CURS 14 Organizarea Genomului Uman
22
http://dgilber2.files.wordpress.com/2008/02/human-genome.jpg Organizarea genomului uman
description
UMF Cluj
Transcript of CURS 14 Organizarea Genomului Uman

http://dgilber2.files.wordpress.com/2008/02/human-genome.jpg
Organizarea genomului uman

Organizarea genomului uman
GENOM – totalitatea informaţiei genetice din celulă.
Genom nuclear – 99,995% din genomul unei celule
Genom extranuclear (ADN mitocondrial sau plastidial) – 0,005%
Genomul uman nuclear este distribuit în cei 24 de tipuri de cromozomi din nucleu.
Nucleul celulelor somatice conţine 22 de perechi de autozomi şi 2 cromozomi sexuali (XX sau XY).
Cromozomii sunt clasificaţi în dependenţă de mărimea lor, poziţia centromerului, comform modelului benzilor rezultate în urma diferitor tipuri de colorare a cromozomilor.
Caracteristica cea mai importantă a genomului uman este prezenţa ADN necodant, acesta ocupând peste 90%.

Organizarea genomului uman
AD�mt
ADNmt deţine doar o mică parte din informaţia genetică din celulă – 0,005%.
În genomul mitocondrial sunt: 2 gene pentru ARNr, 22 gene pentru ARNt şi 13 gene care codifică polipeptide, care nu sunt suficiente pentru funcţionarea independentă a mitocondriei. Marea majoritate a genelor ce codifică enzime necesare funcţionării mitocondriei se află în genomul nuclear, acestea sunt transcrise în nucleu, traduse în citoplazmă şi apoi importate în mitocondrie.
Genomul mitocondrial (secvenţa stabilită de Anderson şi col, 1981) are 16 569pb, este alcătuit dintr-o singură moleculă circulară de ADN şi constă 44% din G+C.
Cele două catene ale ADNmt:
- Catena grea (heavy H), bogată în G
- Catena uşoară (light L) bogată în C

http://a248.e.akamai.net/7/248/430/20080327144032/www.mercksource.com/ppdocs/us/common/dorlands/dorland/images/map_gene%20m.(1).jpg
Organizarea genomului uman

http://www.nature.com/nrg/journal/v6/n5/images/nrg1606-f3.jpg
Organizarea genomului uman
Heteroplasmie – prezenţa mai multor tipuri de genomuri mitocondraile într-o celulă.

Organizarea genomului uman
ADNmt are o porţiune triplucatenară – regiunea 7S, cauza este replicarea dublă a unei porţiuni din catena H.
ADNmt reprezintă 1/8000 dintr-un cromozom de măriem medie, dar ADNmt din toate mitocondriile unei celule somatice poate alcătui 0,5% din ADN total al celulei.
Spermatozoizii contribuie cu 50% la formarea genomul nuclear al zigotului, dar nu şi la alcătuirea genomului mitocondrial. Zigotul moşteneşte mitocondriile doar din ovul, deci exclusiv pe cale maternă. Atât bărbaţii cât şi femeile moştenesc mitocondriile de la mamă, dar bărbaţii nu participă la transmiterea acestora urmaşilor. Mitocondriile sunt împărţite randomic în timpul diviziunilor mitotice.
Genomul mitocondrial conţine 37 gene, dintre care 28 sunt codificate de catena H şi 9 de cea L. 24 de gene din cele 37 codifică ARN funcţional (ARN transcris care nu este tradus).
Genele mitocondriale pentru ARNr:
-ARNr 16S şi ARNr 12S

Organizarea genomului uman
13 gene mitocondriale ce codifică polipeptide sunt transcrise şi ARNm este tradus de ribosomii mitocondriali. Proteinele rezultate fac parte din complexul respirator, participă la producerea de ATP. În sistemul oxidativ de fosforilare al mitocondriei participă aproximativ 100 de proteine, deci marea majoritate a genelor codificatoare se află în nucleu.
Pentru traducerea ARNm, mitocondria îşi poate asigura ARNt şi ARNr, iar celelalte componente sunt importate din citoplazmă (proteinele din ribosomi, amino-acil ARNt sintetazele etc).
Cei 22 de tipuri de ARNt sunt capabili să recunoască toţi codonii din ARNm mitocondrial, deci unii dintre ei recunosc mai multi codoni.
ARNt:
-8 tipuri de ARNt recunosc familii de codoni alcătuiţi din 4 codoni care diferă numai prin al 3-lea nucleotid
-14 tipuri de ARNt recunosc perechi de codoni, care au primele două nucleotide identice, iar a 3-ea este fie o purină fie o pirimidină
Cele 22 de tipuri de ARNt pot recunoaşte astfel 60 de codoni (8×4 + 14×2), ceilalţi 4 codoni nu codifică nimic şi acţionează ca şi codoni stop (UAG, UAA, AGA, AGG)

