UNIVERSITATEA „ŞTEFAN CEL MARE” SUCEAVAsilvic.usv.ro/cursuri/cbcp_div_ecol_sp.pdf ·...
65
UNIVERSITATEA „ŞTEFAN CEL MARE” SUCEAVA FACULTATEA DE SILVICULTURĂ MASTERAT CONSERVAREA BIODIVERSITĂŢII ŞI CERTIFICAREA PĂDURILOR DIVERSITATEA ECOLOGICĂ A SPECIILOR LEMNOASE Conf. dr. ing. Florin CLINOVSCHI - 2013 –
Transcript of UNIVERSITATEA „ŞTEFAN CEL MARE” SUCEAVAsilvic.usv.ro/cursuri/cbcp_div_ecol_sp.pdf ·...

UNIVERSITATEA „ŞTEFAN CEL MARE” SUCEAVA FACULTATEA DE SILVICULTURĂ
MASTERAT CONSERVAREA BIODIVERSITĂŢII ŞI CERTIFICAREA PĂDURILOR
DIVERSITATEA ECOLOGICĂ
A SPECIILOR LEMNOASE
Conf. dr. ing. Florin CLINOVSCHI
- 2013 –

Diversitatea ecologică a speciilor forestiere
CUPRINS
1. Prezentarea generală a speciilor forestiere lemnoase ……………………………….. 3 Generalităţi ……………………………………………………………………………… 3 Bazele morfologice ale studiului plantelor lemnoase …………………………………... 3 Sistematica speciilor forestiere ………………………………………………………….. 8 Ecologia speciilor forestiere …………………………………………………………….. 10
2. Răspândirea speciilor forestiere lemnoase …………………………………………... 12 Introducere ……………………………………………………………………………… 12 Areal. Noţiuni generale …………………………………………………………………. 12 Localizarea. Arealul unui taxon ………………………………………………… 12 Suprafaţa arealului ……………………………………………………………… 13 Endemismele ……………………………………………………………………. 14 Forma arealului …………………………………………………………………. 15 Infrataxoni: subspecii, rase geografice, ecotipuri şi varietăţi …………………… 16 Specii vicariante ………………………………………………………………… 17 Arealul genurilor ………………………………………………………………... 17 Arealul familiilor ………………………………………………………………... 18
3. De la arbore la arboret ………………………………………………………………… 19 Imobilitatea individului – mobilitatea populaţiei ……………………………………….. 19 Modalităţi de recucerire a spaţiului ……………………………………………………... 19 Activităţi ciclice în cadrul ecosistemului forestier ……………………………………… 20
4. Diversitatea ecologică a speciilor de răşinoase ………………………………………. 24 Caractere generale ale răşinoaselor ……………………………………………………... 24 Repartiţie geografică ……………………………………………………………………. 25 Emisfera nordică ………………………………………………………………... 25 Regiunile intertropicale …………………………………………………………. 26 Emisfera sudică …………………………………………………………………. 27 Familia Pinaceae ………………………………………………………………………... 27 Genul Abies ……………………………………………………………………... 27 Genul Keteleeria ………………………………………………………………... 32 Genul Cathaya ………………………………………………………………….. 32 Genul Pseudotsuga ……………………………………………………………... 32 Genul Tsuga …………………………………………………………………...... 33 Genul Picea …………………………………………………………………....... 34 Genul Larix ……………………………………………………………………... 37 Genul Cedrus …………………………………………………………………… 39 Genul Pinus ……………………………………………………………………... 40 Familia Taxodiaceae ……………………………………………………………………. 45 Familia Cupressaceae …………………………………………………………………... 46 Familia Taxaceae ……………………………………………………………………….. 47
1

Diversitatea ecologică a speciilor forestiere
2
5. Diversitatea ecologică a unor specii de foioase ………………………………………. 49 Caractere generale ale foioaselor ……………………………………………………….. 49 Familia Betulaceae ……………………………………………………………………… 49 Genul Carpinus …………………………………………………………………. 50 Genul Corylus …………………………………………………………………... 50 Genul Betula ……………………………………………………………………. 51 Genul Alnus …………………………………………………………………….. 52 Familia Fagaceae ……………………………………………………………………….. 53 Genul Fagus …………………………………………………………………….. 53 Genul Nothofagus ………………………………………………………………. 54 Genul Castanea …………………………………………………………………. 54 Genul Quercus ………………………………………………………………….. 55 Familia Salicaceae ……………………………………………………………………… 57 Genul Populus …………………………………………………………………... 57 Genul Salix ……………………………………………………………………… 58 Familia Ulmaceae ………………………………………………………………………. 59 Genul Ulmus ……………………………………………………………………. 59 Genul Celtis ……………………………………………………………………... 60 Familia Rosaceae ……………………………………………………………………….. 60 Genul Prunus …………………………………………………………………… 60 Familia Aceraceae ………………………………………………………………………. 61 Genul Acer ……………………………………………………………………… 61 Familia Tiliaceae ………………………………………………………………………... 62 Genul Tilia ……………………………………………………………………… 62 Familia Oleaceae ………………………………………………………………………... 63 Genul Fraxinus …………………………………………………………………. 63
Scurtă incursiune în pădurile canadiene ……………………………………………... 64

Diversitatea ecologică a speciilor forestiere
3
Capitolul 1. PREZENTAREA GENERALĂ A SPECIILOR FORESTIERE LEMNOASE
GENERALITĂŢI Studiul diversităţii ecologice la plantele lemnoase presupune, o abordare globală, de
ansamblu, a arborilor şi arbuştilor, căutând să explice prin intermediul exigenţelor ecologice ale plantelor şi al selecţiei naturale modalităţile de apariţie sau dispariţie, substituire sau asociere a acestora.
În fapt este o aprofundare ecologică a dendrologiei, disciplină care se ocupă cu studiul plantelor lemnoase din punct de vedere sistematic, morfologic, corologic (arealogic), ecologic, silvobiologic, economic şi peisager. Astfel, dendrologia se defineşte ca o disciplină de bază a silvicultorilor, o bună cunoaştere a acesteia conferind celor ce vor lucra în domeniul forestier temeinice noţiuni de specialitate cu scopul declarat de a recunoaşte, asocia şi promova speciile lemnoase.
Disciplina fundamentează această specializare şi are conexiuni directe cu botanica, pedologia, fitopatologia şi ecologia forestieră, având un caracter iniţiatic pentru discipline ca silvicultura, împăduririle, staţiunile forestiere, dendrometrie, amenajarea pădurilor.
Metodele de cercetare în domeniu sunt tipice domeniilor dendro-ecologice, dintre care amintim: − metoda morfologică şi metoda anatomică, ce sunt bazate pe studii comparative între specii
şi unităţi intraspecifice, − metoda investigaţiilor corelative şi deductive morfo-ecologice şi anatomo-ecologice, − metode de cercetare ecologice cantitative şi experimentale.
BAZELE MORFOLOGICE ALE STUDIULUI PLANTELOR LEMNOASE
Arbori, arbuşti, subarbuşti: definiţii, caractere generale Arborii sunt acele plante lemnoase care cresc obişnuit în păduri, au cel puţin 7 m în înălţime,
care produc lemn şi alte materii prime importante (coajă, răşini, flori) şi care exercită o influenţă benefică asupra ambientului.
Drept categorii de arbori, conform literaturii de specialitate amintim: arborii forestieri care produc lemn şi sunt parte integrantă din ecosistemul forestier, arborii ornamentali ce prezintă interes din punctul de vedere al aspectului, precum şi pomii fructiferi, destinaţi să producă fructe comestibile.
Forma arborilor diferă de la o specie la alta. Aceasta diferă în primul rând, după locul unde se creşte şi se dezvoltă exemplarele plantelor lemnoase. Se deosebesc astfel: - forma specifică (habitus), caracteristică arborilor crescuţi izolaţi, şi care au o tulpină scurtă, puternic îngroşată la bază, coroana mult dezvoltată în lăţime şi care coboară până aproape de sol - forma forestieră, întâlnită la arbori crescuţi în masiv, arbori ce au tulpina dreaptă, cilindrică, elagată cu coroana restrânsă spre vârf.
După înălţimea realizată, arborii sunt de mărimea I, când depăşesc 25 m, de mărimea a II-a când au între 15 şi 25 m şi de mărimea a III-a, când au între 7 şi 15 m.
Arbuştii sunt plante lemnoase cu înălţimea de până la 7 m, de regulă cu mai multe tulpini lemnoase ramificate de la bază, sub formă de tufă. Arbuştii cu înălţimi de sub 1 m intră în categoria arbuştilor pitici (afin, merişor, smirdar etc.).
Subarbuştii sunt plante ce au tulpina lemnoasă numai la bază, părţile superioare sunt erbacee, degerând peste iarnă (drobul, cătina mică etc.).

Diversitatea ecologică a speciilor forestiere
4
Rădăcina Rădăcina este un organ vegetativ lipsit de frunze cu geotropism pozitiv şi fototropism negativ
şi are ca funcţii: - fixează planta în sol şi asigură rezistenţa la vânt şi zăpadă, - absoarbe apa şi substanţe nutritive din sol transmiţându-le altor organe, - poartă raporturi de simbioză cu microorganismele din sol, - regenerarea vegetativă din muguri adventivi, drajoni şi butaşi.
În tinereţe se distinge o rădăcină principală care ulterior se ramifică în rădăcini laterale, acesta diferind de la specie la specie şi alcătuieşte sistemul de înrădăcinare. Ca tipuri, se remarcă: - pivotant, la care se distinge o rădăcină principală ca un pivot puternic ce poate depăşi 1 m (ex. brad, stejar), - trasant sau superficial, unde pivotul principal este slab dezvoltat, iar rădăcinile laterale se ramifică radial aproape de suprafaţa solului până la 50 cm (ex. molid), - pivotant-trasant sau mixt, unde pivotul principal este dezvoltat, iar rădăcinile laterale sunt puternice, dezvoltându-se fie către suprafaţa solului, cum este cazul la carpen, ulm, tei, paltin, anin alb, fie se dezvoltă în profunzime, cum este cazul laricelui, fagului şi al aninului negru.
Ca regulă generală, sistemele pivotant şi pivotant-trasant asigură arborilor o bună ancorare în sol. De asemenea, trebuie de precizat că sistemul de înrădăcinare variază cu vârsta şi după însuşirile fizice ale solului (profunzime, compactitate, umiditate etc.).
Rădăcinile adventive apar din muguri adventivi ce se formează pe tulpini, ramuri sau rădăcini şi stau la baza înmulţirii vegetative prin butaşi, marcote, drajoni. Tulpina
Tulpina este partea aeriană de deasupra coletului, cu creştere obişnuit verticală, cu formă de ax principal, în care se acumulează cea mai mare parte din masa lemnoasă produsă. Ea creşte datorită mugurilor terminali la ramificaţia monopodială sau a mugurilor laterali la ramificaţia simpodială. Arbuştii au tulpina care se ramifică de la bază, iar la liane care au un ţesut mecanic slab dezvoltat, este nevoie de suport pe care să se agaţe sau să se înfăşoare.
După direcţia de creştere, tulpinile pot fi: drepte, nutante, geniculate, tortuoase, volubile, scadente, radicante, repente, procumbente, ascendente etc.
Secţiunea transversală a tulpinii poate fi circulară, eliptică sau neregulat ondulată (canelată).
Scoarţa. Ritidomul Plantele lemnoase tinere au totdeauna scoarţa netedă şi poate fi în mod deosebit colorată, în
funcţie de specie. Odată cu vârsta, ţesuturile moarte ale scoarţei rezultate din activitatea felogenului, cumulat cu parenchimul şi liberul, generează ritidomul.
Ritidomul se poate exfolia circular (mesteacăn, cireş), în fâşii longitudinale (curpen, tuia), în solzi (molid, măr, platan). Ritidomul nu se exfoliază întotdeauna şi atunci formează nişte crăpături caracteristice, cum este cazul la ulm, cer, nuc, sau sub forma unor excrescenţe de suber, cum întâlnim la stejarul de plută şi arborele de plută de Amur. Unele specii nu formează ritidom, având toată viaţa scoarţa netedă (carpen, fag).
Coroana
Coroana este partea superioară a arborelui alcătuită din ramuri, frunze, flori şi fructe. Aceasta are o anumită formă, în funcţie de modul de aşezare al mugurilor, după desimea şi poziţia lujerilor şi după unghiul de inserţie a ramurilor pe tulpină: globuloasă, ovoidă, conică, columnară, tabulară, pendentă. Lujerii
Lujerii sunt porţiuni de tulpină sau ramuri formate în ultima perioadă de vegetaţie şi care poartă frunzele şi mugurii şi nu sunt ramificaţi. Lujerii de mai mulţi ani nu au muguri şi frunze şi se numesc ramuri.

Diversitatea ecologică a speciilor forestiere
5
Locul de inserţie al frunzelor, respectiv mugurilor, în lungul lujerilor se numesc noduri, iar porţiunile dintre noduri, internoduri.
Tipuri de lujeri După modul de creştere deosebim lujeri lungi – macroblaste – cu creşteri normale şi lujeri
scurţi – microblaste – cu creşteri de câţiva mm. Brachiblastele sunt ramuri rezultate din suprapunerea microblastelor, au un aspect inelat, noduros datorită cicatricelor frunzelor; poartă obişnuit flori, sunt terminate cu muguri şi uneori cu spini.
După poziţie, distingem lujeri terminali, crescuţi în prelungirea lujerului din anul precedent şi lujeri laterali, ce se formează lateral pe lujerii din anul precedent.
După provenienţă, deosebim lăstari care apar la baza tulpinii sau pe cioată din muguri adventivi/proventivi, lujeri lacomi ce apar de-a lungul tulpinii şi duc la coronarea arborelui şi drajoni care apar din mugurii adventivi ai rădăcinii.
După forma de creştere, lujeri pot fi drepţi (răşinoase), geniculaţi (ulm, tei, carpen), neregulat-curbaţi (soc).
După grosime, lujerii pot fi groşi (oţetar fals, roşcov de Canada, nuc), subţiri şi foarte subţiri (cătina roşie, floarea miresii).
După suprafaţa scoarţei, lujerii sunt netezi (majoritatea), striaţi, cu crăpături fine ca nişte zgârieturi (ulm), brăzdaţi sau sulcaţi cu nişte brazde longitudinale, muchiaţi (salbă moale), aripat-muchiaţi, ce au nişte excrescenţe suberoase de forma unor aripi (ulm de câmp, mături).
Culoarea scoarţei este în general brună, cu diferite nuanţe, dar mai poate fi: verde (sofora, afin), argintie (sălcioară), purpurie, discoloră (roşu şi verde ca la sânger). Uneori culoarea poate fi mascată de o brumă albăstruie (arţar american).
Scoarţa lujerilor este prevăzută cu lenticele de diferite mărimi şi forme (liniare, eliptice, rotunde), cu glande ceroase – verucozităţi (mesteacăn, salbă râioasă) – sau cu peri.
La unele specii lujerii se transformă în spini persistenţi (porumbar, glădiţă, maclura), cârcei (viţă-de-vie), filocladii (Ruscus sp.).
Prin zdrelire sau în tăietură proaspătă lujerii secretă un suc lăptos (familia Moraceae), iar alţii au un miros caracteristic (scumpie, mălin).
Spinii Unele specii au lujerii prevăzuţi cu spini. Aceştia pot lua naştere din modificarea:
- epidermei scoarţei, când sunt neregulat împrăştiaţi şi se desprind uşor de la bază (măceş, mur),
- stipelelor, când stau câte doi lateral faţă de mugure (salcâm), - nervurilor frunzei, când stau sub mugure (dracilă, agriş), - lujerilor, când sunt lateral faţă de mugure (păducel) sau deasupra mugurelui (glădiţă), - vârfului lujerilor lungi (verigariu) sau al brachiblastelor (păr),
Măduva Aceasta se observă cel mai bine prin secţiuni transversale sau longitudinale prin lujer. Ea
poate fi de culoare albă-gălbuie (majoritatea), roşcată (soc de munte), verzuie (păr), etc. şi este continuă (majoritatea), lamelar întreruptă (nuc), absentă (lujeri fistuloşi – caprifoi).
Mugurii
Mugurii sunt organe ce dau naştere la frunze, flori sau lujeri. Ei se formează încă din vară şi îşi ating deplina dezvoltare la sfârşitul perioadei de vegetaţie.
După organele la care vor da naştere, mugurii se clasifică în foliari (vegetativi), florali (floriferi) şi micşti. Ca provenienţă, mugurii sunt normali, cândapar încă din timpul verii, numai pe lujerii anuali, proventivi (dorminzi), ce nu se dezvoltă la exterior în anul următor formării lor, sunt acoperiţi de scoarţă şi pot rămâne timp îndelungat în stare latentă şi adventivi (întâmplători) care se formează neregulat pe tulpină şi rădăcină, generând lujeri sau rădăcini adventive.
După poziţia lor, deosebim muguri terminali (la vârful lujerilor) şi muguri laterali sau axilari (de-a lungul lujerilor).

Diversitatea ecologică a speciilor forestiere
6
După modul de repartizare, mugurii sunt alterni, aşezaţi izolat la diferite distanţe, distici, aşezaţi altern dar în acelaşi plan, opuşi, câte doi faţă în faţă, verticilaţi, câte trei sau mai mulţi, la acelaşi nivel pe lujer.
După modul de grupare, se disting muguri solitari, câte unul deasupra cicatricei, muguri suprapuşi, unul deasupra altuia (Lonicera sp.), muguri colaterali, câte 2-3 unul lângă altul (corcoduş).
După modul de inserţie pe lujer, mugurii se clasifică în sesili (majoritatea) şi pedicelaţi (anin negru, anin alb).
După mărime, mugurii sunt foarte mari, când au peste 2 cm (fag, magnolia, castan porcesc), mari, de 1-2 cm (scoruş păsăresc), mijlocii, sub 1 cm (majoritatea), mici, sub 1 mm (gărdurariţă), şi ascunşi (salcâm).
Mugurii sunt prevăzuţi la exterior cu solzi – catafile – în număr variabil (la sălcii unul aparent, la tei 2-3 solzi, la stejari numeroşi solzi) sau sunt nuzi (cruşân, dârmox).
Solzii pot fi largi, înguşti, acuţi, acuminaţi, rotunjiţi, cu margini întregi sau fin dinţate. Suprafaţa solzului poate fi glabră sau păroasă, uneori acoperită cu o substanţă vâscoasă. Frunza
Frunza este organul vegetativ cu structură dorso-ventrală şi simetrie bilaterală. Ea este alcătuită din limb, peţiol, teacă, la bază uneori cu stipele.
După lungimea peţiolului, frunzele sunt lung peţiolate, scurt peţiolate, sesile, decurente şi amplexicaule sau conate.
După organizare, frunzele sunt simple sau compuse. Cel compuse sunt de tip penat (pari- sau imparipenat) sau palmat.
După forma limbului, frunza poate fi rotundă, subrotundă, eliptică, ovată, obovată, oblongă, lanceolată, liniară, romboidală, triunghiulară, cordată sau reniformă.
Vârful limbului poate fi acut, acuminat, obtuz, rotunjit, trunchiat, emarginat, mucronat sau spinos.
Baza limbului poate fi rotunjită, cordată, reniformă, sagitată, hastată, trunchiată, îngustată, cuneată, asimetrică sau auriculată.
Marginea limbului este întreagă, simplu sau dublu serată, simplu sau dublu dinţată, crenată, sinuată, revolută, lobulată, ciliată.
Suprafaţa frunzei poate fi netedă, rugoasă (cu mici ridicături neregulate), plisată (vălurată), lucitoare sau nelucitoare, păroasă, scabră (aspră, datorită perilor rigizi, scurţi), glabră (lipsită complet de peri), glabrescentă (aproape lipsită de peri), glaucă sau glaucescentă (albăstrui).
După durata rămânerii pe ramuri, frunzele sunt caduce (căzătoare), persistente (sempervirescente), când ţin 2-12 ani şi marcescente, când se usucă toamna dar cad în primăvară (familia Fagaceae).
Floarea
O floare completă are următoarele părţi componente: învelişul floral sau periantul (caliciu şi corolă), elementele reproducătoare (androceu şi gineceu), receptaculul (axa florală).
Florile incomplete sunt acele flori cărora le lipsesc anumite elemente. Astfel, acestea pot fi apetale (învelişurile florale lipsesc parţial) sau nude (învelişurile lipsesc total), ca exemplu amintind că la familia Salicaceae locul învelişurilor florale este preluat de o bractee.
După repartiţia sexelor, florile sunt hermafrodite (bisexuate) şi unisexuate, acestea putând fi monoice (pe acelaşi individ apar flori ♀ şi ♂) şi dioice (florile ♀ şi cele ♂ sunt pe indivizi diferiţi). Drept cazuri speciale amintim arborii poligami (pe acelaşi individ flori bisexuate şi unisexuate) şi arborii trioici (pe exemplare diferite flori ♂, ♀ şi bisexuate).
Majoritatea gimnospermelor au flori unisexuat monoice lipsite de înveliş floral, unde florile mascule sunt grupate sub forma unor conuleţe sau amenţi cu stamine solziforme, iar florile femele au numeroşi solzi carpelari nesudaţi (nu formează ovar) dispuşi spiralat în jurul axului, iar ovulele rămân descoperite.

Diversitatea ecologică a speciilor forestiere
7
Inflorescenţa Florile pot fi aşezate câte una sau reunite mai multe, formând o grupare numită inflorescenţă.
La aceasta deosebim una sau mai multe axe, simple sau ramificate, pe care se prind florile cu ajutorul pedicelilor, iar întreaga inflorescenţă este purtată de un peduncul. Aceştia pot fi prevăzuţi la bază cu bractei, care uneori sunt aşezate la acelaşi nivel, formând un involucru.
Ca tipuri de inflorescenţe distingem inflorescenţe monopodiale, la care axul principal are creştere continuă, iar axele laterale se termină cu o floare şi inflorescenţe simpodiale, la care axul principal are o creştere definită terminându-se cu o floare, la fel şi cele secundare.
Tipurile de inflorescenţe întâlnite la plantele lemnoase sunt: - spicul, - amentul (mâţişorul) întâlnit la genurile Salix, Populus, Alnus, Corylus, Quercus etc., - racemul de la Prunus padus, Robinia pseudacacia, - paniculul la Sophora japonica, Ailanthus altissima, Rhus typhina, - corimbul la Acer platanoides, Sorbus aria, - umbela la Cornus mas, Spiraea ulmifolia, - fasciculul la Prunus avium, - capitulul la Fagus sylvatica (flori mascule), - cima la Euonymus europaeus. Fructul
După polenizare, la angiosperme are loc transformarea ovarului în fruct şi a ovulului în sămânţă.
De reţinut că, la gimnosperme, conul este doar organ de fructificaţie. După originea lor, după modificările suferite în timpul formării şi după modul de deschidere,
fructele se împart în următoarele categorii: - simple monocarpice, care pot fi dehiscente (Spiraea, Robinia) şi indehiscente (Sophora), - simple sincarpice, care, după consistenţa pericarpului, de subîmpart în uscate dehiscente (silicva, capsula), uscate indehiscente (achena, samara) şi fructe cărnoase (baca, drupa), - multiple (poliachenă la Clematis, polifoliculă la Spiraea, polidrupă la Rubus) - false (poama la genurile Malus, Pyrus, Cydonia şi Mespilus), - compuse (la genurile Morus, Ficus, Maclura şi Platanus). Maturaţie, maturitate, periodicitate
Maturaţia este perioada de timp ce trece de la polenizare şi până la coacerea fructelor. Aceasta poate avea loc la sfârşitul primăverii - începutul verii, în lunile mai-iunie (ulmii, plopii, sălciile), în toamna anului în care specia înfloreşte (majoritatea speciilor) sau în toamna celui de-al doilea an (Quercus cerris, Pinus sp.).
Maturitatea o constituie perioada de timp ce trece până când arborele începe să fructifice. Ea se poate realiza la vârste mici (5-6 ani la salcâm) sau, dimpotrivă, la vârste mai mari (50-70 ani la stejar, gorun).
Periodicitatea fructificaţiei se defineşte ca fiind numărul de ani care trec între două fructificaţii succesive, ea variind de la anual şi abundent (plopi, sălcii, salcâm, ulmi) la 7-12 ani (stejari). De specificat că la speciile cu o periodicitate mare, cum este cazul speciilor din familia Fagaceae, între două fructificaţii succesive abundente, pot avea loc fructificaţii mai slabe numite stropeli.
Diseminaţie, germinaţie, putere germinativă
Diseminarea fructelor se poate realiza prin: - simpla cădere şi rostogolire, - vânt (anemochor) – ulmi, tei, frasin, - apă (hidrochor) – anini, sălcii, plopi, - animale (zoochor) – vâsc, nuc, stejar, fag.

