv ajung la
destinaţie.
A. Lanţul simpatic.
Lanţul ganglionar simpatic latero-vertebral este alcătuit din 22-23 ganglioni alungiţi,
uniţi prin cordoane de substanţă albă ca un şirag. În ganglioni se găsesc neuroni
postganglionari pentru partea somatică şi viscerele supradiafragmatice ca şi neuronii receptori.
Aferenţele lui sunt reprezentate de fibrele senzitive de la viscere şi de ramuri
comunicante albe. Eferenţele pleacă prin ramul comunicant cenuşiu şi prin nervii splanhnici.
Din punct de vedere funcţional i se descriu mai multe porţiuni:
-partea cervicală alcătuită din 3 ganglioni, superior, mijlociu şi inferior. Fibrele lor
eferente se distribuie la viscerele capului, gâtului şi ale mediastinului anterior. Ultimile
realizează un plex cardiac la care se adaugă şi o componentă parasimatică vagală.
-partea toracică este alcătuită din 10-11 ganglioni. Primii 5 ganglioni toracici trimit
eferenţe la viscerele toracice din mediastinul posterior formând plexurile pulmonar, esofagian
şi aortic. Ultimii 6 dau fibre abdominale sub forma nervului mare şi mic splanhnic abdominal
ce merg spre plexul celiac.

-partea lombară este formată din 3-4 ganglioni ale căror eferenţe merg la plexul aortic
şi apoi cu ramurile aortei ajung ca plexurile perivasculare la viscere.
-partea pelvină este formată din 3-4 ganglioni. Nervii splanhnici sacrali, merg în plexul
hipogastric şi de aici cu vasele laperiferie.
B. Plexurile prevertebrale.
La nivelul aortei abdominale se descrie în partea superioară plexul celiac, iar inferior şi
în continuarea lui plexul hipogastric.
Plexul celiac este alcătuit din ganglionii celiaci, mezenteric superior, aortico-renali şi
mezenteric inferior legaţi între ei prin cordoane nervoase. Sunt formaţi din neuronii
postganglionari simpatici şi din neuronii receptori. Aferenţele sunt reprezentate de fibrele
preganglionare din marele şi micul nerv splanhnic abdominal, de fibre senzitive, dar şi din
fibre preganglionare parasimpatice vagale ce trec fără oprire spre a face sinapsă cu neuronul
postganglionar care este situat intramural. Eferenţele merg ca plexuri parietale spre viscere
fiind alcătuite din fibre simpatice postganglionare şi parasimpatice preganglionare.
Plexul hipogastric are componenta simpatică adusă de nervii splanhnici pelvini
(erectori). {i aici fibrele parasimpatice trec prin plex fără oprire, iar cele ale splanhnicilor
simpatici sacrali fac sinapsă cu neuronul postganglionar. Calea spre viscere este tot ca plex
imtramural.Sistemul nervos parasimpatic.
După situaţia neuronului preganglionar se descrie un parasimpatic cranian şi altul
sacrat.
A. Parasimaticul cranian.
Centrii preganglionari sunt reprezentaţi de nucleii din trunchiul cerebral: nucleul
accesor al nervului oculomotor (Eddinger-Westphal) din mezencefal, nucleii lacrimali şi
salivator superior din tegmentul pontin, nucleul salivator inferior şi nucleul dorsal al vagului
din bulb.
Fibrele preganglionare merg pe calea nervilor cranieni formând aşa-zisele cabluri
vegetative. De exemplu de la nucleul Eddinger-Westphal merg pe calea oculomotorului; de la
nucelul salivator superior spre glandele submandibulară şi sublinguală pe calea nervului facial,
coarda timpanului şi apoi ramul mandibular al trigemenului, iar de la nucleul salivator inferior
pe calea nervului glosofaringian.

Nervii postganglionari sunt situaţi în nişte ganglioni de lângă nervul oftalmic în orbită;
ganglionul pterigopalatin lângă nervul maxilar şi ganglionul otic şi submandibular alăturaţi
mandibularului.
Fibrele postganglionare merg fie ca nervi proprii, fie sub forma unor fascicule
vegetative spre organele efectoare.
Vagul are o situaţie deosebită deoarece fibrele lui preganglionare merg să facă sinapsă
cu neuronii postganglionari situaţi în imediata vecinătate a viscerelor inervate de el sau chiar în
pereţii acestora.
B. Parasimpaticul sacrat are centrii preganglionari în nucleii micţiunii, erecţiei şi
defecaţiei din măduva sacrată. Fibrele preganglionare trec prin cornul anterior, rădăcina
anterioară, nervii sacrali şi se desprind din aceştia din urmă dub numele de nervi errectori
(nervul splachnici pelvini), care se duc la plexul hipogastric. În continuare la plexuri
perivasculare (perirectale, perivezicale etc.), împreună cu fibrele postganglionare simpatice
ajung la rect în vezică, vasele peniene etc.

Unitatea de curs 9 Analizatorii
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului analizatorilor.
Obiectivele operaţionale
7. Prezentarea structurii analizatorilor
8. Prezentarea clasificării analizatorilor.
9. Prezentarea rolului şi căilor de conducere ale analizatorilor
Analizatorii sunt formaţiuni anatomo-funcţionale prin care sistemul nervos
recepţionează informaţiile din mediul înconjurător sau din interiorul organismului şi le
integrează în centrii nervoşi, transformându-le în senzaţii. Analizatorii reprezintă canalele
informaţionale ale sistemului nervos şi anatomic. Se compun din 3 părţi:
- segmentul periferic sau organul de simţ propriu-zis, care recepţionează stimulii;
- segmentul intermediar cu rol de conducere a excitaţiilor spre centrii nervoşi;
- segmentul central reprezentat de centrul nervos cortical care integrează informaţiile şi le
transformă în senzaţii.
Segmentul periferic. Receptorii periferici percep stimulii fizici sau chimici din mediul
intern sau extern şi îi transformă în impuls nervos. După locul de unde preiau informaţiile se
disting trei categorii de receptori: exteroceptori, proprioceptori şi interoceptori.
- Exteroceptorii culeg stimulii din mediul extern şi se împart în: exteroceptorii de contact
(receptorii cutanaţi tactili, termici, de presiune, durere, olfactivi, gustativi), exteroceptorii de la
distanţă sau telereceptori (receptorul vizual şi receptorul acustic).
- Proprioceptorii sau receptorii profunzi ai aparatului locomotor, sunt reprezentaţi de fusurile
neuromusculare, corpusculii tendinoşi Golgii, corpusculii Vater-Pacini; ei dau informaţii
privind poziţia corpului sau a segmentelor lui în repaus şi în mişcare.
- Interoceptorii (visceroceptori) culeg stimulii de la organele interne (viscere) şi sunt
reprezentaţi de terminaţii nervoase intraepiteliale, din mucoasa organelor cavitare, de
terminaţiile nervoase difuze din seroase şi vase sanguine, de corpusculii Vater-Pacini din
viscere, de corpusculii genitali din organele genitale externe.

Segmentul intermediar (de conducere) este format din căile nervoase prin care influxul
nervos este transmis la scoarţa cerebrală. Căile ascendente sunt directe şi indirecte. Căile de
conducere directe, cu sinapse puţine, conduc rapid impulsurile ce sunt proiectate pe o arie
corticală specifică fiecărui analizator. Calea de conducere indirectă, utilizează sistemul
reticulat ascendent activator, prin care impulsurile nervoase sunt conduse lent şi proiectate
cortical, ţn mod difuz şi nespecific.
Segmentul central este reprezentat de aria din scoarţa cerebrală, la care ajunge calea de
conducere şi la nivelu căreia excitaţiile sunt transformate în senzaţii conştiente specifice. În
mijlocul segmentului central există o zonă de specializare maximă (arie primară); lezarea ei
duce la pierderea simţului respectiv. Zona din jur este o zonă asociativă şi este denumită arie
psiho-senzorială (arie secundară); lezarea ei duce la pierderea posibilităţii de asociere a
senzaţiei, cu informaţii de la ceilalţi analizatori. Date recente au stabilit că din centrii corticali
ai analizatorului prin sisteme de fibre cortico-fugale, poate fi influenţată direct activitatea
celulelor receptoare, modificând pragul de excitabilitate al acestora.
9.1. Analizatorul cutanat.
Pielea
Segmentul periferic a analizatorului cutanat este situat în piele. Pielea este învelişul
extern la organismului, de natură conjunctivo-epitelială, care îl separă de mediul extern şi se
continuă la nivelul orificiilor naturale cu mucoasele. Pielea este un organ de protecţie faţă de
agenţii fizici, chimici, microbieni; organ de excreţie prin secreţie sudorală, eliminând apă, uree
şi produşi de catabolism; organ de absorbţie a unor substanţe medicamentoase; organ cu rol în
termoreglare; organ cu funcţii hemodinamice, depozitând sânge în reţeaua capilară. Pe lângă
toate acestea, datorită receptorilor pe care-i conţine, pielea are un important rol de organ de
simţ.
De la suprafaţă spre profunzime, pielea este alcătuită din 3 straturi: epidermul, dermul
şi hipodermul (ţesutul subcutanat). Pielea are o dublă origine embrionară provenită din
ectoderm (epidermul) şi din mezoderm (dermul şi hipodermul). Suprafaţa pielii la omul adult
este de cca. 1,5 m2, iar greutatea cca. 18 kg. Mobilitatea pielii pe planurile subiacente este mai
mare pe feţele de extensie ale articulaţiilor (cot, genunchi, partea dorsală a mâinii) şi redusă sau
chiar absentă pe feţele de flexiune (plica cotului, palma, planta piciorului).

Epidermul este format dintr-un epiteliu pluristratificat pavimentos, cheratinizat, aşezat
pe o membrană bazală. În partea sa profundă se află pătura germinativă (constituită din stratul
bazal şi stratul spinos), iar deasupra ei pătura cornoasă (ce curpinde stratul glanular, lucid şi
stratul cornos).
- Stratul bazal format dintr-un singur rând de celule prismatice, aşezate pe membrana bazală
care le desparte de derm.
- Stratul spinos format din 6-20 de rânduri de celule poliedrice, legate între ele, prin prelungiri
în formă de spini (desmozomi). În ambele straturi se găsesc celule cu pigment melanic produs
de melanocite, care sunt localizate în derm. Pigmentul melanic dă culoarea mai închisă sau mai
deschisă a pielii. Celulele din pătura germinativă se divid activ, asigurând reînnoirea straturilor
superficiale ale epidermului.
- Stratul granular este format din 1-5 rânduri de celule aplatizate, care conţin granulaţii de
keratină în citoplasmă. Nucleii se fragmentează deoarece la nivelul acestui strat celulele
epidermului încep să moară.
- Stratul lucid este format din celule turtite, cu nucleul degenerat, cu citoplasmă omogenă şi
transparentă datorită eleiedinei, pe care o conţine şi care provine din degradarea keratinei.
- Stratul cornos conţine celule foarte turtite, cu un înveliş de keratină, o proteină bogată în sulf,
rezistentă şi elastică; acest strat este foarte gros pe palme şi pe planta piciorului. Celulele
keratinizate sunt celule moarte, iar legăturile dintre ele slăbesc încet şi se descuamează (se
desprind şi cad). În epiderm nu pătrund vase de sânge, în schimb se găsesc numeroase
terminaţii nervoase libere.
Dermul (corionul) este stratul de sub epiderm, mai gros decât acesta şi format din ţesut
conjunctiv dens. Dermul este alcătuit dintr-un strat papilar superficial şi un strat rticular
profund.
- Stratul papilar ridică spre epiderm o serie de proeminenţe conice, numite papile dermice, cu
aspect ondulat pe suprafaţa de secţiune. Pe suprafaţa degetelor, în palmă şi în talpa piciorului
papilele sunt mai proeminente şi formează ridicături cunoscute sub numele de creste papilare, a
căror întinpărire constituie amprentele digitale. Stratul papilar este format din ţesut conjunctiv
cu substanţă fundamentală abundentă, fibroblaşti, melanoblaşti, fibre elastice şi de reticulină,
capilare şi fibre nervoase.