Organizarea genomului uman
Codul genetic al mitocondriei are câţiva codoni diferiţi de codul genetic universal:
- codonul standard stop UGA este codonul pentru Trp
- codonii standard pentru Arg, AGA, AGG sunt codoni STOP
- codonul standard AUA pentru Ile este un codon pentru Met
Codul genetic mitocondrial are 2 codoni pentru Trp şi 2 pentru Met (codul universal include câte un codon pentru fiecare aminoacid).
Pe catena H ce conţine gena pentru ATP-ază subunităţile 6 şi 8 se suprapun parţial şi au cadru diferit de citire.
ADNmt este foare compact, 93% din genom reprezintă secvenţe codificatoare. Toate genele sunt lipsite de introni, unele gene sunt separate între ele doar prin 1-2 nt. Unele gene sunt lipsite de codoni stop, ei fiind introduşi doar după transcriere.
ADNmt are 2 origini de replicare, OH şi OL pentru fiecare catenă în parte. În timpul replicării se formează o structură de tip D. Replicarea începe la una din originile de replicare şi după ce este copiată aproape toată molecula de ADNmt, începe sinteza celeilate catene de la a doua origine de replicare, formându-se o structură asemănătoare literei D.
Boli mitocondriale: neuropatii, deficienţe ale auzului, scleroză multiplă.

Organizarea genomului uman
Genele umane sunt distribuite neuniform în genom, fiecare cromozom conţine regiuni cu o densitate mai mare sau mai mică de gene. Această densitate variabilă corelează cu benzile cromozomilor şi cu conţinutul de GC.
http://www.glowm.com/resources/glowm/graphics/figures/v5/0730/002f.jpg
Bandarea cromozomilor se efectuează în metafaza mitozei când cromozomii sunt peternic condensaţi. Cromozomii sunt mai întâi trataţi cu tripsină, urmată de colorare Giemsa. Tripsina are rolul de a reduce din proteinele din cromozomi. Benzile mai intens colorate sunt bogate în A şi T. Bandarea cromozomilor contribuie la depistarea unor modificări structurale cromozomale. Colorantul Giemsa este specific pentru grupările fosfat ale ADN. Colorantul constă dintr-un amestec de albastru de metilen şi eozină.

Organizarea genomului uman
Genomul uman conţine circa 23 000 de gene codificatoare de proteine, care reprezintă 1,5%, restul fiind ADN reglator, introni, gene pentru ARN şi ADN necodant care nu are aparent o funcţie importantă.
Numărul de gene este doar de 2 ori mai mare decât cea la musculiţa de oţet, o sursă suplimentară de variabilitate proteică se datorează procesării alternative a ARNm.
Genomul uman conţine mii de gene pentru ARN funcţional: ARNr, ARNt, microARN. Pe lângă cele 22 gene pentru ARNt din mitocondrie, în genomul nuclear au fost identificate 497 gene pentru ARNt citoplasmatic şi 324 pseudogene pentru ARNt. Genele pentru ARNt se află pe toţi cromozomii cu excepţia cromozomilor 22 şi Y. Cea mai mare parte a genelor pentru ARNt sunt pe cromozomul 1 şi 6 (140). Aceste gene sunt transcrise de ARN polimeraza III.
În genomul uman există 5 regiuni ce cuprind genele pentru ARNr:13,14, 15, 21 şi 22. Genele pentru ARNr 5S sunt transcrise de ARN polimeraza III, iar celelalte de ARN polimeraza I. Genele pentru ARNr 28S, 5,8S şi 18S sunt grupate şi sunt transcrise într-o singură moleculă de ARN. Aceste gene grupate în 5 grupuri se găsesc în mai multe copii – 30-40 fiecare. Genele pentru ARNr 5S se află pe cromozomul 1 (1q41-42 ) şi sunt 200-300 copii.
Secvenţele reglatoare se află în apropierea genelor şi participă la reglarea lor.