Diversitatea ecologică a speciilor forestiere
8
Germinaţia poate avea loc imediat (plopi, sălcii), în primăvara următoare (majoritatea speciilor) sau în primăvara celui de-al doilea an (tei, carpen, frasin).
Germinaţia poate fi epigee (cotiledoanele sunt deasupra solului) cum este cazul la fag, carpen, ulmi sau hipogee (cotiledoanele rămân în sol), ca la stejari, castanul bun, ginkgo.
Puterea germinativă – definită ca fiind capacitatea seminţelor de a încolţi în condiţii mai mult sau mai puţin propice – variază de la 80-90% (molid) la 20-40% (mesteacăn, brad, magnolia).
PLANUL DE PREZENTARE AL SPECIILOR LEMNOASE
• Caractere morfologice (iarnă şi vară) 1. Provenienţa (indigen, exotic) 2. Mărimea, dimensiuni realizate 3. Înrădăcinarea 4. Tulpina 5. Scoarţa, ritidomul 6. Lemnul 7. Coroana 8. Lujerii
9. Mugurii 10. Frunzele 11. Florile 12. Fructele, conurile, seminţele, maturaţie 13. Maturitate, putere germinativă,
periodicitate 14. Creşteri, productivitate 15. Longevitate
• Vătămători (insecte, ciuperci, factori de mediu)
• Arealul - general - în România
• Cerinţe ecologice - sol - climă - staţiuni - factori limitativi
• Variabilitate morfologică, ecologică (ecotipuri, climatipuri, edafotipuri)
• Înmulţire
• Însuşiri silviculturale, importanţă
SISTEMATICA SPECIILOR FORESTIERE Din punct de vedere al diversităţii biologice, dar şi ecologice, specia este un concept definit
plecând de la un număr de n populaţii componente, fiecare din ele acumulând un număr de n indivizi.
Privind prin prisma unor concepte de diversitate, specia este caracterizată plecând de la cel mai mare număr posibil de caractere morfologice şi de la mai multe populaţii pentru fiecare din cei n indivizi. Astfel, - delimitările morfologice se fac în raport cu alte specii, - se decelează principalele variaţii, fapt ce duce la identificarea unităţilor intraspecifice, - se exprimă un tablou al situaţiei filogenetice în raport cu alte specii, ceea ce duce la
clasificarea propriu-zisă. Aceste aspecte duc, în final, la recunoaşterea fiecărui individ plecând de la caracterele de
diagnoză, în principal florile, dar şi alte caractere specifice.
Diversitatea ecologică a speciilor forestiere
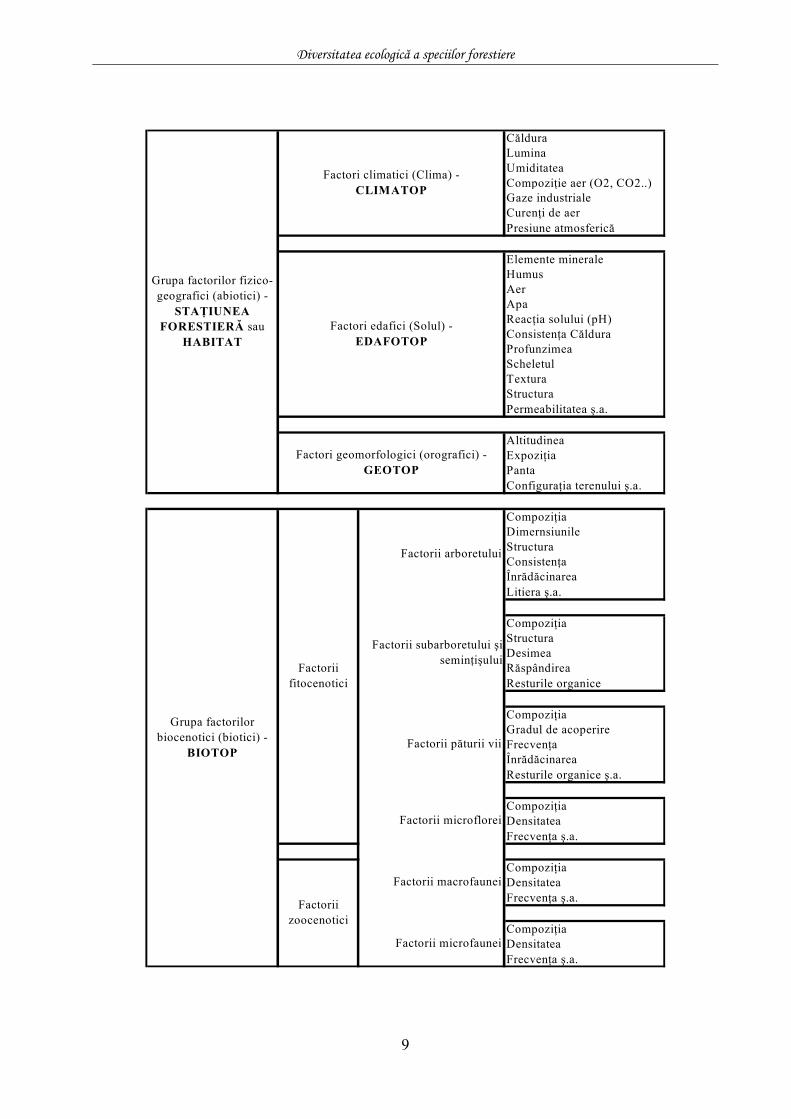
CălduraLuminaUmiditateaCompoziţie aer (O2, CO2..)Gaze industrialeCurenţi de aerPresiune atmosferică
Elemente mineraleHumusAerApaReacţia solului (pH)Consistenţa CălduraProfunzimeaScheletulTexturaStructuraPermeabilitatea ş.a.
AltitudineaExpoziţiaPantaConfiguraţia terenului ş.a.
CompoziţiaDimernsiunileStructuraConsistenţaÎnrădăcinareaLitiera ş.a.
CompoziţiaStructuraDesimeaRăspândireaResturile organice
CompoziţiaGradul de acoperireFrecvenţaÎnrădăcinareaResturile organice ş.a.
CompoziţiaDensitateaFrecvenţa ş.a.
CompoziţiaDensitateaFrecvenţa ş.a.
CompoziţiaDensitateaFrecvenţa ş.a.
Grupa factorilor biocenotici (biotici) -
BIOTOP
Factori geomorfologici (orografici) - GEOTOP
Factori edafici (Solul) - EDAFOTOP
Factori climatici (Clima) - CLIMATOP
Grupa factorilor fizico-geografici (abiotici) -
STAŢIUNEA FORESTIERĂ sau
HABITAT
Factorii subarboretului şi seminţişului
Factorii arboretului
Factorii zoocenotici
Factorii fitocenotici
Factorii microfaunei
Factorii macrofaunei
Factorii microflorei
Factorii păturii vii
9

Diversitatea ecologică a speciilor forestiere
10
ECOLOGIA SPECIILOR FORESTIERE
Din punct de vedere forestier, ecologia speciilor lemnoase se împarte în două diviziuni: - autecologia - sinecologia.
Autecologia se ocupă cu studiul caracteristicilor ecologice ale speciilor, adică adaptările lor la acţiunea factorilor mediului abiotic şi biotic. Altfel spus, autecologia este reprezentată prin studiul pentru fiecare populaţie a exigenţelor indivizilor vis-à-vis de climat, sol, lumină.
Şi asta pentru că cele n populaţii care constituie o specie trăiesc într-un mediu care se diferenţiază prin climat, sol, într-un mod mai mult sau mai puţin sensibil. Populaţiile ce trăiesc în medii diferite oferă adaptări diferenţiate prin selecţia naturală.
În acest sens, există un aşa numit „genic pool” corespondent la fiecare tip de mediu (staţiune), ceea ce induce existenţa unei corelaţii cu genetica speciei. Aici rezidă, în scop interpretativ, adaptările ce caracterizează un ecotip. Ca aplicaţie practică a acestui fapt demonstrat ştiinţific, este consilierea şi transpunerea în teren a diferitelor provenienţe adaptate la un anumit tip de mediu cu anumite caracteristici staţionale şi care posedă calităţile studiate. Bineînţeles că studiul provenienţelor, selecţia lor şi izolarea de clone constituie un câmp de cercetare destul de puţin dezvoltat şi nici pe departe epuizat. După unii autori, aceste aspecte genetice ale speciilor forestiere, dar şi a celorlalte specii întrunesc o altă latură a ecologiei şi anume genecologia.
Autecologia se bucură de tradiţie, toate tratatele de dendrologie fiind în fapt sinteze autecologice.
Componentele mediului care exercită diferite acţiuni asupra arborelui sunt considerate condiţii de viaţă, deoarece condiţionează activităţile metabolice fundamentale, diminuând sau intensificând efectele factorilor ecologici propriu-zişi. Ar fi cazul unor elemente constituente ale mediului fizico-geografic, ca de exemplu mişcările de aer, căldura din sol, aciditatea solului, textura, structura solului, conţinutul de schelet etc.
Diferenţierea elementelor fizico-geografice şi biotice după importanţa acţiunii lor în factori de mediu şi condiţii de mediu rămâne arbitrară, deoarece şi factorii ecologici condiţionează efectul altor factori. Aşa se întâmplă, de exemplu, cu apa din sol care influenţează hotărâtor accesibilitatea substanţelor minerale sau cu radiaţia luminoasă, de intensitatea căreia sunt strâns legate mişcările stomatelor şi deci, accesibilitatea CO2 etc.
Pe de altă parte, specializarea ecologică a speciilor lemnoase fiind însoţită şi de o serie de reacţii fiziologice şi morfologice de răspuns la acţiunea factorilor de mediu, adaptările ecofiziologice şi morfoecologice care au apărut sub presiunea selecţiei naturale, trebuie avute în vedere.
Explicarea tuturor trăsăturilor morfologice, ale arborilor şi arbuştilor forestieri, prin
implicarea lor pe plan ecologic cauzal, nu este însă posibilă, deoarece caracterele fizionomice sunt, în fond, reflectarea însuşirilor ereditare ale speciilor apărute în cursul evoluţiei filogenetice.
Mecanismul fundamental de perfecţionare adaptativă a speciilor constituindu-l selecţia naturală este evident că variantele optime de formă şi structură (de exemplu forma conică a coroanei molidului în climatele cu zăpezi dese şi abundente) sunt rezultatul unei evoluţii de lungă durată.
Sinecologia se ocupă de raporturile de convieţuire ale indivizilor şi populaţiilor în cadrul
comunităţii şi de reacţiile lor neadaptative faţă de mediul ecosistemului. Un mediu silvestru adăposteşte un anumit număr de specii, fiecare reprezentată printr-o populaţie de n indivizi.
Conceptul central al acestei diviziuni a ecologiei pleacă de la ideea că un ansamblu floristic dat se regăseşte de fiecare dată acelaşi când condiţiile de mediu sunt identice. Este evident faptul că aceasta presupune aplicarea în câmp interdisciplinar a mai multor domenii.
Diversitatea ecologică a speciilor forestiere
Dovada acestei afirmaţii constă în faptul că este posibil să fie caracterizate şi delimitate diverse medii sau staţiuni prin prezenţa unui anumit cortegiu floristic, fiind astfel definită fitosociologia.
La modul practic, sinecologia este definită prin faptul că staţiunile, la modul general, condiţiile de vegetaţie fiind circumscrise studiului factorilor ecologici (climat, sol, lumină, factori limitativi) permit în sinteză stabilirea de corelaţii între vegetaţia forestieră şi aceşti factori.
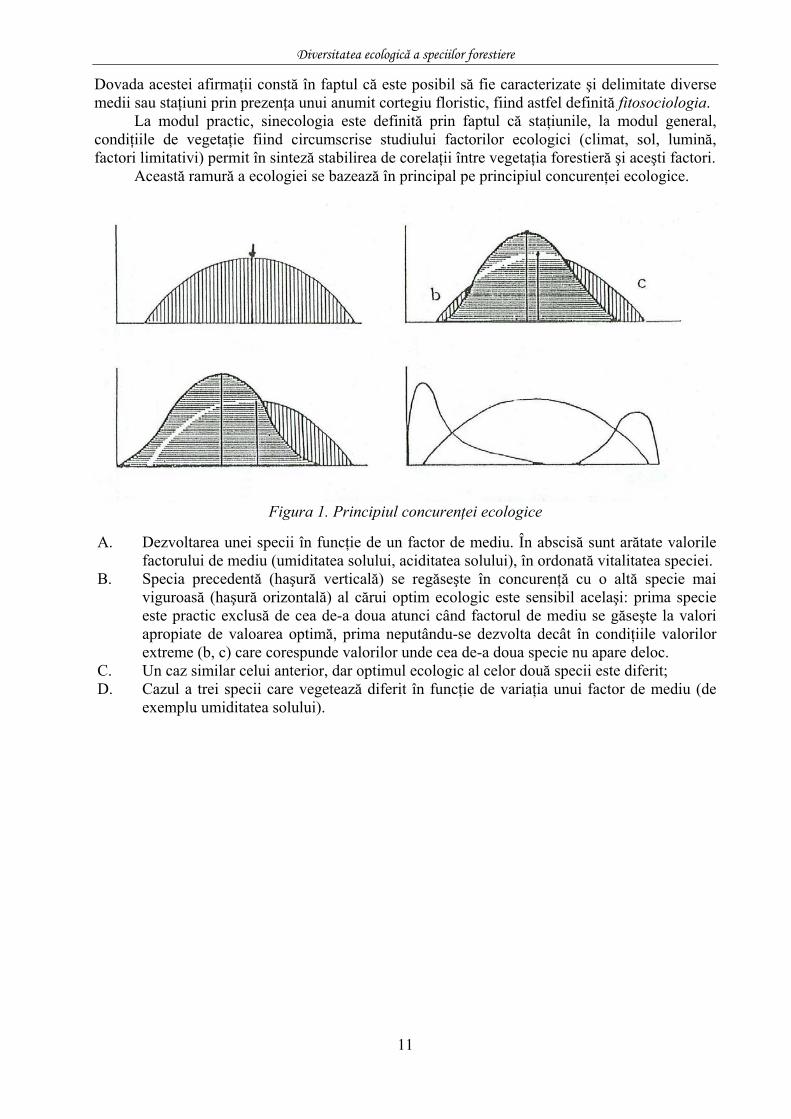
Această ramură a ecologiei se bazează în principal pe principiul concurenţei ecologice.
Figura 1. Principiul concurenţei ecologice
A. Dezvoltarea unei specii în funcţie de un factor de mediu. În abscisă sunt arătate valorile
factorului de mediu (umiditatea solului, aciditatea solului), în ordonată vitalitatea speciei. B. Specia precedentă (haşură verticală) se regăseşte în concurenţă cu o altă specie mai
viguroasă (haşură orizontală) al cărui optim ecologic este sensibil acelaşi: prima specie este practic exclusă de cea de-a doua atunci când factorul de mediu se găseşte la valori apropiate de valoarea optimă, prima neputându-se dezvolta decât în condiţiile valorilor extreme (b, c) care corespunde valorilor unde cea de-a doua specie nu apare deloc.
C. Un caz similar celui anterior, dar optimul ecologic al celor două specii este diferit; D. Cazul a trei specii care vegetează diferit în funcţie de variaţia unui factor de mediu (de
exemplu umiditatea solului).
11

Diversitatea ecologică a speciilor forestiere
CAPITOLUL 2.
RĂSPÂNDIREA SPECIILOR FORESTIERE LEMNOASE
INTRODUCERE
Pentru a justifica menţinerea şi prosperitatea unei specii sau raritatea sa, trebuie anvizajat comportamentul ecologic al taxonului, adică exigenţele sale faţă de climat şi sol şi de asemenea, reacţiile sale pentru condiţiile create prin prezenţa altor vegetale.
Dar, condiţii de mediu comparabile nu sunt suficiente pentru a determina specii identice. Exemplu: pădurile canadiene şi pădurile scandinave au condiţii de mediu apropiate,
fizionomia este aceeaşi, dar speciile sunt diferite. Ecologia nu poate, deci, explica aceste diferenţe: - pentru ca o specie să existe şi să prospere într-un anume loc, trebuie ca ea să fi putut să
parvină, trebuie deci, să facem apel la corologie (chora = loc) - pentru HAECKEL (1866) această disciplină înglobează toată documentaţia relativă la
repartiţia geografică a speciilor vegetale. Se disting: - autocorologia – repartiţia geografică a unităţilor taxonomice (specii, genuri, familii), - sincorologia – repartiţia geografică a grupărilor vegetale.
Într-un cuvânt, este ştiinţa dispersiei şi repartiţiei organismelor vii. Aceasta are două laturi: studiul static al repartiţiei geografice (areografie) a taxonilor şi elementelor floristice sau
faunistice, studiul dinamic al dispersiei (origine şi schimbări).
Fiecare speciei vegetală (sau animală) ocupă o parte din suprafaţa Globului bine delimitată la un moment dat, dar care poate varia – regresa sau se întinde – în decursul timpului, acesta fiind arealul de repartiţie geografică.
Printre cauzele foarte complexe care explică arealele actuale ale speciilor, se disting: - cele care favorizează extinderea arealului
• migraţiile supuse unui dinamism intern = variabilitate genetică şi puterea de extindere a organismului,
• migraţiile supuse unui dinamism extern = acţiunea favorizantă sau inhibitoare a mediului;
- cele care limitează arealele • cauzele actuale geografice şi ecologice, • cauze vechi (nu se pot interpreta arealele geografice fără a cunoaşte
liniile directoare ale istoriei speciilor vegetale).
AREAL. NOŢIUNI GENERALE
1. Localizarea. Arealul unui taxon Punctul de plecare elementar este localitatea – locul geografic ocupat de populaţiile unei
specii susceptibile de a fi reprezentată pe o hartă (THURMANN, 1849). Noţiunea de localitate este diferită de noţiunea de staţiune – mediu biologic normal al unei
unităţi sistematice – taxoni sau grupări vegetale (Du RIETZ, 1930). Conform specialiştilor anglo-americani, staţiune = habitat. Suprafaţa totală pe care se constată existenţa în stare spontană a unei specii sau unei grupe de
specii. Pentru a determina arealul unei specii se marchează pe o hartă toate localităţile cunoscute unde există această specie, apoi se trasează curba care înglobează aceste puncte.
12

Diversitatea ecologică a speciilor forestiere
În funcţie de scopul cercetării, scara acestor hărţi poate varia considerabil: - hărţi la scară mare şi foarte mare (1:5000 şi mai mare) întocmite pentru studiul dinamicii
anumitor populaţii animale, - hărţi la mică şi foarte mică scară întocmite pentru reprezentarea distribuţiei unei specii
date pe un teritoriu sau pe tot arealul de răspândire. Precizia acestor hărţi este foarte variabilă, în funcţie de specie şi ţări, depinzând de nivelul de cunoaştere al localităţilor şi speciilor.
În general arealul de răspândire al speciilor forestiere arborescente este mai bine cunoscut decât cel al speciilor arbustive sau ierboase ce nu sunt direct utilizate de către factorul antropic.
Metoda cea mai precisă constă în a reprezenta pe hartă fiecare localitate printr-un punct: - la scară mare pentru speciile rare şi cu extindere geografică restrânsă, - pentru o mică parte a unei areal vast. Cel mai adesea, se foloseşte o metodă de cadrilaj asupra teritoriului, marcând cu un semn
pătratele unde taxonul este reprezentat. Această metodă este folosită în Scandinavia, Marea Britanie, Olanda, Belgia şi Franţa pentru plantele vasculare în vederea comparării repartiţiei speciilor în interiorul ţării.
Trebuie să se facă distincţia între arealul real de arealul potenţial. Arealul real este suprafaţa geografică unde o specie este efectiv prezentă, adesea circumscrisă
prin conturul arealului populaţiilor din care ea este constituită. Astfel, arealul real acoperă zone de unde specia a dispărut din cauza acţiunii omului sau zone în care anumite cerinţe ecologice ale speciei nu sunt îndeplinite.
Arealul potenţial este ansamblul zonelor unde specia considerată nu există sau nu mai există în stare spontană din cauza diseminării sale, migrării din decursul timpului, dar, unde ea poate să crească, să se dezvolte şi să se reproducă în mod natural când ea va fi introdusă; ea devine astfel subspontană. Cunoaşterea arealului potenţial are o mare importanţă pentru anumite specii în cazul importării acestora sau în cazul reîmpăduririi.
Arealul speciilor diferă foarte mult de la specie la specie, atât din punct de vedere a întinderii, cât şi a formei.
2. Suprafaţa arealului Nu există multe specii vegetale care să fie ubicviste în sensul etimologic al cuvântului. Dar,
acest termen este utilizat pentru a desemna speciile care se pot dezvolta în climate foarte variate, însă într-un anumit mediu.
A. de CANDOLLE care a studiat foarte mult repartiţia speciilor vegetale, estimează că un număr foarte mic de specii de fanerogame pot fi calificate ca fiind cosmopolite – reprezentate peste tot pe suprafaţa Globului – dintre speciile forestiere Pteridium aquilinum ocupând un areal foarte vast: Europa, Asia, Africa, America de Nord, acolo unde condiţiile de mediu îi sunt favorabile (mai ales condiţiile de sol, mai mult decât cele de climat). Cu toate acestea, specia lipseşte câteodată pe teritorii întinse atunci când condiţiile ecologice par să îi fie favorabile.
Nici o specie lemnoasă nu figurează printre speciile vegetale cu areal foarte întins, mai mult nici un arbore nu posedă un areal care să cuprindă emisfera boreală şi cea australă.
Printre speciile lemnoase care au arealul cel mai mare şi continuu este ienupărul comun (Juniperus communis), adică cea mai mare parte a Eurasiei şi America de Nord, acest areal fiind calificat ca fiind circumterestru holarctic.
Există un foarte mic număr de specii lemnoase a căror areal se întinde atât în Europa, Asia, cât şi în America de Nord. Acestea sunt în general specii de talie mică, ce vegetează în regiunile reci (boreale sau montane înalte): mesteacănul pitic (Betula nana) şi specii pitice de salcie (Salix glauca, S. reticulata, S. herbacea).
Printre deja numeroşii arbori ce au o întindere în Europa şi Asia putem aminti mesteacănul pufos (Betula pubescens), plopul tremurător (Populus tremula), aninul negru (Alnus glutinosa), ulmii (Ulmus minor, U. glabra), sâmbovina (Celtis australis).
Dar principalele specii forestiere lemnoase au un areal care nu vizează decât un continent, în speţă Europa, sau nu depăşesc decât puţin limitele continentului. Exemplificăm prin prezenţa următoarelor specii: stejarul şi gorunul (Quercus robur, Q. petraea), fagul (Fagus sylvatica), carpenul (Carpinus betulus).
13

Diversitatea ecologică a speciilor forestiere
Speciile secundare, cum ar fi unele specii de stejar (Quercus suber, Q. pyrenaica) au un areal restrâns, de mică întindere.
3. Endemismele Există un mare număr de specii lemnoase care au areale foarte restrânse, aceste specii
numindu-se endemice şi ele sunt caracteristice unei anumite regiuni. Se constată că endemismul este foarte dezvoltat în anumite regiuni ale Globului, în mod
special în insule, câteva lanţuri muntoase şi în câteva puncte particulare din zona de coastă a litoralului continental, adică acolo unde sunt create zone de izolare.
Este adesea cazul speciilor ancestrale (de exemplu coniferele) care marchează o anumită regresie.
a. Endemismele insulare În Oceania, existând numeroase insule, sunt foarte multe endemisme de acest tip. Noua Zeelandă, Noua Caledonie, Tazmania, posedă fiecare din ele, specii endemice din
familiile Podocarpaceae (Podocarpus, Dacrydium, Phyllocladus) şi Araucariaceae (Araucaria, Agathis) care sunt, în fapt, gimnosperme primitive.
În Noua Zeelandă trei conifere endemice fac obiectul unor exploatări forestiere importante: Kauri (Agathis australis) care ocupă un areal restrâns la extremitatea septentrională a Insulei Nordului, Rimu (Dacrydium cupressinum) şi Kahikatea (Podocarpus dacrydivides) ceva mai larg răspândite în cele două insule care formează Noua Zeelandă, o ţară ce este un important producător de lemn.
Araucaria excelsa, plantă de apartament în climatul nostru, este endemică în insula Norfolk, fiind un foarte frumos arbore ce atinge 50-70 m înălţime (Virginia, la sud de Washington).
În insulele Canare specii endemice sunt Pinus canariensis şi Juniperus cedrus, în Cipru Cedrus brevifolia (ocupă circa 250 hectare), iar în Sicilia Abies nebrodensis îşi are arealul doar pe Mont Cervo, în regiunea Polizzo.
În cazul general al ansamblului florei, trebuie de precizat că în: - Noua Zeelandă 77% din floră este endemică; - Hawai 82% din floră este endemică; - Madagascar 66% din floră este endemică; - Noua Caledonie 80% din floră este endemică. b. Endemismele montane În Balcani există trei specii cunoscute care sunt endemice şi anume Pinus peuce, P.
leucodermis şi Picea omorika. În Spania specia de brad Abies pinsapo vegetează în masivul Sierra de Ronda, Andaluzia,
unde formează trei păduri distincte destul de îndepărtate una de alta. În America de Nord, în Munţii Stâncoşi există numeroase specii endemice, dintre care se
remarcă Sequoiadendron giganteum (California, versanţii vestici ai munţilor Sierra Nevada) între 36-39° latitudine nordică unde există o serie de 31 de populaţii ce acoperă doar 23 000 hectare ce conţin circa 90 000 arbori.
c. Endemismele de coastă Exemplul tipic al acestui tip de endemism este regiunea Monterey, pe coasta Oceanului
Pacific, la 140 km sud de San Francisco. Acolo apar trei specii endemice: - Abies bracteata care apare pe versantul vestic a lanţului muntos de coastă Santa Lucia, la
sud de Monterey şi care are un areal dispersat pe circa 70 km de-a lungul coastei oceanice;
- Pinus insignis (=P. radiata) care apare pe un promontoriu stâncos care domină acelaşi oraş, dar la vest, fiind localizat într-o staţiune litorală de 50 km2 şi încă alte două mici staţiuni de o parte şi de alta a oraşului Monterey;
- Cupressus macrocarpa ce apare pe acelaşi promontoriu, specia însumând câteva zeci de hectare şi câteva mii de exemplare.
Plecând de la aceste arborete naturale, Pinus radiata şi Cupressus macrocarpa au fost larg răspândite prin reîmpăduriri.
Speciile endemice sunt frecvent capabile de interfertilitate cu speciile vecine, ceea ce denotă desprinderea lor sistematică, în raport cu speciile cu care sunt interfertile, din aceeaşi tulpină.
14

Diversitatea ecologică a speciilor forestiere
4. Forma arealului a. Areal compact Acest tip de areal este cel mai simplu: punctele ce figurează localităţile sunt numeroase şi
apropiate. Astfel, curbele de contur sunt uşor de trasat, partea subiectivă, de interpretare a arealului fiind puternic minimizată.
Pe măsură ce arealul creşte ca întindere, pe atât creşte riscul arealului de a fi mai puţin compact. Cu toate acestea, unele din marile noastre specii indigene – stejarul (Quercus robur), gorunul (Quercus petraea), carpenul (Carpinus betulus) – sunt exemple elocvente de specii cu areal compact.
b. Areal difuz Adesea, există în zonele marginale staţiuni forestiere sau localităţi mai mult sau mai puţin
depărtate, astfel încât traseul de contur este dificil de materializat pe hartă, dar depinde totuşi de scara la care este întocmită aceasta.
c. Areal insular Dacă diferite staţiuni forestiere ale speciei studiate se grupează în mai multe insule, atunci
arealul este insular. Ca exemple, se pot cita două specii: - Jugastrul de Banat (Acer monspenssulanum) apare insular în câteva zone: în Franţa
(denumirea populară este arţar de Montpellier) apare în zona centrală şi de est a ţării, apoi îl găsim la 400 km mai la est în bazinul Rinului, apoi pe valea râului Main, aproape de Frankfurt şi apoi în România, în zona Banatului.
- Molidul (Picea abies) are o răspândire compusă dintr-un areal compact în zonele nordice ale Europei, dar şi dintr-un areal insular în zonele montane ale Europei Central şi de Est, ca şi în zona de sud a Alpilor.
d. Areal disjunct Dacă staţiunile forestiere sau zonele de areal sunt separate prin distanţe considerabile atunci
arealul este disjunct. Ca exemplu putem da zâmbrul (Pinus cembra) care are un areal foarte vast în Siberia, dar
apare şi în staţiuni îndepărtate din zonele înalte de origine vulcanică din Alpi şi Carpaţi. Un alt exemplu foarte interesant este cel al tisei (Taxus baccata). Specia apare în: - cea mai mare parte a Europei, în Asia Mică şi în Africa de Nord, - tot sud-estul Asiei, din Himalaya până în Japonia şi chiar în zonele montane din Filipine, - vestul Americii de Nord (Columbia Britanică şi California), - estul Canadei şi al Statelor Unite ale Americii. Un areal disjunct dovedeşte adesea faptul că specia este în regresie şi că insulele actuale sunt,
cu certitudine, relicvele unui areal vechi mult mai vast. În egală măsură, arealul disjunct poate traduce exigenţe ecologice particulare, aici având ca exemplu cazul speciilor montane.
5. Infrataxoni: subspecii, rase geografice, ecotipuri şi varietăţi În timp ce unele specii au un areal disjunct, frecvent se constată diferenţe mai mult sau mai
puţin mari între indivizii din diverse zone ale arealului, diferenţe care justifică separarea de subspecii, varietăţi sau rase geografice.
Trebuie de menţionat că între aceşti infrataxoni creaţi se găsesc în general mult forme intermediare.
Aşa de exemplu este specia colectivă Pinus nigra, specie care are un areal circummediteranean reprezentat în diferite părţi ale arealului său prin populaţii fenotipic asemănătoare, dar din punct de vedere ecologic şi genetic foarte diferite.
Conform diferiţilor autori ce s-au ocupat de această specie, aceste forme sunt considerate ca varietăţi, subspecii sau chiar specii. În realitate, sunt unităţi intraspecifice a unei specii colective polimorfe.
15

