
GLICOLIZA Calea Embden – Meyerhof · GLICOLIZA Calea Embden–Meyerhof 1. Definiţie, localizare,...
71
Curs 2: •GLUCIDE - aspecte generale •GLICOLIZA •Ciclul acidului LACTIC ( ciclul CORI) •Căi alternative ale PIRUVATULUI
Transcript of GLICOLIZA Calea Embden – Meyerhof · GLICOLIZA Calea Embden–Meyerhof 1. Definiţie, localizare,...

Curs 2: •GLUCIDE - aspecte generale
•GLICOLIZA
•Ciclul acidului LACTIC ( ciclul CORI)
•Căi alternative ale PIRUVATULUI

GLUCIDE - aspecte generale
1. Definiţie, roluri
2. Clasificare, structură, proprietăţi, izomerie
3. Digestia, absorbţia şi transportul glucidelor
4. Importanţa medicală
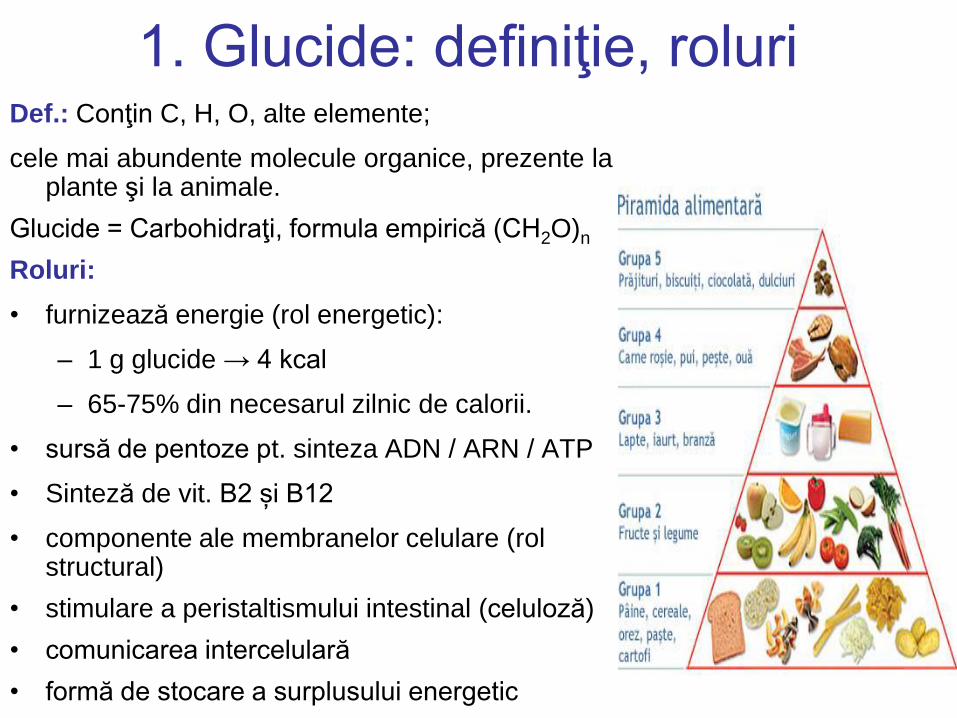
1. Glucide: definiţie, roluri Def.: Conţin C, H, O, alte elemente;
cele mai abundente molecule organice, prezente la plante şi la animale.
Glucide = Carbohidraţi, formula empirică (CH2O)n
Roluri:
• furnizează energie (rol energetic):
– 1 g glucide → 4 kcal
– 65-75% din necesarul zilnic de calorii.
• sursă de pentoze pt. sinteza ADN / ARN / ATP
• Sinteză de vit. B2 și B12
• componente ale membranelor celulare (rol structural)
• stimulare a peristaltismului intestinal (celuloză)
• comunicarea intercelulară
• formă de stocare a surplusului energetic
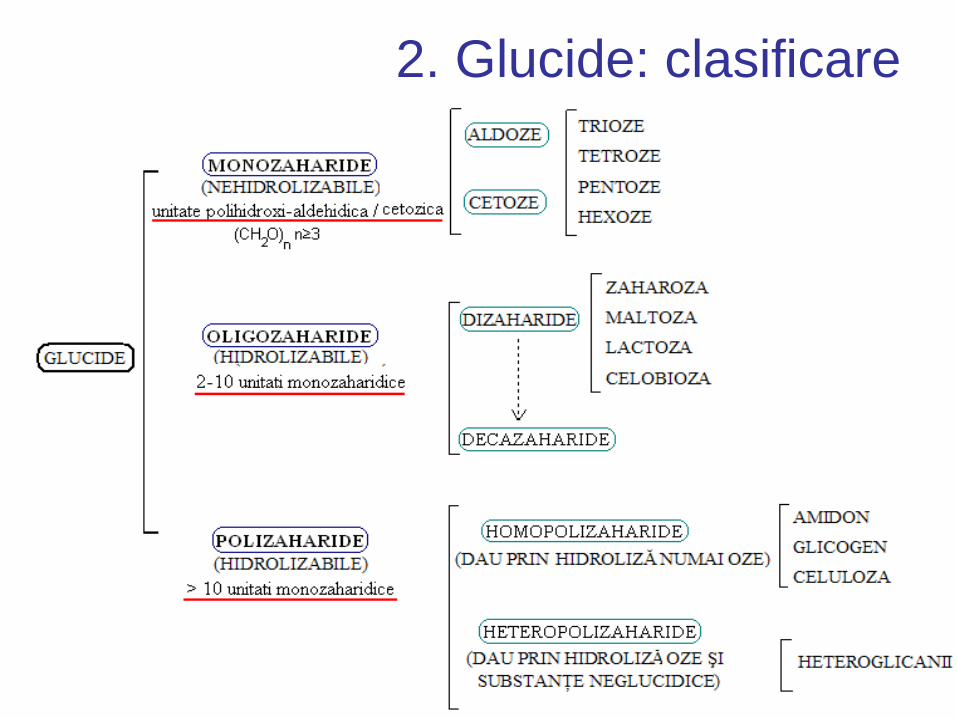
2. Glucide: clasificare

2. Glucide: structură Monozaharide:
– Aldoze = conţin gruparea aldehidică
• ex. gliceraldehida
– Cetoze = conţin gruparea cetozică
• ex. dihidroxiacetona, fructoza, xiluloza
• au formula generală (CH2O)n, n≥3
• Monozaharidele ≥ 5 atomi de C au structuri predominant ciclice
• În funcție de numărul de atomi de C, monozaharide sunt denumite:
– trioze (C3: ex. gliceraldehida),
– tetroze (C4: ex. eritroza),
– pentoze (C5: ex. riboza),
– hexoze (C6: ex. glucoza), etc.
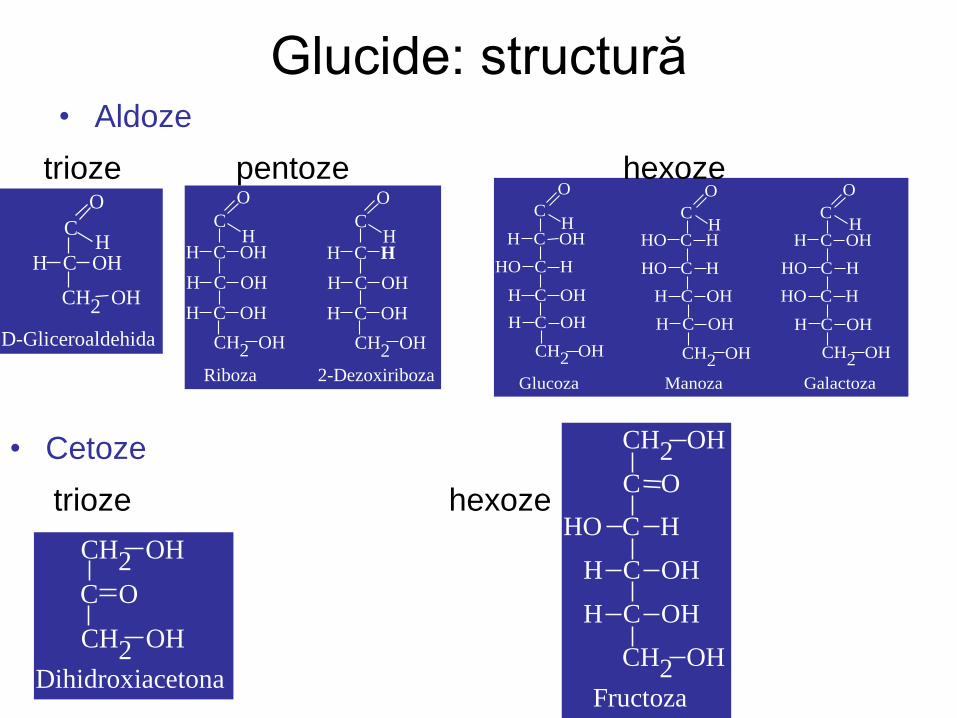
Glucide: structură • Aldoze
H C OH
C
CH2 OH
O
H
D-Gliceroaldehida
H C OH
C
O
H
H C OH
H C OH
CH2 OH
Riboza 2-Dezoxiriboza
H C H
C
O
H
H C OH
H C OH
CH2 OH
H C OH
C
O
H
HO C H
H C OH
CH2 OH
H C OH
C
O
H
HO C H
H C OH
CH2 OH
H C OH
HO C H
Glucoza Manoza Galactoza
H C OH
C
O
H
HO C H
CH2 OH
H C OH
HO C H
trioze pentoze hexoze
• Cetoze
CH2 OH
C O
CH2 OH
Dihidroxiacetona
HO C H
CH2 OH
H C OH
C O
CH2 OH
H C OH
Fructoza
trioze hexoze
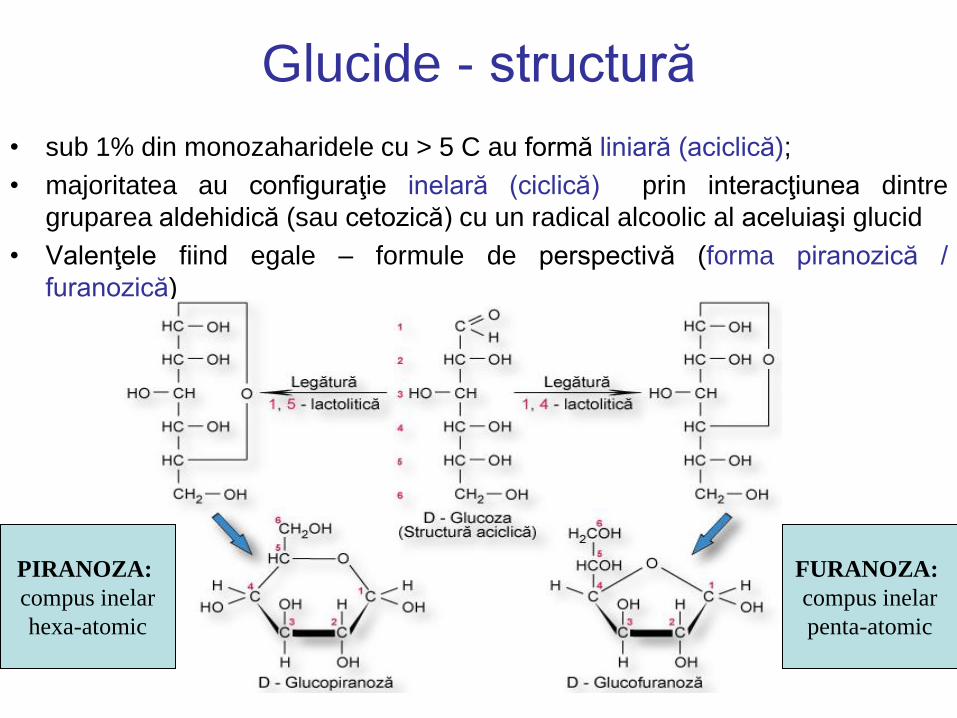
Glucide - structură
• sub 1% din monozaharidele cu > 5 C au formă liniară (aciclică);
• majoritatea au configuraţie inelară (ciclică) prin interacţiunea dintre
gruparea aldehidică (sau cetozică) cu un radical alcoolic al aceluiaşi glucid
• Valenţele fiind egale – formule de perspectivă (forma piranozică /
furanozică)
PIRANOZA:
compus inelar
hexa-atomic
FURANOZA:
compus inelar
penta-atomic
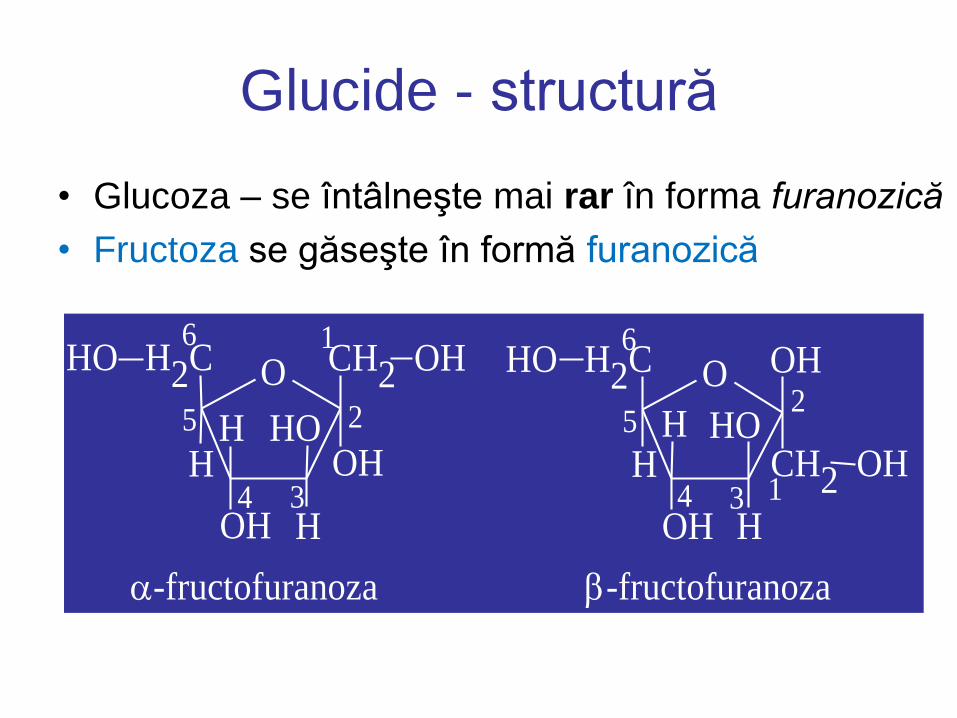
Glucide - structură
• Glucoza – se întâlneşte mai rar în forma furanozică
• Fructoza se găseşte în formă furanozică
O
HO
HO H2C
OH
OH H
H
-fructofuranoza -fructofuranoza
H
CH2 OH1
2
34
5
6
OHO H2C
OH
OH
H H
H
CH2 OH1
2
34
5
6
HO

