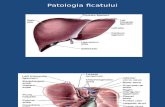
2.1_Caile nervoase de reglare a func+úiilor ficatului

of 15
-
Upload
gabriel-ionescu -
Category
Documents
-
view
212 -
download
0
Transcript of 2.1_Caile nervoase de reglare a func+úiilor ficatului
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
1/15
Controlul nervos al func iilor hepatice
1
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
2/15
Cile nervoase de reglare a funciilor ficatului: axa creier-ficat
Letiia Adela Maria Streba, Cristin Constantin Vere
Creierul primete informaii referitoare la funciile ficatului (metabolic, imun etc)(14) i, la rndul su, regleaz aceste funcii pe cale nervoas i endocrin (1, 5, 6).Comunicarea nervoas dintre ficat i sistemul nervos central este bidirecional i serealizeaz prin intermediul fibrelor nervoase vegetative, simpatice i parasimpatice,existnd o cale aferent (senzitiv) i una eferent (motorie). (1)
Calea aferent
Receptorii hepatici
O serie de autori au descris existena la nivelul ficatului a mai multor tipuri dereceptori nervoi: osmoreceptori, ionoreceptori, baroreceptori i receptori metabolici.Exist, ns, i studii care nu au evideniat prezena unora dintre aceti receptori. (7)
Osmoreceptorii
Existena osmoreceptorilor hepatici a fost demonstrat n special prin metodeelectrofiziologice. (8) Majoritatea studiilor au fost efectuate in situ, pe ficat perfuzat,nregistrndu-se activitatea nervilor hepatici afereni dup administrarea de soluiihiperosmolare. Cercetrile au artat c intensitatea stimulilor nervoi nregistrai lanivelul fibrelor vagale hepatice este direct proporional cu osmolaritatea soluieiperfuzate. (811)
IonoreceptoriiExistena ionoreceptorilor hepatici este controversat. Dei unii autori neag
prezena acestor receptori la nivelul ficatului, o serie de experimente aduc o serie deargumente n favoarea existenei ionoreceptorilor hepatici. Astfel, Passo i colab.demonstreaz existena receptorilor pentru NaCl prin experimente efectuate pe pisici. Eiau observat c efectul natriuretic obinut prin administrarea de soluie salin hiperton nvena port este superior celui realizat prin infuzia de sucroz hiperton n vena femural,ceea ce ridic problema existenei ionoreceptorilor hepatici. (12) Andrews i Orbach, prinexperimente efectuate pe iepuri, au adus i ei argumente n favoarea existeneireceptorilor hepatici pentru NaCl, concluzionnd c acetia sunt diferii deosmoreceptorii hepatici. (13)
Baroreceptorii
Primele argumente cu privire la existena baroreceptorilor hepatici au fost aduse deAndrews i Palmer n urma cercetrilor efectuate pe cini. (14) Niijima a localizat aceti
baroreceptori la nivelul ramurilor intrahepatice ale venei porte. n plus, el a artat c ocretere a presiunii n circulaia hepatic determin intensificarea impulsurilor nervoasen fibrele aferente hepatice, ceea ce constituie un argument n favoarea existeneibaroreceptorilor la nivelul ficatului. (15)
Receptorii metabolici
Unii cercettori au sugerat prezena receptorilor metabolici la nivelul ficatului. (16)Russek a fost primul care a sugerat rolul receptorilor senzitivi ai ficatului n controlulingestiei de alimente, precum i aciunea glucozei de la nivel hepatic de inducere a
2
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
3/15
saietii. (16) Nu s-a stabilit nc dac receptorii senzitivi hepatici sunt stimulai numaide glucoz sau i de ali metabolii. (16)
Studiile electrofiziologice efectuate de Niijima pe ficat izolat de porcuor de guineaau demonstrat sensibilitatea ficatului la glucoz. Autorul a nregistrat activitatea electrica ramurii hepatice a nervului vag, observnd c infuzia de glucoz reduce activitatea
nervoas, spre deosebire de alte hexoze care nu au nici un efect. (15) Acelai cercettor aevideniat creterea activitii fibrelor nervoase aferente hepatice n timpul terapiei cuinsulin, n timp ce activitatea nervoas este deprimat de colecistokinin i glucagon.(15)
Ali autori au artat c ficatul detecteaz nu numai modificrile metabolismuluiglucozei, dar i ale altor metabolii. (16) Astfel, Langhans i colab. au observat c o seriede metabolii, cum sunt glicerolul, 3-hidroxibutiratul, maleatul, lactatul i piruvatul,determin scderea apetitului, efect dependent de integritatea ramurii hepatice a nervuluivag. (2) Infuzia portal de lizin la pui deprim apetitul, n timp ce vagotomia local,hepatic i pancreatic, inhib aciunea lizinei. (17)
Studiile histochimice efectuate au indicat c informaiile culese de la nivelul
ficatului de ctre receptorii senzitivi sunt transmise la nivelul sistemului nervos central pecalea aferenelor vagale hepatice ajungnd la nucleul tractului solitar. (18) Ali receptoriFicatul conine i ali receptori, cum sunt cei pentru temperatur i durere. (16)
Sawchencko i Friedman au constatat c o cretere a temperaturii ficatului determinreducerea ingestiei de hran, efect care se obine i prin secionarea nervilor splahnici.(19) Lewis susine ipoteza existenei algoreceptorilor artnd c durerea resimit demajoritatea bolnavilor cu hepatopatii, n absena inflamaiei hepatice, nu se datoreazdistensiei capsulei hepatice, fiind deci posibil prezena acestor receptori n plus,Sawchencko i Friedman au afirmat c vagul i nervii splahnici ar putea fi implicai ntransmiterea la nivel central a senzaiei de durere. (19)
Nervii afereni
n componena cii aferente intr dou tipuri de fibre nervoase: vagale ispinale.Fibre nervoase vagale au originea n bulbul rahidian, n nucleul dorsal al vagului.