Diversitatea ecologică a speciilor forestiere
• Pinus nigra ssp. clusiana var. salzmanni – pin de Salzmann (Pirineii Orientali) var. clusiana – f. hispanica pin de Clusius (Spania – Cuenca, Andaluzia) var. mauretanica – pin de Mauritania (Algeria – masivul Babor)
• Pinus nigra ssp. laricio var. laricio – pin laricio de Corsica var. calabrica – pin laricio de Calabria
• Pinus nigra ssp. pallasiana var. banatica var. bulgarica var. pindica var. pontica var. caraminica var. fenzlii
• Pinus nigra ssp. nigricans var. austriaca – pin negru de Austria var.illyrica var.dalmatica var. italica var. gocensis
Pe baza caracterelor morfologice şi anatomice ale frunzelor, anumiţi autori nu recunosc decât două subspecii distincte:
- ssp. occidentalis (clusiana şi laricio de Corsica), - ssp. orientalis (nigricans, laricio de Calabria şi pallasiana).
Un alt exemplu este tisa (Taxus baccata). Există, la această specie, câteva diferenţe de ordin morfologic între exemplarele din regiuni mai îndepărtate una de alta, diferenţe de port în principal.
Anumiţi autori precizaţi mai jos constituie specii diferite: - RHEDER – 5 specii - Taxus baccata,
- T. chinensis (vestul Chinei), - T. cuspidata (Japonia, Coreea, Manciuria), - T. canadensis (America de Nord, coasta de est), - T. brevifolia (America de Nord, coasta de vest).
- DALLMORE şi JACKSON susţin aceleaşi cinci divizări, dar recunoscând că, practic, nu există diferenţe de ordin morfologic, dar estimează că separarea clară din punct de vedere arealogic este suficientă pentru a stabilire această scindare.
- PILGER consideră că există o singură specie şi nu conferă formelor geografice decât valoare de subspecie.
Cu toate acestea, dispersarea pe continente îndepărtate conduce la ideea că separarea celor cinci specii este cea mai aproape de adevăr.
6. Specii vicariante Dacă diferenţele morfologice existente între formele ce populează areale diferite sunt
evidente, atunci aceste forme devin specii diferite, dar foarte apropiate unele de altele cărora li se atribuie numele de specii vicariante.
În esenţă vicarianţa reprezintă înlocuirea unui taxon prin altul diferit, dar alăturat: - fie în staţiuni asemănătoare, din două regiuni geografice diferite = vicarianţă
corologică sau geovicarianţă; - fie staţiuni diferite ale aceleiaşi regiuni geografice = vicarianţă ecologică sau
ecovicarianţă. Aceste forme de vicarianţă sunt în mod special între flora Europei Occidentale şi Centrale şi
flora Americii de Nord, partea de nord-est.
16

Diversitatea ecologică a speciilor forestiere
Specii europene Specii americane Fagus sylvatica Fagus grandifolia
Carpinus betulus Carpinus caroliniana Populus tremula Populus tremuloides
Ulmus minor Ulmus fulva Sorbus aucuparia Sorbus americana
Alnus incana Alnus rugosa Alnus viridis Alnus crispa
Vicarianţa există de asemenea şi la speciile de arbuşti, subarbuşti şi specii ierboase.
Specii europene Specii americane
Viburnum lantana Viburnum lantanoides Viburnum opulus Viburnum americanum
Sambucus racemosa Sambucus pubens Clematis vitalba Clematis virginiana
Cornus sanguinea Cornus stolonifera Athyrium filix femina Athyrium angustum
Hepatica nobilis Hepatica americana Oxalis acetosella Oxalis montana Fragaria vesca Fragaria americana
Majanthemum bifolium Majanthemum canadense
7. Arealul genurilor Noţiunea de areal se aplică în egală măsură şi grupelor sistematice de ordin mai mare decât
specia. Astfel, un exemplu de gen cu areal cosmopolit este genul Salix. Multe din genurile speciilor lemnoase se caracterizează prin un areal circumterestru, adică se
întinde în jurul Globului, dar rămâne între limite latitudinale precise. Aşa, spre exemplu, este genul Betula, care are un areal continuu, de mare întindere, dar care se limitează în cea mai mare parte a emisferei nordice în intervalul 30-75° latitudine nordică.
Un areal holarctic presupune existenţa unui taxon în emisfera boreală, la nord de Tropicul Racului. Exemple:
- genul Quercus are un areal foarte vast, meridional cu limită sudică destul de neregulată care atinge Ecuatorul în Asia (Siam, Malaiezia) şi coboară până în Columbia, în America Centrală;
- genul Pinus are un areal continuu în cea mai mare parte a emisferei nordice, având totuşi două specii care fac excepţie, fiind localizate în emisfera sudică (Sumatra, Java, Borneo, Filipine)
Frecvent însă, genurile au un areal fărâmiţat, insular, diferite părţi ale arealului putând fi de întindere variabilă.
Genul Fagus (fagi boreali): - o specie în Europa - F. sylvatica - o specie în Asia Mică - F. orientalis - 2 specii în America - F. grandifolia, F. mexicana - 7 specii în Extremul Orient - F. japonica, F. crenata etc. Genul vicariant Nothofagus (fagi australi): - 8 specii în America de Sud (Anzii Cordilieri) - 6 specii în Noua Zeelandă - 2 specii în Tazmania şi sud-estul Australiei
17

Diversitatea ecologică a speciilor forestiere
8. Arealul familiilor Familiile care din punct de vedere numeric sunt importante au un areal de răspândire pe toată
suprafaţa Globului: Asteraceae, Poaceae, Orchidaceae etc. Familia Fabaceae grupează un mare număr de plante lemnoase ce vegetează în zonele calde.
Dintre familiile cele mai importante din punct de vedere forestier exemplificăm: - familia Betulaceae, cu un areal continuu, foarte întins în cea mai mare parte a emisferei
nordice, cu excepţia câtorva specii de anin (Alnus sp.) care sunt răspândiţi în zonele montane ale Americii Centrale şi a jumătăţii nordice din Anzii Cordilieri;
- familia Fagaceae, cu un areal foarte întins, dar disjunct, ca urmare a existenţei genului Nothofagus în emisfera sudică;
- iedera (Hedera helix) este singurul reprezentant european al unei familii numeroase şi exclusiv tropicale: familia Araliaceae.
Familiile sunt grupări sistematice mult mai vaste ca întindere decât genurile sau speciile, fiind grupe mai „solide” decât genurile.
Un foarte mic număr de familii pot fi considerate ca endemice. Nici una din ele nu cuprinde specii lemnoase importante din punct de vedere forestier. Cu titlu de exemplu, poate fi citată familia Cneoraceae, endemică, din regiunea mediteraneană, familie cu un singur gen şi o singură specie: Cneorum tricoccum (arbust ce apare pe stâncării calcaroase).
18

Diversitatea ecologică a speciilor forestiere
Capitolul 3.
DE LA ARBORE LA ARBORET
IMOBILITATEA INDIVIDULUI – MOBILITATEA POPULAŢIEI Încă din Neolitic se succed numeroase cicluri de rotaţie a vegetaţiei, cicluri care debutează
printr-o perioadă de intensificare a activităţilor agricole şi se termină printr-o perioadă de reîmpădurire naturală. Spaţiul rural prezintă zone de tranziţie care suferă în mod alternativ astfel de fenomene, în funcţie de evoluţia demografică. În afara perioadelor de abandon, arborii care au fost păstraţi asigură recucerirea activă a spaţiilor deschise şi reconstituirea progresivă a ecosistemului forestier graţie diferitelor strategii de reproducere ce conferă puterea de a migra, mai mult sau mai puţin eficient.
Puterea de a migra depinde, înainte de toate, de numărul seminţelor produse şi de maniera în care are loc diseminaţia. Anumite specii investesc cea mai mare parte a energiei în producţia unei cantităţi mari de seminţe, şi asta în fiecare an, cum este cazul la mesteacăn, salcie, anini, ulmi, plopi. Acestea au din capul locului un avantaj incontestabil asupra speciilor care produc o cantitate redusă de fructe, seminţe şi cu o periodicitate mare.
Diseminarea la distanţă mare face apel la intervenţia unui agent exterior. Vântul este cu certitudine agentul cel mai activ din Europa, cu atât mai eficace cu cât seminţele sunt mai uşoare sau că ele prezintă suprafaţă mare în raport cu volumul lor. Cele mai multe posedă adaptări morfologice în acest sens: smocuri de peri la sălcii şi plopi, aripioare la ulmi, mesteceni, anini, frasini, arţari, pini, molid, larice, bracteea inflorescenţei de la tei etc.
Păsările asigură răspândirea seminţelor de la fructelor cărnoase pe care vântul este incapabil să le transporte, prin simpla ingerare: speciile de scoruş, cireş, călin, corn, ienupăr ş.a. în plus, nu numai că facultatea germinativă este pierdută, din contră, pentru anumite specii ea este activată, cum este cazul păducelului. În pădurile tropicale, unde arborii produc în majoritatea lor fructe cărnoase, acţiunea păsărilor este determinantă.
Seminţele şi fructele uscate ale unor specii lemnoase sunt dispersate de către animale care acumulează provizii pentru iarnă (seminţe pierdute în timpul transportului sau neutilizate): ghinde, castane, nuci, alune, coconari, sâmburi ale unor drupe etc.
Pe un metru pătrat se pot găsi până la 400 samare de mesteacăn, 500-700 seminţe de pin silvestru sau molid, 400-500 bucăţi de jir sau seminţe de brad în timpul unor ani de fructificaţie abundentă. Experienţa arată că între 50 şi 100 de bucăţi de seminţe sau fructe sunt necesare şi suficiente pentru a asigura o reînsămânţare naturală mulţumitoare, restul constituie un enorm bagaj cheltuit inutil.
Aceste seminţe constituie banca de seminţe in situ care, după ani de aşteptare a unor condiţii favorabile poate deveni capabilă să recucerească terenul pierdut de către ecosistemul forestier.
MODALITĂŢI DE RECUCERIRE A SPAŢIULUI
Seminţele germinează şi se dezvoltă în mod liber în timp ce blocajul creat de om este ridicat şi presiunea este în continuă scădere, presiune exercitată cu scopul de a elimina pădurea. O selecţie se operează asupra regenerării: singurele specii heliofile în stadiul juvenil vor urma avântul din acest stadiu. Speciile pioniere posedă caracteristici specifice care le favorizează în cucerirea terenului deschis: numărul mare de seminţe diseminate adesea prin vânt şi capacitatea de a se regenera în plină lumină (mesteacăn, plopi, sălcii, anini, pini, frasin, ulm, tei, arţari). Ele transformă frecvent mediul, îmbogăţind solul şi creând condiţii propice pentru instalarea speciilor mai exigente din punct de vedere trofic şi necesită luminozitate diferenţiată în raport cu vârsta (la început umbră, apoi din ce în ce mai multa lumină). Aceste specii intră în categoria driadelor, adică a speciilor ce definesc ecosistemul forestier: gorun, stejar, fag, brad.
Reinstalarea pădurii se efectuează conform diferitelor procese. Colonizarea frontală este, de regulă, când subzistă o anume presiune din punct de vedere al păşunatului. În timpul presiunii
19

Diversitatea ecologică a speciilor forestiere
antropice, obişnuit a păşunatului intensiv, reîmpădurirea naturală se face prin intermediul speciilor arbustive (ex. ienupăr, alun, măceş, păducel, sânger, corn etc.), care în momentul slăbirii proceselor antropice aceste specii sfârşesc prin a se uni, iar la adăpostul lor vin speciile pioniere (mesteacăn, plop tremurător, salcie căprească). Câteodată, acolo unde condiţiile permit, speciile arborescente se instalează direct în mediul ierbos: pinul silvestru, mesteacăn, paltin de munte, molid, frasin.
Oricare ar fi modelul de recolonizare forestieră, acest tip de vegetaţie apare plecând de la existenţa unor suprafeţe acoperite de vegetaţie forestieră, cum ar fi la periferia pădurilor, cu plecare din liziera acestora, tufărişurilor, desişuri formare din specii pioniere deja instalate.
ACTIVITĂŢI CICLICE ÎN CADRUL ECOSISTEMULUI FORESTIER Ecosistemul forestier este în permanentă schimbare sub influenţa ritmurilor naturale zilnice
sau sezoniere sau impuse de către om în arborete. Fiinţele vii şi-au adaptat diversele activităţi pe măsura zilelor şi anotimpurilor, ceea ce are drept consecinţă limitarea fenomenelor de concurenţă între specii şi exploatarea maximă a biotopului.
1. Activităţi în funcţie de anotimp În regiunea temperată, iarna este cea a repausului obligatoriu pentru vegetaţie din cauza
temperaturilor joase şi a zilelor scurte. Plantele au de ales între numeroase strategii pentru a rezista acestor rigori. Coniferele (mai puţin laricele) îşi păstrează acele şi se mulţumesc de a-şi limita nivelul de activitate. Majoritatea foioaselor îşi pierd complet frunzele toamna; mugurii sunt puşi la adăpost înaintea anotimpului mohorât în învelişurile lor. Stejarul, gorunul şi fagul, în etapa tinereţii îşi păstrează frunzele moarte până primăvara. Unii arbuşti (iarba neagră, afin) pot fi acoperiţi de un strat de zăpadă protector care îi apără de gerurile mari. Câteva graminee rămân verzi, ascunzându-şi înfrigurate mugurii la baza solului. Dar numeroase specii au devenit invizibile; ele îşi ascund mai mult sau mai puţin în sol organele perene (tuberculi, bulbi, rizomi) sau la suprafaţa humusului (seminţe ale plantelor anuale).
La sfârşitul iernii şi înaintea înfrunzirii arborilor ale căror frunze împiedică să aibă un acces suficient la lumină, apare primul val de flori cu ghiocei mai întâi, apoi viorelele, stânjeneii galbeni, brebeneii. Aceste specii de primăvară îşi formează vara primordiile florale; pentru a se alungi, ele trebuie să suporte succesiv o perioadă de frig şi o încălzire progresivă; astfel, ele pot înflori foarte precoce.
Lemnoasele cum sunt sălcile, plopii tremurători, alunul se deschid, cornul se acoperă de flori galbene şi pădurea se înmiresmează. La sfârşit de martie, înflorirea se succede într-un ritm accelerat.
Între sfârşitul lui aprilie şi începutul lui mai, seva se îndreaptă spre mugurii care pocnesc şi îşi arată frunzele încă îngheţate şi florile: în sfârşit, a venit primăvara.
Din iunie până în august înfloresc succesiv gramineele. Plantele de primăvară au încheiat ciclul anual, făcând să dispară organele lor aeriene.
Toamna soseşte marcată de îmbrăţişarea frunzişurilor (purpurii ale arţarilor şi ale sorbilor, ocru şi stacojiu ale fagilor) maturarea şi dispersarea fructelor şi dezvoltarea explozivă a ciupercilor.
Frunzele cad. Durata nopţii joacă un rol important în această cădere. Acest proces este precedat de migrarea sărurilor minerale şi a substanţelor carbonice sau azotate spre ramurile lemnoase; produsele vor apărea în primăvară. Căderea frunzelor protejează copacul de riscurile secetei de iarnă(apa nu mai este disponibilă în solul îngheţat).
2. Exigenţele şi toleranţele speciilor faţă de factorii de mediu Arborele îşi îndeplineşte tot ciclul într-o staţiune caracterizată de climă şi sol. Dezvoltarea lui
se efectuează în contact cu alţi indivizi, cu alte specii cu care el intră în competiţie pentru apă, hrană, lumină şi spaţiu vital.
20

Diversitatea ecologică a speciilor forestiere
Cunoaşterea comportamentului fiecărei specii (nevoi, exigenţe, toleranţe), faţă de aceşti factori este indispensabilă silvicultorului care vrea să instaleze arborete stabile, în echilibru cu mediul.
Temperamentul speciilor (umbra şi lumina) Fiecare fază a dezvoltării necesită un minim de lumină. În stadiul adult, esenţele au aceeaşi
nevoie de lumină. În schimb, la stadiile tinere germinarea şi dezvoltarea seminţişului, temperamentele diferă. Anumite specii au nevoie atunci de multă lumină (specii de lumină sau de semi-lumină, numite heliofile ca mesteacănul, plopul tremurător, laricele, majoritatea pinilor, stejarul pedunculat, frasinul, stejarul pubescent, ulmii, aninii). Altele tolerează un luminiş aerisit sau îl caută (specii de semi-umbră sau de umbră numite sciafile: castanul, carpenul, fagul, bradul, molidul, tisa); situaţia de adăpost pe care ele o preferă le protejează şi de îngheţuri târzii şi de secetă.
Distincţia se estompează cu vârsta; dar indivizii din speciile de lumină, dominate de vecinii lor, dispar foarte repede din arborete atunci când esenţele de umbră subzistă mai mult timp.
Exigenţele termice (căldura şi frigul) Temperatura joacă un rol important în repartizarea spaţială a speciilor; ea acţionează prin
modalităţi (anuale, lunare, ale perioadei de vegetaţie) şi mai ales prin extreme care exercită un efect limitator, chiar eliminator. Pagubele cauzate de iernile excepţionale, destul de rare în pădurile compuse din esenţe spontane pot fi catastrofice în reîmpăduririle compuse din specii exotice sau cu specii de provenienţă indigenă prost adaptate climatului local.
Frigul poate provoca moartea totală sau parţială a arborilor sau să inducă pagube de ordin mecanic sau fiziologic. Speciile cele mai sensibile sunt bineînţeles speciile meridionale, iar printre speciile cele mai rezistente enumerăm zâmbrul (Pinus cembra), jneapănul (Pinus mugo), pinul silvestru (Pinus sylvestris), laricele (Larix decidua).
Frigurile puternice provoacă adesea pagube mecanice precum crăparea tulpinilor stejarilor (gelivuri). Sub efectul contrastelor termice, crăpăturile mai mult sau mai puţin adânci se deschid adesea la baza trunchiului, atacând coaja rămasă.
Îngheţurile de primăvară sunt nocive unor specii când intervin în momentul dezvoltării mugurilor (distrugând frunzele tinere, florile şi lujerii tineri), ele perturbă astfel creşterea (despicarea frasinului), viitoarea reproducere, chiar distrug sămânţa (fag).
Exigenţele edafice (calcar sau siliciu, sol neutru sau acid, troficitate) Anumite esenţe nu suportă prezenţa calcarului în solul fin şi de aceea se întâlnesc pe suprafeţe
cu siliciu; le numim calcifuge (pinul negru, castanul, molidul). Din contră, alte specii populează solurile cu calcar unde găsesc căldură mai mare şi condiţii mai uscate (stejarul pubescent, jugastrul de Banat) Multe sunt indiferente, dar calcarul din sol le perturbă adesea creşterea (pinul silvestru).
În general, agricultura a abandonat solurile cele mai sărace în favoarea pădurii. Numeroase specii se acomodează aici datorită cumpătării lor (mesteacăn, pin silvestru). În schimb, altele necesită o alimentaţie minerală importantă; ele nu se regăsesc decât pe solurile bogate; este cazul frasinului, cireşului sălbatic, al ulmilor.
Exigenţele hidrice (soluri uscate sau umede) Adusă de precipitaţii, apa este conservată de sol mai mult sau mai puţin bine (în funcţie de
adâncimea şi elementele sale) şi se menţine inegal, în funcţie de condiţiile de păstrare. Arborele este sensibil la rezultanta acestor diferiţi factori, adică la bilanţul hidric. Speciile xerofite tolerează o alimentaţie slabă în apă (stejarul brumăriu, stejarul pubescent); fagul, bradul, frasinul, stejarul preferă mediile cu bilanţ hidric favorabil.
Excesul de apă care determină o pânză mai mult sau mai puţin temporară elimină numeroase specii; doar aninii şi sălciile suportă solurile îmbibate mereu cu apă.
Problema apei arată că arborele reacţionează în realitate la un ansamblu de factori (legaţi de climat şi de sol). Un factor care ar trebui eliminat se găseşte adesea compensat de un alt factor favorabil. De pildă, solurile foarte filtrante ale unui versant sunt căutate de către fag, de paltinul de munte, de brad, când acesta se găseşte în situaţia unei umidităţi a aerului foarte ridicată şi a unei evapotranspiraţii limitate.
21

Diversitatea ecologică a speciilor forestiere
3. Concurenţa şi caracteristicile intrinseci ale speciei Exigenţele şi toleranţele variază, printre altele, considerabil în funcţie de intensitatea
concurenţei exercitate de alte specii. Astfel, frasinul şi cireşul sălbatic se instalează în lizierele pădurilor de fag şi brad. Dispar foarte repede de pe solurile sărace de îndată ce bradul ajunge la nivelul lor. Stejarul pubescent rezistă pe soluri calcaroase superficiale, dacă alte specii nu sunt atât de performante şi dinamice.
Comportamentul unei specii variază în funcţie de provenienţă. Plantaţiile de larice din Alpi nu rezistă în etajul dealurilor, dar populaţiile din Polonia generează arbori foarte bine adaptate acestor condiţii.
De multă vreme, în funcţie de cerinţele şi toleranţele lor, speciile s-au repartizat în diverse regiuni naturale în combinaţii deosebite şi originale care depind de condiţiile oferite.
Legat de fenomenele de competiţie, se poate afirma cu certitudine că încă de la naştere şi pe durata vieţii, arborele forestier este supus unei concurenţe implacabile a vecinilor săi, vegetale şi animale, specii şi talii diferite, la fel ca în sol pentru a căuta apă şi elemente nutritive precum şi în aer pentru lumină. Ca în orice bătălie, există învingători şi învinşi, unii ies neatinşi, alţii „mutilaţi” sau eliminaţi. Munca silvicultorului este de a urma cu atenţie diferitele modalităţi de competiţie între indivizi şi între specii pentru a introduce evoluţia care să corespundă obiectivelor sale.
Acest lucru începe de la germinaţie: plantulele îşi înfig prima rădăcină fragilă într-un sol deja plin de rădăcinile arborilor adulţi si de cele ale plantelor ierboase; lupta se dă imediat pentru a obţine apă şi hrană. Apare reducerea rapidă a mlădiţelor care au nevoie de o nutriţie minerală ridicată (frasinul sau arţarul). Această competiţie e mai acerbă întrucât puieţii dezvoltă o înrădăcinare superficială.
Dar acest lucru devine aleatoriu, chiar imposibil, când solul e acoperit cu plante ierboase sociale cum sunt Calamagrostis sp., Brachypodium sp., Festuca sp., Molinia sp., Poa sp., Elymus sp.. Pentru a evita un asemenea blocaj care apare după expunerea la lumină, silvicultorul, înainte de a exploata arboretul, poate să aştepte ca o regenerare abundentă să se fi dezvoltat sub strat. Dacă nu, nu îi va rămâne decât să planteze şi, în acest caz, va trebui să reducă concurenţa covorului verde erbaceu care împiedică creşterea plantelor.
În aer, lupta e la fel de crâncenă pentru lumină şi spaţiu. Puieţii se alungesc datorită activităţii mugurelui terminal; cele mai mari, cele mai robuste, cele mai puţin pretenţioase la lumină vor domina şi vor sufoca pe celelalte.
La marginea pădurii dense şi întunecoase, puieţii nu au nici o şansă de a rezista. Dacă adăpostul se luminează încet, regenerările speciilor de umbră vor apărea şi vor supravieţui; puieţii speciilor de lumină mor mai repede când solul e sărac sau când concurenţa subterană este ridicată. Laricele nu tolerează nici o limitare de la lumină de-a lungul existenţei sale. Într-o pădure densă întunecoasă, sub coroana exemplarelor mature, puieţii de stejar pedunculat persistă 2-3 ani, apoi dispar. Singurele specii capabile să suporte o umbră de durată sunt bradul şi tisa care pot atinge uşor mai mult de 50 ani la umbra arboretului matern cu o creştere practic nulă şi care sunt capabili apoi de a răspunde unei luminări reluându-şi dezvoltarea.
Pentru a reuşi, operaţiile de regenerare naturală, silvicultorul operează dozări subtile de lumină prin tăieri adaptate speciilor prezente.
În arborete, vecinătatea privează ramurile joase de lumina necesară frunzelor lor; ele putrezesc şi cad mai mult sau mai puţin rapid. Acest elagaj natural este complet la foioase. În cazul coniferelor, descompunerea ramurilor moarte este mai lentă; pot rămâne cioturi şi lasă noduri depreciind scoarţa (silvicultorul poate remedia aceasta prin elagajul artificial). Curăţirea naturală încetează spre 80 de ani: arborele îşi arată atunci o formă forestieră fără ramuri pe jumătatea înălţimii sale prin opoziţie arborelui izolat. La indivizii adulţi, lupta este acerbă la nivelul coronamentului: în funcţie de densitatea arboretului, coroanele acestora devin largi sau zvelte, dezvoltaţi uniform în toate direcţiile sau asimetrici. Creşterea în înălţime şi mai ales în diametru îi influenţează; densitatea inelului anual va fi cu atât mai mare cu cât vârful va fi mai dezvoltat.
22

Diversitatea ecologică a speciilor forestiere
23
Concurenţa joacă şi ea un rol în fructificaţie: producţia de seminţe este cu atât mai mare cu cât coronamentul este mai impunător şi puternic expus la soare.
Forma definitivă a arborelui (cu diametru, înălţime, număr şi amploare a ramificaţiilor) rezultă din aceste fenomene de concurenţă şi din jocul luminii cu umbra. La aceeaşi vârstă, arborele izolat atinge un diametru mai important decât al colegului său pierdut în arboret. Dar prezintă şi numeroase defecte (formă conică, prezenţa nodurilor). De aceea, silvicultorul alege înălţarea arborilor în masiv; obţine trunchiuri lungi şi cilindrice, ale căror inele anuale mai strâmte oferă un arboret care este adesea mai bună din punct de vedere calitativ şi tehnic.
Reglând densitatea arborilor, silvicultorul comandă deci creşterea în diametru, calitatea lemnului şi utilizările lui ulterioare.
Conducerea unui arboret consistă în a grăbi efectele concurenţei selecţionând exemplarele valoroase, eliminându-le pe celelalte şi apoi aducând progresiv populaţii de arbori cu densitate căutată.