Organizarea genomului uman
Alte componente ale genomului uman sunt:
ADN repetitiv în tandem (ADN satelit – minisateliţi şi microsateliţi)
ADN repetitiv eterogen (LINEs şi SINEs)
Transpozoni
Pseudogene
AD� satelit – secvenţe scurte nulceotidice care se repetă. În dependenţă de numărul de nucleotide din unitatea repetitivă, ADN satelit poate fi împărţit în minisateliţi sau microsateliţi.
Microsatelţii au unitatea repetitivă alcătuită doar din câteva nucleotide.

Organizarea genomului uman
Minisateliţii au unitatea repetitivă alcătuită din 10-60nt.
Există mai mult de 1000 de loci pentru minisateliţi în genomul uman.
Unii sateliţi conţin o secvenţa centrală GGGCAGGA�G, unde N este orice nucleotidă. Se presupune că aceşti minisateliţii favorizează schimbarea ADN.
Datorită poliformismului ridicat la nivelul minisateliţilor, ei au fost intens studiaţi pentru utilizarea lor în analize de înlănţuire a genelor şi în calitate de markeri genetici. S-a constatat că minisateliţii au rol în reglarea transcrierii unor gene.
Minisateliţii hipervariabili au unitatea centrală (repetitivă) alcătuită din 9-24pb şi se află in regiunile centromerice. Minisateliţii telomerici constau din 6pb repetitive. 90% din minisateliţii din genomul uman se află în regiunile subtelomerice.

Organizarea genomului uman
Microsateliţii sunt secvenţe simple repetitive (Simple Sequence Repeats SSR ), unitatea repetitivă a cărora constă din 1-6pb. Locii pentru microsateliţi sunt polimorfi şi pot fi atât nucleari cât şi mitocondriali. Microsateliţii sunt neutri, codominanţi şi sunt utilizaţi în cercetările de genetica populaţiilor.
Un exemplu de microsatelit este (CA)n, unde n este numărul de repetiţii şi variază între alele. Secvenţele repetitive pot fi alcătuite din 2, 3, 4 nucleotide, iar numărul de repetiţii poate fi de la 10 la 100.
La nivelul microsateliţilor se pot observa o rată mare a mutaţiilor, o cauză fiind alunecarea catenelor în timpul replicării.
Microsateliţii se pot întâlni în secvenţele necodificatoare, introni, transpozoni sau chiar exoni.

Organizarea genomului uman
Amprenta genetică
99,9% din genom este identic între persoane diferite. Diferenţierea persoanelor din punct de vedere genetic se poate realiza cu ajutorul amprentei genetice, alcătuite din secvenţe repetitive numeric variabile (Variable �umeric Tandem Repeats VNTRs).
Amprenta genetică umană a fost propusă de Alec Jeffreys în 1985.
În 1986 amprenta genetică a fost folosită pentru prima dată ca dovadă în instanţă, devenind în continuare o metodă sigură şi eficientă în medicina legală.
http://upload.wikimedia.org/wikipedia/commons/thumb/7/7a/Codis_profile.jpg/350px-Codis_profile.jpg
Acordul general este că cei 13 loci utilizaţi în crearea amprentei genetice umane sunt loci STR (Short Tandem Repeats) – secvenţe scurte repetitive, majoritatea lor conţin secvenţe repetitive alcătuite din 4nt.
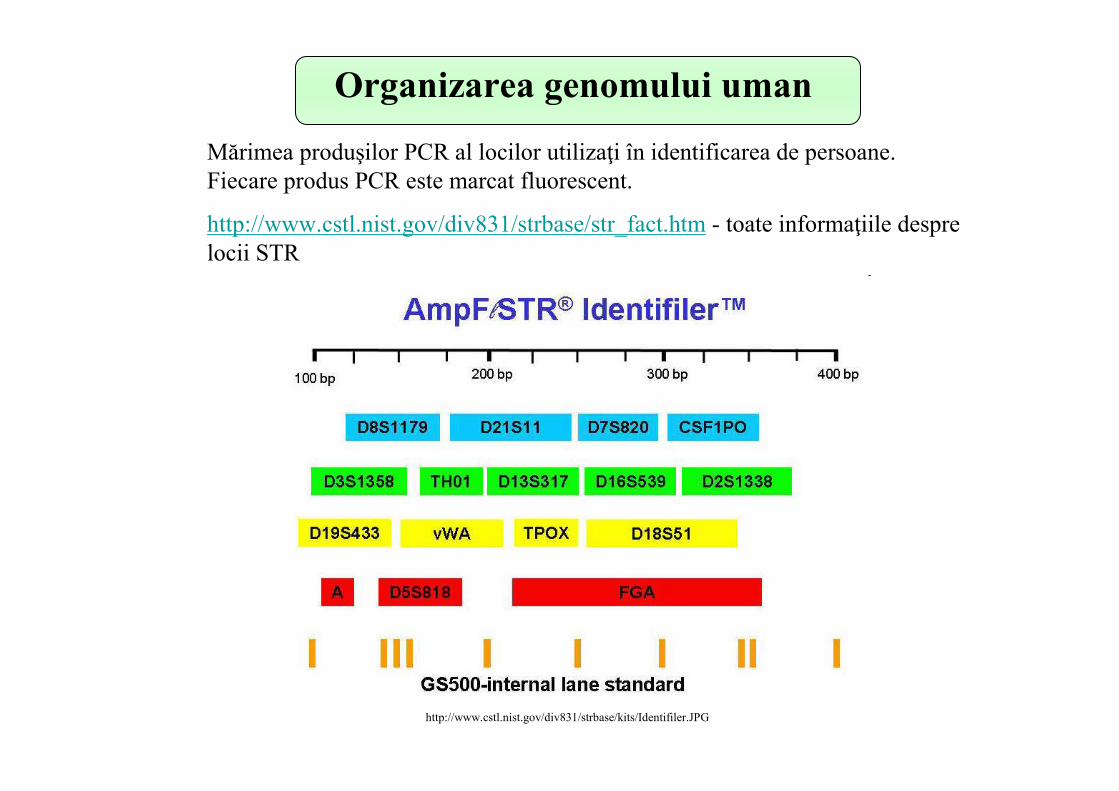
Organizarea genomului uman
http://www.cstl.nist.gov/div831/strbase/kits/Identifiler.JPG
Mărimea produşilor PCR al locilor utilizaţi în identificarea de persoane. Fiecare produs PCR este marcat fluorescent.
http://www.cstl.nist.gov/div831/strbase/str_fact.htm - toate informaţiile despre locii STR