- Stratul reticular continuă stratul precedent, care este mai bogat în fascicule de fibre colagene
şi lame elastice, care dau rezistenţă pielii. Conţine de asemenea capilare sanguine şi limfatice,
fibre nervoase.
În derm sunt glande sebacee, canalele de excreţie ale glandelor sudoripare, partea
superficială a foliculilor piloşi, reţeaua vasculară a dermului şi numeroşi receptori nervoşi.
Hipodermul (stratul subcutanat) este format din ţesut conjunctiv lax, bogat în celule
adipoase, grupate în lobuli adipoşi (paniculi adipoşi) printre care se găsesc fibre conjunctive.
În stratul subcutanat se află glomerulul glandelor sudoripare, partea profundă a foliculilor
piloşi, reţeaua vasculară subcutanată, nervii subcutanaţi, receptorii nervoşi (corpusculii Vater-
Pacini, Ruffini, Golgi-Mazzoni).
Anexele pielii sunt cornoase (firele de păr şi unghiile) şi glandulare (glandele
sudoripare, sebacee, glandele mamare).
Receptorii cutanaţi.
Receptorii cutanaţi (tactili, dermici, dureroşi şi de presiune) sunt exteroceptori de
contact situaţi la nivelul pielii. Din punct de vedere structural receptorii cutanaţi sunt de 2
tipuri: terminaţii nervoase libere şi receptorii incapsulaţi.
Terminaţii nervoase libere sunt arborizaţii dendtritice ale neuronilor senzitivi din
ganglionii spinali, distribuite printre celulele păturii geminative a epidermului. La om au fost
descrise două varietăţi morfologice de terminaţii nervoase intraepidermice:
- reţeaua intraepidermică e formată din fibre amielinice situate în profunzimea epidermului
din care se desprind expansiuni nervoase ce se termină la suprafaţa celulelor sub formă de
butoni ce recepţionează excitaţiile dureroase;
- expansiunile iederiforme (discurile tactile Merkel) fac trecerea către receptorii incapsulaţi şi
sunt reprezentate prin fibre mielinice provenite din plexul nervos din derm, care se termină sub
forma unui disc neurofibrilar concav în care este aşezată o celulă epitelială modificată;
recepţionează stimulii tactili.
Receptorii incapsulaţi sunt expansiuni nervoase în care terminaţiile senzitive sunt
izolate în interiorul unei capsule conjunctive, ansamblul numindu-se corpuscul senzitiv.
Capsula este formată din mai multe lamele concentrice, iar porţiunea axială este reprezentată
de una sau mai multe fibre nervoase amielinice.

- Corpusculii Meissner sunt situaţi în papilele dermice, mai numeroşi la nivelul palmei şi
plantei piciorului. Au formă ovoidă şi o lungime de cca. 100μ (microni); recepţionează
excitaţiile de presiune şi tactile.
- Corpusculii Krausse sunt situaţi la nivelul papilelor dermice, au formă sferoidală, în interiorul
lor fibra nervoasă este bogat ramificată realizând aspect de reţea. Recepţionează stimulii
termici reci.
- Corpusculii Ruffini sunt localizaţi în dermul profund şi în hipoderm, fibra nervoasă, cu
ramificaţii fine este cuprinsă într-o stromă conjuctivă acoperită la exterior de o capsulă subţire
din lamele concentrice. Recepţionează stimulii termici calzi, dar şi tracţiunile şi deformările
care se exercită asupra fibrelor conjunctive din straturile profunde ale dermului.
- Corpusculii Vater-Pacini sunt corpusculi lamelari, de formă ovoidală, cu lungimea de 1-5
mm., vizibili şi cu ochiul liber. Recepţionează stimulii daţi de presiunea şi tracţiunea pielii şi
sunt foarte numeroşi la nivelul pulpei degetelor. Sunt formaţi din multiple lame concentrice în
axul lung urcând fibra nervoasă.
- Corpusculii Golgi-Mazzoni au o structură asemănătoare celor precedenţi, dar sunt mai mici,
localizaţi în hipodermul pulpei degetelor. Recepţionează stimulii daţi de presiuni mai mici
decât corpusculii Vater-Pacini.
Segmentul de conducere. Sensibilitatea tactilă protopatică, grosieră se transmite prin
fasciculele spinotalamice anterioare, iar cea tactilă epicritică, pe calea fasciculelor Goll şi
Burdach. Sensibilitatea dureroasă este condusă pe calea fasciculuui spinotalamic lateral. Toate
acest căi de conducere după încrucişare ajung la talamus, unde este al treilea neuron.
Segmentul central este situat în aria senzitivă primară, din lobul parietal, de partea
opusă celei receptoare.
9.2. Analizatorul kinestezic.
Segmentul periferic e reprezentat de proprioreceptori situaţi în muşchi, tendoane,
capsule articulare şi periost. Receptorii kinestezici din capsule articulare, tendoane, periost şi
articulaţii sunt corpusculii Vater-Pacini, identici cu cei din piele; sunt sensibili la mişcări şi
modificări de presiune.
-Corpusculii Ruffini sunt situaţi în stratul superficial al capsulei articulare şi recepţionează
informaţii în legătură cu poziţia şi mişcările din articulaţii.

-Terminaţiile nervoase libere se ramifică în toată grosimea capsulei articulare şi transmit
sensibilitatea dureroasă articulară cauzată de amplitudinea excesivă a mişcării.
-Corpusculii neuro-tendinoşi Golgi sunt situaţi la limita dintre corpul muşchiului şi tendon.
Sunt formaţi dintr-o reţea de fibre nervoase terminate sub formă de butoni, printre fasciculele
tendinoase. La exterior fasciculele sunt înconjurate de o capsulă subţire conjunctivă. În
corpuscul pătrund 1-3 fibre mielinice, care la intrare îşi pierd teaca de mielină (fig. 34).
Terminaţiile nervoase sunt excitate de întinderea puternică a tendonului în timpul contracţiei
musculare.
- Fusurile neuro-musculare sunt formate din 5-10 fibre musculare modificate numite fibre
intrafusale, învelite într-o capsulă conjunctivă. Fibrele musculare intrafusale sunt de 2 tipuri:
fibre cu sac nuclear, lungi, groase cu o parte centrală, dilatată, fără striaţii, ce conţin 40-50
nuclei şi fibre cu lanţ nuclear, subţiri şi scurte cu nucleii aşezaţi în şir pe toată lungimea lor.
Capetele fusului neuro-muscular, care conţin fibre extrafusale contractile, primesc terminaţii
nervoase motorii.
Fusurile au inervaţie senzitivă şi motorie.
Inervaţia senzitivă este asigurată de dentritele neuronilor senzitivi din ganglionul
spinal. Unele dintre ele se numesc anulospirale şi se rulează în jurul ecuatorului fibrelor cu sac
nuclear, alte terminaţii nervoase "în buchet" se termină pe ecuatorul cu lanţ nuclear.
Inervaţia motorie este asigurată de axonii motoneuronilor γ (gama) din cornul anterior
al măduvei. Excitarea neuronului γ, activează zonele polare ale fibrelor intrafusale, care prin
contracţie excită receptorul situat în zona ecuatorială.
Segmentul de conducere. Impulsurile aferente de la proprioceptori sunt conduse pe 2
căi:
-pentru sensibilitatea kinestezică (simţul poziţiei şi al mişcării în spaţiu), prin fasciculele
spinobulbare (sensibilitatea proprioceptivă conştientă); excitaţiile sunt colectate de la
corpusculii Golgi, Ruffini, Pacini şi terminaţiile nervoase libere;
-pentru sensibilitatea proprioceptivă de control a mişcării (simţul tonusului muscular), prin
fasciculele spinocerebeloase ventrale şi dorsale (sensibilitatea proprioceptivă inconştientă);
receptorii acestei căi sunt fusurile neuro-musculare;

9.3 Analizatorul olfactiv.
Segmentul periferic este situat în mucoasa olfactivă ce acoperă lama ciuruită a
etmoidului, faţa medial a cornetului nazal superior şi o mică parte din septul nazal. Epiteliul
mucoasei olfacive este format din celule de susţinere, celule nervoase senzitive bipolare şi
celule bazale. Celulele bipolare au rol de receptor sunt fusiforme, iar dendrita lor se termină la
suprafaţa mucoasei cu un buton olfactiv de la care pleacă 5-6 cili olfactivi. În corionul
mucoasei sunt glande seroase care menţin umedă mucoasa.
Segmentul de conducere. Axonii celulelor olfactive străbat lama ciuruită se
înmănunchează şi formează 10-20 nervi olfactivi, ce fac sinapsă cu deutoneuronul căilor
olfactive reprezentat de celulele mitrale multipolare din bulbul olfactiv. Axonii acestor neuroni
mitrali formează tractul olfactiv, care se termină prin trigonul olfactiv, de la care pleacă stria
olfactivă medială, laterală ce delimitează substanţa perforată anterioară. Calea olfactivă nu are
legături directe cu talamusul.
Segmentul central. Centrii primari unde ajung fibrele deutoneuronilot sunt formaţi de
trigonul olfactiv, substanţa perforată anterioară şi nucleii septali. Aceşti centri au legătură cu
centrii olfactivi secundari, din hipocamp (lobul temporal) şi nucleul amigdalian.
9.4. Analizatorul gustativ.
Segmentul periferic. Receptorii analizatorului gustativ sunt reprezentaţi de mugurii
gustativi situaţi în mucoasa linguală (papilele caliciforme, fungiforme şi foliate). Mugurii
gustativi se prezintă ca formaţiuni ovoide formate de celeule de susţinere, între care se găsesc
5-20 de celule senzoriale, ce au la polul apical câte 3-6 cili, care pătrund în orificiul de la
suprafaţă numit porul gustativ al mugurului; cilii vin în astfel în contact cu substanţele sapide
dizolvate. La polul bazal celulele gustative vin în contact cu terminaţiile nervoase senzitive ale
nervilor facial, glosofaringian şi vag.
Segmentul intermediar. Primul neuron se află situat în ganglionii de pe traiectul
nervului VII (ganglion geniculat), IX (ganglion extracranian) şi X (ganglion plexiform al
vagului). Axonii protoneuronilor ajung la nucleul solitar din bulb, unde fac sinapsă cu
deutoneuronii, iar al treilea neuron este în talamus.