Diversitatea ecologică a speciilor forestiere
CAPITOLUL 4.
DIVERSITATEA ECOLOGICĂ A SPECIILOR DE RĂŞINOASE
CARACTERE GENERALE ALE RĂŞINOASELOR Dintre toate plantele vasculare cu seminţe, gimnospermele constituie un grupa caracterizat
în esenţă prin structura particulară a aparatului reproducător femel: ovulele sunt grupate pe un solz fertil, nu sunt închise într-o cameră carpelară şi sunt nude.
Embrionul, apărut din dezvoltarea ovulului fecundat, marchează o fază de stopare din evoluţia sa în interiorul seminţei. Toate coniferele sunt specii lemnoase, frecvent arbori de talie mare şi mai rar arbuşti sau subarbuşti.
Tulpina este adesea dreaptă, datorită unui tip de ramificaţie de tip monopodial: ramificaţiile secundare rămân laterale în timp de axul principal se alungeşte nedefinit. Ramificaţia poate lua aspectul unei false verticilări datorită localizării mugurilor axilari (exemplu Pinus sp.) ramurile dezvoltă creşteri mai mult sau mai puţin active, astfel că se pot distinge mai multe tipuri de ramuri: lungi, scurt şi pitice. De exemplu la speciile de Abies, Picea, Pseudotsuga nu există decât ramuri lungi, la Cedrus şi Larix există atât ramuri lungi (macroblaste), cât şi scurte (microblaste), iar la Pinus sp. există ramuri lungi şi ramuri pitice. Frunzele sunt adesea aciculare sau solziforme, cu un limb îngustat, rar cu un limb dezvoltat (Araucariaceae, Podocarpaceae).
Trebuie adăugat că există un anumit polimorfism al frunzelor conform cu vârsta sau poziţia pe ramuri.
Coniferele conţin în ţesuturile lor celule secretoare izolate sau grupate, în pungi şi canale, producând oleorezine, de unde termenul de răşinoase adesea folosit.
Organele reproducătoare sunt inflorescenţe unisexuate, dar speciile sunt fie monoice (flori mascule şi flori femele pe acelaşi individ), fie dioice (flori mascule şi flori femele pe indivizi diferiţi).
După fecundaţie solzii carpelari devin fie lemnoşi şi constituie un con tipic, fie cărnoşi şi constituie o după falsă.
Speciile existente de conifere sunt repartizate în şapte familii principale a căror caractere principale sunt:
1. Pinaceae – aparat femel în conuleţ, lemnos la maturitate, solzii conului poartă câte 2 ovule,bractee independentă de solz, frunze aciculare;
2. Taxodiaceae – aparat femel în conuleţ, lemnos la maturitate, solzii conului poarte 1-9 ovule la maturitate, bractee sudată parţial de solz, frunze aciculare sau solziforme;
3. Cupressaceae – ramificaţii opuse sau verticilate, conuri cu solzi opuşi sau verticilaţi lemnoşi sau cărnoşi, bractei şi solzi sudaţi în întregime, frunze aciculare sau solziforme;
4. Taxaceae – aparat femel cu un singur ovul, aparent terminal, galbulus cărnos, sămânţă învelită de un aril cărnos, traheide spiralate, frunze aciculare;
5. Cephalotaxaceae – aparat femel foarte contractat, pe un peduncul, „fruct” cărnos, cu tegumentul extern al seminţei cărnos, frunze aciculare;
6. Araucariaceae – aparat femel în con lemnos dezarticulant, bractei şi solzi sudaţi, seminţe sudate de solzi, frunze cu limbul lat;
7. Podocarpaceae – aparat femel cu ovul înconjurat de piese ce devin cărnoase şi constituie epimatium, frunze cu limb larg sau solziforme.
24

Diversitatea ecologică a speciilor forestiere
REPARTIŢIE GEOGRAFICĂ Coniferele ocupă în peisajul vegetal un rol primordial. Ele constituie elementele esenţiale a
anumitor tipuri de păduri, în special în emisfera nordică, specii din familia Pinaceae (Picea, Abies, Pinus, Larix) formează cele mai mari rezerve forestiere din regiunile septentrionale ale Eurasiei şi Americii de Nord, în etajele montane a regiunilor temperate ca şi în regiunile temperate cu secetă estivală accentuată.
În emisfera sudică răşinoasele joacă un rol mai puţin important, speciile fiind reprezentate adesea prin areale disjuncte şi care aparţin unor genuri proprii acestei emisfere.
Sintetic, zonele unde apar coniferele sunt:
Macroregiuni climatice, zone de răspândire şi regiuni floristice I. Emisfera nordică 1. Europa şi Asia (Eurasia) 1.1. Regiunile septentrionale
1.2. Regiunile temperate reci 1.3. Regiunile temperate calde
2. America de Nord II. Regiunile intertropicale III. Emisfera sudică
I. EMISFERA NORDICĂ I.1. Eurasia I.1.1. Regiunile septentrionale Pădurea de conifere constituie o bandă continuă din nordul Europei (Scandinavia) până pe
litoralul Pacificului de Nord. În Europa speciile sunt puţin numeroase: Pinus sylvestris, Picea abies, Picea obovata, Larix sibirica. În Siberia se adaugă Abies sibirica şi Pinus sibirica. Dincolo de Enisei, Larix sibirica este înlocuit de Larix gmelini.
I.1.2. Regiunile temperate reci Coniferele joacă un rol mai puţin important la altitudini joase, dar constituie esenţialul
pădurilor montane. În Europa occidentală şi centrală principalele specii sunt Pinus sylvestris, Abies alba, Picea
abies şi Larix decidua, iar în etajul subalpin Pinus cembra şi diferite forme geografice de Pinus montana.
În Caucaz se găsesc Abies nordmanniana şi Picea orientalis, în munţii Tianshan Picea schrenkiana.
În etajele montane ale masivului Himalaya, coniferele au un rol preponderent: diferite genuri ale familiei Pinaceae sunt reprezentate prin specii endemice, în special genul Pinus prin speciile Pinus griffithii, genul Abies prin speciile Abies pindrow şi A. spectabilis, genul Picea prin specia Picea morinda, genul Larix prin Larix griffithii, genul Tsuga prin Tsuga dumosa şi genul Cedrus prin Cedrus deodara.
În masivele montane din vestul Chinei, există păduri de conifere de altitudine, cu numeroase specii din genurile Abies şi Picea.
În Himalaya, ca şi în partea de sud-vest a Chinei, pădurile de conifere se succed în altitudine pădurilor de foioase cu afinităţi tropicale.
I.1.3. Regiuni temperate calde Anumite genuri importante din regiunile boreale, cum ar fi genurile Picea şi Larix nu mai
sunt reprezentate. Dar, noi genuri apar, aşa cum este cazul genului Cupressus. În regiunile circummediteraneene, anumite genuri sunt tipic montane cu specii colective, până la subspecii şi rase geografice. Este cazul peciile de brad mediteranean cu areal disjunct în Africa de Nord
25

Diversitatea ecologică a speciilor forestiere
(Abies numidica, A. marocana), în Spania Abies pinsapo, în Balcani Abies cephalonica, în Asia Mică Abies nordmanniana, A. bornmulleriana, A. cilicica. În egală măsură, este cazul pinilor din grupa speciei Pinus nigra şi a speciilor de cedru (Cedrus libani în Asia Mică, Cedrus atlantica în Africa de Nord).
Pe de altă parte, anumite specii caracterizează etajele montane aride, cum este Juniperus thurifera şi J. phoenicea. La altitudini ceva mai joase, anumite specii de pin sunt elemente esenţiale a pădurilor adaptate la climatul estival secetos: este vorba despre Pinus pinaster, P. halepensis, P. brutia, P. eldarica şi P. pinea.
Chiparosul (Cupressus sempervirens) este un element mediteranean caracteristic, dar el a fost răspândit de către om pe scară largă, astfel încât populaţiile spontane nu sunt cunoscute decât pentru anumite forme geografice. În fine, o specie din Africa de Nord, Tetraclinis articulata, este interesantă, deoarece ea aparţine unei subfamilii din familia Cupressaceae, subfamilia Callitrodeae, din care toate speciile se găsesc în emisfera sudică.
În regiunile Asiei de Sud-Est, cu condiţii climatice total diferite de cele mediteraneene, genul Pinus este bine reprezentat prin numeroase specii.
Japonia prezintă un mare interes fitogeografic, deoarece există în egală măsură specii ce se regăsesc şi în Asia continentală (Coreea, Manciuria, China), cum ar fi Pinus densiflora, Biota orientalis, dar multe specii sunt endemice: Picea polita, Abies firma, A. veitchii, A. homolepis, Larix leptolepis, Tsuga sieboldii, Thujopsis dolabrata.
I.2. America de Nord În regiunea boreală, pădurea de răşinoase ocupă o bandă neîntreruptă care merge de la
litoralul atlantic şi până aproape de litoralul pacific. Principalele specii sunt de molid (Picea mariana, P. glauca), un brad (Abies balsamea) şi un larice (Larix laricina).
Mai la sud, continentul nord-american trebuie divizat în două regiuni florale, nici o specie de răşinoase nefiind comună în cele două regiuni.
Regiunea vestică, a Pacificului, este domeniul unde coniferele ajung la cea mai puternică dezvoltare spaţială. În regiunea septentrională a litoralului pacific speciile preponderente sunt molidul de Sitka (Picea sitchensis), tuia uriaşă (Thuja plicata) şi o specie de tsuga (Tsuga heterophylla). Mai la sud, intrăm în domeniul duglasului (Pseudotsuga menziesii), a anumitor specii de brad (Abies procera, A. lowiana) şi în fine, specii din familia Taxodiaceae, cum ar fi Sequoia sempervirens şi Sequoiadendron giganteum. În altitudine, pădurile montane sunt constituite din Abies lasiocarpa, Larix occidentalis, Picea engelmanni, Abies concolor. În regiunea meridională, în lanţul muntos de coastă a Munţilor Stâncoşi, climatul mai cald, favorizează mai bine speciile de pin (Pinus ponderosa, P. jeffreyi). În California, numeroase specii sunt endemice (Pinus radiata, Abies bracteata, Cupressus macrocarpa, Calocedrus decurrens).
În regiunea floristică de Est, răşinoasele în număr mult mai mic şi nu joacă decât un rol modest în regiunea temperată. Aici apar pinul strob (Pinus strobus), tsuga (Tsuga canadensis), molidul roşu (Picea rubens) şi o singură specie de brad (Abies fraseri).
Din contră, în sud-estul Statelor Unite ale Americii, de-a lungul litoralului atlantic şi a Golfului Mexic, arborete de pin acoperă teritorii vaste (Pinus taeda, P. palustris, P. echinata, P. elliotii).
II. REGIUNILE INTERTROPICALE
În aceste zone, coniferele joacă un rol secundar în peisajul vegetal, dar numărul speciilor întâlnite aici este destul de mare.
Dintre genurile familiei Pinaceae, numai genurile Abies şi Picea nu sunt reprezentate, şi acestea dacă nu se află printre speciile de brad aflate în etajul montan al Mexicului şi Guatemalei (Abies religiosa, A. guatemalensis).
26

Diversitatea ecologică a speciilor forestiere
Din contră, genul Pinus este foarte numeros în această regiune, pe de o parte în zona tropicală americană şi pe de altă parte în zona tropicală a Asiei de Sud-Est. Astfel, - Pinus caribaea constituie arborete întinse în Mexic, Honduras, Nicaragua şi Guatemala, - Pinus merkusii există în Sumatra, Malaiezia, Indochina şi Filipine, - Pinus khasya are un areal întins în Birmania, Tailanda, Indochina şi Filipine.
Genurile Tsuga, Pseudotsuga şi Keteleeria nu sunt reprezentate decât prin specii endemice cu areal restrâns la limita zonei tropicale (Taiwan).
Din familia Taxodiaceae, de asemenea nu există decât puţine specii tropicale, aici putând fi date ca exemplu speciile Cunninghamia lanceolata şi Taiwania cryptomerioides.
Familia Araucariaceae, cu genurile Araucaria şi Agathis şi familia Podocarpaceae cu genurile Podocarpus şi Dacrydium au cel mai mare număr de specii care vegetează în această regiune.
Trebuie de precizat că în zona intertropicală, Asia şi America sunt cele mai bogate în specii de conifere, ca urmare a existenţei punţilor ancestrale a Cordilierilor pe de o parte şi a indopacificului pe de altă parte, care au permis migraţia speciilor în faza fluctuaţiilor climatice. Din contră, Africa tropicală este foarte săracă în specii de conifere, datorită separării totale de zona temperată printr-o zonă deşertică neîntreruptă.
III. EMISFERA SUDICĂ
În emisfera sudică familia Pinaceae este în totalitate absentă, familia Taxodiaceae este reprezentată prin două specii endemice în Tasmania, iar familia Cupressaceae este reprezentată prin subfamilia Callitroideae.
Din contră, familia Araucariaceae este în totalitate apartenentă emisferei sudice. În Africa australă, coniferele nu sunt reprezentate decât prin câteva specii de
Podocarpaceae (genul Podocarpus) şi de Cupressaceae (genul Widdringtonia). În Australia, coniferele sunt de asemenea puţin numeroase. În regiunea pădurilor dese de
pe coasta de Est unde este cuprinsă şi zona tropicală apar specii de Araucariaceae (Agathis robusta, Araucaria bidwilli, A. cunninghamii). Speciile de Callitris se regăsesc atât în regiunile pădurilor ombrofile, cât şi în regiunile aride.
În Tasmania, speciile de conifere aparţinând genurilor Dacrydium, Phyllocladus din familia Podocarpaceae şi Athrotaxis din familia Taxodiaceae sunt endemice.
În Noua Zeelandă există câteva specii care constituie importante pădure de conifere: Agathis australis, A. robusta, Podocarpus dacrydioides, P. totara, Dacrydium cupressinum.
În America de Sud, speciile de conifere sunt puţin numeroase, putând cita pe: - Araucaria angustifolia sau pinul de Parana care constituie importante păduri în sudul
Braziliei şi nordul Argentinei, - În zona montană din Chile apar Araucaria araucana, trei specii din familia
Cupressaceae, subfamilia Callitroideae (Fitzroya patagonica, Austrocedrus chilensis, Pilgerodendron uviferum) şi două specii din familia Podocarpaceae (Saxegothaea conspicua, Podocarpus andinus).
FAMILIA PINACEAE Lindl.
Această familie cuprinde arbori şi arbuşti preponderent răspândiţi în emisfera nordică, conţinând speciile următoarelor genuri: Abies, Keteleeria, Cathaya, Picea, Larix, Pseudolarix, Pinus, Tsuga, Pseudotsuga, Cedrus.
I. Genul Abies Mill. Cuprinde peste 40 specii răspândite preponderent în zonele muntoase ale emisferei nordice.
Sunt arbori de talie mare cu înrădăcinare pivotantă. Coroană este piramidală deasă, umbroasă, scoarţa mult timp netedă cu pungi de răşină. Lujerii sunt netezi, iar mugurii dispuşi terminal
27

Diversitatea ecologică a speciilor forestiere
întotdeauna câte trei. Prezintă frunze aciculare lăţite, pe dos cu 2 dungi albicioase de stomate, persistente, se schimbă la 6-15 ani.
Conurile sunt erecte, cilindrice, cu bractei obişnuit vizibile şi răsfrânte; solzul carpelar cade la maturitate odată cu seminţele, iar pe lujer rămâne axul erect. Seminţe relativ mari, triunghiulare, cu pungi de răşină pe tegument şi prinse strâns de aripioară.
Genul Abies este reprezentat în toată emisfera nordică, în etajul montan a regiunilor meridionale şi la joasă altitudine şi regiunile septentrionale.
Speciile genului Abies pot fi încadrate ca areal în trei mari grupe de zone geografice: - Europa şi regiunea circummediteraneană, - Asia, - America de Nord.
1. Europa şi regiunea circummediteraneană Genul Abies este reprezentat: - în munţii Europei Centrale prin Abies alba, - în Grecia prin A. cephalonica, - în Asia Mică prin A. nordmanniana, A. bornmulleriana şi A. cilicica, - în sudul Spaniei prin A. pinsapo, - în Africa de Nord prin A. numidica etc.
Afinitatea dintre diferite specii de Abies este pusă în evidenţă prin hibridarea naturală care este observată prin asocieri de specii ale genului în diferite arboretumuri, colecţii dendrologice etc. De exemplu, Abies×vilmorinii = A. pinsapo × A. cephalonica, A.×insignis = A. pinsapo × A. nordmanniana ş.a.
a. Abies alba – răspândire şi diversitate ecologică Areal general al bradului este neregulat, putând fi continuu (deseori arborete pure în munţii
Vosgi, Pădurea Neagră) sau insular (munţii Pirinei). Arealul este exclusiv european: munţii Jura, Vosgi, Masivul Central Francez, Pădurea
Neagră, Alpi, Apenini, Balcani, Carpaţi, Pirinei; în Polonia este arbore de câmpie. În nord, dincolo de 55° latitudine nordicăşi spre est, în afara arealului carpatic bradul lipseşte.
Ca rase geografice sunt de precizat: Abies alba Podolica (în nordul arealului) A. a. Carpatica (în Carpaţii Româneşti), A. a. Bihariense (Munţii Apuseni), A. a. Illyrica (Alpii Dinarici), A. a. Moesiaca (Balcani), A. a. Apenninica (Munţii Apenini).
În România, bradul este al doilea conifer ca pondere, după molid, ocupând 5% din suprafaţa păduroasă a ţării. El este bine reprezentat în Carpaţii Orientali (clina estică) în intervalul altitudinal 400-1200 m, regăsindu-se în amestecuri cu fagul şi molidul sau în arborete pure.
În Carpaţii Meridionali formează păduri de amestec, în special la vest de Olt; altitudinea maximă în Carpaţii Sudici este de 1700-1750 m. În Apuseni, apare frecvent în Munţii Gilăului şi Munţii Bihorului. Apare frecvent în Munţii Banatului, pe valea Nerei coborând până la 192 m.
Diversitatea ecologică a speciei Abies alba trebuie analizată la nivel regional, pentru fiecare rasă geografică fiind necesare studii ecologice aprofundate. Astfel, pentru bradul românesc se pot evidenţia următoarele ecotipuri autohtone: - bradul de mică altitudine (Subcarpaţii Moldovei, Depresiunea Tg. Secuiesc), adaptat la
climate reci şi precipitaţii puţine, - bradul relativ termofil (Munţii Banatului) adaptat la soluri argiloase formate pe substrat
calcaros, - bradul de soluri nisipoase (gresia de Tarcău), - bradul de pe soluri cu exces de umiditate (amfigleice din Bucovina).
28

Diversitatea ecologică a speciilor forestiere
b. Abies nordmanianna – răspândire şi diversitate ecologică Specie răspândită în partea occidentală a Munţilor Caucaz, la altitudini de peste 400 m în
Georgia şi 800 m în estul Transcaucaziei, putând ajunge chiar şi la 2000 m. El apare şi în nord-estul Turciei, în lanţul muntos pontic. De altfel, în nord-vestul Turciei
apar două specii vecine, dar care au un areal distinct: - Abies bornmulleriana, care constituie arborete pure sau amestecate în Vestul lanţului
muntos pontic, între 1100-1800 m altitudine, - Abies equi trojani, care are un areal mult mai restrâns în masivele Mont Gargari şi
Mont Ida în nord-vestul Turciei.
c. Abies cephalonica – răspândire şi diversitate ecologică Specia apare în Grecia meridională, între 700-1700 m altitudine, mai exact în Peloponezia
(masivele Taygete, Parnon, Mainalon), insula Cephalonia şi sudul Greciei continentale (Attica, Eubeea, Beotia, Focid şi Etolia).
În munţii din nordul Greciei şi sudul Bulgariei, între 700-1500 m (masivul Pind, muntele Olimp, muntele Athos, insula Thasos), genul este reprezentat printr-o specie considerată hibrid fixat între Abies alba şi A. cephalonica, cunoscută sub numele de Abies borisii regis.
d. Abies cilicica Această specie este răspândită în nordul Libanului (pădurea Ehden, Wadijahannam, Dkebel
Ganmmoua), între 1200 şi 1700 m altitudine. Ea mai apare şi în nord-vestul Siriei (Alouites) în masivele Taurus, Antitaurus, Amamus, între 1300 şi 2000 m.
e. Abies numidica Această specie de brad are un areal restrâns în masivul Babors în Kabylia Orientală în
Algeria, între 1300-2000 m, în etajul montan umed, în amestec cu o specie de stejar (Quercus mirbecki). Numele îi vine de la vechea denumire Numidia, dată de către romani regiunii de nord-est a Algeriei.
f. Abies marocana – răspândire şi diversitate ecologică Este o specie endemică răspândită în munţii Rif din Maroc, în regiunile Xauen şi Tazaot,
între 1200-2000 m altitudine. Specia din regiunea Tazaot a fost descrisă ca o specie distinctă sub numele de Abies
tazaotana (S. Cozar, 1948). Bradul de Maroc este considerat de către unii autori (Trabut, Maire) ca o subspecie a
bradului de Spania, Abies pinsapo ssp. marocana. g. Abies pinsapo – răspândire şi diversitate ecologică Bradul de Spania este o specie endemică cu areal foarte restrâns şi fragmentat, în munţii
din sudul Spaniei, în regiunea Ronda, între 1200-1800 m altitudine: Sierra de la Nueve, Yunquera – Sierra del Pinar, Sierras Bermeja şi Estepona.
În munţii Siciliei există o staţiune relictară – Vallone della Madonna degli Angeli – unde a existat o specie de brad cu un areal mult mai întins, Abies nebrodensis.
2. Asia a. Abies spectabilis Alte denumiri ale speciei: A. webbiana, A. densa. Este o specie de etaj montan superior şi subalpin din masivul Himalaya, cu o largă
răspândire din Hindus şi până în Bhutan, între 3000-4200 m altitudine.
29

Diversitatea ecologică a speciilor forestiere
b. Abies pindrow – răspândire şi diversitate ecologică Această specie constituie brădete în etajul montan umed al Himalayei occidentale, din
Afganistan în Nepal, între 2000-3000 m altitudine, adesea asociat cu Picea smithiana şi câteodată cu Cedrus deodara şi Pinus griffithii, alături de alte foioase.
O subspecie cu ace mai scurte a fost descrisă sub numele de Abies gamblei (Hickel 1929), care este sinonim cu Abies pindrow brevifolia (Dall. et Jackson).
c. Abies sibirica – răspândire şi diversitate ecologică Această specie are un areal foarte întins în Siberia, din Rusia europeană şi până la estul
Lacului Baikal, unde este un element esenţial al pădurilor de răşinoase, în amestec cu Picea obovata, Pinus sibirica, Larix sibirica şi Larix gmelini.
O specie asemănătoare a cărui areal a fost izilat în munţii Ciatkali a fost descrisă sub numele de Abies semenovii.
d. Abies nephrolepis – răspândire şi diversitate ecologică Este larg răspândită în Manciuria (bazinul fluviului Amur) şi în Coreea, unde se găseşte în
pădurile de munte, la altitudini de 700-1500 m, în amestec cu Picea ajanensis, Pinus koraiensis şi Larix gmelini.
O specie asemănătoare există în Peninsula Kamceatka, denumită Abies gracilis. e. Abies sacchalinensis – răspândire şi diversitate ecologică Specia există în pădurile de răşinoase în amestec cu Abies nephrolepis şi picea ajanensis,
în provincia maritimă a Siberiei Orientale. Speciile următoare ar trebui considerate ca rase geografice:
- Abies mayriana – importantă specie din insula Hokkaido unde ea constituie păduri de răşinoase, alături de Picea jezoensis şi Picea glehnii, până la 900 m în centrul insulei;
- Abies nemorensis – în insula Hokkaido, - Abies wilsoni – în sudul Sachalinului.
f. Abies holophylla Arealul acestei specii este localizat în sudul teritoriului maritim al Siberiei Orientale,
Manciuria, nordul Coreei şi fragmentar în sudul Coreei, între 300-1200 m altitudine. g. Abies koreana Această specie de brad apare pe un areal restrâns şi fragmentat în zona montană a Coreei
de Sud, la altitudini de peste 1000 m. h. Abies firma este o specie endemică ce apare în Japonia (insulele Kyushu, Shikoku, Honshu), cu un
optim de vegetaţie între 300-800 m, dar poate ajunge şi la 1600 m în sudul insulei Honshu. i. Abies homolepis Specie endemică de brad în Japonia, în etajul montan, între 1000-2200 m în insula Honshu,
sporadic în insulele Kyushu şi Shikoku. j. Abies mariesii Specie endemică în Japonia, în uona montană din centrul şi nordul insulei Honshu, la
1800-2500 m altitudine. k. Abies veitchii De asemenea specie endemică în Japonia, în insula Honshu, la 1600-1900 m altitudine.
30

