
acizii nucleici
266
Acizii nucleici
-
Upload
dumitru-nuca -
Category
Documents
-
view
29 -
download
3
description
AN
Transcript of acizii nucleici

Acizii nucleici

Obiectivele:1. Tipurile de acizi nucleici, funcţiile şi repartizarea lor în celulă.2. Constituienţii acizilor nucleici; bazele azotate, pentozele, acidul fosforic.3. Nucleozidele şi nucleotidele. 3, 5- cAMP
4. Acidul Dezoxiribonucleic (ADN): structura şi funcţiile. Dublul helix şi conformaţiile lui de tip B, A, Z.
5. Nivelurile de compactizare a moleculei de ADN la procariote (nucleoidul) şi eucariote (nucleozomii, cromatina şi cromozomii).
6. Proprietăţile fizico-chimice ale ADN. Denaturarea şi renaturarea. Hibridizarea.
7. Replicarea ADN la procariote – matricea, substraturile, enzimele şi factori proteici. Mecanismul biochimic şi etapele biosintezei ADN.
8. Particularităţile replicării la eucariote. Telomerele şi telomeraza. Structura telomerazei. Rolul biomedical al telomerazei.
9. Mecanismele biochimice ale reparaţiei ADN. Enzimele implicate.
10. Mecanismele biochimice ale genezei mutaţiilor punctiforme. Rolul biomedical al mutaţiilor

Acizi nucleici Acizi nucleici – sunt polinucleotide,
alcătuite din mononucleotide, unite prin legături 3’, 5’-fosfodiesterice.
1. ADN - acidul dezoxiribonucleic; 2. ARN - acidul ribonucleic.

ADN
Localizarea: a. 97-99% - concentrat în nucleu b. 1-3% - situat în MC. Rolul: deţine, păstrează şi transmite
informaţia genetică.

ARN Localizarea: 11% - în nucleu 15% -în mitocondrii 50% - în ribosomi 24% - în hialoplasmă Deosebim:
1. ARN mesager
2. ARN ribozomal
3. ARN de transport ARN nuclear heterogen ARN nuclear cu moleculă mică

ARN mesager (mARN) Localizat -în nucleu şi citozol. prezintă copia sectorului de ADN şi conţine
informaţia despre structura catenei polipeptidice a proteinei.
Rolul:Transmite informaţia de la ADN spre ribozomi, unde o traduce în sinteza de proteine.
ARN ribozomal (rARN) constituie 60% din totalul ARN-ului. Localizat- în ribozomii citoplasmei. Rolul - formează scheletul ribozomilor. Joacă un
rol auxiliar în procesul de asamblare a proteinelor.

ARN de transport (tARN)
constituie 15% din totalul ARN-lui. Localizat: în citoplasmă, ribosomi,
mitocondrii.Rolul: participă la activarea şi
transportul AA spre ribozomi şi asamblarea lor în polipeptide.

Structura chimică a AN La hidroliză AN degradează în
mononucleotide (MN), MN la hidroliza completă degradează în
BA, pentoze şi acid fosforic. ADN----A; G; C; T + dR+ H2PO3
ARN---- A; G; C; U+ R+ H2PO3

Bazele azotateBA se clasifică în: 1. majore:a. purinice: A, G b. pirimidinice: C,T,U2. minore:a. purinice (2metil A; 1 metilG) b. pirimidinice (5 metil C;5 hidroximetil C)
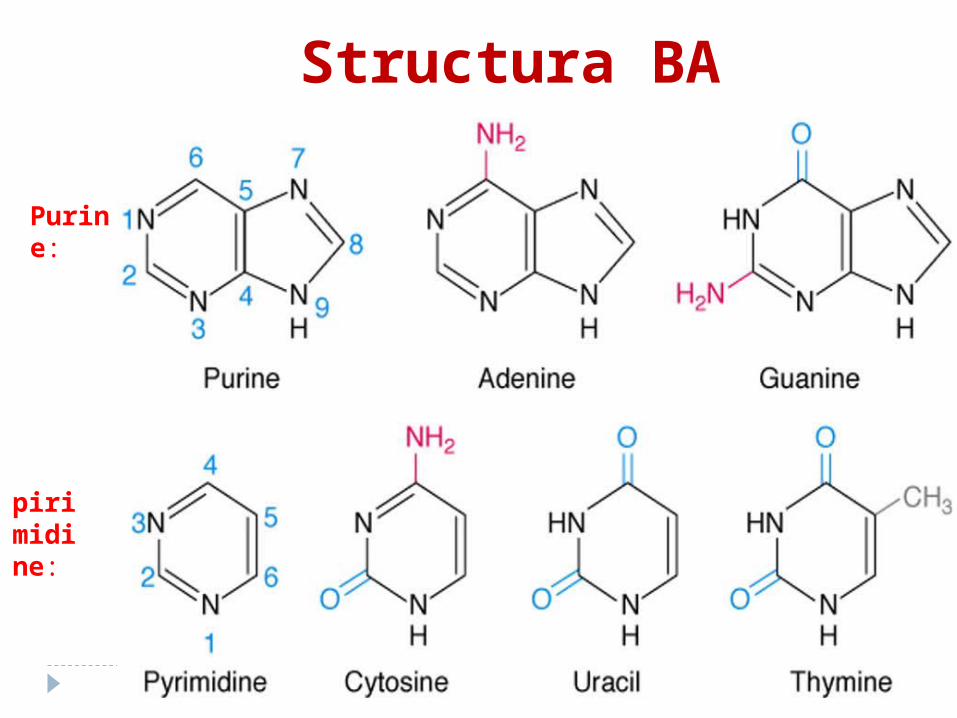
Purine:
Structura BA
pirimidine:
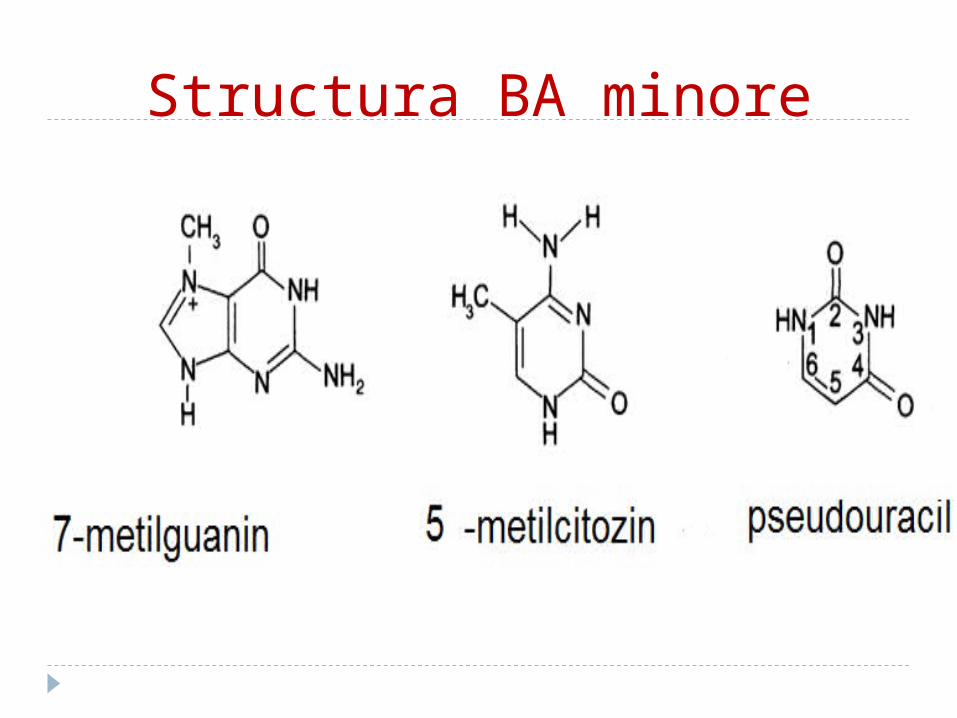
Structura BA minore

Proprietăţile BA:1. Sunt slab solubile în H2O2. BA pirimidinice prezintă fenomenul de
tautomerie: forme lactim (enol OH) –lactam (ceto C=O)
3. Sunt responsabile de informaţia genetică4. BA purinice- au structură plană; cele
pirimidinice- aproape plană, puţin plată5. max capacităţii de absorbţie în ultraviolet
este între 260-280 nm
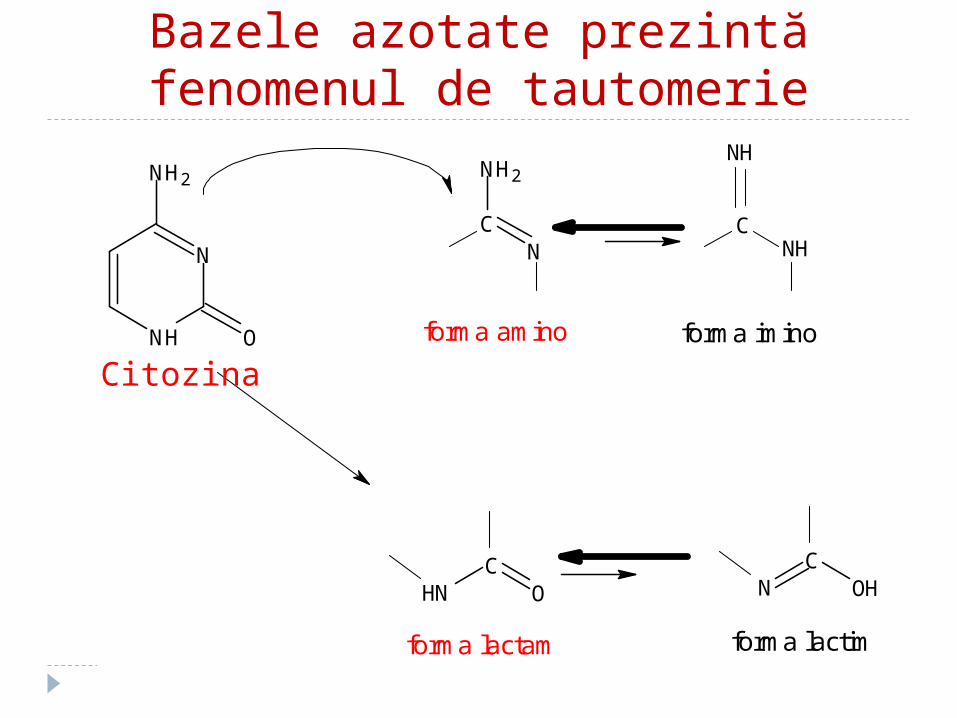
Bazele azotate prezintă fenomenul de tautomerie
N
NH
NH2
O
NC
NH2NH
CNH
forma lactam forma lactim
forma amino forma imino
CN OH
CHN O
Citozina

Bazele azotate prezintă fenonemul de tautomerie
Formele mai stabile:Forma lactam este mai stabilă decît forma lactim
Forma amino este mai stabilă decît forma imino
În acizii nucleici există formele lactam şi amino
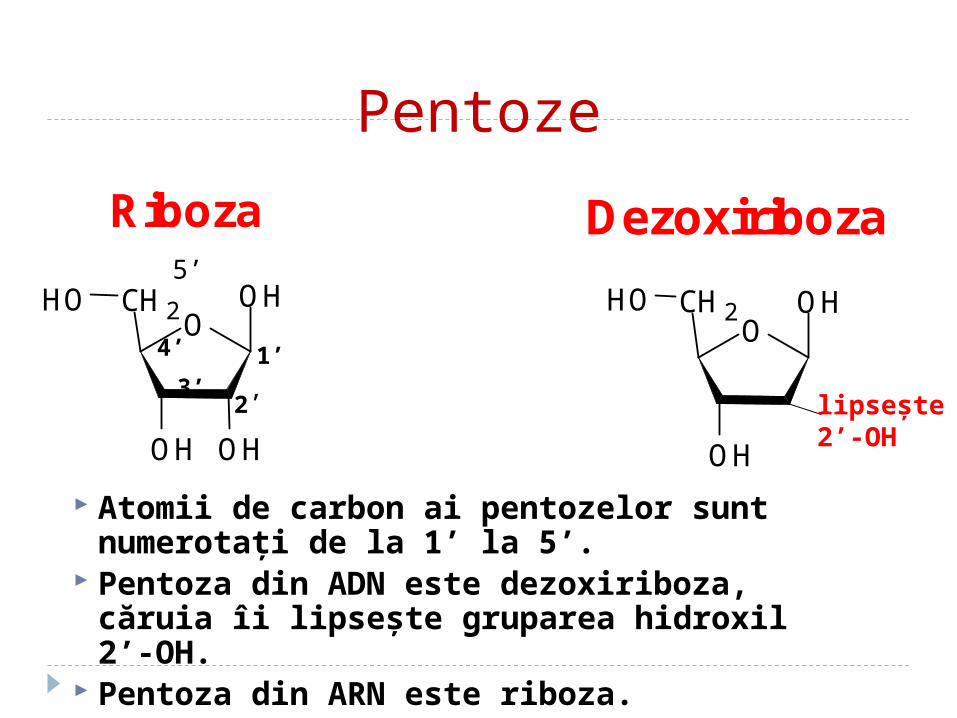
Pentoze
Atomii de carbon ai pentozelor sunt numerotaţi de la 1’ la 5’.
Pentoza din ADN este dezoxiriboza, căruia îi lipseşte gruparea hidroxil 2’-OH.
Pentoza din ARN este riboza.
OHO
CH2OH
OHOH
OHO
CH2OH
OH
Riboza Dezoxiriboza
1’
2’3’
4’
5’
lipseşte2’-OH
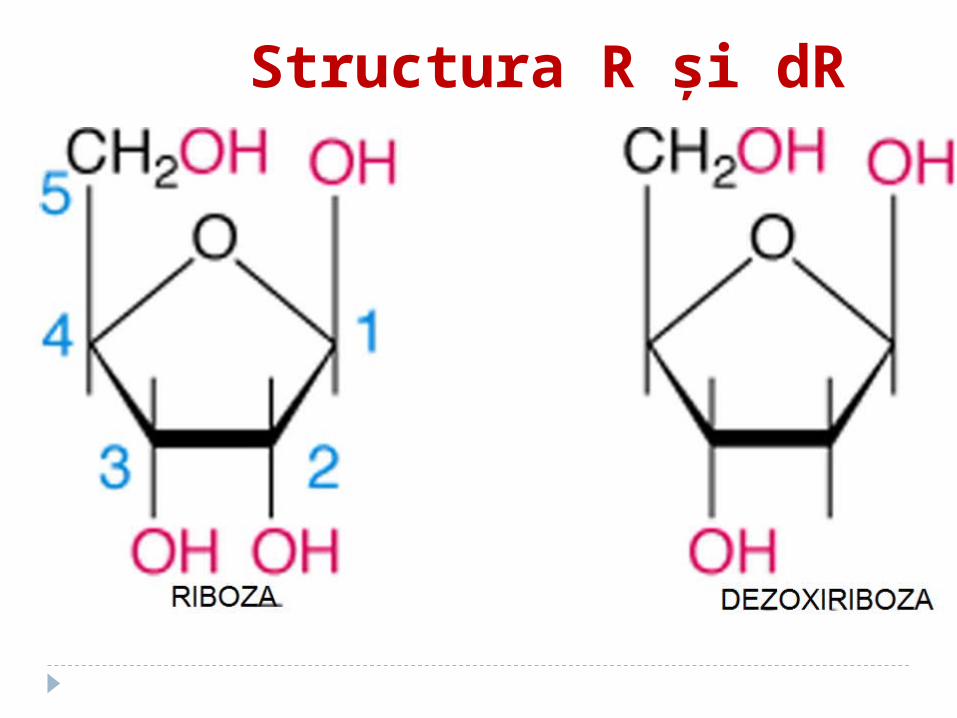
Structura R şi dR
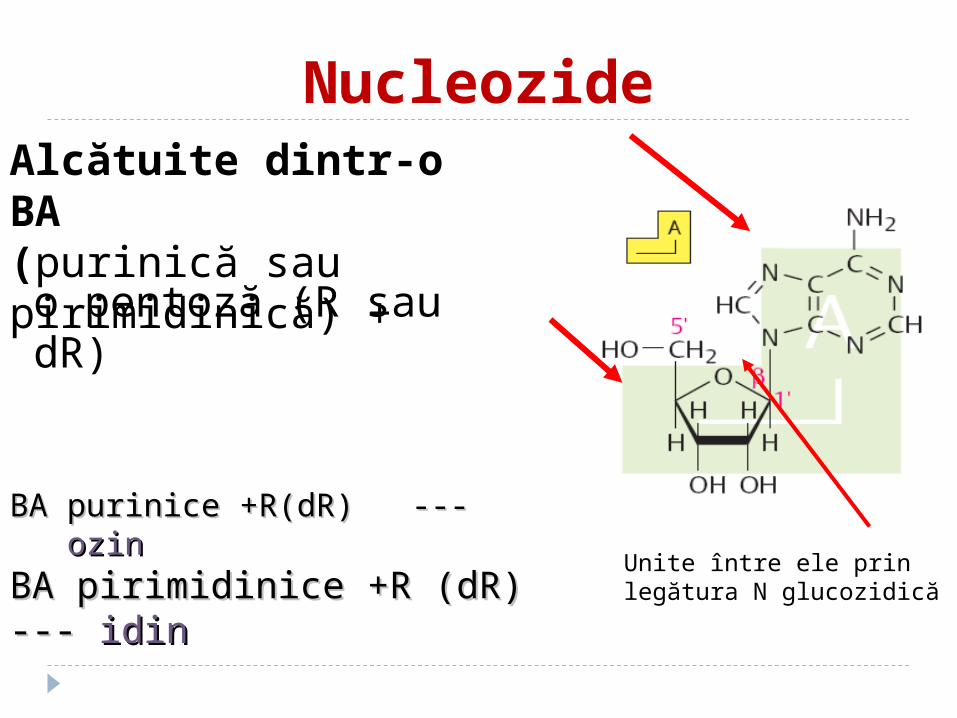
NucleozideAlcătuite dintr-o BA (purinică sau pirimidinică) +
o pentoză (R sau dR)
BA purinice +R(dR)BA purinice +R(dR) ------ ozinozin
BA pirimidinice +R (dR) --- BA pirimidinice +R (dR) --- idinidin
Unite între ele prin legătura N glucozidică
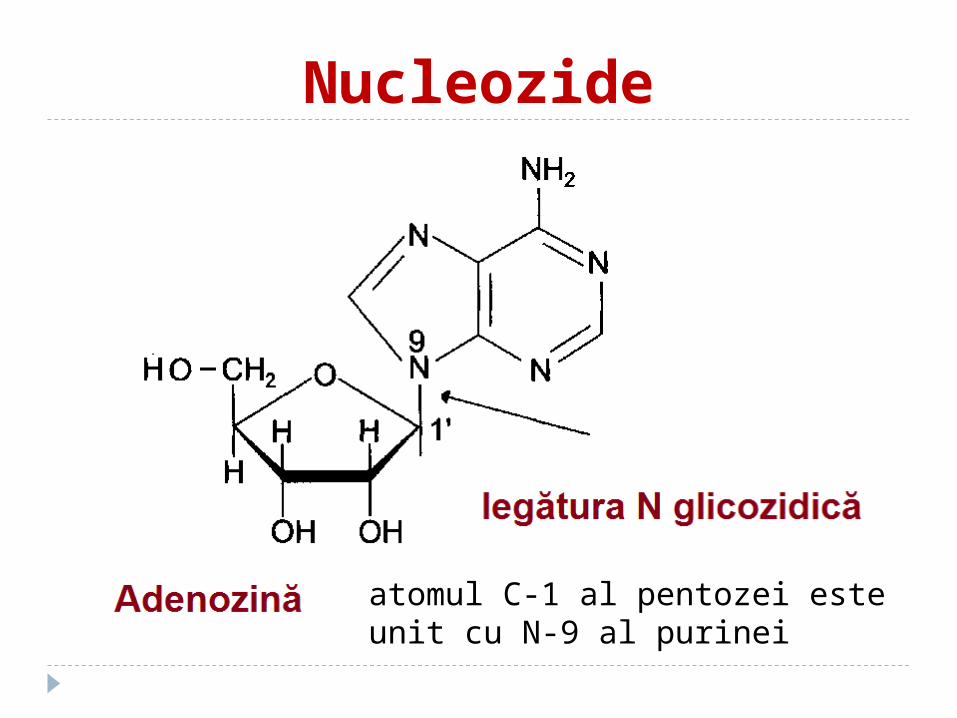
Nucleozide
atomul C-1 al pentozei este unit cu N-9 al purinei
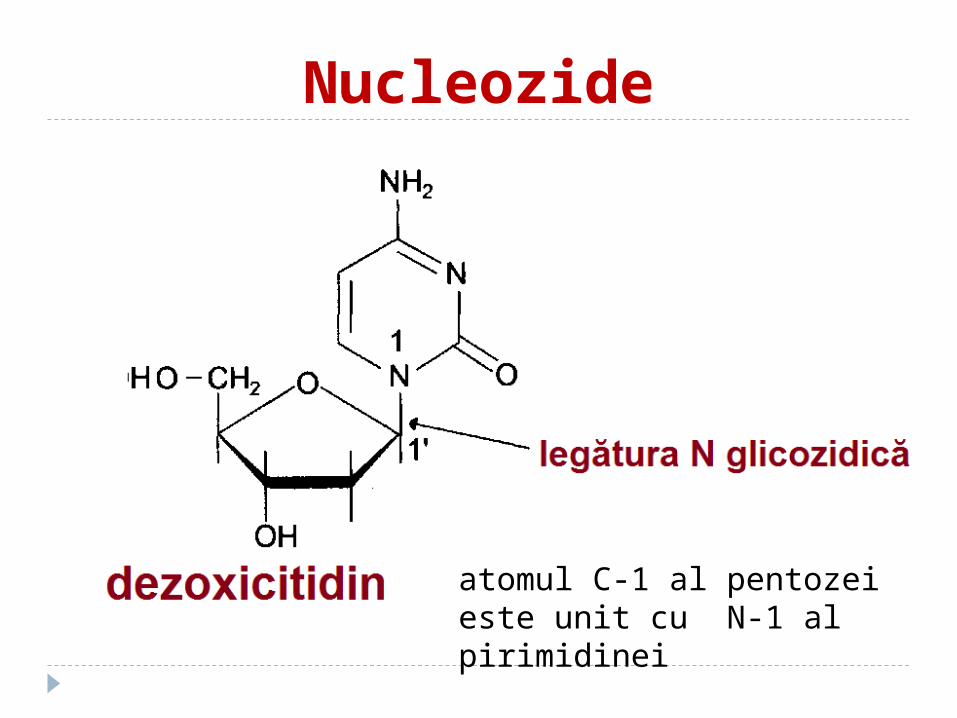
Nucleozide
atomul C-1 al pentozei este unit cu N-1 al pirimidinei
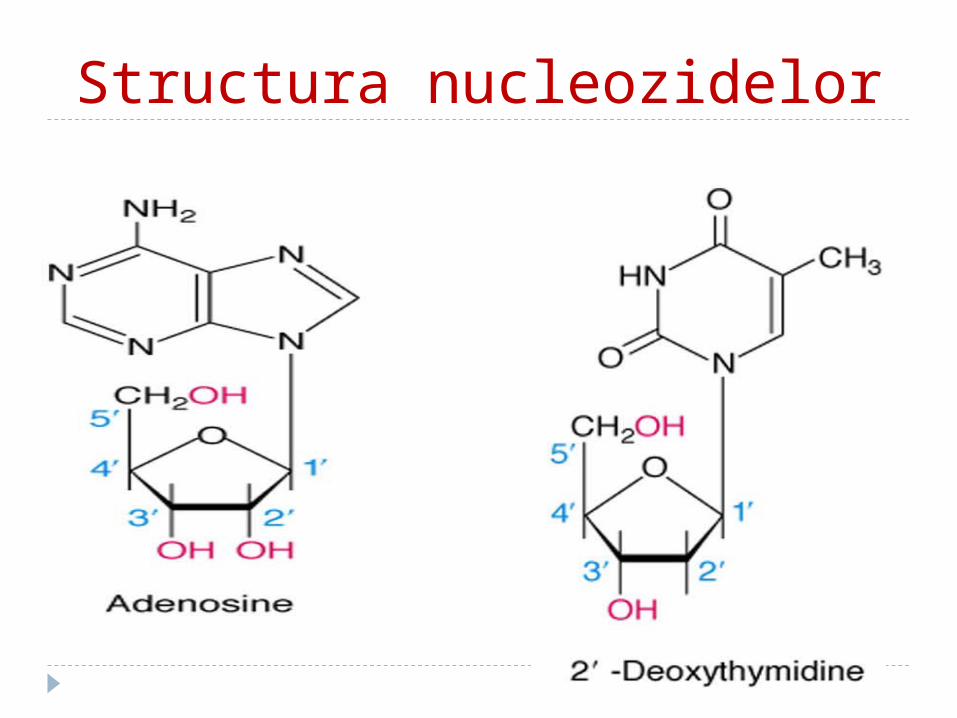
Structura nucleozidelor
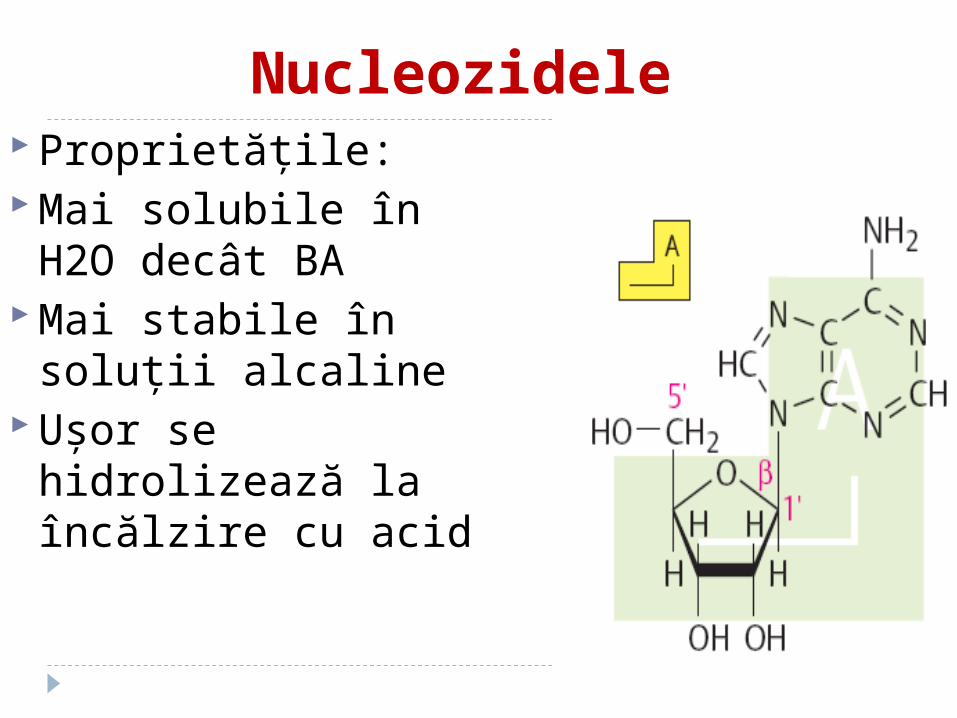
Nucleozidele Proprietăţile: Mai solubile în H2O
decât BA Mai stabile în soluţii
alcaline Uşor se hidrolizează la
încălzire cu acid
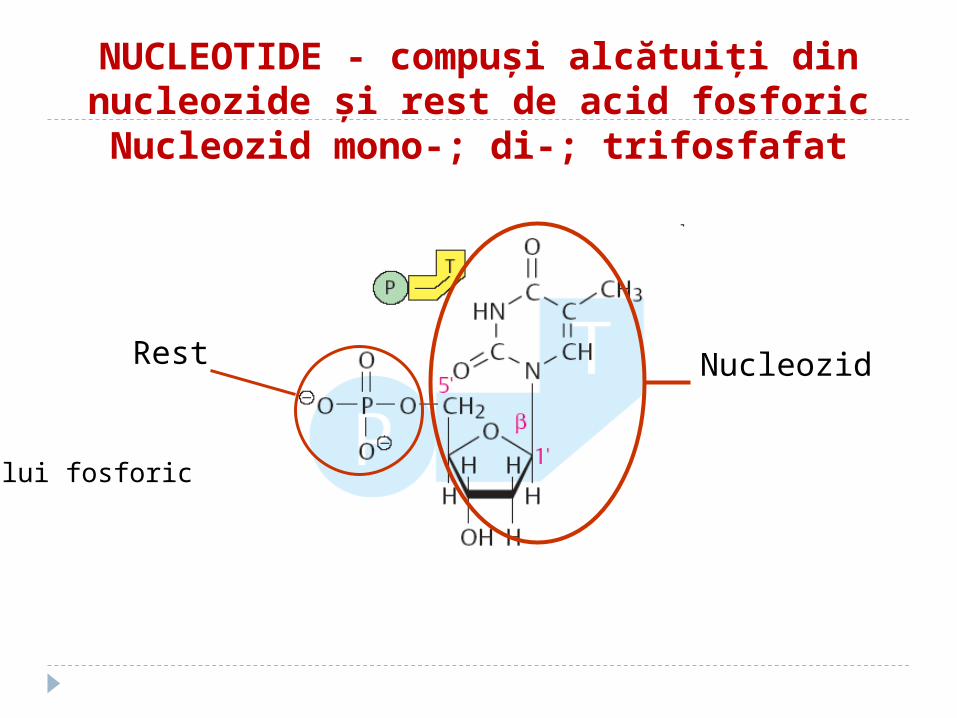
NUCLEOTIDE - compuşi alcătuiţi din nucleozide şi rest de acid fosforicNucleozid mono-; di-; trifosfafat
Rest al
acidului fosforic
Nucleozid
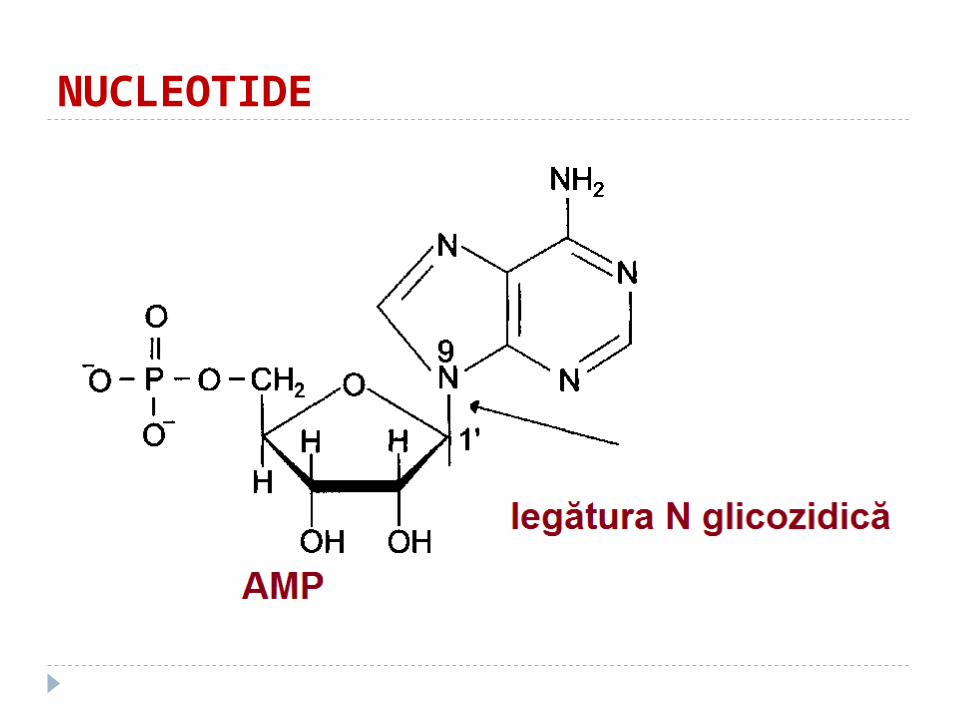
NUCLEOTIDE
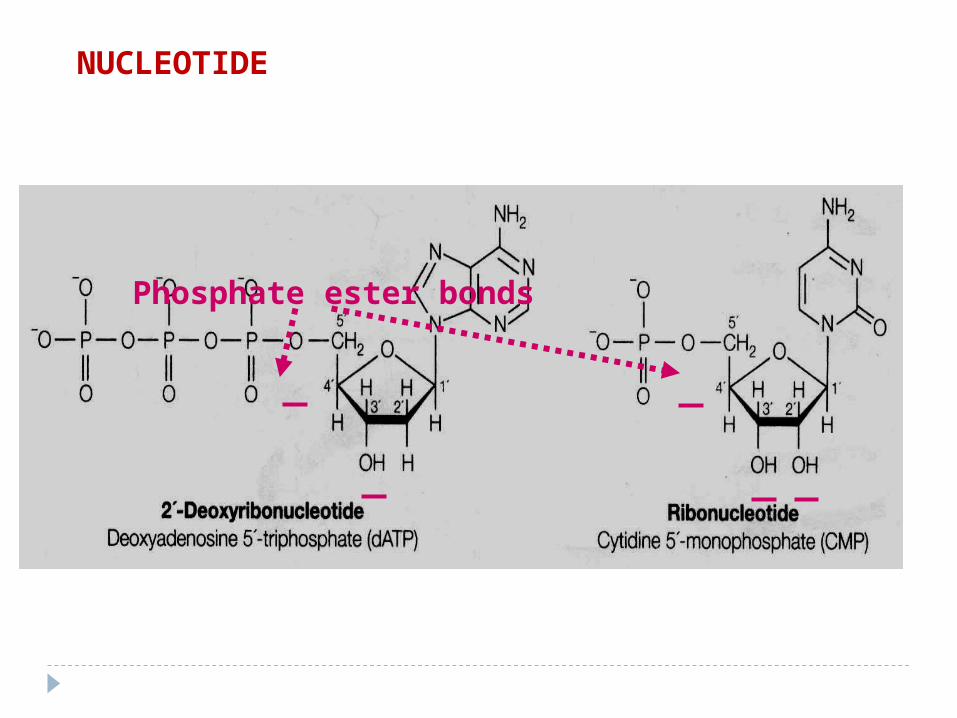
Phosphate ester bonds
NUCLEOTIDE
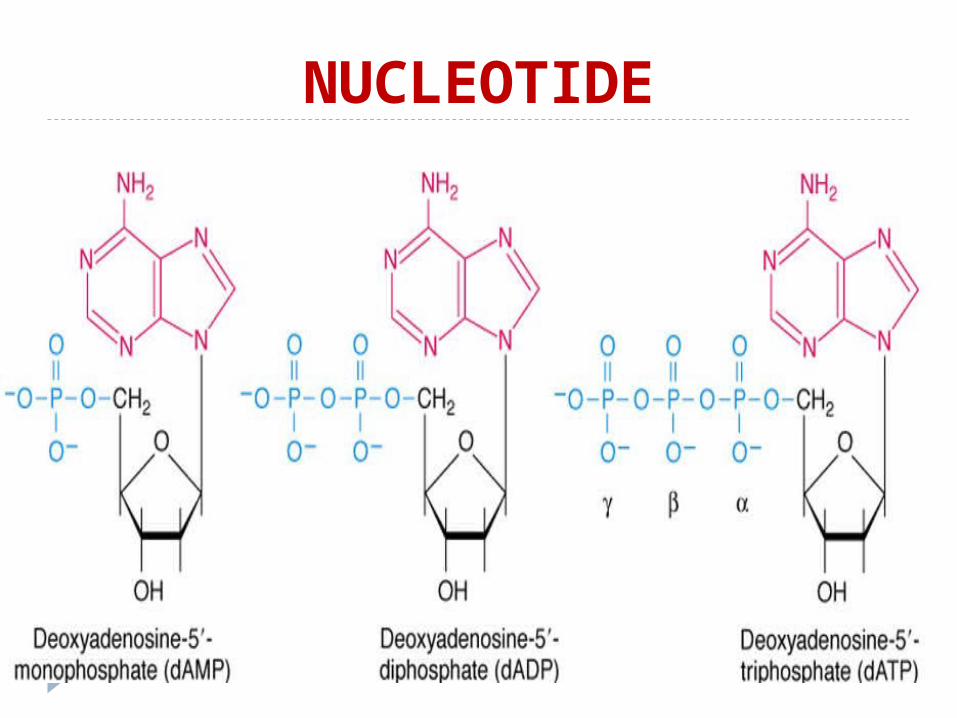
NUCLEOTIDE
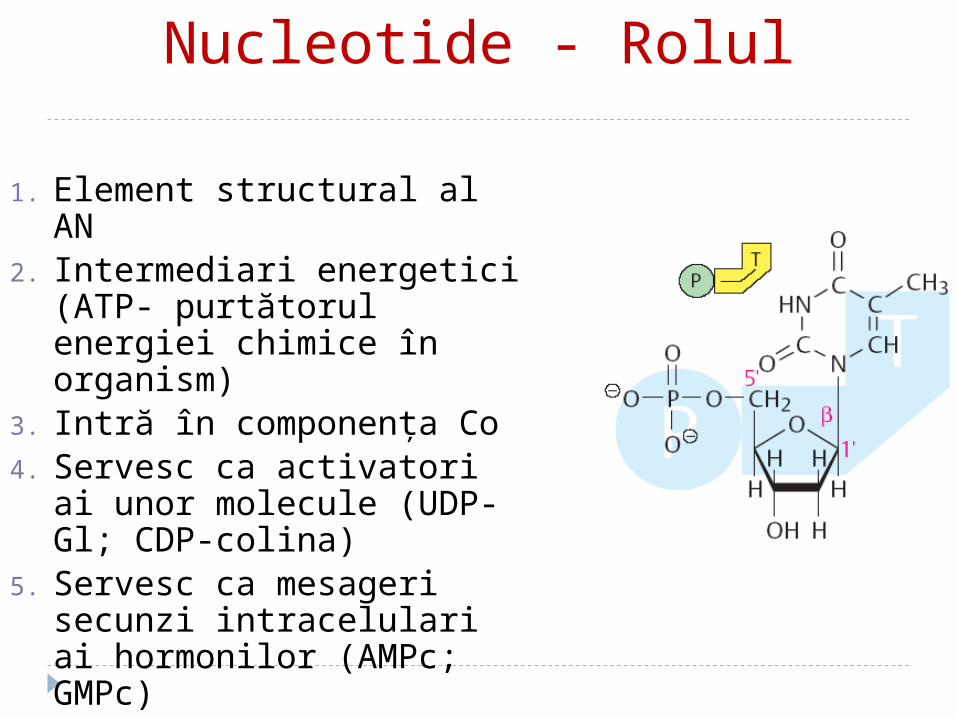
Nucleotide - Rolul
1. Element structural al AN2. Intermediari energetici (ATP-
purtătorul energiei chimice în organism)
3. Intră în componenţa Co4. Servesc ca activatori ai unor
molecule (UDP-Gl; CDP-colina)
5. Servesc ca mesageri secunzi intracelulari ai hormonilor (AMPc; GMPc)
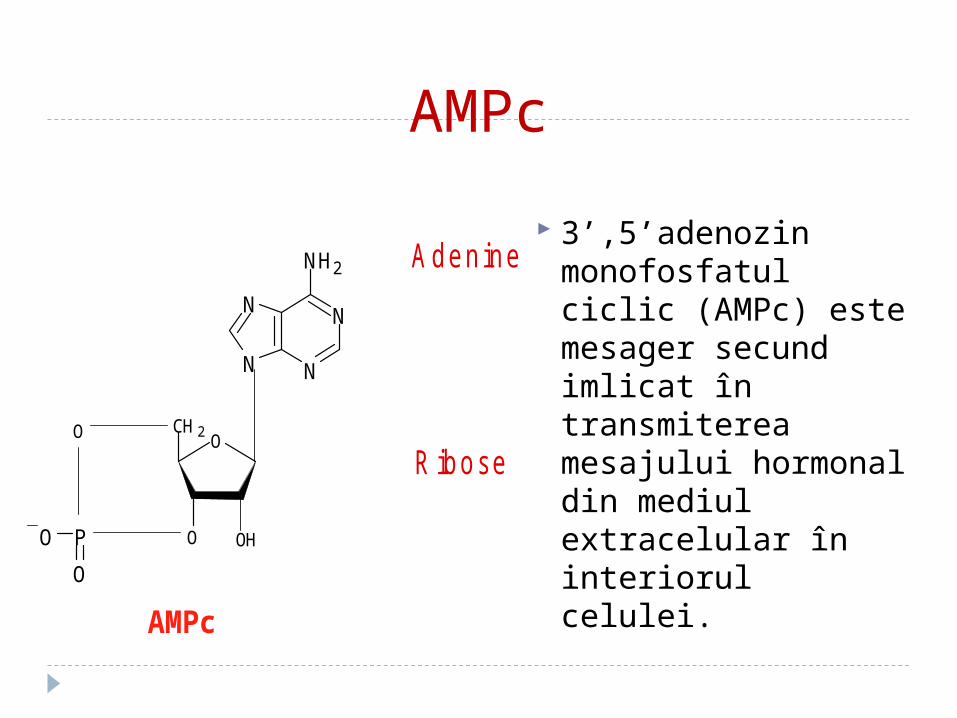
AMPc
OCH2
OP
O
O
A den ine
N
N
N
N
NH2
O
OH
R ibose
AMPc
3’,5’adenozin monofosfatul ciclic (AMPc) este mesager secund imlicat în transmiterea mesajului hormonal din mediul extracelular în interiorul celulei.
NUCLEOTIDE

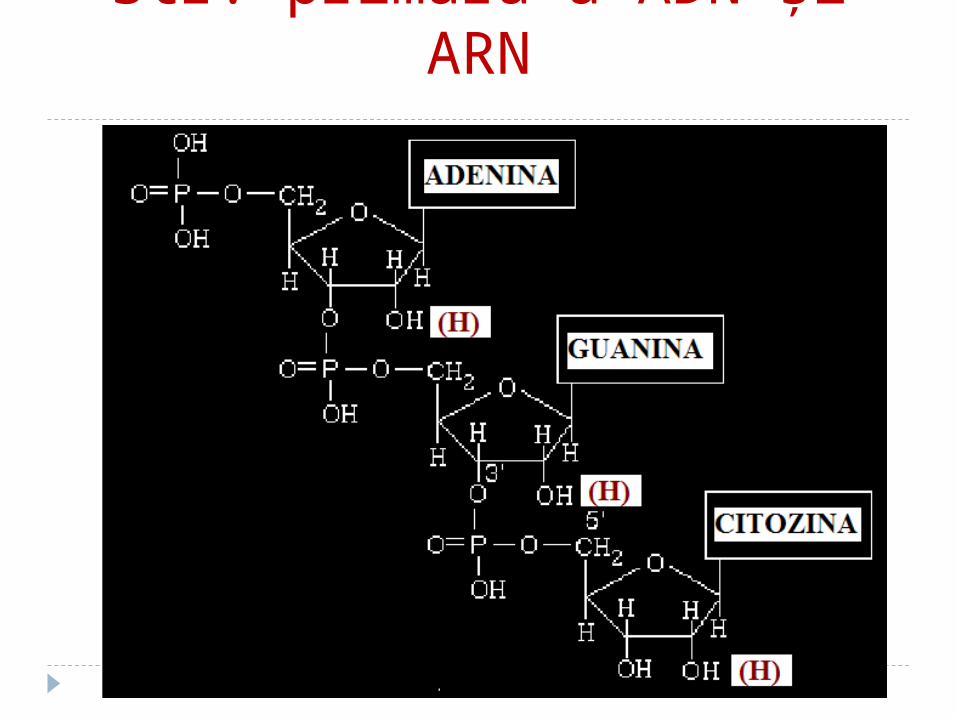
Structura primară a AN Reprezintă secvenţa mononucleotidelor în
lanţul polinucleotidic liniar, legate între ele prin legăturile 3' - 5' fosfodiesterice
Catenele au două capete: 5‘ – nucleozid tri fosfatul; 3‘ – gr. OH liberă
Str. primară a ADN şi ARN
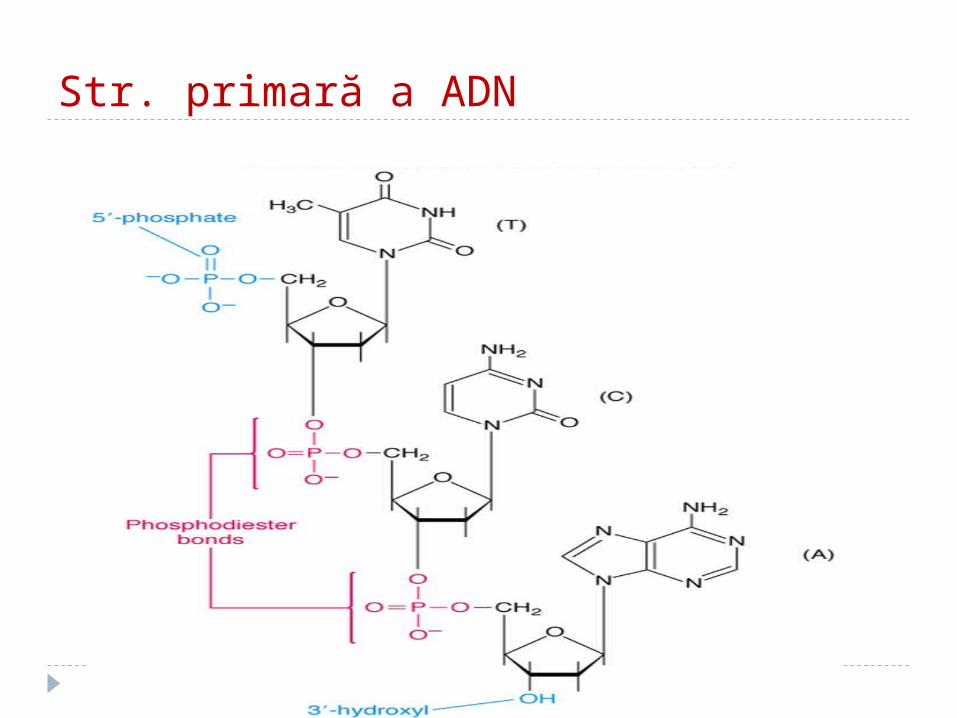
Str. primară a ADN