2. Glucide - proprietăţi
• Statusul atomului de oxigen legat de atomul anomeric (fostul C carbonilic) dictează caracterul reducător sau nereducător al unui glucid, restul grupărilor OH nefiind implicate
• testele colorimetrice ce utilizează reactivi cromogeni detectează prezenţa glucidelor în urină
• Identificarea exactă a glucidului necesită teste adiţionale specifice
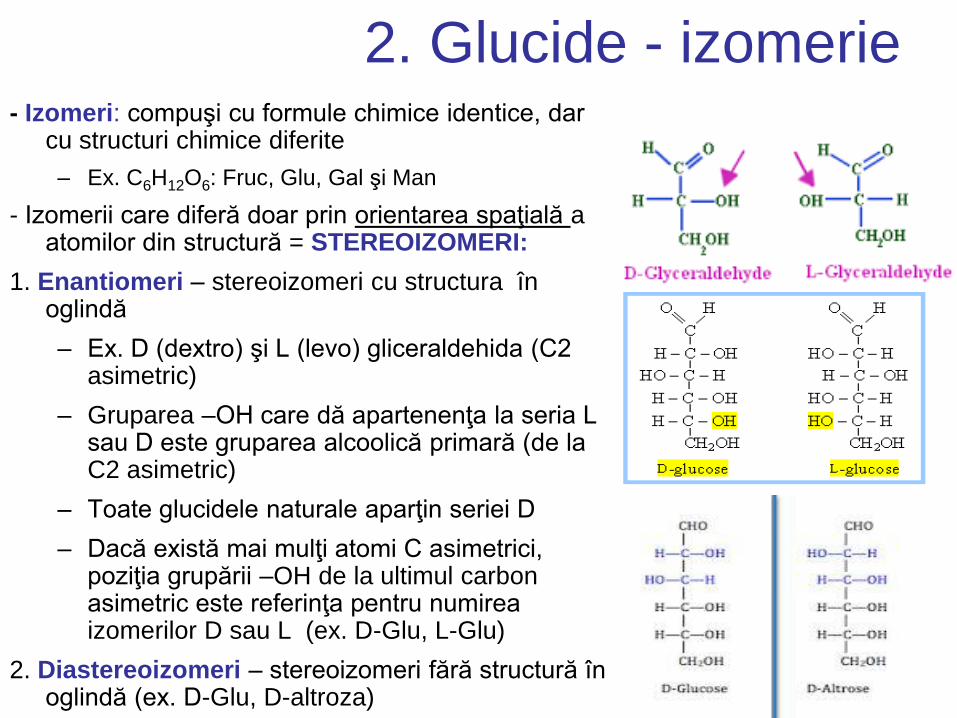
2. Glucide - izomerie - Izomeri: compuşi cu formule chimice identice, dar
cu structuri chimice diferite
– Ex. C6H12O6: Fruc, Glu, Gal şi Man
- Izomerii care diferă doar prin orientarea spaţială a atomilor din structură = STEREOIZOMERI:
1. Enantiomeri – stereoizomeri cu structura în oglindă
– Ex. D (dextro) şi L (levo) gliceraldehida (C2 asimetric)
– Gruparea –OH care dă apartenenţa la seria L sau D este gruparea alcoolică primară (de la C2 asimetric)
– Toate glucidele naturale aparţin seriei D
– Dacă există mai mulţi atomi C asimetrici, poziţia grupării –OH de la ultimul carbon asimetric este referinţa pentru numirea izomerilor D sau L (ex. D-Glu, L-Glu)
2. Diastereoizomeri – stereoizomeri fără structură în oglindă (ex. D-Glu, D-altroza)
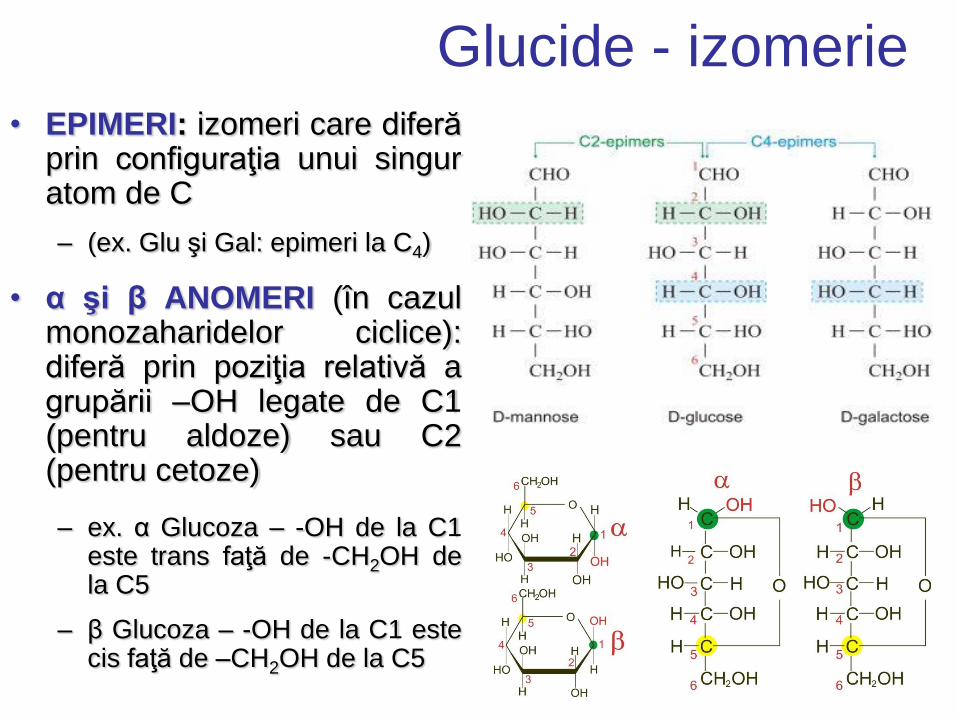
Glucide - izomerie • EPIMERI: izomeri care diferă
prin configuraţia unui singur atom de C
– (ex. Glu şi Gal: epimeri la C4)
• α şi β ANOMERI (în cazul monozaharidelor ciclice): diferă prin poziţia relativă a grupării –OH legate de C1 (pentru aldoze) sau C2 (pentru cetoze)
– ex. α Glucoza – -OH de la C1 este trans faţă de -CH2OH de la C5
– β Glucoza – -OH de la C1 este cis faţă de –CH2OH de la C5
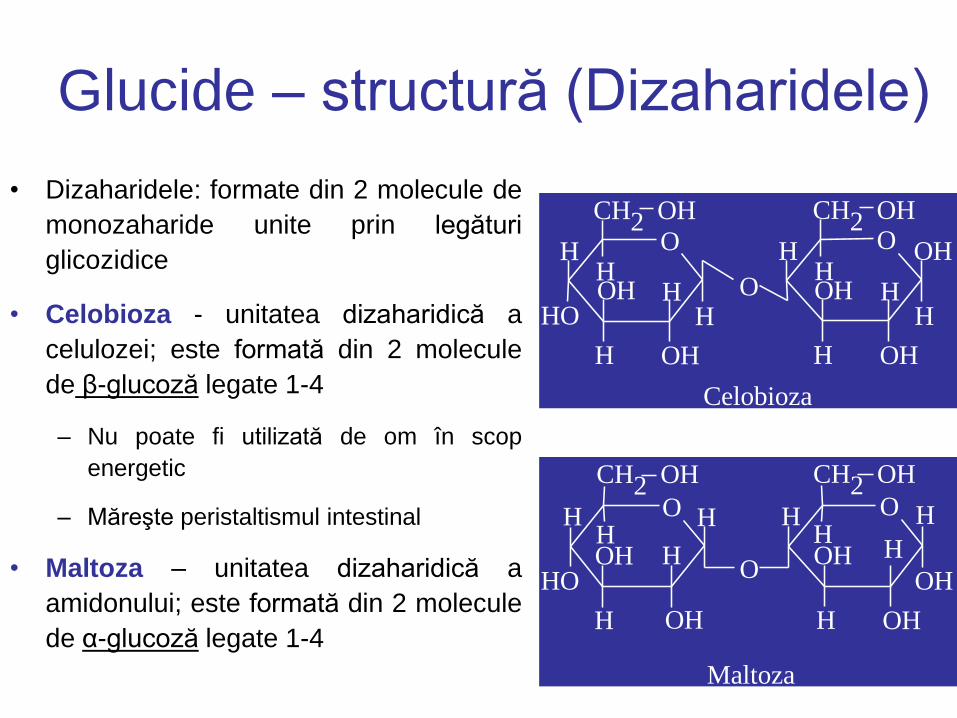
Glucide – structură (Dizaharidele)
• Dizaharidele: formate din 2 molecule de
monozaharide unite prin legături
glicozidice
• Celobioza - unitatea dizaharidică a
celulozei; este formată din 2 molecule
de β-glucoză legate 1-4
– Nu poate fi utilizată de om în scop
energetic
– Măreşte peristaltismul intestinal
• Maltoza – unitatea dizaharidică a
amidonului; este formată din 2 molecule
de α-glucoză legate 1-4
CH2 OH
OH
OH H
HHO
H
H
H
CH2 OH
OH
OH H
HO
H
H
H OH
Celobioza
O O
CH2 OH
OH
OH H
HHO
H
H H
CH2 OH
OH
OH H
HO
H
H H
OH
Maltoza
O O
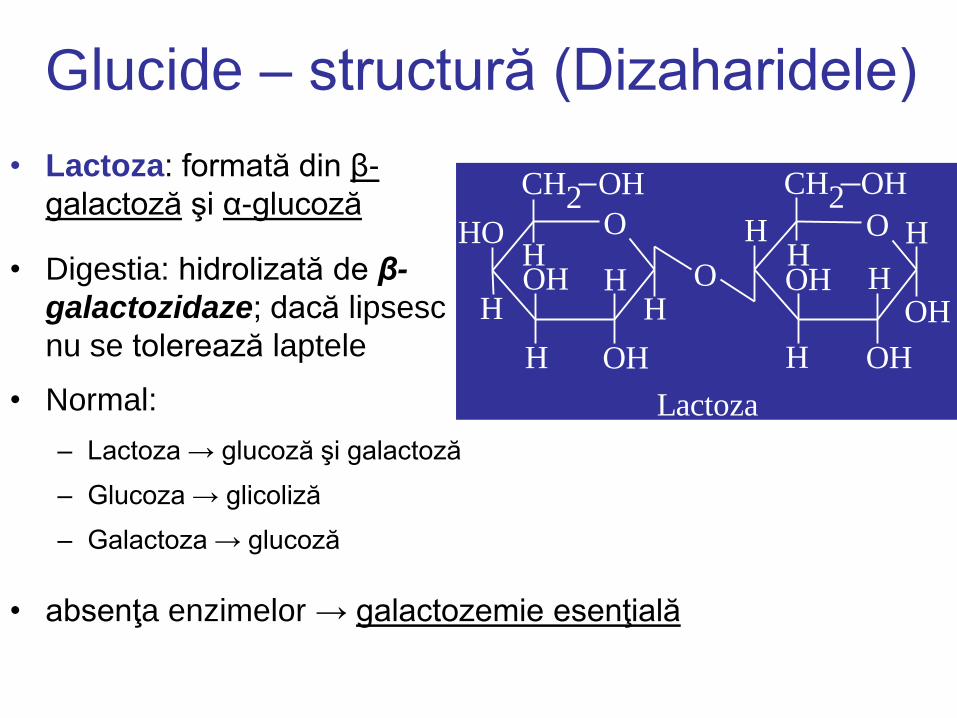
Glucide – structură (Dizaharidele)
• Lactoza: formată din β-
galactoză şi α-glucoză
• Digestia: hidrolizată de β-
galactozidaze; dacă lipsesc
nu se tolerează laptele
• Normal:
– Lactoza → glucoză şi galactoză
– Glucoza → glicoliză
– Galactoza → glucoză
CH2 OH
OH
OH H
H
HO
H
H H
CH2 OH
OH
OH H
HO
H
H H
OH
Lactoza
OO
• absenţa enzimelor → galactozemie esenţială
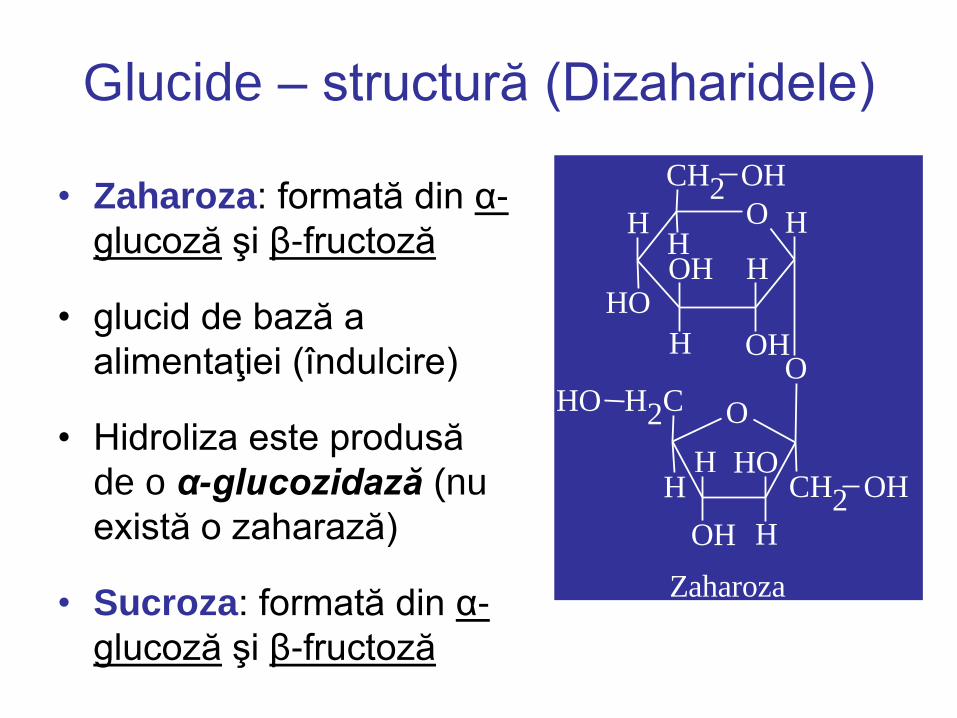
Glucide – structură (Dizaharidele)
• Zaharoza: formată din α-
glucoză şi β-fructoză
• glucid de bază a
alimentaţiei (îndulcire)
• Hidroliza este produsă
de o α-glucozidază (nu
există o zaharază)
• Sucroza: formată din α-
glucoză şi β-fructoză
HO H2C
OH
H
H
H
Zaharoza
HOCH2 OH
CH2 OH
OH
OH H
H
H
H H
HO
O
O
O