Diversitatea ecologică a speciilor forestiere
Pe teritoriul Chinei, vegetează o serie de specii aparţinând genului Abies, acestea având un rol important în constituirea pădurilor de răşinoase de mare altitudine, cu o ecologie şi chiar o morfologie destul de imprecisă şi puţin studiată: Abies ernestii, A. chensiensis, A. recurvata, A. squamata, A. delavayi, A. faxoniana, A. sutchuenensis.
3. America de Nord a. Abies balsamea Este o specie tipică a pădurii boreale din nordul S.U.A. şi mai ales din estul şi centrul
Canadei, del anivelul mării şi până la 1600 m altitudine. Această specie de brad vegetează în arborete pure sau amestecate alături de Picea rubens, P. glauca şi P. mariana, de laricele american Larix laricina, precum şi de o serie de specii de foioase Populus tremuloides, P. grandidentata şi Betula papyrifera.
b. Abies fraseri Specie cu un areal restrâns în Virginia, Carolina de Nord şi Tennessee, la 1200-1800 m
altitudine. c. Abies concolor Specia are un areal vast în Munţii Stâncoşi, din Idaho şi Wyoming în nord şi până în Mexic
în sud, la 1800-3200 m altitudine, fiind întâlnită alături de duglasul brumăriu (Pseudotsuga glauca) şi pinul galben (Pinus poderosa) şi mai rar Picea engelmanni şi Picea pungens.
d. Abies lowiana Această specie este considerată în SUA ca fiind forma de coastă a speciei anterioare. Ea se
găseşte în Oregon (900-1800 m) şi California (Sierra Nevada). Apare alături de duglasul verde (Pseudotsuga menziesii), Pinus lambertiana şi Pinus ponderosa.
e. Abies grandis Specia are un areal destul de întins în regiunile maritime vestice, pe coasta pacifică a
Canadei, în partea interioară umedă a Columbiei Britanice şi a insulei Vancouver, iar în SUA de-a lungul coastei pacifice, până în zona San Francisco.
f. Abies amabilis Specie din zona de coastă a Pacificului, din Alaska de Nord şi până în Oregon. În
Columbia Britanică această specie se găseşte alături de Picea sitchensis, Tsuga heterophylla, Pseudotsuga menziesii şi Pinus monticola, iar în SUA (Washington, Oregon) apare la 1800 m alături de Abies lowiana şi A. lasiocarpa.
g. Abies lasiocarpa – răspândire şi diversitate ecologică Specie cu un areal extrem de vast în vestul Americii de Nord, ea fiind prezentă din Alaska
şi până în Arizona şi New Mexico şi formează amestecuri cu Tsuga mertensiana şi Picea engelmanni.
O varietate morfologică din zonele sudice a fost descrisă ca fiind specie separată, Abies arizonica.
h. Abies magnifica Acest brad creşte în lanţul muntos din Oregon şi California, constituind arborete pure la
altitudini superioare celor unde vegetează Abies lowiana, cu un optim între 1500-3000 m, sau arborete de amestec, alături de Pseudotsuga menziesii, Pinus lambertiana, Pinus monticola, Pinus ponderosa şi Sequoiadendron giganteum.
31

Diversitatea ecologică a speciilor forestiere
i. Abies bracteata Specie diseminată în California, cu un areal foarte restrâns, ce apare la 600-1500 m. j. Abies procera Specie diseminată ce vegetează între 800-1800 m, în zona Cascadelor
(Washington→California), alături de Pseudotsuga menziesii, Tsuga heterophylla, Abies amabilis, Pinus monticola, Abies lowiana şi Pinus lambertiana.
i. Abies religiosa Specia are cea mai largă răspândire în Mexic, formând arborete între 2600-3500 m în
munţii din centrul regiunii (Jalisco, Michoacan, Mexico, Hidalgo, Guerrero). Alături de această specie mai apar în zona Mexicului următoarele specii de brad: Abies hickeli, A. guatemalensis, A. durangensis, A. vejari, A. mexicana.
II. Genul Keteleeria Carr. Cuprinde arbori cu frunze aciculare persistente, asemănătoare celor de brad, cu areal
exclusiv asiatic, în regiuni cu climat temperat cald. Principalele specii sunt: - Keteleeria davidiana – China (Hou-pe, Kouei-chu, Se-Chuan), etajul temperat cald, - Keteleeria evelyniana – China (Yunnan, Si-Kang), etajul temperat cald, - Keteleeria fortunei – China de Sud (Che-Kiang, Yunnan), - Keteleeria roulletii – Laos şi Vietnam (800-1500 m), arborete pure sau cu Pinus
khasya, Pinus merkusii şi Quercus griffithii.
III. Genul Cathaya Chun et Kuang (1958). Cuprinde arbori de talie mijlocie, frunze aciculare, localizate exclusiv în China. Singurele
două specii descrise sunt: - Cathaya argyrophylla în regiunea Kuang Si, - Cathaya nanchuanensis în regiunea Se-Chuan.
IV. Genul Pseudotsuga Carr (1867). Cuprinde specii de arbori originare din America de Nord şi Extremul Orient. Aceştia au
dimensiuni mari, ritidom gros, ramificaţii regulat verticilate, scoarţa mult timp netedă cu pungi de răşină. Mugurii sunt mari, ovoconici, ascuţiţi, roşiatici, dispuşi terminal pe lujerii laterali întotdeauna câte unul. Acele sunt liniare, turtite, ascuţite, aşezate pe un umeraş scurt, nedecurente. Conurile sunt pendente, şi prezintă bractei lungi trifidate.
1. America de Nord a. Pseudotsuga menziesii – răspândire şi diversitate ecologică Duglasul verde apare în zona coastei Pacificului, într-un climat maritim, în Columbia
Britanică şi statele Washington, Oregon, în lanţurile muntoase şi în Sierra Nevada în California. În pădurile de duglas, la altitudini mici şi mijlocii, până la 350 m, el este asociat cu tuia gigantă (Thuja plicata), Tsuga heterophylla, Abies grandis. La altitudini mai ridicate, duglasul apare alături de Abies amabilis, A. procera şi Pinus monticola, iar în partea septentrională apare alături de Picea sitchensis. Din contră, în partea meridională apare alături de Pinus ponderosa, P. lambertiana, Calocedrus decurrens, iar în regiunea de coastă californiană formează amestecuri cu Sequoia sempervirens.
În Europa s-a introdus acum 150 ani, iar la noi acum 100 ani pe suprafeţe apreciabile pe raza ocoalelor silvice Aleşd, Marghita, Dobreşti, bazinul Nădrag, Crisbav, Râşnov, Săcele, Fântânele etc. (aprox. 30 000 ha). În condiţiile ţării noastre, cele mai bune rezultate au dat provenienţele montane, cu caracter relativ continental.
32

Diversitatea ecologică a speciilor forestiere
b. Pseudotsuga glauca – răspândire şi diversitate ecologică Această specie considerată de unii autori (Franco, 1950) ca varietate a speciei precedente
(Pseudotsuga menziesii var. glauca), se găseşte în centrul şi sudul Munţilor Stâncoşi, în Canada în interiorul Columbiei Britanice şi la vest de Alberta in SUA, din Montana şi Wyoming până în Colorado, Utah, Arizona şi New Mexico.
El formează arborete amestecate cu Pinus ponderosa, Picea engelmanni, Abies concolor, până la 1800 m altitudine.
În afară de specia tipică, în sudul Munţilor Stâncoşi şi în munţii din Mexic, între 2300-2900 m altitudine, există forme de Pseudotsuga care au fost ridicate la rang de specie şi care au o morfologie, dar mai ales un areal puţin studiat. Acestea sunt:
- Pseudotsuga macrolepis – Mexic (Durango, Nuevo Leon), - Pseudotsuga flahaulti – Mexic (Sonora, Chihuahua, Durango, Zacatecas, Coahuila), - Pseudotsuga guinieri – Mexic (Chihuahua, Durango), - Pseudotsuga rehderi – Mexic (Chihuahua, Durango, Nuevo Leon),
c. Pseudotsuga macrocarpa – răspândire şi diversitate ecologică Această specie este răspândită destul de puţin, în sudul Californiei, într-un climat arid
(350-500 mm anual), la altitudini de 500-2000 m. La altitudini joase vegetează alături de Pinus ponderosa şi Pinus jeffreyi, iar la altitudini mari alături de Calocedrus decurrens şi Abies concolor.
2. Asia Genul are specii reprezentative în China, Japonia şi Taiwan, acestea fiind: - Pseudotsuga sinensis – China (Yunnan, Si-Kang), 1500-2400 m, în amestec cu Tsuga
chinensis, T. yunnanensis, Picea complanata şi foioase diverse; - Pseudotsuga japonica – Japonia, sudul insulei Honshu (400-1100 m) şi în insula
Shikoku (500-900 m); - Pseudotsuga wilsoniana – Taiwan, specie endemică, vegetează la 2800 m altitudine; - Pseudotsuga forrestii – China, valea Mekong, vestul Yunnan, 3000 m altitudine.
V. Genul Tsuga (Endl) Carr (1855). Cuprinde specii de arbori originare din America de Nord şi Asia. Ramificaţia nu este
verticilată datorită creşterilor rezultate din mugurii laterali, frunzele sunt aciculare, persistente, clar peţiolate, turtite dorso-ventral, dispuse pectinat. Conuri mici, întregi, pendente, cu bractei mici, nevizibile. Genul conţine specii din America de Nord şi Asia.
1. America de Nord a. Tsuga heterophylla Specie foarte importantă de-a lungul coastei Pacificului, din Alaska şi până în zona San
Francisco, fiind întâlnită de la nivelul mării şi până în etajul montan superior, la 2400 m altitudine.
În Columbia Britanică poate constitui arborete pure sau în amestec cu Picea sitchensis în nord, Thuja plicata în centrul continentului, iar în altitudine cu Abies amabilis. PE mari suprafeţe este asociată cu duglasul verde (Pseudotsuga menziesii).
b. Tsuga mertensiana Specie de regiune pacifică maritimă, din Alaska şi până în Oregon. Este caracteristică
zonei subalpine, în interiorul continentului fiind în arborete pure sau în amestec cu Abies lasiocarpa, Larix lyalli, Picea engelmanni, şi Pinus albicaulis, iar în zona de coastă pecia este însoţită de Abies amabilis şi Chamaecyparis nootkatensis.
33

Diversitatea ecologică a speciilor forestiere
c. Tsuga canadensis Specie foarte răspândită în Canada (Provence maritime, Quebec, Ontario) şi în SUA în
munţii Appalachi, alături de Pinus strobus, Abies balsamea, Picea rubens, Picea glauca, iar dintre foioase, Acer saccharum şi Betula lutea.
d. Tsuga caroliniana Specie răspândită în Appalachi, din Virginia în Georgia. 2. Asia a. Tsuga chinensis Specie foarte răspândită în centrul şi vestul Chinei, în arborete pure sau amestecate, între
2000-3000 m altitudine. b. Tsuga diversifolia Specie montană endemică, ce apare în Japonia, în centrul insulei Honshu (Nikko,
Chichibu, Alpii japonezi) şi mai rar în insulele Shikoku şi Kyushu. Genul mai are specii reprezentative în China, Japonia şi Taiwan, acestea fiind: - Tsuga sieboldii – Japonia, - Tsuga yunnanensis – sud-vestul Chinei, 300-1400 m altitudine, - Tsuga dumosa – larg răspândită în centrul şi estul masivului Himalaya, 2400-3500 m.
VI. Genul Picea A. Dietr. (1854). Cuprinde circa 40 specii de arbori de talie mare, originare din Europa, America de Nord şi
Asia. Speciile acestui gen se întâlnesc la altitudine mică în regiunile septentrionale, unde anumite specii constituie limita zonei forestiere boreale în Asia şi America, şi, de asemenea, în etajele montan şi subalpin în regiunile meridionale.
Sunt specii mult mai puţin pretenţioase faţă de condiţiile staţionale în raport cu speciile genului Abies, dar mai exigente decât pinii: rezistenţi la ger, nu suportă seceta şi nu pot rezista la vânt.
Frunzele sunt persistente, aciculare, tetramuchiate (secţia Eupicea) sau turtite dorso-ventral (secţia Omorica), ascuţite, aşezate spiralat pe lujer pe perniţe proeminente ce rămân pe lujer după căderea acelor dând acestora un aspect zgrăbunţos.
Conurile sunt pendente, au solzi pieloşi, persistenţi şi bractei ascunse. Seminţe sunt prinse într-o aripioară ca într-o linguriţă.
Speciile sunt repartizate în diferite regiuni floristice: Eurasia septentrională, regiunea himalayană, vestul Chinei şi Taiwan, insulele de nord-vest asiatice, estul Americii de Nord, vestul Americii de Nord.
1. Regiunea Europa-Asia septentrională a. Picea abies – răspândire şi diversitate ecologică Arealul general al molidului este exclusiv european, mai întins decât al bradului, cu un
contur foarte neregulat, apărând în următoarele masive muntoase central şi est europene: Alpi, Jura, Vosgi, Pădurea Neagră, Harz, Carpaţi, Alpii Dinarici, Rhodopi. Lipseşte din Pirinei şi Apenini.
Din Alpi urcă în Scandinavia, spre nord-est merge până în regiunea Cazan unde se întâlneşte cu Picea obovata, coboară spre Carpaţi (evită zonele stepei ruseşti de la nord de Marea Neagră); se întinde de-a lungul Alpilor Dinarici, apare insular în munţii Apuseni şi Balcani; în nordul arealului devine arbore de câmpie. În nordul continentului este zona majoritară a arealului: Scandinavia şi nordul Rusiei.
34

Diversitatea ecologică a speciilor forestiere
Ca rase geografice amintim: - molidul scandinav (rezistent la ger şi la ciuperci parazite, cu creşteri lente şi productivitate
redusă), - molidul subalpin, rasa carpatică (mare capacitate de creştere şi adaptare), - rasele din Alpi (diferă prin culoarea scoarţei şi forma solzilor la conuri).
În România molidul este specie montană şi subalpină, constituind subzona fitoclimatică a molidului. El ocupă 22% din suprafaţa fondului forestier (peste 1 milion de hectare) şi apare în întregul lanţ carpatic, cu excepţia Semenicului, Almăjului, Locvei.
Limita superioară altitudinală în Carpaţii Orientali este de 1500-1550 m, în partea nordică, iar în Carpaţii Meridionali este de 1700-1750 m, max. 1860 m în Munţii Sebeşului (coincide cu limita superioară a pădurii). Ca arbore izolat (port în drapel) urcă la 2000 m în munţii Rodnei şi Călimani. Maximum de extindere îl înregistrează în Carpaţii Orientali, unde apare concentrat în partea nordică şi pe clina transilvăneană.
Limita inferioară de apariţie a molidului este la (600) 700-800 m în Carpaţii Orientali şi 900-1000 m în Carpaţii Meridionali. În lanţul vulcanic Călimani-Harghita, atinge altitudinea minimă (500 m în nordul Moldovei, 538 m la Gurghiu). Creşterile maximale din punct de vedere zonal le înregistrează pe valea Bistriţei, la Tulgheş, Broşteni, Cârlibaba, Ciuc, unde molidul îşi manifestă caracterul de specie pionieră, având pe alocuri caracter invadant.
Ca şi climatipuri, trebuie de precizat: • insulele de molid din Apuseni şi Poiana Ruscăi, • molidul din depresiunile carpatice transilvănene ale Mureşului şi Oltului, adaptate la
climat rece, cu precipitaţii sărace (limita inferioară a suportabilităţii speciei), • populaţiile vestice adaptate la un climat mai cald, molidul din Carpaţii de Curbură, unde
acesta găseşte un climat adăpostit cu ierni lungi şi precipitaţii abundente, • molidul din Carpaţii de Nord, adaptat la un climat rece cu sezon scurt de vegetaţie şi
zăpezi abundente. Din punct de vedere ecologic se diferenţiază ecotipuri edafice, cum ar fi:
molidul de stâncării calcaroase, molidul de luncă, molidul de turbării. b. Picea obovata Specie cu un areal vast în nordul Eurasiei, din nordul Scandinaviei până în Manciuria şi la
Marea Okotsk. Este una din speciile de bază din pădurile siberiene, vegetând alături de Abies sibirica şi Pinus sibirica.
c. Picea koyamai – răspândire şi diversitate ecologică Specie cu un areal foarte limitat, fiind întâlnită în Japonia, în insula Honshu, (munţii
Yatsugataka), la 1500-2000 m. Există opinia că molidul de Coreea, Picea koraiensis, poate fi raportat la aceeaşi specie.
d. Picea omorica Specie răspândită doar în Balcani, în sud-vestul Serbiei, pe valea Drinei, între 600-1400 m. e. Picea orientalis – răspândire şi diversitate ecologică Molidul oriental este o specie distinctă de toate alte specii ale genului Picea, ea fiind
caracteristică munţilor Caucaz, partea lor occidentală, la 700-2100 m altitudine, în amestec cu Abies nordmanniana, dar apare şi în masivul Taurus în nord-estul Turciei (600-2000 m).
În Extremul Orient, dar în partea septentrională, există o serie de specii ale genului Picea,
mai puţin cunoscute şi cu un areal destul de limitat:
35

Diversitatea ecologică a speciilor forestiere
36
- Picea schrenkiana – munţii Thianshan (Kirghizistan, China), - Picea ajanensis – sudul regiunii litorale a mării Okotsk, munţii Stanovoi, Kamceatka,
nordul Sacchalinului, - Picea microsperma – insula Hokkaido, centrul şi sudul Sacchalinului, insulele Kourile, - Picea komarovii – sudul Manciuriei şi Coreei de Nord, - Picea hondoensis – Japonia (insula Honshu), la 1400-2600 m altitudine.
2. Regiunea Himalaya a. Picea smithiana Specie răspândită din Afganistan până în Nepal, între 2000-4000 m, în amestec cu Cedrus
deodara, Abies pindrow, Abies spectabilis, şi Pinus griffithii. b. Picea spinulosa Specie din estul Himalayei, din Sikkim şi Bhutan. 3. Vestul Chinei şi insulele din nord-estul Asiei În vestul Chinei există o serie de specii ale genului Picea, nu pe deplin studiate, a căror
morfologie şi corologie nu sunt foarte bine cunoscute: - Picea asperata - Picea hetrolepis - Picea retroflexa - Picea aurantiaca - Picea gemmata - Picea meyerii - Picea neoveitchii
- Picea likiangensis - Picea balfouriana - Picea hirtella - Picea montigena - Picea yunnanensis - Picea wilsonii - Picea purpurea
- Picea brachytyla - Picea ascendens - Picea complanata - Picea sargentiana - Picea morrisonicola
Pe de altă parte, insulelor din nord-estul Asiei le sunt caracteristice specii endemice din genul Picea, specii care nu apar în partea continentală: - Picea glehnii - Picea bicolor - Picea maximowiczii - Picea polita
4. America de Nord a. Picea rubens (Picea rubra) Specie comună în Canada, în Noua Scoţie, Insula Prince Edouard, sudul Quebecului, dar şi
în SUA. În Maine, New York, Massachussets, munţii Appalachi. Cele mai multe arborete cu molidul roşu sunt localizate la mai puţin de 350 m altitudine, alături de Acer saccharum, Betula lutea, Abies balsamea, Tsuga canadensis şi Picea glauca.
b. Picea glauca – răspândire şi diversitate ecologică Este unul din coniferele cele mai importante din Canada, unde specia apare în arborete
pure sau în amestec cu Picea mariana şi Abies balsamea, de pe coasta atlantică şi până în Alaska. Alături de Picea mariana şi Larix laricina, Picea glauca constituie limita septentrională a vegetaţiei forestiere.
În SUA specia apare în statele din nord-est, cele din centru-nord, precum şi de-a lungul Munţilor Stâncoşi.
O subspecie regională a fost identificată sub numele de Picea albertiana, fiind în fapt considerată un hibrid între Picea glauca şi Picea engelmanni.
Picea glauca este foarte utilizat ca specie ornamentală, existând numeroase varietăţi: aurea, coerulea, coniqua, nana, pendula etc.

Diversitatea ecologică a speciilor forestiere
c. Picea mariana (Picea nigra) Specia este răspândită de pe litoralul atlantic şi până în Columbia Britanică şi Alaska şi
constituie alături de Picea glauca şi Larix laricina, limita septentrională a vegetaţiei forestiere.
d. Picea engelmanni Este o specie de etaj montan, fiind întâlnită în zona interioară vestică a continentului nord-
american, la altitudini de 300-1200 m în nordul arealului (Columbia britanică) şi de 1000-4000 m în sudul arealului (sudul Munţilor Stâncoşi).
e. Picea pungens Specie cu un areal vast, dar foarte fragmentat, situat în întregime în cadrul Munţilor
Stâncoşi. Este o specie diseminată între 1800-3300 m altitudine, în amestec la altitudini mari cu Picea engelmanni şi Abies lasiocarpa, iar la altitudini mici cu Pseudotsuga glauca, Abies concolor şi Pinus ponderosa.
f. Picea chihuahuana Specie endemică situată în Munţii Mexicului, la altitudini de 2350-2500 m. g. Picea sitchensis Molidul de Sitka este o specie cu areal tipic maritim, fiind situat în zona litorală a coastei
Pacificului unde umiditatea atmosferică este foarte mare, fiind întâlnit din California şi până în Alaska. Este o specie foarte importantă în Columbia Britanică. Formează arborete pure sau în amestec cu Tsuga heterophylla şi Pseudotsuga menziesii, iar mai în sud vegetează alături de Chamaecyparis lawsoniana. În zona montană molidul de Sitka creşte împreună cu Abies amabilis, Abies procera şi Pinus monticola.
h. Picea breweriana Specie cu un areal restrâns, limitat la lanţul Munţilor Siskiyon, în sud-estul statului Oregon
şi nord-vestul Californiei, între 1600-2100 m altitudine. VII. Genul Larix Miller (1754). Genul Larix cuprinde peste 10 specii de arbori din regiunile septentrionale şi înalte din
emisfera nordică, repartiţia lor fiind condiţionată de compensarea altitudinii cu latitudinea. Astfel, la latitudini mari, în zonele septentrionale speciile genului Larix sunt arbori de joasă altitudine, iar în zonele meridionale sunt caracteristice etajului montan superior şi subalpin.
Arborii au coroana rară cu verticile neregulate, prezentând lujeri lungi şi lujeri scurţi. Acele sunt liniare, moi şi caduce, pe lujerii lungi solitare, inserate spiralat, iar pe lujerii scurţi grupate în rozete sau smocuri. Florile sunt monoice, conurile au solzi pieloşi, persistenţi, iar maturaţia anuală.
Speciile de Larix sunt mai pretenţioase decât molizii, dar mai puţin exigente decât brazii, au temperamentul cel mai de lumină dintre toate răşinoasele şi sunt rezistente la ger, dar sensibili la secetă.
1. Europa şi Asia
a. Larix decidua – răspândire şi diversitate ecologică Laricele este o specie exclusiv europeană, cu un areal fragmentat şi concentrat numai pe
teritorii montane, cu centrul cel mai important în Alpi: în Alpii Occidentali ai Franţei şi Elveţiei ajunge la 2200 m (în amestec cu Pinus cembra şi Pinus uncinata), iar la altitudini mici în Alpii Centrali şi Orientali coboară la 400 m (subspecia decidua).
În Cehia şi Slovacia formează arborete de amestec cu fagul (300-800 m), iar în Polonia, de-a lungul Vistulei, ajunge la 150-600 m, devenind arbore de câmpie.
37

Diversitatea ecologică a speciilor forestiere
Laricele din România aparţine subspeciei carpatica şi se localizează în 5 centre muntoase: Ceahlău, Ciucaş, Bucegi, Lotru, Apuseni (Trascău-Vidolm); maximul răspândirii sale îl înregistrează în bazinul Latoriţei (Lotru), pe munţii Târnovul Mare şi Târnovul Mic.
Altitudinile minime se înregistrează în Apuseni (650 m), iar cele maxime sunt de circa 1820 m în Ceahlău şi de 2050 m în Bucegi.
Formează arborete pure numai în Ceahlău la Poliţa cu Crini sau în amestec cu Picea abies, Pinus cembra, Fagus sylvatica, Betula pendula etc.
Laricele este o specie relativ unitară din punct de vedere morfologic, dar se remarcă totuşi, ca urmare a compensării altitudinii cu latitudinea două rase geografice, fiecare cu ecotipurile ei: rasa vestică şi rasa estică.
Dintre ecotipuri amintim: − Ecotipul austriac al laricelui de Alpi caracteristic altitudinilor mici (350-800 m) de la
care s-a utilizat sămânţa pentru multe din plantaţiile de la noi; − Ecotipul austriac al laricelui de Alpi de stâncărie întâlnit natural la altitudini mari (2000-
2200 m); − Ecotipul laricelui de Sudeţi, ecotip de mică altitudine, caracterizat prin rezistenţă la
umbrire şi un potenţial ridicat de creştere; − Laricele din Polonia, cu ecotipuri de mică altitudine (150-600 m); − Laricele carpatic (Larix decidua ssp. carpatica), ce vegetează la altitudini mari, în
staţiuni puternic însorite, pe soluri scheletice; ca excepţii de la această regulă se semnalează ecotipurile de mică altitudine ce apar în Apuseni şi ecotipurile edafice (de exemplu, cele de soluri argiloase de pe terasele Sighişoarei).
b. Larix sibirica – răspândire şi diversitate ecologică Este o specie cu un areal foarte clar delimitat, fiind răspândită din nordul Rusiei până la
fluviul Enisei. De precizat faptul că în partea europeană Larix sibirica a fost descris sub denumirea de Larix sukaczewii, fiind separată într-o altă specie de sine stătătoare.
c. Larix gmelini – răspândire şi diversitate ecologică arealul acestei specii este foarte întins, din Siberia Centrală până în partea nordică a
peninsulei Kamceatka şi Coreea de Sud. Astfel, au fost decelate mai multe rase geografice, ridicate la rang de subspecie:
- ssp. japonica – insulele Kouriles, Sacchalin şi Kamceatka, în arborete pure sau în amestec cu Abies sacchalinensis şi Picea ajanensis;
- ssp. olgensis – estul Siberiei, împărţită la rândul ei în două forme, Larix komarovii (coasta mării Japoniei) şi Larix middendorfi (litoralul mării Okotsk, Sacchalin şi Kamceatka);
- ssp. principis rupprechtii – nordul Chinei şi Coreea. d. Larix griffithii Specia apare în estul Himalayei, mai exact în estul Nepalului, Sikkim şi Bhutan, în etajul
montan, la 2700-3700 m altitudine. e. Larix leptolepis – răspândire şi diversitate ecologică specia este endemică în insula japoneză Honshu, unde are un areal destul de restrâns. Cele
mai întinse arborete se găsesc l a1200-2400 m altitudine, unde formează arborete pure sau în amestec cu alte răşinoase: Abies homolepis, Abies veitchii, Picea hondoensis, Tsuga diversifolia.
Este o specie cu o plasticitate ecologică remarcabilă şi este imună la atacul cancerului provocat de Dasyscypha wilkomii, motiv pentru care Larix leptolepis a fost hibridat cu Larix decidua. Astfel, s-a reuşit transferul genei ce determină imunitatea sus-menţionată, hibridul
38