Organizarea genomului uman
Pentru identificare umană mai sunt utilizaţi loci mitocondriali sau loci de pe cromozomul Y. Locii mitocondriali urmăresc linia maternă de moştenire, iar cei de pe Y transmiterea exclusiv paternă. Atât locii mitocondriali cât şi cei Y sunt de tip STR.
Atunci când ADN este vechi şi degradat este mai dificilă obţinerea unor amplificări pentru toţii locii nucleari. În aceste cazuri se utilizează loci STR mitocondriali (ADNmt este mai mic şi este suficientă prezenţa unei singure molecule). Locii mitocondriali sunt din regiunea HV1 şi HV2.
Astfel pentru identificarea rămăşiţelor este suficientă o cantitate mică de ADN şi este necesar un ADN de control de la urmaşii ce au moştenit acelaşi ADN mitocondrial pe linie maternă. Dificultăţile în cazul locilor mitocondriali sunt rata mare a mutaţiilor, datorită faptului că mitocondria nu dispune de un sistem reparator al ADN.
ADNmt a fost utilizat la identificarea Annei Anderson ca impostoare a ţarinei Anastasia.

Organizarea genomului uman
Secvenţe repetitive eterogene – din această clasă cele mai cunoscute elemente sunt transpozonii.
Un transpozon este o secvenţă de ADN care poate să-şi schimbe poziţia în genomul aceleliaşi celule. Transpozonii fac parte din clasa elementelor genetice mobile.
Clasa I de elemente gentice mobile cuprinde retranspozonii.
Retrotranspozonii îşi schimbă poziţia prin: transcriere, reverstranscriere şi inserare la un locus nou în genom.
Clasa II de elemente genetice mobile îşi schimbă poziţia în genom prin tăierea de la locul iniţial şi inserarea la un locus nou (mecanism de tip cut-paste). Această schimbare are loc datorită participării la transpoziţie a enzimelor numite transpozaze.