3. Digestia glucidelor
- Digestia carbohidraţilor alimentari se desfăşoară în:
○ cavitatea orală
○ intestin
• Procesul are loc rapid şi se încheie când chimul
gastric ajunge la joncţiunea duodeno-jejunală
• Enzime implicate:
○ Endoglicozidazele: hidroliza oligo- şi polizaharidelor
○ Dizaharidazele: hidroliza dizaharidelor
○ Glicozidazele specifice: hidroliza legăturilor glicozidice
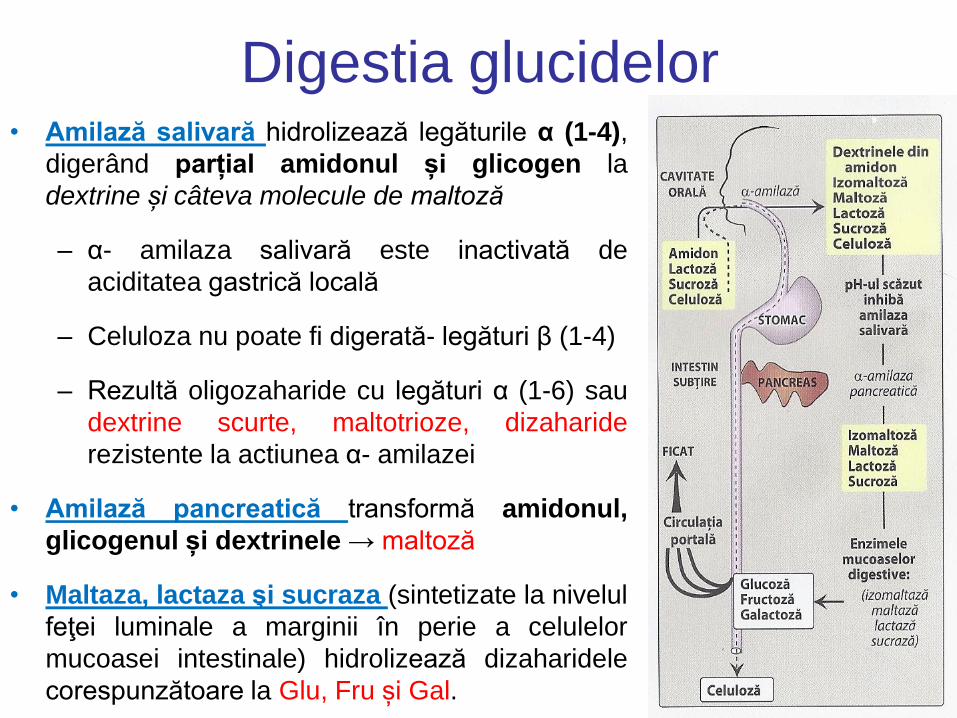
Digestia glucidelor • Amilază salivară hidrolizează legăturile α (1-4),
digerând parțial amidonul și glicogen la
dextrine și câteva molecule de maltoză
– α- amilaza salivară este inactivată de
aciditatea gastrică locală
– Celuloza nu poate fi digerată- legături β (1-4)
– Rezultă oligozaharide cu legături α (1-6) sau
dextrine scurte, maltotrioze, dizaharide
rezistente la actiunea α- amilazei
• Amilază pancreatică transformă amidonul,
glicogenul și dextrinele → maltoză
• Maltaza, lactaza şi sucraza (sintetizate la nivelul
feţei luminale a marginii în perie a celulelor
mucoasei intestinale) hidrolizează dizaharidele
corespunzătoare la Glu, Fru și Gal.
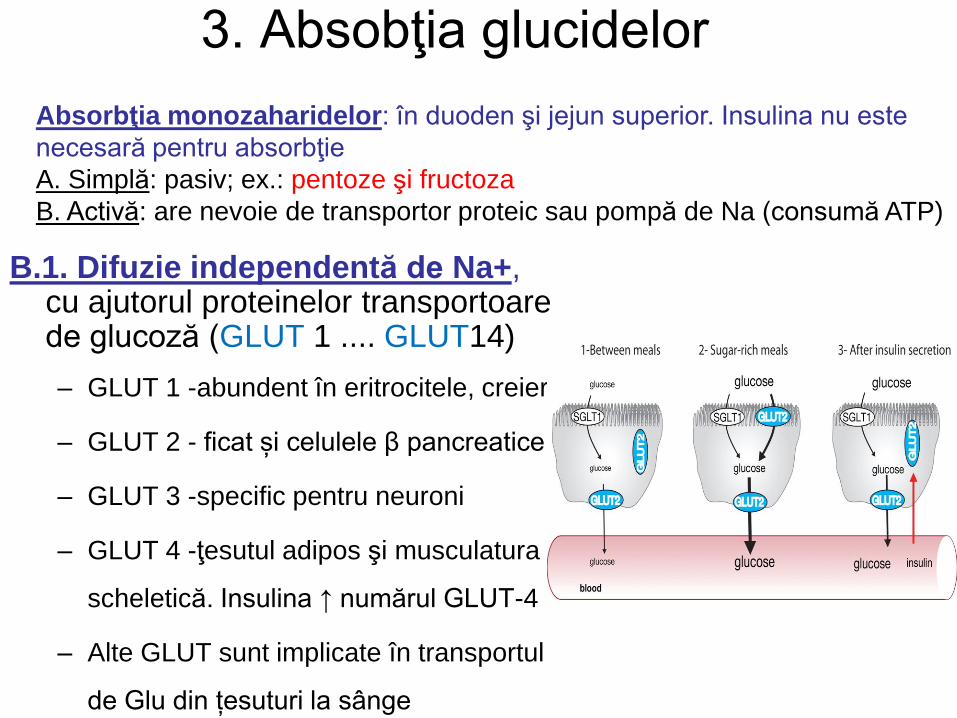
3. Absobţia glucidelor
Absorbţia monozaharidelor: în duoden şi jejun superior. Insulina nu este
necesară pentru absorbţie
A. Simplă: pasiv; ex.: pentoze şi fructoza
B. Activă: are nevoie de transportor proteic sau pompă de Na (consumă ATP)
B.1. Difuzie independentă de Na+, cu ajutorul proteinelor transportoare de glucoză (GLUT 1 .... GLUT14)
– GLUT 1 -abundent în eritrocitele, creier
– GLUT 2 - ficat și celulele β pancreatice
– GLUT 3 -specific pentru neuroni
– GLUT 4 -ţesutul adipos şi musculatura
scheletică. Insulina ↑ numărul GLUT-4
– Alte GLUT sunt implicate în transportul
de Glu din țesuturi la sânge
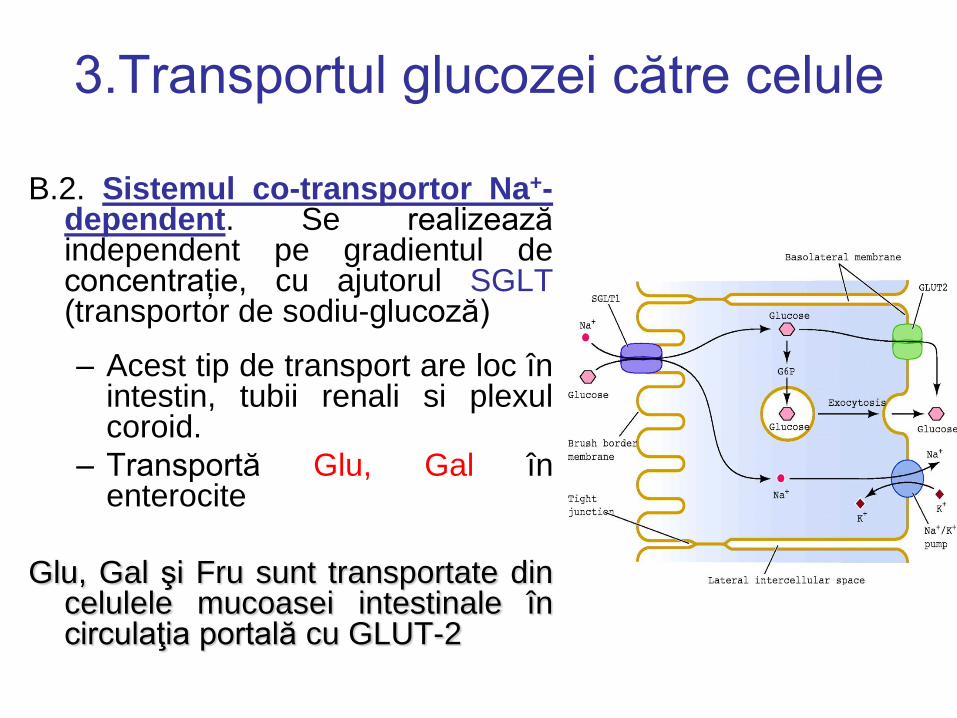
3.Transportul glucozei către celule
B.2. Sistemul co-transportor Na+-dependent. Se realizează independent pe gradientul de concentrație, cu ajutorul SGLT (transportor de sodiu-glucoză)
– Acest tip de transport are loc în intestin, tubii renali si plexul coroid.
– Transportă Glu, Gal în enterocite
Glu, Gal şi Fru sunt transportate din celulele mucoasei intestinale în circulaţia portală cu GLUT-2

Soarta glucozei după absorbţie
• Oxidare în principal prin glicoliză și ciclul Krebs în CO2 și H2O, cu eliberare de energie stocată în ATP.
• Sinteza altor carbohidraţi:
– Pentoză prin șuntul pentozelor
– Acid glucuronic prin calea acidului uronic
– Galactoză și lactoză
– Mucopolizaharide
• Excesul este transformat în glicogen prin glicogeneză
• Un exces marcat de glucoză, duce la stocarea lipidelor prin lipogeneză
• Sinteza aminoacizi neesențiali din intermediarii de glucoză.

4. Importanţa medicală
- Glucoza – absolut necesară organismului
– Nivelul glucozei în sânge = glicemie
– Interval biologic de referinţă: 70-105 mg/dl (→100 mg/dl)
– Valori ↓= hipoglicemie; valori sub limita inferioară – coma
hipoglicemică
– Valori ↑= hiperglicemie; se întâlnesc în diabet zaharat
- Excesul = depus sub formă de glicogen (glicogenogeneză)
- Mobilizarea Glu din depozite = glicogenoliză

Importanţa medicală
- tulburări ale depozitării sau mobilizării Glu →
glicogenoze
- Glu = substrat energetic major pentru SNC.
Când rezerva de Glu a ficatului este epuizată →
gluconeogeneză
- Concluzie: prin glicogenogeneză, glicogenoliză şi
gluconeogeneză – aportul de Glu este continuu

GLICOLIZA
Calea Embden–Meyerhof
1. Definiţie, localizare, generalităţi
2. Glicoliza: etape, reacţii, reglare
3. Bilanţ energetic
Otto Fritz Meyerhof 1884-1951
1922– Premiul Nobel
Gustav Embden
1874-1933

1. Glicoliza – definiţie, localizare
• Definiţie: secvenţă de reacţii enzimatice prin
care, în aerobioză, 1 moleculă Glu (6C) → 2
molecule piruvat (3C) şi metaboliţi
intermediari utilizaţi în alte căi metabolice
• Cale metabolică importantă: furnizează
energie (ATP)
~120 g Glu/zi sunt utilizate de creierul adultului pentru necesarul de ATP
• Localizare: toate celulele, citosol
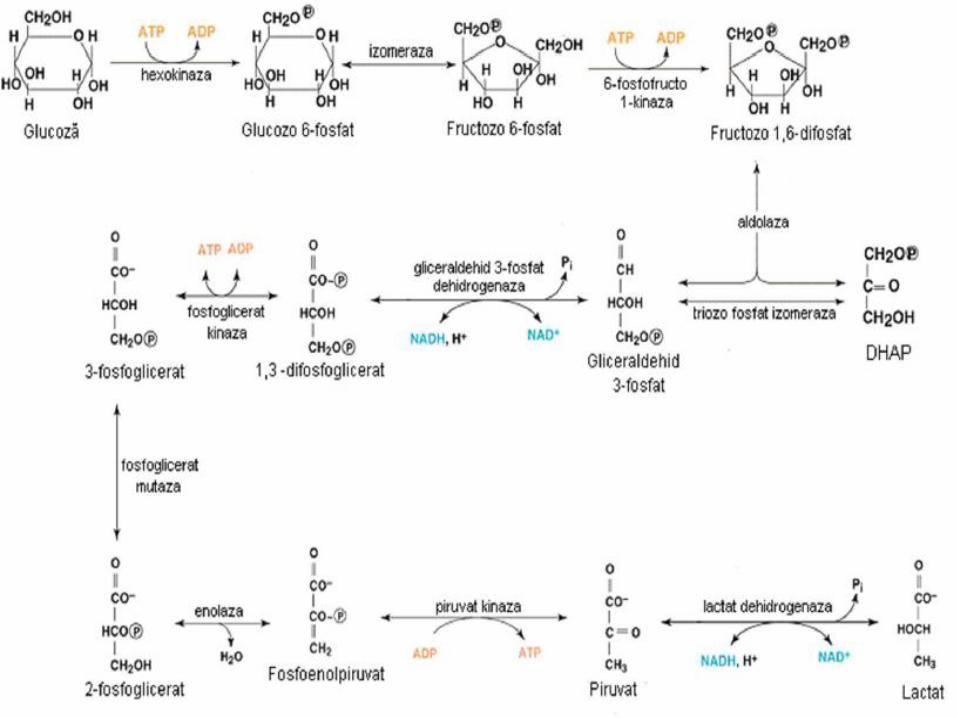

Glicoliza - generalităţi
• Glicoliza aerobă:
– secvenţă de 10 reacţii: produs final = Piruvat (Py)
– Furnizează energie (2 ATP şi 2 NADH)
– Localizare: celule care deţin mitocondrii şi aport adecvat de O2
– O2= indispensabil reoxidării NADH rezultat din oxidarea GA-3P
– pregăteşte teren pentru decarboxilarea oxidativă Py→ acetil-CoA (sursă de energie a ciclului Krebs)
– Prin metabolizarea acetil-CoA la nivelul ciclului Krebs şi a lanţului respirator mitocondrial rezultă 36 sau 38 moli ATP / 1 mol Glu

Glicoliza - generalităţi
• Glicoliza anaerobă:
• secvenţă de 11 reacţii:
produs final = Lactat
(←reducerea Py)
• permite sinteza de ATP
(2 ATP / 1 moleculă Glu)
în ţesuturi cu aportul de
O2 insuficient

Ţesuturi dependente de glicoliză
pentru furnizarea ATP
• Hematii, cornee, cristalin şi anumite regiuni ale retinei
– nu deţin mitocondrii (lactatul = produs final al glicolizei)
• Rinichi (medulara), testicule, leucocite si fibre
muscular netede – conţin puţine mitocondrii (aproape
total dependente de glicoliză ca sursă de ATP)
• La adulţii sănătoşi, aceste ţesuturi consumă
aproximativ 40 g Glu / zi pentru producerea ATP
• Sursa majoră de ac. lactic în sânge = hematiile şi
muşchii
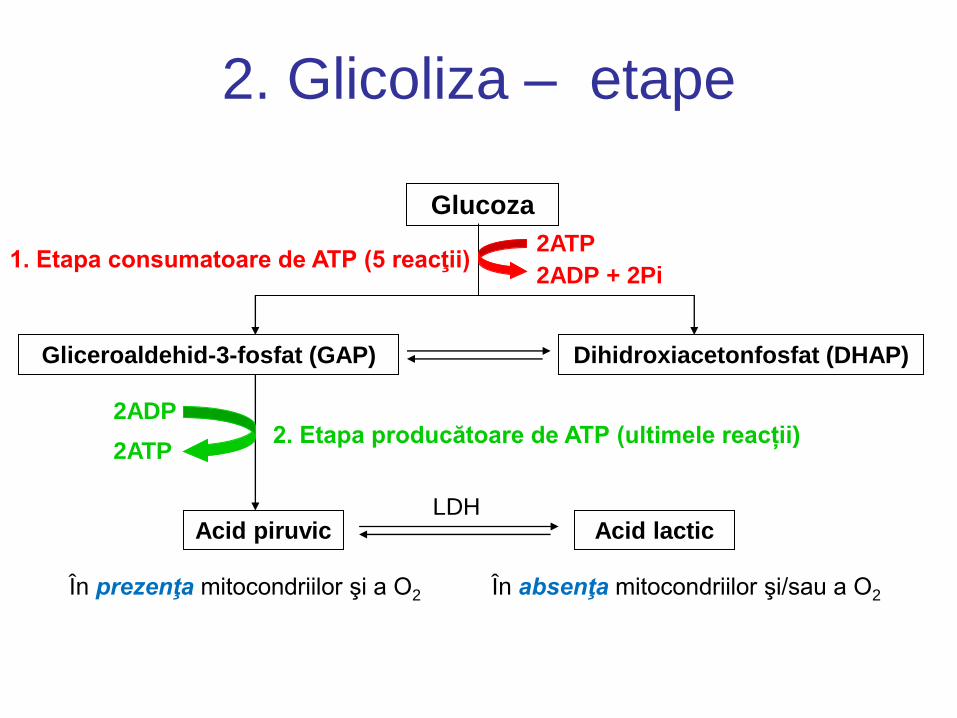
2. Glicoliza – etape
Glucoza
2ATP
2ADP + 2Pi 1. Etapa consumatoare de ATP (5 reacţii)
Gliceroaldehid-3-fosfat (GAP) Dihidroxiacetonfosfat (DHAP)
2ADP
2ATP 2. Etapa producătoare de ATP (ultimele reacții)
Acid piruvic
În prezenţa mitocondriilor şi a O2
LDH Acid lactic
În absenţa mitocondriilor şi/sau a O2