Diversitatea ecologică a speciilor forestiere
caracterizându-se morfologic prin caractere intermediare între genitori. Hibridul se numeşte Larix × eurolepis.
2. America de Nord a. Larix laricina – răspândire şi diversitate ecologică Specia are o repartiţie transcontinentală în America de Nord, de pe coasta Atlanticului şi
până în Alaska, unde o subspecie – ssp. alaskensis – ajunge la limita boreală a vegetaţiei forestiere. Limita sudică este trece prin SUA (Maine, Pensilvania) şi Canada (Saskatchewan, Alberta). Constituie arborete amestecate alături de Abies balsamea, Picea glauca, Picea mariana, Populus tremuloides, Betula papyrifera, Betula lutea.
b. Larix occidentalis Specie de etaj montan, între 600-2000 m, în vestul Americii de Nord, fiind cu răspândire
limitată la sud de Columbia Britanică, în Canada şi SUA. Apare alături de Pseudotsuga menziesii, Pinus monticola, Pinus contorta, Picea engelmanni, Abies subalpina.
c. Larix lyalli Specie de etaj subalpin, în vestul Americii de Nord, ajunge la limita vegetaţiei forestiere în
Munţii Stâncoşi, la 2500-3500 m. VIII. Genul Cedrus Trew (1757). Genul Cedrus cuprinde arbori de talie mare cu frunze aciculare persistente inserate izolat
pe ramurile lungi şi în buchete pe ramurile scurte. Înfloreşte la sfârşitul verii. Conuri mari, ovoide a căro solzi se desfac la maturitate de pe ax. Genul Cedrus este reprezentat prin patru specii din etajul montan din regiunile mediteraneană şi himalayană.
a. Cedrus atlantica – răspândire şi diversitate ecologică Specie montană din Africa de Nord: - în Maroc apare în Munţii Rif, unde apare la peste 1400 m pe versanţii atlantici şi peste
1600 m pe versanţii mediteraneeni., în amestec cu Abies marocana şi Quercus mirbecki. În Munţii Atlas cedrul apare la 1600-2200 m;
- în Algeria, arealul este mult mai fragmentat, unde vegetează alături de Abies numidica şi Quercus mirbecki.
b. Cedrus libani Specie de zonă montană din regiunea mediteraneană orientală. În Liban apare în câteva
staţiuni la 1500-1700 m, iar în Siria se întâlneşte dincolo de 1000 m altitudine, alături de specii de stejar cu frunze căzătoare: Quercus cerris, Q. libani, Q. infectoria. La peste 1200 m specia este însoţită de Abies cilicica. În Turcia arealul speciei este foarte fărâmiţat, apărând în sudul Anatoliei, în masivele Taurus şi Maras, la 1250-2000 m, în amestec cu Pinus nigra ssp. pallasiana, Juniperus excelsa, Juniperus foetidissima, Abies cilicica, iar la altitudini mai joase alături de specii de foioase (Fraxinus ornus, Ostrya carpinifolia, Acer monspessulanum).
c. Cedrus brevifolia Este o specie endemică ce apare în Cipru, la 1400-1700 m, în amestec cu o specie de stejar
endemică, Quercus alnifolia. d. Cedrus deodara Specie importantă a pădurilor montane de răşinoase din vestul Munţilor Himalaya, din
Afganistan până în Nepal, la 1700-3000 m altitudine. Astfel, arealul este vast, iar specia ocupă diferite tipuri de păduri:
39

Diversitatea ecologică a speciilor forestiere
- păduri umede (2200-2700 m), alături de Pinus griffithii, deasupra etajului de Pinus roxburghii,
- la peste 2700 m cedează locul speciilor Abies pindrow şi Picea smithiana, - păduri mai puţin umede, specia este abundentă în etajul montan mijlociu, deasupra
etajului de Pinus gerardiana, - între 2300-2700 m apare în arborete pure, acest etaj fiind ocupat de păduri cu Abies
pindrow, Picea smithiana şi Betula utilis. VIII. Genul Pinus L. (1753). Acest gen cuprinde un număr mare de specii lemnoase din emisfera nordică, peste 120
specii de arbori, rar arbuşti, cu numeroşi infrataxoni. Pe lângă speciile din Emisfera Nordică, mai sunt specii şi în sud-vestul Asiei, precum şi câteva specii în regiunile ecuatoriale.
Prezintă frunze aciculare persistente, grupate câte două, trei sau cinci într-o teacă membranoasă, astfel că speciile genului sunt structurate în două subgenuri, trei grupe şi şapte secţii, după cum urmează1:
GENUL PINUS
Subgenul DIPLOXYLON
Koehne
Cu 2 ace
1. Secţia Eupitys Spach
P. sylvestris L. P. montana Mill. P. nigra Arn. P. merkusii
2. S. Banksia Mayr. P. banksiana Lamb. P. pinaster Ait.
3. S. Pinea Endl. P. pinea L.
Cu 3 ace 4. Secţia Taeda Spach. P. rigida Mill. 5. S. Pseudostrobus P. ponderosa Laws.
Subgenul HAPLOXYLON
Koehne Cu 5 ace
1. S. Strobus Sweet
P. strobus L. P. excelsa Vall. P. peuce Grieseb P. monticola Dougl.
2. S. Cembrae Spach P. cembra L. P. sibirica Mayr.
Florile sunt unisexuat-monoice, din care cele mascule grupate în amenţi, iar amenţii în buchete, iar cele femele sunt conuleţe. Conurile au solzi îngroşaţi la vârf (apofiza), cu o proeminenţă (umbelic) prevăzută uneori cu un ghimpe; la unele specii pe apofiză se observă o muchie transversală (carenă). Bracteile sunt avortate, iar seminţele sunt obişnuit aripate, prinse în aripioară ca într-un cleşte. Maturaţia este bienală, iar diseminaţia are loc în primăvara celui de-al treilea an.
Speciile genului Pinus sunt rezistente la ger, secetă, nepretenţioase faţă de condiţiile edafice, reuşind să pună în valoare solurile oligotrofe şi staţiunile extreme.
SUBGENUL HAPLOXYLON
1. Europa şi Asia a. Pinus cembra – răspândire şi diversitate ecologică Arealul general al zâmbrului este exclusiv european, fiind caracteristic etajului subalpin al
Alpilor şi Carpaţilor, vegetând alături de Picea abies, Larix decidua, Pinus uncinata, Pinus mugo, Juniperus communis ssp. nana.
În România vegetează la limita superioară a zonei forestiere, în staţiunile fostelor circuri glaciare din Munţii Rodnei, Călimani, Bucegi, Făgăraş, Parâng, Ţarcu, Godeanu, Retezat. În Retezat coboară pe văile umede la 1200 m, în Călimani coboară diseminat în molidişurile de limită la 1400 m şi urcă în jnepenişuri până la 1800 m.
O altă specie asemănătoare, Pinus sibirica, înlocuieşte zâmbrul în Siberia, între Urali şi lacul Baikal, la altitudini mici. 1 Sunt precizate doar câteva specii reprezentative
40

Diversitatea ecologică a speciilor forestiere
Dincolo de Lacul Baikal există o specie similară, Pinus pumila, ce constituie arborete la limita superioară a vegetaţiei forestiere în munţii din nord-estul Asiei (Manciuria, Kamceatka, Sacchalin, Japonia – Honshu şi Hokkaido).
b. Pinus peuce Specie endemică în Balcani (Muntenegru, Albania, Macedonia, Bulgaria), fiind întâlnită în
etajul montan superior şi subalpin până la 2200 m altitudine, alături de Picea abies sau în arborete pure.
c. Pinus griffithii (Pinus wallichiana) Specia este originară din Himalaia, unde vegetează la altitudini de 1600-4000 m, fiind un
arbore montan, întâlnit în arborete pure sau în amestec cu Cedrus deodara, Abies pindrow şi Abies spectabilis.
d. Pinus koraiensis Specie răspândită în Manciuria, China şi Coreea, la altitudini de 600-1200 m, iar în Japonia
apare în insulele Honshu şi Shikoku, între 1200-2600 m.
e. Pinus parviflora Specie endemică ce apare în zona montană din Japonia, la altitudini de 1300-2000 m, în
insula Honshu. În sudul Chinei apare o altă specie, Pinus kwangtungensis, în nordul Japoniei (Hokkaido) Pinus pentaphylla, iar în Vietnam apare Pinus dalatensis.
Tot în China, mai exact în centrul şi sudul ţării, apare Pinus armandi, specie foarte larg răspândită, între 300-3200 m.
2. America de Nord
a. Pinus strobus - – răspândire şi diversitate ecologică Pinul strob este una din principalele specii de răşinoase din estul Americii de Nord, fiind
întâlnit din sudul Canadei şi al SUA, zona Marilor Lacuri şi până în estul Munţilor Appalachi. Specia vegetează în arborete pure sau în amestec cu Pinus resinosa, Pinus banksiana, Abies balsamea, Picea rubens, Picea glauca, Betula papyrifera, sau alături de Tsuga canadensis, Acer saccharinum şi Betula lutea.
b. Pinus monticola Specia apare învestul Americii de Nord, cu temperament montan (Vancouver, Columbia
Britanică, Montana, California etc.), până la 3000 m latitudine. În partea septentrională a Munţilor Stâncoşi specia apare alături de Larix occidentalis, Picea engelmanni, Pinus contorta sau alături de Pseudotsuga menziesii şi Tsuga heterophylla sau Abies magnifica.
c. Pinus lambertiana Acest pin este specific vestului Americii de Nord, este specie numai de amestec, fiind
însoţit de Pseudotsuga menziesii, Tsuga heterophylla, Chamaecyparis lawsoniana şi Pinus ponderosa în nordul arealului (Oregon) şi de Abies lowiana, Pinus jeffreyi şi Sequoiadendron giganteum în sudul arealului (Mexic).
d. Pinus albicaulis Specie de etaj subalpin, în vestul Americii de Nord (asemănător zâmbrului), fiind specie
numai de amestec, alături de alte specii montane-subalpine: Pinus monticola, Pinus contorta, Tsuga mertensiana şi Picea engelmanni.
e. Pinus flexilis Specia este destul de răspândită, dar are un areal foarte fărâmiţat, la altitudini mari, între
1200-3000 m (Columbia Britanică) şi 2000-4000 m în Munţii Stâncoşi şi Mexic (Chihuahua).
41

Diversitatea ecologică a speciilor forestiere
De mică întindere, mai apar în America de Nord, ca şi în Asia: - Pinus cembroides – Mexic, sudul Californiei (1300-2250 m); - Pinus monophylla – munţi cu climat deşertic (SUA, Mexic); - Pinus quadrifolia – (California); - Pinus balfouriana – California, Sierra Nevada (1500-3500 m); - Pinus aristata – sudul Munţilor Stâncoşi (peste 2400 m); - Pinus bungeana – China; - Pinus gerardiana – nord-vestul Himalayei (1800-3000 m).
SUBGENUL DIPLOXYLON
1. Europa a. Pinus sylvestris – răspândire şi diversitate ecologică Pinul silvestru este o specie cu areal vast, ocupând peste 145 milioane hectare, ceea ce
înseamnă circa 3.7% din pădurile globului. El apare de la vest la est, de la Oceanul Atlantic la Oceanul Pacific (Europa, Asia), iar de la
nord la sud, de la peste 70° latitudine nordică (nordul Scandinaviei) la sub 40° latitudine (Peninsula Iberică).
În vastul areal, apare compensarea latitudinii cu altitudinea, astfel încât în regiunile nordice şi vestice apare frecvent ca arbore de câmpie (Scandinavia, Siberia, ţinuturile baltice), iar la est şi sud doar în regiunile montane (Pirinei, Alpi, Balcani, Carpaţi). În concluzie, la nivelul arealului general, limita altitudinală creşte de la nord la sud şi de la vest la est.
În România este puţin răspândit comparativ cu alte ţări, vegetând spontan, insular, în lungul Carpaţilor şi în Apuseni, pe stâncării aride, turbării etc. Cert este că actualele staţiuni sunt refugii ale pinetelor de dinaintea glaciaţiunii (circa 9000 ha, adică 0.14% din fondul forestier românesc), acolo unde alte specii lemnoase nu au reuşit să se instaleze datorită condiţiilor vitrege.
Zona de maximă răspândire este în bazinul Trotuşului, unde formează arborete pure sau în amestec cu molidul, fagul ,bradul, gorunul, precum şi bazinele Buzăului, Râmnicului, Putnei, Oituzului (ocoalele silvice Nehoi, Nehoiaşi, Dumitreşti, Vintilă Vodă, Năruja ş.a.)
Altitudinal, pinul silvestru apare între 300 m în Subcarpaţii Moldovei şi pe valea Oltului şi 1700 m în Retezat (1900 m în bazinul Gemenele).
În Cheile Bicazului, vegetează pe stâncării lipsite aproape de sol mineral, în Bucovina apare în bazinul Bistriţei (Barnar, Zugreni), precum şi în bazinul Moldovei la Pojorâta şi în masivul Răchitişul Mare (rezervaţie cu Arctostaphyllos uva-ursi). De asemenea, pinul silvestru apare şi în turbăriile oligotrofe din Depresiunea Dornelor (Poiana Stampei, Coşna, Grădiniţa, Şaru Dornei etc.).
Fiind o specie cosmopolită, pinul silvestru este reprezentat în zonele arealului său prin numeroase rase geografice. Acestea sunt: - Pinus sylvestris rigensis – pinul silvestru nordic, - Pinus sylvestris lapponica – pinul de Laponia (dincolo de 62° latitudine), - Pinus sylvestris vindelica – pinul din Austria de Jos (de mare altitudine), - Pinus sylvestris carpatica – pinul nord-carpatic (pe stâncării), - Pinus sylvestris rumunica – pinul silvestru românesc (Carpaţii Orientali), - Pinus sylvestris balcanica – pinul de Balcani, - Pinus sylvestris sibirica – pinul siberian (Munţii Altai), - Pinus sylvestris hamata – pinul de Caucaz.
Ca ecotipuri şi climatipuri autohtone, sunt de menţionat: - ecotipul de mare altitudine din Retezat, - climatipul est-carpatic din Carpaţii Orientali,
42

Diversitatea ecologică a speciilor forestiere
- climatipul vest-carpatic, ecotipul de soluri silicioase, oligotrofice (Cheile Bicazului, Răchitişul Mare),
- ecotipul de turbării (Poiana Stampei, Lucina, Luci). b. Pinus nigra – răspândire şi diversitate ecologică Specie colectivă a cărui areal este fragmentat, dar localizat în jurul bazinului Mediteranei.
Numeroase subspecii au fost ridicate la rang de specie, iar rasele geografice l arang de subspecie:
- ssp. Clusiana – Africa de Nord, Spania, sudul Franţei, - ssp. Laricio – Corsica, Sicilia, Calabria, - ssp. Nigricans – Austria, Serbia, Italia, Balcani, Grecia, - ssp. Pallasiana – Crimeea, Turcia.
c. Pinus montana – răspândire şi diversitate ecologică Specie cu areal fragmentat în etajul subalpin al munţilor din Europa, cu rase geografice
diferite din punct de vedere morfologic şi corologic pentru a putea fi considerate specii diferite:
- Pinus uncinata – Pirinei, Alpii meridionali (1600-2200 m), Jura, - Pinus mugo – Alpii Orientali (1500-2200 m), Carpaţi, Rodopi, - Pinus pumilio – Alpii Centrali (Boemia).
d. Pinus leucodermis Specie endemică în Balcani (Albania, Serbia, Grecia, Bulgaria) şi Calabria, la altitudini de
1500-2200 m. 2. Asia a. Pinus densiflora Una din principalele specii forestiere din Japonia, în toate cele patru mari insule (Honshu,
Shikoku, Kyushu, Hokkaido), de la nivelul mării şi până la 1400 m. Apare şi în China şi Manciuria.
b. Pinus thunbergii apare doar în zona litorală, pe dune, în Japonia, Taiwan şi Coreea de Sud.
c. Pinus massoniana Importantă specie cu areal destul de vast, din regiunea temperată caldă a Chinei,
Vietnamului şi Taiwanului.
d. Pinus tabulaeformis (Pinus sinensis) Specie forestieră importantă pentru centrul, nordul şi vestul Chinei. Alături de această
specie mai apare şi Pinus yunnanensis (1000-2300 m).
3. America de Nord
a. Pinus resinosa Este o specie destul de răspândită în nord-estul Americii de Nord, în regiunea atlantică , în
amestec cu Pinus strobus şi Pinus banksiana.
b. Pinus tropicalis Specie endemică din parte occidentală a Cubei, în regiunea tropicală.
c. Pinus ponderosa – răspândire şi diversitate ecologică Importantă specie din vestul Americii de Nord, cu un areal întins şi fragmentat, din partea
continentală a Columbiei Britanice şi până în New Mexico şi Arizona, fiind întâlnită în
43

Diversitatea ecologică a speciilor forestiere
amestec cu Pseudotsuga menziesii sau Pinus jeffreyi, Pinus lambertiana, Calocedrus decurrens şi Abies concolor.
Fiind o specie cu areal foarte vast, există câteva rase geografice, ridicate la rang de specie: Pinus scopulorum în centrul Munţilor Stâncoşi, Pinus arizonica în Arizona şi New Mexico.
d. Pinus jeffreyi Această specie de pin a fost considerată odinioară ca subspecie a lui Pinus ponderosa şi are
un areal limitat la munţii de pe coasta Pacificului, cele mai dezvoltate populaţii fiind în Sierra Nevada.
e. Pinus torreyana Areal restrâns numai la sudul Californiei (San Diego, Insula Santa Rosa). În zona tropicală a Americii de Nord şi a Mexicului, sunt următoarele specii de pin:
- Pinus rudis – Mexic, Guatemala (2200-4000 m), - Pinus michoacana – sudul Mexicului, - Pinus hartwegii – sudul Mexicului (3000-3800 m), - Pinus cooperi – Mexic (areal restrâns), - Pinus montezumae – Mexic, Guatemala, Honduras, - Pinus pseudostrobus – Mexic, Guatemala, Honduras, Nicaragua, Salvador, - Pinus oaxana – Mexic, Guatemala, Honduras (2100-3000 m), - Pinus tenuifolia – Mexic, Guatemala (climat subtropical, 1000-2100 m), - Pinus teocote – Mexic (1400-3000 m), - Pinus lawsoni – Mexic (1300-3000 m).
f. Pinus pinaster (Pinus maritima) – răspândire şi diversitate ecologică Pinul maritim are un areal destul de întins în regiunea mediteraneană occidentală, precum
şi pe ţărmul Oceanului Atlantic, în partea sud-vestică a continentului european. În cadrul acestui areal s-au distins câteva rase geografice:
- Rasa atlantică, răspândită în regiunea de sud-vest a Europei, în Franţa, Spania şi nordul Portugaliei. Ea a constituit rasa cea mai utilizată pentru împădurirea şi reîmpădurirea landurilor din Gasconia şi Bretania.
- Rasa mediteraneană, răspândită în Peninsula Italică, sud-estul Franţei şi nordul Africii. În Alpii maritimi urcă până la 1200 m, iar în Corsica până la 1000 m. În Africa de Nord, specia este prezentă în masive izolate de-a lungul litoralului algero-tunisian, precum şi în câteva zone muntoase unde ajunge la 700 m altitudine.
- Rasa de Maroc, unde pinul maritim constituie arborete montane în munţii Rif şi Atlas, localizându-se între 1000 şi 2000 m altitudine.
g. Pinus baksiana Arealul pinului banksian este cantonat în părţile reci ale Americii de Nord, unde se întinde
până dincolo de cercul polar. Specia apare în arborete pure sau în amestec cu Populus tremuloides şi Betula papyrifera.
h. Pinus contorta– răspândire şi diversitate ecologică Areal vast în nord-vestul Americii de Nord, de pe litoralul Californiei şi până în Columbia
Britanică, în etajul montan superior al munţilor Sierra Nevada şi Munţilor Stâncoşi. Ca rase geografice ridicate la rang de subspecie precizăm:
- ssp. contorta – Washington, Columbia Britanică, - ssp. bolanderi – California, - ssp. murrayana – amestecuri cu Abies magnifica şi Pinus flexilis, - ssp. latifolia – amestecuri cu Pseudotsuga menziesii şi Picea engelmanni.
44

Diversitatea ecologică a speciilor forestiere
45
i. Pinus rigida Estul Americii de Nord, cu areal restrâns în Canada şi SUA. j. Pinus palustris Importantă specie din câmpiile din zona de coastă din sud-estul SUA. k. Pinus elliotii Areal limitat la zona caldă şi umedă în Florida, Carolina de Sud şi Louisiana. Specia creşte
în amestec cu Pinus palustris şi specii de Quercus cu frunze persistente şi palmieri. l. Pinus taeda Răspândit în sud-estul SUA, la altitudini mijlocii şi joase, în arborete pure sau în amestec
cu Pinus echinata, P. serotina, P. elliotii. m. Pinus echinata Specie cu areal destul de mare, în tot sud-estul SUA, până la 900 m altitudine, alături de
Quercus velutina, Quercus marilandica, Quercus stellata, Quercus prinus şi Carya cordiformis.
Alte specii ale genului Pinus mai puţin importante din sud-estul SUA sunt: - Pinus serotina, - Pinus glabra, - Pinus virginiana, - Pinus clausa, - Pinus pungens, - Pinus patula, - Pinus oocarpa,
- Pinus gregii, - Pinus occidentalis, - Pinus attenuata, - Pinus muricata, - Pinus radiata (Pinus insignis), - Pinus coulteri, - Pinus sabiniana.
La acestea se adauga câteva specii din Mexic, la fel cu areal restrâns la câteva zone: - Pinus leiophylla, - Pinus chihuahuana, - Pinus lumhotzii.
n. Pinus halepensis Specie circummediteraneană, în Maroc (munţii Rif, Atlas), Algeria, Atlasul saharian,
Tunisia, Spania (Gibraltar, Catalonia), Franţa (Languedoc), Italia, Grecia. o. Pinus brutia Specia cu areal în regiunea mediteraneană orientală (Grecia orientală, Creta, Cipru, Turcia,
Siria). În regiunea insulelor Canare, într-un climat subtropical, apare Pinus canariensis, iar în
masivul himalayan, tot climat tsubtropical Pinus roxburghii (Pinus longifolia) (Afganistan, Birmania de Nord).
p. Pinus pinea Specie cu repartiţie circummediteraneană, din Portugalia până în Siria, cultivată de mult
timp pentru producţia de seminţe utilizate în patiserie.
FAMILIA TAXODIACEAE F.N.Neger.
În această familie sunt incluse specii de arbori din regiunile calde şi temperate ale emisferei nordice (Asia, America de Nord), de talie mare, cu frunze persistente sau caduce, aciculare sau solziforme şi care aparţin celor zece genuri:

Diversitatea ecologică a speciilor forestiere
- Taxodium, - Sequoia, - Cryptomeria, - Sciadopitys, - Metasequoia,
- Cunninghamia, - Taiwania, - Athrotaxis, - Sequoiadendron, - Glyptostrobus.
FAMILIA CUPRESSACEAE F.N.Neger.
Familia include circa 140 specii de arbori sau arbuşti cu frunze opuse sau verticilate, solziforme sau aciculare, persistente, cu flori unisexuat monoice sau dioice. Conurile sunt variabile, uscate sau cărnoase, cu solzi aşezaţi imbricat sau valvat, opus sau verticilat.
a. genul Cupressus L. Dintre multitudinea de specii din genul Cupressus fac parte: - Cupressus sempervirens – bazinul Mediteranei, - Cupressus cashmeriana – India, - Cupressus funebris – China, - Cupressus torulosa – Himalaya, China, - Cupressus duclouxiana – China, - Cupressus abramsiana – California, - Cupressus arizonica – Arizona, Texas, New Mexico, - Cupressus bakeri – California, Oregon, - Cupressus macrocarpa – California, cultivat în bazinul Mediteranei.
b. genul Chamaecyparis Spach. Speciile genului sunt răspândite în America de Nord şi Asia: - Chamaecyparis lawsoniana - Chamaecyparis nootkatensis - Chamaecyparis thujoides
- Chamaecyparis obtusa - Chamaecyparis pisifera - Chamaecyparis formosensis
c. genul Thujopsis Sieb. Et. Zucc. Singura speciedin aceste gen este Thujopsis dolabrata, specie endemică din Japonia.
d. genul Thuja L. Speciile acestui gen se localizează, ca origine, în America de Nord şi Extremul Orient:
- Thuja plicata (T. gigantea) - Thuja occidentalis - Thuja orientalis (Biota orientalis)
e. genul Juniperus - Juniperus drupacea – Grecia, Orientul Apropiat - Juniperus communis – Europa, Asia de Nord, America de Nord, cu
numeroase rase geografice, varietăţi sau subspecii: var. hemisphaerica – regiunea mediteraneană var. nana – Europa în regiunile înalte sau septentrionale var. depressa – America de Nord var. nipponica – Japonia
- Juniperus oxycedrus – bazinul Mediteranei - Juniperus rigida – Japonia, Coreea - Juniperus conferta – Japonia - Juniperus formosana – China, Taiwan - Juniperus sabina – Europa
46

Diversitatea ecologică a speciilor forestiere
- Juniperus horizontalis – America de Nord - Juniperus phoenicea – bazinul Mediteranei - Juniperus thurifera – Spania, Africa de Nord - Juniperus excelsa – Balcani, Asia Mică - Juniperus polycarpos – Asia Mică - Juniperus foetidissima – Asia Mică - Juniperus procera – Africa Orientală (Etiopia, Somalia, Kenya, Tanganika) - Juniperus silicicola – sud-estul SUA - Juniperus osteosperma – sudul Munţilor Stâncoşi, Mexic - Juniperus monticola – sudul Munţilor Stâncoşi, Mexic - Juniperus california – sudul Munţilor Stâncoşi, Mexic - Juniperus flaccida – sudul Munţilor Stâncoşi, Mexic - Juniperus deppeana – sudul Munţilor Stâncoşi, Mexic - Juniperus pachyplaea – sudul Munţilor Stâncoşi, Mexic - Juniperus virginiana – estul Americii de Nord - Juniperus scopulorum – vestul Americii de Nord - Juniperus chinensis – China, Coreea, Japonia
FAMILIA TAXACEAE S.F.Gray
Această familie cuprinde arbori şi arbuşti ce aparţin la 5 genuri: Taxus, Torreya, Amenotaxus, Austrotaxus, Nothotaxus.
Sub aspect morfologic se caracterizează prin frunze liniare, decurente, persistente, inserate spiralat şi dispuse pectinat. Florile sunt obişnuit dioice, cele mascule izolate la subsuoara frunzelor, iar cele femele solitare, cu câte un singur ovul. După fecundare, baza ovulului generează un aril care acoperă parţial sau total sămânţa. Plantula are două cotiledoane, iar lemnul nu conţine canale rezinifere.
a. genul Taxus Genul nu conţine decât o singură specie colectivă – Taxus baccata L. – cu o largă
răspândire în Emisfera nordică. Diferite specii descrise în literatura de specialitate ca fiind cu areal european, asiatic sau din America de Nord trebuie considerate ca şi rase geografice apropiate unele de altele.
Tisa europeană are un areal foarte fragmentat, fiind întâlnită de la Oceanul Atlantic până la sud de Marea Caspică, în bazinul Mediteranei, centrul Europei şi până în nordul Africii.
În România apare diseminat de la coline până în zona montană, în făgete sau amestecuri de fag cu răşinoase; la Cazane coboară la 90 m altitudine.
Ea apare în zone cu relief accidentat, pe stâncării, grohotişuri sub formă de exemplare izolate sau buchete.
În trecut avea o răspândire largă, dar datorită lemnului preţios şi frunzelor otrăvitoare (taxina) tisa a fost exterminată, astfel încât actualmente este declarată specie monument al naturii.
În Asia, trei forme principale au fost descrise: - Taxus cuspidata – Japonia, Coreea - Taxus chinensis – China - Taxus wallichiana – Himalaya
În America de Nord, patru specii principale au fost distinse: - Taxus brevifolia – coasta montană a Pacificului - Taxus canadensis – coasta montană a Atlanticului - Taxus floridana – Florida (endemism) - Taxus globosa – Mexic
47

Diversitatea ecologică a speciilor forestiere
48
b. genul Amenotaxus Genul este endemic, caracteristic Extremului Orient, conţine o singură specie, Amenotaxus
argotaenia, din China şi Taiwan. c. genul Austrotaxus Gen cu o singură specie, Austrotaxus spicata, endemism din Noua Caledonie. d. genul Nothotaxus Şi acest gen conţine tot o singură specie, Nothotaxus chienii, originară din China.