Dublu helixul ADN Watson şi Crick
(1953) au postulat modelul structural al moleculei de DNA - dublul helix (spirală dublă)

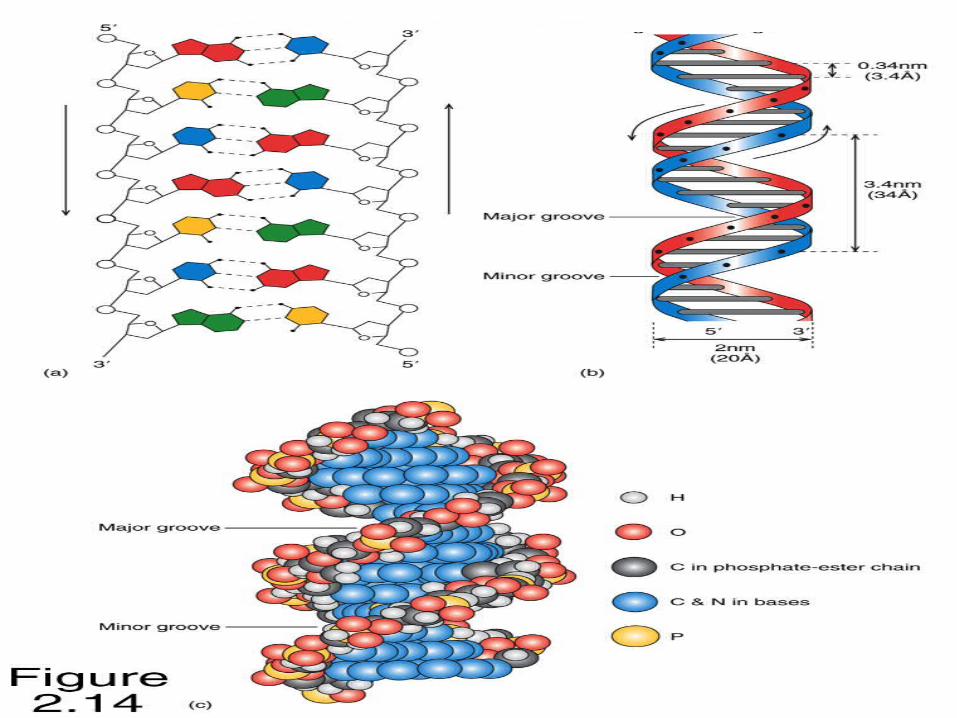
Caracteristicile dublei spirale:
1.2 lanţuri polidezoxiribonucleotidice se răsucesc helicoidal în jurul unui ax comun, formând dublul helix cu orientare spre dreapta;
2.Cilindrul ce încadrează dublul helix are d=2nm
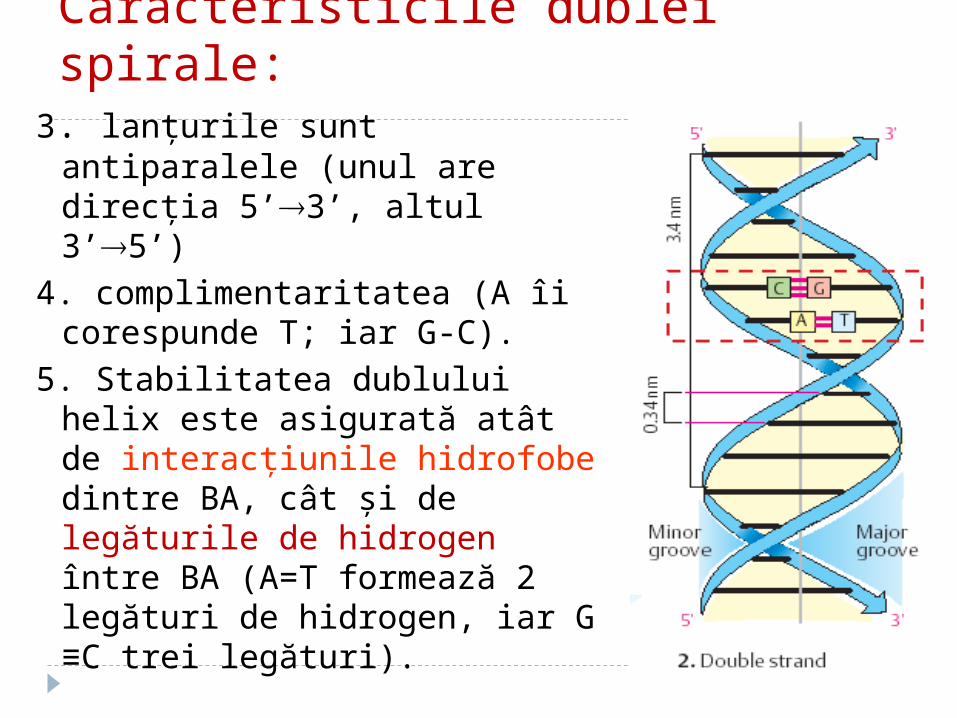
Caracteristicile dublei spirale:3. lanţurile sunt antiparalele (unul
are direcţia 5’3’, altul 3’5’)
4. complimentaritatea (A îi corespunde T; iar G-C).
5. Stabilitatea dublului helix este asigurată atât de interacţiunile hidrofobe dintre BA, cât şi de legăturile de hidrogen între BA (A=T formează 2 legături de hidrogen, iar G ≡C trei legături).
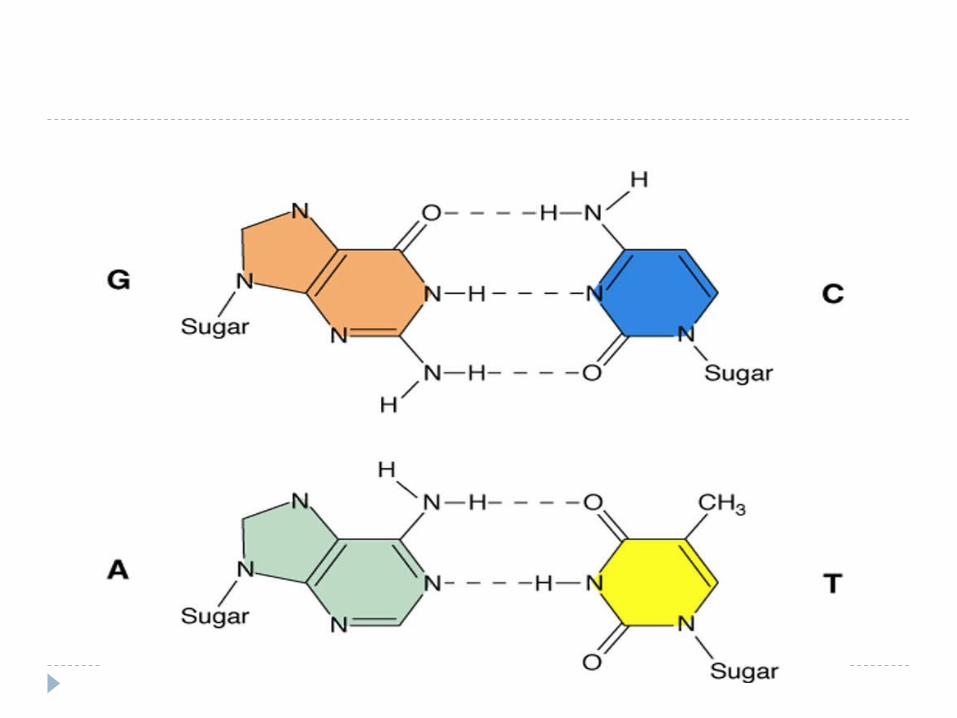

Caracteristicile dublei spirale:
6. BA hidrofobe sunt situate în interiorul spiralei duble şi aranjate sub formă de stive, pe cînd complexul pentozofosfat este situat la exteriorul spiralei duble, bine interacţionează cu apa, de aceia molecula gigantă de DNA se dizolva în apă.

Caracteristicile dublei spirale:
7. Spirală este regulată (fiecare spiră cuprinde 10 nucleotide). Distanţa dintre BA învecinate este de 0,34 nm, perioada de identitate (pasul) – 3,4 nm.
8. La pH=7 grupele fosfat sunt ionizate, poarta sarcini negative, deaceia DNA prezintă acid puternic.
9. Dublul helix este de tip plectonemical (transversal în acelaşi plan), dar nu paranemical (longitudional)
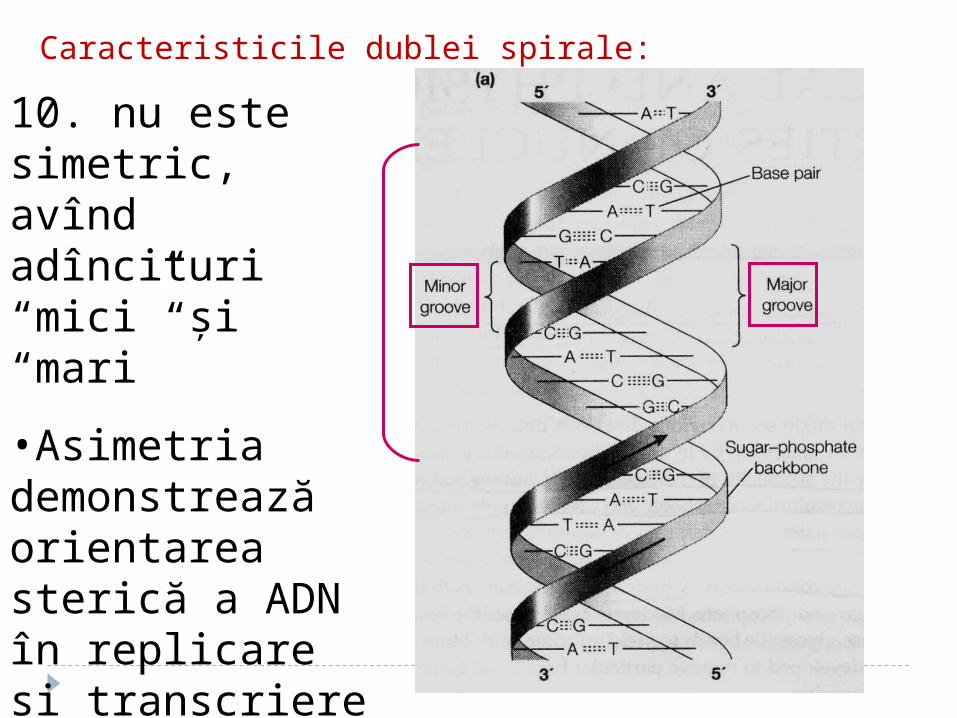
10. nu este simetric, avînd adîncituri “mici” şi “mari”
•Asimetria demonstrează orientarea sterică a ADN în replicare şi transcriere
Caracteristicile dublei spirale:

Legităţile lui Chargaff
1. Conţinutul A=T, iar al G=C2. În orice preparat de DNA, independent de
specie, suma bazelor purinice este egală cu cea a bazelor pirimidinice (A+G=T+C)
3. Preparatele de DNA separate din diferite ţesuturi a uneia şi aceeiaşi specie de organisme sunt absolut identice privind componenţa nucleotidică.
4. Componenţa nucleotidică a DNA la aceeaşi specie nu se modifică odată cu vârstă, nu depinde de regimul alimentar şi modificările mediului.

Legităţile lui Chargaff5. dacă A+T este mai mare decît G+C avem
DNA de tip AT6. dacă G+C este mai mare decît A+T avem
DNA de tip GC7. t de topire este mai mica cînd predomină
perechile A-T8. t de topire este mai mare cînd predomină
perechile G-C 9. la eucariote DNA mitocondrial este
circular
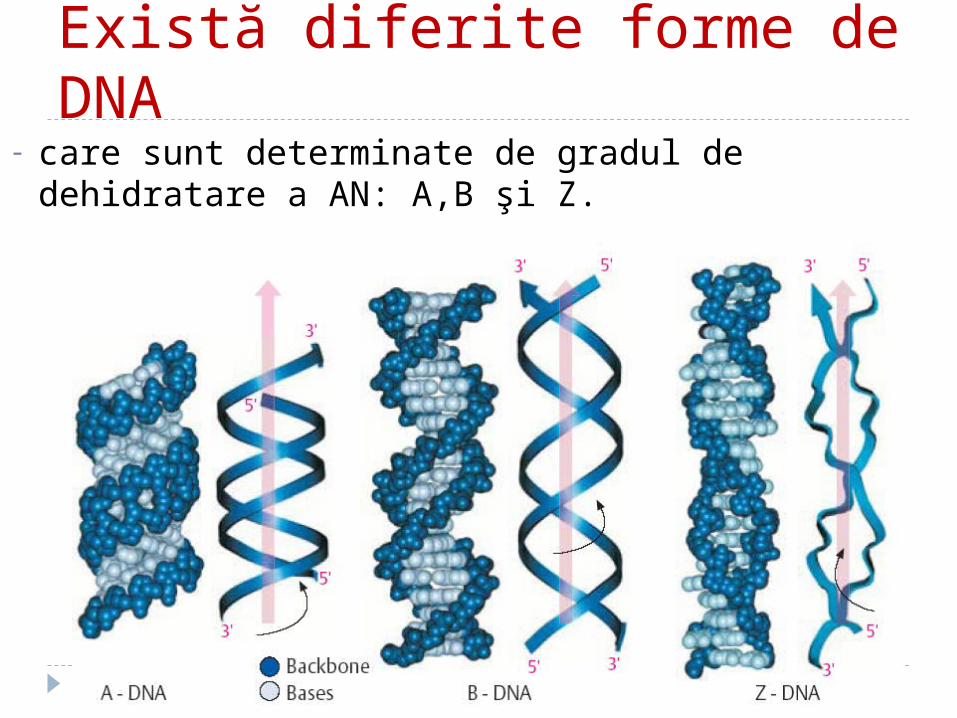
Există diferite forme de DNA- care sunt determinate de gradul de dehidratare a AN: A,B
şi Z.

- forma A:- conţine 11 resturi la o spiră, - este răsucită spre dreapta.- Pasul spirei este de 32 A forma clasică B:- conţine 10,4 perechi de baze per spiră.- este răsucită spre dreapta.- 10 nucleotide ocupă 34 A (3,4 nm).- o nucleotidă cuprinde 3,4A (0,34 nm).• Conformația Z este răsucită spre stînga. conţine 12 baze per spiră pasul spirei este de 45 A
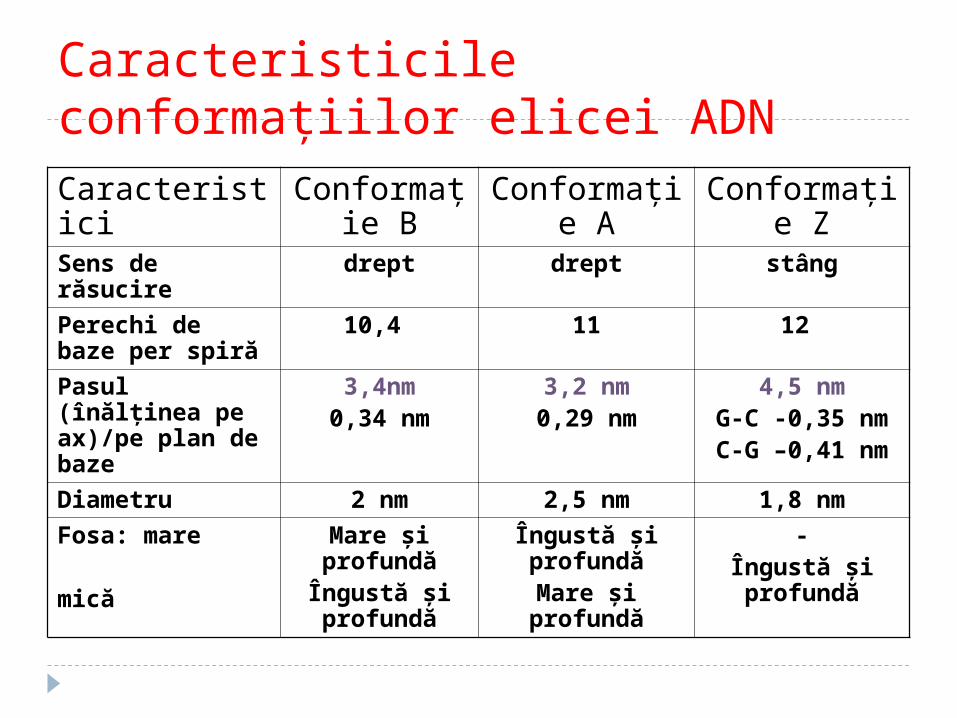
Caracteristicile conformaţiilor elicei ADNCaracteristici Conformaţi
e BConformaţi
e AConformaţi
e ZSens de răsucire
drept drept stâng
Perechi de baze per spiră
10,4 11 12
Pasul (înălţinea pe ax)/pe plan de baze
3,4nm0,34 nm
3,2 nm0,29 nm
4,5 nmG-C -0,35 nmC-G –0,41 nm
Diametru 2 nm 2,5 nm 1,8 nm
Fosa: mare mică
Mare şi profundăÎngustă şi profundă
Îngustă şi profundăMare şi
profundă
-Îngustă şi profundă
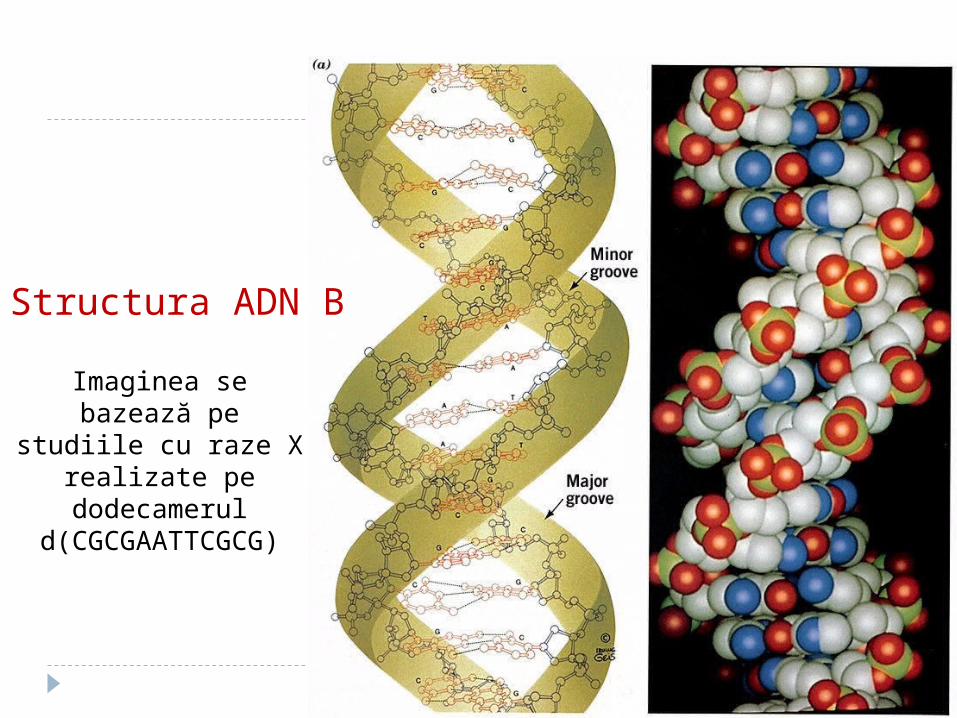
Structura ADN B
Imaginea se bazează pe studiile cu raze X realizate
pe dodecamerul d(CGCGAATTCGCG)
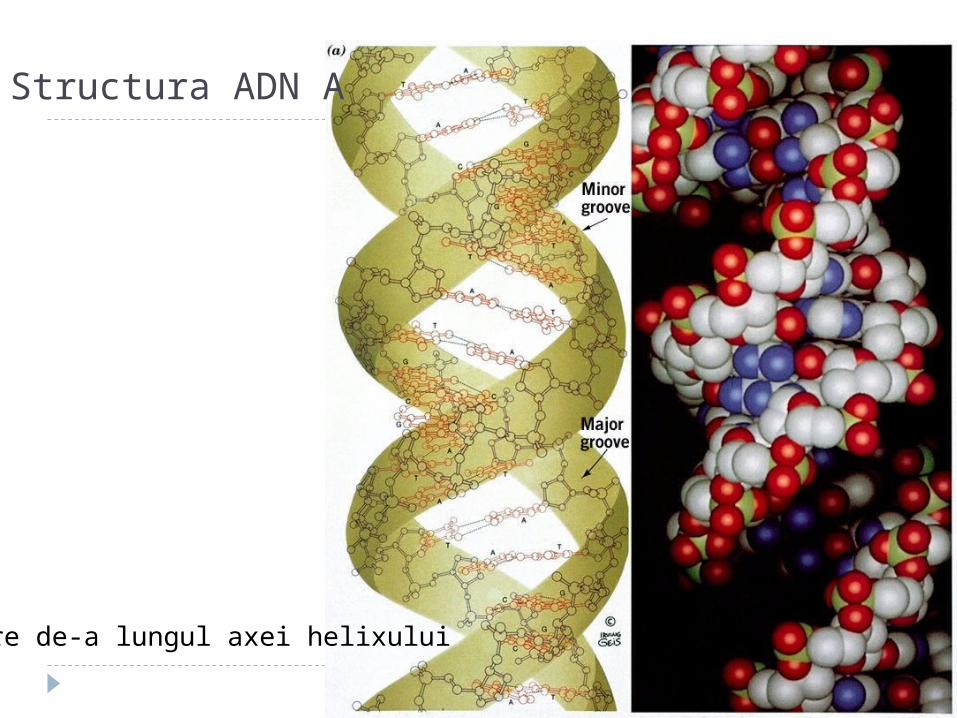
Structura ADN A
Vedere de-a lungul axei helixului
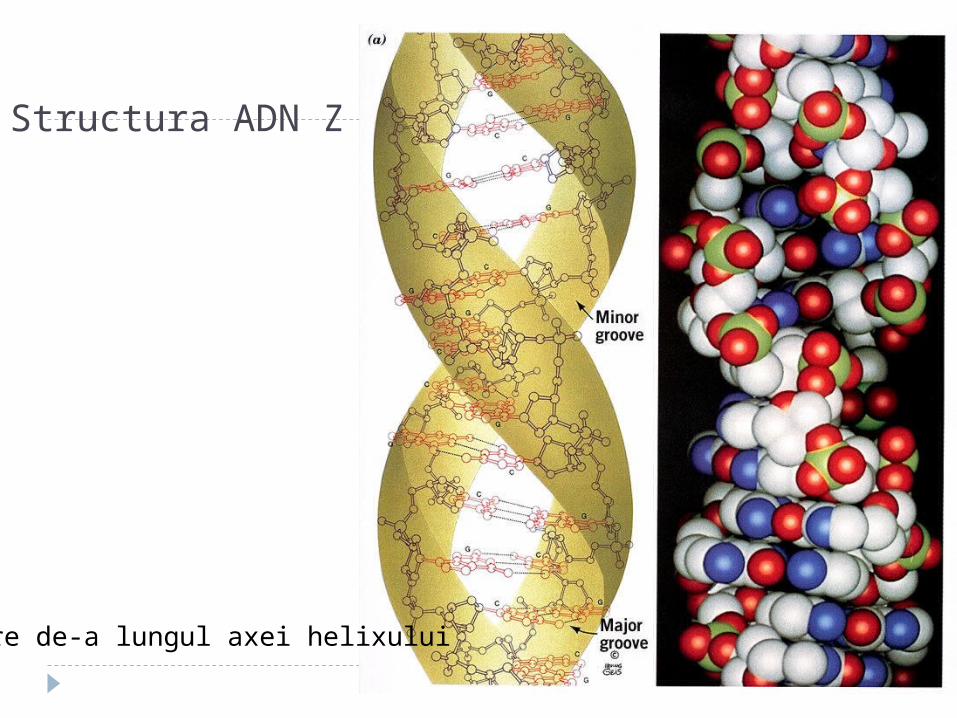
Structura ADN Z
Vedere de-a lungul axei helixului
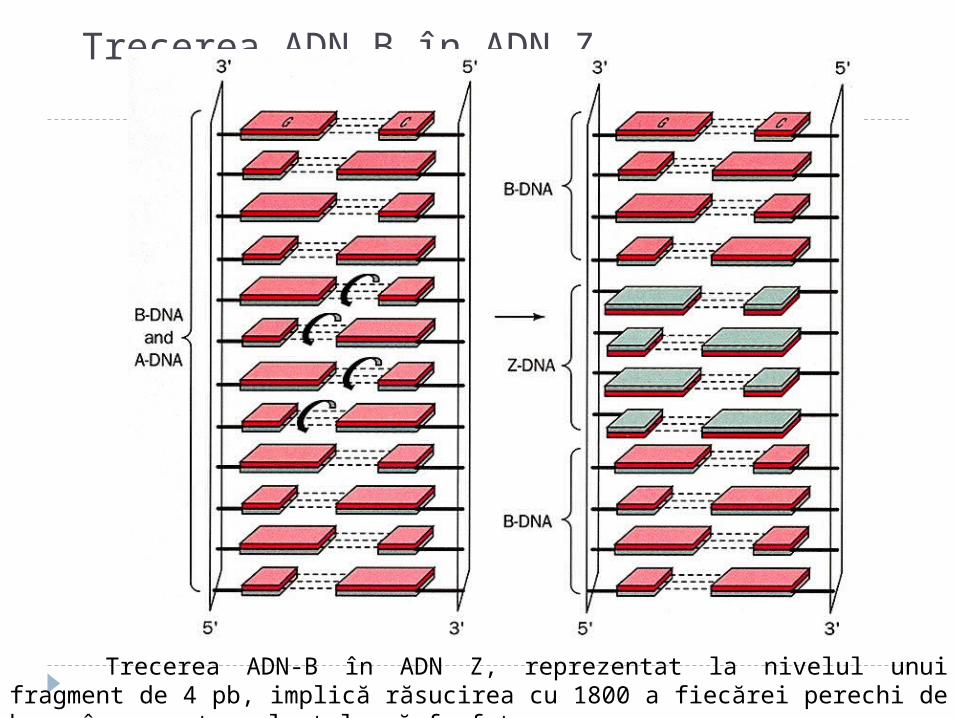
Trecerea ADN B în ADN Z
Trecerea ADN-B în ADN Z, reprezentat la nivelul unui fragment de 4 pb, implică răsucirea cu 1800 a fiecărei perechi de baze în raport cu lanţul oză-fosfat

ADN circular
La bacterii, viruşi – cele 2 extremităţi ale unor molecule de ADN dublu catenare se pot lega covalent formând ADN circular

De la ADN la cromozomi In eucariote ADN-ul este stocat în nucleu. Deoarece nu este suficient spaţiu iar
molecula ADN-ului este extrem de mare (lungimea ADN de la o singură celulă umană e 2m!!!), ADN-ul trebuie compactat.

Organizarea materialului cromosomial la eucariote
În perioada de repaus a celulei, aproape tot ADN se află în cromatina nucleului, care formează cromosomi înaintea mitozei.
Cromatina conţine: ADN (35%); ARN (5%) proteinele bazice – histone (H1, H2A, H2B,H3, H4) şi proteine nehistonice- hertone.
Unitatea structurală a cromatinei este nucleosomul

Nucleosomul – este un octamer histonic (2H2A; 2H2B;
2H3; 2H4) înfăşurat de aproximativ de 2 ori de ADN (dublul helix, cu lungimea de 146 perechi de nucleotide).
Între 2 nucleosomi se găseşte ADN de legătură (linker) asociat cu H1
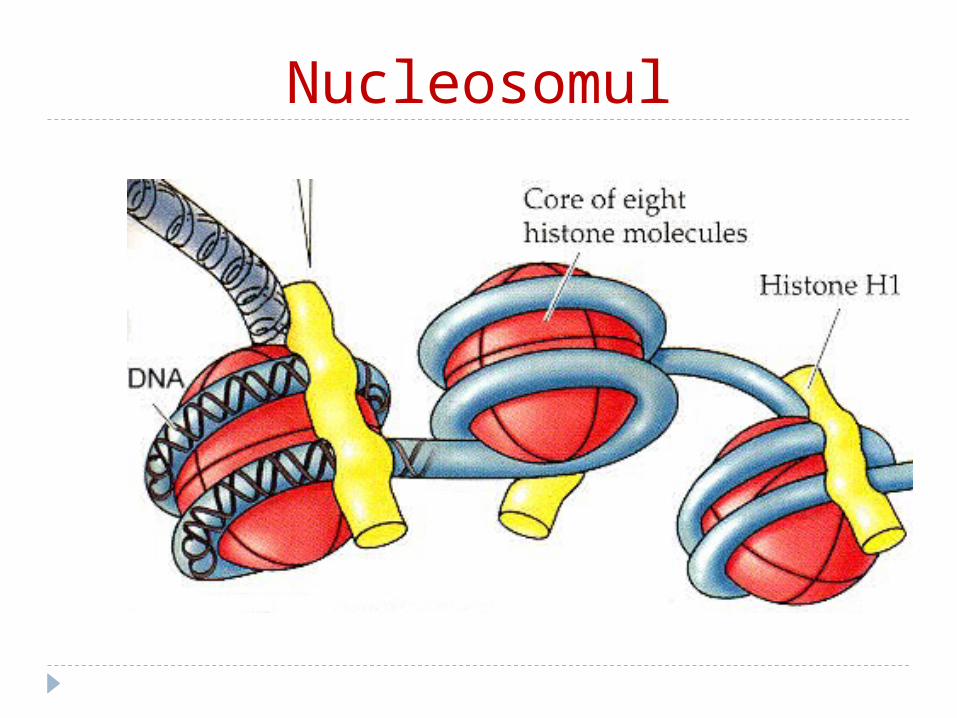
Nucleosomul
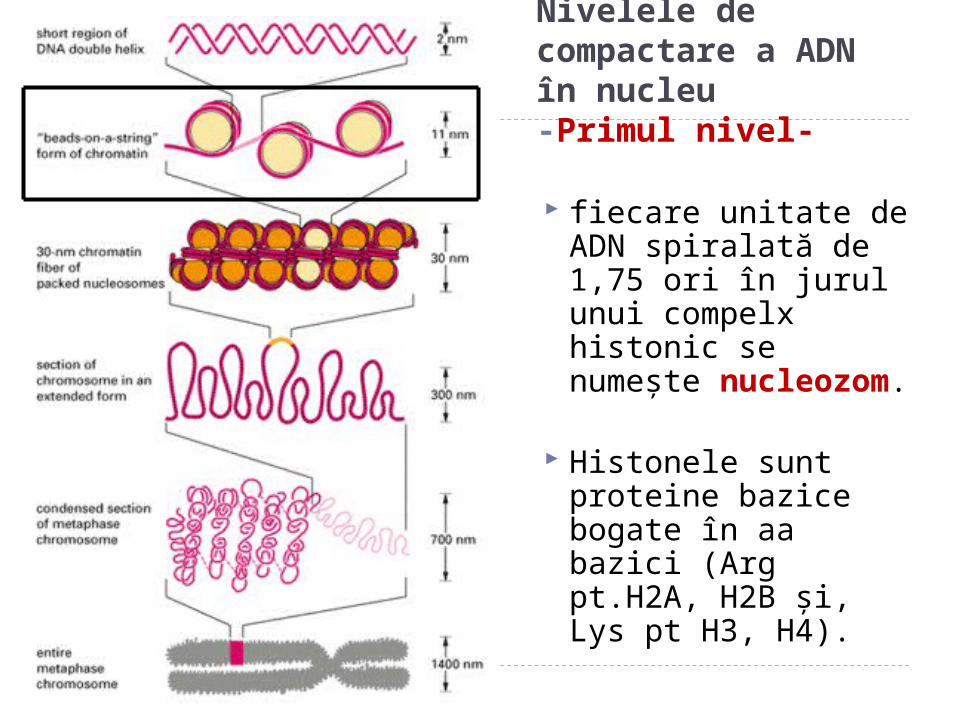
Nivelele de compactare a ADN în nucleu-Primul nivel-
r fiecare unitate de ADN spiralată de 1,75 ori în jurul unui compelx histonic se numeşte nucleozom.
Histonele sunt proteine bazice bogate în aa bazici (Arg pt.H2A, H2B şi, Lys pt H3, H4).
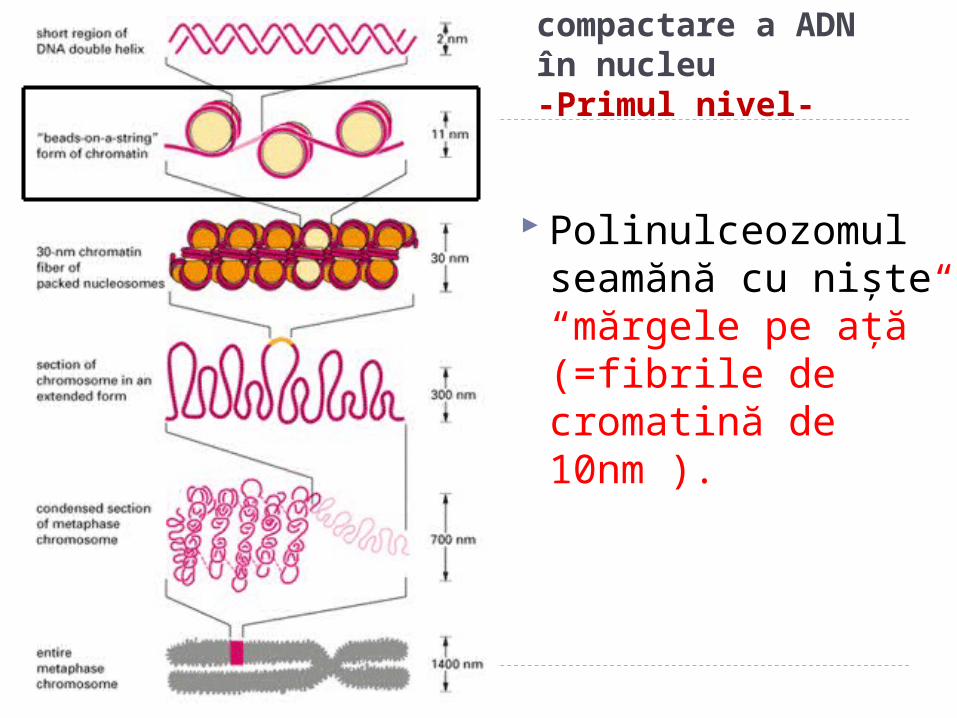
Nivelele de compactare a ADN în nucleu-Primul nivel-
r Polinulceozomul
seamănă cu nişte “mărgele pe aţă” (=fibrile de cromatină de 10nm ).

Mărgele de ametist
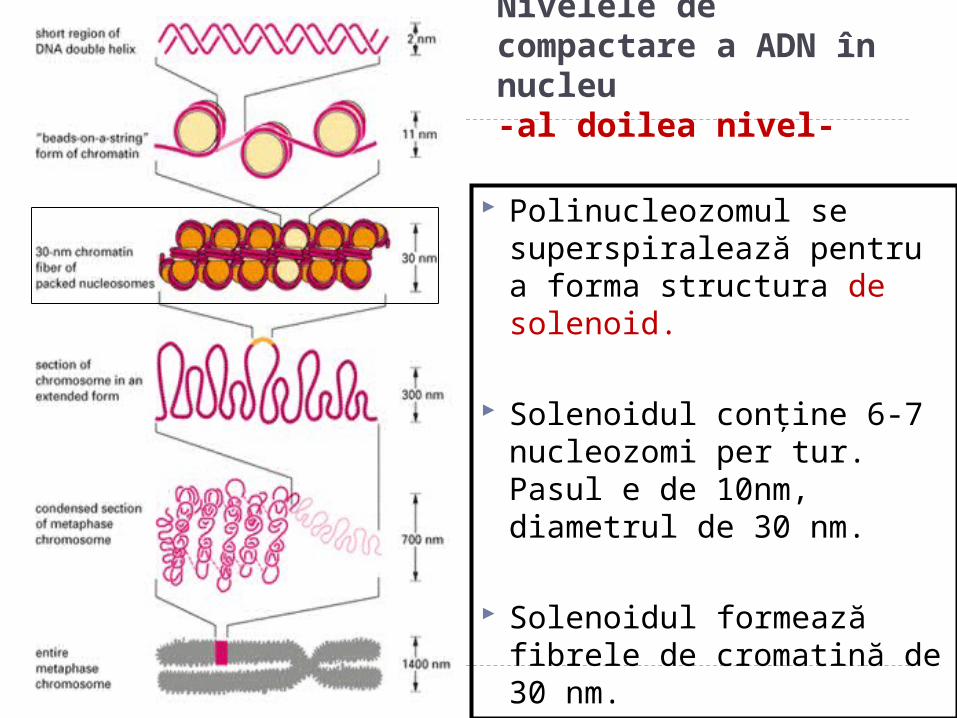
Nivelele de compactare a ADN în nucleu-al doilea nivel-
Polinucleozomul se superspiralează pentru a forma structura de solenoid.
Solenoidul conţine 6-7 nucleozomi per tur. Pasul e de 10nm, diametrul de 30 nm.
Solenoidul formează fibrele de cromatină de 30 nm.
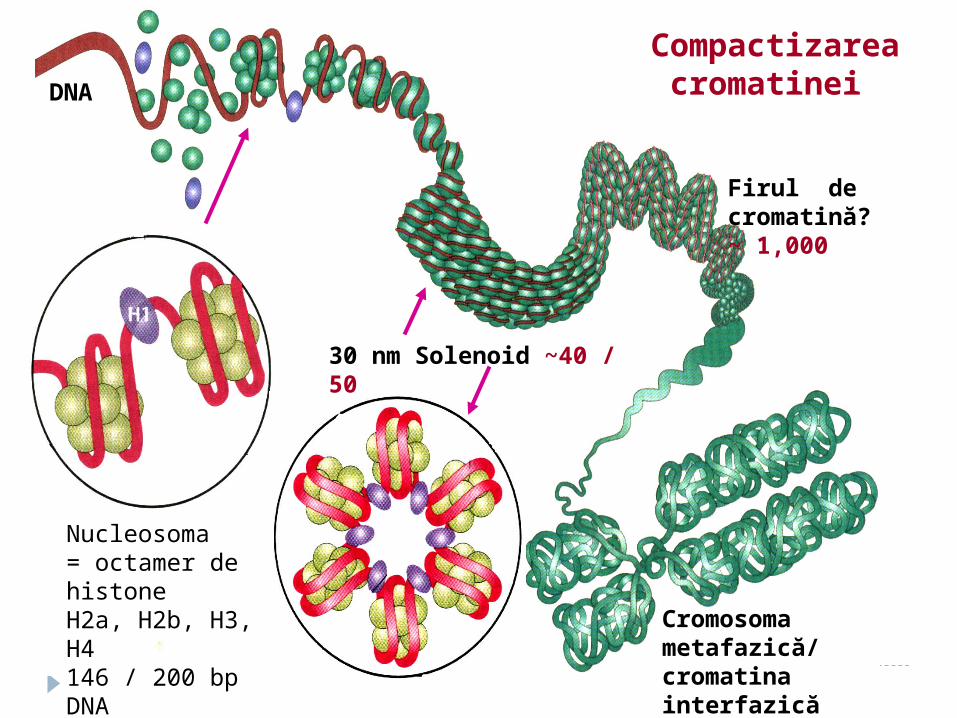
30 nm Solenoid ~40 / 50
DNA
Firul de cromatină? ~ 1,000
Cromosoma metafazică/ cromatina interfazică ~ 10,000
Compactizarea cromatinei
Nucleosoma = оctamer de histoneH2a, H2b, H3, H4 146 / 200 bp DNACompactizare ~10 ori
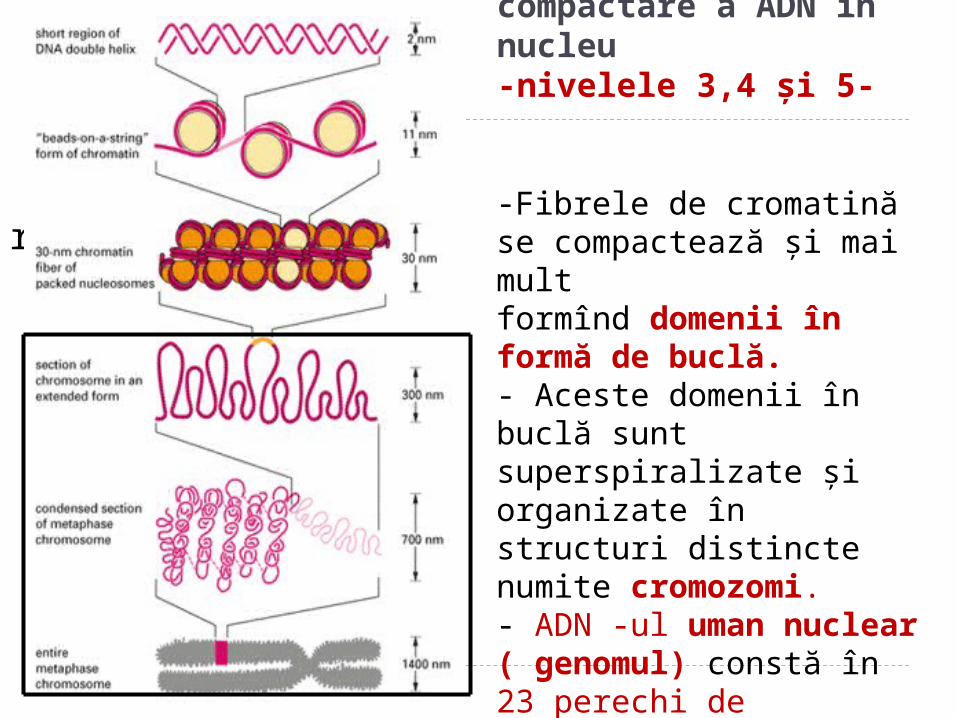
Nivelele de compactare a ADN în nucleu-nivelele 3,4 şi 5-
r
-Fibrele de cromatină se compactează şi mai mult formînd domenii în formă de buclă.- Aceste domenii în buclă sunt superspiralizate şi organizate în structuri distincte numite cromozomi.- ADN -ul uman nuclear ( genomul) constă în 23 perechi de cromozomi.
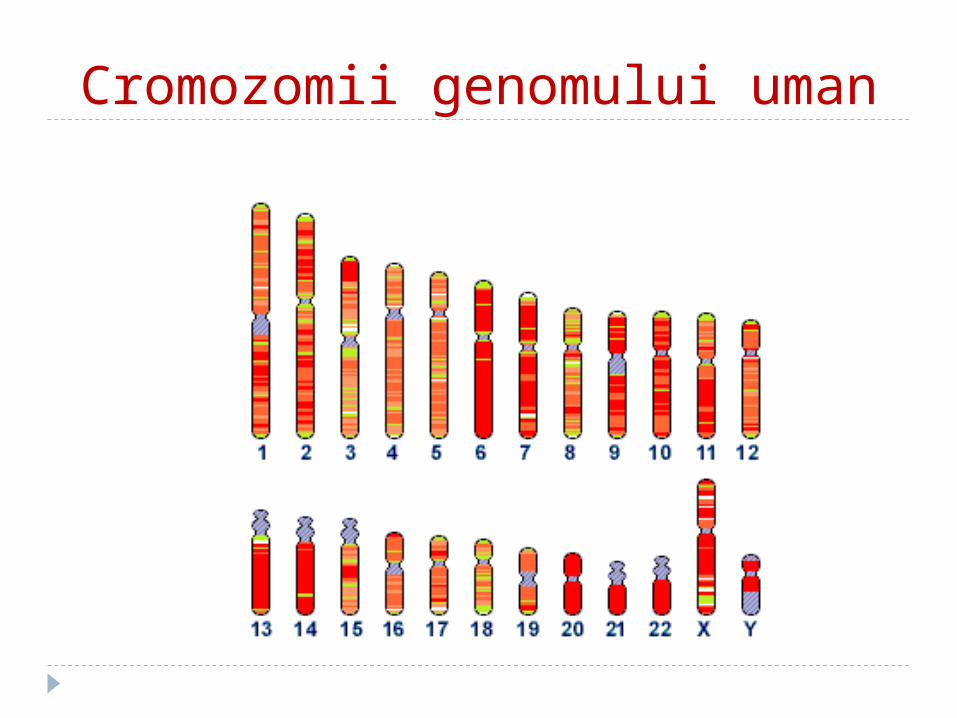
Cromozomii genomului uman
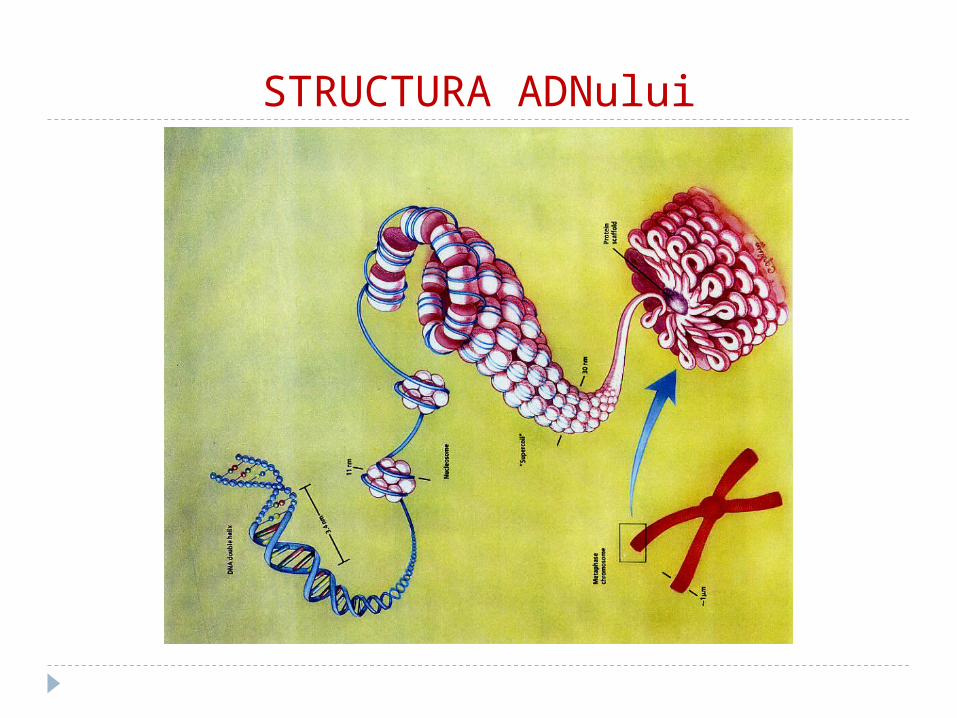
STRUCTURA ADNului