Ele prsesc bulbul pe calea nervului vag i ajung la nivelul ficatului, unde fac sinaps cuneuronii postganglionari, a cror localizare hepatic nu este foarte bine precizat. (20)Terminaiile nervoase vagale nu ajung la nivelul lobulilor hepatici, fiind dispuse nstraturile externe ale pereilor cilor biliare intra- i extrahepatice de calibru mare i nadventicea venei porte. (21, 22) Prin intermediul acestor fibre nervoase sunt transmise lahipotalamus informaii despre concentraia plasmatic a metaboliilor, hormonilor icitokinelor. (2327)
Nervii afereni spinali au originea n ganglionii de pe traseul rdcinilorposterioare ale mduvei toracale inferioare (T7T12). (28, 29) Prin studiiimunohistochimice s-a demonstrat c aferenele spinale au distribuie intrahepaticsimilar fibrelor vagale. (21) Se consider c fibrele nervoase spinale au rolul de atransmite informaii nociceptive de la nivel hepatic la creier (ex. inflamaii). (22)
Sistemul nervos central Centrii vegetativi
3
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
4/15
n ordinea apariiei lor filogenetice se disting centrii nervoi inferiori n trunchi imduv, centrii supraetajai subcorticali n hipotalamus i centrii vegetativi corticali.
- Centrii vegetativi inferiori se numesc i centrii preganglionari. De la ei pleac spreperiferie calea eferent. Se disting centrii parasimpatici n trunchiul cerebral i n mduvasacrat i centrii simpatici n mduva toracolombar.
- Centrii vegetativi subcorticali sunt situai n hipotalamus, cei parasimpatici lanivelul regiunii lui antero-laterale, iar cei simpatici n partea posterioar. Hipotalamusuleste principalul centru subcortical de reglare a activitii simpaticului i parasimpaticului.El primete informaii de la centrii inferiori prin ci ascendente nespecifice,multineuronale, iar influena lui se exercit prin cile descendente, ca fasciculul dorsalSchtz, i prin substana reticulat. De la hipotalamus informaiile se proiecteaz pescoara cerebral direct sau prin intermediul nucleilor rostrali talamici. Hipotalamusuleste controlat de cortexul cerebral prin ci descendente corticohipotalamice.
- Centrii vegetativi corticali. n scoara cerebral au fost identificai centriivegetativi n ariile 13, 14, 24, 25, 32, de pe feele inferioare i interne alelobilor frontali, precum i n hipocamp. Stimularea electric a ariilor 24, 25, determin o
rrire a frecvenei cardiace, efecte respiratorii, piloerecie, dilatarea pupilei,modif icr i tensionale. Stimularea ariilor 13, 14 poate opri micrile respiratorii,modific tensiunea arterial i motilitatea gastrointestinal.
Nu au fost identificai nc centrii corticali implicai n controlul funciilor hepatice.n prezent se consider c n scoara cerebral exist centri n raport cu inervaiasimpatic i parasimpatic. (30)
Hipotalamusul
Dintre centrii vegetativi subcorticali cel mai important este hipotalamusul, careprezint legturi strnse cu hipocampul prin intermediul talamusului i altrigonului cerebral. De asemenea, hipotalamusul este conectat cu scoara
cerebral prin intermediul corpilor striai. Activitatea hipotalamusului estecontrolat de scoara cerebral, iar la rndul su, hipotalamusul constituiepr incipalul centru subcortical de reglare a activitii simpatice i parasimpatice. (22)
Hipotalamusul poate fi mprit n trei poriuni: lateral, medial iperiventricular. (22) Se consider c aria hipotalamic lateral are rol n reglareaactivitii parasimpatice, deoarece stimularea acestei arii diminueaz apetitul ideclaneaz o serie de reacii anabolice, cum este creterea secreiei de insulin, careintensific lipogeneza la nivelul esutului adipos. (3133) n reglarea activitii simpaticepare a fi implicat nucleul hipotalamic ventromedial, care face parte din aria hipotalamicmedial. (3336) Stimularea acestui nucleu determin creterea apetitului i activeazunele reacii catabolice, cum sunt hiperglicemia, lipoliza la nivelul esutului adipos iintensificarea secreiei de glucagon. ntre aria hipotalamic lateral i nucleul hipotalamicventromedial exist influene funcionale reciproce. (22)