2. Glicoliza – reacţii
• Toţi intermediarii glicolizei (de la Glu la Py) sunt fosforilaţi
• Gruparea fosfat a intermediarilor glicolizei are 2 roluri:
1. Menținerea moleculelor energetice în celulă (compuşi hidrofili)
- cu un grup polar negativ sunt incapabili să traverseze
membrana celulară prin simpla difuziune la pH-ul fiziologic.
2. interacţiunea enzimă-substrat mai bună
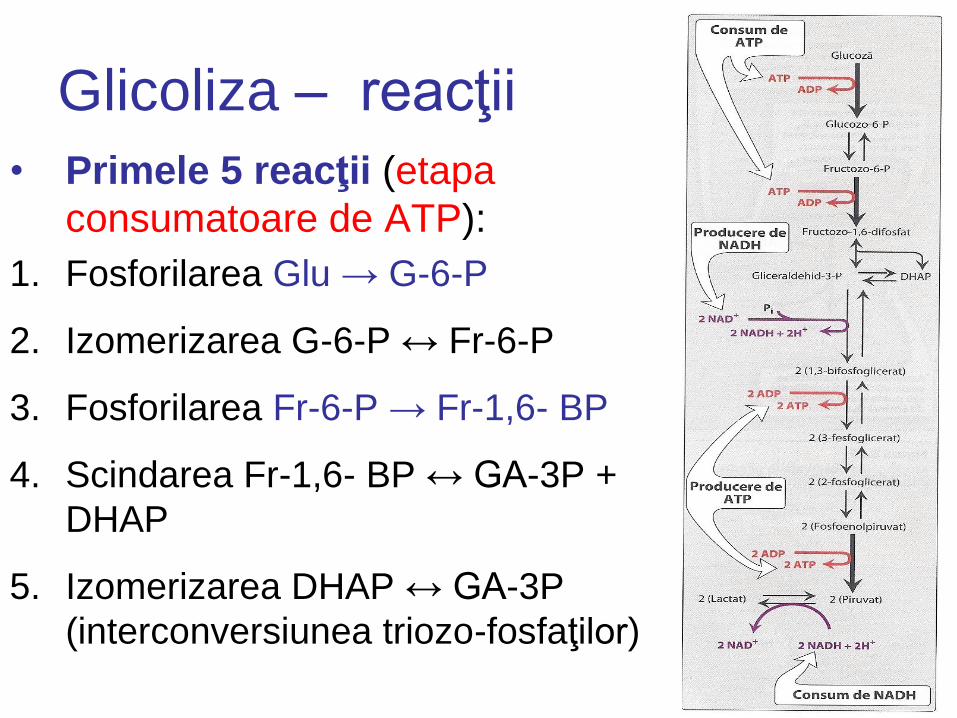
Glicoliza – reacţii
• Primele 5 reacţii (etapa
consumatoare de ATP):
1. Fosforilarea Glu → G-6-P
2. Izomerizarea G-6-P ↔ Fr-6-P
3. Fosforilarea Fr-6-P → Fr-1,6- BP
4. Scindarea Fr-1,6- BP ↔ GA-3P +
DHAP
5. Izomerizarea DHAP ↔ GA-3P
(interconversiunea triozo-fosfaţilor)
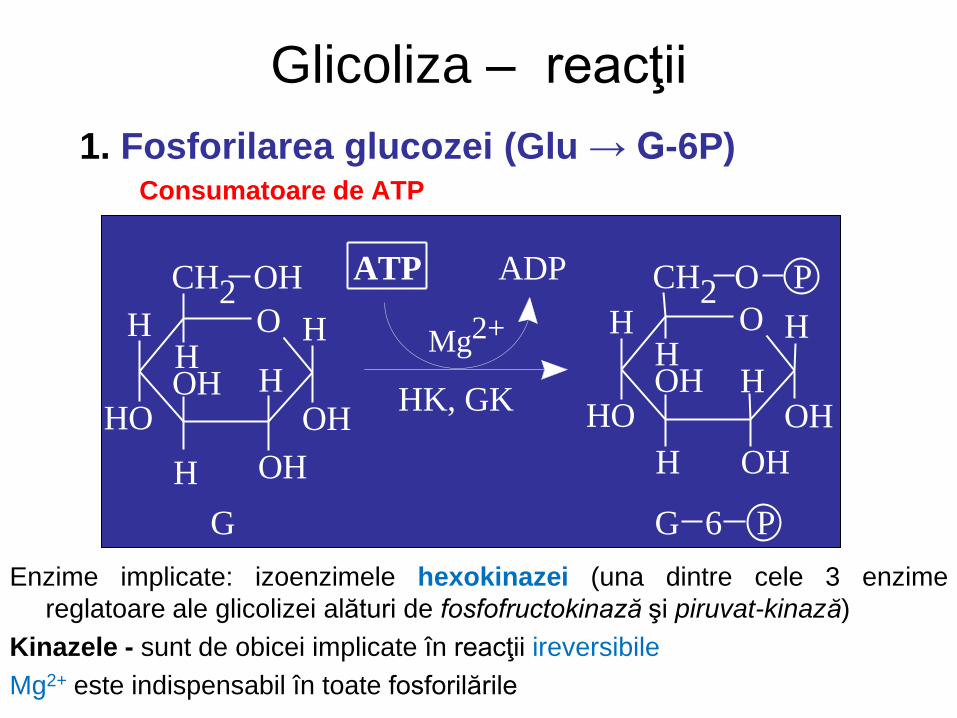
Glicoliza – reacţii
1. Fosforilarea glucozei (Glu → G-6P) Consumatoare de ATP
OH
CH2 OH
OHOH H
HHO
H
H
OH
G G 6 P
OH
CH2 O P
OHOH H
HHO
H
H
OH
ATP ADP
Mg2+
HK, GK
Enzime implicate: izoenzimele hexokinazei (una dintre cele 3 enzime
reglatoare ale glicolizei alături de fosfofructokinază şi piruvat-kinază)
Kinazele - sunt de obicei implicate în reacţii ireversibile
Mg2+ este indispensabil în toate fosforilările

Glicoliza – reglare
Hexokinaza (HK)– prezentă în toate ţesuturile
• specificitate largă pt substrat (fosforilează diverse hexoze, nu doar Glu)
• prima enzimă alosterică a glicolizei
• Reglare: inhibată prin mecanism feed-back de produsul final de
reacţie G-6-P (se acumulează când rata de metabolizare scade)
• Km ↓ pt Glu (are afinitate ↑ pt Glu)
• Vmax ↓ pt Glu (nu poate reţine hexozele fosforilate în interiorul
celulei şi nu poate fosforila o cantitate de glucide > decât necesarul)
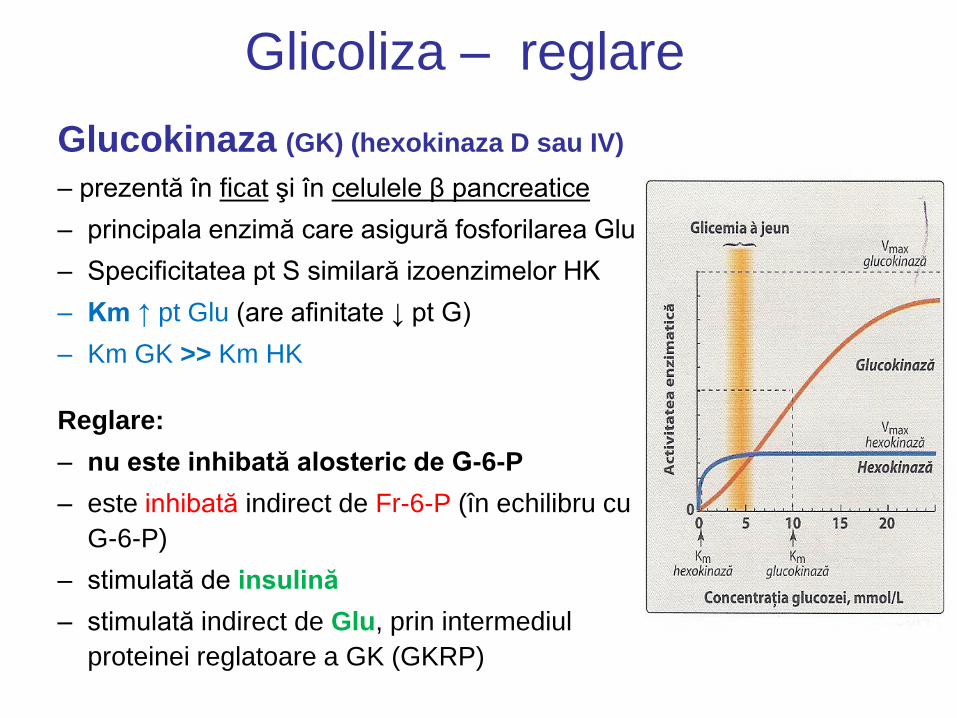
Glicoliza – reglare
Glucokinaza (GK) (hexokinaza D sau IV)
– prezentă în ficat şi în celulele β pancreatice
– principala enzimă care asigură fosforilarea Glu
– Specificitatea pt S similară izoenzimelor HK
– Km ↑ pt Glu (are afinitate ↓ pt G)
– Km GK >> Km HK
Reglare:
– nu este inhibată alosteric de G-6-P
– este inhibată indirect de Fr-6-P (în echilibru cu
G-6-P)
– stimulată de insulină
– stimulată indirect de Glu, prin intermediul
proteinei reglatoare a GK (GKRP)

Glicoliza – reglare
Glucokinaza Roluri - intervine în controlul homeostaziei Glu
plasmatice prin:
– În celulele β pancreatice
• detector de Glu
• determină pragul secreţiei de insulină
– în ficat
• reduce glicemia din perioada de absorţie (GLUT-2)
• permite ficatului să elimine eficient fluxul masiv de Glu
adus prin sângele portal după alimentaţie
• previne supraâncărcarea cu Glu a circulaţiei sistemice

Glicoliza – reacţii
2. Izomerizarea G-6-P ↔ Fr-6-P
• Enzima implicată: fosfohexoizomeraza (fosfoglucoizomeraza)
• Reacţia este complet reversibilă
• Nu constituie o etapă limitantă de viteză
O
HO H
OH
H
H
OHP O H2C
CH2 OH
OH
CH2 O P
OHOH H
HHO
H
H
OH
PHI
G 6 P Fr 6 P

Glicoliza – reacţii 3. Fosforilarea Fr-6-P → Fr-1,6-BP
Consumatoare de ATP
• Enzima implicată:
Fosfofructokinaza-1 (PFK-1)
– enzimă “cheie” (a 2-a enzimă
alosterică a glicolizei)
Cea mai importantă etapă
limitantă de viteză a
glicolizei
O
HO H
OH
H
H
OHP O H2C
CH2 OH
Fr 6 P
PFrK
ATP
Mg2+
O
HO H
OH
H
H
OHP O H2C
CH2 O P
Fr 1,6 BP
ADP
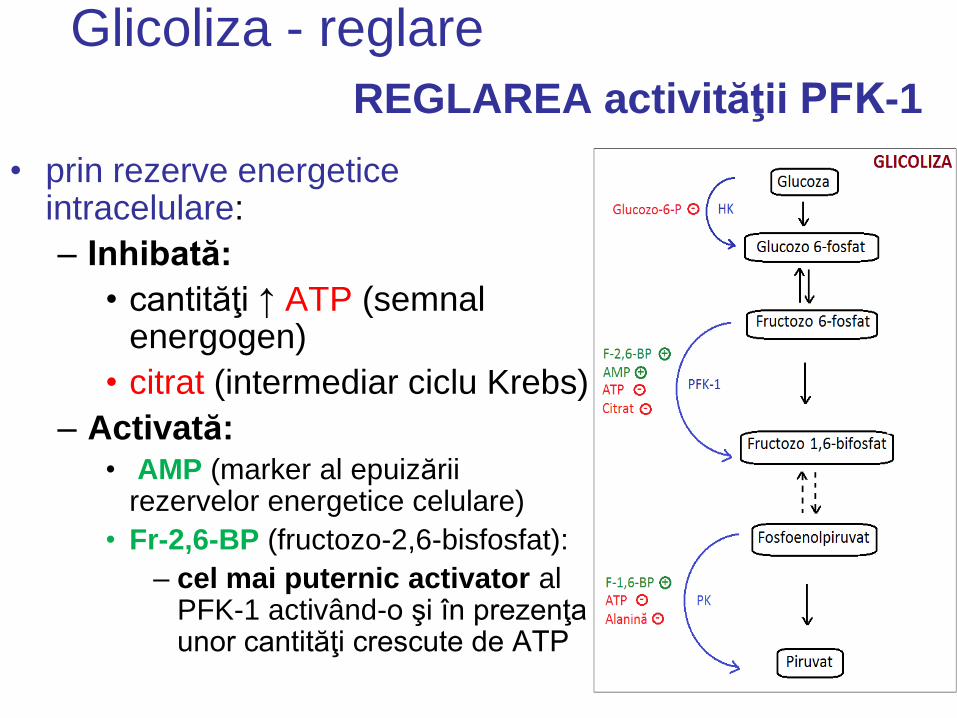
REGLAREA activităţii PFK-1
• prin rezerve energetice intracelulare:
– Inhibată:
• cantităţi ↑ ATP (semnal energogen)
• citrat (intermediar ciclu Krebs)
– Activată: • AMP (marker al epuizării
rezervelor energetice celulare)
• Fr-2,6-BP (fructozo-2,6-bisfosfat):
– cel mai puternic activator al PFK-1 activând-o şi în prezenţa unor cantităţi crescute de ATP
Glicoliza - reglare
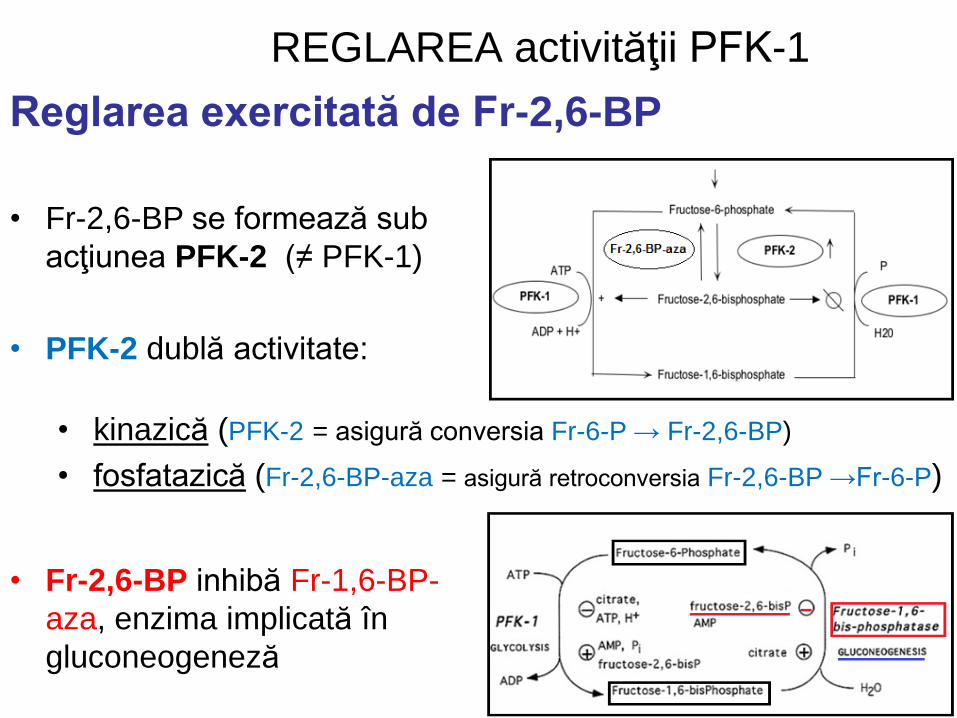
REGLAREA activităţii PFK-1
• Fr-2,6-BP se formează sub
acţiunea PFK-2 (≠ PFK-1)
• PFK-2 dublă activitate:
Reglarea exercitată de Fr-2,6-BP
• Fr-2,6-BP inhibă Fr-1,6-BP-
aza, enzima implicată în
gluconeogeneză
• kinazică (PFK-2 = asigură conversia Fr-6-P → Fr-2,6-BP)
• fosfatazică (Fr-2,6-BP-aza = asigură retroconversia Fr-2,6-BP →Fr-6-P)