Segmentul central se găseşte în aria 43 din lobul parietal din porţiunea inferioară a
girusului postcentral.
9.5. Analizatorul vizual.
Segmentul periferic al analizatorului vizual este alcătuit din globul ocular şi organele
anexe ale acestuia.
Globul ocular
Este alcătuit dintr-un perete format din 3 tunici concentrice şi o cavitate în care se află
mediile refringente (transparente) ale ochiului.
Tunica externă este fibroasă şi formată din 2 porţiuni inegale: posterior se află
sclerotica este opacă şi mai mare ca întindere şi anterior corneea, redusă ca suprafaţă şi
transparentă. Sclerotica este membrana albă-sidefie, dură, care prezintă în partea postero-
mediană lama ciuruită, o zonă perforată prin care trec fibrele nervului optic. Corneea este o
membrană conjunctivă transparentă, care nu are vase de sănge, dar conţine fibre nervoase
amielinice. Între sclerotică şi cornee, se află şanţul sclerocorneean, în profunzimea căruia se
află canalul Schlemm, prin care trece umoarea apoasă spre sistemul venos.
Tunica medie sau vasculară este bogată în celule pigmentare şi vase de sânge, cuprinde
3 părţi: coroida, corpul ciliar şi irisul.
- Coroida este o membrană abundent vascularizată şi are un rol important în nutriţia globilor
oculari. Coroida căptuşeşte sclerotica şi prin celule pigmentare pe care le conţine contribuie la
formarea camerei obscure. Posterior, coroida prezintă un orificiu cu diametrul de 1,5 mm, prin
care iese nervul optic; el corespunde lamei ciuruitei a sclerei.
- Corpul ciliar continuă anterior coroida şi este o formaţiune conjunctivo-vasculară; în
structura sa intră procesele ciliare şi muşchii ciliari. Procesele ciliare sunt alcătuite din cute
vasculare în număr de 60-80, printre care se găseşte ţesut elastic; ele secretă umoarea apoasă.
- Muşchii ciliari sunt alcătuite din fibre musculare netede dispuse longitudinal, circular şi
radial, ce acţionează asupra cristalinului prin intermediul ligamentului suspensor (Zinn), care
se întinde de la corpul ciliar la cristalin.
- Irisul reprezintă partea anterioară a tunicii mijlocii şi are forma unui disc, în centrul căruia se
află orificiul numit pupilă. Muşchii irisului sunt alcătuiţi din fibre musculare netede, circulare
şi radiale, cu rol în micşorarea sau mărirea diametrului pupilei. În iris se găsesc celule

conjunctive ce conţin pigment care dă culoarea ochiului: un număr mare de celule pigmentare
realizează culoarea brună, în timp ce o cantitate mică de pigment determină culoarea albastră.
Tunica internă sau nervoasă este reprezentată de retină (de origine nervoasă),
responsabilă de recepţia şi transformarea stimulilor luminoşi în influx nervos. Din punct de
vedere funcţional i se disting 2 regiuni: retina oarbă care vine în raport cu corpul ciliar şi cu
irisul şi nu conţine celule fotosensibile; retina vizuală situată în partea posterioară, ce prezintă
2 regiuni importante (pata galbenă şi pata oarbă).
Pata galbenă (macula luteea) situată în dreptul axului vizual prezintă în centrul său o scobitură
de 1,5 mm.2 numită foveea centralis în care se găsesc numai celule cu conuri.
Pata oarbă (papila optică) situată medial şi inferior de pata galbenă, reprezintă locul de ieşire
a nervului optic din globul ocular şi de intrare a arterelor; aici nu există elemente fotosensibile.
Retina are o structură stratificată datorită aşezării în lanţ a neuronilor şi a legăturilor lor
sinaptice. Se deosebesc 10 straturi (fig. 37) în care se găsesc 3 feluri de celule funcţionale,
aflate în relaţii sinaptice: celule fotoreceptoare cu prelungiri în formă de con şi celule cu
prelungire în formă de bastonaş, celule bipolare şi celule multipolare. În afara acestora se mai
găsesc celule de susţinere şi celule de asociaţie. Începând de la coroidă, cele mai importante
sunt:
-stratul celulelor pigmentare alcătuit dintr-un rând de celule încărcate cu pigment melanic; ele
trimit prelungiri citoplasmatice ce se dispun în jurul cornurilor şi bastonaşelor, formând camere
obscure;
-stratul celulelor vizuale constituit din elemente fotoreceptoare (celule cu bastonaşe şi celule
cu conuri).
Celulele cu bastonaşe sunt celule nervoase modificate în număr de 125 milioane. Sunt
mai numeroase în partea periferică a retinei, iar în pata galbenă numărul lor este mic, lipsind în
foveea centralis. Celulele cu bastonaşe sunt adaptate pentru vederea nocturnă, la lumină slabă.
Mai multe celule cu bastonaşe fac sinapsă cu o celulă bipolară şi mai multe celule bipolare fac
sinapsă cu o celulă multipolară (fenomenul de convergenţă a impulsurilor) ceea ce explică
faptul că la o celulă multipolară corespund circa 200 celule cu bastonaşe. Celulele cu bastonaşe
conţin o substanţă fotosensibilă numită rodopsină.
Celulele cu conuri şi ele sunt celule nervoase modificate în număr de 7 milioane; mai
numeroase în pata galbenă, iar în foveea centralis sunt numai cele cu conuri. Fiecare celulă cu

conuri face sinapsă cu o singură celulă bipolară, iar aceasta cu o singură celulă multipolară;
acestea permit vederea diurnă, colorată, la lumină intensă. Celulele cu conuri conţin o
substanţă fotosensibilă numită iodopsina.
- stratul neuronilor bipolari, format din celule bipolare care reprezintă primul neuron al căii
optice. Dendrtitele lor fac sinapsă cu prelungirile interne ale celulelor cu conuri şi bastonaşe.
Axonii acestor celule fac sinapsă cu dendritele celulelor multipolare;
- stratul neuronilor multipolari, format din celule multipolare care reprezintă al doilea neuron
al căii optice. Dendritele lor fac sinapsă cu axonii celulelor bipolare, iar axonii lor formează
fibrele nervului optic.
Mediile refringente au rolul de a refracta razele de lumină, proiectând imaginea pe
retină. În afară de cornee, care face parte din tunica externă a globilor oculari, mediile
transparente sunt: umoarea apoasă, cristalinul şi corpul vitros.
Umoarea apoasă este un lichid transparent, secretat de procesele ciliare care umple
camera anterioară a ochiului între cristalin şi iris şi de aici prin pupilă ajungând în spaţiul dintre
cornee şi iris.
Cristalinul este organul cel mai important al aparatului dioptric, el are forma unei
lentile biconvexe aşezat în plan frontal, între iris şi corpul vitros. La exterior este acoperit de o
membrană subţire dar rezistentă numită capsula cristalinului. Marginile capsulei sunt fixate
prin intermediul ligamentului suspensor de procesele ciliare. Muşchiul ciliar prin intermediul
ligamentului suspensor, modifică curbura cristalinului (în special a feţei anterioare) intervenind
astfel în acomodarea la distanţă.
Corpul vitros este o substanţă gelatinoasă, omogenă şi transparentă, secretată tot de
procesele ciliare. Ocupă camera posterioară a globilor oculari dintre cristalin şi retină. La
exterior el prezintă un strat dens, subţire ce se numeşte membrană heloidă.
Anexele globului ocular.
Se împart în anexe de mişcare şi anexe de protecţie.
Anexe de mişcare asigură mişcarea globilor oculari în diferite direcţii. Ele sunt
reprezentate de muşchii extrinseci ai globilor oculari care sunt muşchi striaţi (spre deosebire de
muşchii intrinseci (irisul şi corpul ciliar) care sunt muşchi netezi). Sunt 4 muşchi drepţi şi 2
muşchi oblici.

Muşchi drepţi (superiori, inferiori, externi şi interni) au formă de trunchi de con, cu
baza (inserţia) pe sclerotică, înaintea zonei ecuatoriale a globilor oculari şi vârful (originea) pe
un inel tendinos comun (tendonul Zinn) fixat în vecinătatea orificiului optic din orbită.
Muşchi oblici superiori au aceeaşi origine cu muşchii drepţi, dar trec mai întâi printr-un
inel fibro-cartilaginos situat în unghiul intern al orbitei şi se inseră în partea latero-superioară a
scleroticii. Muşchi oblici inferiori au originea pe osul lacrimal şi inserţia pe partea latero-
inferioară a scleroticii.
Mişcările globilor oculari sunt conjugate prin acţionarea bilaterală a unui număr de
muşchi. Mişcarea de lateralitate, dreaptă sau stângă, se efectueză prin contracţia dreptului
extern de la un ochi împreună cu dreptul intern de la ochiul opus. Mişcarea de convergenţă a
ochilor se realizează prin contracţia ambilor muşchi drepţi interni. Mişcarea în sus se realizează
prin contracţia muşchilor drepţi superiori şi a celor oblici inferiori, bilateral. Mişcarea în jos se
realizează prin contracţia drepţilor inferiori şi a oblicilor superiori, bilateral.
Nervul III (oculomotor) inerează muşchii: muşchii drept superior, drept inferior, drept
intern şi oblic inferior. Nervul IV (tohlear) inervează muşchiul oblic superior, iar nervul VI
(abducens) inervează muşchiul drept extern.
Anexe de protecţie sunt reprezentate de sprâncene, pleoape, conjunctiva, aparatul
lacrimal.
Sprâncenele sunt formate din fire de păr situate deasupra orbitei, care împiedică
scurgerea transpiraţiei pe globul ocular.
Pleoapele sunt două cute (superioară şi inferioară) musculo-fibroase acoperite de piele
şi căptuşite pa faţa internă de conjunctivă cu rolul de a proteja corneea transparentă. Pe
marginile pleoapelor se găsesc genele ce reţin particulele de praf. Pleoapele delimitează
deschiderea palpebrală şi conţin glande sebacee modificate (Mebomius) dispuse la baza
genelor, glande sudoripare modificate (Moll) şi glande ciliare care se deschid pe marginea
liberă a pleoapelor.
Conjunctiva este o membrană fină, transparentă ce tapetează faţa posterioară a
pleoapelor şi cea anterioară a globilor oculari, până la periferia corneei, unde se continuă cu
epiteliul acesteia. În unghiul intern al ochiului acoperă o proeminenţă numită caruncula
lacrimală, iar lateral de aceasta se află plica semilunară (rest al celei de a treia pleoapă de la
păsări).

Aparatul lacrimal cuprinde glandele lacrimale şi conductele lacrimale. Glandele
lacrimale sunt localizate în partea latero-superioară a orbitei. Sunt glande tubulo-acinoase ce
secretă lacrimile eliminate prin 18-20 canalicule excretoare la nivelul conjunctivei. Prin
mişcarea pleoapelor lacrimile sunt dirijate către unghiul intern al pleoapelor, în lacul lacrimal,
de unde sunt preluate de canalele lacrimale care încep pe marginea liberă a pleoapelor;
canalele lacrimale conduc lacrimile spre sacul lacrimal şi de aici prin canalul nazo-lacrimal
ajung în fosele nazale, la nivelul meatului nazal inferior.
Segmentul intermediar al analizatorului vizual (calea optică) este alcătuit din
înlănţuirea a 3 neuroni: protoneuronul este reprezentat de celule bipolare, care primesc
excitaţiile de la celulele cu conuri şi cu bastonaşe, iar deutoneuronul este reprezentat de celule
multipolare. Axonii celulelor multipolare formează nervul optic. La nivelu chiasmei optice,
fibrele ce vin din jumătatea nazală a retinei se încrucişează, mergând în tractul optic în partea
opusă. Fibrele din jumătatea temporală a retinei, nu se încrucişează şi merg în tractul optic de
aceeaşi parte. Deci, nervul optic conţine fibre de la un singur ochi, iar tractul optic conţine fibre
de la ambii ochi. Axonii deutoneuronilor (din tractul optic) ajung la metatalamus în corpul
geniculat extern, unde fac sinapsă cu al treilea neuron.
Segmentul central. Axonii neuronilor din corpul geniculat extern se îndreaptă spre
scoarţa cerebrală şi se termină în lobul occipital, de o parte şi de alta a scizurii calcarine, în
câmpurile 17, 18, 19, ce reprezintă aria vizuală, în care excitaţiile luinoase se transformă în
senzaţii vizuale.
9.6. Analizatorul acustico-vestibular.
Analizatorul acustic (pentru auz) şi analizatorul vestibular (pentru poziţia spaţială a
corpului în repaos şi mişcare) sunt două aparate receptoare situate în urechea internă. Fiecare
au însă receptorii, căile de conducere şi segmentele centrale separate; receptorii sunt însă
situaţi împreună în urechea internă. Perfecţionarea aparatului acustic a determinat dezvoltarea
unor anexe importante (urechea externă şi urechea medie) care nu au nici o relaţie cu aparatul
vestibular. Urechea este alcătuită din 3 părţi cu structură şi funcţii distincte: urechea externă,
urechea medie şi urechea internă.
Urechea externă.
Această porţiune este alcătuită din pavilionul urechii şi contuctul auditiv extern.