Proprietăţile fizico-chimice ale acizilor nucleici
- masa moleculară mare.- proprietatile coloidale si osmotice, tipice pentru
toţi compuşii macromoleculari.- proprietăţile lor hidrofile depind de fosfaţi.- viscozitatea şi densitatea înaltă a soluţiilor,- capacitatea de denaturare.- la pH fiziologic toti AN sunt polianioni (-)

Denaturarea şi renaturareaDenaturarea –sub acţiunea agenților
denaturanți (temperaturii, pH, substanţelor chimice) are loc ruperea legăturilor de hidrogen şi forţelor hidrofobe ce stabilizează structura AND -ului.
ADN îşi pierde proprietăţile biologice.
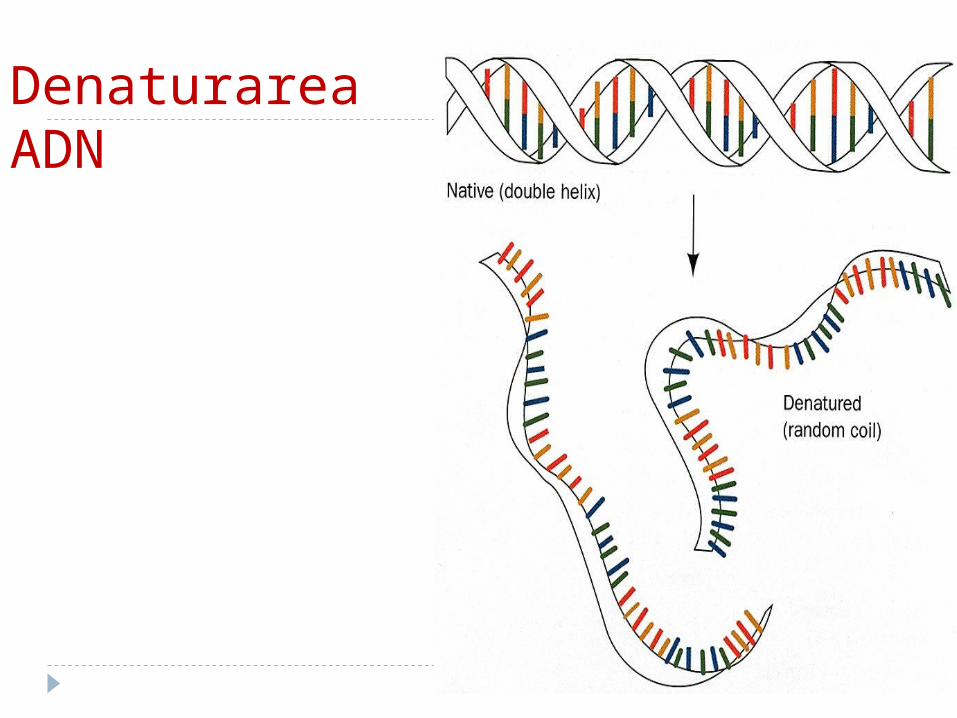
Denaturarea ADN

Renaturarea La răcirea treptată catenele din nou se
reunesc după principiul complementaritătii, formînd spirala dublă nativă.
Acest fenomen se numeşte renaturare (atunci cînd t e mai mică decît cea de topire).
La racirea bruscă renaturarea nu are loc. Denaturarea şi renaturarea AN este însoţită
de schimbarea activitătii lor optice.
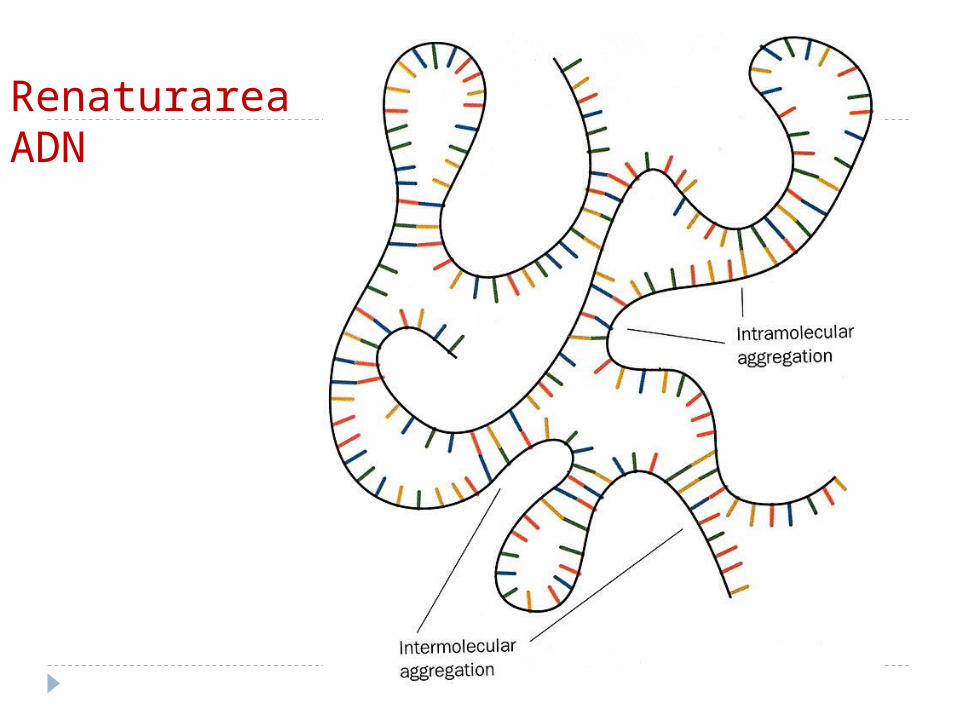
Renaturarea ADN

Hibridizarea AN
Pe capacitatea de renaturare a AN este bazată metoda de determinare a gradului de înrudire a AN, care poartă denumirea de hibridizare moleculară.
La baza ei stă împerecherea complementară a sectoarelor unicatenare ale AN cu formarea unui heteroduplex

Hibridizarea AN1. AN se denaturează separat;2. se incubează împreună ambele tipuri
de DNA (ori DNA şi RNA). 3. În condiţiile unui grad relativ crescut
de complementaritate a acestora se formează moleculele hibride (DNA-DNA sau DNA-RNA). Aceste molecule constau din sectoare spiralate şi nespiralate. Cu cît gradul de înrudire este mai înalt, cu atît hibridizarea este mai complectă.
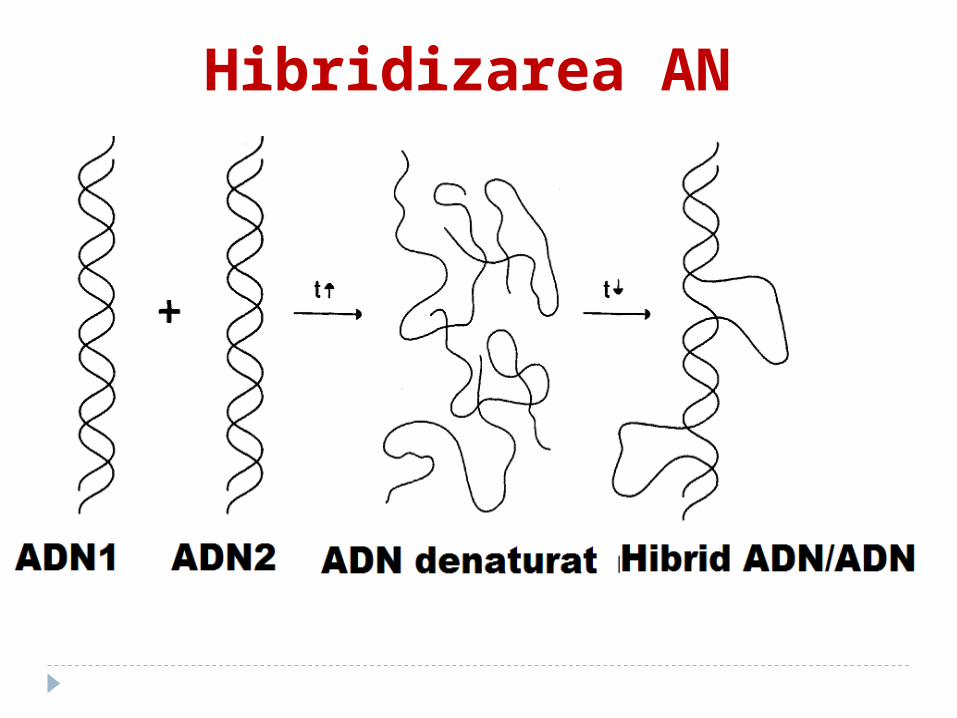
Hibridizarea AN
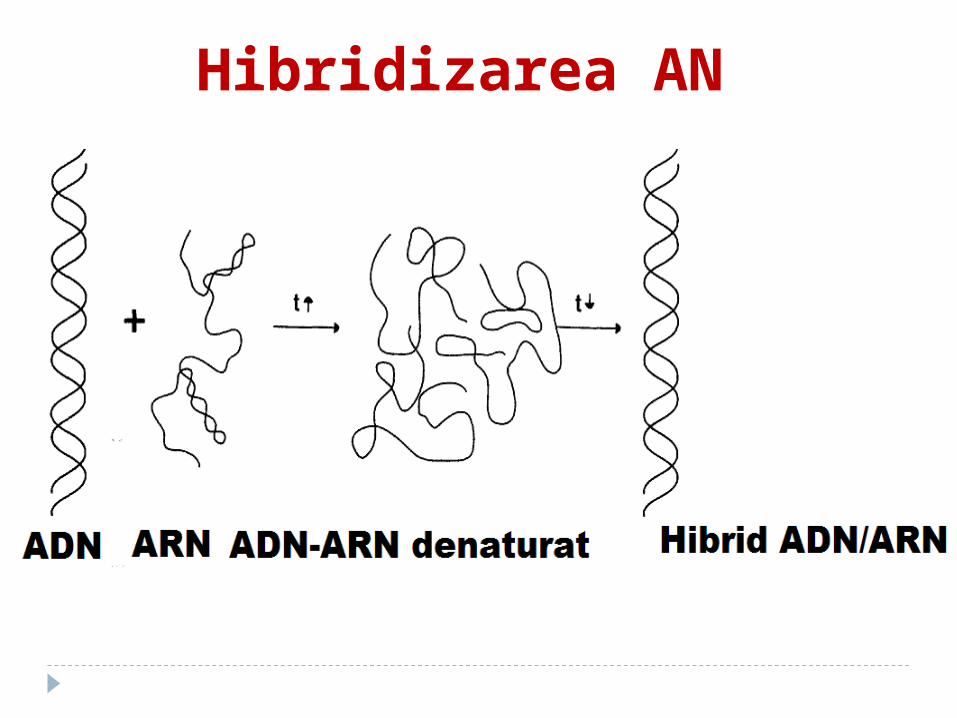
Hibridizarea AN

Această metodă a permis descoperirea particularităţilor structurii primare a DNA.
S-a stabilit, că în componenţa DNA a animalelor se află sectoare cu o succesiune nucleotidică identică, care de multe ori se repetă.
Hibridizarea decurge foarte repede. Restul DNA este prezentat printr-o
succesiune unicală a nucleotidelor, care nu se dublează.
Hibridizarea AN

REPLICAREA TRANSCRIPȚIA
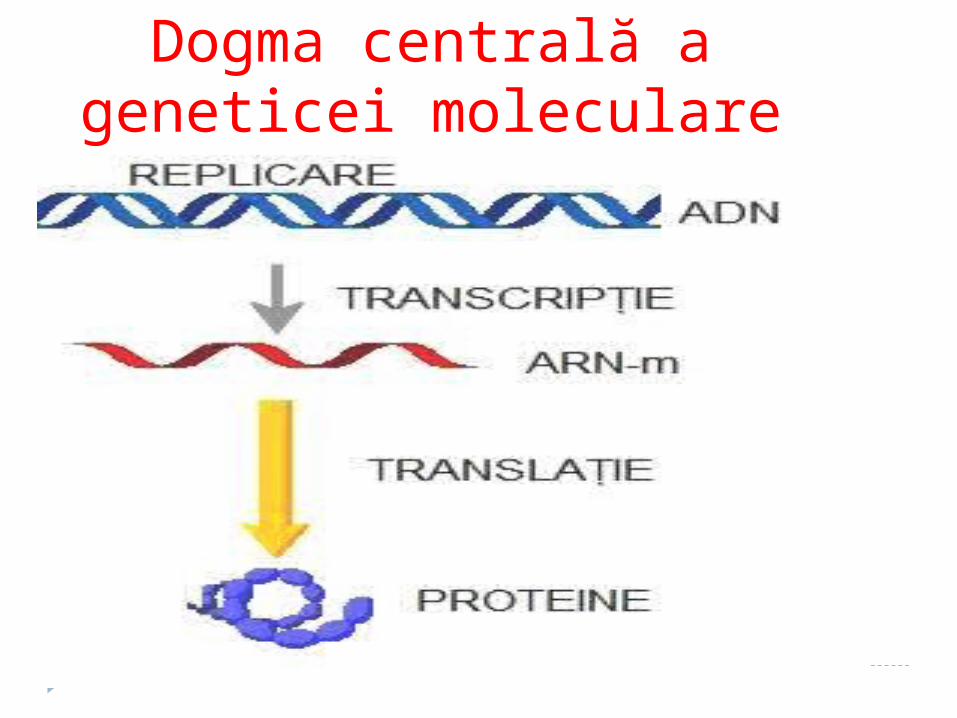
Dogma centrală a geneticei moleculare
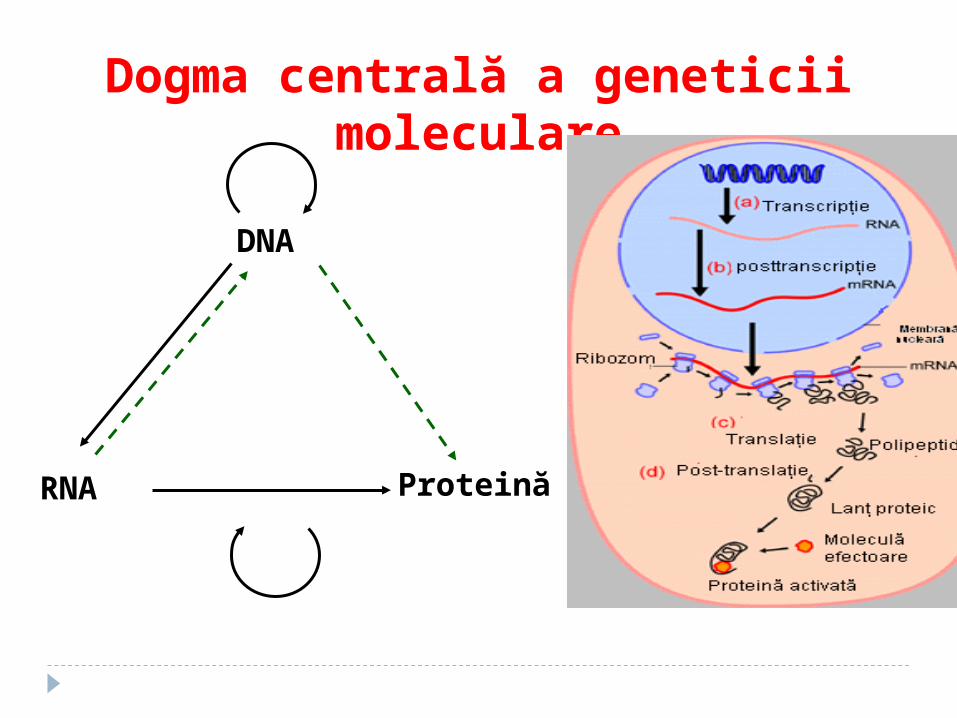
Dogma centrală a geneticii moleculare
DNA
RNA Proteină
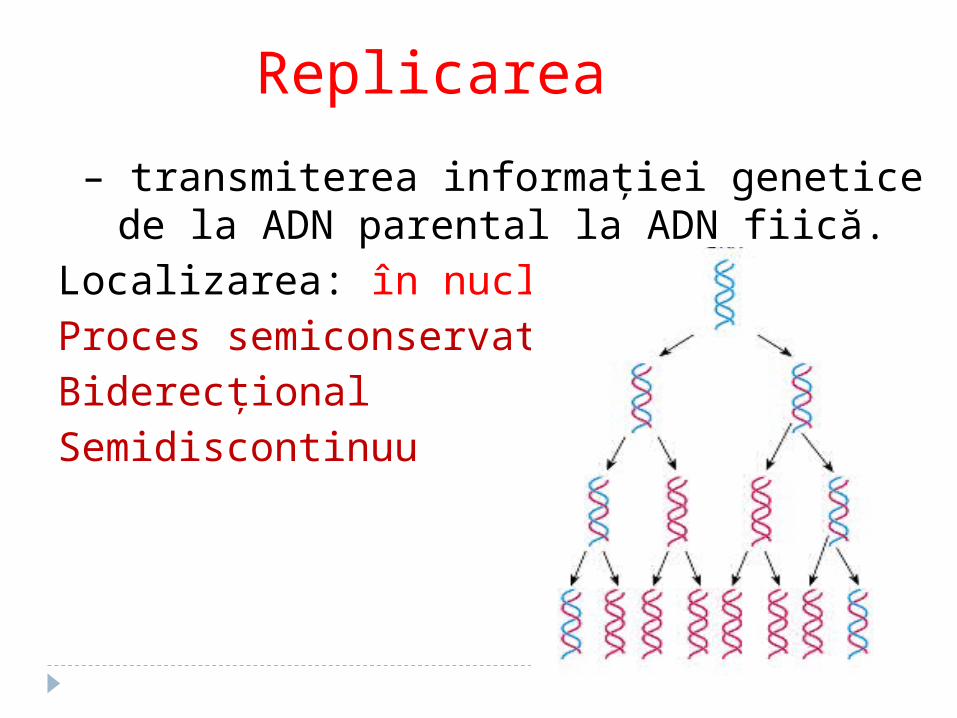
Replicarea
– transmiterea informaţiei genetice de la ADN parental la ADN fiică.
Localizarea: în nucleolProces semiconservativBiderecționalSemidiscontinuu
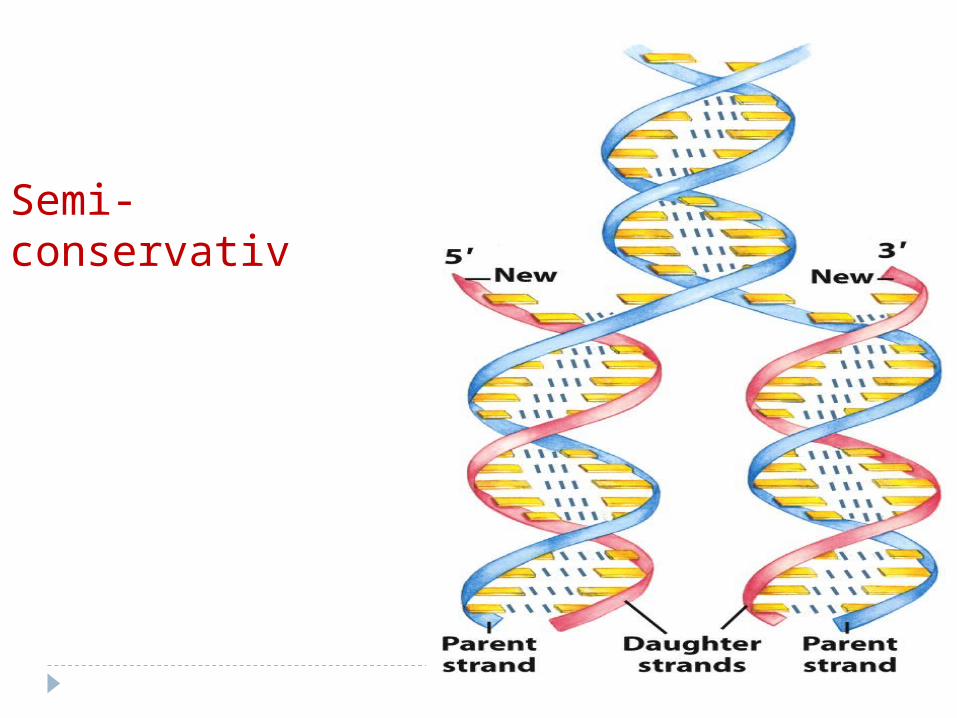
Semi-conservativ
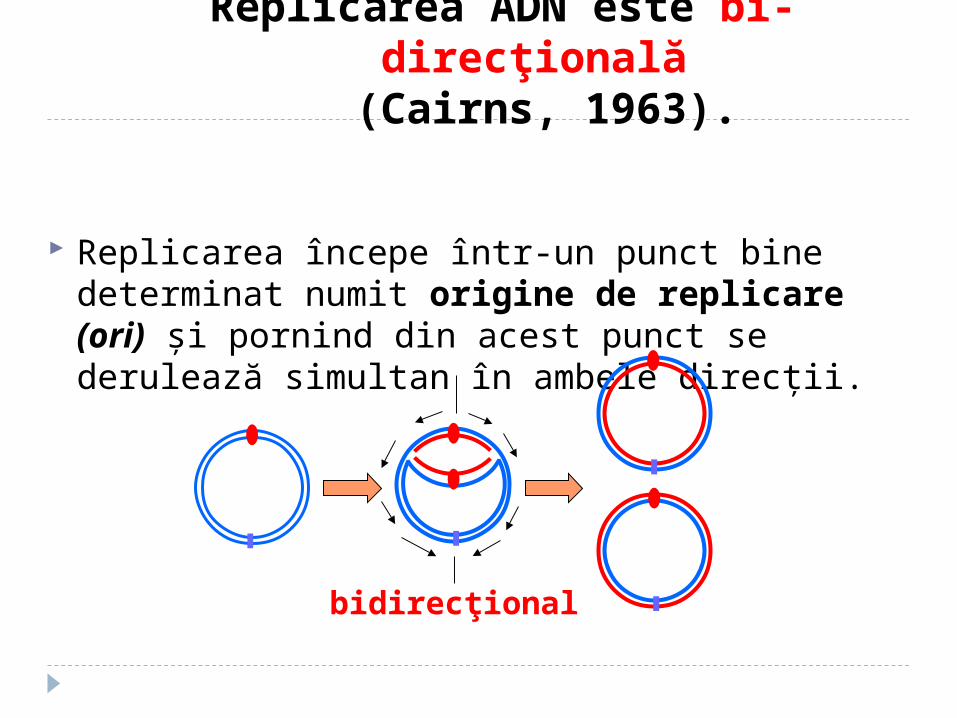
Replicarea ADN este bi-direcţională
(Cairns, 1963).
Replicarea începe într-un punct bine determinat numit origine de replicare (ori) şi pornind din acest punct se derulează simultan în ambele direcţii.
bidirecţional
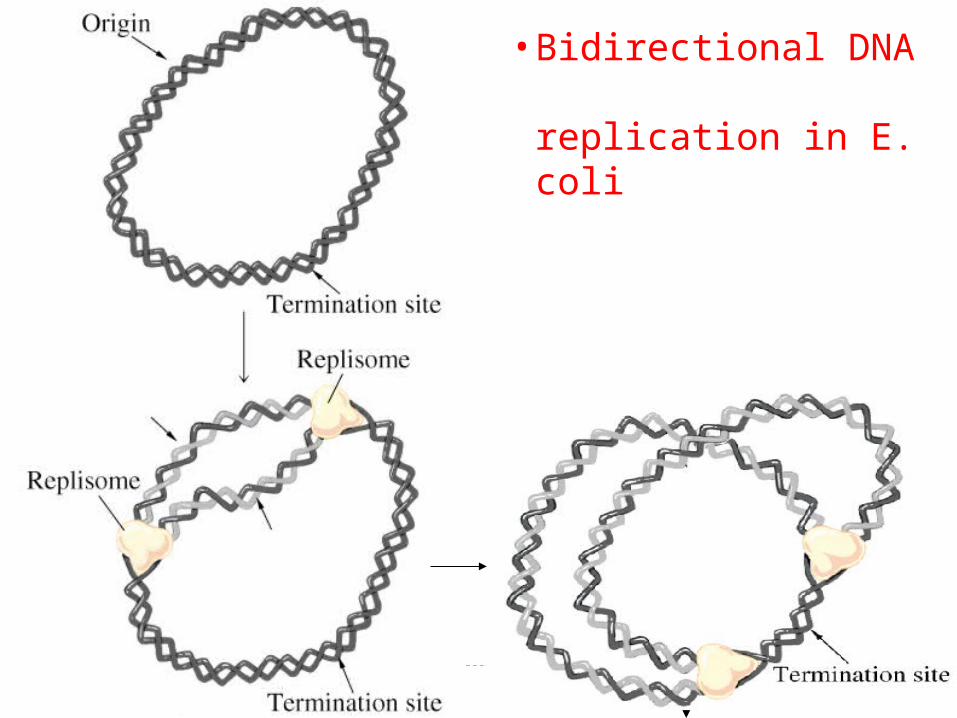
•Bidirectional DNA replication in E. coli
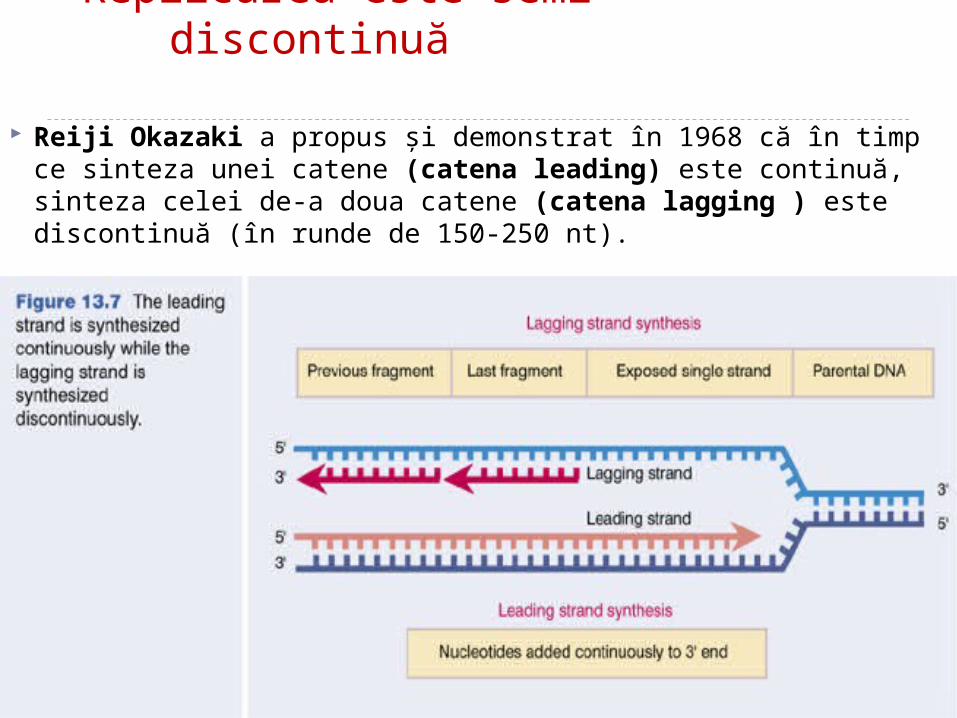
Replicarea este semi-discontinuă
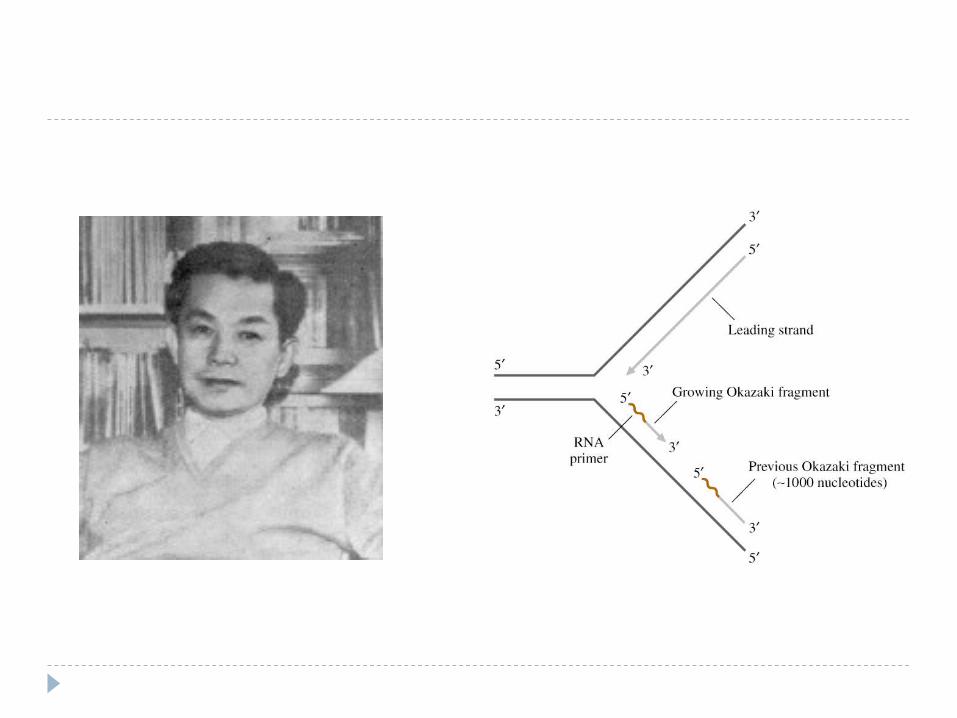
Reiji Okazaki a propus şi demonstrat în 1968 că în timp ce sinteza unei catene (catena leading) este continuă, sinteza celei de-a doua catene (catena lagging ) este discontinuă (în runde de 150-250 nt).

Caracteristicile:
prezenţa praimerului este obligatorie replicarea este cuplată cu desfăşurarea
DNA parental (necesită energie) replicarea decurge în ambele direcţii cu
aceeaşi viteză. Pe catena întîrziată se sintetizează
fragmentele Okazaki.

Caracteristicile:
Este bazată pe împachetarea complementară a BA
Catena-fiică este antiparalelă cu catena parentală dar nu identică după secvenţa nucleotidică
Forţa motrice a procesului este hidroliza pirofosfatului

Componentele necesare replicării:
1. Matriţă - ADN bicatenar2. Substrat:a. dezoxiribonucleozid trifosfaţii: dATP, TTP, dGTP,
dCTP; b. ribonucleozid trifosfaţii: ATP, GTP, CTP, UTP3. prezenţa ionilor de Mg, Mn; Zn4. Sistemul multienzimatic complex

Sistemul multienzimatic complex
1. Helicaza (Proteina DnaB) - desfacerea dublului helix, treptat, pe porţiuni mici (necesită energie; consumă 2 molecule de ATP per 2 legături de hidrogen desfăcute).
2. Single stranded proteins (SSBs) - se leagă de cele două catene ADN şi le menţine separate

Sistemul multienzimatic complex
3. 3. TopoisomeraTopoisomerazelezele - I şi II – înlătură supertorsiunile ADN, rezolvă problemele topologice apărute în cursul desfacerii lui.
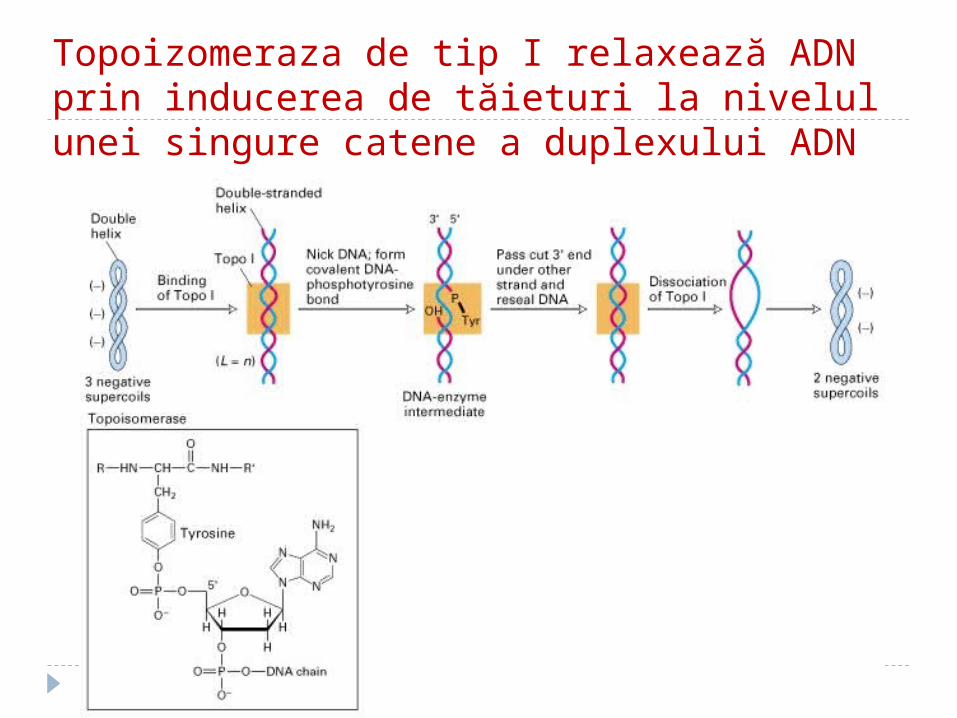
Topoizomeraza de tip I relaxează ADN prin inducerea de tăieturi la nivelul unei singure catene a duplexului ADN
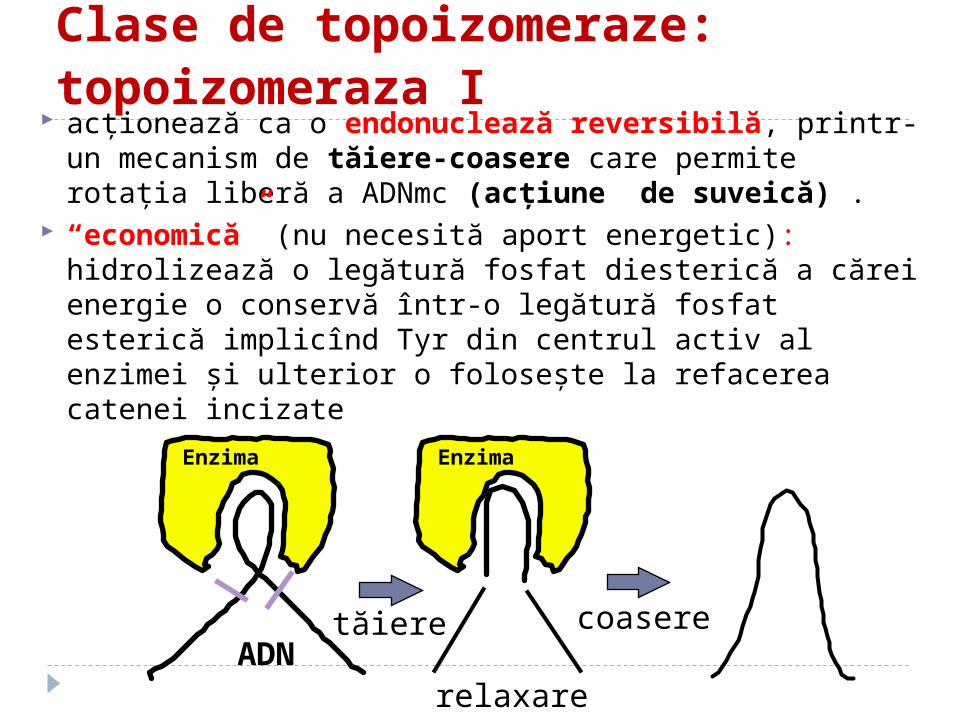
Clase de topoizomeraze: topoizomeraza I
acţionează ca o endonuclează reversibilă, printr-un mecanism de tăiere-coasere care permite rotaţia liberă a ADNmc (acţiune de suveică) .
“economică” (nu necesită aport energetic): hidrolizează o legătură fosfat diesterică a cărei energie o conservă într-o legătură fosfat esterică implicînd Tyr din centrul activ al enzimei şi ulterior o foloseşte la refacerea catenei incizate
Enzima
ADN
Enzima
tăiere
relaxare
coasere

topoizomeraza II
ADN-giraza Introduce în molecula de ADN supertorsiuni (-), spre
stînga, în sens contrar celor create de înaintarea bifurcaţiei de replicare; astfel compesează în avans constrîngerile impuse de desrăsucirea duplexului de ADN
Necesită energie (consum de ATP)

ADN giraza înlătură super-răsuricirile pozitive care apar în mod normal la nivelul furcii de replicare
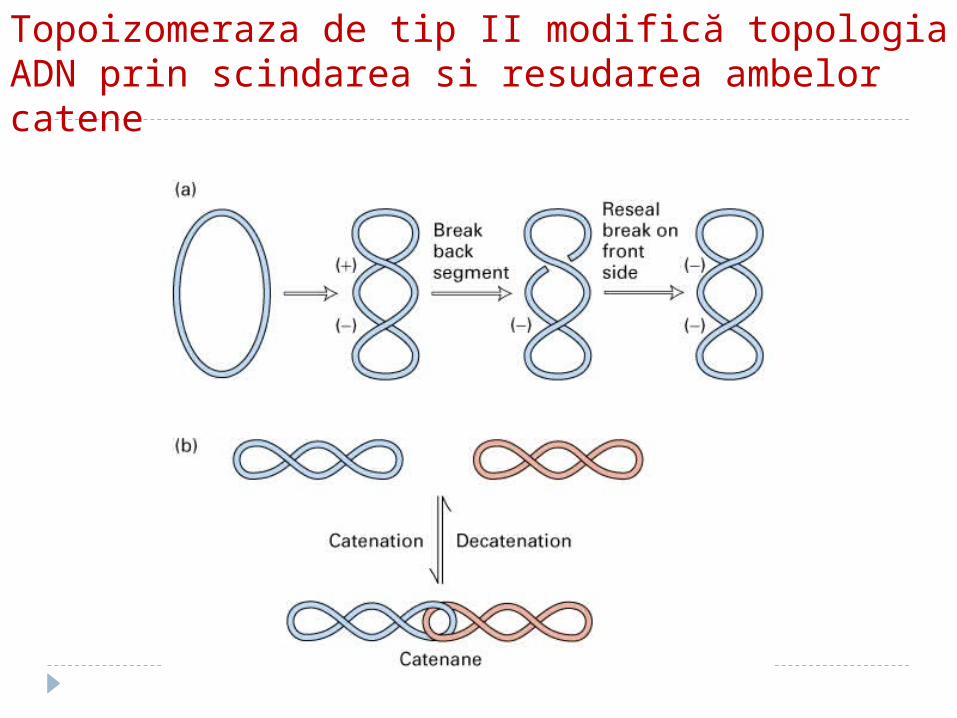
Topoizomeraza de tip II modifică topologia ADN prin scindarea si resudarea ambelor catene
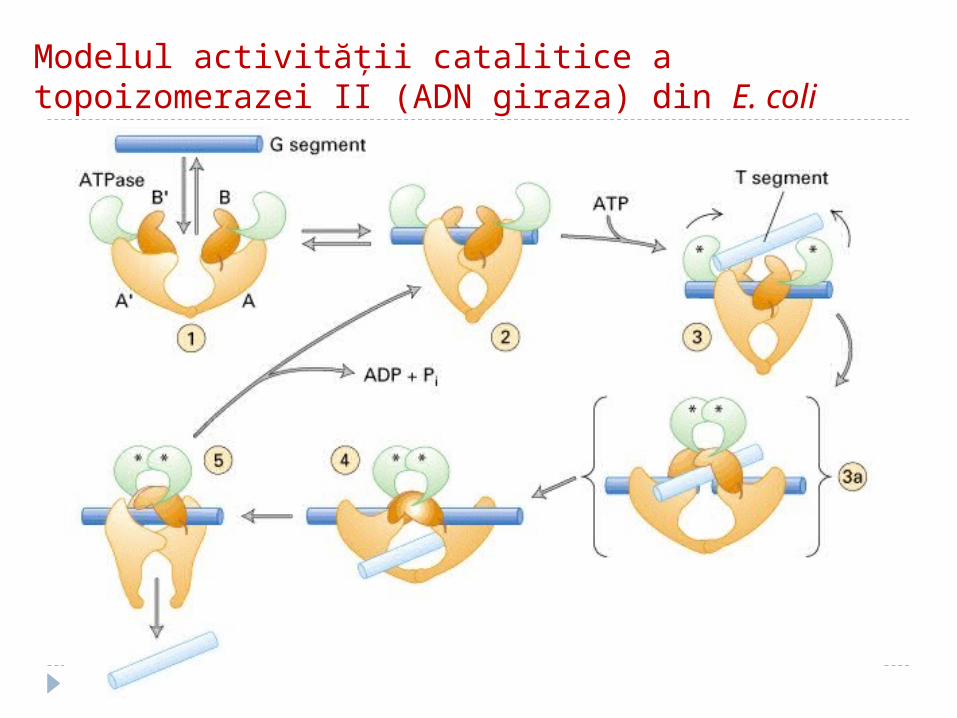
Modelul activităţii catalitice a topoizomerazei II (ADN giraza) din E. coli

Sistemul multienzimatic complex
4. 4. . P. Primază - sintetizează primerul în direcţia 5'- 3‘ .