Reglarea exercitată de Fr-2,6-BP - continuare
• Acţiunea reciprocă a Fr-2,6-BP de
activare a glicolizei şi de inhibare a
gluconeogenezei împiedică crearea unui
ciclu inutil (Glu → Py în paralel cu Glu ← Py)
• Există un control hormonal asupra PFK-2 / Fr-2,6-BP-aza :
Postprandial:
• ↓[Glucagon] şi ↑[insulină]→ stimularea PFK-2 , deci ↑ [Fr-2,6-BP],
cu activarea glicolizei
• Fr-2,6-BP= semnal intracelular ce indică prezenţa unei ↑[Glu]
à jeun:
• ↑[Glucagon] şi ↓[insulină]→ stimularea Fr-2,6-BP-aza, deci ↓ [Fr-
2,6-BP] intrahepatocitar cu ↓ ratei globale a glicolizei

Glicoliza – reacţii 4. Scindarea Fr-1,6- BP ↔ GA-3P + DHAP
Enzima implicată: aldolaza
• Transformarea Fr-1,6-BP în triozo-fosfaţi: dihidroxiacetonfosfat (DHAP) şi
gliceraldehid-3-fosfat (GA-3P sau GAP)
• reacţie reversibilă şi necontrolată
HO C H
H C OH
C O
H C OH
CH2 O P
CH2 O P
Fr 1,6 BP
89%
C O
CH2 OH
CH2 O P
96%
DHAP
Aldolazã
H C OH
C
O
H
CH2 O P
4%
GAP
11%TIM

Glicoliza – reacţii
5. Izomerizarea DHAP ↔ GA-3P
(Interconversiunea triozo-fosfaţilor)
Enzima implicată: triozofosfatizomeraza (TIM)
• Numai GA-3P este degradat în faza a II-a a glicolizei
• DHAP este utilizat după conversia la GA-3P
HO C H
H C OH
C O
H C OH
CH2 O P
CH2 O P
Fr 1,6 BP
89%
C O
CH2 OH
CH2 O P
96%
DHAP
Aldolazã
H C OH
C
O
H
CH2 O P
4%
GAP
11%TIM

Glicoliza – reacţii • Ultimele 5 reacţii (etapa producătoare
de energie)
– Formează Py (sau lactat în anaerobioză)
– Generează 2 ATP pt. fiecare moleculă GA-3P
6. Oxidarea GA-3P ↔ 1,3-BPG (1,3 bifosfoglicerat)
7. Sinteza 3- PG (transferul grupării fosfat pe
ADP cu sinteza de ATP)
8. Izomerizarea 3- PG ↔ 2-PG (translocarea
fosfatului de pe C 3 pe C 2)
9. Deshidratarea 2- PG ↔ PEP (fosfoenolpiruvat)
10. Sinteza Py (transferul fosfatului de pe
PEP pe ADP cu formare de ATP)

Glicoliza – reacţii
• Enzima implicată:
GAPDH
(gliceroaldehidfosfat
dehidrogenaza)
• prima reacţie de
oxidoreducere a
glicolizei →
formarea NADH, H+
• Reacţia – mai multe
etape
6. Oxidarea GA-3P ↔ 1,3-BPG
H C OH
C
O
H
CH2 O P
GAP
HS-Enz-NAD+
H C OH
C
OH
H
CH2 O P
ES I
S Enz NAD+
H C OH
C
CH2 O P
S Enz NADH,H+
ES II
(contine o legãturã )
O
GAPDH
H C OH
C
CH2 O P
ES III
S Enz NAD+ O NAD+
NADH,H+
H C OH
C O P
CH2 O P
Acid 1,3-bisfosfogliceric
(Acid 1,3-BPG)
O
HS-Enz-NAD+
Pa
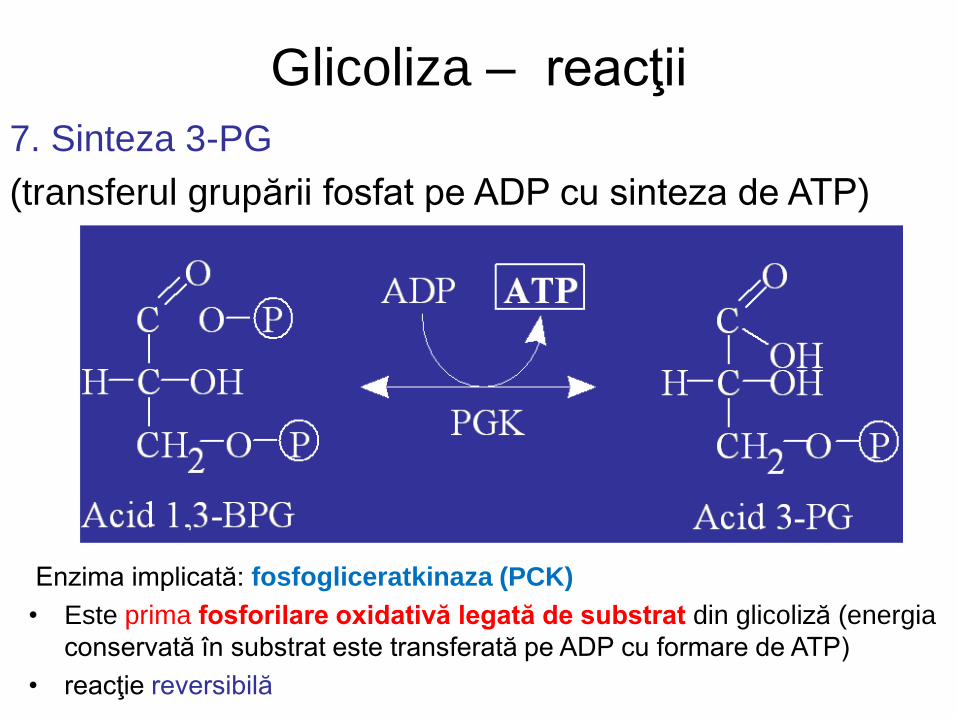
Glicoliza – reacţii
7. Sinteza 3-PG
(transferul grupării fosfat pe ADP cu sinteza de ATP)
Enzima implicată: fosfogliceratkinaza (PCK)
• Este prima fosforilare oxidativă legată de substrat din glicoliză (energia
conservată în substrat este transferată pe ADP cu formare de ATP)
• reacţie reversibilă

Ciclu Rapaport-Leubering (şuntul 2,3-DPG)
• Serveşte ca mecanism de disipare
a excesului de energie produs în
hematii
• 1,3-DPG ↔ 2,3-DPG (conţine
grupare fosfat cu energie mai mică)
• 2,3 DPG ↔ 3-PG (poate continua
reacţiile glicolizei)
• Importanţa 2,3-DPG:
- modulator al afinităţii Hb pentru O2 (se
leagă de deoxiHb, împiedică legarea
O2 de Hb, deci previne hipoxia tisulară
prin disponibilitatea crescută de O2
pentru ţesuturi)
!! 2,3-DPG scade
afinitatea Hb
pentru O2
(stabilizează
starea T),
favorizând
oxigenarea
țesuturilor