n aria hipotalamic periventricular au fost evideniai mai muli nuclei, cum suntnucleul hipotalamic periventricular i nucleul suprachiasmatic. Nucleul hipotalamicperiventricular reprezint cea mai important component a ariei hipotalamiceperiventriculare, primind majoritatea informaiilor de la alte arii i nuclei aihipotalamusului. n componena acestui nucleu intr dou tipuri de celule. Primul tip decelule controleaz activitatea sistemului nervos vegetativ, reglnd inervaia simpatic i
4
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
5/15
parasimpatic a organelor abdominale (ficat, pancreas i glande corticosuprarenale). (22,3739) Al doilea tip de celule este reprezentat de neuronii neurosecretori, care intervin nreglarea activitii hipofizei. (40) Unii dintre aceti neuroni, care controleaz activitatealobului posterior al hipofizei, sunt celule neurosecretoare magnocelulare. (41) Acetineuroni sunt localizai n nucleul supraoptic din aria hipotalamic lateral. (41) Axonii
celulelor neurosecretoare coboar pn n lobul posterior al hipofizei i secretneurohormonii oxitocin i vasopresin. n nucleul supraoptic exist celuleneurosecretorii parvocelulare, care controleaz activitatea lobului hipofizar anterior. (40)Spre deosebire de celulele neurosecretoare magnocelulare, axonii neuronilorparvocelulari nu ajung pn la hipofiz. Ei secret hormoni hipofiziotropi care ajung prinintermediul tractului hipotalamo-hipofizar n lobul anterior al hipofizei, unde stimuleazactivitatea celulelor int. Acestea secret ase hormoni: hormonul de cretere sausomatotrop (STH), hormonul adenocorticotrop (ACTH), hormonul tireotrop (TSH),hormonii gonadotropi (FSH i LH) i prolactina. Hormonii hipofizari acioneaz asupraaltor glande endocrine, care, la rndul lor, secret hormoni ce determin o serie de efecte,cum sunt modificarea volumului i compoziiei fluidelor organismului, a greutii
corporale, a funciei sexuale i a rspunsului la stress. n plus, nucleul hipotalamicperiventricular determin o serie de efecte sistemice prin stimularea sistemului nervosvegetativ i prin stimularea secreiei unor hormoni hipofizari. (22)
Nucleul suprachiasmatic este localizat n aria hipotalamic periventricularimediat deasupra chiasmei optice. Are un rol important n reglarea bioritmuluiorganismului. (4244) Nucleul suprachiasmatic primete influxuri nervoase direct de laretin i realizeaz conexiuni cu diverse arii hipotalamice, influennd ritmul circadian alsecreiei hormonale i activitatea sistemului nervos vegetativ. De asemenea, nucleulsuprachiasmatic intervine n controlul variaiilor circadiene ale valorilor glicemiei,nivelul maxim fiind fazei diurne. (45)
NeuropeptideNeuropeptidele sunt distribuite att la nivelul sistemului nervos central, ct i nnervii periferici, avnd rol de neurotransmitori i neuromodulatori.
Experimentele efectuate pe modele animale au demonstrat c o serie de centrinervoi vegetativi din nevrax pot interveni n controlul activitii ficatului, indirect, ncazul stimulrii cu unele neuropeptide. Acestea acioneaz specific asupra centrilor dinnevrax i controleaz funciile gastrointestinale prin intermediul sistemului nervosvegetativ.
Recent s-a demonstrat c ntre neuropeptidele centrale i funciile hepatice existo interrelaie. Astfel, tireotropina (TRH) acioneaz la nivel bulbar, asupra complexuluinuclear vagal dorsal stng, determinnd creterea fluxului sanguin hepatic, stimularearegenerrii hepatice i protejnd ficatul, prin mecanism vagal i ci colinergice mpotrivaleziunilor induse experimental.
Corticotropina injectat intracistenal determin inhibiia fluxului sanguin hepatici exacerbeaz leziunile hepatice, prin mecanism simpatic i ci noradrenergice.
Neuropeptidul Y acioneaz la nivelul complexului dorsal vagal stng, nparticular asupra receptorilor Y1, stimulnd secreia biliar.