Sistemul multienzimatic complex
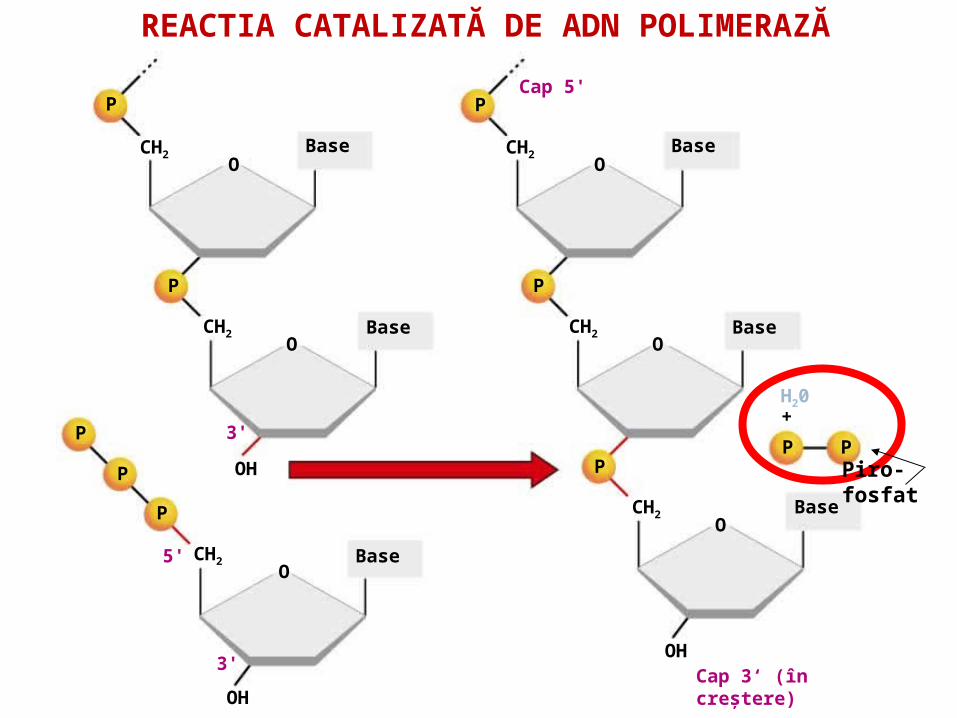
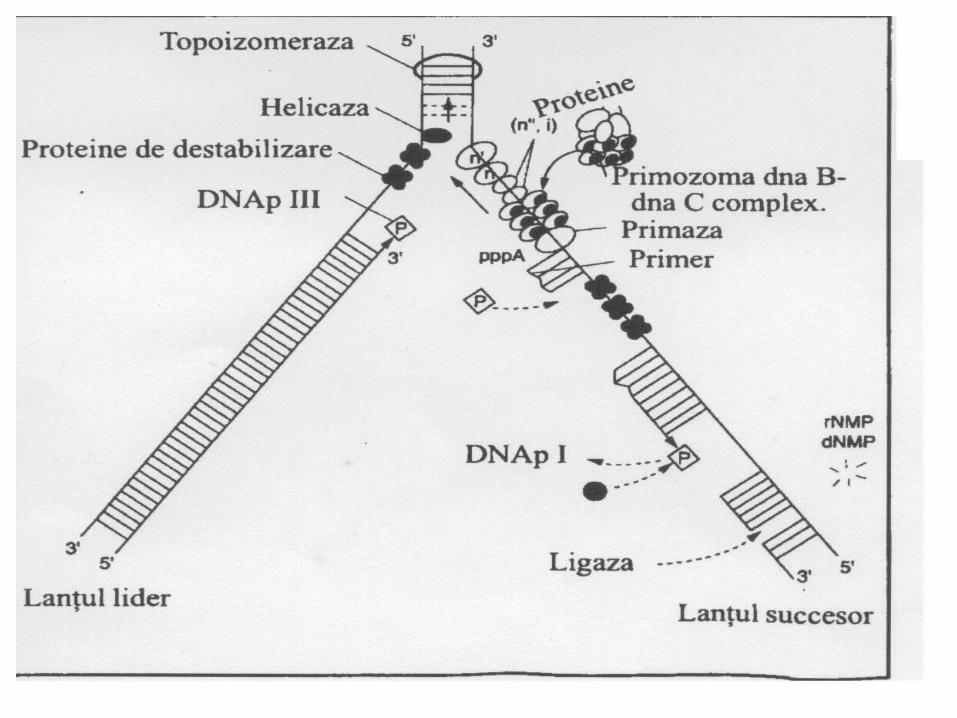
55. . ADN polimeraza ( ADNp I, II, III)- sinteza catenei fiice în direcţia 5'→3' . - ADN p III: – acţiune polimerazică (5'- 3‘)- acţiune exonucleazică (3'- 5‘) - ADN p II - este implicată în reparea leziunilor ADN (are activitate 3' -> 5‘ exonucleazică. - ADN p I - posedă activitate 5'- 3‘ exonucleazică, înlătură primerul şi-l înlocuieşte cu fragmente de ADN
P
P
P
P
P
P
P
PP P
CH2
CH2
CH2
OH
OH
O
O
OBase
Base
Base
CH2
CH2
CH2
OH
O
O
OBase
Base
Base
Cap 5'
Cap 3‘ (în creştere)3'
5'
3'
H20+
REACTIA CATALIZATĂ DE ADN POLIMERAZĂ
Piro-fosfat

ADN polimerazele la procariote (continuare)
ADN pol IV- Pol IV este o ADN polimerază predispusă la erori de împerechere (nu are activitate de proof-reading ca la pol III, pol II şi pol I)
- Pol IV este responsibilă de 50% din mutaţiile adaptative.
ADN pol V- Pol V aparţine unei căi speciale de reparare (un fel de “ultima barieră“) denumită calea SOS de reparare a ADN.- Pol V este sintetizată cînd celula este expusă la doze mari de radiaţii sau de mutageni, cînd pot să apară lez. majore ADN.

Sistemul multienzimatic complex
6. ADN ligaza- uneşte fragmentele Okazaki de pe catena întîrziată. Catalizează formarea unei legături fosfat diesterice între 3'-OH a unui fragment de ADN şi extremitatea 5' monofosfat al altuia.

Enzimele implicate în replicarea ADN
Helicase desface dublu helix
SSB stabilizează ADNmc
ADN polimeraza IIIleagă nucleotidele pt. a forma noul ADN
Ligaza leagă Fragmentele Okazaki).
Primaza sintetizează un ARN primer pe matriţă de ADN
ADN polimeraza I (exonucleaza) înlocuieşte ARN primercu ADN
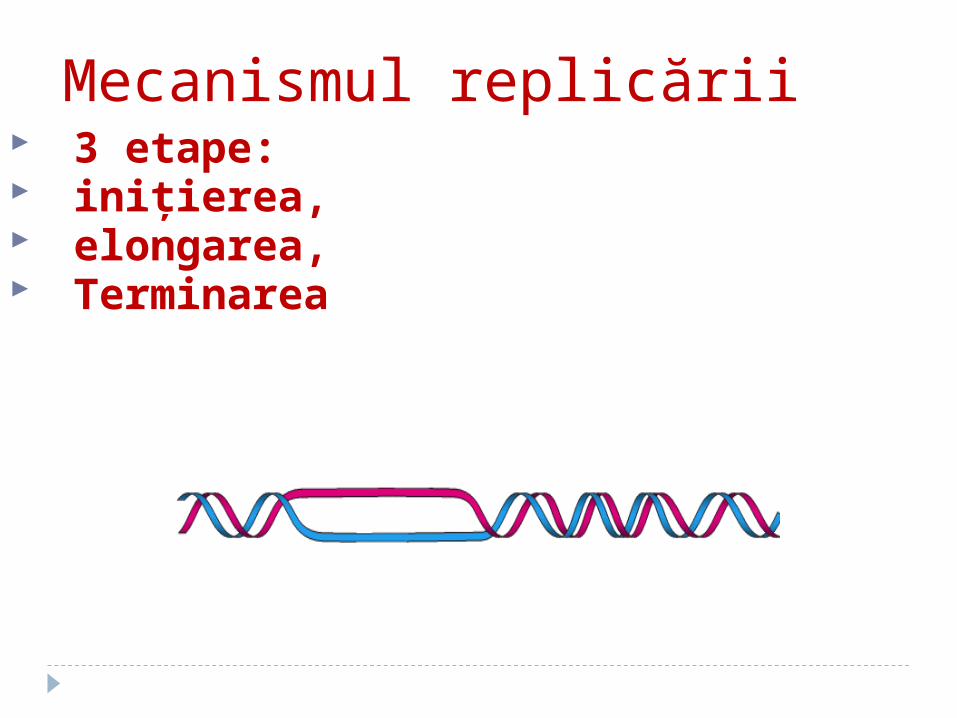
Mecanismul replicării 3 etape: iniţierea, elongarea, Terminarea

ETAPELE REPLICĂRII ADN(şi elementele-cheie ale fiecărei etape)
Iniţierea: primozomul- un complex de proteine şi originea replicării ori
Elongarea: replizomul- un complex de proteine, asociate cu ADN cu rol în sinteza
catenelor noi de ADNTerminarea: proteina Tus - situsul de terminare ter
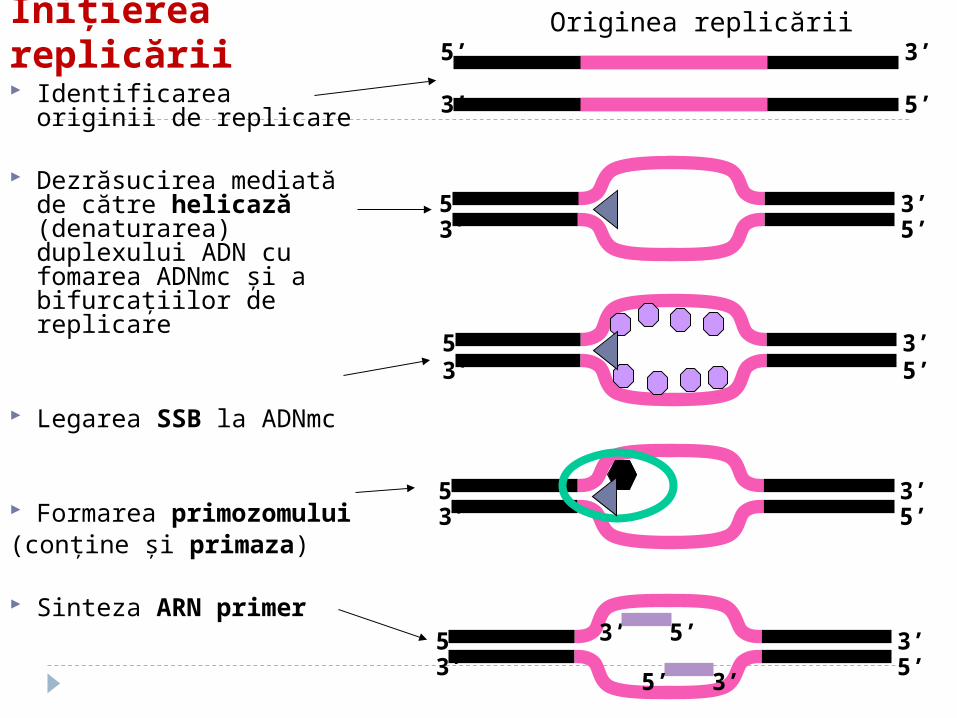
Iniţierea replicării Identificarea originii de
replicare
Dezrăsucirea mediată de către helicază (denaturarea) duplexului ADN cu fomarea ADNmc şi a bifurcaţiilor de replicare
Legarea SSB la ADNmc
Formarea primozomului (conţine şi primaza)
Sinteza ARN primer
Originea replicării
5’
3’
3’
5’
5’5’3’
3’
5’5’3’
3’
5’5’3’
3’
5’5’3’
3’3’ 5’
3’5’

Originea replicării Originea replicării
(oriC) este o regiune de 245 perechi baze bogată în AT
OriC este recunoscută de către proteina dnaA care se leagă la oriC, şi care împreună cu proteina dnaB (helicaza) desface ADNdc (folosind ATP).
oriC

Iniţierea parcurge două etape:
a. Formarea furcii de replicaţie- sub acţiunea helicazelor are loc desfacerea duplexului parental (la scindarea leg. de H dintre BA - se utilizează min 2 mol. de ATP).
La desfacerea duplexului parental apar regiuni superhelicoidale, care se reglează cu ajutorul girazei (topoizomerazei).
Topoizomeraza efectuează rupturi monocatenare apoi sudează legătura fosfodiesterică şi favorizează relaxarea structurii DNA
b. Sinteza primerului
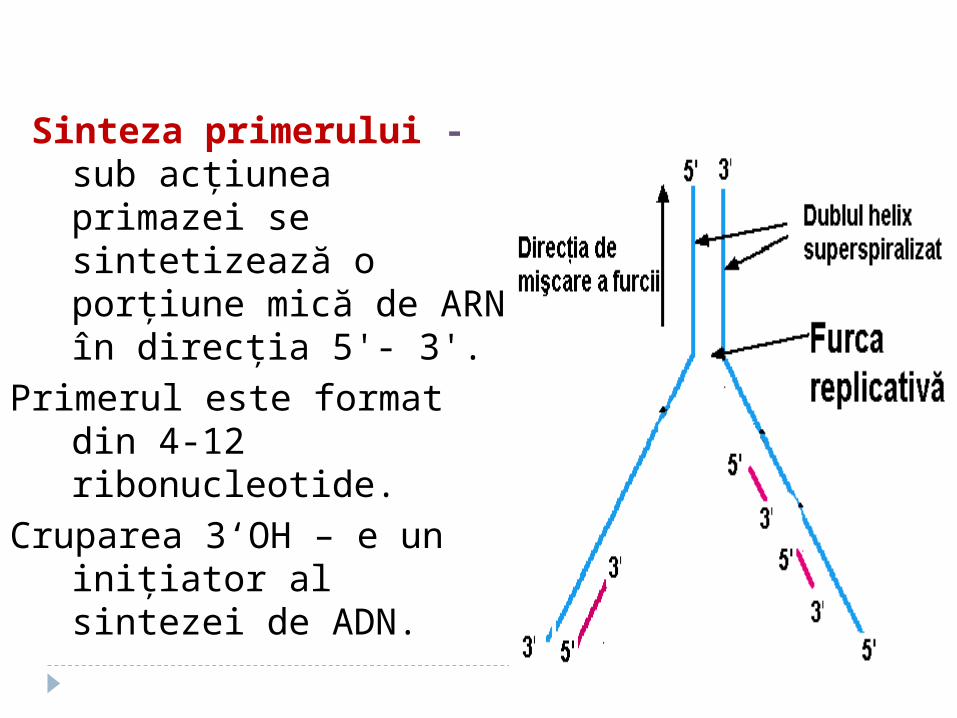
Sinteza primerului - sub acţiunea primazei se sintetizează o porţiune mică de ARN în direcţia 5'- 3'.
Primerul este format din 4-12 ribonucleotide.
Cruparea 3‘OH – e un iniţiator al sintezei de ADN.

2. ETAPA DE ELONGARE
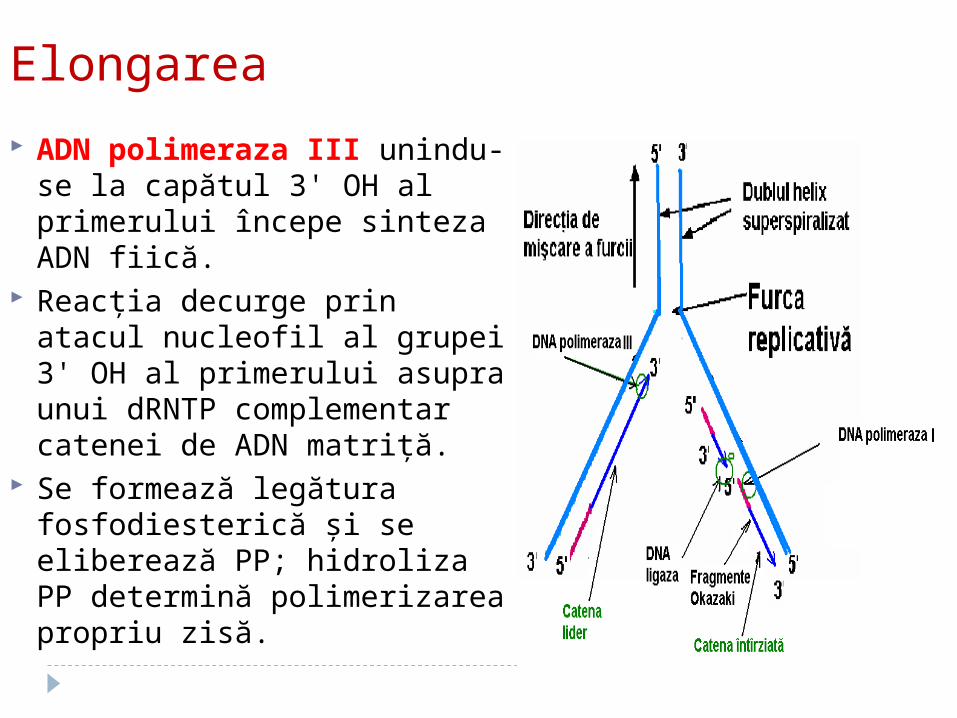
Elongarea
ADN polimeraza III unindu-se la capătul 3' OH al primerului începe sinteza ADN fiică.
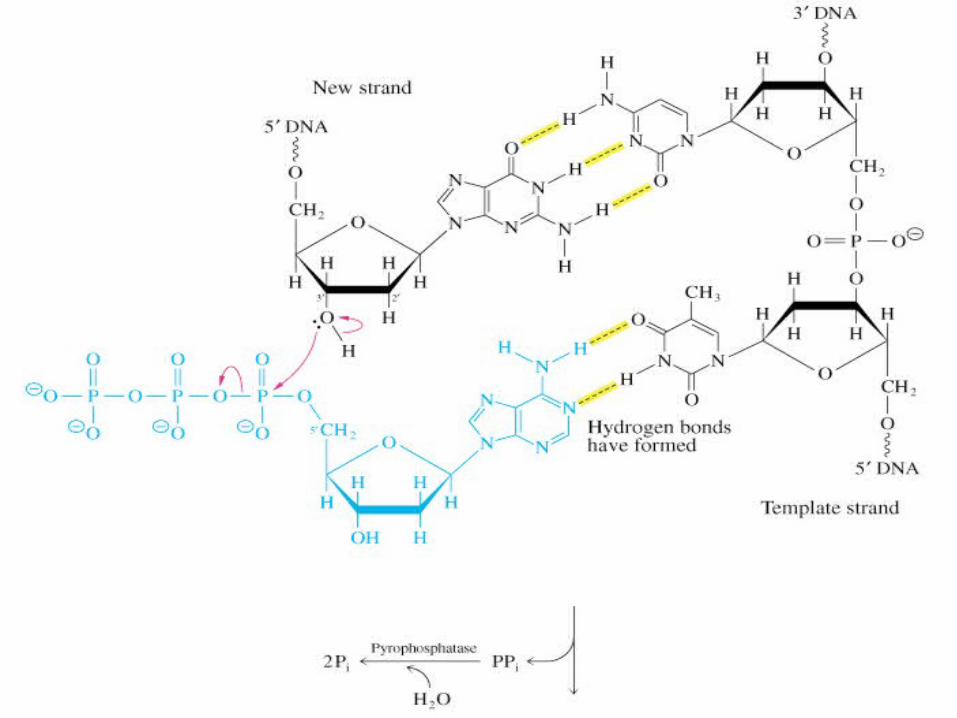
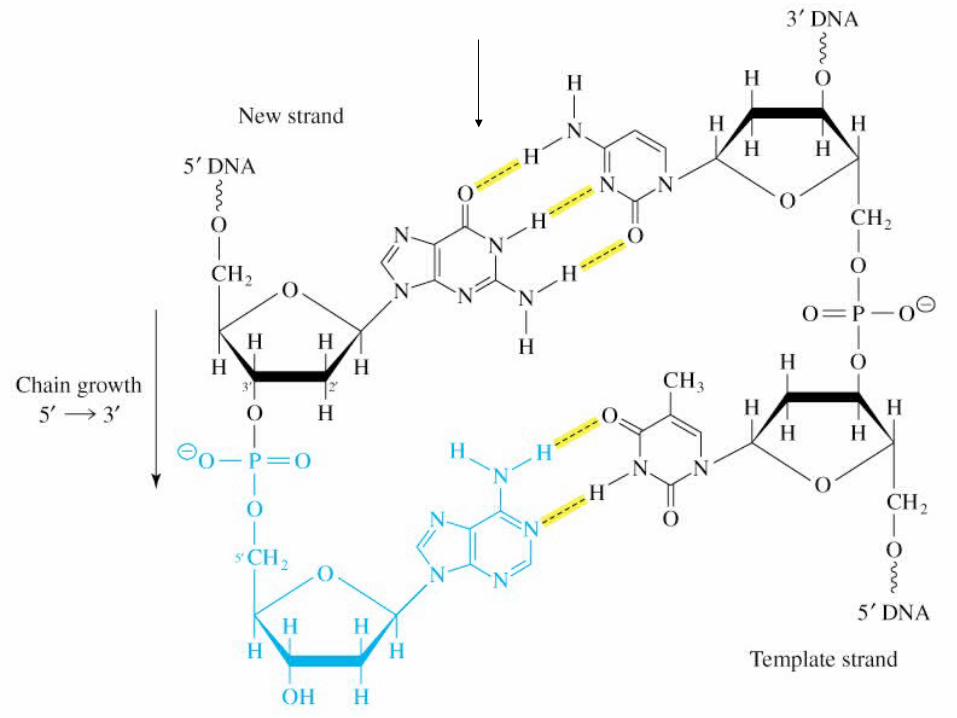
Reacţia decurge prin atacul nucleofil al grupei 3' OH al primerului asupra unui dRNTP complementar catenei de ADN matriţă.
Se formează legătura fosfodiesterică şi se eliberează PP; hidroliza PP determină polimerizarea propriu zisă.
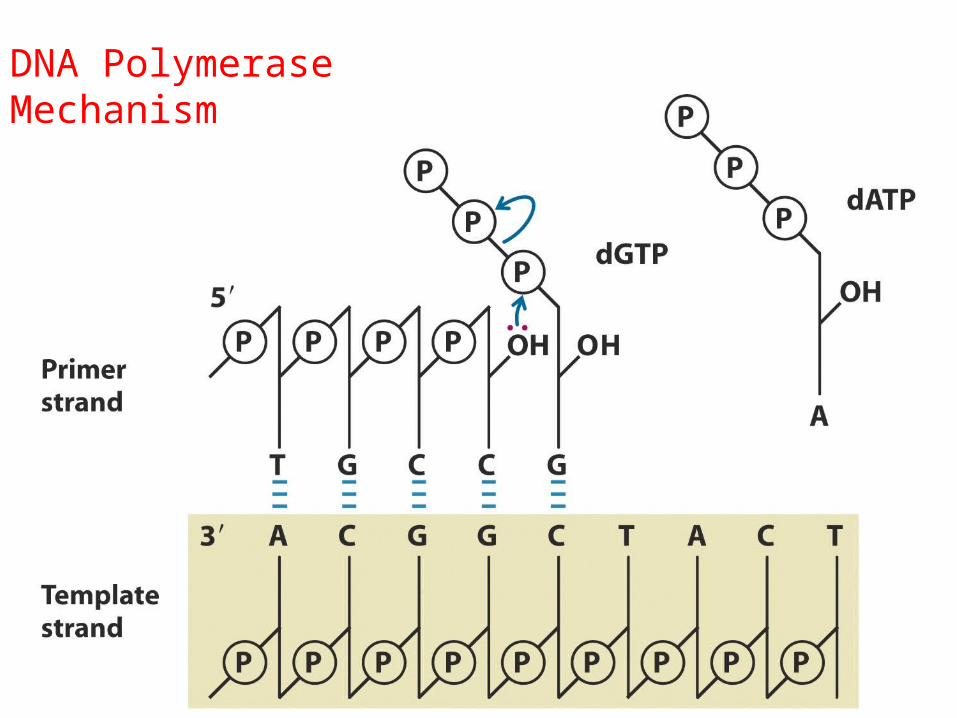
DNA Polymerase Mechanism
Elongarea
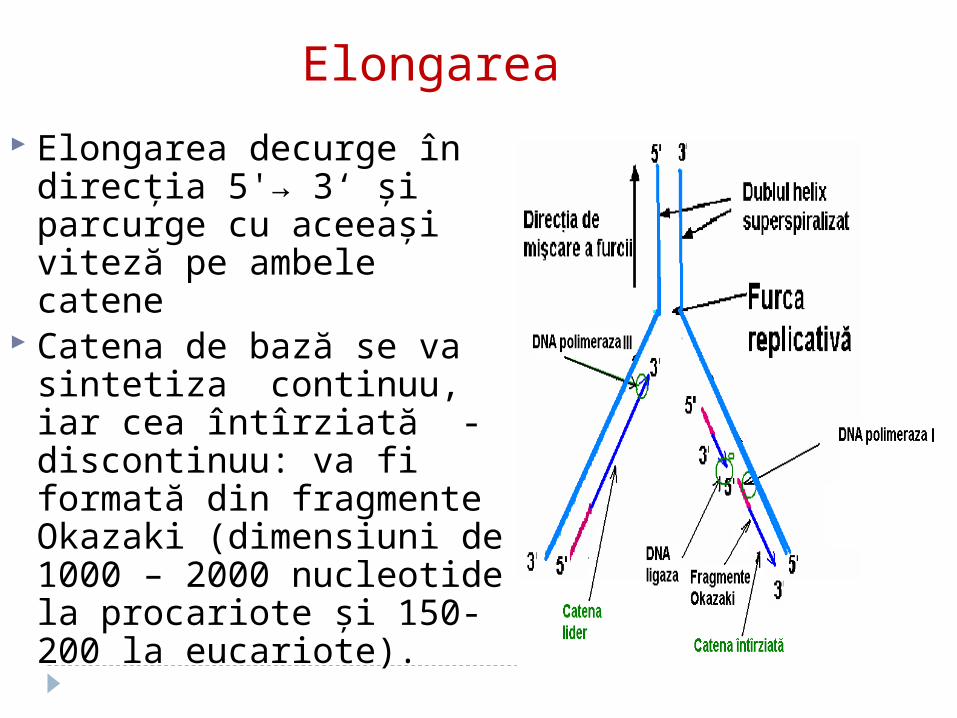
Elongarea decurge în direcţia 5'→ 3‘ şi parcurge cu aceeaşi viteză pe ambele catene
Catena de bază se va sintetiza continuu, iar cea întîrziată - discontinuu: va fi formată din fragmente Okazaki (dimensiuni de 1000 – 2000 nucleotide la procariote şi 150-200 la eucariote).
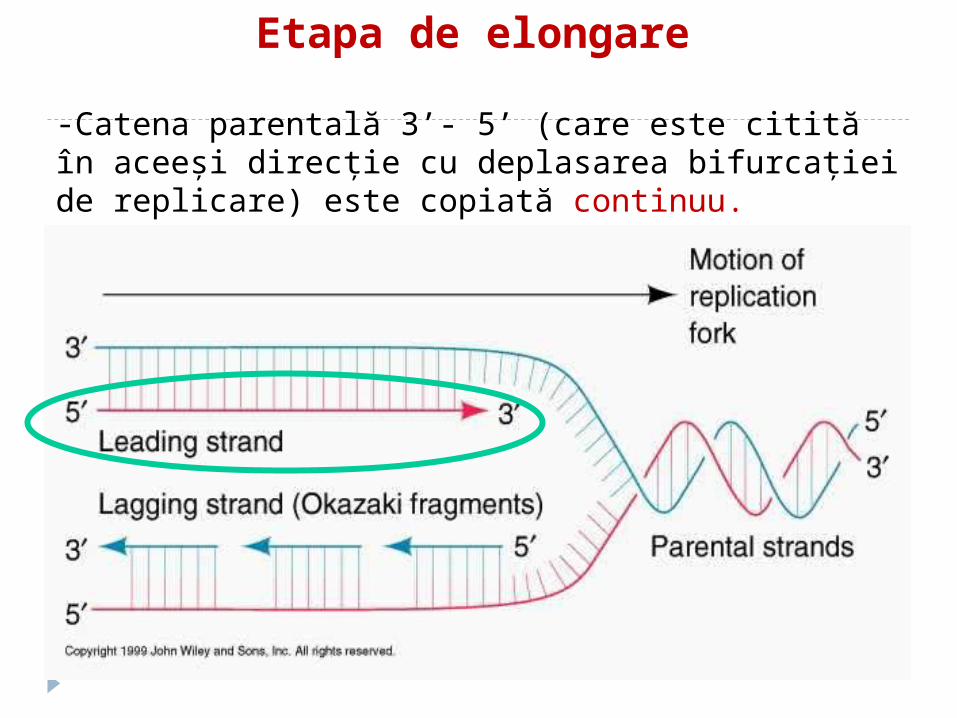
Etapa de elongare
-Catena parentală 3’- 5’ (care este citită în aceeşi direcţie cu deplasarea bifurcaţiei de replicare) este copiată continuu.-Catena nouă astfel sintetizată este catena “leading”.
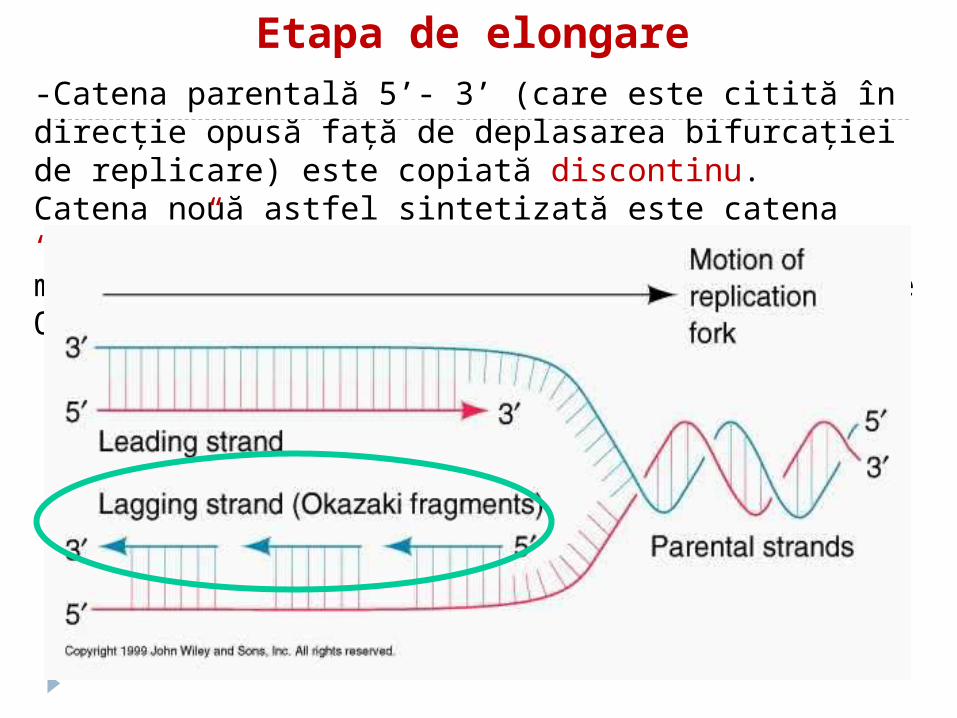
Etapa de elongare -Catena parentală 5’- 3’ (care este citită în direcţie opusă faţă de deplasarea bifurcaţiei de replicare) este copiată discontinu.Catena nouă astfel sintetizată este catena “lagging” şi este sintetizată în fragmente mici (1fragment per rundă sinteză)-fragmentele Okazaki
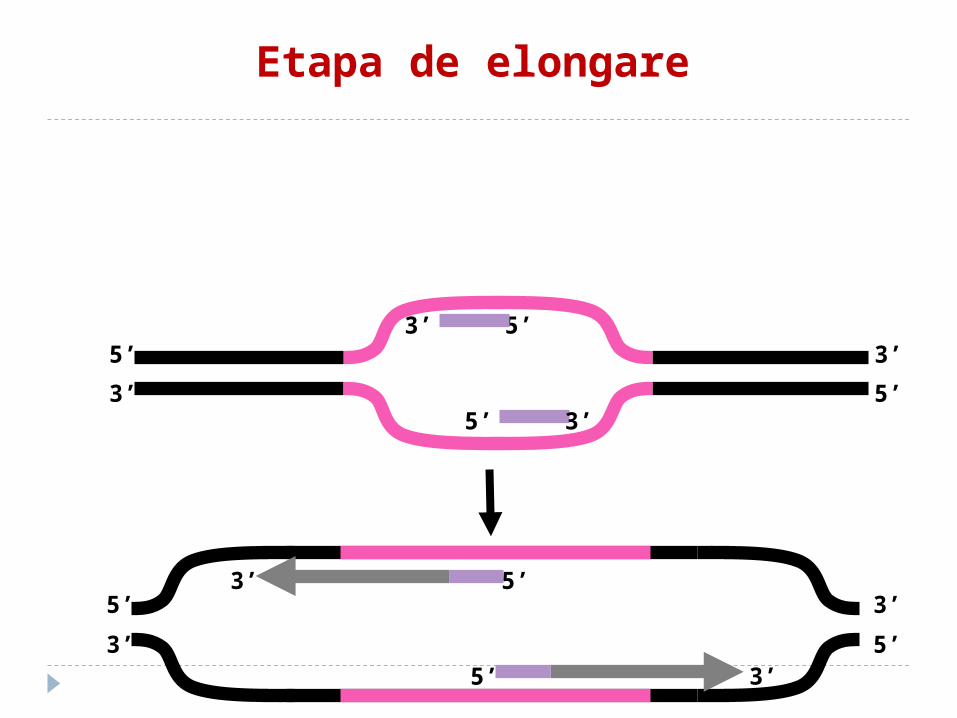
Etapa de elongare
5’
5’
3’
3’
5’3’
5’
5’
5’
3’
3’3’
3’ 5’
3’5’

Etapa de elongare ADN polimeraza I exclude primerii şi sintetizează
complementar ADN. Fragmentele sînt unite cu enzima ADN ligaza
(necesită ATP la eucariote şi NAD la procariote).
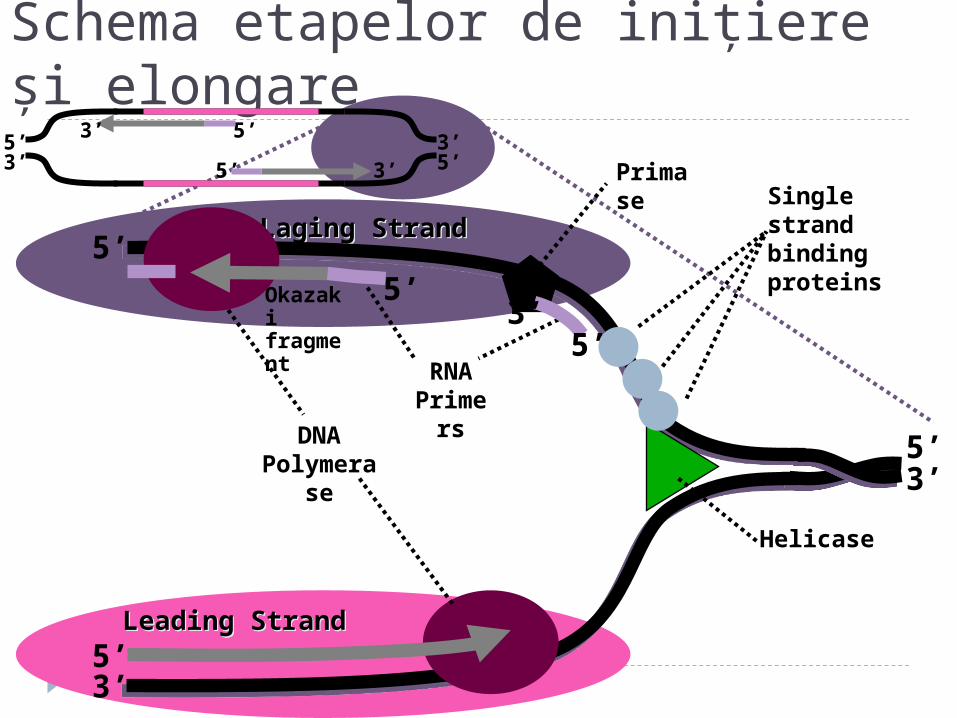
Leading StrandLeading Strand
Laging StrandLaging Strand
3’
5’3’
5’
Schema etapelor de iniţiere şi elongare
5’
5’5’3’
3’
5’3’3’
5’
Single strand binding proteins
DNA Polymerase
Okazaki fragment
RNA Primers
Primase
5’3’
5’
Helicase