Ciclu Rapaport-Leubering (şuntul 2,3-DPG)
• Aplicaţie medicală:
- în deficienţa de PK (piruvatkinază) → ↑ nivel 2,3-DPG
(afinitatea Hb faţă de O2 < decât în mod normal; → o
mai bună oxigenare a ţesuturilor).
- în deficitul HK (hexokinază) → ↓ nivel 2,3-DPG cu
70% din normal (afinitatea Hb faţă de O2 > decât în
mod normal; → eliberare mai dificilă a O2 către
ţesuturi
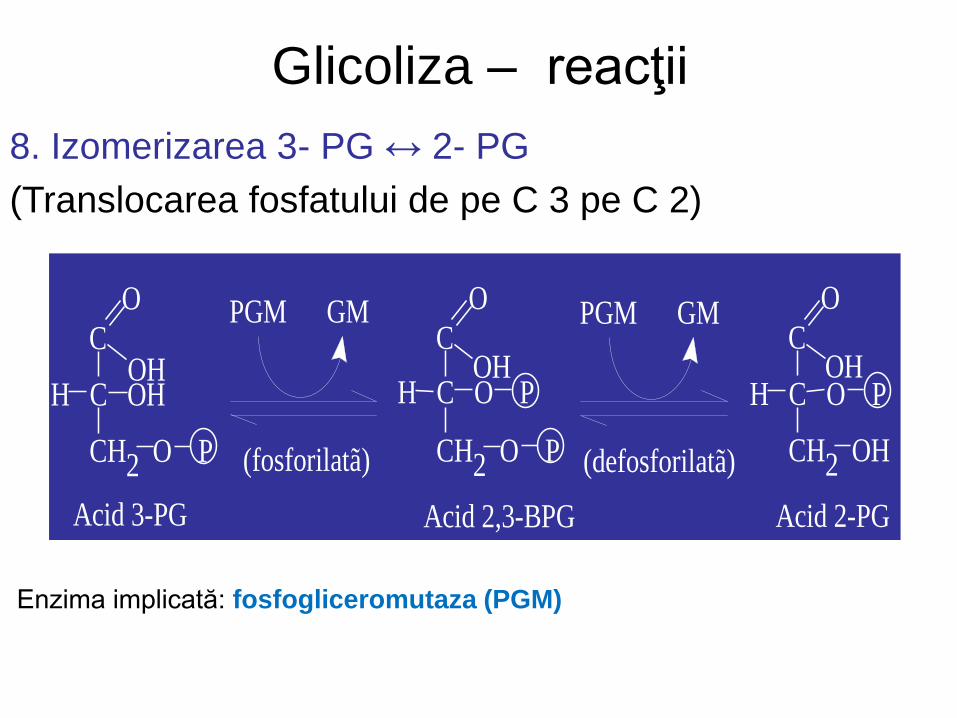
Glicoliza – reacţii
8. Izomerizarea 3- PG ↔ 2- PG
(Translocarea fosfatului de pe C 3 pe C 2)
Enzima implicată: fosfogliceromutaza (PGM)
H C OH
C
O
OH
CH2 O P
Acid 3-PG
H C O P
C
O
OH
CH2 O P
Acid 2,3-BPG
PGM GM
(fosforilatã)
PGM GM
(defosforilatã)
H C O P
C
O
OH
CH2 OH
Acid 2-PG
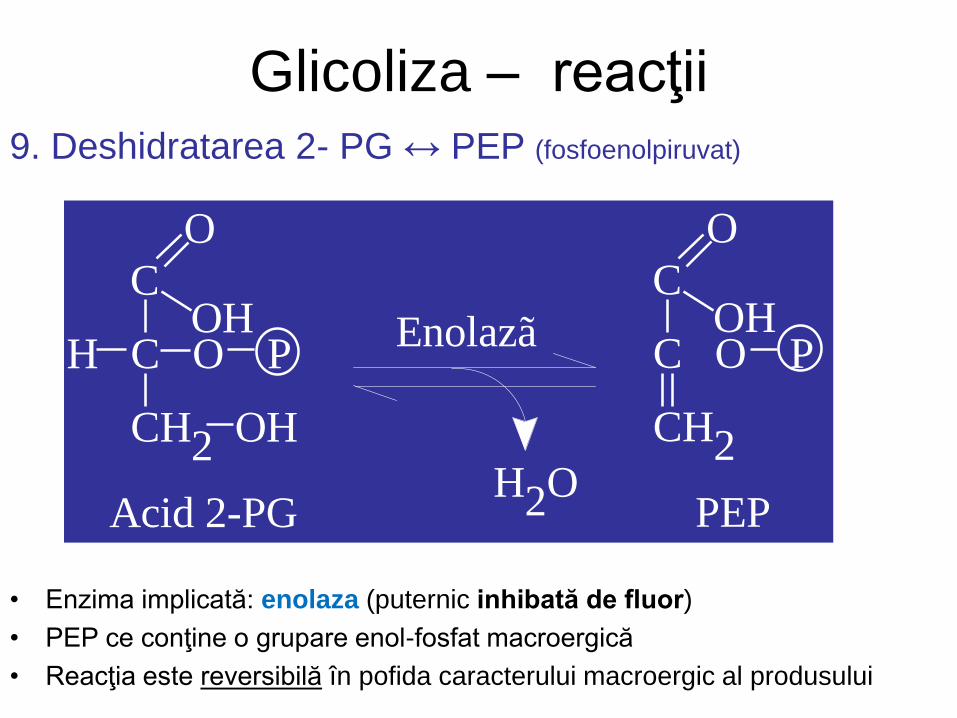
Glicoliza – reacţii 9. Deshidratarea 2- PG ↔ PEP (fosfoenolpiruvat)
• Enzima implicată: enolaza (puternic inhibată de fluor)
• PEP ce conţine o grupare enol-fosfat macroergică
• Reacţia este reversibilă în pofida caracterului macroergic al produsului
H C O P
C
O
OH
CH2 OH
Acid 2-PG
C O P
C
O
OH
CH2
PEP
Enolazã
H2O
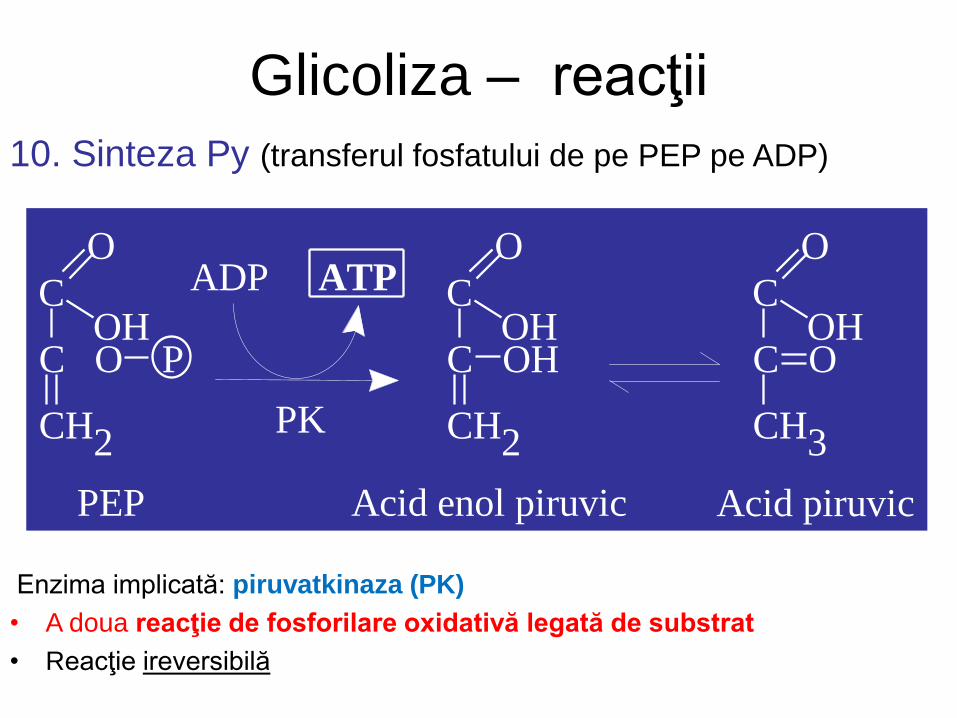
Glicoliza – reacţii
10. Sinteza Py (transferul fosfatului de pe PEP pe ADP)
Enzima implicată: piruvatkinaza (PK)
• A doua reacţie de fosforilare oxidativă legată de substrat
• Reacţie ireversibilă
C O P
C
O
OH
CH2
PEP
PK
ADP ATP
C OH
C
O
OH
CH2
Acid enol piruvic
C O
C
O
OH
CH3
Acid piruvic
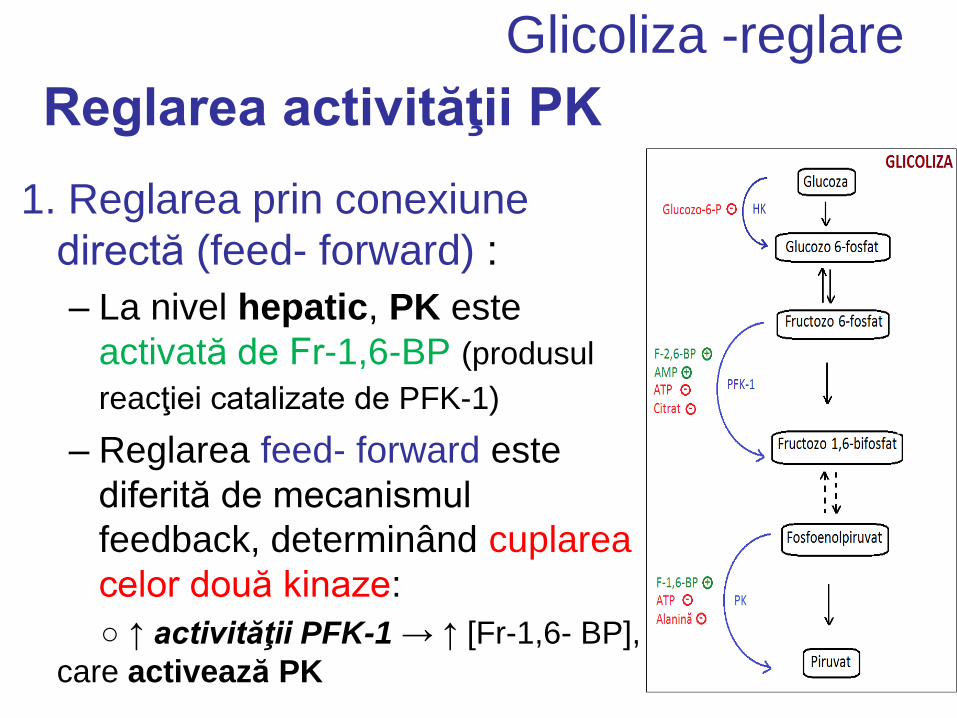
Reglarea activităţii PK
1. Reglarea prin conexiune
directă (feed- forward) :
– La nivel hepatic, PK este
activată de Fr-1,6-BP (produsul
reacţiei catalizate de PFK-1)
– Reglarea feed- forward este
diferită de mecanismul
feedback, determinând cuplarea
celor două kinaze:
○ ↑ activităţii PFK-1 → ↑ [Fr-1,6- BP],
care activează PK
Glicoliza -reglare
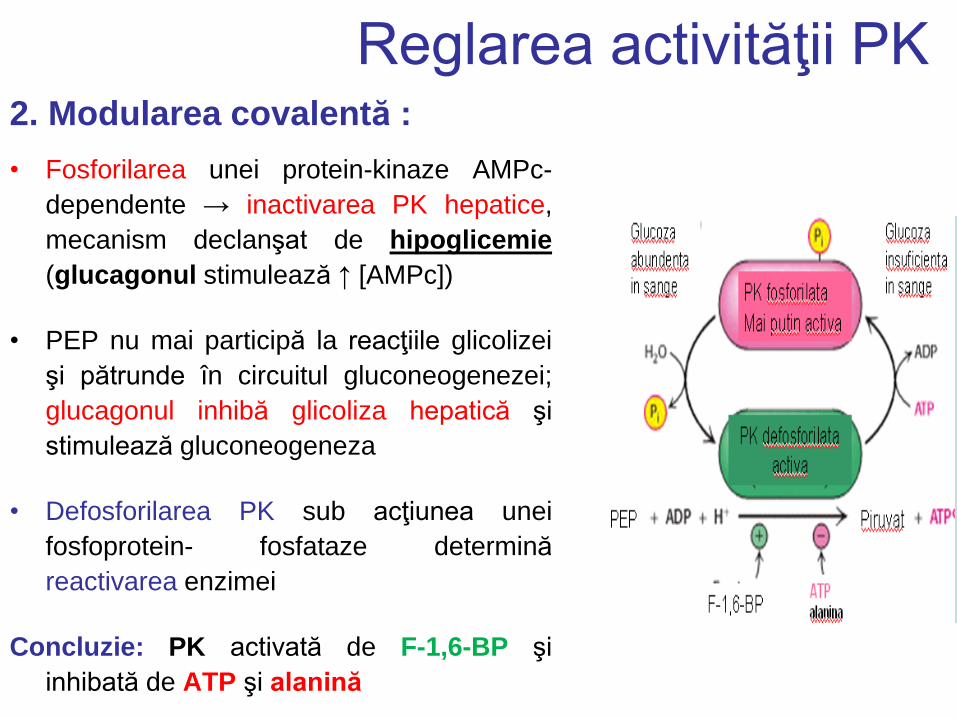
Reglarea activităţii PK 2. Modularea covalentă :
• Fosforilarea unei protein-kinaze AMPc-
dependente → inactivarea PK hepatice,
mecanism declanşat de hipoglicemie
(glucagonul stimulează ↑ [AMPc])
• PEP nu mai participă la reacţiile glicolizei
şi pătrunde în circuitul gluconeogenezei;
glucagonul inhibă glicoliza hepatică şi
stimulează gluconeogeneza
• Defosforilarea PK sub acţiunea unei
fosfoprotein- fosfataze determină
reactivarea enzimei
Concluzie: PK activată de F-1,6-BP şi
inhibată de ATP şi alanină

Deficienţă congenitală (AR) care conduce
la anemie hemolitică cronică
• ↓ numărul eritrocitelor datorită scăderii
aportului energetic;
• eritrocitele nu îşi menţin integritatea
peretelui celular datorită dezechilibrului
ionic → hemoliza celulară.
• Eritrocitele devin rigide şi deformate
• Există o marcată reticulocitoză şi
poikilocitoză
Deficienţa de PK
• Manifestări clinice: icter la nou-născuţi, splenomegalie
• Tratament: transfuzii de sange şi splenectomie;
• ! Poate apărea litiază biliară şi hemosideroză de la transfuzii multiple
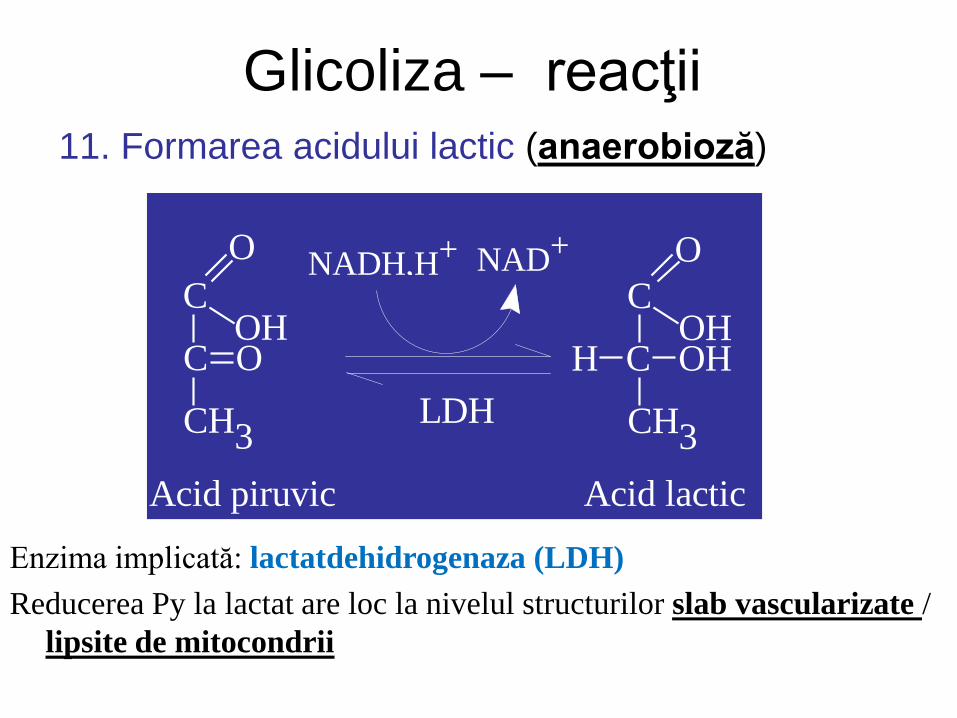
Glicoliza – reacţii 11. Formarea acidului lactic (anaerobioză)
Enzima implicată: lactatdehidrogenaza (LDH)
Reducerea Py la lactat are loc la nivelul structurilor slab vascularizate /
lipsite de mitocondrii
C O
C
O
OH
CH3
Acid piruvic
NAD+NADH,H+
LDH
H C OH
C
O
OH
CH3
Acid lactic

Glicoliza – reglare
La nivelul enzimelor reglatoare = alosterice
– Hexokinaza (HK), glucokinaza (GK)
– Fosfofructokinaza1 (PFK1)
– Piruvat kinaza (PK)
• PFK este considerată peacemaker-ul glicolizei.
• De ce PFK este mai importantă decât HK?
– Pe lângă glicoliză, G-6-P este intermediar şi al altor căi metabolice, de
aceea inhibarea HK va conduce la inhibarea şi a celorlalte căi.
– Prima reacţie ireversibilă caracteristică glicolizei este reacţia
catalizată de PFK. Deci, alterarea activităţii PFK va influenţa
activitatea doar a unei căi metabolice (glicoliza).