Organizarea genomului uman
Retrotranspozonii conţin gena pentru transcriptază inversă necesară transpoziţiei. Retrotranspozonii au o structură similară retrovirusurilor, cum ar fi HIV, ceea ce a condus la ipoteza despre originea acestor virusuri.
Retrotranspozonii sunt împărţiţi în 3 clase:
- de tip viral, au gena pentru transcriptază inversă, au secvenţe repetitive lungi (LTR) şi prezintă asemănări cu virusurile
- LI�Es, codifică transcriptaza inversă, sunt absente LTR, sunt transcrise de ARN polimeraza II
- de tip nonviral, nu codifică reverstranscriptaza şi sunt transcrise de ARN polimeraza III
Transpozonii ADN – diferă de retrotranspozoni prin faptul că pentru transpoziţie nu necesită transcriere. Transpozazele sunt enzimele responsabile de tăeirea ADN şi religarea lui în alt locus.
Clasa I ţi II de transpozoni pot sa-şi piardă capacitatea de transpoziţie datorită mutaţiilor, dar alte elemente genetice mobile pot produce transpozaze sau reverstranscriptaze necesare transpoziţiei.

Organizarea genomului uman
http://www.nature.com/nrg/journal/v8/n4/images/nrg2072-i1.jpg
Tipul I de transpozoni –retrotranspozonii sunt numeroşi în genomul uman datorită mecanismului de transpoziţie. Elementul non-LTR LINE1 ocupă 17% din genomul uman.
ADN transpozonii au la capete secvenţe terminale repetitive inversate (Terminal Inverted Repeats ).
Atât retrotranspozonii cât şi ADN transpozonii pot fi clasificaţi în elemente autonome sau neautonome.
Elementele neautonome sunt nişte transpozoni ancestrali, care au suferit mutaţii.

Organizarea genomului uman
Elementele neautonome în cele mai dese cazuri sunt nişte transpozoni reduşi, care constau doar din regiunile TIR terminale, care flanchează un fragment de ADN deseori copiat dintr-o regiune codificatoare. Unele elemente neautonome numite MITES (Miniature Inverted-Repeat Transposable Elements) constau doar din 2 regiuni TIR. Numărul de elemente neautonome depăşeşte numărul celor autonome. Numărul de elemente repetitive Alu este estimat la 1,2 mln în genomul uman. Elementele Alu sunt nonLTR-retrotranspozoni neautonomi.
Transpozonii pot crea mutaţii. Aceste mutaţii se datorează transpoziţiei elementelor în regiunile codificatoare ale genelor. După transpoziţia unui element în alt locus, acesta nu va fi corect reparat. Secvenţele repetitive multiple pot contribui la un crossing-over inegal în timpul meiozei (secvenţele Alu spre exemplu).
Maladii cauzate frecvent de elemente transpozabile sunt hemofilia A şi B, porfiria, predispoziţiile pentru cancer, distrofia musculară Duchenne. Unii transpozoni au propriii promotori care contribuie la transcrierea transpozazelor. Aceşti promotori pot infleunţa şi transcrierea genelor adiacente.

Organizarea genomului uman
Pseudogenele sunt gene care şi-au pierdut activitatea, nu sunt transcrise şi traduse.
Unele pseudogene nu au introni şi nici promotori şi sunt numite pseudogene procesate. Originea lor se presupune a fi prin inserarea ARNm în cromozomi.
Unele din aceste gene au caracteristici comune genelor cum ar fi promotori, situsuri de procesare, dar sunt considerate nefuncţionale din cauza absenţei unor elemente importante cum ar codonii STOP, modificarea cadrului de citire, lipsa transcrierii.
Pseudogenele sunt considerate ADN inutil. Pseudogenele oferă o informaţie foarte importantă din punct de vedere evolutiv, deoarece au o origine comună cu genele funcţionale.
Pseudogenele se caracterizează prin omologie şi nefuncţionalitate. Ele prezintă o asemănare cu unele gene funcţionale, dar sunt incapabile de a avea un produs (incapabile de transcriere).

Organizarea genomului uman
Principalele tipuri de pseudogene:
- procesate (retrotranspuse) prin intermediul retrotranspozonilor, care s-a dovedit că in vitro au proprietatea de a retrotranspune şi gene întregi. O caracteristică a acestui tip este lipsa intronilor şi prezenţa cozii poli-A. Deoarece aceste pseudogene sunt lipsite de promotor, ele devin inactive imediat dupa transpunere.
- neprocesate (sau duplicate), cauza este duplicarea genelor în cromozom, aceste gene sunt identice cu cele funcţionale – au aceeaşi structură de introni/exoni şi au promotor. Aceste gene devin nefuncţionale după achiziţia unor mutaţii.
- inerte sau unitare, diverse mutaţii pot opri transcrierea sau traducerea şi o genă poate deveni nefuncţională dacă o asemenea mutaţie se stabileşte în populaţie.
Se presupune că pseudogenele contribuie la reglarea transcrierii, siARN (small interfering RNA) este derivat din pseudogene.