Diversitatea ecologică a speciilor forestiere
CAPITOLUL 5. DIVERSITATEA ECOLOGICĂ A UNOR SPECII DE FOIOASE
CARACTERE GENERALE ALE FOIOASELOR Angiospermele sunt plante mai tinere filogenetic decât gimnospermele, descinzând dinunle
gimnosperme dispărute (ex. Benetitalele). Actualmente, angiospermele lemnoase sunt reprezentate printr-un număr mult mai mare de
specii decât gimnospermele, răspândite în toate zonele climatice al Globului. Sunt plante lemnoase şi ierboase. Arborii au tulpini simpodiale sau monopodiale, ramuri
neverticilate, port foarte divers, mai frecvent coroana fiind ovoidă sau globuloasă. Frunzele sunt căzătoare sau persistente, în formă de limb lăţit sau foaie, ceea ce şi făcut ca speciile respective să fie denumite foioase (excepţie Gingko biloba). Nervaţiunea frunzelor este ramificată.
Florile, foarte diverse în ceea ce priveşte poziţia relativă a elementelor sexuate, sunt frecvent unisexuat monoice, dar şi dioice, bisexuate, poligame. Caracteristic este faptul că prezintă învelişuri florale simple sau duble, deşi la o serie de unităţi sistematice acestea sunt mult mai reduse, avortează imediat după fecundare sau lipsesc complet (familia Betulaceae, Salicaceae etc.). Florile extrem de variate ca structură, formă, culoare, dimensiuni, stau grupate în inflorescenţe diverse sau izolate.
Caracteristic pentru foioase (angiosperme) este faptul că florile femele au marginile carpelelor concrescute alcătuind o cavitate ovariană în care se formează şi stau închise ovulele. Ovarul este prevăzut întotdeauna cu un stigmat care primeşte polenul, iar seminţele ce se dezvoltă din ovule rămân închise (acoperite) în fruct (gr. aggeion=înveliş, iar spermum=sămânţă).
Fructele, de asemenea, extrem de diversificate, cărnoase adeseori, dar frecvent şi uscate, dehiscente sau indehiscente la maturitate etc. Embrionul, de obicei, are un cotiledon sau două cotiledoane.
Arealul foioaselor atinge cotele cele mai ample, fiind dezvoltat deopotrivă în zonele reci şi temperate ale celor două emisfere, ca şi în zonele ecuatoriale, tropicale şi subtropicale.
Angiospermele se împart în două clase, în funcţie de numărul cotiledoanelor embrionului, şi anume: dicotiledonate şi monocotiledonate.
FAMILIA BETULACEAE S.F.Gray
Cuprinde peste 100 specii de arbori şi arbuşti din regiunile temperate şi reci din emisfera nordică, mai rar din cea sudică şi regiunile tropicale.
Morfologic, speciile acestei familii prezintă frunze caduce, simple, alterne, dinţate sau slab lobate, flori monoice, din care cele mascule sunt grupate în amenţi ce se formează în toamnă, iar cele femele amenţi sau fascicule. Fructul este achenă sau samară, cu maturaţie anuală, iar germinaţia este epigee.
Speciile din această familie se grupează în două triburi: • tribul Coryleae Meisn., cu flori ♂ce au trei stamine concrescute cu o bractee şi flori ♀ce au
perigon (Carpinus, Corylus, Ostrya); • tribul Betuleae Döll, cu flori ♂ce au perigon format din 2-4 lacinii, flori ♀lipsite de perigon,
involucru fructifer alipit de fruct provenit din concreşterea bracteei cu bracteolele (Alnus, Betula).
49

Diversitatea ecologică a speciilor forestiere
I. Genul Carpinus L. Cuprinde 18 specii de arbori şi arbuşti din regiunile temperate şi subtropicale din Europa,
China, Japonia, America de Nord. Aceştia prezintă scoarţa netedă, cu ritidom doar la bătrâneţe, muguri alterni, frunze ovat-eliptice, serate şi flori monoice grupate în amenţi.
Fructul este o achenă muchiată, la vârf cu urme de perigon şi stile persistente ce stă la baza unui involucru fructifer generat de unirea a două bracteole cu o bractee. Germinaţia este epigee.
a. Carpinus betulus – răspândire şi diversitate ecologică Arealul speciei este Europa şi Asia de sud-vest, mergând de la Oceanul Atlantic până în
Crimeea, Caucaz, nordul Mării Negre, iar în est ajunge până la Don, evitând stepele ruseşti şi româneşti; de la sud la nord, carpenul se întinde de la Marea Mediterană şi până dincolo de 55� latitudine nordică, în Danemarca şi sudul Suediei. Centrul răspândirii este Europa centrală.
În România este specie de câmpie şi deal şi participă la constituirea pădurilor de şleau, alături de stejar, gorun, ulm de câmp, jugastru etc.
Optimul altitudinal este în intervalul 100-450 m, dar urcă şi în zona amestecurilor de fag cu răşinoase şi a brădetelor, ajungând până la 1000-1200 m în Carpaţii Meridionali.
Graniţele arealului românesc sunt silvostepa Moldovei şi a Munteniei, aceastea constituind, în acelaşi timp, limita estică a arealului european.
Ca ecotipuri, remarcabile sunt: - ecotipul nordic de Bucovina (climă rece, soluri compacte, grele), - ecotipul sud-vestic din Banat (climat cald, soluri calcaroase), - ecotipul de mare altitudine de pe versanţii sudici ai munţilor Căpăţânii (şisturi) - ecotipul de soluri grele din piemonturile subcarpatice (soluri pseudogleizate).
b. Carpinus orientalis – răspândire şi diversitate ecologică Specia are un areal submediteranean, mai sudic decât al carpenului, din Italia, Peninsula
Balcanică şi până în Asia Mică şi Caucaz. La noi apare spontan la câmpie şi coline în staţiuni calde şi relativ uscate din zona
forestieră sau silvostepă. Apare frecvent în Dobrogea, sudul Banatului, vestul Olteniei, dealurile Buzăului, Câmpia Munteniei, silvostepa sudică a Moldovei; apare la Roşcani-Iaşi în silvostepa nordică a Moldovei, aceasta constituind limita nordică a arealului de la noi.
Formează tufărişuri pe coastele însorite împreună cu Quercus pubescens, Prunus mahaleb, Fraxinus ornus, Cotinus coggygria, Syringa vulgaris, intrând în compoziţia formaţiunilor de silvostepă numite şibliacuri.
Alte specii ale genului răspândite în afara Europei sunt: - Carpinus caroliniana (Carpinus americana) - America de Nord; - Carpinus japonica – Japonia - Carpinus cordata – Japonia - Carpinus faginea – Himalaya - Carpinus viminea – Himalaya II. Genul Corylus L. Genul cuprinde peste 15 specii de arbuşti, mai rar arbori, din regiunile temperate sau reci
ale emisferei nordice. Din punct de vedere morfologic, aceste prezintă frunze ovate sau rotund-ovate, pubescente, dublu serate, flori unisexuat monoice, din care cele mascule sunt grupate în amenţi formaţi din anul precedent, iar cele femele aşezate mai multe în mugurele florifer, iar primăvara devreme apar dintre catafile doar stilele roşii. Fructele sunt achene înconjurate de un involucru fructifer format din concreşterea bracteolelor şi au margini dinţate sau laciniate.
50

Diversitatea ecologică a speciilor forestiere
a. Corylus avellana Alunul este răspândit în toată Europa, lipsind în părţile nord-estice; este mult întâlnit în
pădurile de stejar din Crimeea, Caucaz şi Asia Mică. În România este considerat un însoţitor frecvent al stejarului şi gorunului, la câmpie şi
deal, constituind un element important al subarboretului. b. Corylus colurna Alunul turcesc creşte spontan dinn Peninsula Balcanică, Asia Mică şi Caucaz, până în
Himalaia. În România apare în vestul Olteniei şi sudul Banatului, pe coastele însorite, calcaroase.
Alte specii ale genului Corylus răspândite în alte zone ale Globului: - Corylus americana – America de Nord - Corylus heterophylla – Mongolia - Corylus maxima – Balcani, Asia Mică - Corylus sieboldiana – Japonia
III. Genul Betula L. Cuprinde circa 90 specii de arbori şi arbuşti şi subarbuşti din emisfera nordică, ce
vegetează până spre tundră, la limita vegetaţiei lemnoase. Scoarţa este cu periderm suberos alb cretaceu sau gălbui, exfoliabil în fâşii circulare.
Florile sunt unisexuat monoice dispuse în amenţi: cele femele după fecundare se transformă într-un fel de con de fructificaţie alungit care poartă la subsuoara bracteolelor trifidate câte trei samare mici biaripate; florile mascule sunt grupate în amenţi ce se formează din vara precedentă, rar primăvara odată cu cele femele.
În acest gen intră specii nepretenţioase faţă de climă şi sol, intrând în categoria speciilor pioniere (primă împădurire naturală).
a. Betula pendula – răspândire şi diversitate ecologică Mesteacănul are un areal larg, comparabil cu al plopului tremurător, în Europa de la
Atlantic până în Siberia şi Altai, iar în nord urcă până dincolo de Cercul Polar; în sud coboară până în Apenini şi Balcani, iar spre est ajunge până în stepa rusească.
În România apare frecvent în parchete tăiate ras, locuri deschise, pe stâncării, din zona colinară până în zona montană superioară. La munte poate depăşi 1500 m altitudine, iar la câmpie poate coborî sub 250 m (Filiaşi, Piscu-Tunari, Hanu Conachi, Ciurea-Iaşi).
Optimul de vegetaţie este în pădurile de deal, făgetele montane, amestecurile fag-răşinoase mai ales în Carpaţii Orientali.
Fiind o specie cu un areal atât de larg, mesteacănul este reprezentat prin intermediul a câtorva rase geografice: cele din ţinuturile siberiene, nordice, atlantice, baltice, din Apenini, Balcani, sudul Rusiei. Acestea se pot identifica din punct de vedere ecologic prin intermediul ecotipurilor climatice: • mesteacănul de câmpie, • mesteacănul de mare altitudine. Ca ecotipuri edafice, trebuie de precizat:
- mesteacănul de soluri relativ uscate - mesteacănul de turbărie.
b. Betula pubescens Specia este destul de larg răspândită, din Europa până în părţile estice ale Siberiei,
mergând chiar până la limita polară a vegetaţiei forestiere. În România, mesteacănul pufos aparţine subspeciei carpatica (lujeri tineri glabri, rar
pubescenţi, frunze mai mici), fiind un endemism carpatic.
51

Diversitatea ecologică a speciilor forestiere
c. Betula nana Mesteacănul pitic este răspândit în nordul Eurasiei, Groenlanda, Alaska, în tundra arctică şi
regiunile montane. În România este limita sudică a arealului mondial al speciei, fiind semnalată în două turbării: la Luci, în judeţul Harghita şi la Lucina, în judeţul Suceava.
d. Betula humilis Această specie este un element boreal, circumpolar, răspândit în Alpi, nord-estul Europei,
trecând dincolo de Urali, până în Altai. Apare în câteva staţiuni şi în România.
Alte specii ale genului Betula cu un areal remarcabil sunt: - Betula populifolia - Betula davurica - Betula papyrifera – America de Nord - Betula platyphylla - Manciuria - Betula pumila - Betula fruticosa - Betula maximowicziana - Japonia - Betula lutea – America de Nord - Betula nigra - Betula lenta - Betula albo-sinensis – nordul Chinei - Betula ermani - Betula costata – nord-vestul Chinei
IV. Genul Alnus Miller Cuprinde circa 40 specii de arbori şi arbuşti din emisfera nordică, câteva din ele ajung în
America de Sud, iar unele specii se ridică dincolo de limita superioară a vegetaţiei lemnoase, în zona alpină.
Morfologic, se individualizează prin nodozităţile de pe rădăcinile tinere, muguri alterni, obişnuit pedicelaţi, frunze simple, dublu serate, flori monoice grupate în amenţi, cei masculi pendenţi, grupaţi în fascicule, iar cei femeli mai scurţi, grupaţi în raceme, ambele categorii formându-se în anul precedent înfloririi, mai rar în primăvară; speciile genului înfloresc înaintea înfrunzirii, cu unele excepţii.
După fecundaţie amentul femel se transformă într-un conuleţ numit “rânză” ce rămâne pe arbore mai mult timp după diseminaţie, iar fructele sunt samare.
Din punct de vedere ecologic au exigenţe reduse faţă de sol, preferând pe cele cu umiditate ridicată, motiv pentru care majoritatea cresc în lunci, pe lângă ape. Sunt specii rezistente la ger şi pretenţioase faţă de lumină.
a. Alnus glutinosa Arealul aninului negru este larg, exclusiv european, lipsind din regiunile septentrionale
(dincolo de 63-64° latitudine nordică) şi din stepele de la nord de Marea Neagră şi Marea Caspică. În România apare pe văile râurilor din zona de câmpie şi de dealuri. Sporadic, urcă în zona montană a făgetelor (800-900 m) sub formă de pâlcuri, iar ca indivizi izolaţi urcă până la 1300 m în Munţii Harghitei. În Lunca Dunării apare rar, iar în Deltă se găseşte pe grindurile C.A.Rosetti şi Ivancea.
b. Alnus incana Aninul alb are un areal vast, în Europa depăşeşte mult graniţa septentrională a aninului
negru, ajungând dincolo de Cercul Polar; mai apare şi în Asia estică şi America de Nord. Ocupă regiuni mai reci – altitudinal şi latitudinal – decât aninul negru. În România preia locul aninului negru la altitudini mai mari, frecvent în luncile montane; urcă în Carpaţii Orientali la
52

Diversitatea ecologică a speciilor forestiere
53
1300 m (aninişuri pure), dar coboară şi în zona de dealuri (subzona stejarului). Lipseşte în Banat. Aninul alb apare la poalele versanţilor orientali şi meridionali ai Carpaţilor, constituind limita sud-estică a arealului său european.
c. Alnus viridis Aninul verde creşte spontan în Alpi şi Carpaţi, la altitudini mari, la limita superioară a
pădurilor, prin rarişti şi golul alpin. Ajunge la limita latitudinală a pădurilor din nordul Europei, Siberia, Groenlanda şi America de Nord. În România apare frecvent în etajul subalpin şi alpin asociat cu Pinus mugo.
Alte specii ale genului Alnus sunt: - Alnus hirsuta - Alnus japonica - Japonia - Alnus rugosa - Alnus serrulata
- Alnus cordata – Italia - Alnus rubra – America de Nord - Alnus subcordata – Caucaz
FAMILIA FAGACEAE Dumort.
Familia cuprinde numeroase specii de arbori de talie mare, rar arbuşti, originari din regiunile temperate şi călduroase ale emisferei nordice; excepţie fac câteva specii de Quercus care în Malaesia depăşesc Ecuatorul şi genul Nothofagus specific emisferei australe (America de Sud, Australia, Noua Zeelandă).
Morfologic, aceste specii se caracterizează prin muguri cu solzi imbricaţi, frunze simple, întregi, dinţate sau adânc lobate, caduce sau persistente, flori unisexuat monoice, cu perigon alcătuit din 4-7 lobi, din care cele mascule sunt grupate în amenţi sau capitule cu patru sau mai multe stamine, iar cele femele sunt înconjurate la bază de un involucru format din mai multe bractei, grupate câte 1-3 pe un peduncul comun.
Fructele sunt achene aflate total sau parţial într-o o cupă lemnoasă ce a luat naştere din involucrul bracteal; maturaţia este anuală sau bienală.
Genurile tratate în prezentul curs sunt Fagus, Castanea şi Quercus.
I. Genul Fagus L. Cuprinde circa 10 specii de arbori din zona temperată a emisferei nordice, răspândite în
Europa, Asia Mică, Extremul Orient şi în zonele estice ale Americii de Nord. Înrădăcinarea este pivotant-trasantă, scoarţa netedă fără ritidom, mugurii alterni fuziformi,
frunzele caduce, întregi, iar florile unisexuat monoice, din care cele mascule sunt grupate în capitule la vârful unui peduncul lung, pendent, fiecare floare cu un perigon cu 4-7 lobi şi câte 8-10 stamine, iar cele femele câte două, înconjurate de numeroase bractei unite la bază care formează un involucru cu 4 diviziuni, şi stau pe un peduncul scurt.
Fructul este achenă (popular numită jir) triunghiulară, obişnuit câte două închise complet într-o cupă prevăzută la exterior cu prelungiri subulate, lemnoase sau foliacei.
a. Fagus sylvatica – răspândire şi diversitate ecologică Fagul este specie europeană, fiind răspândită în vestul, centrul şi sudul continentului: de la
Atlantic până în nordul Moldovei, din Pirinei, ţărmul Mării Mediterane şi Grecia, până în sudul Scoţiei şi al Scandinaviei.
În Europa de nord-vest devine arbore de câmpie şi dealuri, iar în sud apare la peste 1500 m (Alpi - 1650 m, Pirinei - 2000 m, Etna - 2160 m).
În România fagul ocupă circa 2 milioane hectare (32%), fiind întâlnit de la 300-500 m la 1200-1400 m, frecvent în zona deluroasă şi montană; pe văi umede coboară la 150-200 m, pe valea Cernei şi a Dunării la 60-100 m.
În vastul areal formează arborete pure pe mari suprafeţe sau amestecuri cu gorunul, carpenul, bradul, molidul.

Diversitatea ecologică a speciilor forestiere
Insular fagul apare în nordul Dobrogei la Luncaviţa, în Câmpia Olteniei (Bucovăţ-Craiova, Stârmina-Drobeta Turnu Severin) şi în Câmpia Munteniei, la Snagov.
În Carpaţii Meridionali întâlnim fagul în Retezat, Vâlcan, Parâng, iar în Munţii Apuseni, limita superioară a făgetelor depăşeşte 1400 m (1650m) în Munţii Bihor, Semenic, Cernei, Parâng, Vâlcan, Sebeş etc. • Rase geografice: - F. s. britanica (Marea Britanie),
- F. s. scandinavica (sudul Scandinaviei), - F. s. borealis (litoralul Mării Baltice), - F. s. celtica (nordul Franţei), - F. s. pomeranica (Danemarca); • Climatipuri - F. s. pyrenaica (Pirinei),
- F. s. gallica (Jura), - F. s. alpina (Alpii Vestici), - F. s. austriaca (Alpii Austriei), - F. s. apennina (Apenini), - F. s. hercynica (Europa Centrală - zona montană şi colinară), - F. s. carpatica (Carpaţii Slovaciei), - F. s. silesiaca (Polonia), - F. s. polonica (Polonia), - F. s. transsilvanica (Carpaţii României), - F. s. balcanica (Balcani), - F. s. podolica (Podişul Podolic, Central Moldovenesc);
• Ecotipuri climatice şi edafice: - fagul de Bucovina (de climat rece), - fagul de Banat (climatip termofil), - fagul din Munţii Apuseni (climatip termofil), - fagul dobrogean (climat cald şi uscat), - fagul de mare altitudine (Vâlcan, Parâng, Godeanu), - fagul de mică altitudine (Valea Dunării, Snagov).
b. Fagus orientalis Fagul oriental este răspândit în sud-estul Peninsulei Balcanice, nordul Asiei Mici, Crimeea,
Caucaz. În România, această specie a fost mult timp confundată cu subspecia moesiaca, arealul şi exigenţele ecologice nefiind încă pe deplin clarificate.
Alte specii ale genului Fagus sunt:
- Fagus engleriana – China - Fagus grandifolia – America de Nord - Fagus mexicana – Mexic - Fagus crenata – Japonia - Fagus japonica – Japonia
II. Genul Nothofagus Bl. Gen ce conţine circa 20 specii asemănătoare genului Fagus, nu foarte bine studiate, care
sunt răspândite în emisfera australă (Australia, Noua Zeelandă, America de Sud, Noua Guinee, Noua Caledonie). Ce mai cunoscută şi cea mai răspândită specie Nothofagus antarctica.
III. Genul Castanea Mill. Cuprinde peste 10 specii de arbori răspândite în Europa Meridională, Asia Mică, Algeria,
Asia Orientală şi sud-estul Americii de Nord. Sunt specii termofile, cu frunze lanceolate şi
54

Diversitatea ecologică a speciilor forestiere
55
ascuţit dinţate, cu flori unisexuat monoice, cele mascule cu caliciu campanulat cu 6 diviziuni şi 10-12 stamine, grupate în amenţi lungi, erecţi, iar cele femele câte 1-3, înconjurate de un involucru spinos, aşezate la baza amenţilor masculi.
Fructul este o achenă (castana), închise câte 1-3 într-o cupă sferică prevăzută cu spini lungi înţepători; castana este comestibilă, se maturează în toamnă, iar germinaţia este hipogee.
a. Castanea sativa Castanul bun are origine mediteraneană, fiind răspândit în Europa, din Spania până în
Peninsula Balcanică, trece în Asia Mică, Caucaz şi nord-vestul Africii (Maroc, Algeria, Tunisia).
În România, el vegetează în regiunile cu climat blând: depresiunea subcarpatică a Olteniei (Tismana, Polovragi, Baia de Aramă etc.) şi nord-vestul ţării (Baia Mare, Baia Sprie, etc.); cultura castanului a fost extinsă în Transilvania, Oltenia.
Alte specii ale genului Castanea sunt: - Castanea dentata – America de Nord - Castanea pumila - Mexic - Castanea crenata – Japonia
IV. Genul Quercus L. Genul cuprinde peste 200 specii, în majoritate arbori, rar arbuşti, cu areal vast în regiunile
temperate şi subtropicale ale emisferei boreale; numărul stejarilor europeni este redus în comparaţie cu cel al stejarilor din Asia estică şi, mai ales, din America de Nord (ex. în Mexic vegetează peste 80 specii).
Morfologic acestea se caracterizează prin frunze alterne, caduce, marcescente sau persistente, lobate, dinţate rar întregi, flori unisexuat monoice, cele mascule au perigonul divizat în 4-7 lobi şi 4-12 stamine, grupate în amenţi lungi, subţiri, pendenţi, iar cele femele au perigonul slab dinţat, solitare sau câte 2-8 într-o inflorescenţă spiciformă. Fructul este o achenă (ghinda) cilindrică, elipsoidală sau semisferică, susţinută într-o cupă cu numeroşi solzi imbricaţi, liberi sau concrescuţi, cu maturaţie anuală sau bienală şi germinaţie hipogee. Au un temperament de lumină.
La noi în ţară, speciile de stejari ocupă 19% din fondul forestier. Prezintă fenomene de hibridogeneză, fiind semnalate forme intermediare hibride.
a. Quercus rubra Specie din America de Nord, unde vegetează în sud-estul Canadei, formând arborete pure
sau amestecuri cu Pinus strobus şi Juglans nigra. Alte specii de Quercus răspândite în America de Nord sunt:
- Quercus nigra - Quercus phellos - Quercus imbricaria - Quercus acuttissima - Quercus marilandica
- Quercus velutina - Quercus coccinea - Quercus palustris - Quercus falcata
b. Quercus cerris – răspândire şi diversitate ecologică Cerul este răspândit în Peninsula Iberică şi Franţa, de la Oceanul Atlantic până în Asia
Mică, fără a se depărta de ţărmul Mării Mediterane prea mult; în interiorul continentului pătrunde mai adânc doar la nord de Dunăre. În România, el apare în zona de silvostepă din Muntenia, Oltenia, urcând în zona colinară la 500-600 m, iar în vestul Transilvaniei şi Banat devine frecvent. În munţii Apuseni, urcă la 900-1000 m. Izolat, apare în sudul Dobrogei (Canaraua Fetii), Moldova (Rogojeni-Galaţi) şi Podişul Transilvaniei.