Glicoliza – reglare Modalităţi de reglare:
Pe perioade scurte (min→ore):
• Activare / inhibiţie alosterică (vezi slide-uri
anterioare pentru reglarea HK/GK, PFK, PK)
• Fosforilarea / defosforilarea enzimelor
limitante de viteză (vezi slide-uri anterioare
pentru reglarea HK/GK, PFK, PK)
Pe perioade mai lungi (ore →zile):
• Hormonal: insulina, glucagon (produc ↑
de 20-30 ori a activităţii enzimatice)
• Insulina sau mesele bogate în glucide
activează transcripţia genică precum şi ↑
[GK], [PFK] şi [PK] la nivel hepatic cu
activarea glicolizei
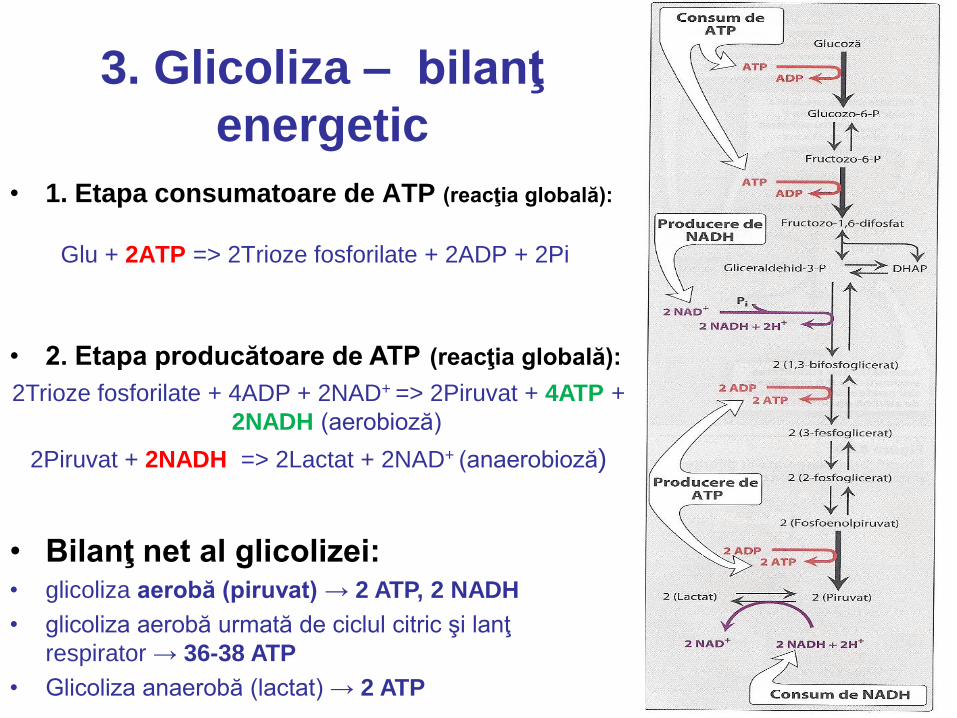
3. Glicoliza – bilanţ
energetic
• 1. Etapa consumatoare de ATP (reacţia globală):
Glu + 2ATP => 2Trioze fosforilate + 2ADP + 2Pi
• 2. Etapa producătoare de ATP (reacţia globală): 2Trioze fosforilate + 4ADP + 2NAD+ => 2Piruvat + 4ATP +
2NADH (aerobioză)
2Piruvat + 2NADH => 2Lactat + 2NAD+ (anaerobioză)
• Bilanţ net al glicolizei: • glicoliza aerobă (piruvat) → 2 ATP, 2 NADH
• glicoliza aerobă urmată de ciclul citric şi lanţ
respirator → 36-38 ATP
• Glicoliza anaerobă (lactat) → 2 ATP
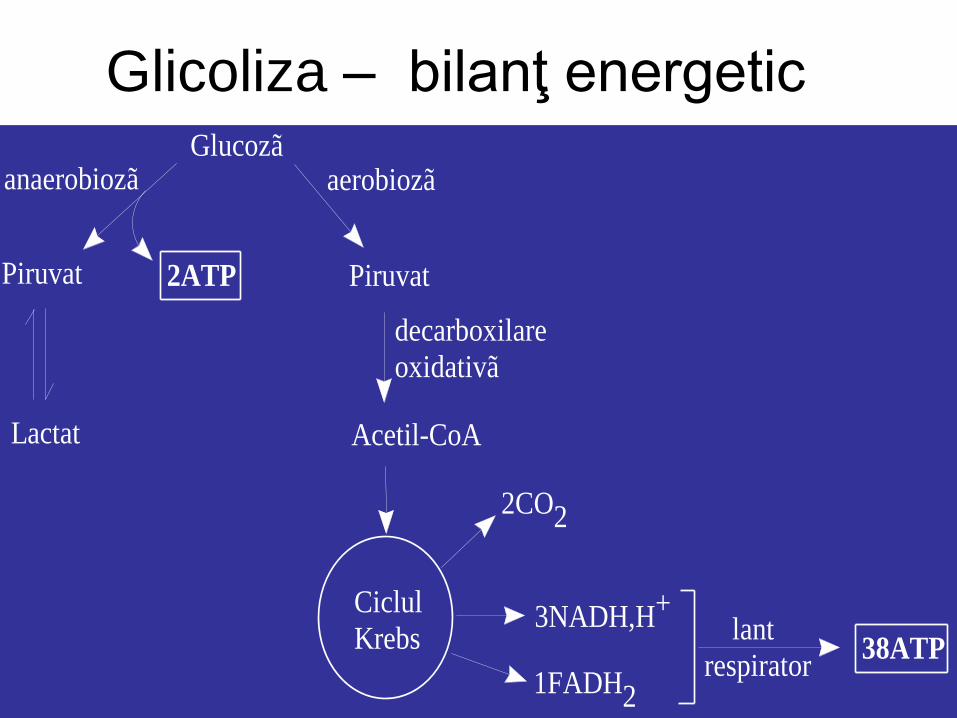
Glicoliza – bilanţ energetic Glucozã
2ATP
anaerobiozã aerobiozã
Piruvat Piruvat
Lactat Acetil-CoA
decarboxilare
oxidativã
Ciclul
Krebs3NADH,H+
1FADH2
2CO2
38ATP lant
respirator
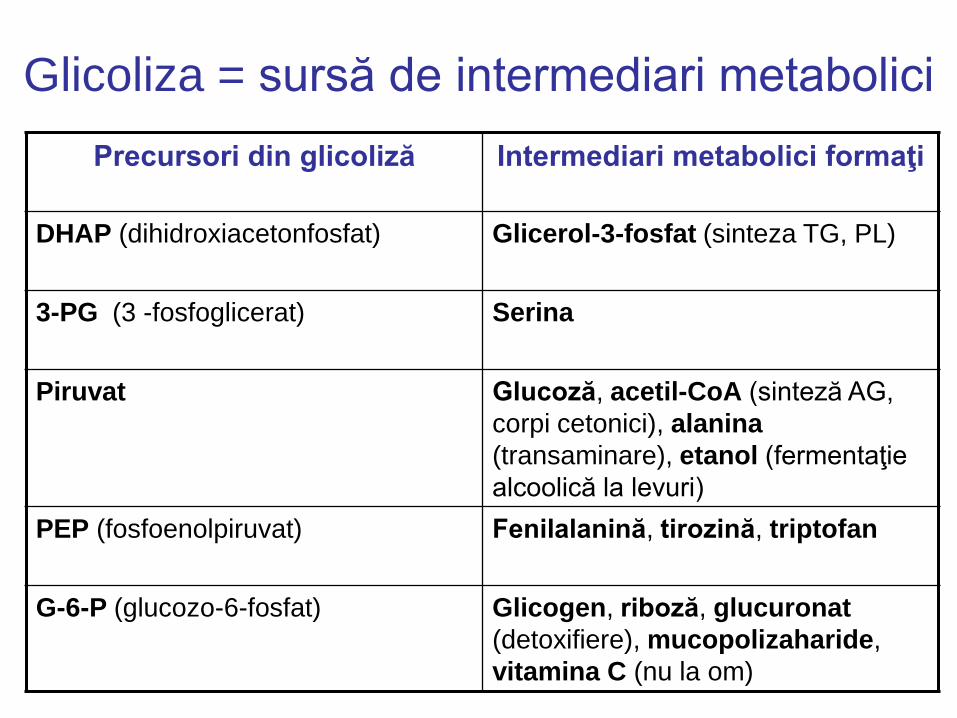
Glicoliza = sursă de intermediari metabolici
Precursori din glicoliză Intermediari metabolici formaţi
DHAP (dihidroxiacetonfosfat) Glicerol-3-fosfat (sinteza TG, PL)
3-PG (3 -fosfoglicerat) Serina
Piruvat Glucoză, acetil-CoA (sinteză AG,
corpi cetonici), alanina
(transaminare), etanol (fermentaţie
alcoolică la levuri)
PEP (fosfoenolpiruvat) Fenilalanină, tirozină, triptofan
G-6-P (glucozo-6-fosfat) Glicogen, riboză, glucuronat
(detoxifiere), mucopolizaharide,
vitamina C (nu la om)
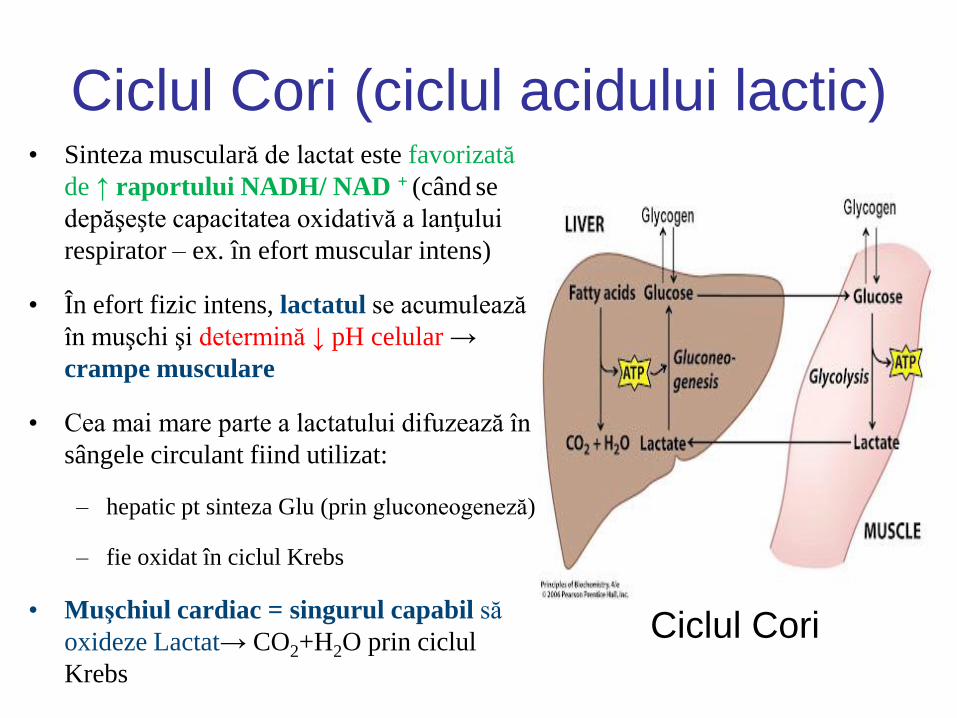
Ciclul Cori (ciclul acidului lactic) • Sinteza musculară de lactat este favorizată
de ↑ raportului NADH/ NAD + (când se
depăşeşte capacitatea oxidativă a lanţului
respirator – ex. în efort muscular intens)
• În efort fizic intens, lactatul se acumulează
în muşchi şi determină ↓ pH celular →
crampe musculare
• Cea mai mare parte a lactatului difuzează în
sângele circulant fiind utilizat:
– hepatic pt sinteza Glu (prin gluconeogeneză)
– fie oxidat în ciclul Krebs
• Muşchiul cardiac = singurul capabil să
oxideze Lactat→ CO2+H2O prin ciclul
Krebs
Ciclul Cori

Acidoza lactică
= ↑[lactat] ce însoţeşte colapsul circulator din
○ infarctul miocardic
○ embolia pulmonară
○ hemoragiile necontrolate
○ şoc
• Excesul de oxigen necesar recuperării după o perioadă în
care aportul de oxigen a fost inadecvat = "datorie de oxigen"
• Lactacidemia permite depistarea rapidă a datoriei de oxigen
şi permite monitorizarea recuperării pacientului cu şoc

Căi alternative ale piruvatului
A. Decarboxilarea oxidativă cu formare de acetil-CoA
B. Reducerea la acid lactic
C. Formarea oxaloacetatului
D. Formarea alaninei
E. Formarea etanolului
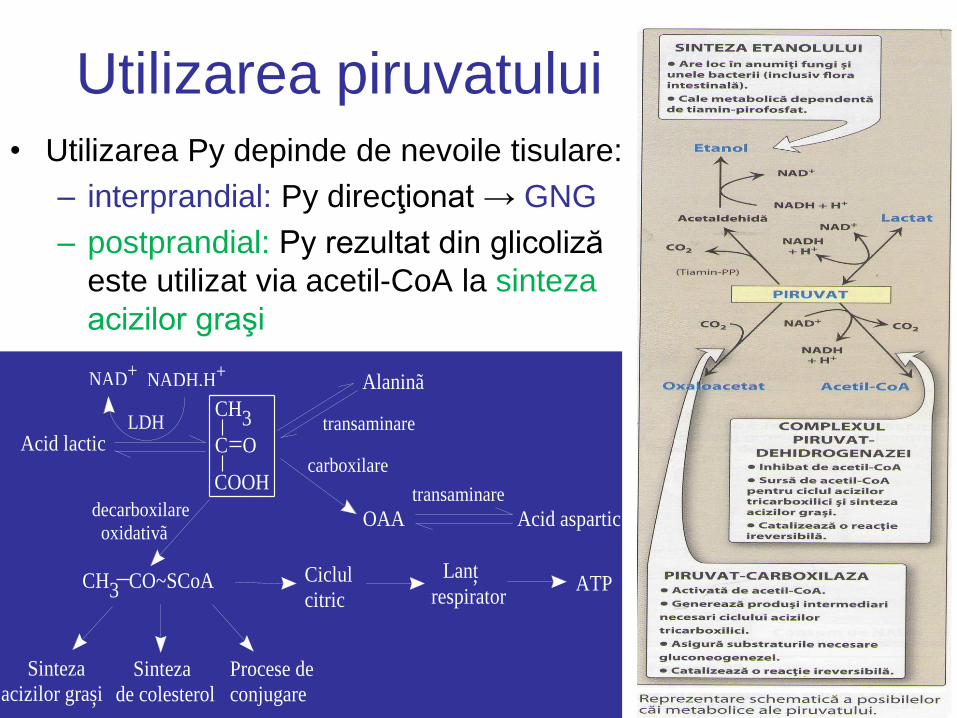
Utilizarea piruvatului • Utilizarea Py depinde de nevoile tisulare:
– interprandial: Py direcţionat → GNG
– postprandial: Py rezultat din glicoliză
este utilizat via acetil-CoA la sinteza
acizilor graşi
C O
COOH
Alaninã
CH3
Acid lactic
NAD+NADH,H+
LDH
OAA Acid aspartictransaminare
transaminare
carboxilare
decarboxilare
oxidativã
CH3 CO~SCoA
Sinteza
acizilor grasi Sinteza
de colesterol
Procese de
conjugare
Ciclul
citric
Lant
respirator ATP,
,
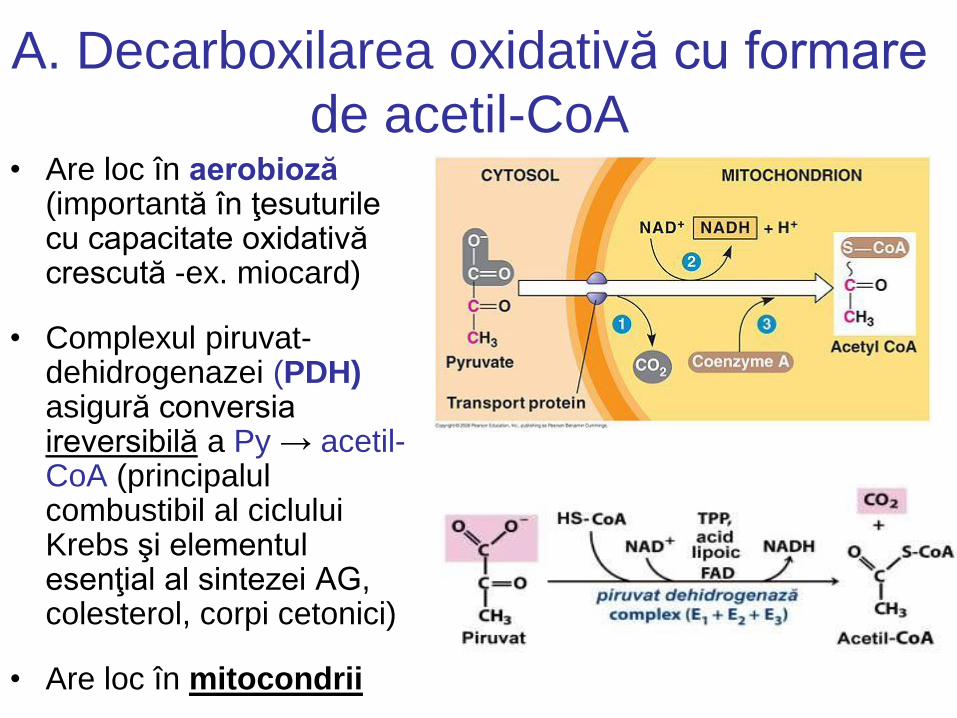
A. Decarboxilarea oxidativă cu formare
de acetil-CoA • Are loc în aerobioză
(importantă în ţesuturile cu capacitate oxidativă crescută -ex. miocard)
• Complexul piruvat- dehidrogenazei (PDH) asigură conversia ireversibilă a Py → acetil- CoA (principalul combustibil al ciclului Krebs şi elementul esenţial al sintezei AG, colesterol, corpi cetonici)
• Are loc în mitocondrii

Complexul PDH Localizare: matricea mitocondrială
Conţine:
• 3 enzime:
E1. PDH (în unele tratate piruvat decarboxilaza) are ca grupare activă (coenzimă) tiaminpirofosfatul (TPP)
E2. DHLTA (dihidrolipoil transacetilaza) cu gruparea activă acidul lipoic, necesită prezenţa CoA
E3. DHLDH (dihidrolipoil dehidrogenaza) conţine FAD (flavin adenin dinucleotidul), necesită prezenţa NAD+.
• 5 coenzime:
1. TPP (tiamin pirofosfat)=coenzima derivată din vit. B1
2. Acidul lipoic (derivat din ALA)
3. CoASH = coenzima A, derivată din acidul pantotenic (un membru al complexului de vit. B)
4. FAD = Flavin adenin dinucleotid, o coenzimă derivată din vit. B2 (riboflavina)
5. NAD = nicotinamid adenin dinucleotid, o coenzimă derivat din acidul nicotinic (un membru al complexului de vit. B)
• o proteinkinază

Reglarea complexului PDH
• formă fosforilată = formă inactivă
• formă defosforilată = formă activă
• Activarea complexului PDH este coordonată de PDH fosfataza
– stimulată de Mg2+, Ca2+ și insulină
• Pentru inactivarea complexului PDH este responsabilă PDH kinaza:
– inhibată de piruvat şi scăderea raportului ATP-ADP
– stimulată de produşii de reacţie: excesul de acetil COA şi NADH + H+, precum și de creşterea raportului ATP / ADP.