Pavilionul urechii este situat în partea laterală a capului în spatele articulaţiei temporo-
mandibulare, având o formă de pâlnie şi prezintă pe suprafaţa sa proeminenţe şi depresiuni cu
rolul de a dirija undele sonore spre conductul auditiv extern. În centrul său se găseşte o
depresiune numită concă. În structura sa are un schelet fibrocartilaginos, învelit pe ambele feţe
de piele. În partea inferioară există o prelungire numită lobul urechii, lipsită de cartilaj.
Pavilionul este legat de oasele din jur de către muşchii auriculari (extrinseci), care la om sunt
rudimentari; mai conţine ligamente şi muşchi intrinseci (muşchii tragusului, helisului etc.) care
au rol în menţinerea pliurilor.
Contuctul auditiv extern are o lungime de 3 cm. şi se întinde de la conca auriculară la
membrana timpanică. În partea laterală, are o structură fibro-cartilaginoasă, iar în partea
medială, osoasă. Este acoperit cu piele, ce prezintă peri şi glande ceruminoase (glande sebacee
modificate) al căror produs de secreţie, cerumenul, are rol de protecţie împotriva pătrunderii
corpurilor străine.
Urechea medie.
Este o cavitate situată în interiorul stâncii temporalului, care prezintă central cavitatea
timpanică, cu rolul de a transmite vibraţiile aerului spre urechea internă. Anterior cavitatea
timpanică comunică cu nazofaringele, prin trompa lui Eustachio, ce contribuie la egalizarea
presiunii de o parte şi de alta a timpanului. Peretele posterior are un orificiu prin care cutia
timpanică comunică cu cavităţi din interiorul apofizei mastoide, numite celule mastoidiene.
Peretele medial corespunde urechii interne şi prezintă în centru o proeminenţă numită
promontoriu, care separă 2 orificii: superior, fereastra ovală (vestibulară), acoperită de talpa
scăriţei, iar inferior fereastra rotundă.
Peretele extern este reprezentat de membrana timpanică, formată dintr-un strat fibros cu
fibre circulare şi radiare, învelit lateral spre conductul auditiv de piele, iar medial de mucoasa
cavităţii timpanice. Pe timpan se inseră mânerul ciocanului. Între timpan şi fereastra ovală se
află un lanţ de 3 oscioare articulate între ele: ciocan, nicovală şi scăriţă. Ciocanul se sprijină cu
mânerul pe membrana timpanică şi se articulează cu nicovala, iar aceasta cu scăriţa care se
fixează cu talpa pe membrana ferestrei ovale. Rolul acestor oscioare este de a transmite
vibraţiile timpanului spre membrana feretsrei ovale şi de aici prin perilimfă, receporilor
acustici.

Urechea internă.
Este formată dintr-un sistem de cavităţi ce comunică între ele, săpate în stânca
temporalului care în ansamblu alcătuiesc labirintul osos, în interiorul căruia, mulându-se pe el,
este labirintul membranos, în care sunt receptorii acustici şi vestibulari. Între labirintul osos şi
membranos se află un lichid numit perilimfă, iar în labirintul membranos, endolimfă.
Labirintul osos este format din vestibul, canale semicirculare şi melcul osos.
- Vestibulul este o cavitate centrală, cu şase pereţi. Pe peretele lateral se găseşte fereastra ovală
(acoperită de talpa scăriţei) şi rotundă (acoperită de membrana elastică). Peretele posterior şi
superior prezintă 5 orificii de deschidere ale canalelor semicirculare. Vestibulul comunică şi cu
melcul osos.
- Canalele semicirculare osoase sunt în număr de 3, în formă de potcoavă, dispuse
perpendicular între ele, în cele 3 planuri ale spaţiului; canalul anterior şi posterior sunt
verticale, iar cel lateral este orizontal. Din cele 2 extremităţi ale unui canal, una este mai
dilatată şi se numeşte ampulă, capetele neampulare (lipsite de dilataţii) ale canalelor anterior şi
posterior, sunt fuzionate aşa încât în vestibul sunt 5 orificii.
- Melcul osos este situat anterior de vestibul şi este format dintr-un canal spiralat, care descrie 2
spire şi jumătate în jurul unui ax central, numit columelă (modiol). De pe columelă pleacă o
lamă subţire, numită lama spirală osoasă (mai lată la bază şi îngustată spre vârf) ce separă
incomplet canalul osos într-un compartiment superior, numit rampa vestibulară şi altul inferior
numit rampa timpanică. Separarea celor 2 rampe este completată de o formaţiune fibroasă
numită membrana bazilară, care se întinde de la marginea liberă a lamei spirale osoase, până la
peretele lateral al melcului osos.
Comunicarea între cele 2 rampe se face printr-un orificiu numit helicotremă, situat la
vârful melcului. La baza lamei spirale este un canal spiral în care se găseşte ganglionul spinal
al lui Corti. În acest canal, se deschid canalicule aferente din lama spirală, prin care trec
dendritele neuronilor receptori şi canale eferente, cu traiect longitudinal în columelă, prin care
plecă axonii ce alcătuiesc ramura cohleară a nervului VIII.
Labirintul membranos este situat în interiorul labirintului osos şi este format din 2
vezicule mici, utricula şi sacula, 3 canale semicirculare membranoase şi melcul membranos
(canalul cohlear).

- Utricula are o formă elipsoidală în ea deschizându-se cele 3 canale semicirculare
membranoase prin 5 orificii. Printr-un canal endolimfatic, comunică cu un mic diverticul, plin
cu endolimfă, numit şi sac endolimfatic care ajunge până la duramater; din acesta pleacă un alt
canal ce stabileşte legătura cu sacula.
- Sacula este o veziculă sferică, mai mică decât utricula şi situată inferior faţă de aceasta.
- Canalele semicirculare membranoase corespund celor osoase deschizându-se prin 5 orificii
în utricule (2 se contopesc) dintre care 3 sunt mai dilatate şi se numesc ampulele canaelor
semicirculare membranoase.
- Melcul membranos (canalul cohlear) descrie ca şi melcul osos 2 spire şi jumîtate. Pe o
secţiune transversală are o formă triunghiulară, cu un perete format din membrana vestibulară
(Reissner), care-l separă de rampa vestibulară, altul format de membrana bazilară care-l separă
de rampa timpanică şi un perete lateral opus lamei spirale osoase. În ambele rampe se găseşte
perilimfă şi comunică între ele prin helicotremă, iar în canalul cohlear se găseşte emdolimfa.
Canalul cohlear cumunică printr-un mic canal cu sacula.
Segmentul periferic.
Receptorii acustici se găsesc în canalul cohlear şi sunt reprezentaţi de organul spiral
(Corti) aşezat pe membrana bazilară. La nivelul său se descriu 2 stâlpi extern şi intern, formaţi
din celule de susţinere înalte, depărtate la bază şi apropiate la vârf, care delimitează tunelul lui
Corti. Celulele senzoriale acustice sunt aşezate în 3 şiruri laterale şi un şir medial faţă de
tunelul Corti, sprijinite pe celule de susţinere mai mici numite celule Deiters; celule de
susţinere internă sunt dipuse pe un singur rând, iar cele externe pe 3 rânduri ca şi celulele
auditive. Celule de susţinere interne sunt continuate spre lama spirală osoasă, de un epiteliu
cubic simplu, în timp ce celulele de susţinere externe sunt continuate spre peretele extern al
canalului cohlear, de celule înalte numite celulele Hensen; acestea la rândul lor diminuă şi se
continuă cu celule cubice numite celulele Claudius.
Celulele auditive sunt mici şi la polul apical prezintă cili (80-100 pentru cele externe şi
40-60 pentru cele interne), care străbat o membrană subţire numită membrana reticulară; iar la
polul bazal sunt înconjurte de terminaţii dendritice ale neuronilor senzitivi din ganglionul spiral
Corti (situat în columelă). Deasupra organului lui Corti se află membrana tectoria, care printr-o

extremitate aderă de lama spirală osoasă, iar cu cealaltă pluteşte liberă în endolimfă. Toate
formaţiunile organului Corti descriu cele 2 spire şi jumătate ca şi canalul cohlear.
Receptorii vestibulari se găsesc în utriculă, saculă şi ampulele canalelor semicirculare
membranoase.
- În utriculă şi saculă se găseşte câte o proeminenţă numită macula utriculei şi macula saculei,
ambele alcătuite dintr-un epiteliu cu celule senzoriale, ciliate aşezate pe celule de susţinere.
Cilii celulelor senzoriale, situate la polul apical, pătrund într-o substanţă gelatinoasă
care se găseşte la suprafaţa maculei şi care prezintă granule de carbonat de calciu numite
otolite, de unde şi denumirea de organe otolitice. La polul bazal al celulelelor senzoriale sosesc
dendritele neuronilor senzitivi din ganglionul vestibular Scarpa. Gravitaţia deformează
membrana otolitică şi astfel, mişcându-se cilii, se excită celulele senzoriale. Maculele
recepţionează deci, stimulii declanşaţi de poziţia capului (simţul static)
- Canalele semicirculare prezintă la nivelul ampulelor nişte proeminenţe transversale pe planul
canalului, numite creste ampulare. Acestea prezintă de asemenea, celule de susţinere şi celule
senzoriale vestibulare cu cili ceva mai groşi. Creasta este acoperită de o masă gelatinoasă sub
formă de cupolă, din care lipsesc otolitele. La polul bazal al celulelor senzoriale sosesc de
asemenea dendrite ale neuronilor din ganglionul vestibular Scarpa. Crestele ampulare
recepţionează stimulii declanşaţi de mişcarea capului, în timpul căreia, datorită inerţiei, în
canalul semicircular apare un curent de endolimfă care înclină cupola ampulară şi îndoaie cilii
celulelor senzoriale care vor fi excitate. Curentul apare doar atunci când canalul semicircular
este deplasat în lungul axului lui.
Segmentul de conducere.
Calea acustică (cohleară). Primul neuron este situat în ganglionul spinal Corti şi
dendritele ale ajung la polul bazal al celulelor auditive din organul Corti; axonii
protoneuronilor formează nervul cohlear, care împreună cu nervul vestibular constituie nervul
VIII cranian (acustico-vestibular), care străbate stânca temporalului, prin meatul auditiv după
care pătrunde în trunchiul cerebral la nivelul şanţului bulbo-pontin, în dreptul şanţului
retroolivar. Axonii se termină în nucleii cohleari (ventral şi dorsal) din planşeulul ventricului
IV, unde se găsesc deutoneuronii căii acustice.

Axonii deutoneuronilor se încrucişează pe linia mediană, formând corpul trapezoid, în
vecinătatea căruia se găseşte nucleul olivar pontin. După încrucişare axonii capătă un traiect
ascendent, formând lemniscul lateral, care se îndreaptă spre coliculul inferior, unde se găseşte
al III neuron. Urmează al patrulea neuron al căii acustice este situat în metatalamus, în corpul
geniculat medial; axonul acestuia formează radiaţiile acustice, ce se proiectează pe scoarţa
cerebrală.
Calea vestibulară. Protoneuronii se găsesc în ganglionul vestibular Scarpa, situat pe
traiectul nervului, în urechea internă. Dendritele primului neuron, merg la celulele senzoriale
vestibulare din macule şi crestele ampulare, iar axonii constituie ramura vestibulară a nervului
VIII cranian, care pătrunde în trunchiul cerebral prin şanţul bulbopontin la nivelul fosetei
retroolivare. Deutoneuronii se găsesc în nucleii vestibulari bulbari (superior, inferior, lateral şi
medial) şi prin axonii lor constituie următoarele fascicule:
- fasciculul vestibulo-spinal, spre măduvă;
- fasciculul vestibulo-cerebelos, spre cerebel;
- fasciculul vestibulo-nuclear, spre nucleii de origine ai nervilor III, IV (din mezencefal) şi VI
din punte;
- fasciculul vestibulo-talamic, spre talamus de unde prin fibrele talamo-corticale se proiectează
spre scoarţa cerebrală (probabil în girusul temporal superior);
Segmentul central.
Centrii corticali ai auzului sunt situaţi în ariile 41, 42 din girusul temporal superior,
unde există o somatotopie (corespondenţă spaţială) între cohlee şi zona corticală. Centrii
auzului au ample conexiuni cu câmpurile 6 şi 8 din lobul frontal şi mai ales cu centrul vorbirii
din aria 44, vorbirea fiind strâns legată de simţul auditiv.
Centrii corticali vestibulari sunt mai puţin cunoscuţi. Unii îi consideră în girurusul
temporal superior, alături de aria auditivă. Cercetări recente au decelat impulsuri vestibulare,
tot cu proiecţie controlaterală în girusul postcentral, ceea ce sugerează că la nivel cortical
proiecţia ar putea fi legată de aferenţele proprioceptive.