Beta-endorfina i bombesina moduleaz proliferarea hepatic i secreia biliar,prin mecanism central. (46, 47)
5
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
6/15
Calea eferent
Nervii efereni
Nervii hepatici simpatici au originea n nucleul ventromedial al hipotalamusului.Fibrele nervoase au un traiect descendent spre poriunile ventrolateral i ventromedial a
formaiunii reticulare medulare, de unde ajung la neuronii din coarnele intermediolateraleale mduvei toracolombare. De la acest nivel, neuronii preganglionari i trimitprelungirile prin rdcina anterioar a nervilor spinali, trec prin lanul ganglionarparavertebral i se grupeaz formnd nervii splahnici (marele, mijlociul i micul nervsplahnic), care fac sinaps cu neuronii din ganglionii vegetativi abdominali (ex.ganglionii celiac i mezenteric superior), unde au originea fibrele nervoasepostganglionare. (21, 22, 48, 49) Fibrele nervoase simpatice postganglionare formeazmpreun cu cele parasimpatice plexurile hepatice anterior i posterior, care nsoescartera hepatic i vena port, prin intermediul crora intr n ficat. Densitatea fibrelorpostganglionare este crescut la nivel portal, fiind sczut de-a lungul sinusoidelor . (21)
Nervii hepatici parasimpatici efereni au originea n aria hipotalamic lateral. Fac
sinaps la nivelul nucleilor vag i ambiguu i cu celule din apropierea mduvei reticulare.De la nucleii dorsal al vagului i ambiguu pornesc spre ficat fibre nervoase parasimpaticepreganglionare eferente, reprezentnd ramura hepatic a nervului vag. (22, 50, 51)Fibrele parasimpatice formeaz un plex, care este separat de cel al fibrelor nervoasecatecolaminergice, (48) i intr n ficat la nivelul hilului, mpreun cu nervii simpatici.Fibrele parasimpatice fac sinaps n ganglionii intramurali i au rol n modulareaactivitii simpatice a ganglionului celiac. (1) Terminaiile fibrelor nervoase colinergicesunt localizate n jurul arterei hepatice, venei porte i ramificaiilor de calibru mare aleacesteia, precum i la nivelul cilor biliare de dimensiuni mari. (21, 22)
Inervaia intrahepatic
Utilizarea tehnicilor de imunohistochimie a permis evidenierea distribuieinervilor la nivelul ficatului. Inervaia intrahepatic difer n funcie de specie, omul fiindmamiferul cu cea mai mare densitate de fibre nervoase la nivelul lobulilor hepatici. (22)
Fibrele nervoase intrahepatice conin neurotransmitori (noradrenalin,acetilcolin) i/sau neuropeptide: substana P (SP), peptidul calcitonin gene-related(CGRP), neuropeptidul Y (NPY), peptidul intestinal vasoactiv (VIP), somatostatin(SOM), glucagon, peptidul glucagon-like (PGL), neurotensin (NT), serotonin igalanin(GAL). Unii nervi adrenergici i colinergici conin, pe lng neurotransmitorii neuropeptide. Se pare c aceti nervi au funcie senzitiv sau de reglare a funciilorhepatice, precum i a microcirculaiei la nivelul ficatului. (22)
Inervaia adrenergicMajoritatea fibrelor nervoase adrenergice au fost identificate n proximitatea vaselorspaiului port, o parte la nivelul ramurilor arterei hepatice i n numr mai redus la nivelulvenelor porte sau a cilor biliare (52, 53).
Fibrele nervoase aminergice sunt prezente n pereii venelor hepatice, dar nu i lanivelul venelor hepatice centrale sau sublobulare (54, 55).
Fibrele nervoase intralobulare urmeaz traiectul spaiilor porte pn la nivelulspaiului Dissese, fiind conectate cu celulele stelate hepatice (CSH), hepatocite,
6
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
7/15
endoteliul sinusoidal i celule Kupffer (56), distribuindu-se neuniform n structuralobulilor hepatici (52).
Datorit dispunerii fibrelor nervoase aminergice n spaiile perisinusoidale, ele potinfluena direct nu numai metabolismul celulelor parenchimatoase hepatice, dar i tonusulperetelui sinusoidal i diametrul endoteliului fenestrat (5759).
Inervaia colinergic
Nervii colinergici realizeaz un plex nervos, de la nivelul cruia fibrele nervoase sedispun adiacent sistemului vascular port, cu un traiect separat i diferit de cel al fibrelornervoase aminergice ().
Unele fibre nervoase colinergice au un traiect adiacent celulelor hepatice localizaten spaiul port, o parte din fibre terminndu-se la acest nivel, ca i proeminene Held. Prinevidenierea celulelor ganglionare acetilcolinesterazo-pozitive n spaiul port, nproximitatea hilului, a fost demonstrat natura eferent a acestor fibre nervoasecolinergice, fr a se putea exclude caracterul senzitiv al unora dintre ele (60, 61).
Inervaia colinergic, ca i cea aminergic, nu este egal distribuit n interiorul
diverilor lobuli hepatici. (48, 59, 62).n ficatul normal, acetilcolina influeneaz funciile metabolice, inclusivgluconeogeneza, prin intermediul receptorilor muscarinici M3. La nivelul celulelorepiteliului biliar, venei porte i colangiocitelor, au fost identificai receptorii M 1, M2 iM3. n celulele hepatice progenitoare, au fost identificai receptorii muscarinici M3.Acetilcolina acioneaz prin intermediul receptorilor si pe aceste celule i influeneazsecreia bilei, fluxul sangvin portal i regenerarea heaptic (63, 64).
n ficatul cirotic, dispoziia fibrelor nervoase colinergice este diferit (65, 66). Lanivelul septurilor fibroase sunt prezente numeroase fibre nervoase colinergice ale crorterminaii realizeaz conexiuni cu miofibroblatii i mastocitele. n nodulii cirotici,terminaiile nervoase realizeaz conexiuni cu miofibroblatii la nivelul sinusoidelor
periseptale. Acetilcolina stimuleaz proliferarea CSH i sinteza de colagen, intervenind napariia fibrozei hepatice (67).
Inervaia peptidergic
n ficat exist fibre nervoase ce conin neurotransmitori de tipul noradrenalinei iacetilcolinei, precum i neuropeptide. n fibrele nervoase aminergice i n celecolinergice, alturi de neurotransmitori au fost identificate neuropeptide de tipul: NPY,SP, PIV, PCGR, PGL, SOM, NT, GAL i serotonin.
Fibrele nervoase ce conin NPY, SP, CGRP i VIP sunt dispuse n special nregiunea periportal, iar cele care posed NPY, SOM i SP sunt prezente la nivelul ariilorintralobulare, unde realizeaz conexiuni cu CSH i celulele sinusoidale hepatice (66, 6870).