3. ETAPA DE TERMINARE
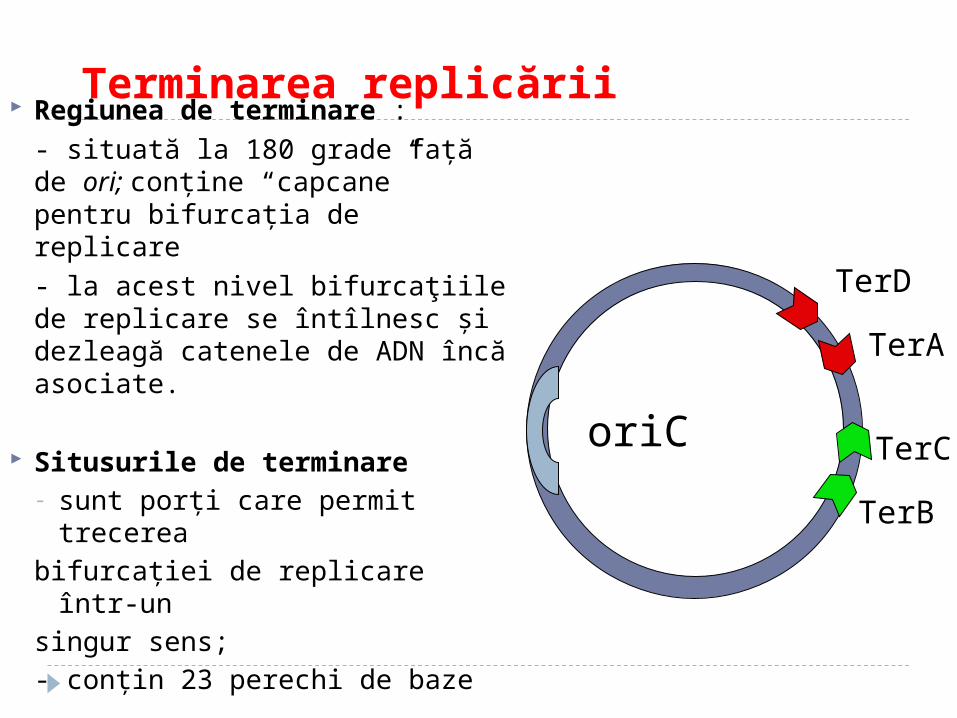
Terminarea replicării Regiunea de terminare :
- situată la 180 grade faţă de ori; conţine “capcane” pentru bifurcația de replicare
- la acest nivel bifurcaţiile de replicare se întîlnesc şi dezleagă catenele de ADN încă asociate.
Situsurile de terminare- sunt porţi care permit trecerea
bifurcaţiei de replicare într-un
singur sens;
- conţin 23 perechi de baze
oriC
TerD
TerC
TerB
TerA
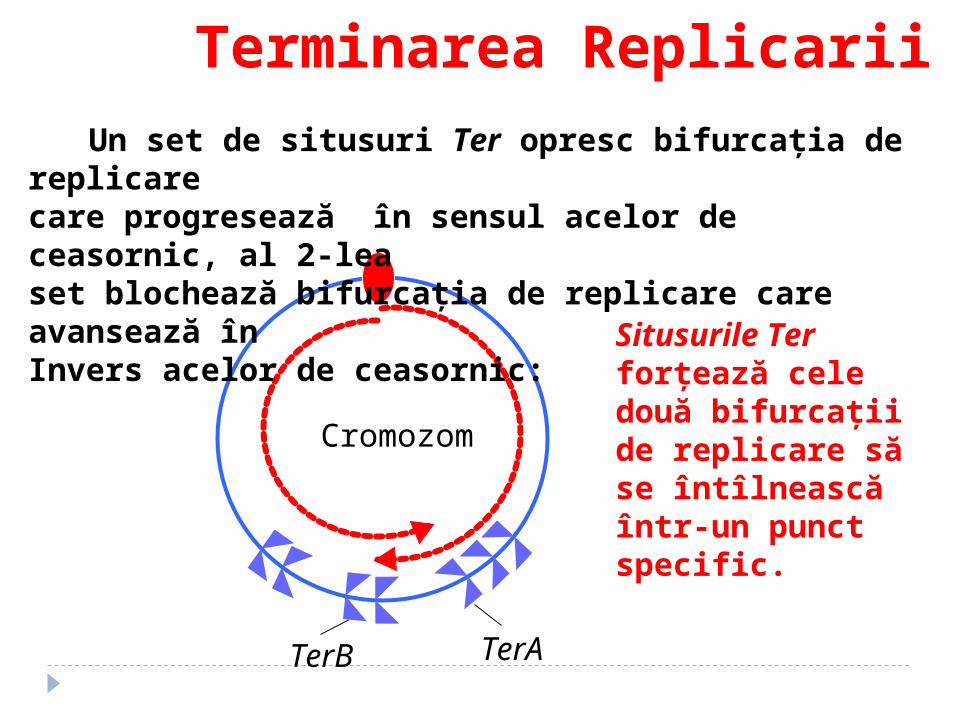
Terminarea Replicarii
TerATerB
Un set de situsuri Ter opresc bifurcaţia de replicare care progresează în sensul acelor de ceasornic, al 2-leaset blochează bifurcaţia de replicare care avansează în Invers acelor de ceasornic:
Cromozom
Situsurile Ter forţează cele două bifurcaţii de replicare să se întîlnească într-un punct specific.
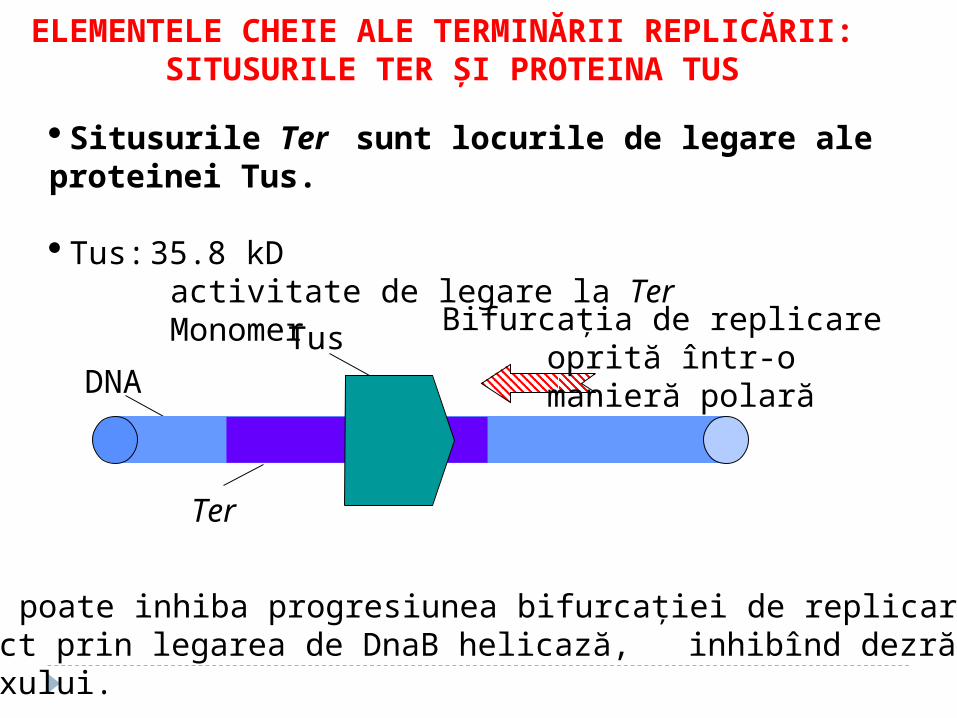
ELEMENTELE CHEIE ALE TERMINĂRII REPLICĂRII: SITUSURILE TER ŞI PROTEINA TUS
Situsurile Ter sunt locurile de legare ale proteinei Tus.
Tus: 35.8 kDactivitate de legare la TerMonomer
TusDNA
Ter
Bifurcaţia de replicare oprită într-o
manieră polară
Tus poate inhiba progresiunea bifurcaţiei de replicare direct prin legarea de DnaB helicază, inhibînd dezrăsucireahelixului.
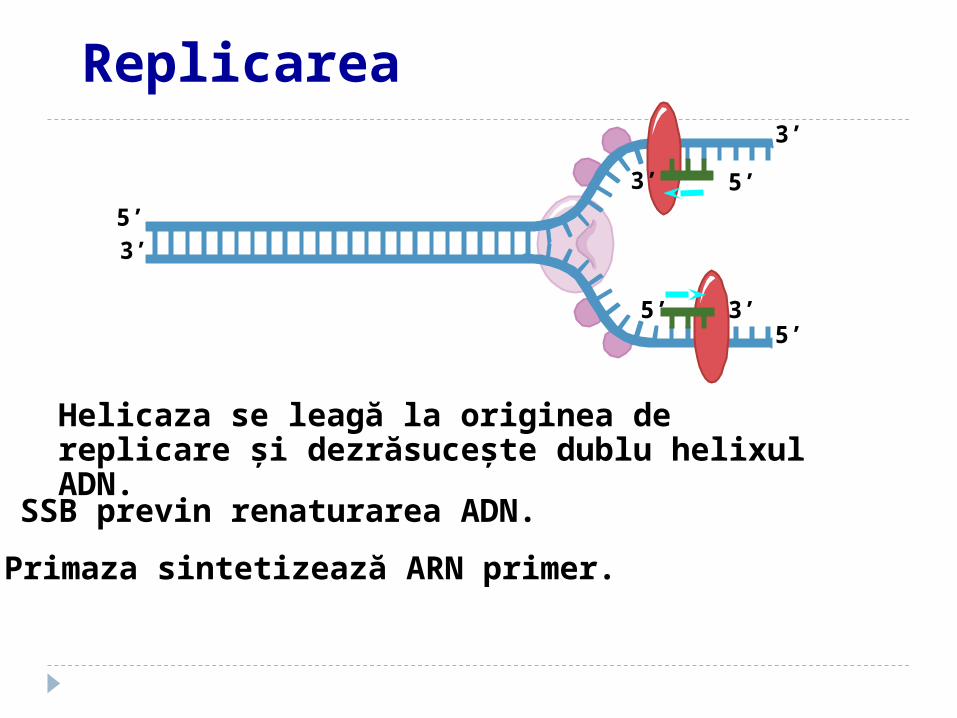
SSB previn renaturarea ADN.
Helicaza se leagă la originea de replicare şi dezrăsuceşte dublu helixul ADN.
5’ 3’
5’
3’
Primaza sintetizează ARN primer.
3’ 5’
5’ 3’
Replicarea
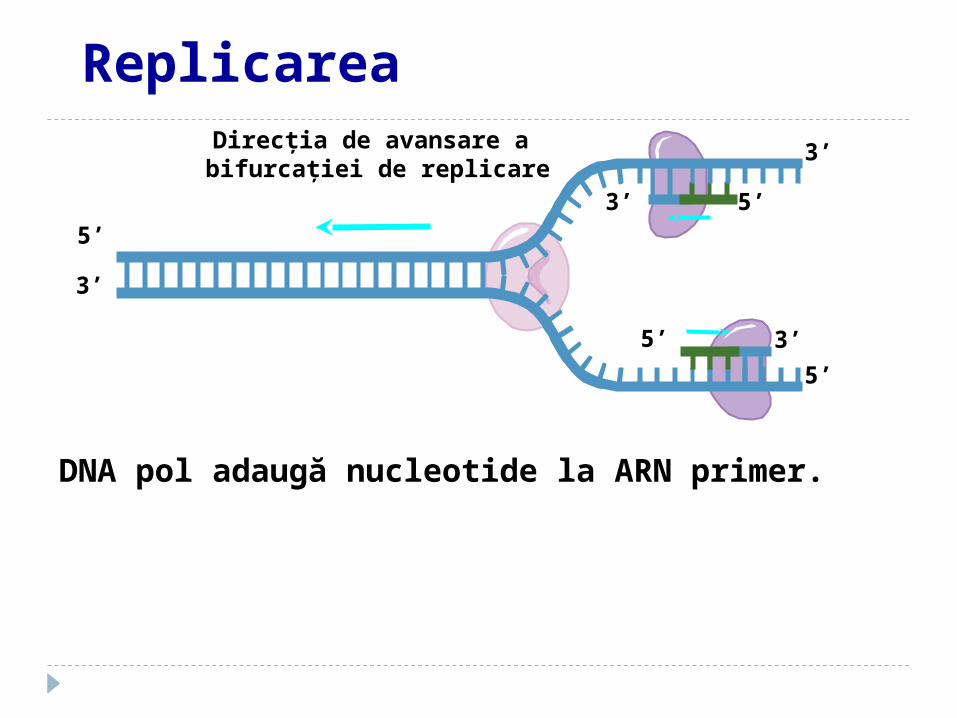
Direcţia de avansare a bifurcaţiei de replicare
5’ 3’
5’
3’
5’
3’
3’ 5’
DNA pol adaugă nucleotide la ARN primer.
Replicarea
5’
5’
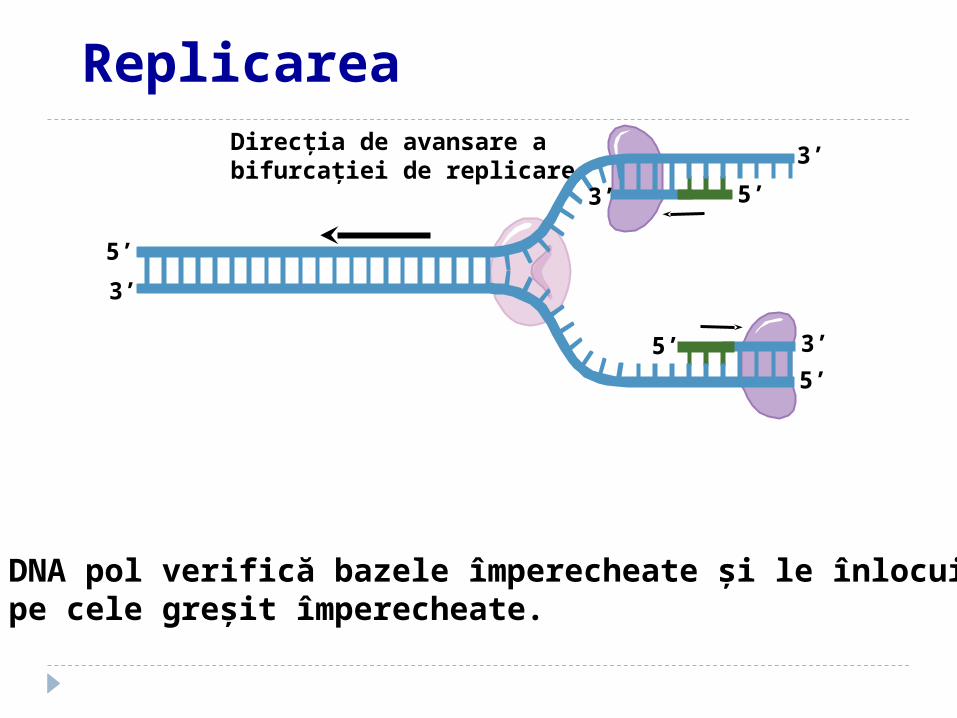
Direcţia de avansare a bifurcaţiei de replicare
5’
3’
5’
3’
3’
3’
DNA pol verifică bazele împerecheate şi le înlocuieşte pe cele greşit împerecheate.
Replicarea
5’
5’ 3’
5’
3’
3’
5’
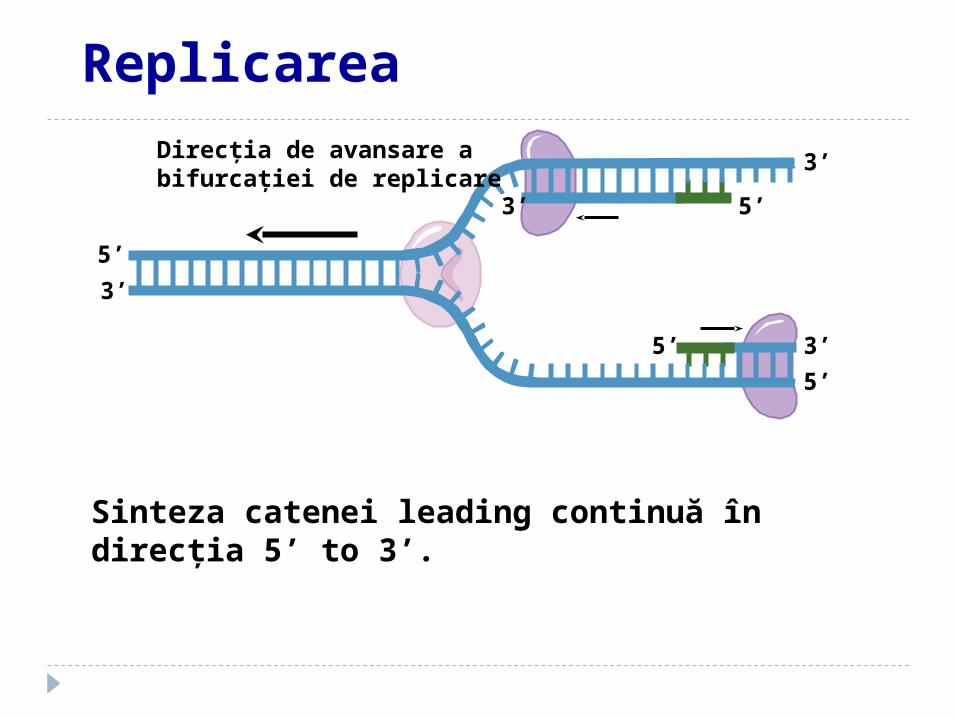
3’Direcţia de avansare a bifurcaţiei de replicare
Sinteza catenei leading continuă în direcţia 5’ to 3’.
Replicarea
3’ 5’ 5’
5’ 3’
5’
3’
3’
5’
3’Direcţia de avansare a bifurcaţiei de replicare
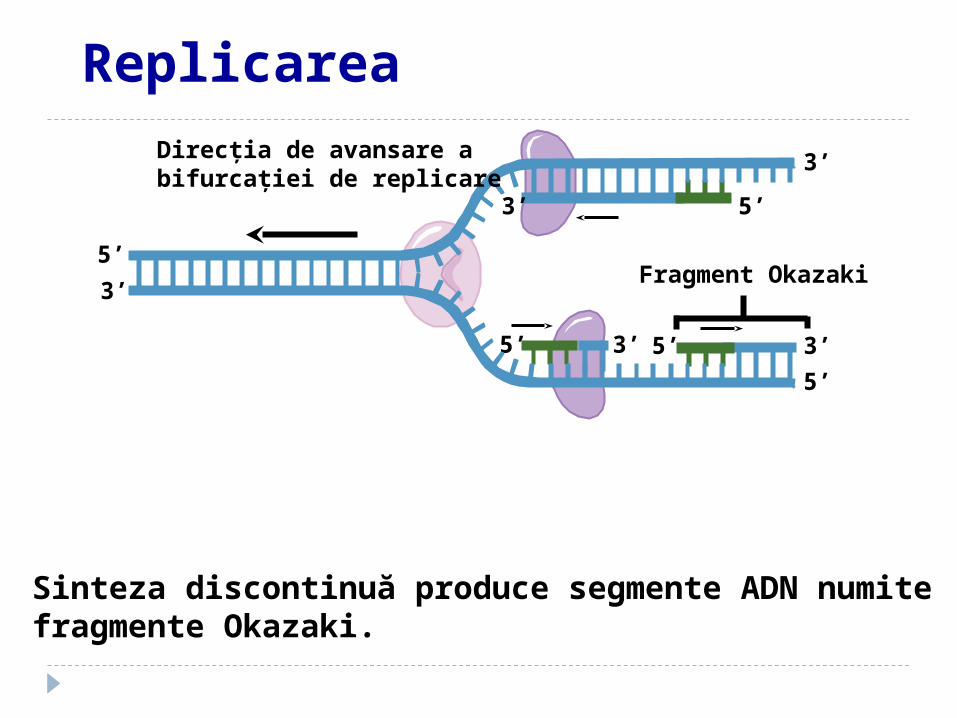
Fragment Okazaki
Sinteza discontinuă produce segmente ADN numite fragmente Okazaki.
Replicarea
5’ 5’
5’ 3’
5’
3’
3’
5’
3’Direcţia de avansare a bifurcaţiei de replicare
3’
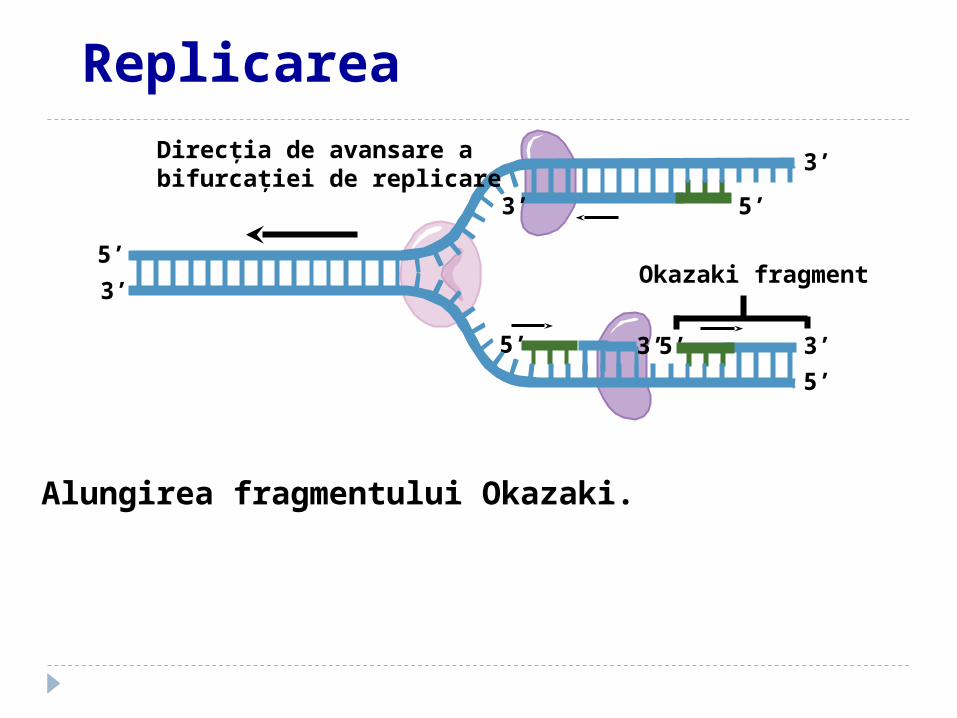
Alungirea fragmentului Okazaki.
Okazaki fragment
Replicarea
5’
5’ 3’
5’
3’
3’
5’
3’
3’
5’ 5’ 3’
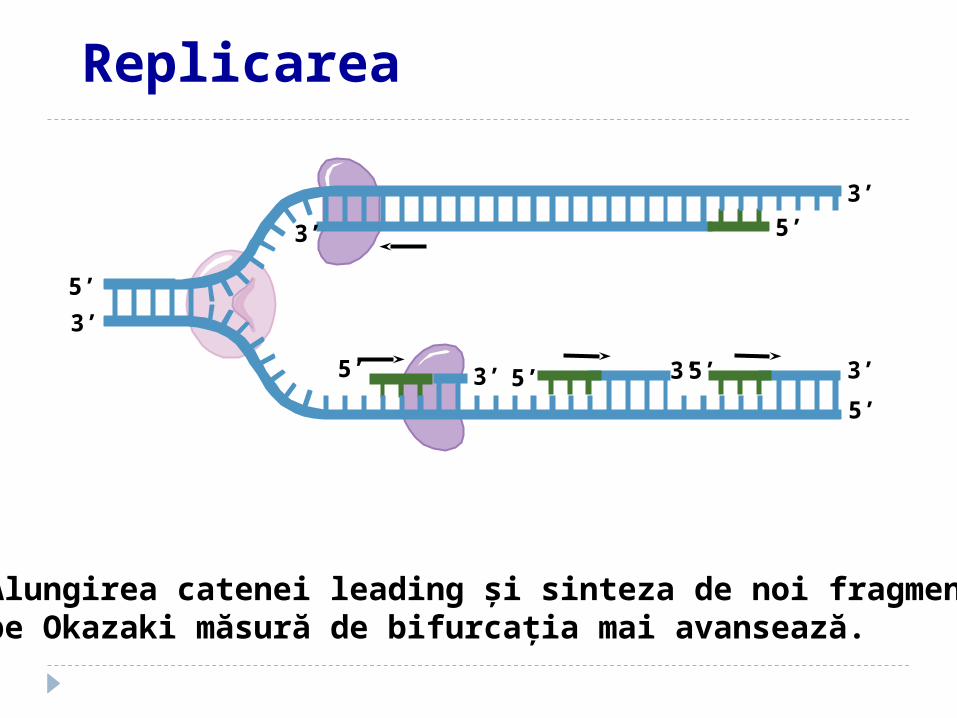
Alungirea catenei leading şi sinteza de noi fragmente pe Okazaki măsură de bifurcaţia mai avansează.
Replicarea
3’
5’
3’
5’
5’ 3’
5’
3’
3’
5’ 5’ 3’
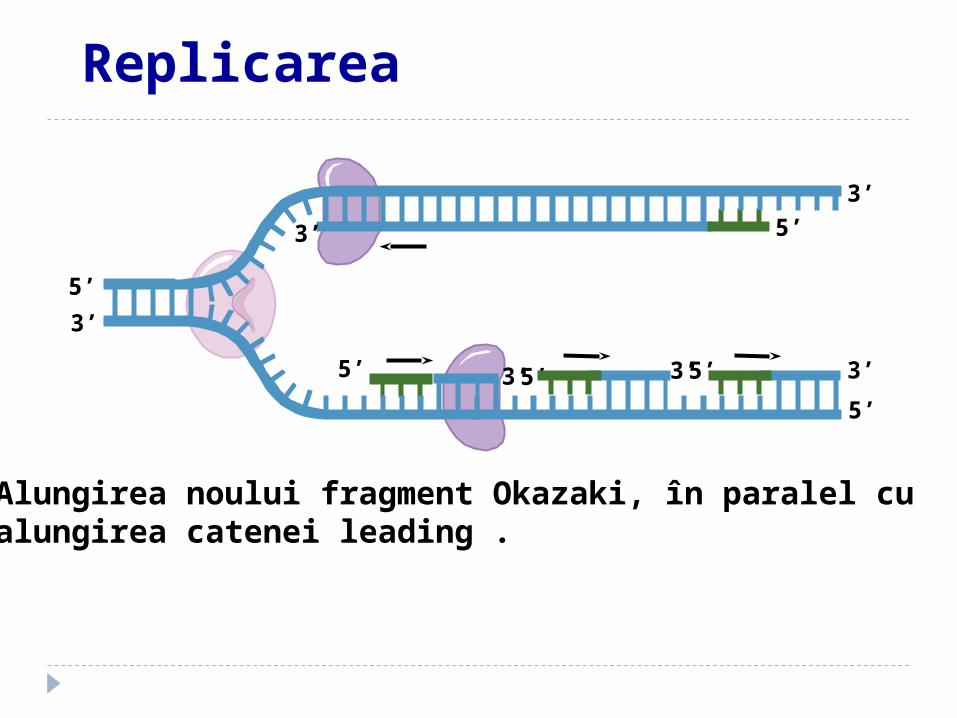
Alungirea noului fragment Okazaki, în paralel cu alungirea catenei leading .
Replicarea
5’
5’
3’ 3’
5’
3’
5’ 3’
5’
3’
3’
5’
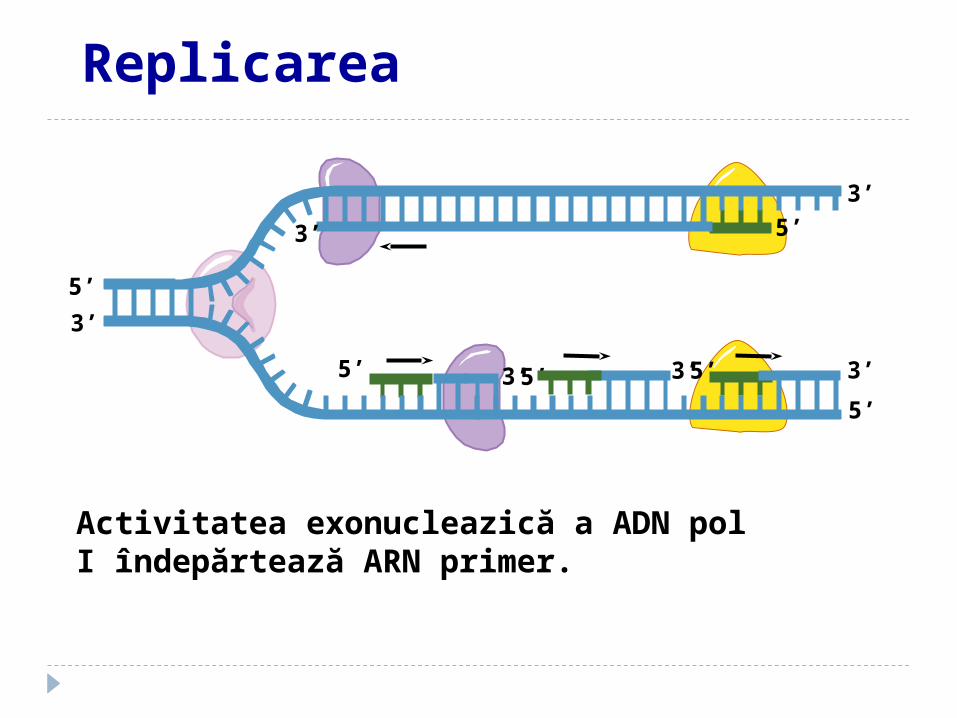
Activitatea exonucleazică a ADN pol I îndepărtează ARN primer.
Replicarea
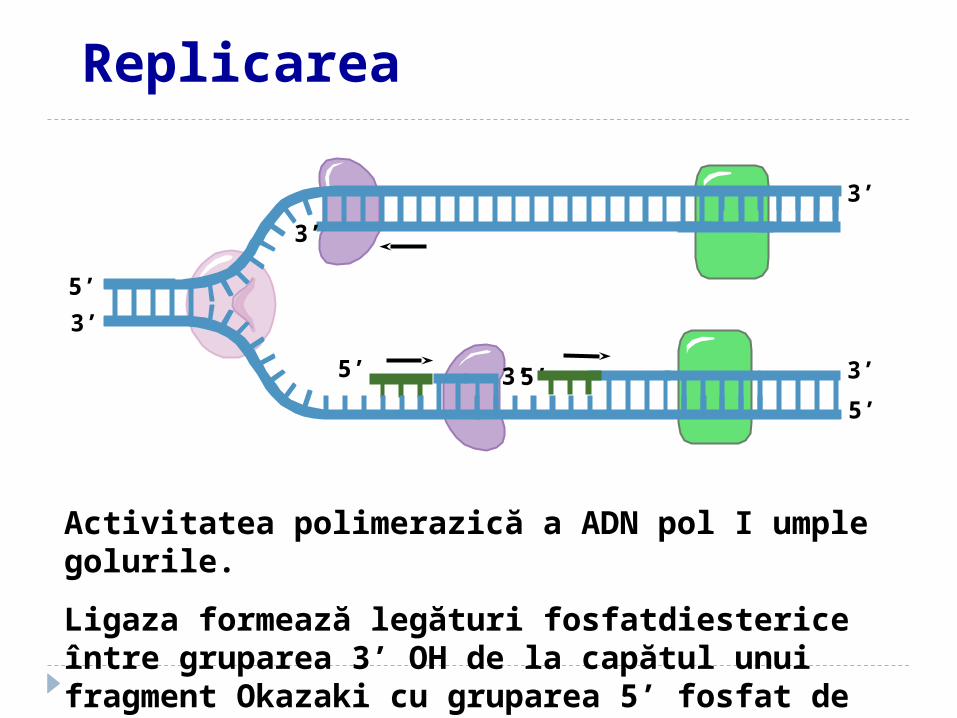
Activitatea polimerazică a ADN pol I umple golurile.
Ligaza formează legături fosfatdiesterice între gruparea 3’ OH de la capătul unui fragment Okazaki cu gruparea 5’ fosfat de la capătul altui fragment Okazaki. .
3’
5’
3’
5’ 3’
5’
3’
3’
5’
Replicarea
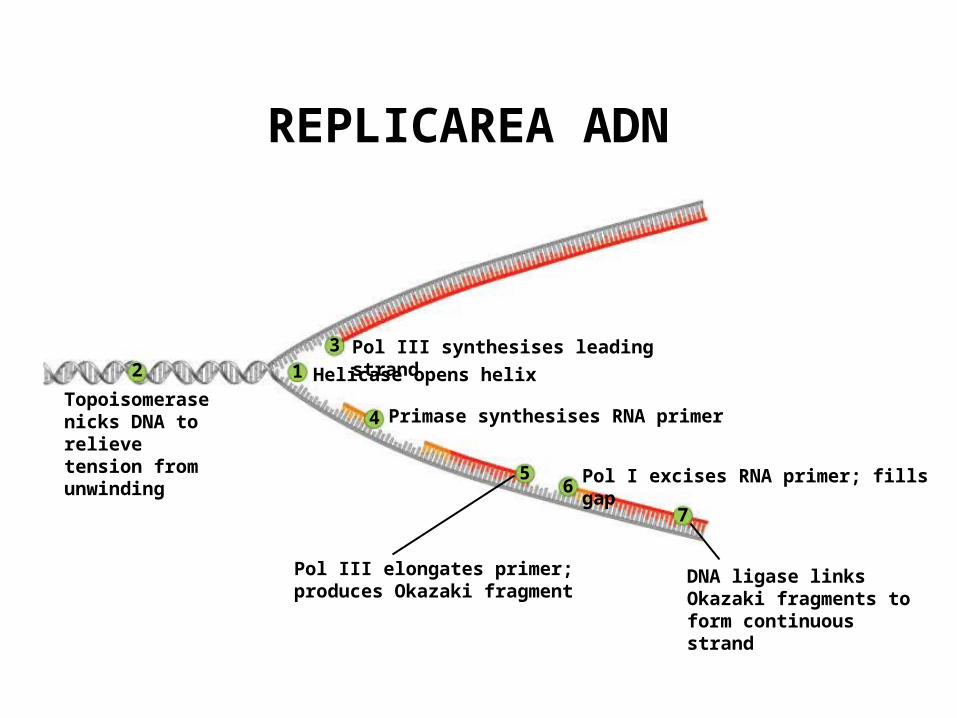
Topoisomerase nicks DNA to relieve tension from unwinding
23
1
4
56
7
Pol III synthesises leading strand
Helicase opens helix
Primase synthesises RNA primer
Pol III elongates primer; produces Okazaki fragment
Pol I excises RNA primer; fills gap
DNA ligase links Okazaki fragments to form continuous strand
REPLICAREA ADN
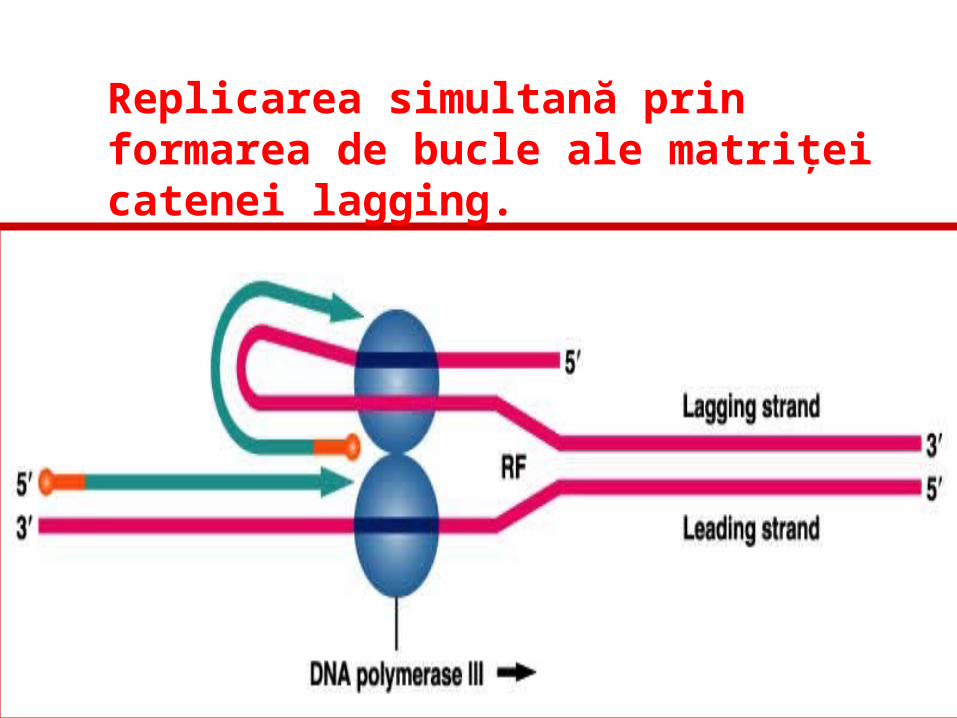
Replicarea simultană prin formarea de bucle ale matriţei catenei lagging.
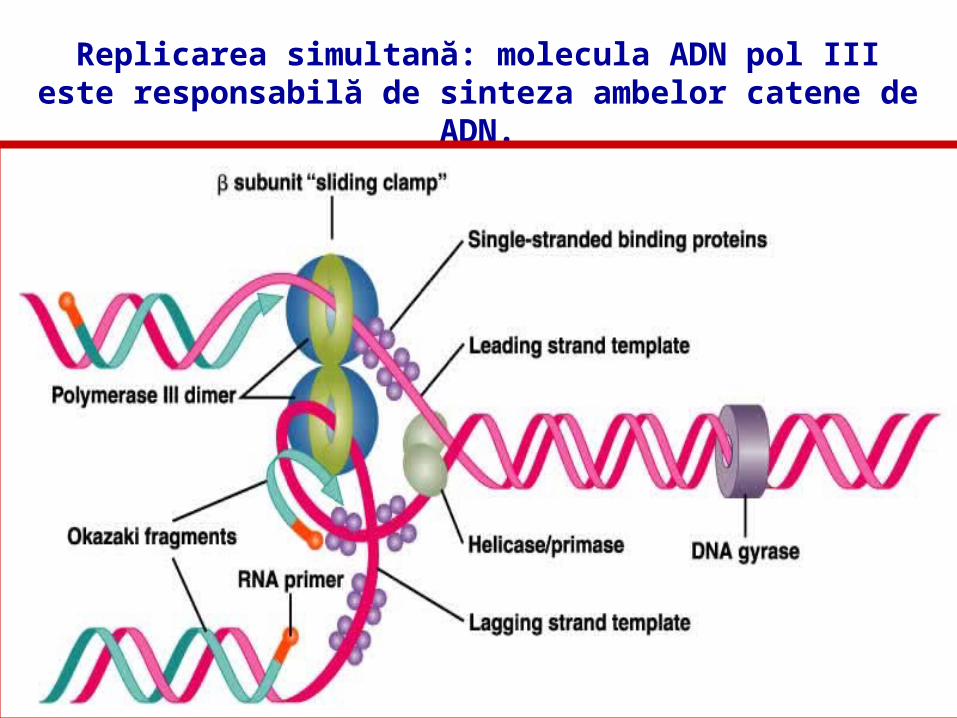
Replicarea simultană: molecula ADN pol III este responsabilă de sinteza ambelor catene de ADN.

Caracteristici ale replicării la eucariote Replicarea este bidirectională Are mai multe origini de replicare
deoarece molecula de ADN este foarte lungă şi viteza de acţiune a replizomului este mică
Histonele vechi se asociază cu duplexul nou care conţine catena leading.
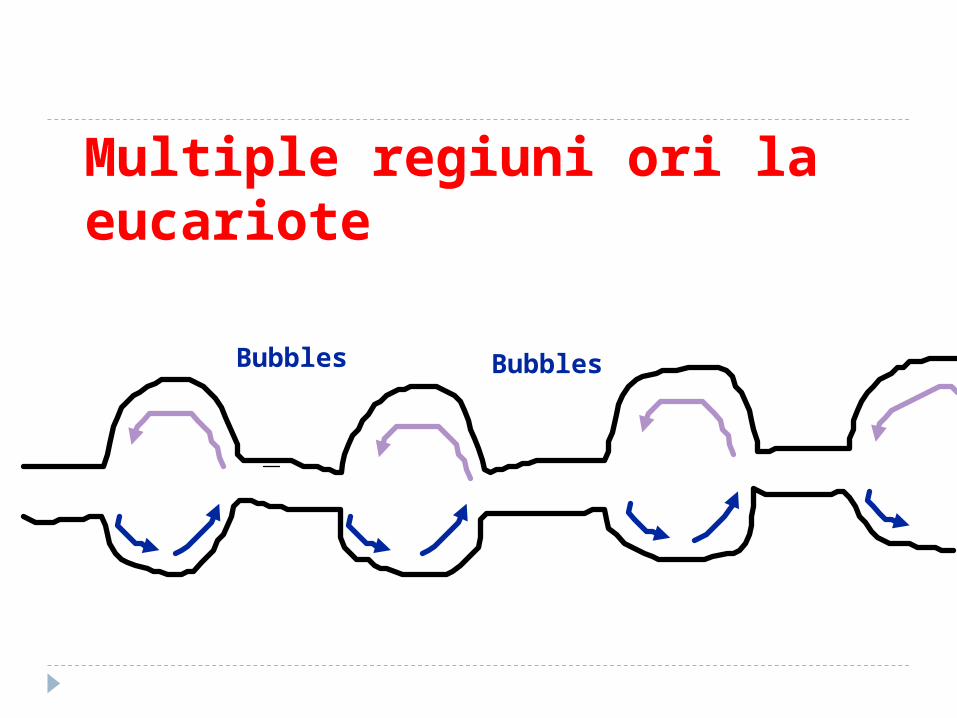
Multiple regiuni ori la eucariote
Bubbles Bubbles

Replicarea la eucariote Particularităţi:
ADN polimerazele: αβγδ α- implicată în replicarea ADN nuclear-
responsabilă de sinteza catenei întârziate – sinteza primerilor
δ – răspunde de sinteza catenei lider. Ea manifestă acţiune exonucleazică
β – implicată în reparaţia ADN γ - implicat în replicarea ADN mitocondrial,
acţiune exonucleazică
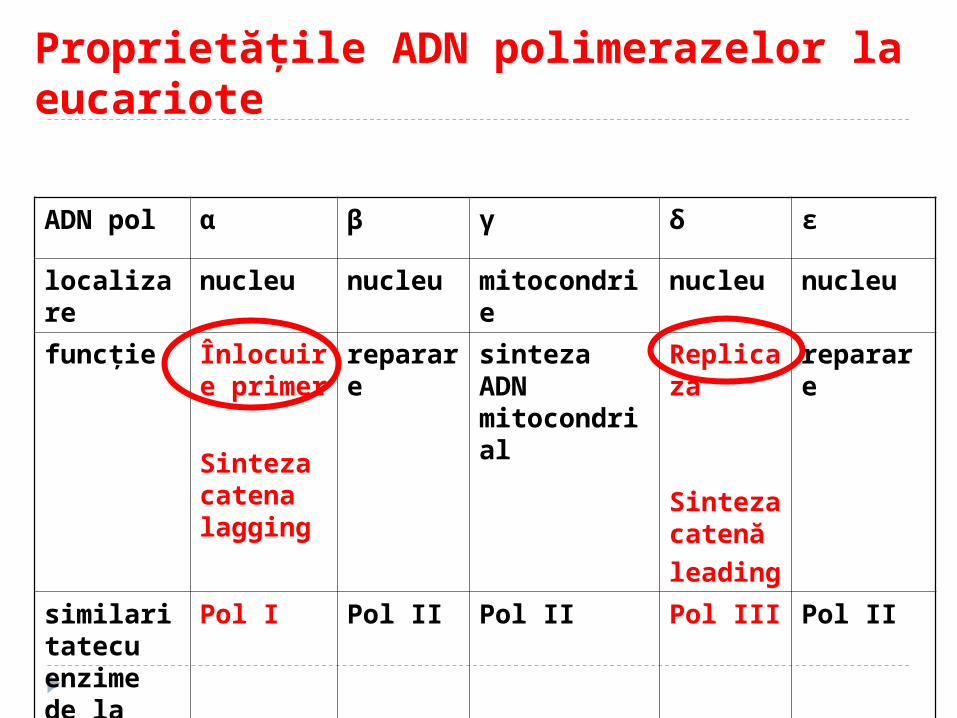
Proprietăţile ADN polimerazelor la eucariote
ADN pol α β γ δ ε
localizare nucleu nucleu mitocondrie nucleu nucleu
funcţie Înlocuire primer
Sinteza catena lagging
reparare sinteza ADN mitocondrial
Replicaza
Sinteza catenă
leading
reparare
similaritatecu enzime de la procariote
Pol I Pol II Pol II Pol III Pol II
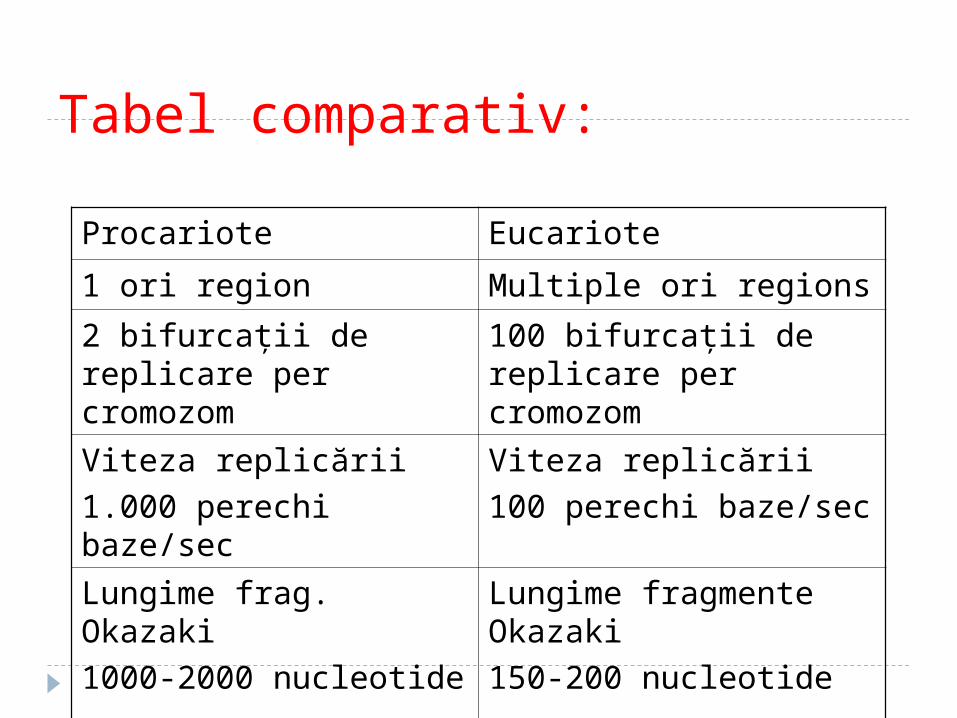
Tabel comparativ:
Procariote Eucariote
1 ori region Multiple ori regions
2 bifurcaţii de replicare per cromozom
100 bifurcaţii de replicare per cromozom
Viteza replicării
1.000 perechi baze/sec
Viteza replicării
100 perechi baze/sec
Lungime frag. Okazaki
1000-2000 nucleotide
Lungime fragmente Okazaki
150-200 nucleotide
1 eoare la 1mil-10mil 1 eroare la 1000 mil

Replicarea la eucariote Bifurcaţia replicii este de 3000 baze pe minut
comparativ cu 16.000 la procariote Pe o moleculă de ADN există mai multe origini
de replicare (3X104 - 3X105 separate prin perechi de baze). În aceste origini multiple de replicare se organizează bifurcaţii- ce se deplasează biderecţional pe cromosomul eucariot în curs de replicare.
Fragmentele Okazaki 150-200 nucleotide

Telomer Telomeraza Replicarea capetelor 5’ ale
catenelor este incompletă (teoria lui Olovnicov, 1971), deoarece după înlăturarea primerului ultimului fragment Okazaki, ADN p I nu e capabilă să completeze aceste goluri.
Astfel la fiecare replicare, capetele ADN se scurtează.

Telomerul Aceasta nu afectează informaţia genetică
deoarece catenele conţin fragmente repetitive neinformative – telomere.
Telomer – complex format din ADN şi proteine - sunr segmente de ADN repetitiv AGGGTT

Telomeraza Telomerele sunt replicate de o E specifică
– telomeraza Telomeraza - reprezintă o
ribonucleoproteidă: ARN şi proteină Subunitatea proteică TRT (telomeraze
revers transcriptase) posedă activitate catalitică

Telomeraza – fiind o revertază (ADN polimeraza ARN dependentă) foloseşte ca matriţă propria coenzimă – un fragment de ARN.
I etapă – are loc asocierea telomerazei la capătul 3’ al catenei lider din regiunea telomerică- TTAGGG
II – E extinde catena, utilizând ca matriţă ARN telomeric (se repetă)
III – Catena complementară a ADN telomeric e sintetizată după principiul catenei întârziate de ADNp

Mecanismul elongării capetelor cromozomului la eucariote


Mecanismul elongării capetelor cromozomului la eucariote
cromozoma GGGTTAG 3’ AUCCCAAUC 5’ Fixarea telomerei TTAGGG elongarea GGGTTAGGGTTAG 5’ AUCCCAAUC
translocarea GGGTTAGGGTTAG 5’
AUCCCAAUC

Structura şi funcţia RNA telomerazice. Structura primară: la majoritatea RNA
telomerice, regiunea matricială se află la depărtarea de 50 nucleotide de la capătul 5’, şi are următoarea succesiune de nucleotide 5’-CUAACCCUA-3’.
Structura secundară e compusă din 4 bucle şi un fragment unicatenar, ce conţine matriţa pentru sinteza DNA telomerice.

Inhibitorii telomerazei oligonucleotidele modificate, complementare
regiunii matrice a RNA – telomerazice. specific se fixează de matriţa RNA –telo a
omului, inhibînd activitatea telomerazică in vitro. In vivo apare problema transportului inhibitorilor prin membrana celulară şi mişcarea dirijată în nucleul celular.
Ca inhibitori au fost testaţi şi inhibitorii reverstranscriptazelor – azidotimidina, didezoxiguanozina.

La om telomeraza e activă numai în celulele embrionale, în epiteliul intestinului, spermatozoizi şi celule canceroase.

Numărul telomerilor determină durata vieţii fiecărei celule şi condiţionează reducerea critică a numărului lor, induce moartea programată a celulei deci pierderea motivelor telomerice este cauza imbătrînirii (telomera conţine mii de motive TTAGGG).
lungimea telomerei este marcherul biologic al îmbătrînirii.