Unitatea de curs 10 Aparatul respirator
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului aparatului respirator.
Obiectivele operaţionale
1. Prezentarea structurii aparatului respirator.
2. Prezentarea clasificării aparatului respirator.
3. Prezentarea rolului şi motilităţii aparatului respirator.
10.1. Structurile şi modalităţile prin care se realizează transportul
gazos
Respiraţia este o funcţie vitală a organismelor vii, prin care se realizează schimbul de
oxigen şi dioxid de carbon al acestora cu mediul înconjurător, astfel asigurându-se homeostazia
gazoasă implicată în metabolismul celular (I. Hăulică).
Componentele
aparatului respirator sunt
reprezentate de către căile
aeriene superioare şi
inferioare şi plămânii.
Căile aeriene
superioare sunt alcătuite din:
cavitatea nazală, faringele
şi laringele. Căile aeriene
inferioare sunt reprezentate
de trahee şi de bronhii.
Toate aceste căi sunt
aerovectoare, cu rol de a
conduce aerul atmosferic în
Figura 16 Căile aeriene superioare plămâni, precum şi de a-l

încălzi şi cu asigurând şi
vid şi cu dublu
rol fun
Î aerul inspirat până la temperatura corpului
Căptuşeala mucoasă are celule
brana mucoasă tinde să se usuce, astfel că organismul secretă mai mult mucus.
torită uscării mucusului.
scade gradul de umezire a
e de la C-3 la C-6
şi, prin contracţia sa rapidă, previne pătrunderea în căile
este un tub elastic, între laringe �i
bronhii
răţa de anumite particule străine, funcţii speciale (fonaţia şi
olfacţia).Nasul şi cavitatea nazală formează primul segment al acestui aparat.
Nasul, formaţiunea mediofacrală cu caracteristici specifice fiecărui indi
cţional (respirator şi olfactiv) îmbracă forma unei piramide cu baza în jos. Este alcătuit
dintr-un schelet osteocartilaginos compus din oasele nazale, cartilajele laterale, cartilajele aripii
nasului (alare mari şi mici), ca şi altele accesorii, mai mici. Fosele nazale comunică cu
exteriorul prin două orificii, numite narine, iar cu faringele, prin două orificii largi, meaturi
nazofaringiene, choane pe schelet. Cavităţile nazale sînt căptuşite de o mucoasă foarte bogat
vascularizatâ şi împărţită funcţional în două regiuni: una în treimea superioară, mucoasa
olfactivă, şi una în partea inferioară, mucoasa respiratorie.
Faringele
• Funcţie
a. ncălzeşte
b. Filtrează şi înlătură impurităţile din aerul inspirat.
care secretă mucus (caliciforme) şi celule ciliate. Cilii şi mucusul reţin particulele străine. Prin
strănut se elimină mare parte din aceste particule. În caz de boală sau de temperatură ridicată a
organismului:
a. Mem
Acest mucus se usucă şi ciclul se reia.
b. Acţiunea cililor este inhibată da
c. Pacientul are tendinţa de a respira pe gură, ceea ce
mucusului şi îi creşte vâscozitatea.
Laringele
1. Se întind
2. Controlează curentul de aer
aeriene a alimentelor, lichidelor şi a obiectelor străine.
Traheea are lungimea de 10-12 cm (C7-T4),
. Pereţii antero-laterali au forma unui C �i sunt alcătuite din cartilaj hialin (inele
cartilaginoase). Peretele posterior închide cartilajul prin muşchi �i ţesut conjunctiv, ca un inel.
Mucoasa prezintă celule cilindrice care prezintă cili vibratili

Plămânii sunt constituiţi din arborele bronşic şi din parenchimul pulmonar. Aceştia sunt
organiz
Arborele bronşic
ee rezultă cele două bronhii principale care pătrund prin hil în
plămân
-funcţională a plămânului şi este alcătuit dintr-o
bronhi
iile lobare şi segmentare (fig.17) au în structura lor schelet cartilaginos, bronhiile
lobular
le primesc aer prin căile respiratorii superioare şi ramificaţiile acestora, dar mai
pot prim
bazală,
format
aţi în lobi, segmente, lobuli şi acini pulmonari.
Prin bifurcaţia trah
i, unde se împart în bronhii lobare (trei pentru plămânul drept şi două pentru plămânul
stâng). Acestea la rândul lor se divid în bronhii segmentare proprii segmentelor pulmonare.
Segmentele au aeraţie, vascularizaţie şi patologie proprie. Bronhii segmentare se divid în
bronhiole lobulare care deservesc lobulii pulmonari. Bronhiolele lobulare dau bronhiole
respiratorii se continuă cu canalele alveolare ai căror pereţi reprezintă dilataţii sub formă de
saci în care se deschid alveolele pulmonare.
Acinul pulmonar este unitatea morfo
olă respiratorie împreună cu canalele alveolare care derivă din ea şi cu alveolele
pulmonare.
Bronh
e şi respiratorii îşi pierd scheletul cartilaginos, pereţii lor fiind fibroelastici şi cu
musculatură netedă circulară, aceasta scăzând treptat în profunzime, la nivelul ductului alveolar
fiind absentă.
Alveole
i şi de la alveolele din cadrul unui acin pulmonar vecin sau de la acelaşi acin.
Peretele alveolar este format dintr-un singur strat celular aşezat pe o membrană
din celule epiteliale, numite pneumocite 1 şi 2. La acest nivel se secretă o substanţă
lipidică denumită surfactant alveolar cu rol important în dinamica respiratorie care va deprima
tensiunea de la suprafaţa alveolelor şi deci colapsul lor va fi prevenit. În inspir nu permite
distensia exagerată a alveolelor ce ar putea fi periculoasă într-un inspir forţat, iar în expir nu
permite colabarea alveolelor.

Figura 17 Căile aeriene inferioare
Schimbul gazos alveolo-capilar
Alveolele sunt înconjurate de o reţea de capilare pericapilară, alcătuind bariera alveolo-
capilară. Aceasta este formată din epiteliul alveolar şi membrana fibroelastică, membrana
bazală a capilarului şi endoteliului capilar.
Molecula de oxigen va străbate straturile:
- surfactantul alveolar;
- celula epitelială alveolară;
- membrana bazală alveolară;
- spaţiul interstiţial;
- membrana bazală a endoteliului capilar;
- celulele endoteliului capilar;
- plasma sanguină;
- membrana eritrocitului.
Trecerea gazelor se face în funcţie de gradientul presional de o parte şi de alta a
membranei. Acestea sunt:
Aer alveolar Sânge capilar
pO2 = 100mm Hg 40 mm Hg
pCO2 = 40 mm Hg 46 mm Hg

Oxigenul trece din aerul alveolar în sângele capilar, iar dioxidul de carbon trece din
sânge în alveole, până ce presiunile se egalizează. Deşi gradientul de presiune este pentru CO2
doar de 6 mm Hg, acesta difuzează de 20 de ori mai repede, deoarece este de 25 de ori mai
solubil decât O2 . astfel, la plămâni soseşte sânge venos, adus de arterele pulmonare şi după
efectuarea schimbului de gaze, sângele încărcat cu CO2 se oxigenează, se transformă în sânge
arterial. Acesta, îmbogăţit cu oxigen, părăseşte plămânul pentru a ajunge în circulaţia
sistemică.
Vascularizaţia plămânilor
Plămânii prezintă o vascularizaţie dublă: nutritivă şi funcţională.
Vascularizaţia nutritivă este parte a marii circulaţii şi este asigurată de arterele şi venele
bronşice. Arterele bronşice, ramuri ale aortei toracice, aduc sânge oxigenat pentru arborele
bronşic, parenchimul pulmonar şi pereţii arterelor pulmonare. Ele pătrund prin hil, însoţind
arborele bronşic până la nivelul bronhiolelor respiratorii, nivel la care se capilarizează. De aici
pornesc venele bronşice care colectează sângele încărcat cu CO2 pe care îl drenează, vărsându-l
în vena cavă superioară prin intermediul venelor azigos.
Vascularizaţia funcţională sau mica circulaţie asigură schimburile gazoase pulmonare.
Originea micii circulaţii este ventriculul drept de unde pleacă trunchiul pulmonar care aduce la
plămân sânge încărcat cu CO2. acesta se împarte în arterele pulmonare dreaptă şi stângă care
pătrund prin hil în plămân, însoţind arborele bronşic până la nivelul alveolelor, unde formează
reţeaua capilară perialveolară. La acest nivel este cedat CO2 şi preluat O2. în continuare, se
formează venele pulmonare, câte două pentru fiecare plămân, care aduc oxigen în atriul stâng.
Inervaţia plămânului provine din plexul bronho-pulmonar. Simpaticul produce
bronhodilataţie şi parasimpaticul are efecte inverse – bronhoconstricţie. Pleura este o seroasă ce acoperă plămânii şi care prezintă o foiţă viscerală, în raport cu
plămânii, şi una parietală care tapetează cavitatea toracică. Între cele două pleure se află un
strat fin de lichid pleural ce le ţine în contact intim şi permite alunecarea lor în timpul
mişcărilor respiratorii. Presiunea din interiorul acestor foiţe este mult mai scăzută decât cea
atmosferică.