NPY intr n componena fibrelor nervoase aminergice ce inerveaz sistemul venosport hepatic, artera hepatic, calea biliar i ariile intralobulare, cu preponderen nregiunea periportal (66, 68, 69)
Fibrele care conin NPY sunt mai numeroase dect cele aminergice i se distribuieuniform n structura lobului hepatic (71).
NPY poteneaz vasoconstricia indus de noradrenalin la nivelul ramurilor veneiporte, putnd genera i singur un efect vasoconstrictor mai mare dect cel produs de
7
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
8/15
noradrenalin (72). NPY stimuleaz direct proliferarea CSH, urmat de fibroz hepatic(67).
Fibrele nervoase ce conin SP sunt dispuse n numr mare n regiunile periportal iintralobular, terminaiilor lor realiznd conexiuni cu CSH (66; 7274).
SP face parte din familia tahikinelor i se afl preponderent n straturile superficiale
ale cordoanelor medulare dorsale i are rolul de a transmite informaii nociceptive,contribuind la apariia strilor dureroase patologice asociate inflamaiei i lezrii nervoase(75). Fibrele nervoase ce conin SP de la nivel hepatic reprezint nervi senzitivi primaricu originea n ganglionul rdcinii dorsale.
Fibrele nervoase care posed SOM se distribuie n jurul venei porte i a artereihepatice n regiunea portal i n regiunea perisinusoidal a ficatului. Terminaiile acestorfibre nervoase realizeaz conexiuni cu celulele sinusoidale (69, 76). n cazul pacienilorcu ciroz hepatic etanolic, numrul acestor fibre este mai redus (69).
SOM ndeplinete mai multe roluri fiziologice printre care controlulneurotransmisiei, a secreiei de proteine, a contractilitii celulelor musculare netede, amotilitii intestinale i a funciei celulelor sistemului imun. SOM acioneaz pe cinci
tipuri de receptori. La nivel hepatic, colangiocitele conin receptorii 1, 2, 3 i 4 pentrusomatostatin, iar CSH activate conin receptorii 1, 2, i 3 (77, 78).La nivelul colangiocitelor, SOM inhib att secreia de bil, ct i proliferarea
acestora (77).Prin intermediul receptorului 1 pentru SOM este inhibat contracia CSH activate
indus de ET-1 (78). SOM influeneaz secreia bilei i intervine direct asupra fluxuluisangvin sinusoidal prin controlul microcirculaiei hepatice.
Fibrele nervoase ce conin CGRP sunt distribuite n zona periportal, dar nu i ncea intralobular, fiind asociate preponderent cilor biliare i mai puin arterei hepatice ivenei porte (69). Neurotransmitorul CGRP este eliberat de fibrele nervoase primareaferente nemielinizate, intervenind n nocicepie (80).
Fibrele nervoase ce conin VIP sunt dispuse n zona periportal, n jurul venei portei a arterei hepatice. VIP este prezent n special n jurul cilor biliare, unde intervine nreglarea secreiei biliare (66).
Fibrele nervoase imunoreactive la CGRP i VIP coexist cu fibrele nervoasecolinergice i senzitive. n numr redus, acestea intr n componena nervilor autonomi ceinerveaz vena port i artera hepatic cu ramurilor lor, dar nu i alte segmente vascularesau cile biliare (80, 81).
Inervaia nitrergicOxidul nitric (NO) este un neuroefector care se formeaz prin catalizarea sintezei
neuronale a oxidului nitric (nNOS). Fibrele nervoase intrahepatice imunoreactive lanNOS formeaz un plex nervos dens n jurul ramurilor mari ale venei porte, a arterelorhepatice i a cilor biliare, reducndu-se numeric la nivelul ramurilor distale ale acestorstructuri (82).
8
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
9/15
Bibliografie
1. Berthoud HR. Anatomy and Function of Sensory Hepatic Nerves. The AnatomicalRecord Part A. 2004; 280A:827835.
2. Langhans W. Role of the liver in the metabolic control of eating: what we know
and what we do not know. Neurosci Biobehav Rev 1996; 20:145153.3. Friedman MI. An energy sensor for control of energy intake. Proc Nutr Soc 1997;56:4150.
4. Goehler LE, Gaykema RP, Hansen MK, Anderson K, Maier SF, Watkins LR. Vagalimmune-to-brain communication: a visceral chemosensory pathway. Auton Neurosci2000; 85:4959.
5. Tracey KJ. The inflammatory reflex. Nature 2002; 420:853859.6. Obici S, Rossetti L. Minireview: nutrient sensing and the regulation of insulin action
and energy balance. Endocrinology 2003; 144: 51725178.7. Lautt, WW. Canadian Journul of Physiology and Pharmacology 1980; 58, 105-123.8. Morita H, Matsuda T, Tanaka K, Hosomi H. Role of Hepatic Receptors in
Controlling Body Fluid Homeostasis. Japanese Journal of Physiology 1995; 45: 355-368.9. Niijima A. Afferent discharges from osmoreceptors in the liver of the guinea pig.