Reparaţia ADN
Erorile în timpul replicării sunt reduse la minimum datorită DNA polimerazei ce posedă funcţie endonucleazică
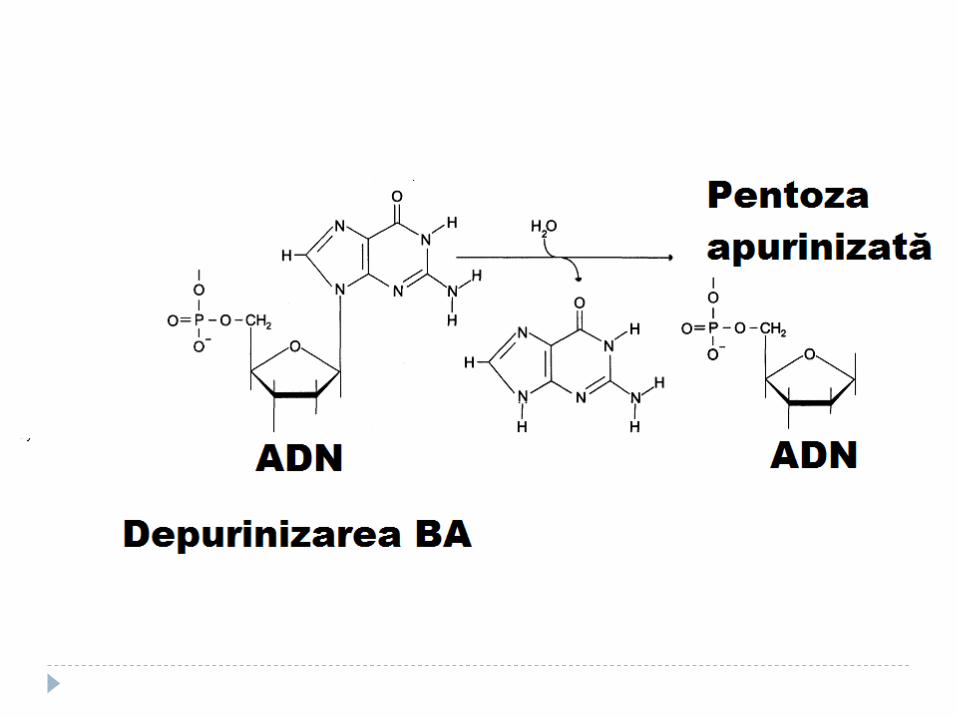
Tipuri de deteriorări:
1. Formarea de breşe
2. Modificarea BA
3. Pierderea de BA
4. Formarea dimerilor de pirimidină sub acţiunea razelor ultraviolete
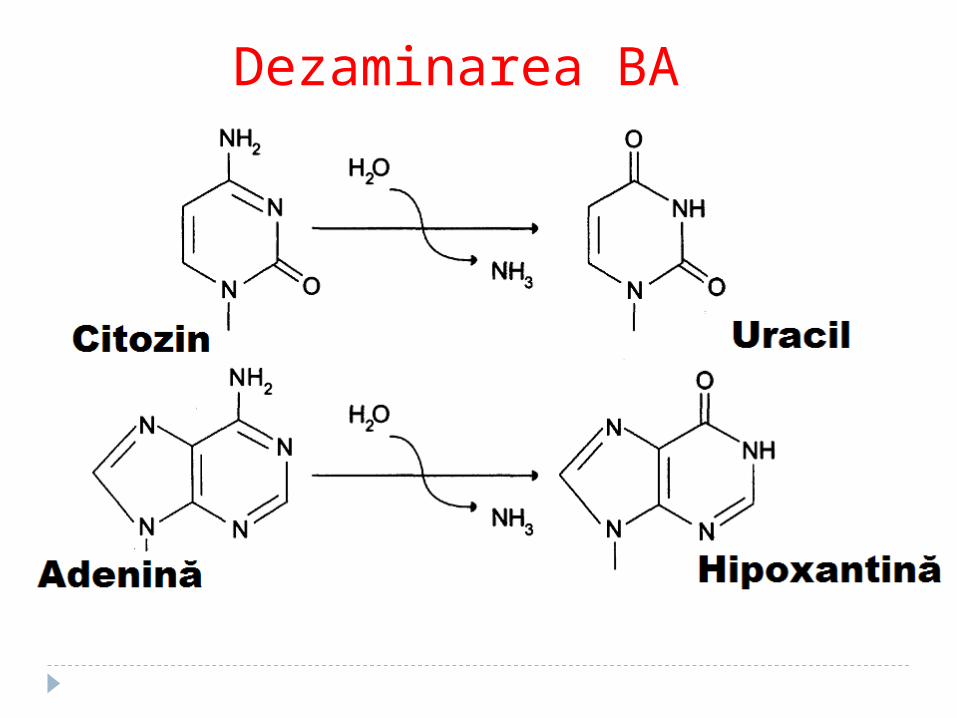
Dezaminarea BA
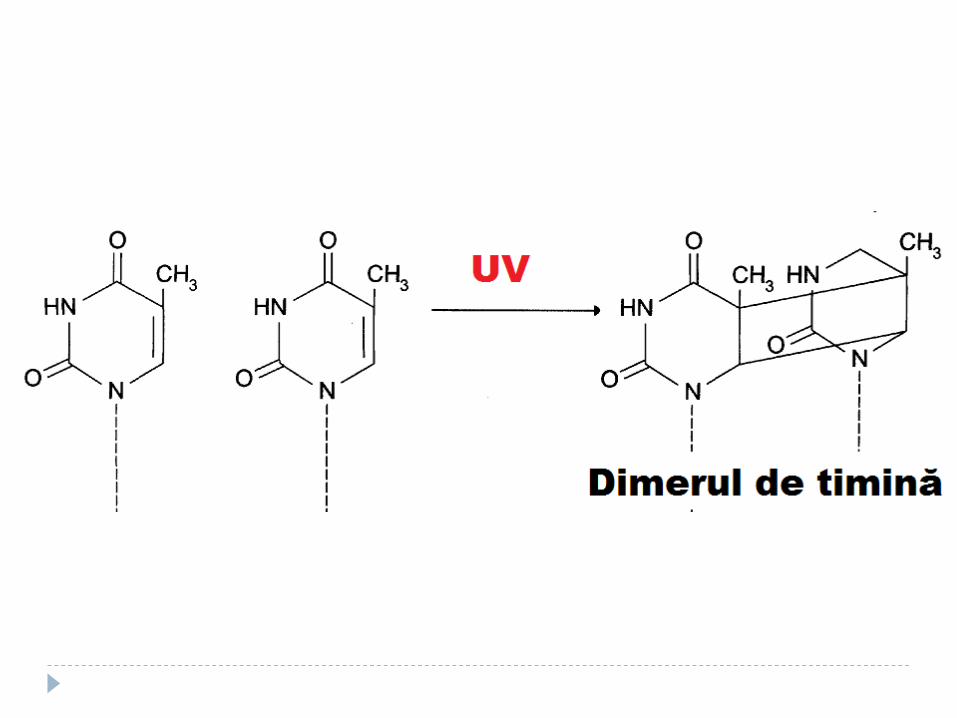

Mecanisme de reparare a ADN Repararea defectelor de împerechere a bazelor
(prin excizia nucleotidelor) – proteinele MUT Repararea defectelor cauzată de lumina UV
(dimerii de timină) Corectarea defectelor BA (excizia bazelor) Repararea rupturilor bicatenare

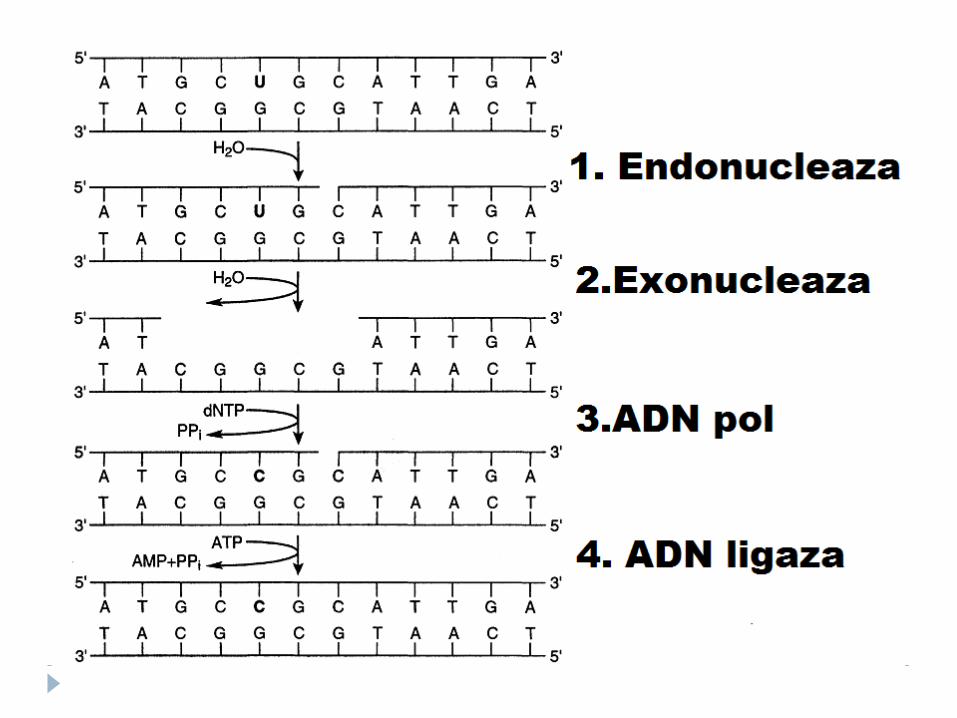
Repararea defectelor de împerechere a bazelor (prin excizia nucleotidelor) – proteinele MUT

Repararea prin excizia dimerilor de pirimidină
Recunoaşterea şi excizarea dimerilor de către endonucleaze UV specifice
SPAŢIU GOL E OCUPAT DE ADN polimerazei I sub acţiunea ADN ligazei, gruparea 3-OH a catenei
nou sintetizate este legată de 5-fosfat a ADN iniţial.
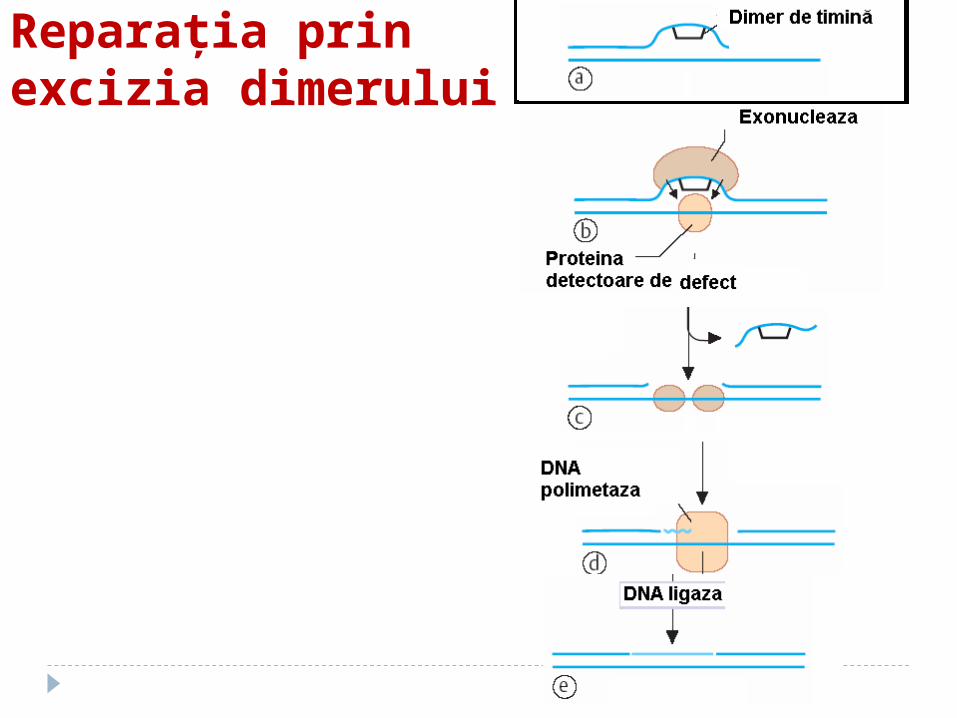
Reparaţia prin excizia dimerului
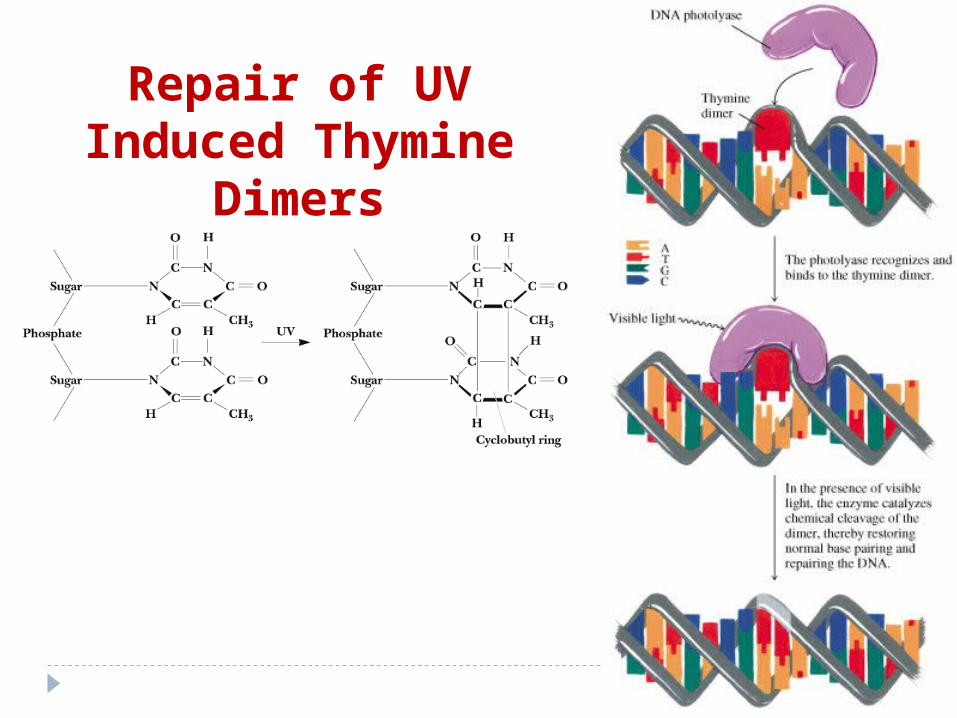
Repair of UV Induced Thymine
Dimers
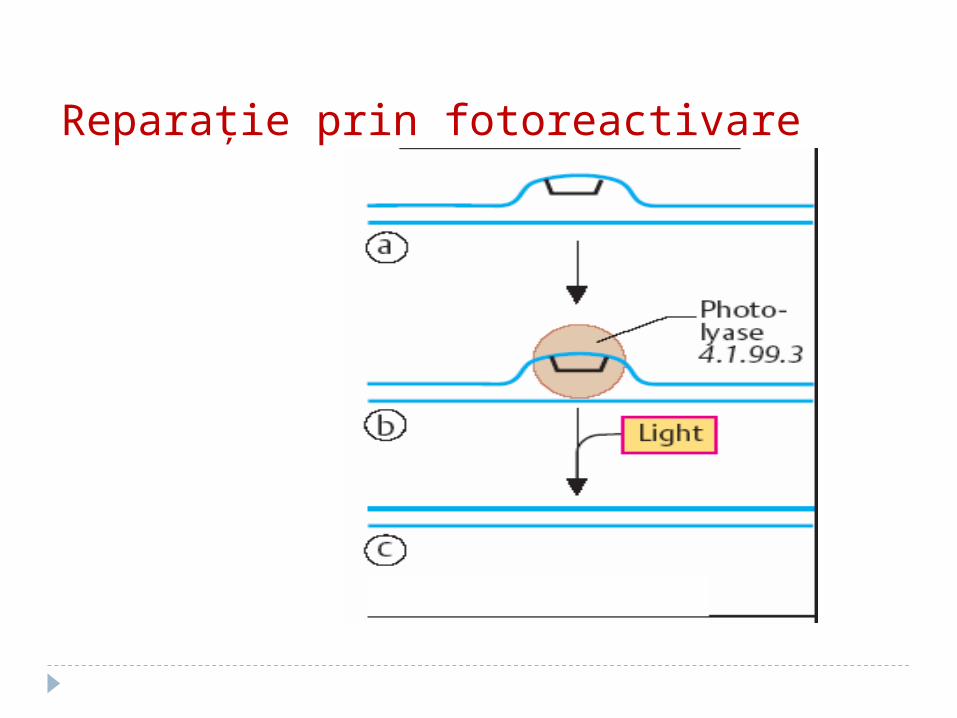
Reparaţie prin fotoreactivare
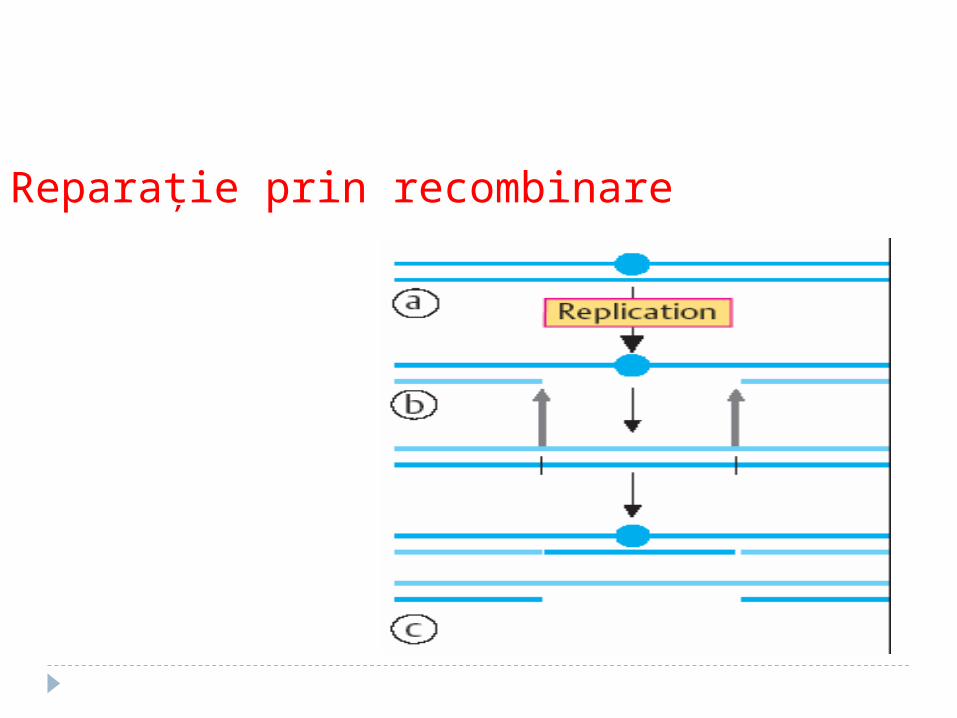
Reparaţie prin recombinare

Mutaţiile
Modificările genomului organismului, care se păstrează şi se transmit prin ereditate
se transmit apoi de la o generaţie la alta.
Modificările pot interesa o pereche de baze (mutatii punctiforme) sau un grup de baze pe una sau pe ambele catene ale unei molecule de DNA.

Mutatiile punctiforme: pot decurge prin:l. Substituţie - misens mutatii, unde deosebim 2 tipuri: a. Tranziţie - o BA purinică este
înlocuită tot cu una purincă, una pirimidinică -tot cu una pirimidinică.
b.Transversie - o pereche de baze purinice este înlocuită cu una pirimidinică sau invers.
2. Inserţie - constă în întroducerea unei perechi de baze suplimentare în catena de DNA.
3. Deleţia - constă în excluderea unei perechi de baze în aşa mod ca ea nu mai poate fî complementară şi la replicare apare "golul" în ambele catene.
Mutaţiile

Mutaţiile
La afectarea segmentelor mari de genă apar mutaţii întinse.
În dependenţă de consecinţele modificărilor deosebim mutaţie:
benignă, neutră, nocivă. Agenţii mutageni pot provoca mutaţiile
spontane cît şi mutaţiile induse.

BIOSINTEZA ARN

Obiectivele: ARN. Tipurile. Structura Dogma centrală a geneticii moleculare. Concepţia: o
genă - un polipeptid. Transcripţia sau biosinteza ARN: matricea,
substratele, enzimele, mecanismul Trsanscripţia inversă. Biosinteza ARN pe matrice de ARN Modificările posttranscripţionale (processing) Inhibitorii sintezei acizilor nucleici.
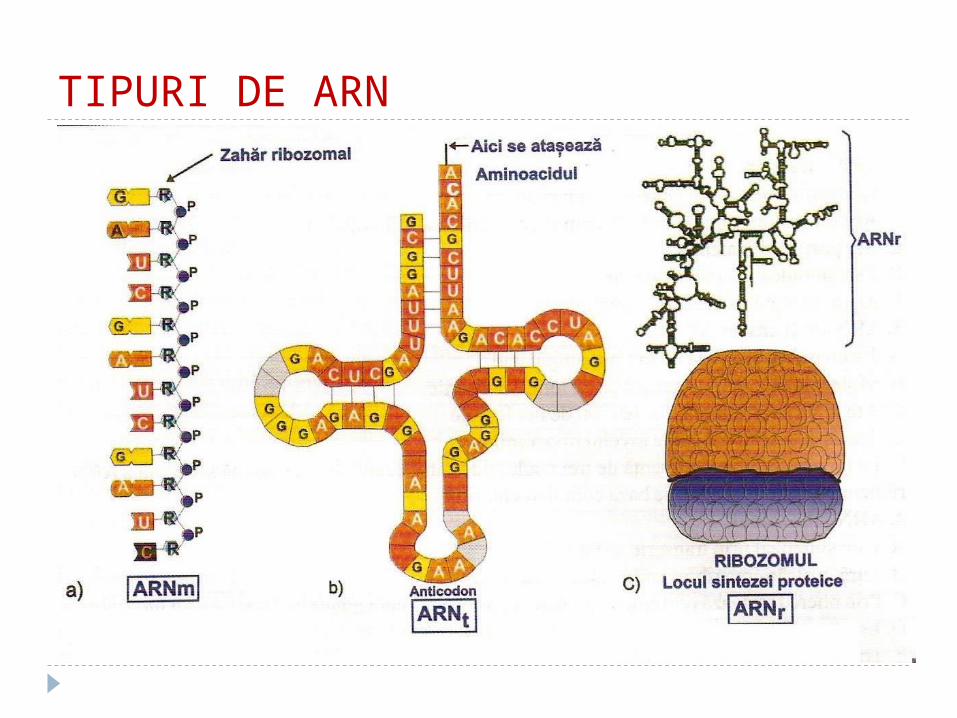
TIPURI DE ARN

Structura secundară şi terţiară a ARNm ARNm – fiecărei gene îi corespunde molecula sa
de ARNm, de aceea el este foarte heterogen Elementul de codificare al ARNm este tripletul
nucleotidic – numit codon. Fiecare codon corespunde unui anumit AA
Structura secundară a ARNm – o catenă curbată Structura terţiară – se aseamănă cu un fir
înfăşurat pe bobină, rolul căreia îl îndeplineşte o proteină de transport numită informer

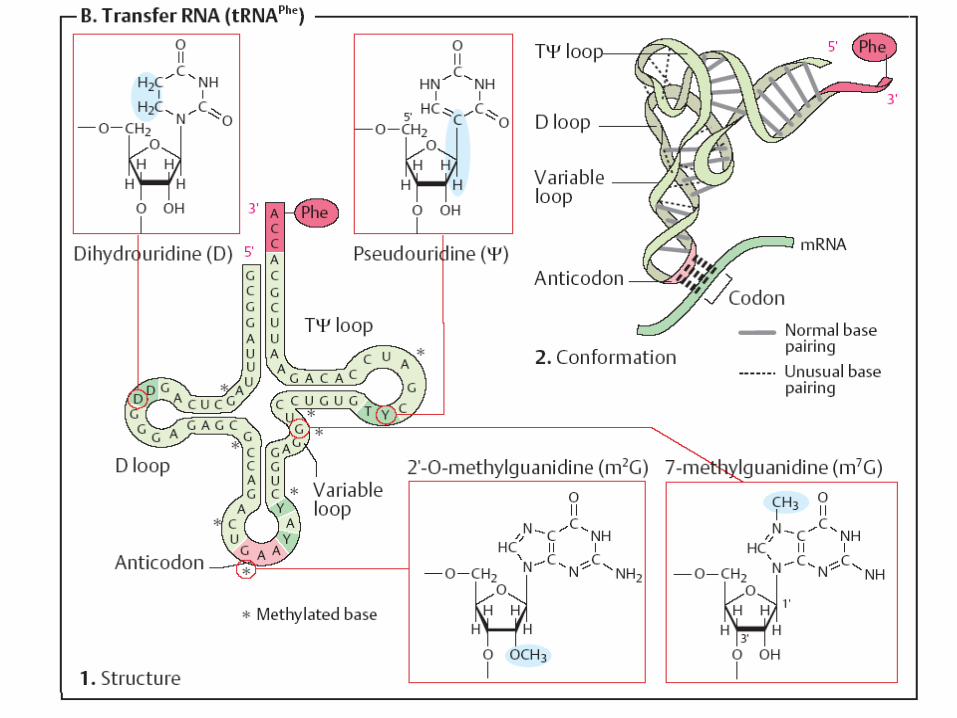
Structura secundară a t-RNA are infăţişarea unei "frunze de trifoi" - se
formează în urma imperecherii complementare intracatenare a nucleotidelor anumitor sectoare.
Sectoarele, care nu sunt încadrate în formarea legăturilor de H formează lanţuri sau bucle
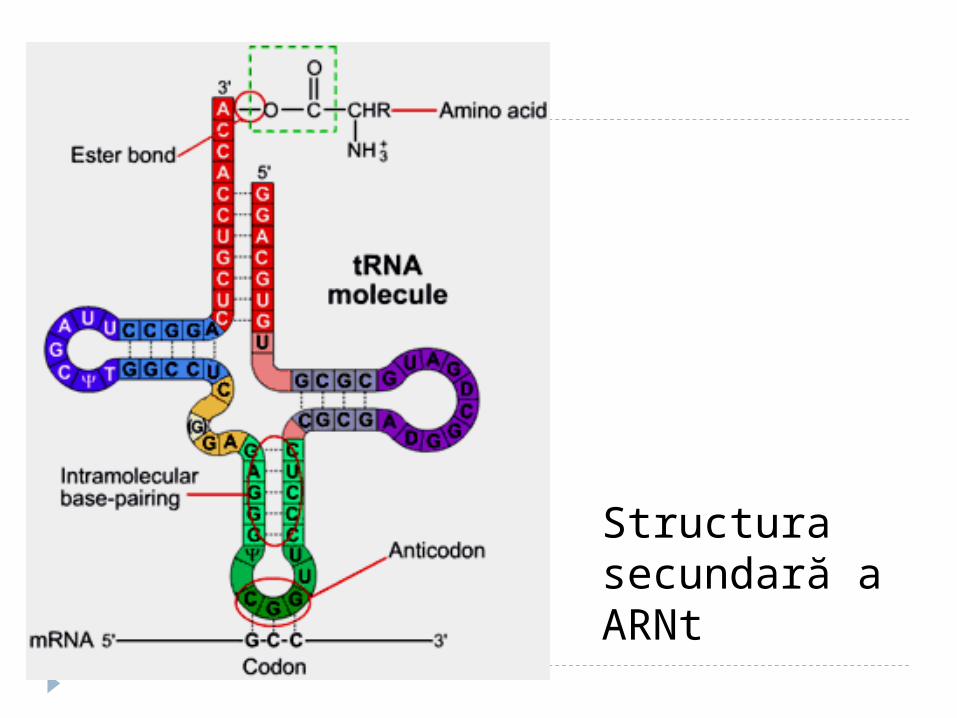
Structura secundară a ARNt

1. Sectorul de acceptare (4 nucleotide, 3 din care au aceeaşi succesiune- CCA cu hidroxilul 3’OH liber la care se fixează grupa COOH al AA.
2. Bucla anticodonică - formată din 7 nucleotide. Conţine un triplet nucleotidic specific pentru fiecare t-RNA numit anticodon. Anticodonul t-RNA după principiul complementaritătii se împerechează cu codonul respectiv din RNAm.
3. Bucla pseudouridilică - constă din 7 nucleotide (restul acidului pseudouridilic este obligatoriu), participă la interacţiunea cu ribozomii.
4. Bucla dihidrouridinică - constă din 8-12 resturi nucleotidice ( resturi de dihidrouridină ), interactiunează cu E- aminoacil-RNAt-sintetaza

Structura terţiară a tRNA Are forma L Include 2 segmente de dublu helix situate
perpendicular (fiecare helix-10 perechi de baze) În afara spiralei bazele formează legături de
hidrogen. Interacţiuni apar între bazele necomplementare (A-A; A-C).Multe baze sunt aranjate în stive (hidrofobe)
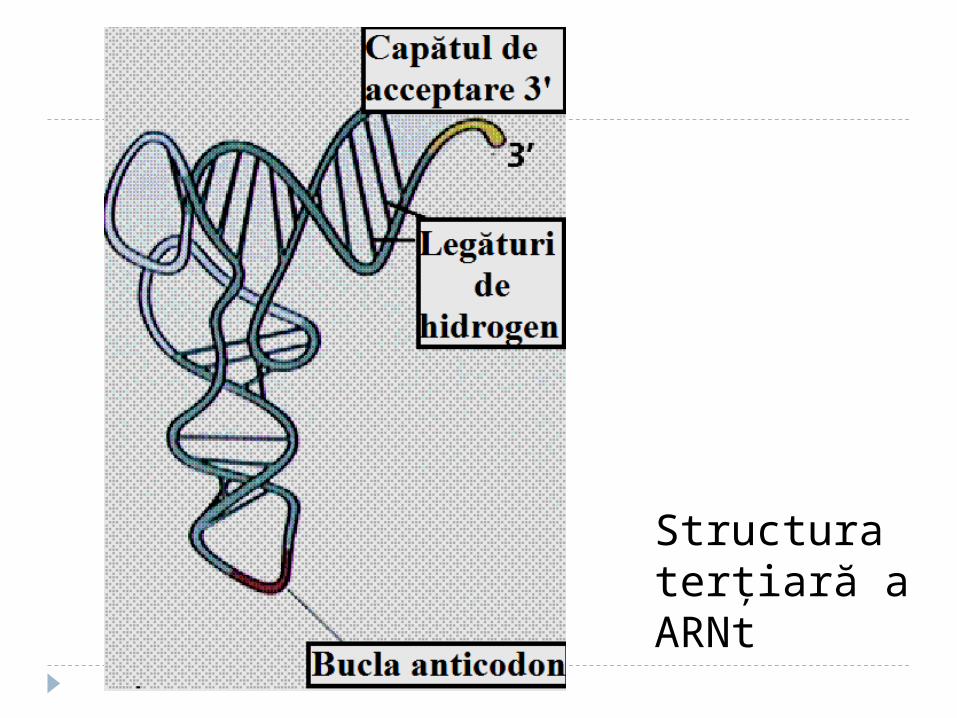
Structura terţiară a ARNt

ARNr Structura secundară – e prezentată prin
sectoare spiralate unite între ele cu ajutorul unei catene curbate
Structura terţiară – prezintă scheletul ribosomului. Are forma unui bastonaş sau ghem pe suprafaţa căruia sunt înfăşurate proteinele ribosomului.

Transcripţia biosinteza ARN pe matriţă de ADN Particularităţi: Matriţă - DNA dublu helicoidal (prezenţa catenei
anticodogene de ADN) (catena+), Substrat - ribonucleozidtrifosfaţi (ATP, GTP, CTP,
UTP) Sinteza are loc în direcţia 5’3’ Este asimetrică – copierea catenei
necodificătoare Este incompletă –are loc copierea doar a unei
porţiuni de ADN (transcripton: promotor, operator, GS, terminator)
Forţa motrice a procesului e hidroliza PP Enzima - ARN polimeraza

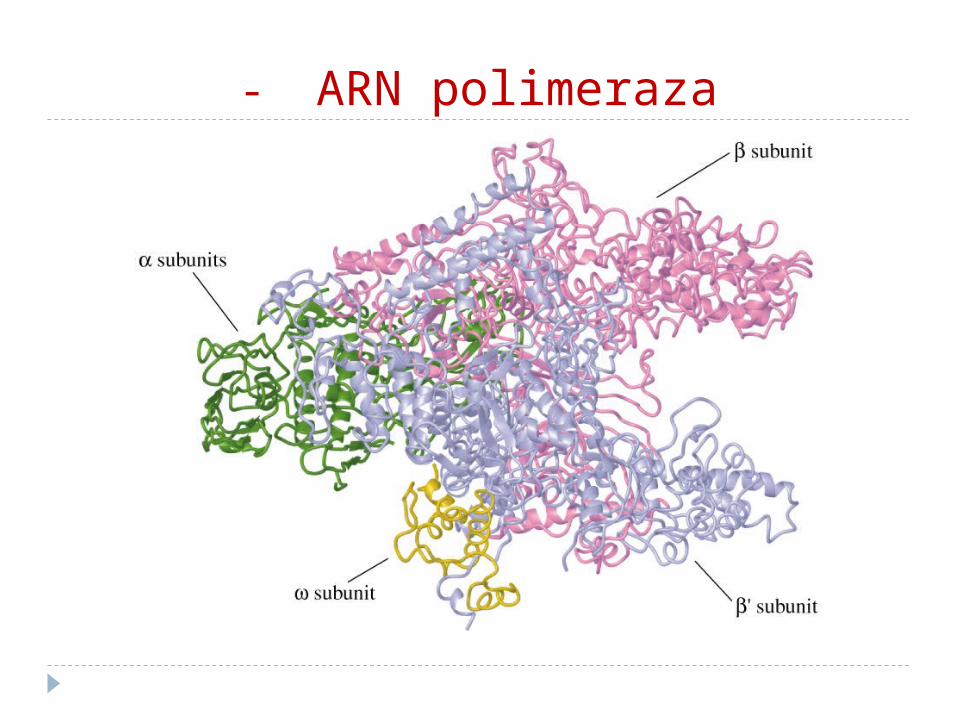
ARN polimeraza
1. este o holoenzimă2. la procariote - este oligomer din 5 protomeri
(2, , 1 şi sigma σ). subunităţile – centre catalitice; - fixează substratul; 1 – se leagă de ADN, σ - are rol în recunoaşterea secvenţelor
matriţei numit promotor, unde aderă enzima3. Nu necesită prezenţa primerului4. Nu posedă funcţie nucleazică, doar
polimerazică5. ADN polimeraza conţine Zn2+ şi necesită
prezenţa în mediu a ionilir de Mg2+, Mn2+
- ARN polimeraza

- ARN polimerazala eucariote: 1. RNA p I sintetizeaza RNA ribozomal
(28S si 18S)2. RNA p II sintetizeaza RNAm•3. RNA p III sintetizeaza RNAt, RNAr
5S şi molecule mai mici

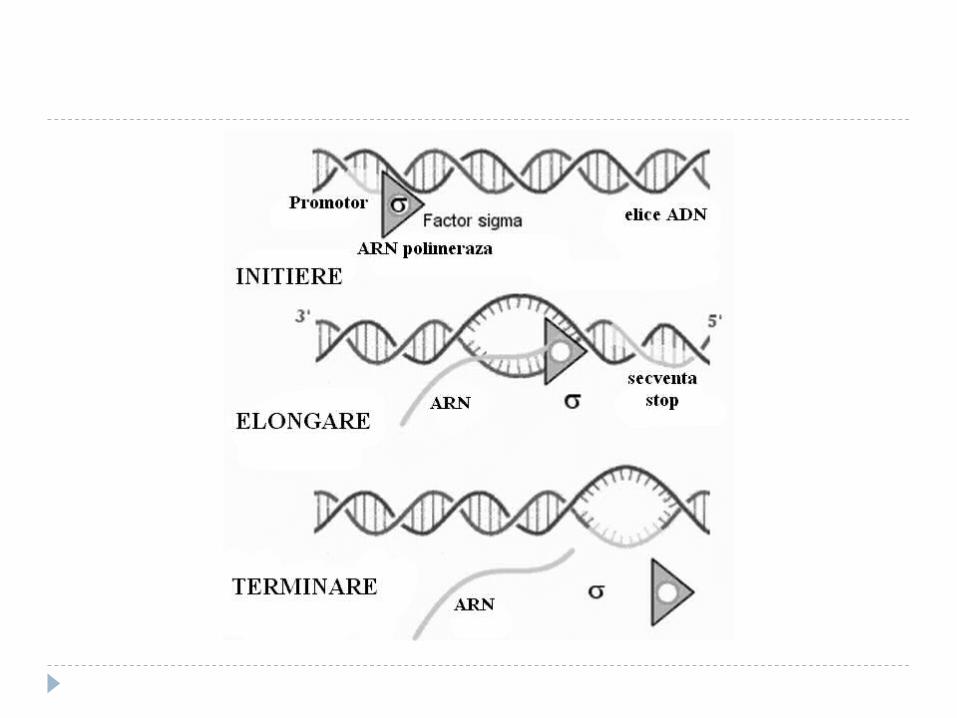
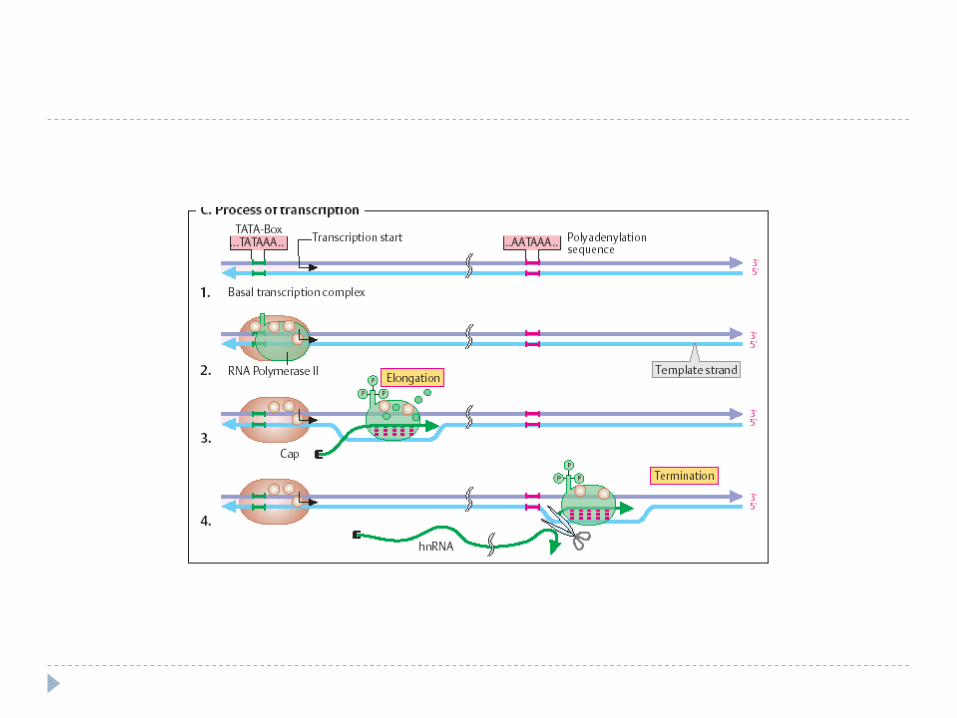
Etapele transcripţiei
Sinteza decurge în 3 etape: iniţierea, elongarea, terminarea. Iniţierea – începe în anumite secvenţe de
ADN numite promotor (P – 40 nucleotide) recunoscută de ARN polimeraza.

CASETELE la procarioteToţi P bacterieni au 2 secvenţe consens situate pe
catena codificătoare: – Caseta -35 - 5’ TTGACA 3’ - locusul de
legare laxă a ARN polimerazei. - Caseta Pribnow (-10) 5’ TATAAT 3’ –locusul
de recunoaştere - responsabilă de iniţierea denaturării locale a ADN.
Locusul de recunoaştere e situat la o distanţă de 25 nucleotide de locusul de legare şi 10 nucleotide de la punctul de iniţiere (+1) – răspunde de exactitatea iniţierii transcripţiei.
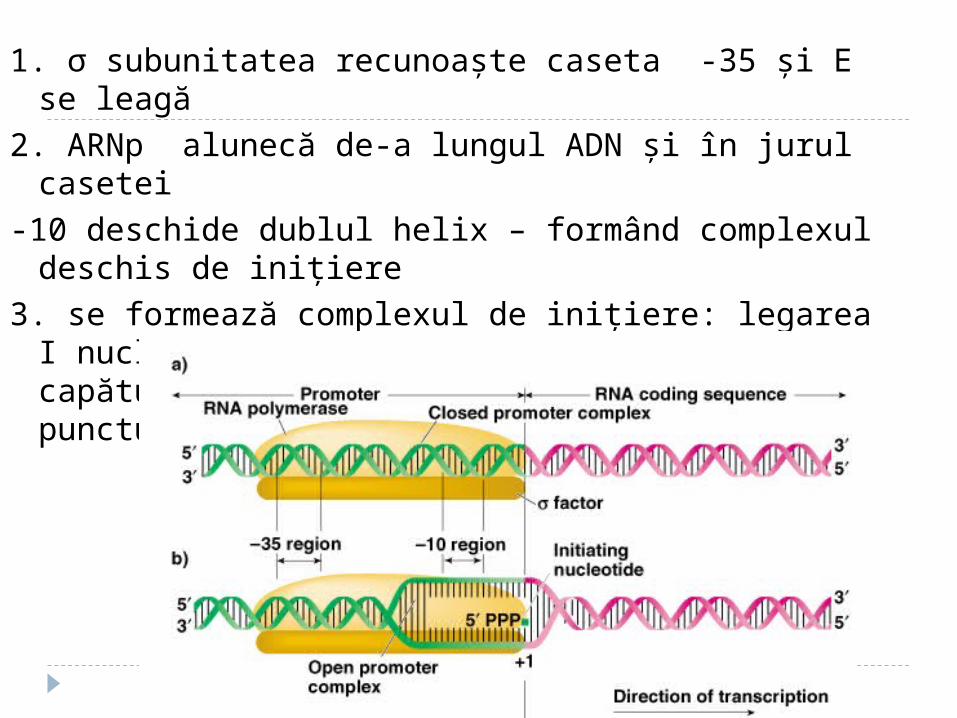
1. σ subunitatea recunoaşte caseta -35 şi E se leagă
2. ARNp alunecă de-a lungul ADN şi în jurul casetei
-10 deschide dublul helix – formând complexul deschis de iniţiere
3. se formează complexul de iniţiere: legarea I nucleotid trifosfat (de regulă purinic - capătul 5’ al ARN). CA al E se află în punctul de start.

4. ARNp catalizează formarea primei leg. fosfodiesterice între nucleotidul +1 şi +2
Sigma subunitatea :1. Activează identificarea secvenţelor de ARN
polimerază2. Ia parte la desfacerea dublului helix 3. Ia parte la formarea primei legături
fosfosiestericeAstfel complexul de iniţiere este format, sigma
subunitatea e disociată de la holoenzimă şi ia parte la iniţierea unui alt ciclu de transcriere
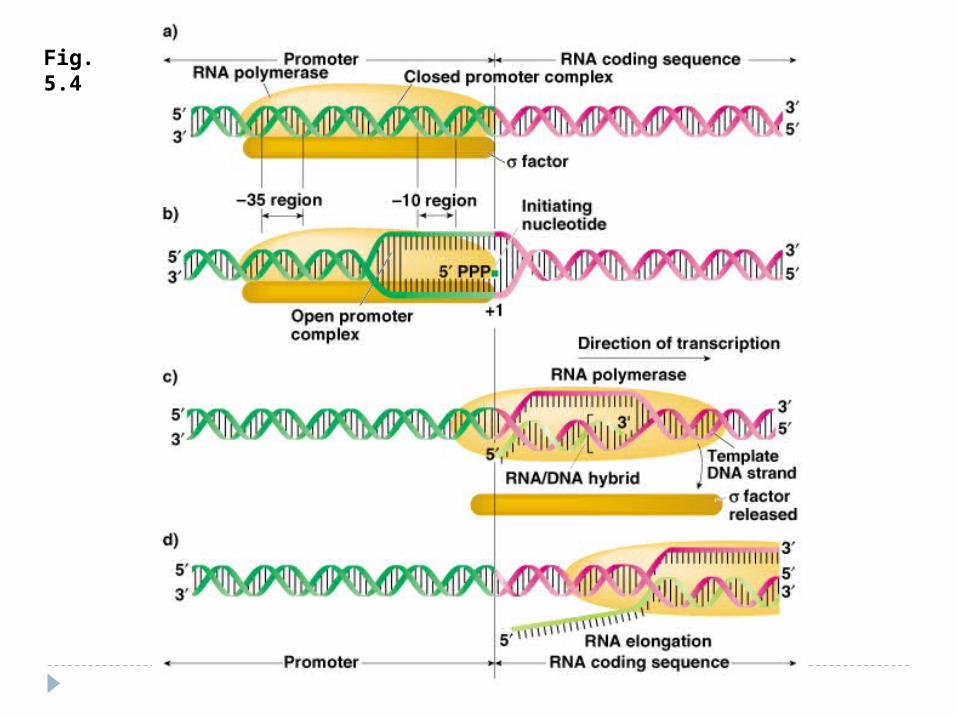
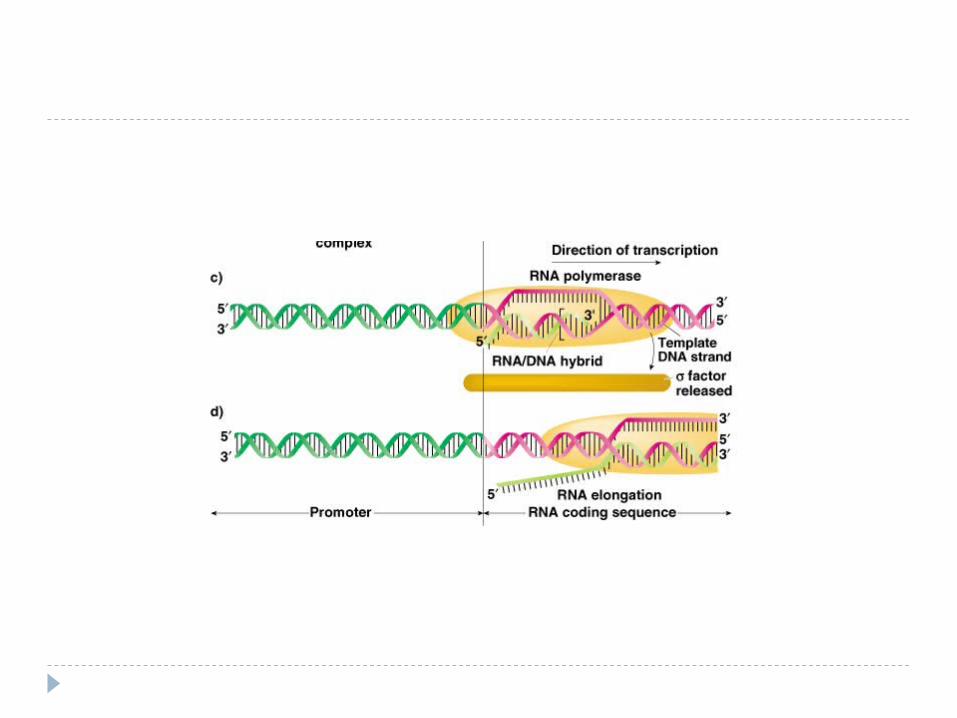
Fig. 5.4

Elongarea şi Terminarea Elongarea - alunecarea ARN polimerazei pe
matrţa de ADN – sinteza transcriptului (50 nucleotide pe secundă) . ARNp nu controlează catena sintetizată – erorile sunt mai multe faţă de replicare. Pe măsură înaintării ARNp are loc desprinderea ARN de la ADN şi refacerea dublului helix
Terminarea – semnal de terminare ARNp recunoaşte secvenţele nucleotidice
specifice de pe ADN, ce conţin un număr mare de G, C .

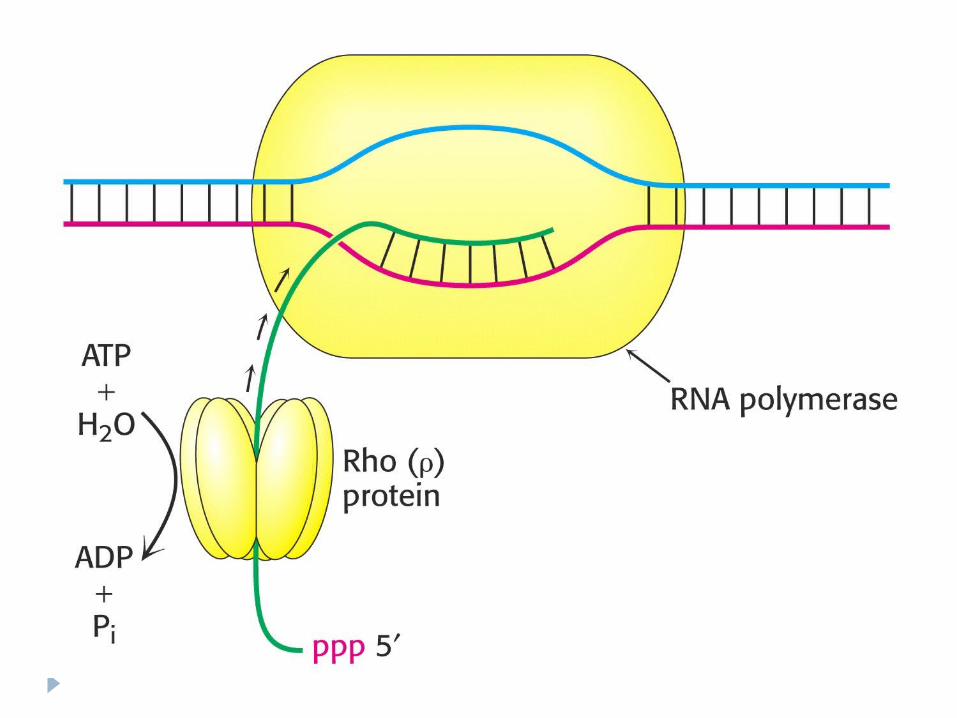
Terminare dependentă de factorul ρ
Proteina ρ - se asociază la E şi se mişcă împreună cu ea, însă la identificarea semnalelor de terminare coboară de pe matriţă şi încetineşte acţiunea E,producând transcriptul cu folosirea energiei – ATP. ADN-se reformează în dublul helix.

Transcripţia la eucariote
1. RNA p alcătuită din 9-11 subunităţi2. Folosesc mai multe tipuri de ARN p:a. ARN polimeraza I – ARNr (18S, 28S,
5,8S; 45S) b. ARN polimeraza II – ARNm; c. ARN polimeraza III –ARNt şi ARNr (5S)d. ARN polimeraza IV (mitocondrială)- toate
tipurile de ARN mitocondrial

Transcripţia la eucariote3. P la eucariote: GC casete GGGCG (-90) CAAT casete – 5-GGCCAATCT -3 (-75) Caseta Hogness –TATAT/AT (-35)Primele 2 sunt responsabile de frecvenţa
transcripţiei (când începe), a 3 - - implicată în iniţiere – semnal (unde).
4. Secvenţele alcătuite din 10-20 nucleotide “enhancers” şi “silencers” – cresc şi scad respectiv V transcrierii.