Sistemul toraco-pulmonar are rolul de a asigura realizarea schimbului de gaze între
mediul extern şi cel intern.
10.2. Motilitatea sistemul toraco-pulmonar
Mişcarea sistemul toraco-pulmonar este asigurată de grupe musculare care prin
contracţia lor scot acest sistem din starea lui de repaus – aceasta fiind considerată poziţia 0 la
nivelul capilarelor reziduale funcţionale (CRF).
Acest sistem se află în echilibru, deşi toracele şi plămânul se dezvoltă inegal, iar forţele
lor elastice tracţionează în sensuri opuse. Pentru a se produce respiraţia este necesar ca grupele
musculare să se contracte, determinând variaţii ale celor trei diametre ale cutiei toracice:
- vertical: cranio-caudal prin coborârea diafragmului;
- sagital: prin ridicarea sternului (muşchii scaleni şi sternocleidomastoidieni –
ridicarea a şase coaste);
- transversal: ridicarea coastelor 2-6 determină ascensiunea şi rotarea pe un ax antero-
posterior a coastelor 7-10.
Coastele 11 şi 12 nu participă la respiraţie, ci constituie punct fix pentru musculatura
care realizează mişcările respiratorii. Coloana vertebrală nu participă la respiraţia obişnuită
ci numai la cea forţată, se extinde în inspir şi se flectează în expir.
Din punct de vedere funcţional, musculatura se va împărţi în patru grupe principale:
1. musculatura inspiraţiei liniştite;
2. musculatura inspiraţiei profunde;
3. musculatura inspiraţiei forţate;
4. musculatura expiraţiei forţate;
1. Musculatura inspiraţiei liniştite
Tradiţional se poate vorbi de o respiraţie diafragmatică şi una costală, dar în practică
se întâlneşte o combinaţie a acestora.
Musculatura inspiraţiei liniştite este reprezentată de diafragm, muşchii intercostali şi
scaleni.
Diafragmul are forma unei cupole, cu o bază de inserţie largă, în jurul bazei toracale,
anterior pe faţa posterioară a apendicelui xifoid, pe faţa internă a ultimelor două arcuri costale,
şi posterior se inseră pe coloana lombară prin două pliuri, dreapta şi stânga. Fibrele converg

spre o formaţiune aponevrotică numită tendon central sau centrul frenic, ce formează o cupolă
ce priveşte antero-inferior (în repaus) în dreptul coastelor 5-7.
Diafragmul este străbătut de artera aortă, vena cavă inferioară şi esofag şi este inervat
de nervul frenic. Când se contractă se produce o coborâre a tendonului central. Mişcările
diafragmului pot fi sistematizate în trei timpi (L. Barnier):
a) fibrele muşchiului sunt relaxate şi centrul frenic atinge cel mai înalt nivel –
expiraţia;
b) stâlpii se contractă şi centrul frenic este uşor tras în jos – începutul inspiraţiei;
c) fibrele anterioare şi laterale se contractă, centrul frenic rămâne imobilizat –
inspiraţie completă.
Muşchii intercostali sunt în număr de 11 perechi de muşchi pentru fiecare hemitorace.
Sunt situaţi între marginea inferioară a coastei supraiacente şi marginea superioară a coastei
subiacente. Se descriu trei categorii de muşchi intercostali: externi, mijlocii şi interni.
Fasciculele externe sunt orientate de sus în jos şi dinapoi – înainte, cele interne tot de sus în jos
şi dinainte – înapoi. Rolul lor este încă discutabil, însă importanţă majoră au în inspiraţie.
Scalenii sunt în număr de trei perechi de muşchi: anteriori, mediani şi posteriori. Au
inserţie posterior, pe apofizele spinoase C2 – C7 şi inserţi este anterioară, pentru fasciculele
anterioare şi mediene pe prima coastă, iar scalenul posterior pe coasta a doua. Pot interveni în
inspiraţia liniştită printr-o contracţie izometrică, fixând toracele şi favorizând mişcările
diafragmului. În inspiraţia forţată contribuie la ridicarea toracelui.
2. Musculatura inspiraţiei profunde
În cadrul inspirului forţat se produce o intensificare a activităţii musculaturii
prezentate anterior, la care se adaugă:
- sternocleidomastoidienii ce acţionează prin ridicarea toracelui, luând punct fix pe
mastoidă;
- ridicătorii coastelor vor ascensiona coastele şi vor extinde coloana vertebrală
mărind amplitudinea inspiraţie;
- micul dinţat postero-superior, ce ridică porţiunea superioară a primelor coaste;
- muşchii toracospinali intervin secundar în inspiraţie, ei extinzând coloana
vertebrală şi contrabalansează aplecarea trunchiului în faţă, în condiţiile în care
există un abdomen mărit de volum sau când este purtată o greutate.

Inspiraţia profundă poate fi definită ca fiind inspiraţia unei persoane sănătoase când
execută un efort mediu.
3. Musculatura inspiraţiei forţate
Inspiraţia forţată apare în condiţiile unui efort de intensitate mare sau la bolnavii
bronhopulmonari.
Muşchii care contribuie la acest tip de inspiraţie sunt reprezentaţi de: marele şi micul
pectoral, trapezii, romboizii şi marele dinţat, având rol în ridicarea coastelor, mărind
diametrul transvers.
4. Musculatura expiraţiei forţate
Expiraţia este un proces pasiv, devenind un proces activ, forţat, în condiţii patologice sau în
cadrul unui efort fizic intens, la o persoană sănătoasă. Cercetări anterioare au demonstrat,
prin plasarea unor electrozi pe grupele musculare inspiratorii, că expiraţia are loc ca urmare
a relaxării treptate a acestei musculaturi.
Aceşti muşchi sunt situaţi la nivelul peretelui abdominal.
- Muşchii transverşi abdominali care inseră pe faţa posterioară a ultimelor două coaste,
pe apofizele transverse L1 – L5, pe faţa internă a crestelor iliace şi 1/3 internă a arcadei crurale,
având ca punct de origine linia albă. Acţiunea sa este sinergică, ca o chingă presând asupra
viscerelor abdominale şi va împinge în sus diafragmul. Când se contractă atât diafragmul cât şi
transversul, respiraţia va fi blocată, situaţie întâlnită în: defecaţie, ridicarea de obiecte grele,
ţipăt, tuse.
- Muşchii drepţi abdominali
- Muşchii oblici interni şi externi
- Muşchiul dinţat mic postero-inferior
- Muşchiul pătratul lombelor
Toţi aceşti muşchi contribuie la coborârea coastelor şi la scăderea diametrul transvers,
facilitând expirul.

Unitatea de curs 11 Aparatul cardiovascular. Inima �i principalele vase de
sânge şi limfatice
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului aparatului cardiovascular.
Obiectivele operaţionale
1. Prezentarea structurii aparatului cardiovascular.
2. Prezentarea clasificării aparatului cardiovascular.
3. Prezentarea rolului şi motilităţii aparatului cardiovascular.
Aparatul circulator sanguin este alcătuit dintr-un organ central, cu rol de pompă
aspiro-respingătoare, inima sau cordul şi un sistem organizat de canale tubulare, numite vase
sanguine, cuprinzând artere şi vene, iar între ele o bogată reţea de vase intermediare foarte
fine, numite vase capilare. Acestea au un rol considerabil în contextul schimburilor nutritive de
la nivelul ţesuturilor, în cadrul metabolismului celular prin importul unor materii folositoare
organismului şi eliminarea altora, spre organele de excreţie. Integritatea sistemului circulator
condiţionează o bună funcţionalitate a organismului.
În cadrul aparatului circulator sanguin, deosebim două circuite deosebite ca
funcţionalitate: unul între cord şi plămâni, care asigură oxigenarea sângelui, reprezentând mica
circulaţie – sau circulaţia pulmonară – şi altul între cord, viscere şi periferia organismului,
reprezentînd marea circulaţie sau circulaţia generală.
Pentru circulaţia limfatică organismul dispune de un sistem de capilare limfatice, vase
şi ganglioni limfatici cu originea la nivelul lichidului interstiţial al ţesuturilor şi cu sens
centripet, prin două trunchiuri colectoare limfatice, spre sistemul venos al marii circulaţii.
Inima este un organ cavitar, eminamente muscular, numit muşchiul cardiac sau
miocardul, căptuşit la interior de endocard şi la exterior de epicard şi care se află în continuă şi
ritmică activitate. Inima este situată în treimea inferioară a mediastinului toracic anterior,
sprijinindu-se pe diafragm şi învelită într-un sac fibros – sacul pericardic. Acesta prezintă la
exterior pericardul fibros, iar în interior pericardul seros, care căptuşeşte, pe de o parte, sacul

fibbros şi formează foiţa parietală seroasă, iar apoi se răsfrânge pe inimă şi formează foiţa
viscerală, denumită epicard.
Inima are forma unui trunchi de con globulos, cu axul lung îndreptat oblic în jos şi spre
stânga, o capacitate medie la adult de 500-600 ml şi o greutate de 250-300g.
Ea prezintă:
- o faţă anterioară (sternocostală),
- una posterioară (pulmonară) şi
- alta inferioară (diafragmatică);
- o bază la nivelul careia se găsesc vasele mari,
- un vârf (apex) care aparţine ventriculului stâng şi care se proiectează în spaţiul V intercostal
stâng şi
- două margini.
Din punct de vedere morfologic, inima este împărţită de un sept longitudinal, în:
- inima dreaptă, conţinând sânge venos pe care îl trimite în mica circulaţie
- inima stângă, cu sânge arterial care este trimis pe calea aortică în marea circulaţie.
De asemenea, un sept transversal împarte fiecare sector în câte două cavităţi:
- una atrială situată spre baza inimii, cu pereţi mai subţiri şi cu o prelungire exterioară –
urechiuşa sau auricula
- una ventriculară, situată spre vârful inimii şi cu pereţii mult mai groşi.
La nivelul septului transversal se află orificiile de comunicare atrioventriculare,
prevăzute cu câte un aparat valvular: la stânga valva bicuspidă (mitrală) cu două valvule şi la
dreapta valva tricuspidă cu trei valvule, de vârful cărora se prind cordajele tendinoase ale
muşchilor papilari.
Ventriculul stâng mai prezintă un orificiu pentru artera aortă, prevăzut cu trei valvule
semilunare, iar ventriculul drept, un orificiu pentru artera pulmonară, prevăzut cu trei valvule
semilunare. Buna funcţionare a aparatului valvular favorizează umplerea cu sânge a inimii în
perioada de repaus (diastolă) şi propulsarea sângelui după contracţie în marea şi mica
circulaţie.
Inima conţine şi un aparat propriu de conducere, format din ţesutul nodal, care asigură
funcţional automatismul cardiac.

Inima este singurul organ muscular care se contractă ritmic, fără întrerupere din viaţa
embrionară şi până la naştere.
a. Miocardul – peretele muscular al inimii.
În structura miocardului se disting două varietăţi de ţesut muscular:
- Ţesut cardiac – formează cea mai mare parte a miocardului. Este format din fibre striate
cardiace; în atrii fibrele sunt dispuse circular iar în ventricule sunt dispuse oblic-
spiralat. Nu există continuitate între miocardul atriilor şi al ventriculelor, separarea fiind
făcută prin ţesut conjunctiv-fibros.
- Ţesut nodal - aparatul de conducere al inimii. În inimă există doi noduli: nodulul
sinoatrial, în atriul drept, în vecinătatea vărsării venei cave superioare şi nodulul
atrioventricular care pleacă din peretele atrioventricular şi se împarte în două ramuri,
stângă ţi dreaptă, ce coboară în ventricule. Cele două ramuri se divid formând reţeaua
Purkinje.
Proprietăţile miocardului sunt comune cu ale muşchilor striaţi scheletici (excitabilitatea,
conductibilitatea, contractilitatea) şi proprii (automatismul).
Excitabilitatea – proprietatea miocardului de a răspunde printr-o contracţie la stimuli
adecvaţi.
Conductibilitatea – proprietatea miocardului , în special a ţesutului nodal de a conduce
unde de contracţie de la nodulul sinoatrial în întreg cordul.
Contractilitatea – proprietatea miocardului de a se contracta atunci când este stimulat
adecvat.Contracţiile se numesc sistole iar relaxările diastole.
Automatismul (ritmicitatea) – proprietatea cordului de a se contracta succesiv ca urmare
a impulsurilor generate de nodulul sinoatrial. Aceste impulsuri sunt urmarea unor modificări
metabolice care au loc în ţesutul excito-conductor.
Vascularizaţia inimii - foarte bogată, asigurată de cele două artere coronare ce se
desprind de la originea aortei.
Inervaţia inimii – realizată prin fibre vegetative simpatixce şi parasimpatice, formând
plexul cardiac.
Vasele sanguine - sunt reprezentate de artere, arteriole, capilare, venule şi vene.
Arterele sunt vase care conduc sângele de la inimă spre ţesuturi.