Science 1967; 166: 1519-1520.10. Adachi A, Niijima A, Jacobs HL. An hepatic osmoreceptors mechanism in the rat:
Electrophysiological and behavioral studies. Am J Physiol 1976; 231: 1043-1049.11. Andrews WHH, Orbach J: Effect of osmotic pressure on spontaneus afferent
discharge in the nerves of the perfused rabitt liver. Pflgers Arch 1975; 361: 89-94.12. Passo SS, Thornborugh JR, Rothballer AB. Hepatic receptors in control of sodium
excretion in anesthetized cats. Am J Physiol 1973; 224: 373-375.13. Andrews WHH, Orbach J. Sodium receptors activating some nerves of perfused
rabbit livers. Am J Physiol 1974; 227: 1273-127514. Andrews WHH, Palmer JF. Afferent nervous discharge from canine liver. QJ ExpPhysiol 1967; 52: 269-276.
15. Niijima A. Afferent disharges from venous pressoreceptors in liver. Am J Physiol1977; 232: C76-C81.
16. Anil M.H, Forb J.M. Neural control and neurosensory functions of the liver.Proceedings ofthe Nutrition Society 1987; 46: 125-133.
17. Rusby, AA. The role of the liver in the control of food intake in the domesticchicken. PhD Thesis, University of Leeds 1985.
18. Norgren, R. Afferent interactions of cranial nerves involved in ingestion. Journalofthe Autonomic Nervous System 1983; 9: 67-77.
19. Sawchenko, PE, Friedman, MI. American Journal OfPhysiology 1979; 236, Rg-Rzo.20. Ganong WF. Review of Medical Physiology. California: Lange, Los Altos, 2001.21. Pschel G.P. Modulation of liver function by hepatic nerves. In Textbook of
Hepatology. From Basic Science to Clinical Practice. Third Edition. Edited by JuanRods, Jean-Pierre Benhamou, Andres Blel, Jurg Reichen, Mario Rizzetto. BlackwellPublishing, 2007, 114-122.
22. Uyama N, Geerts A, Reynaert H. Neural Connections Between the Hypothalamusand the Liver. The Anatomical Record Part A 2004; 280A:808820.
9
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
10/15
23. Sakaguchi T, Iwanaga M. Effects of D-glucose anomers on afferent discharge inthehepatic vagus nerve. Experientia 1982; 38:475476.
24. Niijima A. The afferent discharges from sensors for interleukin 1 beta in thehepatoportal system in the anesthetized rat. J Auton Nerv Syst 1996; 61:287291.
25. Torii K, Niijima A. Effect of lysine on afferent activity of the hepatic branch of the
vagus nerve in normal and L-lysine-deficient rats. Physiol Behav 2001; 72:685690.26. Randich A, Spraggins DS, Cox JE, Meller ST, Kelm GR. Jejunal or portal veininfusions of lipids increase hepatic vagal afferent activity. Neuroreport 2001;12:31013105.
27. Niijima A, Miyata G, Sato T, Meguid MM. Hepato-vagal pathway associated withnicotines anorectic effect in the rat. Auton Neurosci 2001; 93:4855.
28. Magni F, Carobi C. The afferent and preganglionic parasympathetic innervation ofthe rat liver, demonstrated by the retrograde transport of horseradish peroxidase. JAuton Nerv Syst 1983; 8:237260.
29. Barja F, Mathison R. Sensory innervation of the rat portal vein and the hepaticartery. J Auton Nerv Syst 1984; 10:117125.
30. Badiu G, Papari A. Controlul nervos al funciilor organismului Neurofiziologie.Editura Fundaiei Andrei aguna, Constana, 1995.31. de Jong A, Strubbe JH, Steffens AB. Hypothalamic influence on insulin and
glucagon release in the rat. Am J Physiol 1977; 233:E380E388.32. Gutstein WH, Parl F. 1978. Neural factors in experimental degenerative arteriopathy.
Lipids 13:380382. Milam KM, Stern JS, Storlien LH, Keesey RE. Effect of lateralhypothalamic lesions on regulation of body weight and adiposity in rats. Am JPhysiol 1980; 239:R337R343.
33. Yoshimatsu H, Niijima A, Oomura Y, Yamabe K, Katafuchi T. Effects ofhypothalamic lesion on pancreatic autonomic nerve activity in the rat. Brain Res1984; 303:147152.
34. Inoue S, Campfield LA, Bray GA. Comparison of metabolic alterations inhypothalamic and high fat diet-induced obesity. Am J Physiol 1977; 233:R162R168.
35. Niijima A, Rohner-Jeanrenaud F, Jeanrenaud B. Role of ventromedial hypothalamuson sympathetic efferents of brown adipose tissue. Am J Physiol 1984; 247:R650R654.
36. Saito M, Minokoshi Y, Shimazu T. Accelerated norepinephrine turnover inperipheral tissues after ventromedial hypothalamic stimulation in rats. Brain Res1989; 481:298303.
37. Kannan H, Niijima A, Yamashita H. Inhibition of renal sympathetic nerve activity byelectrical stimulation of the hypothalamic paraventricular nucleus in anesthetizedrats. J Auton Nerv Syst 1987; 21:8386.
38. Kannan H, Hayashida Y, Yamashita H. Increase in sympathetic outflow byparaventricular nucleus stimulation in awake rats. Am J Physiol 1989; 256:R1325R1330.