Diversitatea ecologică a speciilor forestiere
c. Quercus petraea – răspândire şi diversitate ecologică Gorunul este răspândit din nord-vestul Europei (Marea Britanie, sudul Scandinaviei) până
la Marea Mediterană şi de la Oceanul Atlantic până în vestul Ucrainei, la gurile Donului şi Nistrului, ajunge la Marea Caspică, nordul Crimeei, Caucaz, trecând şi în Asia Mică.
În România, gorunul urcă mai mult în altitudine decât stejarul, zona lui de optim ecologic fiind în regiunea colinară. Apare frecvent în Carpaţii Orientali, la 400-500 m, în Carpaţii Meridionali la 700-800 (900) m, iar în Apuseni la 600-700 m. Ca arborete rărite urcă la 950-1000 m în Carpaţii Meridionali, pe versanţii transilvăneni. Pe versanţii sudici ai Muntelui Cozia (gneisuri) urcă la 1300 m, izolat apare în sudul Dobrogei şi Moldova, iar în nordul Dobrogei apare pe suprafeţe întinse.
Apare în arborete pure, în amestec cu fagul sau în şleaurile de deal. • Rase geografice: - gorunul de Marea Britanie,
- gorunul de Scandinavia, - gorunul din vestul Europei, - gorunul din ţinuturile baltice, - gorunul balcanic, - gorunul caucazian, - gorunul din Iugoslavia.
• Ecotipuri autohtone: - submezoterm de mare altitudine (Cozia), - ecotipul relativ xerofit (sudul Dobrogei şi Moldovei), - ecotipul terofil din Banat (vestul Apusenilor), - ecotipul de terase şi piemonturi, - ecotipul de soluri rendzinice, stâncării, - ecotipul de soluri podzolice.
d. Quercus robur Stejarul are un areal vast, fiind prezent din regiunile mediteraneene până în nordul Europei,
ajunge în vest în Marea Britanie, avansează în est până în Urali, Crimeea şi Caucaz, până pe ţărmul Mării Caspice. În România ocupă în prezent circa 130 000 ha (2%), alcătuind arborete pure sau amestecuri în regiunea de câmpie şi dealuri joase. În zona dealurilor joase apare frecvent pe terase, platforme şi piemonturi; cele mai întinse suprafeţe sunt în sudul ţării (Piteşti-Alexandria-Comana-Mizil), în câmpia vestică, în nord-estul Moldovei şi în centrul Transilvaniei. În treimea superioară a versanţilor, dealurilor sau spre etajul montan inferior se ridică rar şi numai în staţiunile însorite, calde, suficient de umede (luncile largi). • Rase geografice: - rasa atlantică (adaptată la climat umed şi cald),
- continentală din Rusia, - rasa baltică, - rasa sud-scandinavă (climat rece şi umed), - rasele termofile (sudul Europei, Caucaz, Crimeea);
• Provenienţe valoroase: - stejarul de Spessart (centrul Europei), - stejarul de Slovenia (sud-estul Europei);
• Ecotipuri autohtone: - stejarul de luncă (soluri aluvionare), - stejarul de terasă (substrat argilos, soluri pseudogleizate) - stejarul de lăcovişti (soluri hidromorfe) - stejarul de silvostepă, - stejarul de soluri sărăturoase, alcaline (limita de suportabilitate).
e. Quercus pedunculiflora Stejarul brumăriu are un areal limitat la Peninsula Balcanică, nordul Asiei Mici, Caucaz,
Crimeea. În România, apare în silvostepa Olteniei, Munteniei, Dobrogei şi sudul Moldovei.
56

Diversitatea ecologică a speciilor forestiere
57
f. Quercus frainetto Gârniţa are un areal restrâns, inclus în cel al cerului; specia apare din sudul Italiei, în
Peninsula Balcanică, până în Ungaria, România; spre est avansează puţin, spre nord-vest ajunge în Asia Mică şi Turcia. În România, apare pe arii extinse în Muntenia pe platforma Cotmeana, în silvostepa Olteniei şi Munteniei până în regiunea dealurilor subcarpatice la altitudini de 450 m, unde este în amestec cu cerul (cereto-gârniţete). În vestul ţării (Banat, Transilvania), apare la dealuri, urcând pe alocuri la 500-550 m, iar în Semenic la 660 m.
g. Quercus pubescens Stejarul pufos este un element sud-european, mediteranean şi submediteranean, cu areal
asemănător cu cel al cerului; spre nord urcă până la 50º latitudine, la est merge până la Marea Caspică, iar la sud evită o parte din ţinuturile mediteraneene cuprinse în arealul cerului (sudul Italiei, Sicilia). În România formează păduri rare, pâlcuri sau tufărişuri în regiunea de câmpie şi de silvostepă, în aproape toate regiunile ţării, mai frecvent în silvostepa sudică a Moldovei, Munteniei şi Olteniei; în Dobrogea participă la alcătuirea şibliacurilor, alături de Carpinus orientalis, Fraxinus ornus, Prunus spinosa, Prunus mahaleb, Cotinus coggygria etc.
h. Quercus virgiliana Specie de origine mediteraneană, cu un areal restrâns în Italia şi Peninsula Balcanică. Alte specii ale genului Quercus, specii ce îşi află arealul în Asia, Europa Orientală sau
America de Sud sunt:
- Quercus acutissima - Quercus libani - Quercus trojana - Quercus pontica - Quercus castaneifolia - Quercus lyrata - Quercus stellata - Quercus turneri - Quercus meuhlenbergii - Quercus bicolor
- Quercus michauxii - Quercus prinus - Quercus macranthera - Quercus dentata - Quercus macrocarpa - Quercus pyrenaica - Quercus alba - Quercus schochiana - Quercus ilicifolia - Quercus ilex
FAMILIA SALICACEAE Lindl.
Cuprinde arbori şi arbuşti răspândiţi pe tot globul în cele mai variate condiţii staţionale.
Din punct de vedere morfologic se caracterizează prin frunze simple, alterne, flori unisexuat dioice grupate în amenţi, nude, prevăzute cu câte o bractee (scvamă), cu două sau mai multe stamine şi ovar unilocular cu 2 stigmate. Fructul este o capsulă dehiscentă în 2-4 valve, iar seminţele au smocuri de peri lungi argintii (egretate); fructifică de timpuriu, anual şi abundent.
I. Genul Populus L. Cuprinde circa 25 specii de arbori, în majoritate originari din zonele reci şi temperate ale
emisferei nordice. Lemnul acestora este moale şi putrezeşte repede, lujerii au măduva pentagonal stelată, muguri alterni cu 3-5 solzi şi frunze simple. Florile sunt dioice grupate în amenţi: cele mascule cu numeroase stamine cu anterele roşii, iar cele femele cu 2-4 stigmate. Capsula se desface în 2-4 valve, seminţele au smocuri de peri – egretate (“vată de plopi”); maturaţia are loc vara devreme, diseminaţia prin mai-iunie, iar germinaţia are loc la câteva ore sau zile de la împrăştiere. Puterea germinativă este mare, dar de scurtă durată.

Diversitatea ecologică a speciilor forestiere
a. Populus alba Plopul alb este răspândit în centrul şi sudul Europei, de la Marea Mediterană la 55°
latitudine nordică, trece şi în Africa de Nord, iar spre est ajunge în Asia Centrală şi Siberia. În România, apare în regiunea de câmpie şi dealuri joase, obişnuit în luncile râurilor; în lunca şi Delta Dunării formează arborete pure sau amestecuri împreună cu plopul negru, sălcii şi aninul negru (zăvoaie). Altitudinal specia urcă până la 800 m.
b. Populus tremula Plopul tremurător are un areal larg, în Europa şi Asia, trecând dincolo de 70° latitudine
nordică, iar spre sud ajunge în nord-vestul Africii. În România, apare frecvent în zona montană, în molidişuri la 1600 m altitudine, dar coboară la dealuri şi câmpie.
c. Populus nigra Plopul negru are un areal vast, asemănător cu cel al plopului alb, în Siberia ajungând la 64°
latitudine nordică, adesea fiind înlocuit cu culturi de plopi hibrizi, mult mai productivi. În România, apare în luncile şi depresiunile umede de la câmpie şi coline, pe firul văilor.
Alte specii ale genului Populus răspândite în America de Nord şi Asia sunt:
- Populus tremuloides - Populus lasiocarpa - Populus wilsonii - Populus angustifolia - Populus simonii - Populus balsamifera - Populus szechuanica - Populus trichocarpa - Populus tristis
- Populus laurifolia - Populus berolinensis - Populus maximowiczii - Populus koreana - Populus suaveolens - Populus angulata - Populus deltoides - Populus × canadensis
II. Genul Salix L. Cuprinde circa 300 specii de arbori, arbuşti şi subarbuşti târâtori sau pitici, frecvente pe tot
Globul, în special în climate de tip boreal; avansează mai spre nord şi mai sus altitudinal decât plopii (tundră, ţinuturi alpine).
Lujerii sunt netezi, flexibili, mugurii alterni, cu câte un solz provenit din concreşterea celor doi, acoperind partea internă, păroasă, asemănător unei scufii. Frunzele sunt simple, scurt peţiolate, adesea sesile, întregi sau dinţate, cu stipele persistente. Flori unisexuat dioice, grupate în amenţi erecţi, fiecare floare are la bază o scvamă cu marginea întreagă şi 1-2 glande nectarifere. Fructul este o capsulă bivalvă, ce se maturează prin mai-iunie, iar seminţele sunt prevăzute cu smocuri de peri lungi, albi (egretate) – “vata de salcie”.
a. Salix alba Salcia albă are un areal vast, Europa (fără ţinuturile nordice extreme), ajungând în Caucaz,
Asia Mică, China, Himalaia, Algeria. În România, în zona de câmpie formează arborete pure (sălcete), în prima fază numite “renişuri” sau amestecuri cu plopii (zăvoaie), atât în Lunca Dunării, cât şi pe principalii afluenţi. Urcă în lungul văilor la deal, iar ca exemplare izolate ajunge în regiunea montană inferioară.
b. Salix caprea Salcia căprească are un areal vast, Europa şi Asia, până la 70° latitudine nordică, altitudinal
ajungând până spre limita superioară a vegetaţiei forestiere. În România, apare frecvent în zona colinară, comună în zona montană, rar coboară la câmpie; în etajul molidişurilor, la altitudini mari este înlocuită de Salix silesiaca. Apare frecvent în parchete tăiate ras, în rarişti, poieni unde caracter invadant, alături de Populus tremula şi Betula pendula: SPECIE PIONIERĂ.
58

Diversitatea ecologică a speciilor forestiere
c. Salix cinerea Specia are un areal vast, fiind semnalată în Europa, Asia, Africa de Nord. În România,
creşte în luncile râurilor, prin zăvoaie, locuri mlăştinoase, de la câmpie şi până la munte.
Alte specii ale genului Salix, multe dintre ele indigene, sunt: - Salix purpurea - Salix viminalis - Salix triandra - Salix pentandra - Salix rosmarinifolia - Salix elaeagnos - Salix fragilis - Salix daphnoides - Salix starkeana - Salix aurita - Salix phylicifolia - Salix nigra - Salix lasiandra
- Salix lucida - Salix magnifica - Salix interior - Salix matsudana - Salix babylonica - Salix bockii - Salix elegantissima - Salix aegiptiaca - Salix rigida - Salix caesia - Salix glabra - Salix rehderiana - Salix appendiculata
- Salix nigricans - Salix formosa - Salix myrsinites - Salix hastata - Salix lapponum - Salix helvetica - Salix cordata - Salix discolor - Salix sericea - Salix petiolaris - Salix dasyclados - Salix repens - Salix acutifolia
La acestea, se adaugă o serie de sălcii pitice ce vegetează în zonele de tundră sau în cele alpine:
- Salix myrtilloides - Salix reticulata - Salix retusa
- Salix serpyllifolia - Salix herbacea
FAMILIA ULMACEAE Mirb.
În familia Ulmaceae sunt incluşi arbori şi arbuşti din 3 genuri: Ulmus, Celtis, Zelkowa, ce vegetează în ambele emisfere. Din punct de vedere morfologic acestea se caracterizează prin muguri alterni cu solzi imbricaţi, frunze aşezate distic, cu baza asimetrică, serate, rar întregi, flori mici, hermafrodite, monoice sau poligame, apetale şi fructe de tip samară, achenă sau drupă.
I. Genul Ulmus L. Genul conţine 30 specii de arbori şi arbuşti din zonele subtropicală şi temperată ale
emisferei boreale, Eurasia şi America de Nord, la noi semnalându-se 11 specii (spontane sau cultivate), din care două hibride.
Acestea prezintă lujeri geniculaţi, uneori cu excrescenţe suberoase, frunze căzătoare sau persistente aşezate ± distic, scurt peţiolate, cu baza asimetrică, dublu serate. Florile sunt hermafrodite ce apar primăvara timpuriu înainte de înfrunzire, fiind grupate în fascicule sesile sau scurt pedunculate, mai rar raceme.
Fructul este o samară turtită, orbiculară, eliptică sau ovată, înconjurată de o aripioară membranoasă crestată la vârf şi cu resturi de caliciu la bază; coacerea şi diseminarea are loc la câteva săptămâni după înflorire (mai-iunie la speciile indigene).
Sunt specii exigente faţă de fertilitatea solului, rar constituind arborete pure pe suprafeţe mici, obişnuit fiind specii diseminate în pădurile de amestec.
a. Ulmus minor Areal general este majoritar european: la vest ajunge la Oceanul Atlantic, la est se întinde
până la Marea Caspică, la nord ajunge la 55° latitudine, până în Suedia (fără Marea Britanie), iar la sud trece în Africa de Nord şi Asia (circa 30° latitudine nordică). În România, specia apare din silvostepă şi până în zona forestieră, fiind frecventă în pădurile de câmpie şi dealuri joase. El este specie de amestec, fiind în cvasitotalitatea cazurilor, specie diseminată în pădurile de luncă şi şleauri.
59

Diversitatea ecologică a speciilor forestiere
b. Ulmus glabra Arealul general este mai extins spre nord şi nord-vest faţă de cel al speciei precedente, în
schimb în sud evită ţinuturile mediteraneene (sud Spania, Corsica, Sardinia, Sicilia). La noi, specia apare din zona colinară şi până în regiunea montană (făgete montane, amestecuri), poate apare diseminat sau în pâlcuri până la 1100-1300 m.
c. Ulmus laevis Arealul general este relativ extins, având ca limite: estul Franţei, nordul Italiei, sudul
Peninsulei Balcanice şi munţii Urali; a fost semnalat şi în Siberia (regiunea Tobolsk). În România este mai puţin răspândit faţă de ceilalţi ulmi, fiind sporadic în zona de câmpie şi cea colinară (Oltenia, Banat), legat mai mult de lunci şi zăvoaie.
d. Ulmus pumila Specia vegetează în Siberia Orientală, Mongolia, Asia Centrală, nordul Chinei.
Alte specii ale genului Ulmus sunt: - Ulmus americana - Ulmus rubra - Ulmus carpinifolia
II. Genul Celtis L. Genul cuprinde specii de arbori şi arbuşti, cu ramificaţiile şi frunzele dispuse distic, lujerii
cu măduvă lamelar întreruptă, frunze la bază cu 3 nervuri. Florile sunt poligame (hermafrodite şi mascule), cele mascule find grupate în fascicule, iar cele bisexuate câte una în axila frunzelor. Fructele sunt drupe globuloase cu mezocarp subţire şi sâmbure tare (endocarp sclerificat).
a. Celtis australis Arealul general al speciei este bazinul Mării Mediterane, Caucaz, Asia de Vest. În
România, apare sporadic în Oltenia, Banat şi Dobrogea, pe stâncării însorite, la marginea pădurilor etc.
b. Celtis glabrata Arealul general al speciei este pontic (Crimeea, Caucaz), în România fiind prezentă doar în
Dobrogea: munţii Măcinului, stâncăriile calcaroase de la Hârşova, Gura Dobrogii, Cheia.
În afară de speciile sus amintite mai sunt de precizat specia nord-americană Celtis occidentalis şi Celtis tournefortii.
FAMILIA ROSACEAE Juss.
Familia include numeroase specii de arbori, arbuşti şi plante erbacee, cu flori hermafrodite, pe tip 5. Tipul de fruct este divers: foliculă, achenă, drupă, drupă falsă, acesta fiind criteriul de scindare a familiei în patru subfamilii: Spiraeoideae, Rosoideae, Pomoideae, Prunoideae.
Pentru exemplificare, cel mai cunoscut gen este genul Prunus. În sprijinul acestei idei, putem aduce în discuţie genul Rubus care, numai în flora spontană a ţării noastre, prezintă 325 de specii, astfel încât nu mai poate fi vorba despre vicarianţă sau de specii similare, ci despre un studiu morfo-ecologic aprofundat.
Genul Prunus a. Prunus spinosa Specia are un areal destul de vast, fiind prezent în Europa, Asia Mică, Africa de Nord. În
România, apare frecvent în liziere, rarişti, pe coaste însorite, stâncării, de la câmpie şi până în etajul montan inferior (1000 m); constituie adesea subarboretul stejăretelor xerotermofile (Dobrogea, Moldova).
60

Diversitatea ecologică a speciilor forestiere
61
b. Prunus cerasifera Arealul general al speciei este în Europa de sud-est, Asia de Vest, Caucaz.
c. Prunus avium Specia apare de la Oceanul Atlantic până în Siberia, în Asia Mică şi chiar în Africa de
Nord. În România, este diseminat în pădurile de foioase, de la câmpie şi până în zona montană, pe versanţii însoriţi urcând până la 1000 m.
d. Prunus mahaleb Arealul general al speciei este localizat în Europa Centrală şi Sudică, Asia de Vest, la noi
găsindu-se în păduri rărite din silvostepa Dobrogei (şibliacuri), Moldova de Sud, Banat.
e. Prunus padus Specia apare din Europa şi până în Siberia.
Alte specii ale genului Prunus sunt:
- Prunus serotina - Prunus tenella - Prunus laurocerassus - Prunus lusitanica - Prunus maackii - Prunus virginiana - Prunus incisa - Prunus nipponica - Prunus conradinae - Prunus subhirtella - Prunus sargentii - Prunus yedoensis
- Prunus serrulata - Prunus cerasus - Prunus fruticosa - Prunus serrula - Prunus pennsylvanica - Prunus emarginata - Prunus pumila - Prunus glandulosa - Prunus tenella - Prunus incana - Prunus prostrata - Prunus tomentosa
- Prunus triloba - Prunus argentea - Prunus davidana - Prunus persica - Prunus dulcis - Prunus sibirica - Prunus armeniaca - Prunus mume - Prunus domestica - Prunus nigra - Prunus maritima - Prunus americana
FAMILIA ACERACEAE A. L. Juss. I. Genul Acer L. Genul cuprinde circa 110 specii arbori şi arbuşti, ce vegetează în emisfera nordică. Acestea
prezintă muguri opuşi, cu cicatrici unite printr-o linie stipelară, frunze palmat lobate, penat compuse sau simple, flori poligame, adesea andromonoice (bisexuate şi mascule pe acelaşi exemplar), mai rar dioice sau androdioice, pe tipul 5(4), uneori fără petale şi fructe disamare lung aripate, între aripi cu unghi variabil, dar caracteristic fiecărei specii.
a. Acer pseudoplatanus Arealul general este localizat în Asia de Vest, Europa Centrală şi de Sud, depăşind 55°
latitudine nordică. În România specia se cantonează în regiunea colinară şi montană (până la 1500-1600 m altitudine), diseminat apărând în şleaurile de deal şi molidişuri, izolat în rariştile subalpine, pe grohotişuri montane.
b. Acer platanoides Arealul general al speciei este mai larg decât cel al paltinului de munte: Europa Centrală şi
estică până în munţii Urali, la nord depăşind 60° latitudine nordică în Scandinavia (Finlanda), ajunge la ţărmul Oceanului Atlantic şi al Mării Mediterane şi în Caucaz. La noi, se găseşte diseminat la câmpie şi deal, dar poate apare şi la munte, în făgete şi amestecuri de fag cu răşinoase.
c. Acer campestre Specia apare în toată Europa şi Asia, avansând spre nord-vest până în Anglia, Scandinavia,
mai puţin decât Acer platanoides, iar în sudul arealului dincolo de limita arealului lui Acer pseudoplatanus. La noi este specie comună la câmpie, coline şi silvostepă, fiind o specie de

Diversitatea ecologică a speciilor forestiere
amestec în şleauri, de la marginea silvostepei şi până la limita inferioară a fagului, mai rar în zona montană.
d. Acer tataricum Specia are un areal mai restrâns, în Europa sud-estică, Asia Mică. În România apare la
câmpie şi dealuri, fiind frecvent în subarboretul zonei stejarului (şleauri). Alte specii ale genului Acer, răspândite în special în America de Nord, pe coasta atlantică:
- Acer carpinifolium - Acer negundo - Acer monspessulanum - Acer truncatum - Acer lobelii - Acer cappadocicum - Acer mono - Acer macrophyllum - Acer saccharum - Acer nigrum - Acer opalus - Acer miyabei - Acer ginnala - Acer glabrum - Acer spicatum - Acer velutinum
- Acer heldreichii - Acer trautvetteri - Acer crataegifolium - Acer davidii - Acer capillipes - Acer rufinerve - Acer pensylvanicum - Acer palmatum - Acer sieboldianum - Acer japonicum - Acer circinatum - Acer mandshuricum - Acer griseum - Acer nikoense - Acer cissifolium
FAMILIA TILIACEAE Juss.
I. Genul Tilia L. Genul cuprinde circa 50 de specii din emisfera nordică. Scoarţa este mult timp netedă,
lujerii geniculaţi, mugurii distici, frunzele simple şi florile hermafrodite, cu 5 sepale, 5 petale libere, uneori cu 5-10 staminodii petaloide şi numeroase stamine. Florile sunt grupate în cime pedunculate, iar inflorescenţa este prevăzută cu o bractee liguliformă. Fructul este o achenă globuloasă, cu 1-3 seminţe.
a. Tilia cordata Arealul general al speciei este Europa, de la Marea Mediterană până la 60° latitudine
nordică (Anglia, Scandinavia) şi de la Oceanul Atlantic până în Siberia şi Caucaz. În România apare din regiunea colinară până în etajul montan inferior (900 m altitudine), fiind frecvent şi constant în şleaurile de deal, avansând altitudinal mai mult decât ceilalţi tei, atât la câmpie cât şi la munte; pe alocuri poate forma teişuri pure.
b. Tilia platyphyllos Arealul este limitat la sudul şi centrul Europei, nu trece de 50° latitudine nordică, spre est
trece puţin în Ucraina. La noi apare mai rar decât celelalte două specii – Tilia cordata şi Tilia tomentosa – la câmpie şi coline.
c. Tilia tomentosa Arealul general al speciei este cantonat în Balcani, Asia Mică. La noi este abundent în
şleauri de câmpie, asociat cu stejarii xero-termofili; în estul Moldovei, intră în arboretele de fag sau fag cu gorun. Tot abundent este şi în silvostepa de nord a Moldovei şi în nord-vestul Olteniei, iar în Dobrogea formează pe alocuri teişuri pure destul de întinse.
Alte specii ale genului Tilia sunt:
62

Diversitatea ecologică a speciilor forestiere
- Tilia dasystyla - Tilia mongolica - Tilia americana
- Tilia petiolaris - Tilia heterophylla - Tilia mandshurica
FAMILIA OLEACEAE Lindl.
Familia Oleaceae cuprinde arbori şi arbuşti, cu frunze opuse, simple sau imparipenat compuse, caduce sau persistente, flori bisexuate sau poligame, pe tipul 4 sau 5, grupate în raceme, spice sau cime terminale sau axilare, iar fructul este bacă, drupă, capsulă sau samară.
I. Genul Fraxinus L.
a. Fraxinus excelsior Specia are un areal larg european, dar nu depăşeşte 60° latitudine nordică. În România,
apare frecvent de la câmpie şi până în zona montană, urcând până la 1400 m altitudine. Apare diseminat sau în pâlcuri în toate pădurile de amestec şi, mai rar, în arborete pure.
b. Fraxinus ornus Specia are un areal sudic, mediteranean, fiind răspândit în sudul Europei şi Asiei. Limita
nordică a arealului trece prin ţara noastră pe o linie marcată de Carpaţii Meridionali.
c. Fraxinus pallisae Specia are un areal pontic, ce trece şi în Iugoslavia, la noi vegetând în staţiunile umede din
sudul ţării, mai ales în silvostepa marină, pe grindurile din Delta Dunării.
d. Fraxinus coriariifolia Este un element pontic-caucazian, la noi fiind frecvent în pădurile din silvostepa Dobrogei
şi a Moldovei. Alte specii ale genului Fraxinus sunt:
- Fraxinus bungeana - Fraxinus sieboldiana - Fraxinus paxiana - Fraxinus longicuspis - Fraxinus rhynchophylla - Fraxinus anomala - Fraxinus xanthoxyloides - Fraxinus velutina - Fraxinus platypoda - Fraxinus latifolia
- Fraxinus americana - Fraxinus pennsylvanica - Fraxinus quadrangulata - Fraxinus nigra - Fraxinus mandshurica - Fraxinus holotricha - Fraxinus potamophila - Fraxinus syriaca - Fraxinus angustifolia - Fraxinus biltmoreana
63

Diversitatea ecologică a speciilor forestiere
SCURTĂ INCURSIUNE ÎN PĂDURILE CANADIENE
Pădurile sunt caracteristica dominantă a peisajului canadian. Ele acoperă 398 milioane
hectare adică 43% din suprafaţa totală a teritoriului naţional, restul fiind compus din spaţii naturale: Barren Grounds (tundra canadiană), muskeg (turbării), zone stâncoase şi tufărişuri.
Suprafaţa pădurilor canadiene este în jur de 1,5 ori decât pădurile Europei, este egală cu a pădurilor Asiei şi reprezintă jumătate din cea a pădurilor fostei Uniuni Sovietice. În total, 10% din pădurile Globului se găsesc în Canada.
Pădurile de conifere reprezintă aproape 80% din totalul pădurilor pe picior. Picea glauca şi Picea mariana sunt coniferele cele mai răspândite, celelalte conifere importante aparţinând genurilor Pinus, Abies, Tsuga, Thuja şi Pseudotsuga. În pădurile de foioase predomină plopii (Populus sp.) şi mestecenii (Betula sp.), cărora li se asociază arţarii (Acer sp.), dar şi alte specii.
Mai mult decât jumătate din pădurile canadiene sunt situate în regiunea boreală care se întinde de pe coasta atlantică şi până la Cercul Polar în teritoriile de nord-vest şi aproape atinge Pacificul în Columbia Britanică.
Speciile cele mai comune sunt molidul brumăriu (Picea glauca) şi molidul negru (Picea mariana), bradul de vest (Abies lasiocarpa), bradul balsamifer (Abies balsamea), pinul răsucit (Pinus contorta) şi pinul banksian (Pinus banksiana). De remarcat speciile pioniere Populus tremuloides şi Betula papyrifera.
Pe ansamblul pădurilor, creşterea anuală medie este de 1,5 m3/ha. Cu toate acestea, creşterile cele mai susţinute se înregistrează pe coasta pacifică în Columbia Britanică, unde principalele specii sunt tuia gigantă (Thuja plicata), molidul de Sitka (Picea sitchensis) şi duglasul verde (Pseudotsuga menziesii).
64