Capilarele – formează o reţea foarte fină la nivelul ţesuturilor, realizând legătura între
arteriolă şi venulă.
Venele – colectoare convergente ale sângelui de la periferie spre atriul drept. Venele în
care circulaţia se face în sens opus gravitaţiei (membre şi vena cavă inferioară), prezintă în
interior cute valvulare.
Vasele sanguine prezintă 3 tunici: endotelială (intima), musculo-elastică (media) şi
adventicea (externă).
Sistemul circulator este reprezentat de:
- mica circulaţie
- marea circulaţie
Mica circulaţie sau sistemul circulator pulmonar – conduce sângele venos din
ventriculul drept la plămâni prin artera pulmonară, care se divide în artera pulmonară dreaptă
cu trei ramuri lobare şi artera pulmonară stângă cu două ramuri lobare.
Marea circulaţie sau sistemul circulator aortic – conduce sângele oxigenat din
ventriculul stâng, pe calea arterei aorte în întreg organismul.
Artera aortă prezintă la ieşirea din ventriculul stâng o dilataţie – bulbul aortic – din care
pleacă cele 2 artere coronare dreaptă şi stângă, pentru muşchiul cardiac.
În continuare se află aorta ascendentă, arcul/cârja aortică, apii aorta descendentă cu o
porţiune toracică şi una abdominală. Aorta abdominală ţine pâna la vertebrele L4-L5, unde se
bifurcă în cele 2 artere iliace.
Din trunchiul aortic pornesc:
- Trunchiul brahiocefalic arterial care se bifurcă în: artera carotidă comună dreaptă şi
artera subclaviculară dreaptă;
- Artera carotidă comună care se ramifică în:
- carotida externă, care se ramifică în a.tiroidiană, linguală, facială, auriculară,
faringiană şi occipitală;
- carotida internă, care irigă creierul şi globul occipital.
- Artera subclaviculară irigă: encefalul, gâtul şi umărul.
- Artera axilară dă ramuri pentru umăr şi se continuă cu artera humerală (brahială) care se
bifurcă la nivelul cotului dând artera radială şi ulnară

- Aorta toracică dă ramuri parietale (artere intercostale) şi ramuri viscerale pentru esofag,
pericard, mediastin şi frenice superioare
- Aorta abdominală irigă organele abdominale
- Artera iliacă comună se bifurcă în:
- artera iliacă internă/hipogastrică, cu ramuri parietale şi viscerale
- artera iliacă externă care se continuă cu artera femurală şi irigă membrul
inferior, prin artera poplitee, artera tibială anterioară, artera dorsală a piciorului şi trunchiul
tibio-fibular, arterele plantare, arcul plantar şi arterele digitale.
Sistemul venos al marii circulaţii – conduce sângele venos prin cele 2 vene cave
superioară şi inferioară către atriul drept.
Vena cavă superioară (cranială) – colectează sângele de la cap, gât, membrul superior
şi torace.
Vena cavă inferioră (caudală) – colectează sângele din regiunea supradiafragmatică a
corpului.
Sistemul venos porthepatic – colectează sângele venos de la nivelul tubului digestiv, din
regiunea subdiafragmatică, prin venele mezenterică superioară, mezenterică inferioară şi vena
splenică.
Vena iliacă comună – formată din iliaca internă şi iliaca externă, colectează sângele
venos de la nivelul bazinului şi al membrului inferior.

Unitatea de curs 12 Aparatul digestiv. Glandele anexe ale tubului digestiv
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului aparatului digestiv şi al glandelor anexe.
Obiectivele operaţionale
2. Prezentarea structurii aparatului aparatului digestiv şi al glandelor anexe.
2. Prezentarea clasificării aparatului aparatului digestiv şi al glandelor anexe.
3. Prezentarea rolului şi motilităţii aparatului digestiv şi al glandelor anexe.
Aparatul digestiv este alcătuit din totalitatea organelor în care are loc transformarea
alimentelor în substanţe simple asimilabile de către celule.
Este alcătuit din două părti: tubul digestiv şi glandele anexe.
Tubul digestiv este alcătuit din mai multe segmente: cavitatea bucală, faringele,
esofagul, stomacul, intestinul subţire şi intestinul gros. Aceste segmente se înlănţuie sub forma
unui tub continuu de calibru diferit, începând de la orificiul bucal până la orificiul anal.
De la orificiul bucal până la orificiul anal, tubul digestiv străbate craniul viscerial,
gâtul, toracele(mediastinul), cavitatea abdominală, marele şi micul bazin. Cea mai mare parte a
tubului digestiv este în cavitatea abdominală, care este delimitată de partea superioară de
muschiul diafragm, iar în cea inferioară comunicând cu cavitatea pelviană, de unde şi
denumirea de cavitate abdomino-pelviană.
Glandele anexe ale tubului digestiv.
În tubul digestiv se găsesc numeroase glande, unele în pereţii săi, altele constituind
organe glandulare anexe. Glandele anexe din peretii tubului digestiv sunt răspândite îndeseobi
în tunica mucoasă şi în unele segmente (esofag şi duoden), acestea pătrund până în tunica
submucoasă.

Figura 17 Tubul digestiv şi glandele anexe
Glandele mari dinafara pereţilor
tubului digestiv, numite glande anexe,
comunică cu cavitatea segmentului digestiv
respectiv prin conducte excretoare. În această
categorie intră glandele salivare, ficatul şi
pancreasul. Glandele salivare sunt: mici şi
mari (glanda parotidă, submandibulară,
sublinguală).
Tractul gastrointestinal superior este
format din:
- cavitatea bucală - aici alimentele sunt
marunţite cu ajutorul dinţilor şi limbii şi sunt
amestecate cu salivă. Este delimitată de 5
pereţi: unul anterior – buzele, doi laterali –
obrajii, unul inferior – planşeul bucal şi unul
superior – bolta sau vălul palatin.
- esofag - parte a tractului digestiv, este
un tub musculomucos, lung de 25-30 cm cu
funcţia de transport a hranei şi care leagă
faringele de stomac. Pereţii săi musculari produc contracţii ondulatorii, care ajută la
transportarea alimentelor.
- stomac - aici hrana este transformată în pastă, cu ajutorul acizilor existenţi la acest
nivel.
Tractul gastrointestinal inferior este format din:- intestinul subţire - reprezintă
segmentul aparatului digestiv, situat între stomac şi intestinul gros. Aici sunt absorbite
majoritatea lichidelor şi mineralelor din mâncare. Prezintă o serie de inflexiuni, numite anse
intestinale şi este format din 3 părţi:
o duoden - segment iniţial al intestinului subţire cu lungime de 25 – 30 de cm şi
formă de potcoavă deschisă, ce leagă stomacul de jejun. Aici incepe procesul de

digestie al mâncării, rolul duodenului fiind acela de a neutraliza aciditatea din
hrană.
o jejun - partea centrală a intestinului subţire, fiind situat între duoden şi ileon.
Rolul lui este de a absorbi substantele nutritive;
o ileon - ultima parte a intestinului subţire. Aici are loc absorbţia nutrienţilor
rezultaţi în urma digestiei.
- intestinul gros - parte a aparatului digestiv, cuprins între intestinul subţire şi
anus. Aici are loc absorbţia apei şi a mineralelor neabsorbite. Intestinul gros este format din:
o cecum (cecul) - prima parte a intestinului gros, situat între ileon şi colon. De
cecum este ataşat apendicele. În cecum se află flora intestinală;
o colon (colon ascendent, colon transvers, colon descendent, colon sigmoid) – la
acest nivel este absorbită apa ramasă în alimente;
o rect - canal cu pereţi musculoşi unde sunt depozitate materiile fecale;
- anus - partea finală a rectului, folosit pentru eliminarea materiilor fecale.
Organele anexe ale aparatului digestiv au rol doar în digestie. Prezentăm ficatul, vezica
biliară şi pancreasul:
- ficatul - produce bila, neutralizează toxinele din organism, prelucrează şi depozitează
numeroase vitamine şi o serie de microelemente esenţiale;
- vezica biliară (colecistul) - rezervor temporar al bilei. Este unită cu ficatul, de unde
primeşte bila, şi cu duodenul, unde eliberează bila, necesară digestiei grăsimilor;
- pancreasul (figura 18) - organ care îndeplineşte două funcţii majore: exocrină
(produce sucul pancreatic care conţine enzime digestive) şi endocrină (produce mulţi
hormoni importanţi, incluzând şi insulina).

Figura 18 Pancreasul

Unitatea de curs 13 Sistemul endocrin
Scopul unităţii de curs
Înţelegerea structurii, funcţionalităţii şi rolului sistemului endocrin.
Obiectivele operaţionale
3. Prezentarea structurii aparatului sistemului endocrin.
2. Prezentarea clasificării aparatului sistemului endocrin.
3. Prezentarea rolului şi motilităţii sistemului endocrin.
Glandele endocrine (endo = înăuntru, krino = a secreta) sau glandele cu secreţie
internă sunt organe specializate în secreţia unor substanţe cu acţiune specifică, denumite
hormoni (ormao = a excita), pe care-i varsă direct în sânge. Prin hormoni se înteleg substanţe
elaborate de un anumit organ sau de anumite teritorii celulare, care acţionează la distanţă
asupra altor organe sau ţesuturi, exercitând o acţiune caracteristică metabolică sau
morfogenetică, în sens stimulator sau inhibitor.
Acest sistem prin elaborarea hormonilor specifici, realizează coordonarea hormonală a
funcţiilor organismului, în strânsă dependenţă cu coordonarea nervoasă. Coordonarea nervoasă
este complexă, mult diferenţiată şi rapidă, în timp ce coordonarea endocrină, controlată la
rândul său de sistemul nervos, este auxiliară reglării nervoase, continuuă şi lentă. Buna
funcţionare a glandelor endocrine este o condiţie esenţială pentru realizarea coordonării
umorale. Glandele endocrine sunt: hipofiza, epifiza, tiroida, paratiroidele, timusul,
suprarenalele, partea endocrină a pancreasului şi a glandelor genitale.
Hipofiza.
Glanda hipofiză sau creierul endocrin are un rol important în reglarea funcţiilor
metabolice ale organismului, atât prin legăturile sale intime cu sistemul nervos central, cât şi
prin interrelaţiile cu majoritatea celorlaltor glande endocrine. Ea este situată în cutia craniană,
la baza diencefalului, înapoia chiasmei optice şi înaintea corpilor mamilari, îm şaua turcească a
osului sfenoid. Hipofiza are mărimea unui bob de fasole, este legată de partea bazală a
hipotalamusului (tuber cinereum), printr-o formaţiune numită tija hipofizară şi este formată din

3 lobi (fig. 19): anterior (adenohipofiza), posterior (neurohipofiza) şi lobul intermediar care la
om este rudimentar.
Figura 19 Hipofiza Figura 20 Tiroida
Adenohipofiza.
Adenohipofiza reprezintă partea secretorie a glandei şi cuprinde 75 % din întreaga
glandă. Este alcătuită din cordoane celulare neregulate, ce vin în relaţie strânsă cu reţeaua de
capilare sinusoide dintre ele. În funcţie de afinitatea pentru coloranţi se disting 2 tipuri de
celule: cromofobe şi cromofile. Celulele cromofobe sunt considerate celulele secretorii imature,
iar celulele cromofile sunt celule mature, secretoare ale hormonilor şi sunt la rândul lor
acidofile şi bazofile. Produsul de secreţie al fiecărui tip de celulă este un hormon specific,
numit hormonul trop. Aceşti hormoni influenţează atât creşterea, cât şi activitatea celorlaltor
glande şi sunt reprezentaţi de: hormonul de creştere sau somatotrop (STH), hormonul tireotrop
(TSH), hormonul adenocorticotrop (ACTH) şi hormonii gonadotropi (PSH şi ICSH).
Între adenohipofiză şi hipotalamus există o strânsă corelaţie funcţională, realizată
printr-o legătură vasculară denumită sistemul port-hipotalamo-hipofizar descris în 1929 de
românul Gr. T. Popa şi englezul Aiuna Fielding. Acest sistem îşi are originea într-un plex
capilar din hipotalamus, din care iau naştere vase portale, care la rândul lor se vor capilariza în
hipofiză. Unii neuroni hipotalamici elaborează diferite substanţe chimice (neurosecreţie) pe
care le descarcă în capilarele hipotalamice şi care prin vasele portale, ajung în hipofiza
anterioară, de unde neurosecreţiile ajung în ţesutul gandular. Produşii de neurosecreţie au
proprietatea de a stimula secreţia celulelor adenohipofizare (hormoni hipotalamici de
eliberare) sau de a inhiba secreţia celulelor adenohipofizare (hormoni hipotalamici inhibitori).