39. van Dijk G, Bottone AE, Strubbe JH, Steffens AB. Hormonal and metabolic effectsof paraventricular hypothalamic administration of neuropeptide Y during rest andfeeding. Brain Res 1994; 660:96103.
10
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
11/15
40. Freund-Mercier MJ, Stoeckel ME, Moos F, Porte A, Richard P.Ultrastructural studyof electrophysiologically identified neurones in the paraventricular nucleus of the rat.Cell Tissue Res 1981; 216:503512.
41. Sokol HW, Zimmerman EA, Sawyer WH, Robinson AG. The hypothalamic-neurohypophysial system of the rat: localization and quantitation of neurophysin by
light microscopic immunocytochemistry in normal rats and in Brattleboro ratsdeficient in vasopressin and a neurophysin. Endocrinology 1976; 98:11761188.42. Moore RY, Eichler VB. Loss of a circadian adrenal corticosterone rhythm following
suprachiasmatic lesions in the rat. Brain Res 1972; 42:201206.43. Stephan FK, Zucker I. Circadian rhythms in drinking behavior and locomotor
activity of rats are eliminated by hypothalamic lesions. Proc Natl Acad Sci USA1972; 69:15831586.
44. Karatsoreos IN, Yan L, LeSauter J, Silver R. Phenotype matters: identification oflight-responsive cells in the mouse suprachiasmatic nucleus. J Neurosci 2004; 24:6875.
45. La Fleur SE, Kalsbeek A, Wortel J, Buijs RM. A suprachiasmatic nucleus generated
rhythm in basal glucose concentrations. J Neuroendocrinol 1999; 11:643652.46. Yoneda M, Shimada T, Terano A, Hiraishi H. Regulation of hepatic function bystress-related neuropeptides in the brain. International Congress Series 2006; 1287355 360.
47. Yoneda M, Watanobe H, Terano A. Central regulation of hepatic function byneuropeptides. J Gastroenterol 2001; 36: 361-367.
48. McCuskey RS. Anatomy of efferent hepatic nerves. Anat Rec A Discov Mol CellEvol Biol 2004; 280, 821826.
49. Tache S. Inervaia ficatului i controlul nervos al funciilor hepatice. n Tratat deHepatologie, sub redacia Mircea Grigorescu, Editura Medical Naional, 2004: 86-89.
50. Shimazu T. Innervation of the liver and glucoregulation: roles of the hypothalamusand autonomic nerves. Nutrition 1996; 12:6566.51. Shimazu T. Neuronal regulation of hepatic glucose metabolism in mammals.
Diabetes Metab Rev 1987; 3:185206.52. Goehler LE, Sternini C. Neuropeptide Y immunoreactivity in the mammalian liver:
pattern of innervation and coexistence with tyrosine hydoxylase immunoreactivity.Cell Tissue Res 1991; 265:287295.
53. Lin YS, Nosaka S, Amakata Y, Maeda T. Comparative study of the mammalian liverinnervation: an immunohistochemical study of protein gene product 9.5, dopaminebeta-hydroxylase and tyrosine hydroxylase. Comp Biochem Physiol Part A Physiol1995; 110:289298.
54. Fuller RW, Felten SY, Perry KW, Snoddy HD, Felten DL. Sympatheticnoradrenergic innervation of guinea-pig liver: histofluorescence and pharmacologicalstudies. J Pharmacol Exp Ther 1981; 218:282288.;
55. Burt AD, Tiniakos D, MacSween RNM, Griffiths MR, Wisse E, Polak JM.Localization of adrenergic and neuropeptide tyrosinecontaining nerves in themammalian liver. Hepatology 1989; 9:839845.
56. Scoazec J-Y, Racine L, Couvelard A, Moreau A, Flejou J-F, Bernuau D, FeldmannG. Parenchymal innervation of normal and cirrhotic human liver: a light and electron
11
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
12/15
microscope study using monoclonal antibodies against the neural cell-adhesionmolecule. J Histochem Cytochem 1993; 41:899908.
57. Oda M, Azuma T, Watanabe N, Nishizaki Y, Nishida J, Ishii K, Suzki H, Kaneko H,Komatsu H, Tsukada N, Tsuchiya M. Regulatory mechanism of hepaticmicrocirculation: involvement of contraction and dilatation of sinusoids and
sinusoidal endothelial fenestrae. In: Hammersen MK, editor. Progress in appliedmicrocirculation. 1990: 103128.58. Metz W, Forssmann WG. Comparative morphology of liver innervation. In: Popper
H, Bianchi L, Gudat F, Reutter W, editors. Communications of liver cells. Basel:MTP Press. 1980 p 121127.
59. Amenta F, Cavallotti C, Ferrante F, Tonelli F. Cholinergic nerves in the human liver.Histochem J 1981; 13:419424.
60. Xue C, Aspelund G, Sritharan KC, Wang JP, Slezak LA, Andersen DK. Isolatedhepatic cholinergic deinnervation impairs glucose and glycogen metabolism. J SurgRes 2000; 90:1925.
61. Schafer MK, Eiden LE, Weihe E. Cholinergic neurons and terminal feilds revealed
by immunohistochemistry for the vesicular acetylcholine transporter: II, theperipheral nervous system. Neuroscience 1998; 84:361376.62. McCuskey R.S. Distribution of intrahepatic nerves: an overview. In Liver
Innervation and the neural control of hepatic function edited by Takashi Shimazu,John Libbey & Company Ltd., 1996, 17-22.