Procesingul la eucariote ARN-ul mesager obţinut diferă la procariote faţă
de eucariote. în ARN-ul procariot molecula este continuă deci
intreaga informaţie de aici poate fi folosită pentru sinteza proteinelor.
în ARN-ul eucariot s-a observat prezenţa unor porţiuni repetitive de nucleotide numite introni care nu deţin nici o informaţie pentru sinteza proteică.
ARN-ul mesager primar este mult mai lung decât ARN-ul mesager folosit de ribozom.
Toţi precursorii de ARN în nucleu trec etapa de maturizare posttranscripţională. Pe parcursul procesingului - pre-ARN se transformă în ARN matur.

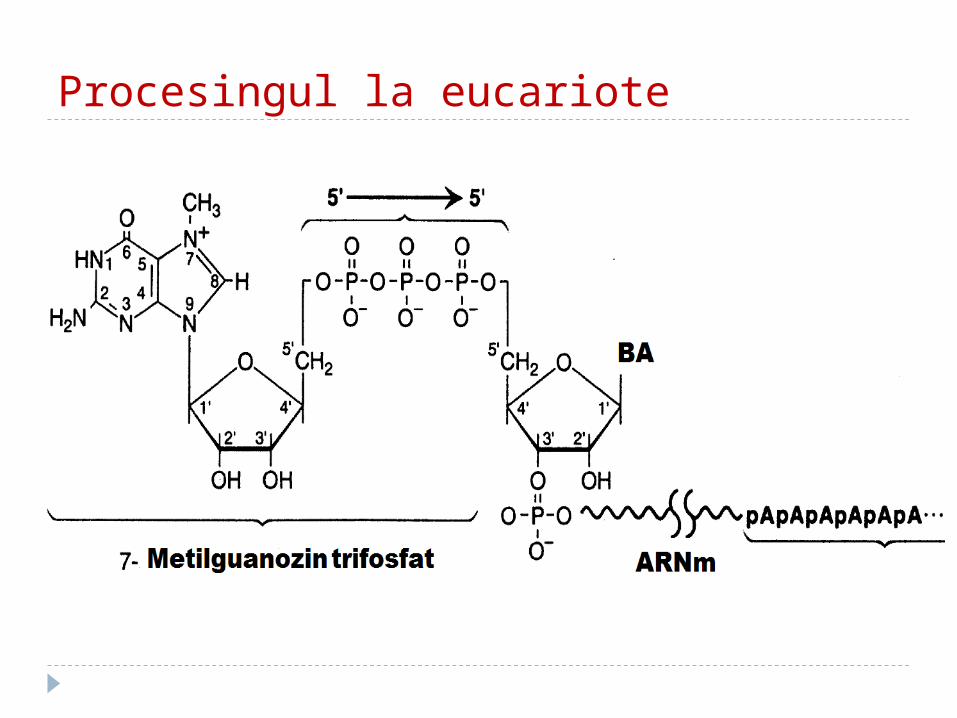
Procesingul la eucariote Procesingul înclude:1. Modificarea fragmentelor terminale 5’ şi 3’
ale ARN: a. “Cap”-area: la capătul 5’ -este adiţionată
guanozina metilată (5’- 5’ trifosfat - protejarea mARN de atacul 5’-exonucleazelor şi pentru recunoaşterea de către ribosomi ca semnal de iniţiere);
b. la capătul 3’ – se adaugă o secvenţă mare de poli A (200 A -coadă). Ea serveşte la exportul moleculelor de ARN din nucleu în citoplasmă.
Procesingul la eucariote
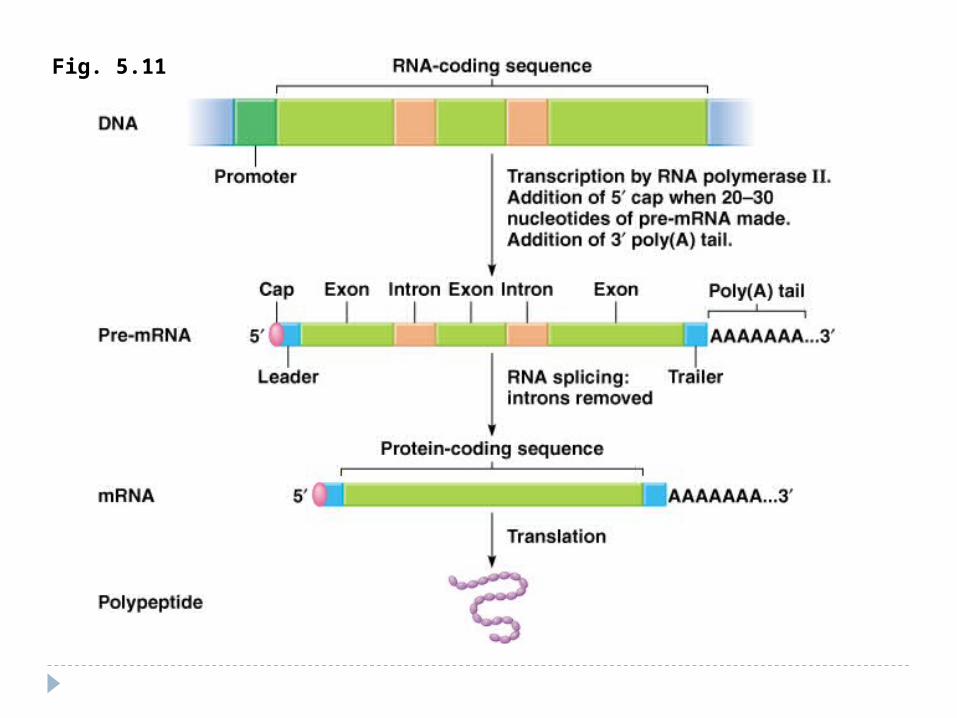
Fig. 5.11

Procesingul la eucariote2.Splisingul - excizia intronilor şi sudarea exonilor.
Aşa numitul splising are loc în nucleul celulei. Excizarea intronilor se poate face in 2 feluri:
Cu ajutorul unor enzime speciale care recunosc capetele fiecarui intron (exemplu: ARN U1) acestia sunt eliminati si capetele libere ale exonilor sunt sudate.Prin autocataliza, fenomen remarcabil care pune pentru prima data in discutie posibilitatea catalitica a unui compus neproteic. Deci chiar ARN-ul ribozomal isi autoelimina intronii printr-un proces care genereaza o bucla (reprezentata de intron), eliminarea acestei bucle cu ajutorul guanozinei si coaserea exonilor.
a.
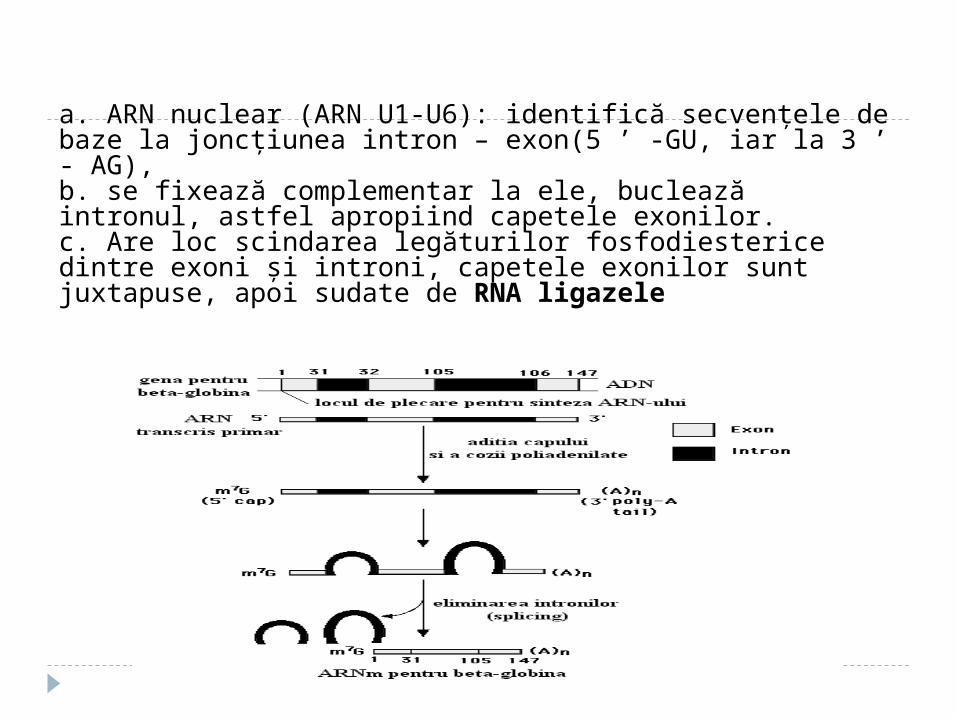
a. ARN nuclear (ARN U1-U6): identifică secvenţele de baze la joncţiunea intron – exon(5 ’ -GU, iar la 3 ’ - AG), b. se fixează complementar la ele, buclează intronul, astfel apropiind capetele exonilor.c. Are loc scindarea legăturilor fosfodiesterice dintre exoni şi introni, capetele exonilor sunt juxtapuse, apoi sudate de RNA ligazele

Mecanismul ARN U1 se leagă la GU a intronului ARN U2 se uneşte la A din interiorul intronului ARN U4,U5, U6 se asociază cu U1 şi U2 formînd
bucla (5” al intronului cu 2” al A) Eliberarea lui U4 din complex – duce la clivarea
capătului 3” al intronului. Intronul se înlătură împreună cu U2,U5, U6
Formarea legăturilor 3”-5”fosfodiesterice între capetele exonilor
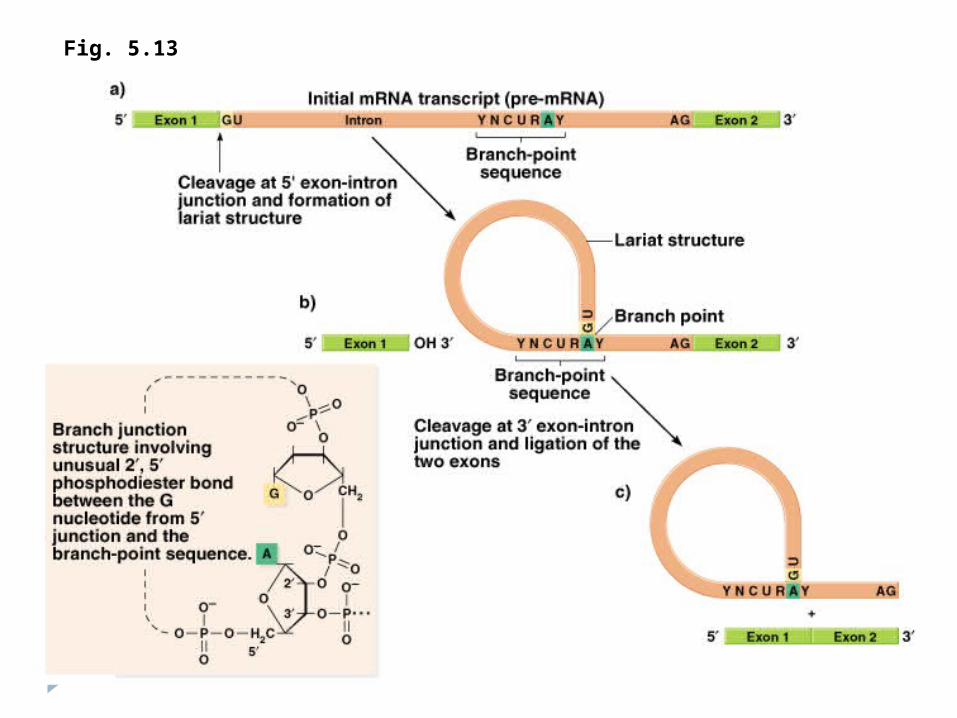
Fig. 5.13
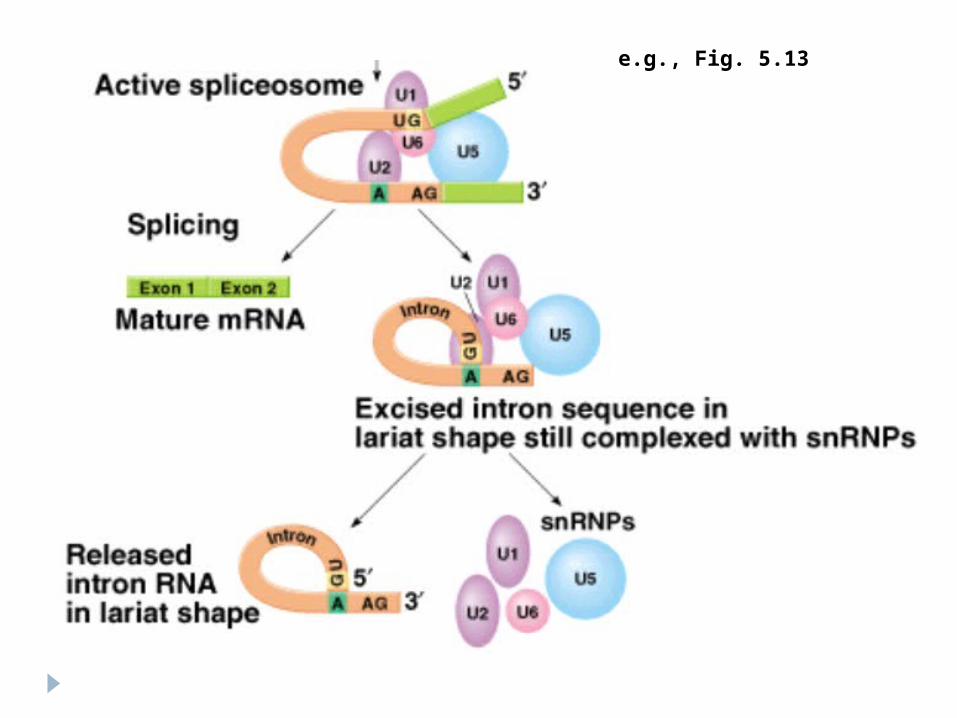
e.g., Fig. 5.13

Transcripţia inversă sinteza ADN pe catena de ARN Matriţa – ARN Substrat – dRNTP:dATP, dGTP, dCTP, TTP Enzima – revers transcriptaza Caracteristic viruşilor oncogeni Mecanismul:a. Revers transcriptaza sintetizează pe ARN viral catena de
ADN- hibrid: ADN_ARNb. Scindarea ARN viral de o nucleazăc. Autoreplicarea ADN – cu formarea unui duplex de ADN

Reglarea sintezei proteinelor Sinteza proteinelor nu e constantă – ea trebuie să se
adapteze cerinţelor vitale Celulel dispun de 3 tipuri de enzime :1. Constitutive - se sintetizează în celulă cu o viteză
constantă.2. Inductible - sunt E a căror c% depinde de prezenţa sau
absenţa în mediu a unui compus denumit inductor. Sunt implicate în căile catabolice,
3. Represible - sunt E a căror c% depinde de prezenţa sau absenţa în mediu a unui compus denumit corepresor. Sunt implicate în căile anabolice.
În mod normal cantitatea de E inductibile în celule este foarte mică,dar ea poate creşte atunci când apare necesitatea utilizării substratului E respective ( S care se comportă ca inductor).
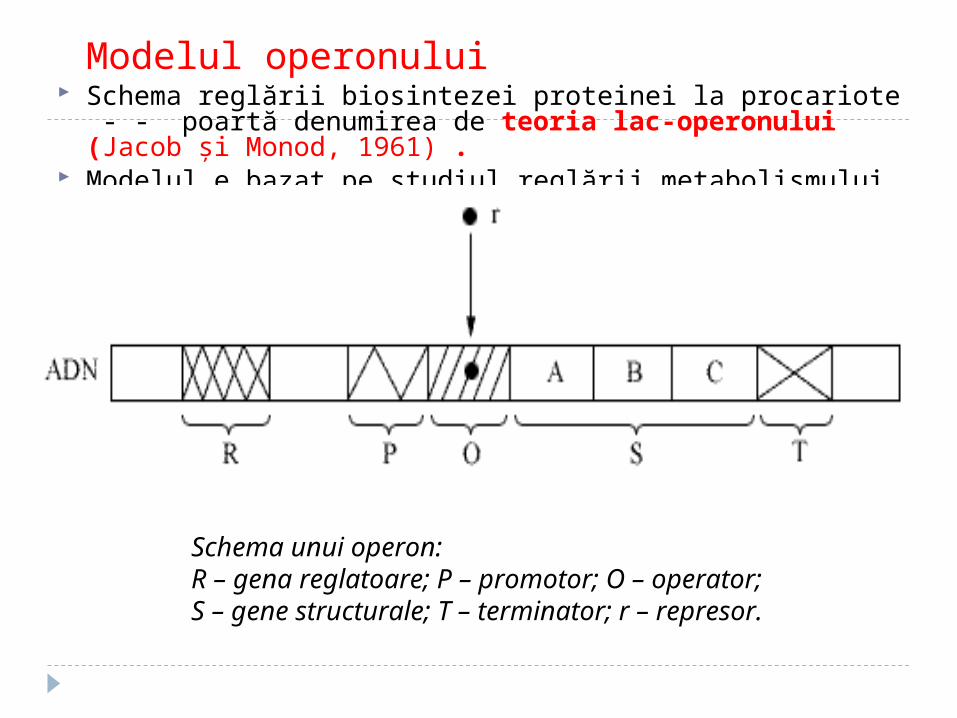
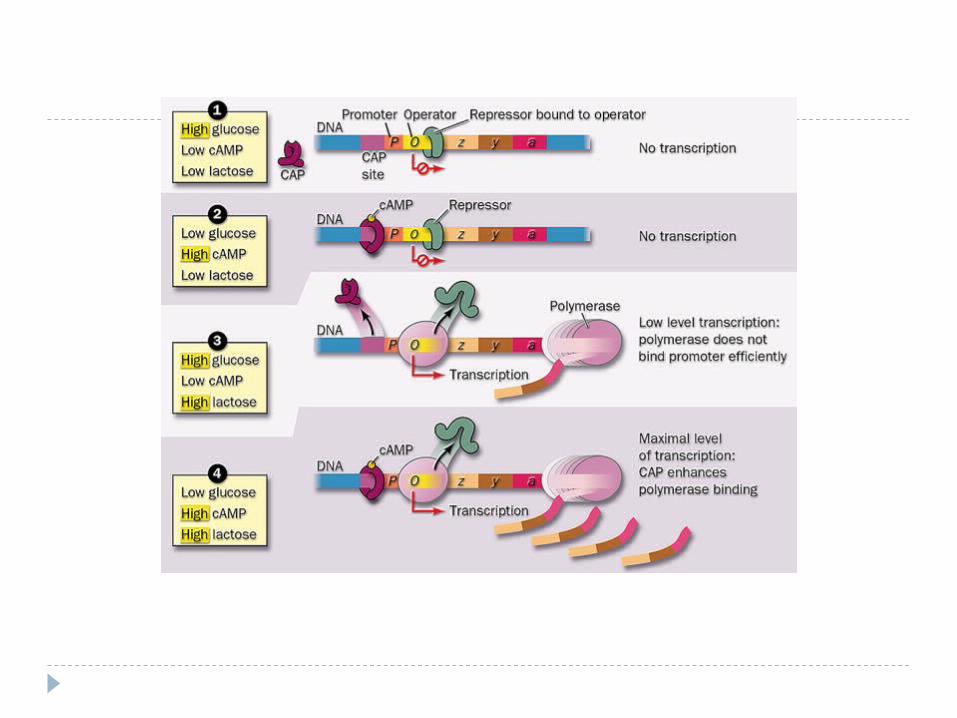
Modelul operonului Schema reglării biosintezei proteinei la procariote - - poartă
denumirea de teoria lac-operonului (Jacob şi Monod, 1961) . Modelul e bazat pe studiul reglării metabolismului lactozei în
Escheria coli. Schema generală a reglării activităţii genelor procariote a
fost propusă de F.Jacob şi J.Monod în 1961.
Schema unui operon:R – gena reglatoare; P – promotor; O – operator;S – gene structurale; T – terminator; r – represor.
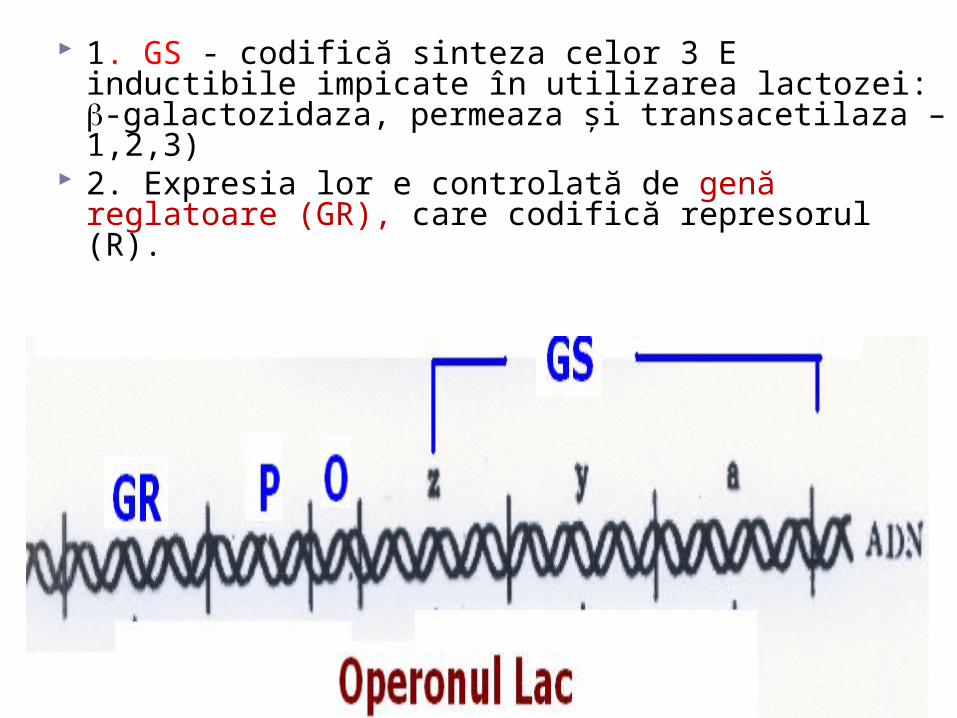
1. GS - codifică sinteza celor 3 E inductibile impicate în utilizarea lactozei: -galactozidaza, permeaza şi transacetilaza –1,2,3)
2. Expresia lor e controlată de genă reglatoare (GR), care codifică represorul (R).

Funcţionarea operonului lac
1. GR determină sinteza unei proteine numită R
2. R se leagă de operator (O). Legarea R la O blochează accesul ARN –
polimerazei la promotor avînd ca rezultat suprimarea transcrierii GS.
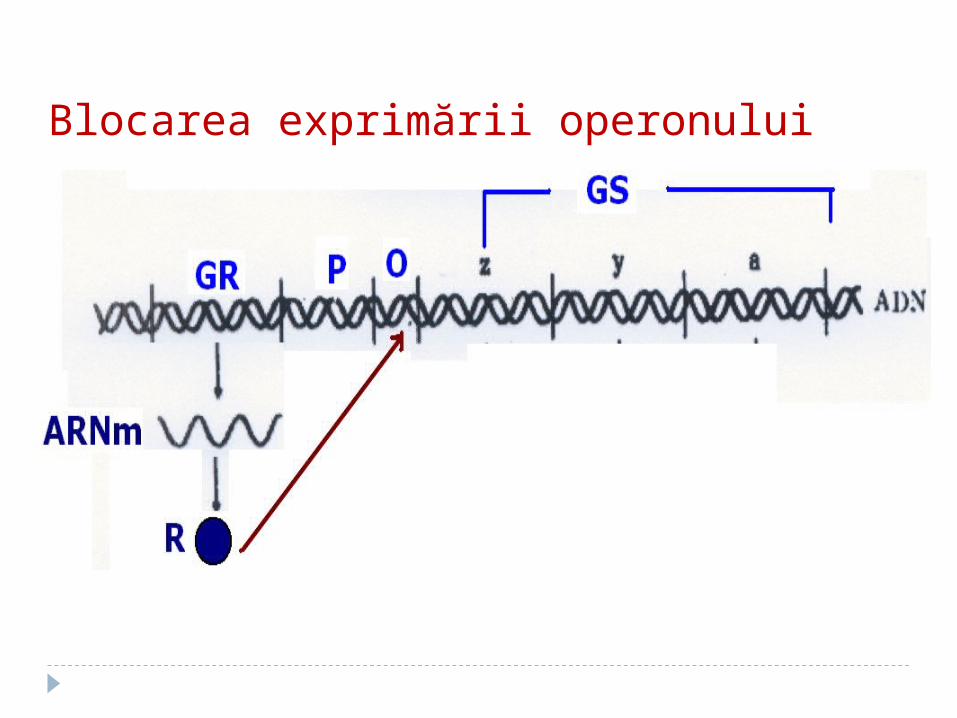
Blocarea exprimării operonului
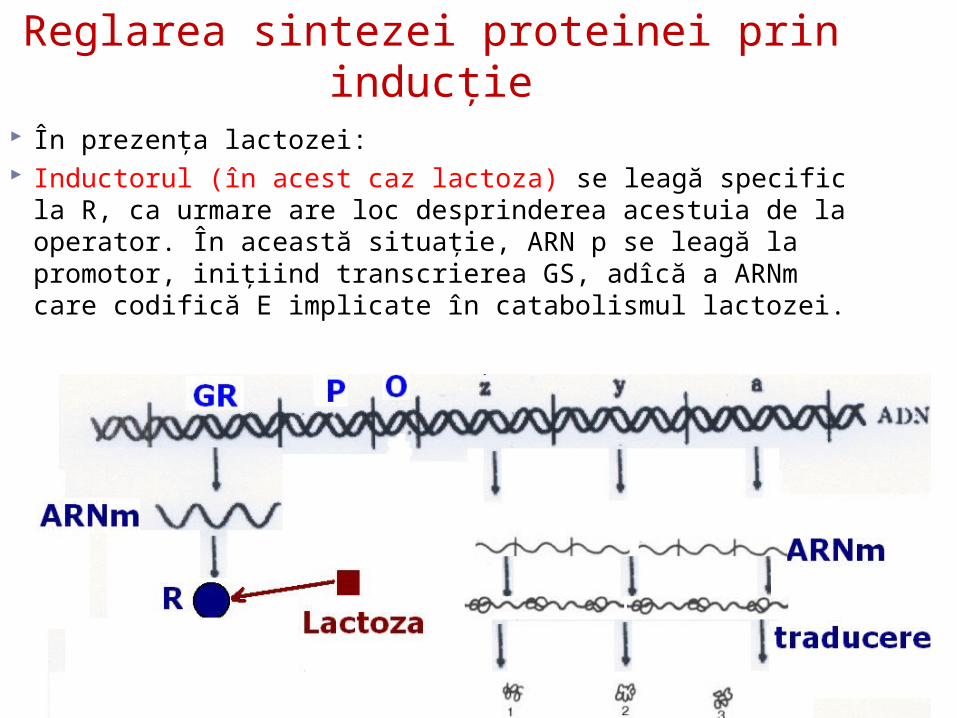
Reglarea sintezei proteinei prin inducţie
În prezenţa lactozei: Inductorul (în acest caz lactoza) se leagă specific la R, ca
urmare are loc desprinderea acestuia de la operator. În această situaţie, ARN p se leagă la promotor, iniţiind transcrierea GS, adîcă a ARNm care codifică E implicate în catabolismul lactozei.
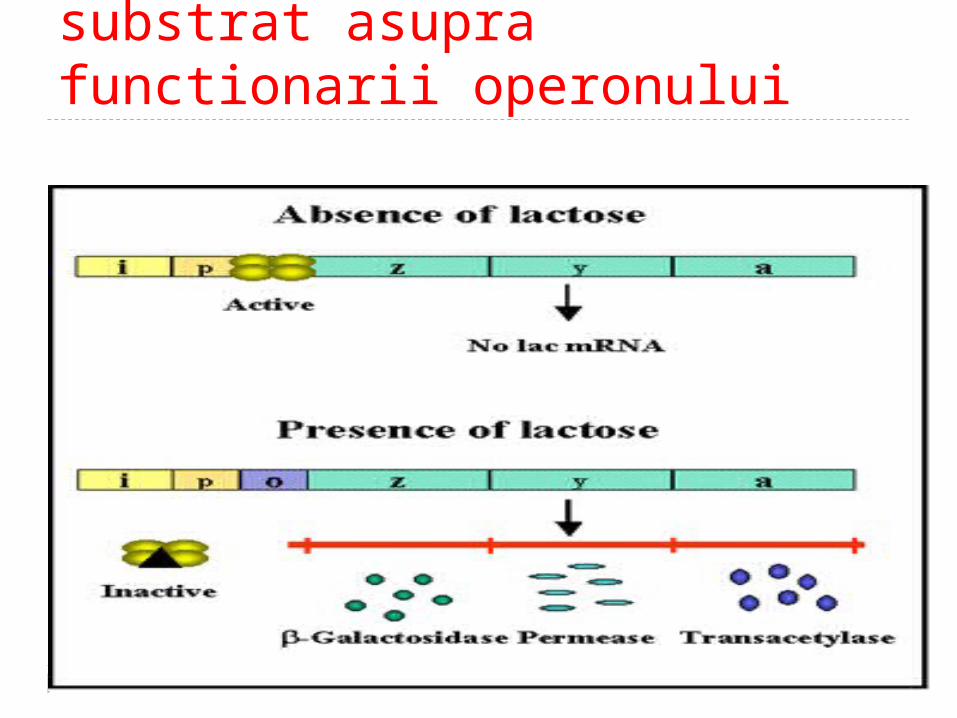
Efectul tipului de substrat asupra functionarii operonului
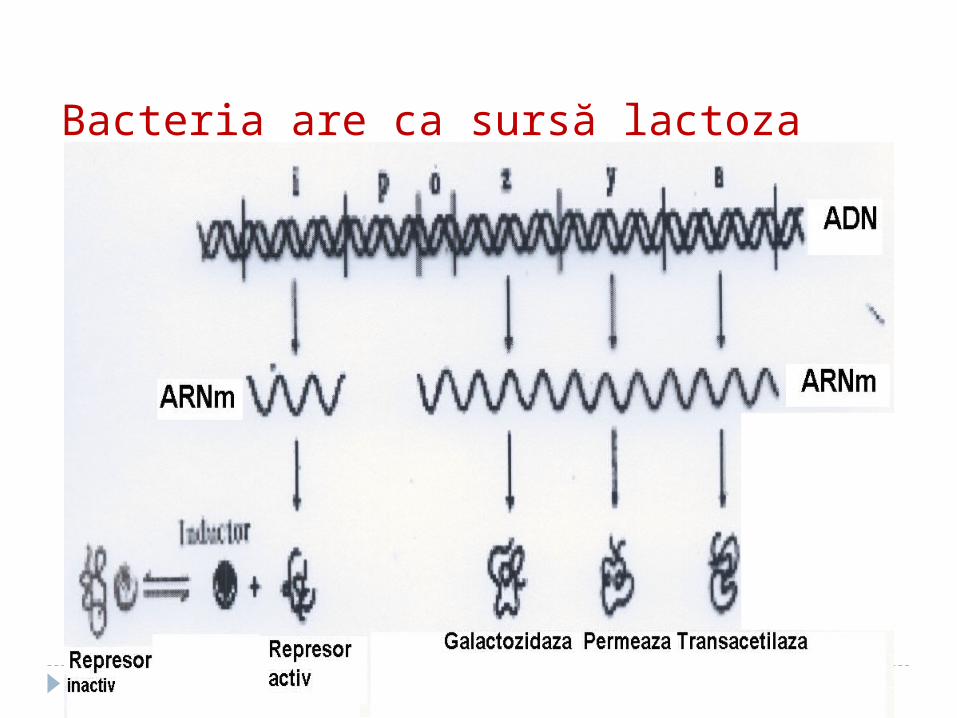
Bacteria are ca sursă lactoza

Ce se întâmplă dacă bacteria dispune simultan de glucoză şi lactoză?
Bacteria nu consumă energie pentru sinteza lac-operonului, atâta timp cât dispune de glucoză.
Bacteria creşte pe seama glucozei - şi numai atunci când c% acesteea devine minimă începe să utilizeze lactoza. Metabolizarea simultană a glucozei şi lactozei sunt excluse.
Cum se comută activitatea bacteriei pe utilizarea lactozei când c% glucozei scade?
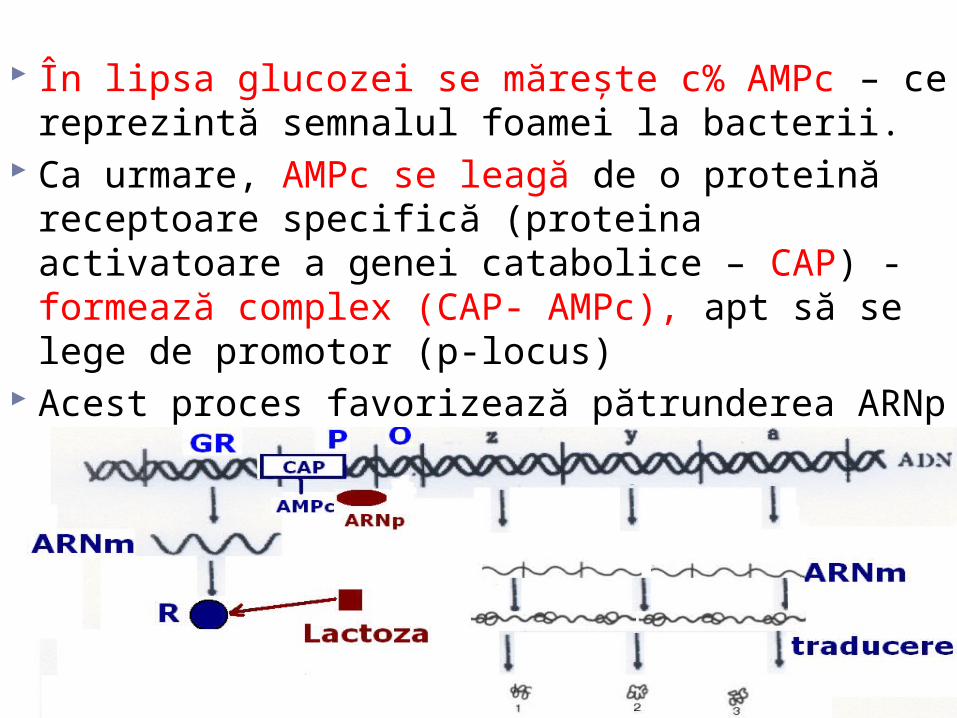
În lipsa glucozei se măreşte c% AMPc – ce reprezintă semnalul foamei la bacterii.
Ca urmare, AMPc se leagă de o proteină receptoare specifică (proteina activatoare a genei catabolice – CAP) - formează complex (CAP- AMPc), apt să se lege de promotor (p-locus)
Acest proces favorizează pătrunderea ARNp în locusul de reglare. Dacă lactoza este prezentă în mediu, operatorul este liber şi ARNp efectuează transcrierea genelor lac.
CAP dispune de 2 centre: pentru AMPc şi pentru ADN

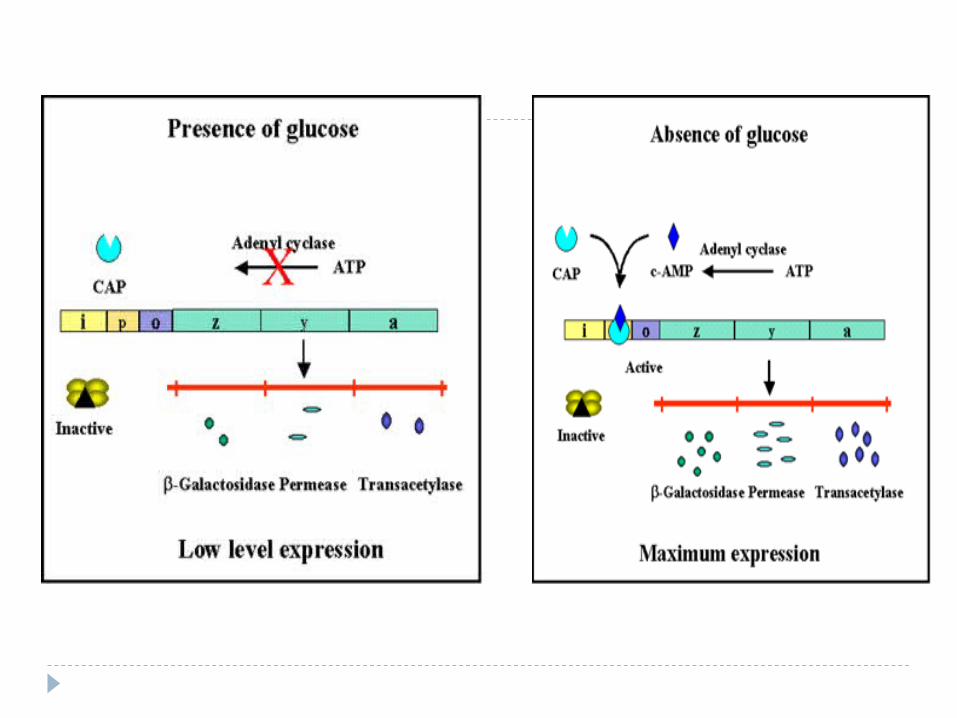
Reglarea lac-operonului într-un mediu ce conţine glucoză
Cu cât c% glucozei e mai mare, c%AMPc – e mai mică. Lipseşte şi complexul CAP- AMPc. În final ARNp nu se leagă de P şi GS nu sunt transcrise,
indiferent dacă există sau nu lactoză, indiferent de faptul dacă operatorul este sau nu ocupat de R.
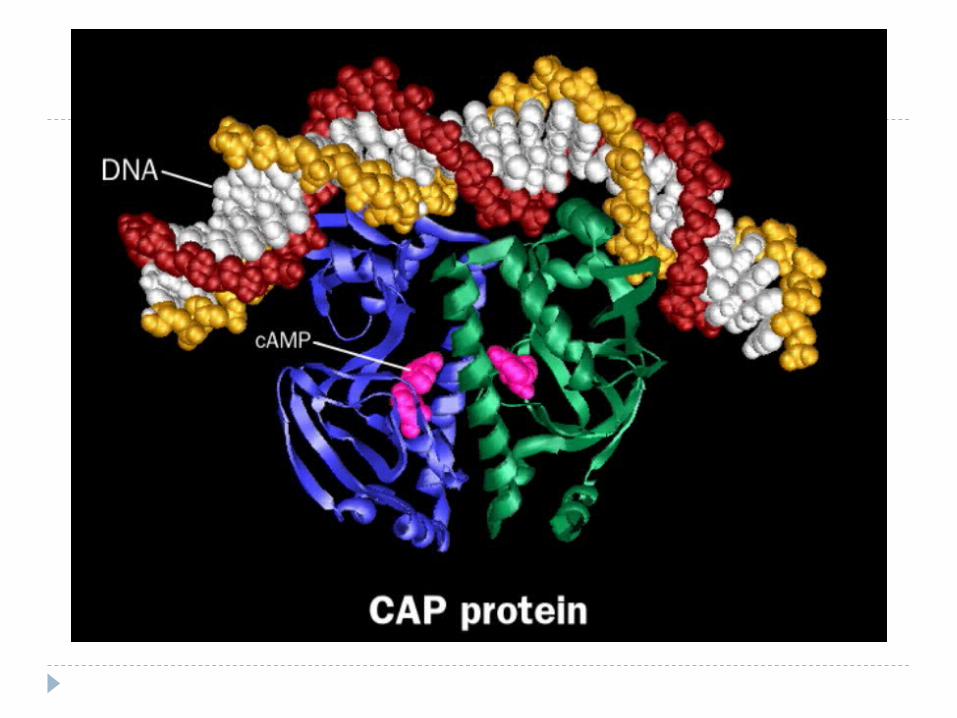
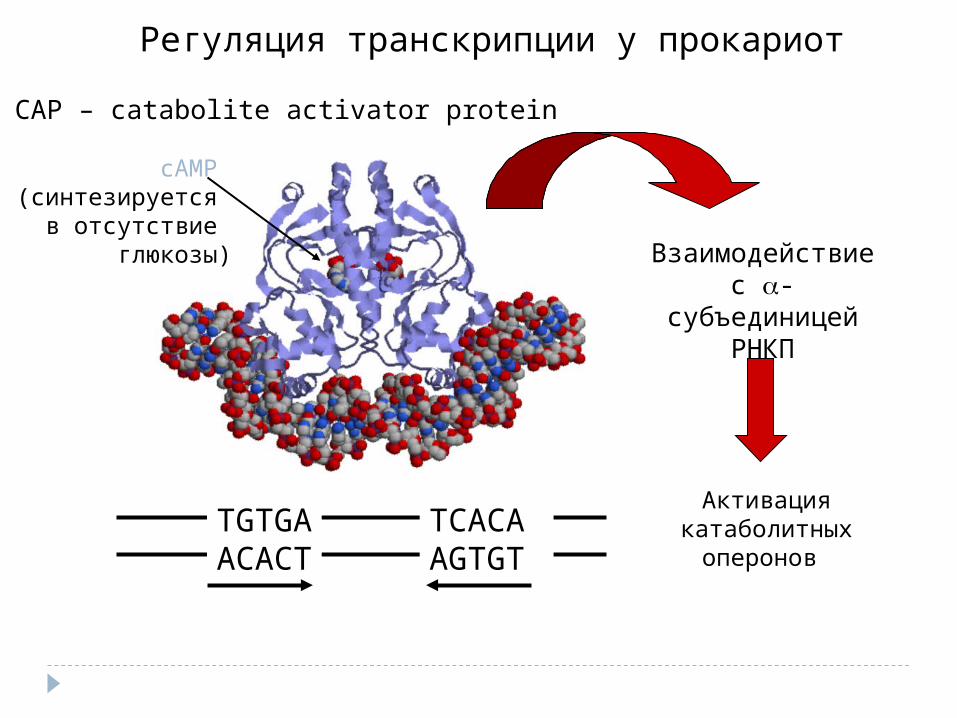
Регуляция транскрипции у прокариот
Взаимодействие с -субъединицей
РНКП
СAP – catabolite activator protein
Активация катаболитных
оперонов
cAMP (синтезируется
в отсутствие глюкозы)
TGTGAACACT
TCACA AGTGT

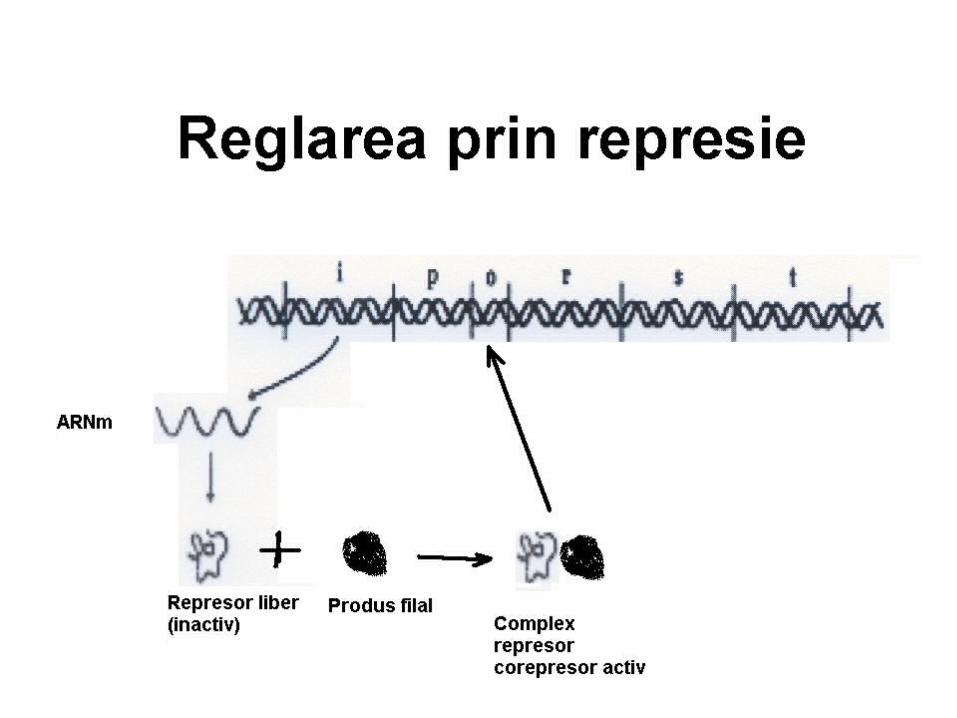
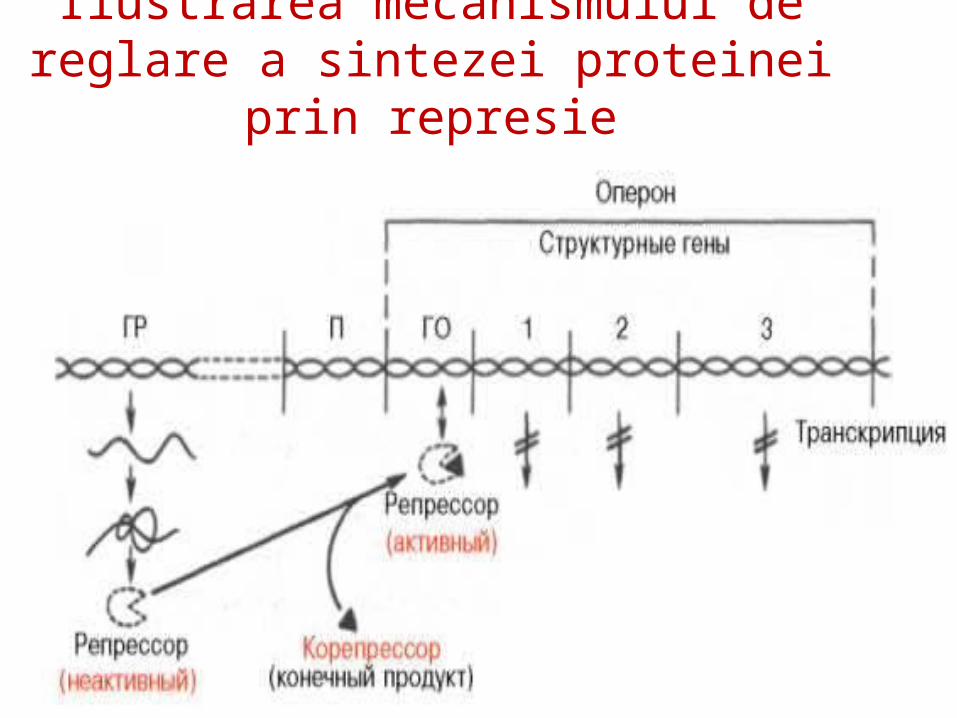
Ilustrarea mecanismului de reglare a sintezei proteinei prin represie
Teoria operonului explică şi represia prin produs final al biosintezei E
Ex: sinteza His: la c% mari de His (corepresor CoR) – se leagă de R, modificându-i conformaţia – activându-l – în rezultat favorizează legarea R la O.
His – produs final, CoR- sistează transcrierea genelor ce codifică E implicate în propria sa sinteză
Ilustrarea mecanismului de reglare a sintezei proteinei prin represie
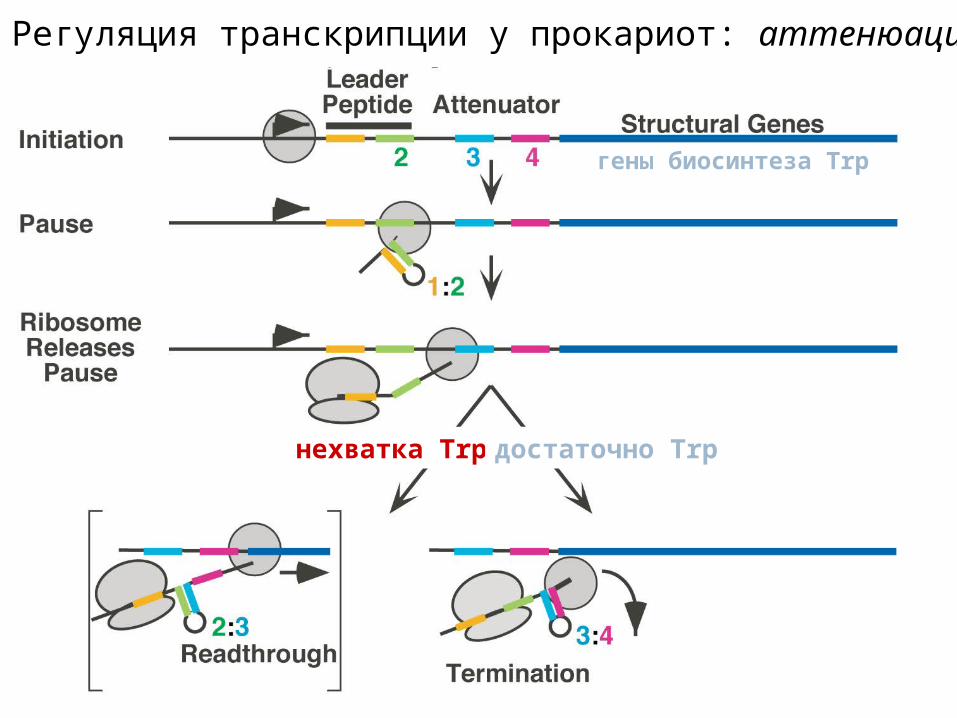
Регуляция транскрипции у прокариот: аттенюация
гены биосинтеза Trp
нехватка Trp достаточно Trp

REZUMĂM: GR controlează exprimarea anumitor GS prin
intermediul unei proteine – R Ra – suprimă sinteza de ARNm, deci de proteine;
R inactivat – permite transcrierea GS şi sinteza proteinei
E inductibile - când în mediul apare I – R se inactivează – are loc sinteza ARNm- proteinei
E represibile – R este inactiv – are loc transcripţia şi translaţia. Când în mediu se acumulează produsul final al căii anabolice (CoR)- R se activează, formarea complexului R-CoR – şi sistarea transcripţiei şi translaţiei.