Deficitul de PDH
• Deficitul E1 (X-linkat dominant) din complexul PDH = cea mai frecventă cauză de acidoză lactică congenitală
• Blochează conversia Py→acetil-CoA, rezultând o şuntare a Py → lactat
• Acest şunt determină: – Tulburări cerebrale (SNC depinde de c. Krebs ca sursă
principală de energie, deosebit de sensibil la acidoză)
– Spasticitate musculară
– Moarte prematură
• Nu există tratament
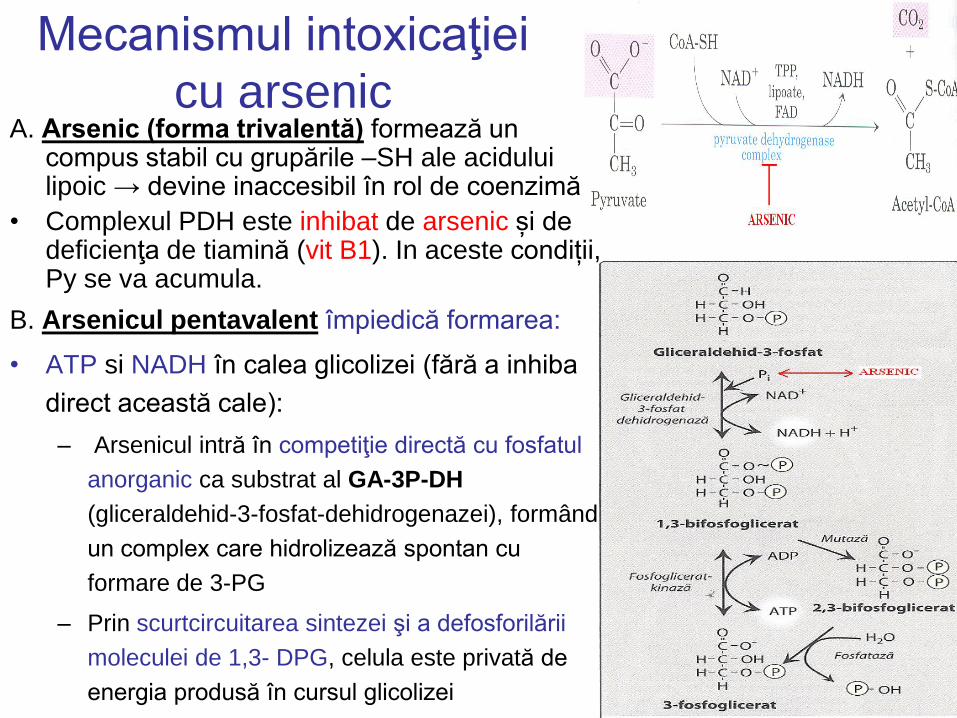
Mecanismul intoxicaţiei
cu arsenic A. Arsenic (forma trivalentă) formează un
compus stabil cu grupările –SH ale acidului lipoic → devine inaccesibil în rol de coenzimă
• Complexul PDH este inhibat de arsenic și de deficienţa de tiamină (vit B1). In aceste condiții, Py se va acumula.
B. Arsenicul pentavalent împiedică formarea:
• ATP si NADH în calea glicolizei (fără a inhiba
direct această cale):
– Arsenicul intră în competiţie directă cu fosfatul
anorganic ca substrat al GA-3P-DH
(gliceraldehid-3-fosfat-dehidrogenazei), formând
un complex care hidrolizează spontan cu
formare de 3-PG
– Prin scurtcircuitarea sintezei şi a defosforilării
moleculei de 1,3- DPG, celula este privată de
energia produsă în cursul glicolizei
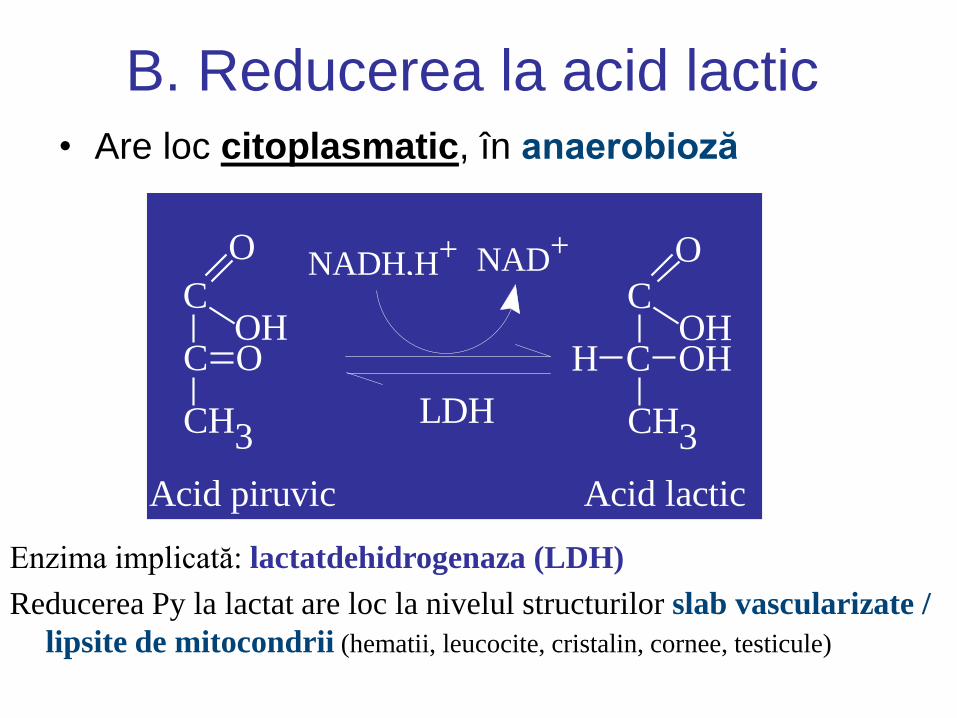
B. Reducerea la acid lactic • Are loc citoplasmatic, în anaerobioză
Enzima implicată: lactatdehidrogenaza (LDH)
Reducerea Py la lactat are loc la nivelul structurilor slab vascularizate /
lipsite de mitocondrii (hematii, leucocite, cristalin, cornee, testicule)
C O
C
O
OH
CH3
Acid piruvic
NAD+NADH,H+
LDH
H C OH
C
O
OH
CH3
Acid lactic
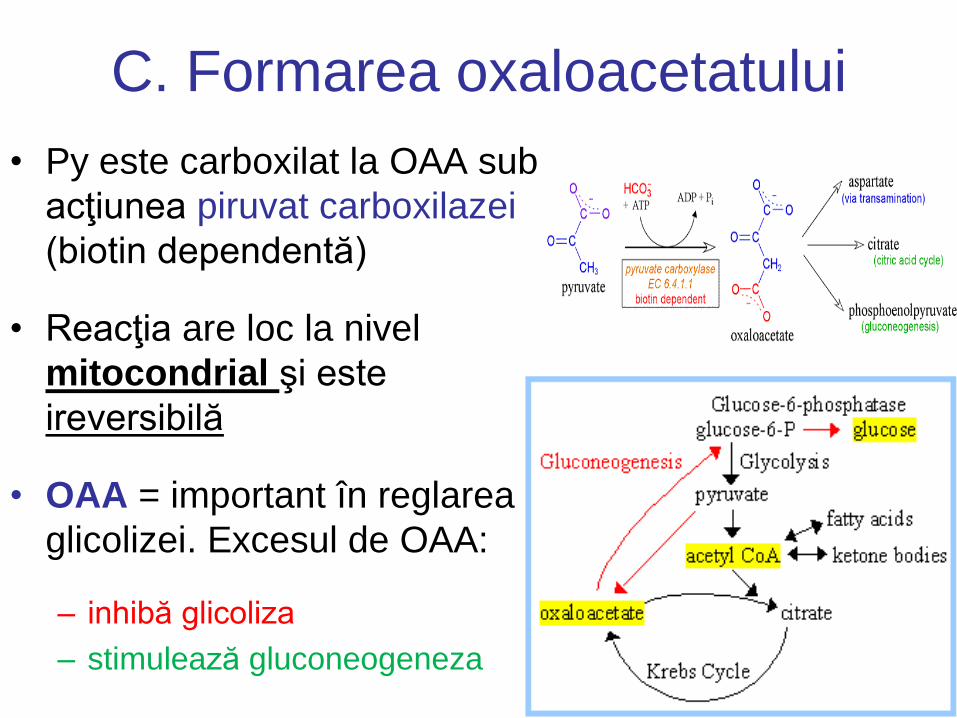
C. Formarea oxaloacetatului
• Py este carboxilat la OAA sub
acţiunea piruvat carboxilazei
(biotin dependentă)
• Reacţia are loc la nivel
mitocondrial şi este
ireversibilă
• OAA = important în reglarea
glicolizei. Excesul de OAA:
– inhibă glicoliza
– stimulează gluconeogeneza
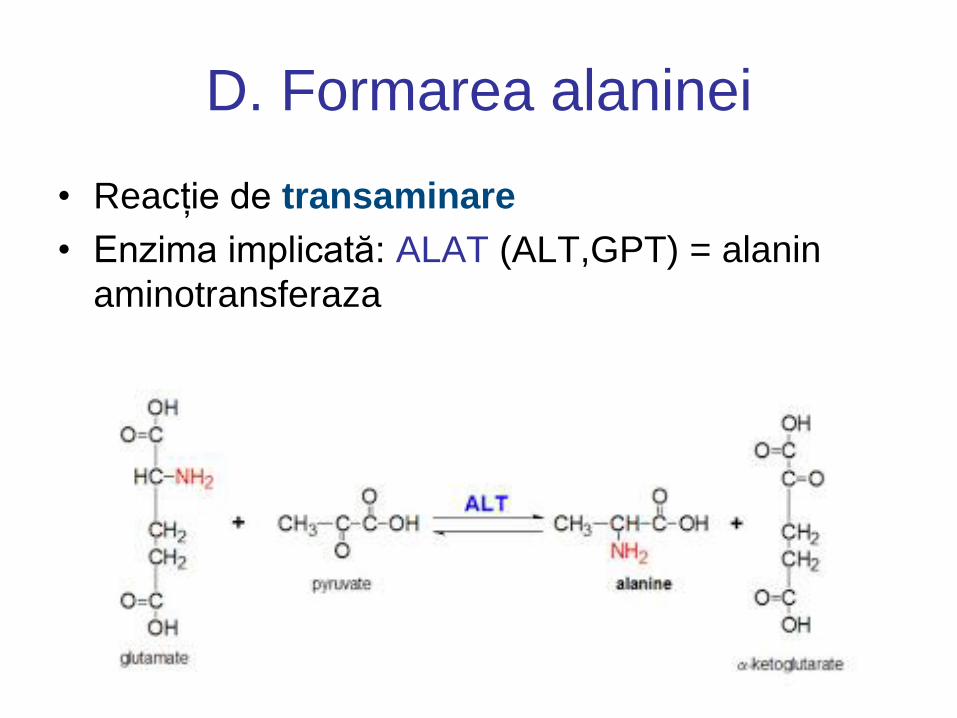
D. Formarea alaninei
• Reacție de transaminare
• Enzima implicată: ALAT (ALT,GPT) = alanin
aminotransferaza
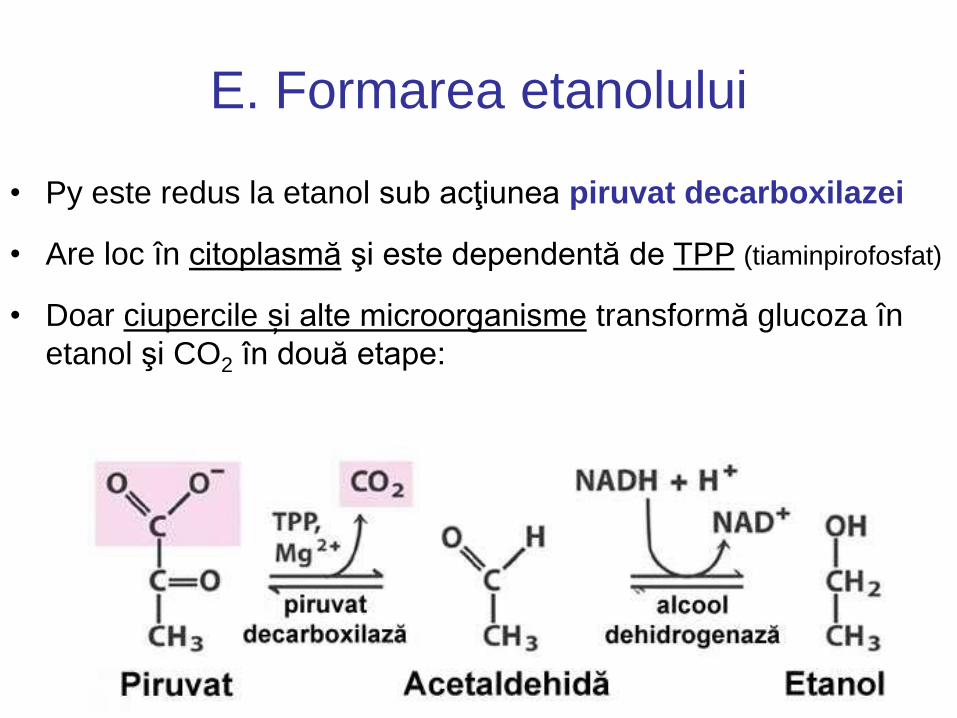
E. Formarea etanolului
• Py este redus la etanol sub acţiunea piruvat decarboxilazei
• Are loc în citoplasmă şi este dependentă de TPP (tiaminpirofosfat)
• Doar ciupercile și alte microorganisme transformă glucoza în
etanol şi CO2 în două etape:


