63. Vatamaniuk MZ, Horyn OV, Vatamaniuk OK, Doliba NM. Acetylcholine affects ratliver metabolism via type 3 muscarinic receptors in hepatocytes. Life Sci 2003;72:18711882.
64. Cassiman D, Libbrecht L, Sinelli N, Desmet V, Denef C, Roskams T. The vagalnerve stimulates activation of the hepatic progenitor cell compartment via muscarinicacetylcholine receptor type 3. Am J Pathol 2002; 161:521530.
65. Akiyoshi H, Terada T. Mast cell, myofibroblast and nerve terminal complexes incarbon tetrachloride-induced cirrhotic rat livers. J Hepatol 1998; 29:112119.;66. Akiyoshi H, Gonda T, Terada T. A comparative histochemical and
immunohistochemical study of aminergic, cholinergic and peptidergic innervation inrat, hamster, guinea pig, dog and human livers. Liver 1998; 18:352359.
67. Oben JA, Yang S, Lin H, Ono M, Diehl AM. Acetylcholine promotes theproliferation and collagen gene expression of myofibroblastic hepatic stellate cells.Biochem Biophys Res Commun 2003; 300:172177.
68. Ding WG, Kitasato H, Kimura H. Development of neuropeptide Y innervation in theliver. Microsc Res Tech 1997; 39:365371.;
69. Stoyanova II, Gulubova MV. Peptidergic nerve fibres in the human liver. ActaHistochem 1998; 100:245256.;
70. Ueno T, Tanikawa K. Intralobular innervation and lipocyte contractility in the liver.Nutrition 1997; 13:141148.
71. Feher E, Fodor M, Gorcs T, Feher J, Vallent K.. Immunohistochemical distributionof neuropeptide Y and catecholamine-synthesizing enzymes in nerve fibers of thehuman liver. Digestion 1991; 50: 194201.
12
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
13/15
72. Pernow J, Saria A, Lundberg JM. Mechanisms underlying preand postjunctionaleffects of neuropeptide Y in sympathetic vascular control. Acta Physiol Scand 1986;126:239249.
73. Tanikawa K. 1995. Hepatic sinusoidal cells and sinusoidal circulation. JGastroenterol Hepatol 10(Suppl 1):S8S11.;
74. Feher E. 1998. Peptidergic innervation of the liver. In: Haussinger D, Jungermann K,editors. Liver and nervous system. Lancaster, U.K.: Kluwer Academic Publisher. p3951.
75. Hokfelt T, Meyerson B, Nilsson G, Pernow B, Sachs C. Immunohistochemicalevidence for substance P-containing nerve endings in the human cortex. Brain Res1976; 104:181186.
76. Feher E, Fodor M, Feher J. Ultrastructural localization of somatostatin- andsubstance P-immunoreactive nerve fibers in the feline liver. Gastroenterology 1992;102:287294.
77. Gong AY, Tietz PS, Muff MA, Splinter PL, Huebert RC, Strowski MZ, Chen XM,LaRusso NF. Somatostatin stimulates ductal bile absorption and inhibits ductal bile
secretion in mice via SSTR2 on cholangiocytes. Am J Physiol Cell Physiol 2003;284:C1205C1214.;78. Reynaert H, Vaeyens F, Qin H, Hellemans K, Chatterjee N, Winand D, Quartier E,
Schuit F, Urbain D, Kumar U, Patel YC, Geerts A. Somatostatin suppressesendothelin-1-induced rat hepatic stellate cell contraction via somatostatin receptorsubtype 1. Gastroenterology 2001; 121:915930.
79. Goehler LE, Sternini C, Brecha NC. Calcitonin gene-related peptideimmunoreactivity in the biliary pathway and liver of the guinea-pig: distribution andcolocalization with substance P. Cell Tissue Res 1988; 253:145150.
80. Berthoud HR, Kressel M, Neuhuber WL. An anterograde tracing study of the vagalinnervation of rat liver, portal vein and biliary system. Anat Embryol 1992; 186:431
442.;81. Goehler LE, Sternini C. Calcitonin gene-related peptide innervation of the rathepatobiliary system. Peptides 1996; 17:209217.
82. Peinado MA, delMoral ML, Jimenez A, Rodrigo J, Esteban FJ. The nitrergicautonomic innervation of the liver. Auton Neurosci 2002; 99:6769.
13
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
14/15
Fig.1. Conexiunile nervoase dintre hipotalamus i ficat.NHVM = nucleu hipotalamic ventral medial. AHL = arie hipotalamic lateral. NHP =nucleu hipotalamic posterior. NSC = nucleu suprachiasmatic. NA = nucleu arcuat. IML =coloan celular intermediolateral. GD = ganglion dorsal. VP = ven port. AH = arterhepatic. CBP = cale biliar principal.
14
-
8/22/2019 2.1_Caile nervoase de reglare a func+iilor ficatului
15/15
Fig.2. Rolul neuropeptidelor n reglarea funciilor ficatului.TRH = hormon eliberator de tireotropin. CRF = factor eliberator de
cotricotropin. NPY = neuropeptid Y.
15