Reglarea sintezei la eucariote Atât la nivelul transcripţiei cât şi la nivelul translaţiei Reglarea hormonală - reglarea biosintezei proteice la
nivelul transcrierii:cortizol- induce sinteza E gluconeogenezei; estrogenii,
androgenii, vitamina D – sinteză de proteine specifice) Amplificare genică – anumite evenimente produse la
nivelul genomului pot modifica numărul de exemplare a unei proteine sau al unui ARNm. Dacă celula are nevoie de o P în cantitate mare, ea creşte nr de copii per genom a genei ce codifică P respectivă (iniţiere repetată în cursul sintezei ADN)
.

Reglarea sintezei la eucariote Reglarea exspresiei genetice prin moleculele
proteice legate de ADN (histonele) – sinteza ARN pe ADN e inhibată prin adaosul de histone
Metilarea ADN-ului – o genă mai puţin metilată are şanse mai mari de a se exprima comparativ cu una metilată.
Reglarea proteinei la nivelul translaţiei – e posibilă prin acţiunea factorilor proteici, care contribuie iniţierea, elongarea, terminarea
Codul geneticTranslaţiaReglarea sintezei proteinei

Obiectivele: Codul genetic. Proprietăţile. Ribozomii - sediul sintezei proteinelor, structura lor. Procesul de translare (sinteza proteinelor).
Modificările posttranslaţionare ale proteinelor. Reglarea biosintezei proteinelor. Inducţia şi represia
enzimelor. Inhibitorii sintezei proteice. Polimorfismul proteinelor (variantele hemoglobinei,
enzimelor, grupelor sanguine). Bolile ereditare şi diagnosticul lor biochimic.

Codul genetic
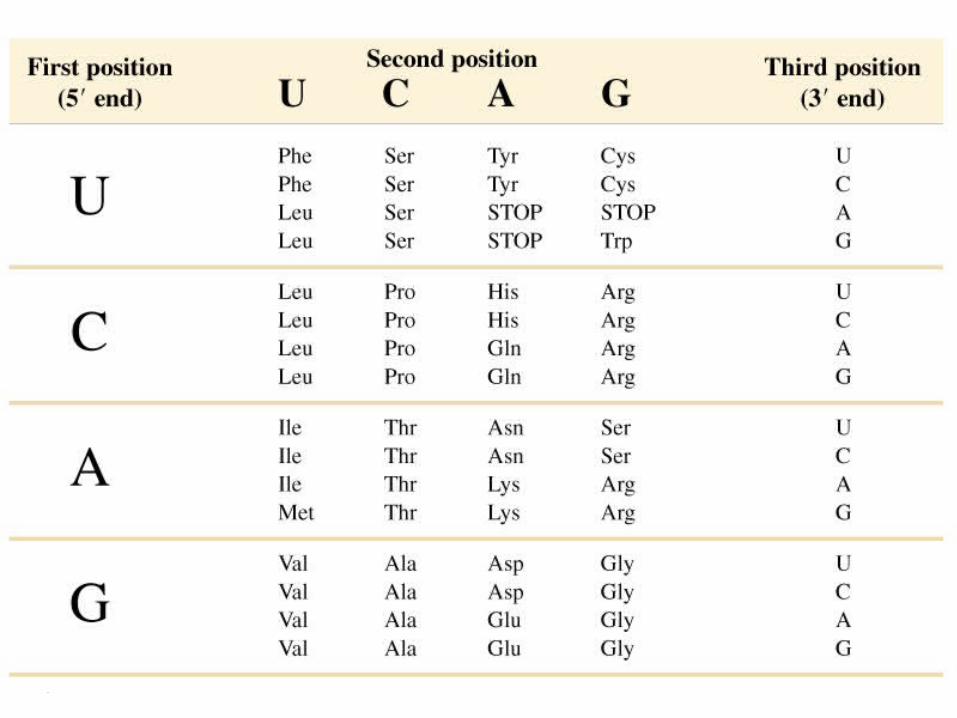
Informaţia genetică referitor la biosinteza proteinelor se transmite cu ajutorul codului genetic - dicţionar ce traduce secvenţa nucleotidelor din ARN în succesiunea AA din lanţul polipeptidic.

Proprietăţile codului genetic Este triplet -64 codoni: 3 nonsens:UAG; UGA; UAA; 61 –
codifică AA corespunzători- este degenerat - unui AA poate să-i corespunda mai
mulţi codoni (Ex. Arg, Leu, Ser - codificate de 6 codoni; Met- şi Trp - un codon). Codonii unui aminoacid sint sinonime. Specificitatea codonului e determinată de primele două litere. Degenerarea se referă la nivelul nucleotidului 3 din codon sau 1 din anticodon care oscileaza.
- nu este ambiguu- acelaşi triplet nu semnifică 2 AA diferiţi
- Are o structură liniară (colinear) – o concordanţă liniară între genă şi proteina codificătoare
- Nu se suprapune (excepţie- viruşii)

Proprietăţile codului genetic
- Este universal – toate veţuitoarele utilizează acelaşi mecanism de traducere (abatere prezintă codul genetic al mitocondriilor);
- nu are virgule, semne de punctuatie - ce ar indica începutul şi sfîrşitul fiecarui codon.
1. AUG - este codonul de initiere 2. UAG, UAA,UGA - codoni stop (non sens) 3. Toţi codonii cu U (în pozitia 2) codifica AA hidrofobi 4. codonii cu A în pozitia -2 codifică AA polari5. Uracilul în poziţia 1 prezintă codonul nonsens 6. dacă în anticodon în directia (5'->3') prima bază nucleotidică
e: a) Citozina sau Adenina, el va citi un singur codon;
b) Uracilul sau Guanina el va citi 2 codoni; c) inozina - respectiv va citi 3 codoni
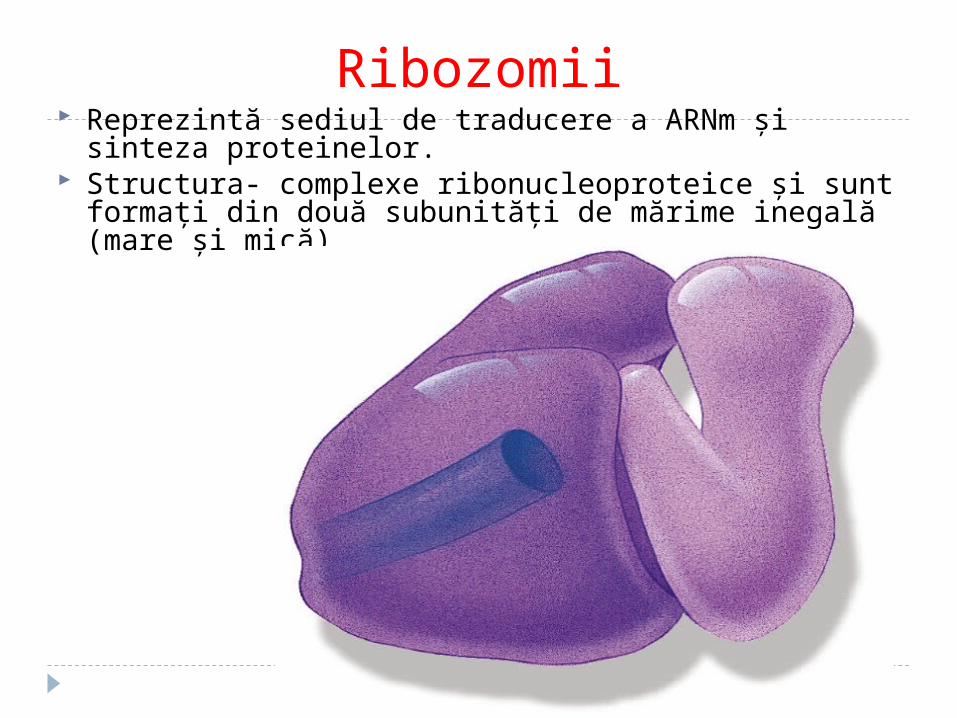
Ribozomii Reprezintă sediul de traducere a ARNm şi sinteza proteinelor. Structura- complexe ribonucleoproteice şi sunt formaţi din
două subunităţi de mărime inegală (mare şi mică)

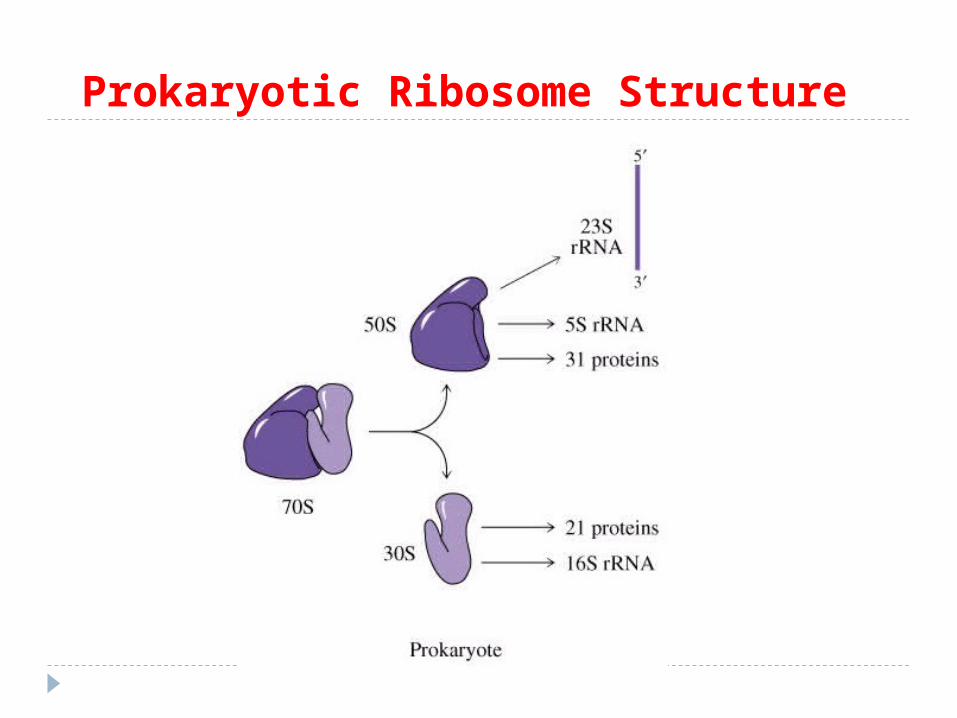
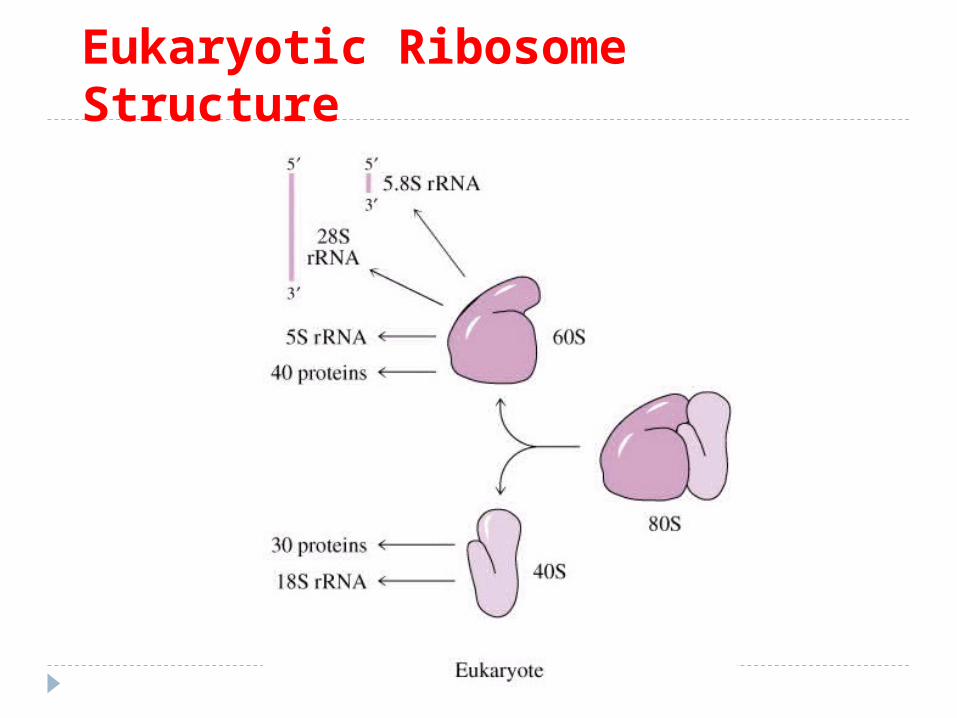
Structura ribozomilor: procariotici:subunitatea 30 S – conţine ARNr 16S şi 21 proteine.subunitatea 50S – ARN r 5S, 23S şi 31 proteine. Sinteza ARNr şi formarea subunităţilor are loc în citoplasmă. eucariotici: subunitatea 40S – ARNr 18S şi 33 proteine. subunitatea 60S – ARN r 5S, 5,8S, 28S şi 49 proteine. ARNr –
se formează în nucleol. Ribozomul va avea constanta de sedimentare 70S la
procariote si 80S la eucariote. S – este coeficientul de sedimentare Svedberg, care depinde
de forma, densitatea şi dimensiunea particulelor.
Prokaryotic Ribosome Structure
Eukaryotic Ribosome Structure

Centrele catalitice ale ribosomilor
Situsul A - aminoacil – responsabil de unirea complexului aminoacil- ARNt
Situsul P – peptidil – găzduieşte ARNt legat de un lanţ polipeptidic deja sintetizat
Situsul E – e responsabil de eliminarea ARNt
În starea complet nedisociată ribozomii sunt activi.
Deplasarea lberă a ribozomilor în diferite sectoare ale celulei, sau combinarea lor în diferite locuri cu membranele reticulului endoplasmatic oferă posibilitatea de asamblare a proteinei în celulă.

POLIRIBOZOMUL Mai mulţi ribozomi
pot citi simultan acelaş ARN mesager pe care il parcurg in acelaş sens.
Se constituie astfel un poliribozom, structură ce permite accelerarea sintezei proteice.

Translaţia Translaţia sau biosinteza proteinelor propriu zisă. Bazele moleculare ale translaţiei:1. m-RNA ca matriţă genetică (determină succesiunea
AA în proteină);2. aminoacil – ARNt;3. ribozomii ca maşini moleculare pentru unirea
succesivă a AA în catena polipeptidică conform programului mRNA;
4. GTP ca sursă de energie;5. “factorii” proteici;6. ca cofactori - unii ioni (Mg 2+, K+).

Etapelese realizeaza in 5 etape: Activarea AA. Iniţierea lanţului polipeptidic. Elongarea lanţului polipeptidic. Terminarea lanţului polipeptidic si eliberarea
acestuia. Prelucrări post traducere ale proteinei sintetizate.

Activarea AA are loc în citozol Sunt necesare:1. AA (procariote – Nformil-Met; la eucariote – Met)2. ARNt3. ATP, Mg, K4. E – aminoacil ARNt sintetaza a. Posedă 4 centre: pentru AA, ATP, ARNt, H2Ob. Specificitatea E e determinată de structura ARNtc. Fidelitatea e asigurată de capacitatea de autocontrold. Conţine grupări libere sulfhidrilice
e. sunt ligazele, care au o specificitate absolută.
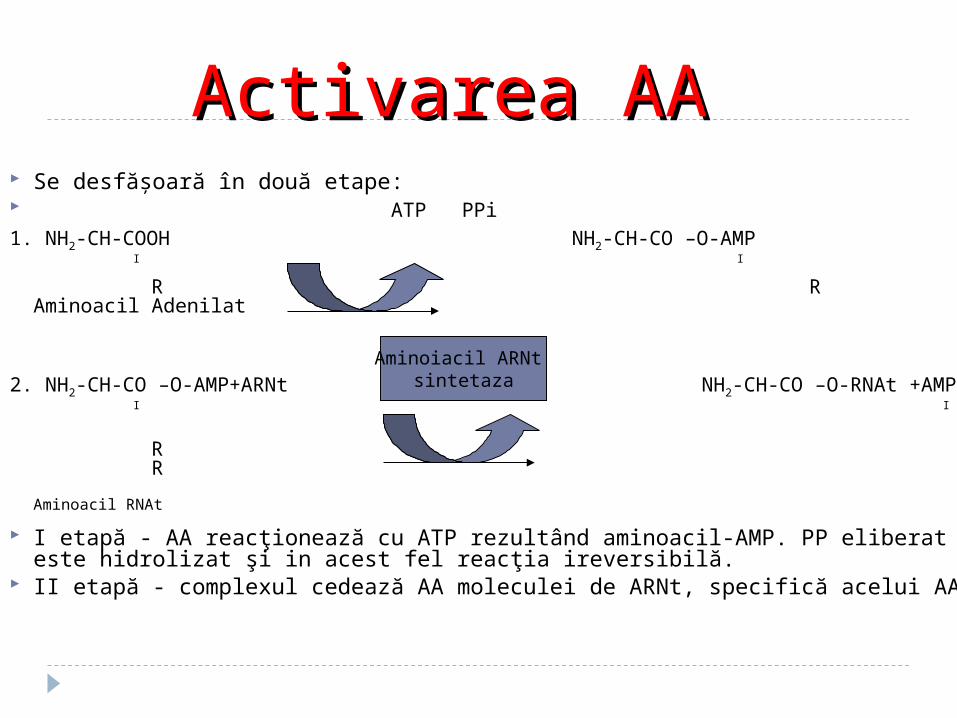
Se desfăşoară în două etape: ATP PPi1. NH2-CH-COOH NH2-CH-CO –O-AMP I I
R R Aminoacil Adenilat
2. NH2-CH-CO –O-AMP+ARNt NH2-CH-CO –O-RNAt +AMP I I
R R Aminoacil RNAt
I etapă - AA reacţionează cu ATP rezultând aminoacil-AMP. PP eliberat este hidrolizat şi in acest fel reacţia ireversibilă.
II etapă - complexul cedează AA moleculei de ARNt, specifică acelui AA
Activarea AA Activarea AA
Aminoiacil ARNt sintetaza
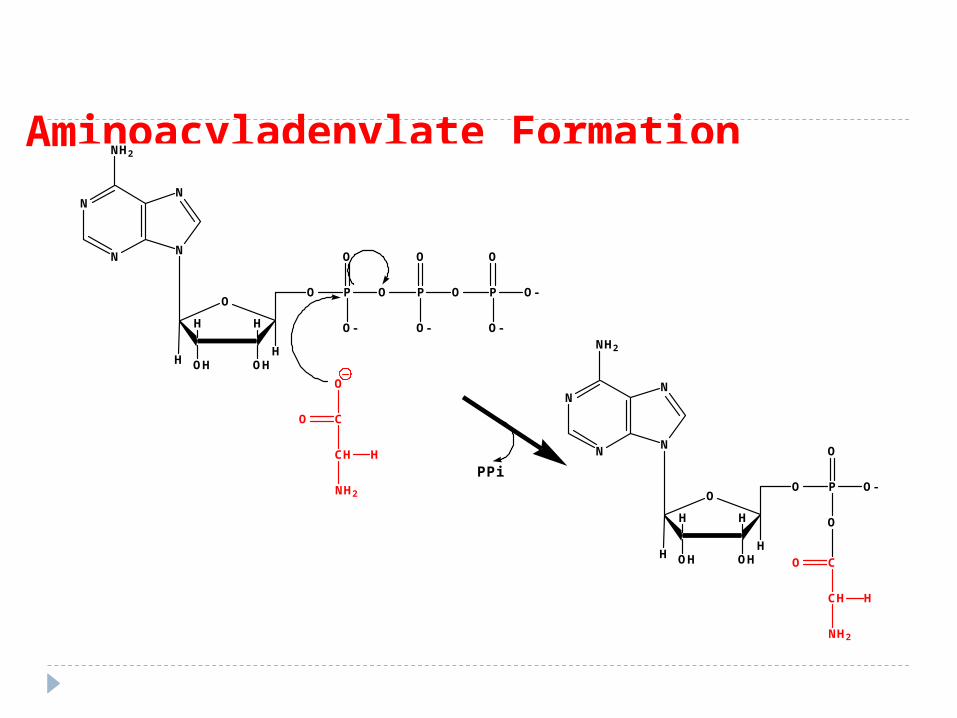
Aminoacyladenylate Formation
O
N
NN
N
NH2
O
OH OH
H H
HH
O P
O-
O
OP
O-
O
O-P
O-
O
NH2
CH
C
H
O
O
PPiO-
N
NN
N
NH2
O
OH OH
H H
HH
O P
O
O
NH2
CH
C
H
O
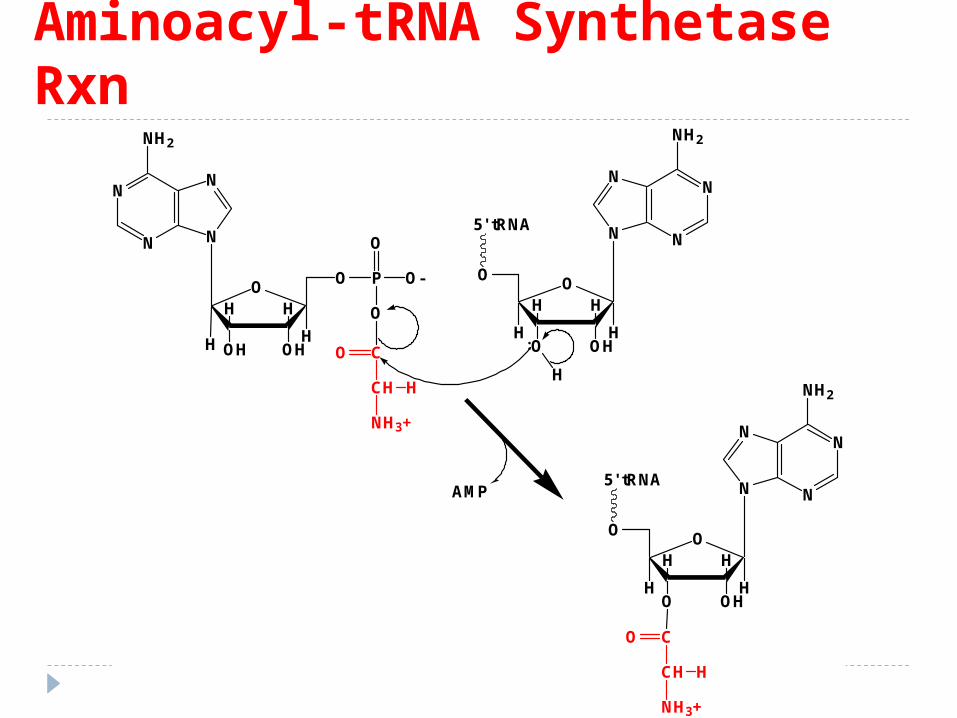
Aminoacyl-tRNA Synthetase Rxn
N
NN
N
NH2
O
OHO
HH
HH
O
5' tRNA
H
N
NN
N
NH2
O
OHO
HH
HH
O
5' tRNA
NH3+
CH
C
H
O
O-
N
N N
N
NH2
O
OH OH
H H
H H
O P
O
O
NH3+
CH
C
H
O
AMP

Activarea AA Esenţa procesului este fixarea AA la ARNt în zona
acceptorie la 3' CCA –OH Activarea AA consumă 2 legături macroergice
E recunoaşte AA greşit fixat pe ARNt - îl elimina şi îl înlocuieşte cu AA corespunzător (prezintă şi un locus hidrolitic).

Translaţia propriu zisă Citirea ARNm se face în direcţia 5‘- 3' iar
proteina se sintetizează de la capătul “N”terminal la “C” terminal
se desting trei etape: Iniţierea Elongarea terminarea.

Iniţierea Necesar:1. ARNm (AUG)2. Ribosomul cu subunităţile
disociate3. ARNt f-met (Met)4. GTP, Mg5. IF1, IF2, IF3 Scopul: formarea complexului de
iniţiere

Formarea complexului de iniţiere: Subunitatea mică leagă IF3 şi
previne reasocierea ribosomilor La subunitate adiţionează ARNm
(AUG) La complex adiţionează IF1 legat de
formil Met-ARNt și IF2 legat de GTP Îndată cum are loc fixarea
anticodonului fMet-ARNt cu codonul AUG (ARNm), are loc hidroliza GTP, eliberarea FI şi unirea subunităţilor
Formil Met-ARNt –e fixată în centrul P
30SFI3
P 50S A
FI1
FI2
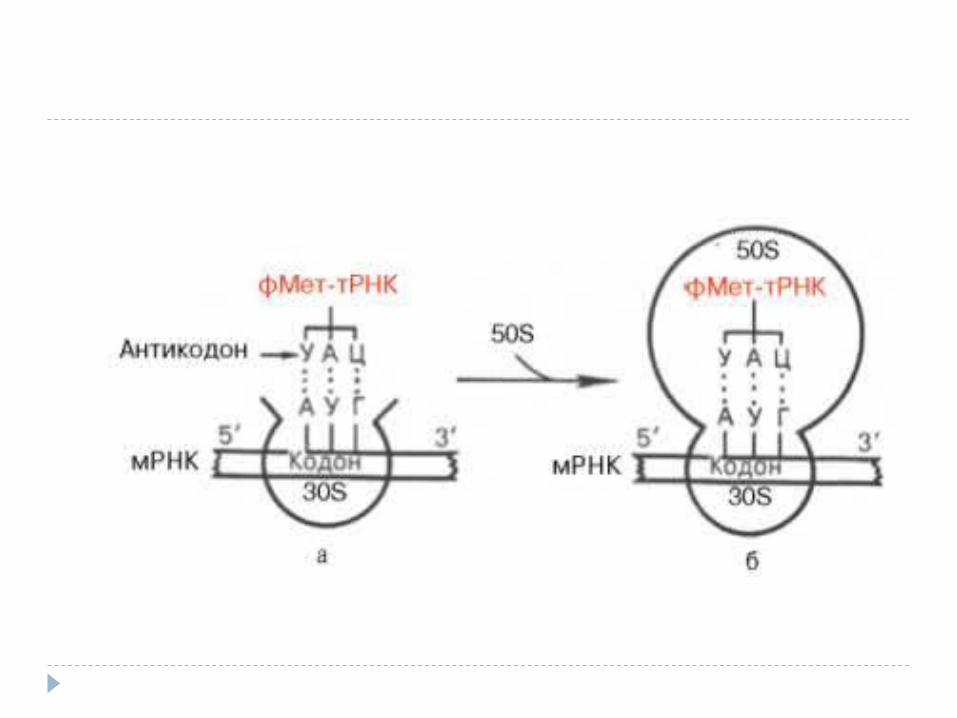

Elongarea Necesar:1. ARNm cu următorul codon2. ARNt cu următorul AA3. GTP4. FE: Tu, Ts, G Elongarea translaţiei include trei etape:1. Legarea aminoacil – ARNt;2. Transpeptidarea- formarea legăturii peptidice,3. Translocarea (deplasarea ARNm cu un codon).
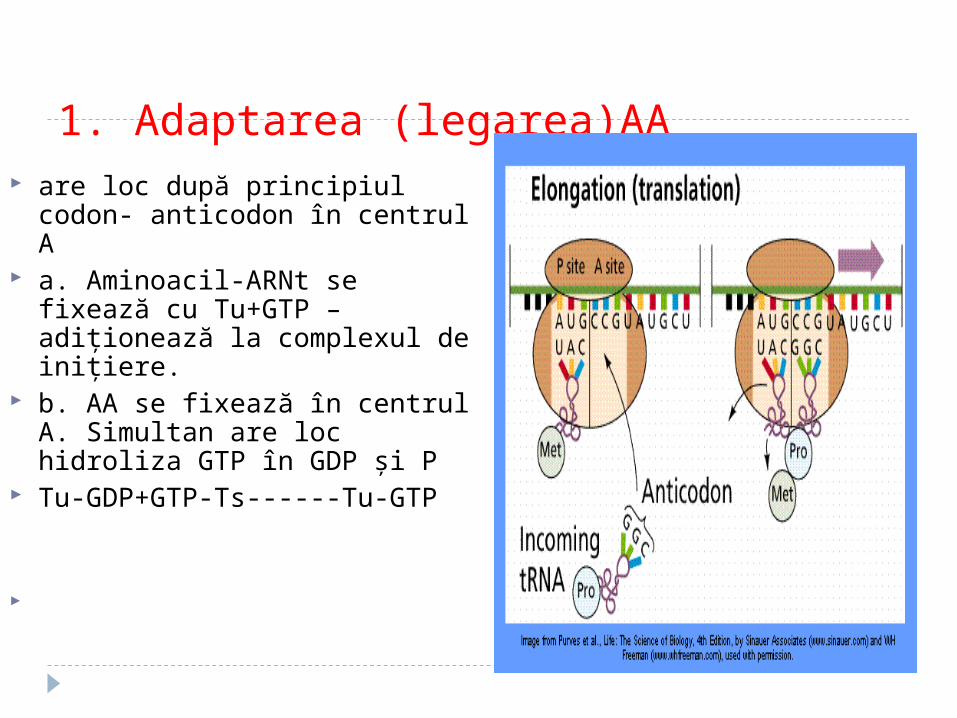
1. Adaptarea (legarea)AA are loc după principiul codon-
anticodon în centrul A a. Aminoacil-ARNt se fixează cu
Tu+GTP – adiţionează la complexul de iniţiere.
b. AA se fixează în centrul A. Simultan are loc hidroliza GTP în GDP şi P
Tu-GDP+GTP-Ts------Tu-GTP
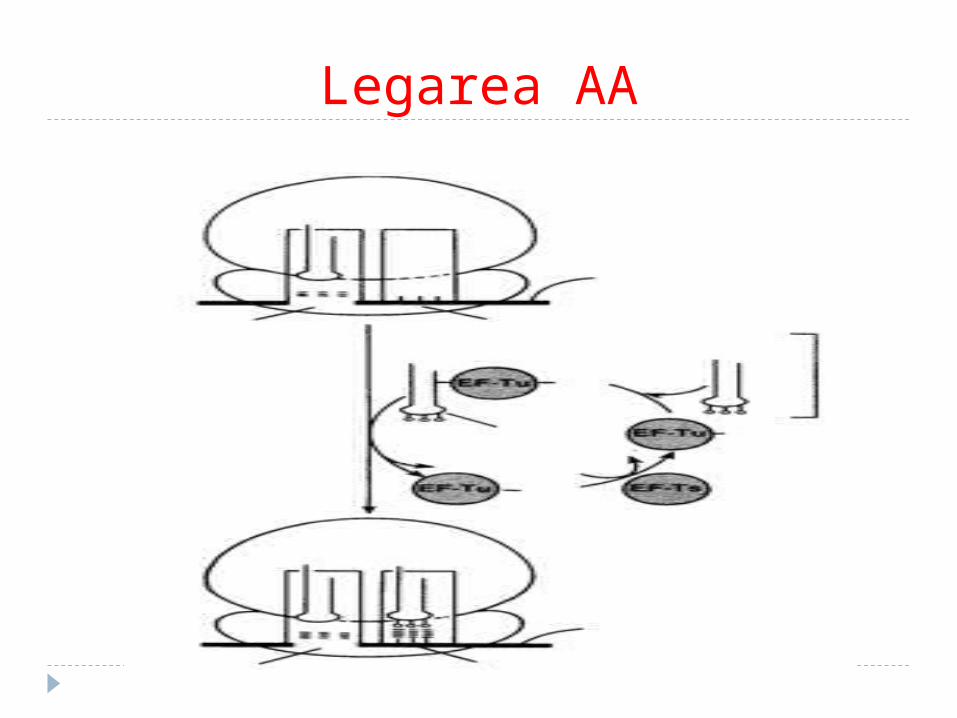
Legarea AA

2. Transpeptidarea este formarea legăturii peptidice între doi
aminoacizi. AA din centrul P sub acţiunea
peptidiltransferazei trece în centrul A. Se formează dipeptida În centrul P rămîne ARNt liber
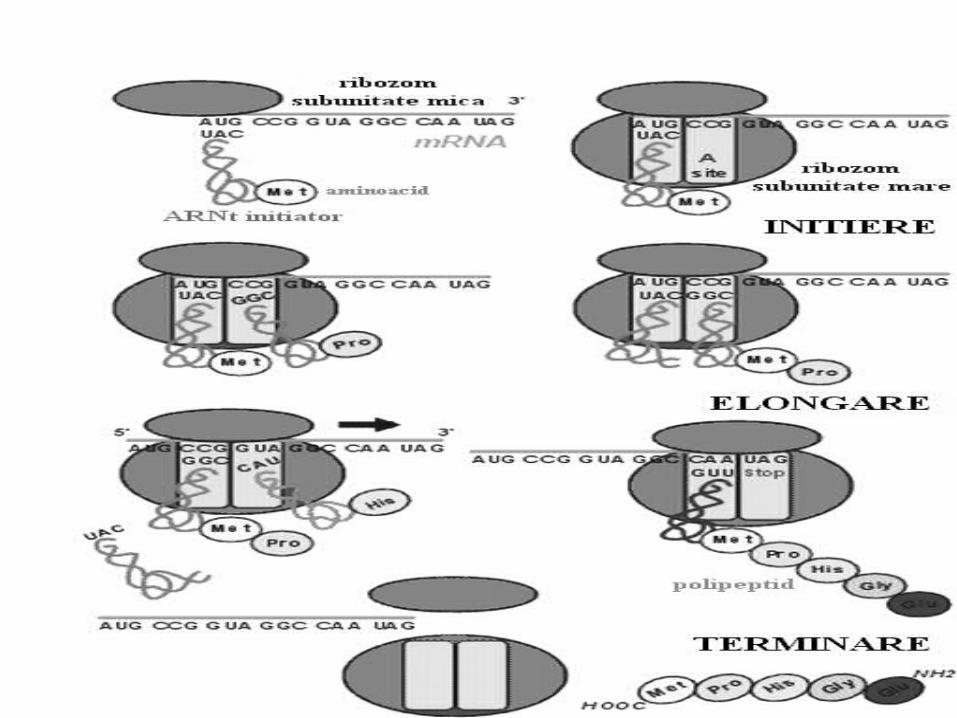

3. Translocarea deplasarea ribosomului pe ARNm cu un
triplet în direcţia 5‘- 3' . Dipeptida din centrul A trece în centrul P
sub acţiunea factorului G (translocazei) şi GTP
ARNt din P părăseşte ribosomul
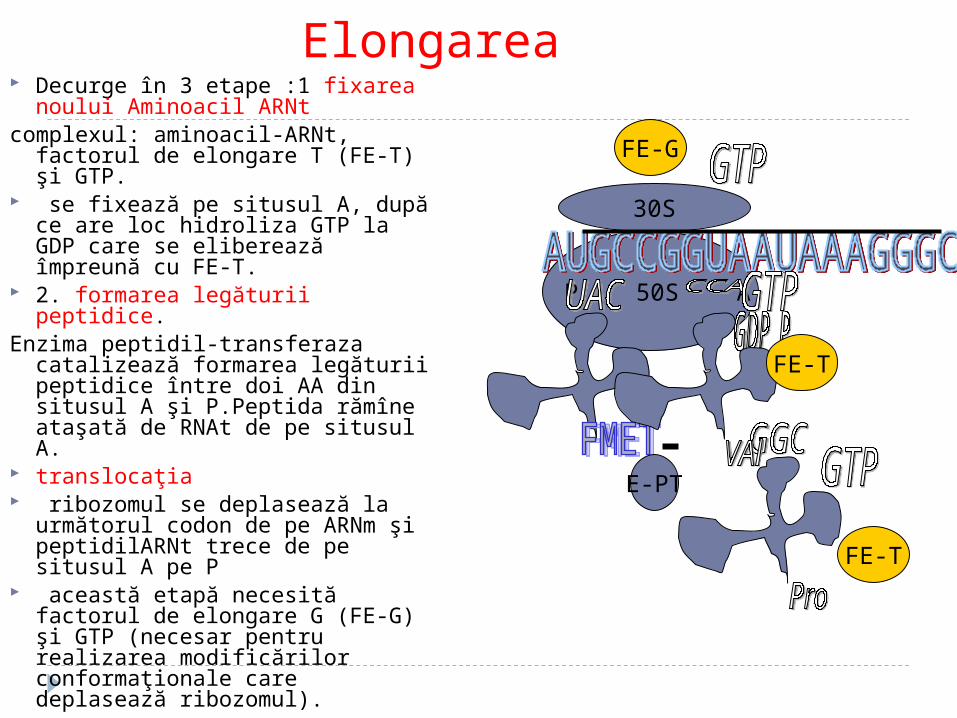
Elongarea Decurge în 3 etape :1 fixarea noului
Aminoacil ARNt complexul: aminoacil-ARNt, factorul de
elongare T (FE-T) şi GTP. se fixează pe situsul A, după ce are
loc hidroliza GTP la GDP care se eliberează împreună cu FE-T.
2. formarea legăturii peptidice. Enzima peptidil-transferaza catalizează
formarea legăturii peptidice între doi AA din situsul A şi P.Peptida rămîne ataşată de RNAt de pe situsul A.
translocaţia ribozomul se deplasează la
următorul codon de pe ARNm şi peptidil ARNt trece de pe situsul A pe P
această etapă necesită factorul de elongare G (FE-G) şi GTP (necesar pentru realizarea modificărilor conformaţionale care deplasează ribozomul).
30S
P 50S A
FE-T
E-PT
FE-G
FE-T

Terminarea are loc cînd sunt întîlniţi codonii UAA, UGA, UAG
şi factorii proteici de terminare: R1, R2, S. Nici un tRNA nu se poate lega cu codonii de
terminare. Factorii de terminare: eliberează lanţul polipeptidic Elimină ARNt din centrul P Disocierea ribosomului în subunităţile respective Degradarea ARNm

DECI:
La formarea unei legături peptidice se consumă patru legături macroergice:
2 în etapa de activare a AA (ATP) şi2 în translația propriu zisă: - GTP.

Prelucrările posttraducere Modificarea capătului N- şi C-terminal; capătul N se
acetilează; Îndepărtarea secvenţei semnalizante cu ajutorul unei
peptidaze; modificarea unor AA: 1. hidroxilarea enzimatică a Pro, Lyz – obţinerea
hidroxiprolinei, hidroxilizinei . 2. Metilarea (Lyz în muşchi)3. Carboxilarea Glu - -carboxil-glutamatului (protrombină) 4. oxidarea reziduuriilor de Cis - cistinei;5. iodurarea reziduurilor de Tir ale tireoglobulinei.

Prelucrările posttraducere ataşarea unor gr. funcţionale: fosfat,
glicozil, metalelor pentru formarea fosfoproteinelor,glicoproteinelor, metaloproteinelor ş.a.
Formarea punţilor disulfurice Proteina se autoasamblează – formând
conformaţia nativă – structura tridimensională

Particularităţile biosintezei proteice la eucariote
AA iniţiator este Met ARNm este monocistronic (un singur
semnalde iniţiere) Ribosomul eucariot “nu sare” peste codonii
terminali Viteza procesului e mai mare FI (eIF1-eIF10); elongare şi terminare.

Inhibitori ai sintezei proteinei
la nivelul replicării: 1. Mitomicina –împiedica separarea catenelor de
ADN
2. Acid nalidixic –inhiba ADN giraza
la nivelul transcriptiei:- Actinomicina D - se fixeaza pe ADN
– Rifampicina - inhiba ARN polimeraza

Inhibitorii sintezei proteinei la nivelul translatiei: Streptomicina –inhiba legarea ARNt iniţiator la
subunitatea 30S Cloramfenicol -inhiba peptidil transferaza Tetraciclina - inhiba legarea ARNt la ribozomi Eritromicina,Azitromicina – blocheaza
subunitatea 50S Puromicina – blocheaza elongarea inhibând
competitiv ARNt Neomicina, Kanamicină - erori în reproducerea
codului genetic Toxina difterică - inhibă translocaza

Polimorfismul proteinelor Tipuri de Hb normale: Hb A – α2 β2 – adultă majoră Hb A2- α2 δ2 – adultă minoră (2-3%) HbF - α2 γ2 – fetală (la făt şi 80% la nou-născuţi) Hb E - α2 ε2 - embrionară Variante genetice ale Hb umane -300-
hemoglobinoze

Hemoglobinoze Hb S- înlocuirea Glu din poziţia a 6 a lanţurilor β
cu Val – deformarea şi aglomerarea hematiilor Modificări ale AA care tapetează buzunarul
hemului – duce la modificări ale oxigenării – formarea methemoglobinei (legarea Fe 3+): scăderea afinităţii pentru oxigen, dispariţia cooperativităţii)
Talazemii – care nu sintetizează lanţurile α sau β.


