Lobul intermediar.
Lobul intermediar este o lamă subţire de ţesut epitelial glandular care aderă de lobul
anterior şi de neurohipofiză şi reprezintă numai 2 % din masa hipofizei. Secretă homonul
stimulator al melanocitelor (MSH).
Neurohipofiza.
Neurohipofiza este alcătuită dintr-o stromă conjunctiv vasculară, din celule gliale şi din
numeroase fibre nervoase care sunt prelungiri axonice ai neuronilor hipotalamici (nucleul
supraoptic şi nucleul paraventricular) care au proprietăţi neurosecretoare. Produsul de
neurosecreţie reprezentat de hormonul antidiuretic (ADH) şi ocitocină, este condus pe cale
axonilor neuronilor secretori ce alcătuiesc tractul hipotalamo-hipofizar, către hipofiza
posterioară, unde este depozitat şi de unde este eliberat îm sânge după necesităţi.
Epifiza.
Epifiza are formă conică asemănătoare unui con de pin (glanda pineală) şi este situată
în şanţul median dintre tuberculii cvadrigemeni superiori. La exterior este învelită într-o
capsulă, formată din piamater, ce trimite în interior septuri ce împart parenchimul glandei în
lobi. Prin baza sa aderă de o regiune diencefalică cu care alcătuieşte epitalamus. Parenchimul
este format din celulele principale (pinocite), celule gliale, celule pigmentare şi celule
nervoase, la care se adaugă celule conjunctive, vase de sânge, fibre vegetative postganglionare
simpatice. După naştere în epifiză au loc depuneri treptate de săruri de calciu şi de magneziu,
astfel că după pubertate ea poate fi calcifiată în întregime. Epifiza secretă mai mulţi hormoni,
dintre care cel mai cunoscut este malatonina.
Glanda tiroidă
Glanda tiroidă (fig. 20) este cea mai mare dintre glandele endocrine, având la adult o
greutate de 40-60 g. (la femeie este puţin mai voluminoasă decât la bărbaţi); situată în regiunea
anterioară şi laterală a gâtului în loja tiroidiană, pe faţa anterioară a laringelui şi traheei. Are
forma literei "H" şi prezintă 2 lobi laterali (stâng şi drept), legaţi printr-o porţiune numită istm.
La exterior prezintă o capsulă fibroasă care trimite la interior septuri conjunctive, ce împart
parenchimul tiroidian în lobuli. În structura unui lobul se găseşte ţesut conjunctiv (stroma

conjunctivă) şi ţesut glandular propriu-zis (parenchimul glandular). Lobulii sunt alcătuiţi din
vezicule sau foliculi tiroidieni. Peretele foliculului este mărginit la periferie de o membrană
bazală pe care spre interior sunt aşezate celule epiteliale tiroidiene, de formă cubică în perioada
de secreţie redusă, dar care devin înalte în perioada de activitate crescută. În cavitatea
foliculului se găseşte o substanţă gelatinoasă, omogenă care poartă numele de holoid, care este
în cantitate redusă când glanda este foarte activă; el conţine iod, care este legat chimic de o
proteină denumită tiroglobulina ce conţine hormonul tiroidian (tiroxina) şi alte substanţe iodate
(diiodotironina şi triiodotironina). În stroma conjunctivă dintre foliculii tiroidieni se găsesc
celule C, ce sintetizează calcitonina (secretată şi de paratiroide).
Glandele paratiroide.
Glandele paratiroide sunt 4 formaţiuni mici, ovoidale, cu diametrul de 0,5 cm., de
culoare galbenă-brună, aşezate perechi pe faţa posterioară a lobilor glandei tiroide, în afara
capsulei acesteia. Două corespund lobului stâng şi două corespund lobului drept al tiroidei.
Glandele paratiroide sunt învelite la exterior de o capsulă fibroasă proprie ce trimite în interior
septuri conjunctive, care împart glanda în lobuli. Celulele glandulare ce alcătuiesc
parenchimuri sunt de două tipuri: celule principale, care secretă hormonul paratiroidian şi
celule oxifine, dispuse în cordoane care probabil sunt formele îmbătrânite ale celor principale.
Există şi celule parafoliculare identice cu celulele C de la tiroidă, secretoare de calcitonină.
Timusul.
Timusul este un organ cu funcţie limfoidă şi endocrină, situat în etajului superior al
mediastinului anterior, retrosternal. El începe să se dezvolte în săptămâna 6 a vieţii
intrauterine, creşte până la pubertate, cănd începe să involueze transformându-se în cea mai
mare parte într-o masă de ţesut adipos, în care se mai păstrează la adult mici zone de ţesut
timic. La naştere timusul are 5-6 cm. lungime, greutate 10-15 g., culoarea roşie-cenuşie şi este
alcătuit din 2 lobi (drept şi stâng) uniţi între ei pe linia mediană şi acoperiţi de o capsulă
conjunctivă care trimite în interior prelungiri delimitând lobuli.
Lobulul timic prezintă o zonă corticală la periferie constituită din limfocite speciale
numite timocite şi o zonă medulară spre interior mai deschisă la culoare în care se află celulele
reticulare şi limfoblaşti. Pe lânga funcţia limfopoetică şi imunitară, timusul are o funcţie

endocrină (deşi nu a fost încă individualizaţ hormonul timic), prin care influenţează creşterea şi
maturarea sexuală.
Pancreasul endocrin.
Pancreasul endocrin este alcătuit din insulele de celule răspândite în interiorul lobulilor
pancreasului exocrin, numite insulele Langerhans. Aceste insule sunt alcătuite din cordoane
celulare care se anastomozează formând o reţea în ochiurile căruia se găsesc capilare sanguine.
Cordoanele glandulare se compun din 2 tipuri de celule: celule alfa, puţine la număr, ce secretă
glucagonul şi celule beta, ce reprezintă 75 % din celulele insulare şi sunt dispuse în zona
periferică a pancreasului endocrin, secretoare de insulină.
Glandele suprarenale.
Glandele suprarenale sunt 2 mase mici (10-12 g), de formă tringhiulară, situate
deasupra polului superior al rinichiului, de care sunt separate printr-un sept conjunctiv. Glanda
este alcătuită din 2 porţiuni: una situată la exterior în zona corticală, denumită
corticosupraranală şi alta în partea internă denumită medulosuprarenală. Corticala este de
origine mezodermică, ca şi glandele sexuale, iar medulara de origine nervoasă (ectodermică),
provenind din celulele crestelor ganglionare din care se dezvoltă şi sistemul nervos simpatic.
La exterior, glanda suprarenală este învelită într-o capsulă fibroasă din care pleacă tracturi
conjunctive în parenchimul glandular.
Corticosuprarenala. Are parenchimul format din celule epiteliale, dispuse în cordoane
separate prin sinusuri venoase. După modul cum sunt dispuse cordoanele, se disting 3 zone
care de la exterior spre interior sunt:
-zona glomerulară, mai subţire, alcătuită din celule mici dispuse în grupe ovoide sau
glomerulare şi secretă hormonii mineralocorticoizi, cel mai activ fiind aldosteronul;
-zona fasciculată, la mijloc, cea mai groasă, alcătuită din celule poliedrice, dispuse în
cordoane paralele şi anastomozate; secretă hormonii glucocorticoizi (cortizolul);
-zona reticulată, alcătuită din cordoane celulare anastomozate, între care se află o
bogată reţea capilară; prezintă cea mai mare dezvoltare la pubertate şi secretă hormonii 17-
cetosteroizi (androgeni şi estrogeni).

Medulosuprarenala. Este formată din cordoane celulare scurte ce reprezintă neuroni
simpatici postganglionari, care nu mai prezintă axoni, dar care secretă adrenalina şi
noradrenalina, pe care le varsă în reţeaua de capilare sinusoide, foarte densă la acest nivel.
Glandele sexuale.
Ovarul endocrin este reprezentat de celulele tecii interne a foliculilor ovarieni, care
secretă hormonii estrogeni - foliculina şi de celulele corpului galben care secretă progesteronul
sau luteina.
Testiculul endocrin este reprezentat de celulele interstiţiale Leydig, care se găsesc în
ţesutul conjunctiv dintre tubii seminiferi contorţi. Totalitatea celulelor interstiţiale secretă
testosteronul.

BIBLIOGRAFIE
Obligatorie:
1. Albu I., Georgia R., Anatomie topografică, Bucureşti, Ed. All, 1998
2. Baciu C., Anatomia funcţională şi biomecanica aparatului locomotor, Bucureşti, Ed. Sport-
Turism, 1977
3. Ifrim M. - Atlas de anatomie umană. vol. III - Sistemul nervos şi organele de simţ, Ed.
{tiinţifică şi Enciclopedică, Bucureşti, 1985
4. Maxim Gh., Anatomie II - Note curs, Bacău, 2003
5. Mişcalencu D., Maxim Gh. şi colab. - Anatomia omului, Ed. Didactică şi Pedagogică,
Bucureşti, 1983
6. Mogoş Gh., Ianculescu A. - Compendiu de anatomie şi fiziologie, Ed. {tiinţifică, Bucureşti,
1973
7. Papilian V., Anatomia omului, Vol. I şi II, Bucureşti, Ed. Didactică şi Pedagogică, 1982
8. Raveica G., Anatomie funcţională, Note de curs, Bacău 2004
9. Ranga V., Teodorescu-Exarcu I. - Anatomia şi fiziologia omului, Ed. Medicală, Bucureşti,
1969
10. Sbenghe T., Bazele teoretice şi practice ale kinetoterapiei, Bucureşti, Ed. Medicală, 1999
11. Voiculescu I.C., Petricu I.C. - Anatomia şi fiziologia omului, Ed. Medicală, Bucureşti,
1971
12. Zaharia C., Îndreptar de anatomie practică şi chirurgicală a membrelor, Bucureşti, Ed.
Paideia, 1994
Suplimentară, facultativă:
1. Moţet D. şi colab., Îndrumător terminologic pentru studenţii secţiilor de kinetoterapie,
Bacău, Ed. Deşteptarea, 1997
2. Popescu M., Artrologie şi biomecanică, Bucureşti, Ed. Scaiul, 1998
3. Pernkopf E., Atlas of topgraphical and applied human anatomy, Vols. 1 and 2, 3rd Ed. W.
Platzer, Munich, Ed. Urban & Schwarzenberg, 1989
4. Sbenghe T., Kinetologie profilactică, terapeutică şi de recuperare, Bucureşti, Ed. Medicală,
1987

5. Colecţia revistei Kinesitherapie scientifique.
Adrese internet: http:// www.ama_assm.org/atlas/newatlab
7. http:// www.bmj.com
8. http://www.ptcentral.com/muscles/musclehead.html
9. http://www.bartleby.com/107/
10. http://www.bio.psu.edu/faculty/strauss/anatomy/biology29.htm
11. http://www.exrx.net/Exercise.html
12. http://www.innerbody.com/tutorial2/tutorial.html
13. http://medlib.med.utah.edu/WebPath/HISTHTML/ANATOMY/ANATOMY.html
14. http://www.ptcentral.com/
15. http://www.rad.washington.edu/anatomy/index.html
16. http://www.scoi.com/index.html
17. http://www.sportsknee.com/kneeanatomy.htm
18. http://sportsmedicine.about.com/
19. http://www.swarminteractive.com/subscriptions/hip/hipânăt.html
20. http://www.wolf-heidegger.com/english/