
VIRUSOLOGIE.pdf
88
1 Intrarea virusului in celula gazda si eliberarea materialului genetic viral Serban MOROSAN, DVM, PhD
-
Upload
oana-bunea -
Category
Documents
-
view
146 -
download
5
Transcript of VIRUSOLOGIE.pdf

1
Intrarea virusului in celula gazda si eliberarea materialului
genetic viral
Serban MOROSAN, DVM, PhD

2
• Particulele virale au o singura misiune: transportul genomului viral de la o celula infectata la o celula neinfectata
• Transportul genomului viral in citoplasma sau in
nucleul unei celule neinfectate

Etapele ciclului de multiplicare
• Atasarea • Penetrarea • Decapsidarea • Multiplicarea intracelulara
– Transcriptie – Traducere – Replicare
• Maturarea • Iesirea
3

• Celula tinta poate fi o celula vecina celei infectate sau o celula dintr-un alt organism (transmiterea infectiei)
• Procesul de infectie parcurge mai multe etape:
1. Intrarea virusului in celula 2. Impachetarea genomului viral si a proteineleor
accesorii (la nivelul celulei infectate) intr-o noua particula virala
3. Eliberarea particulei virale in spatiul extracelular 4. Contactul cu alte celule: atasarea la receptori,
endocitoza, penetrarea membranei, transportul citosolic, import nuclear etc.
5. Eliberarea genomului viral (strategia « calului Troian »)
4
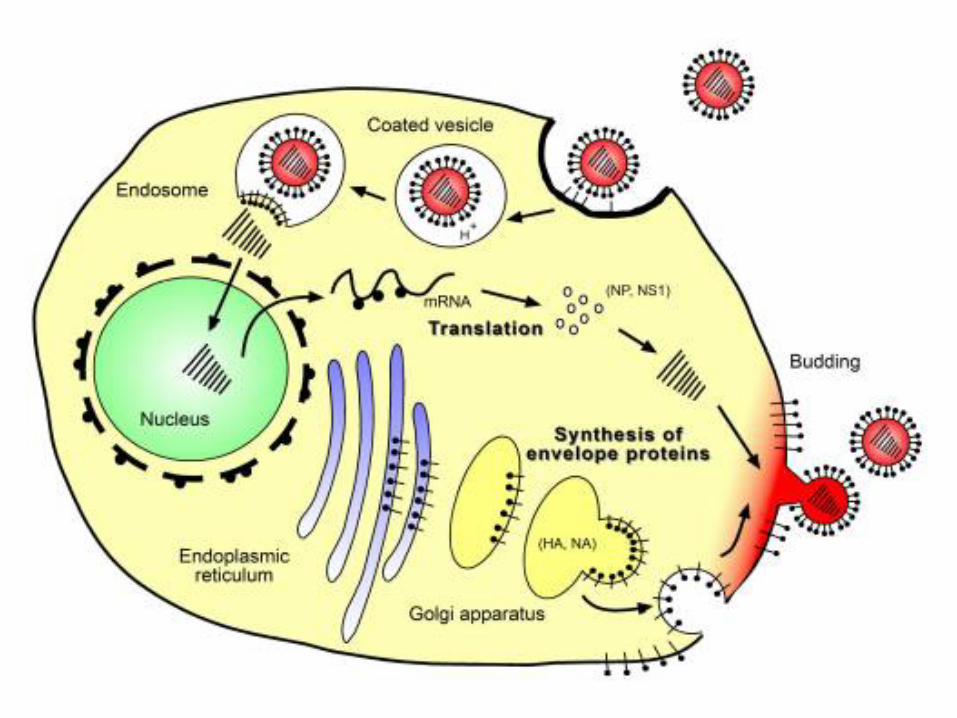
5
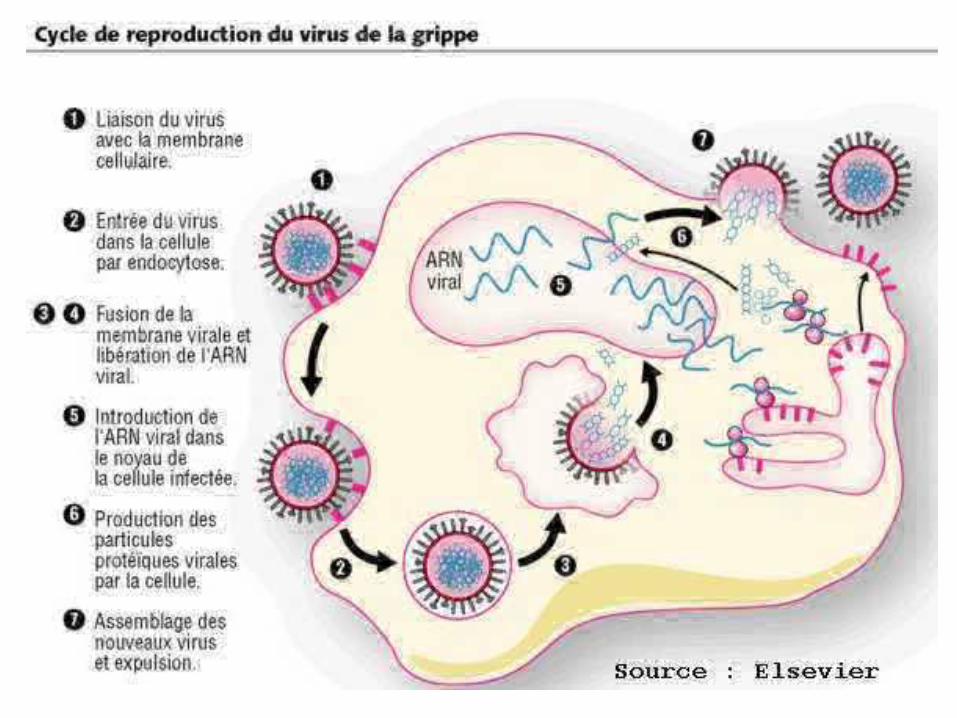
6
7
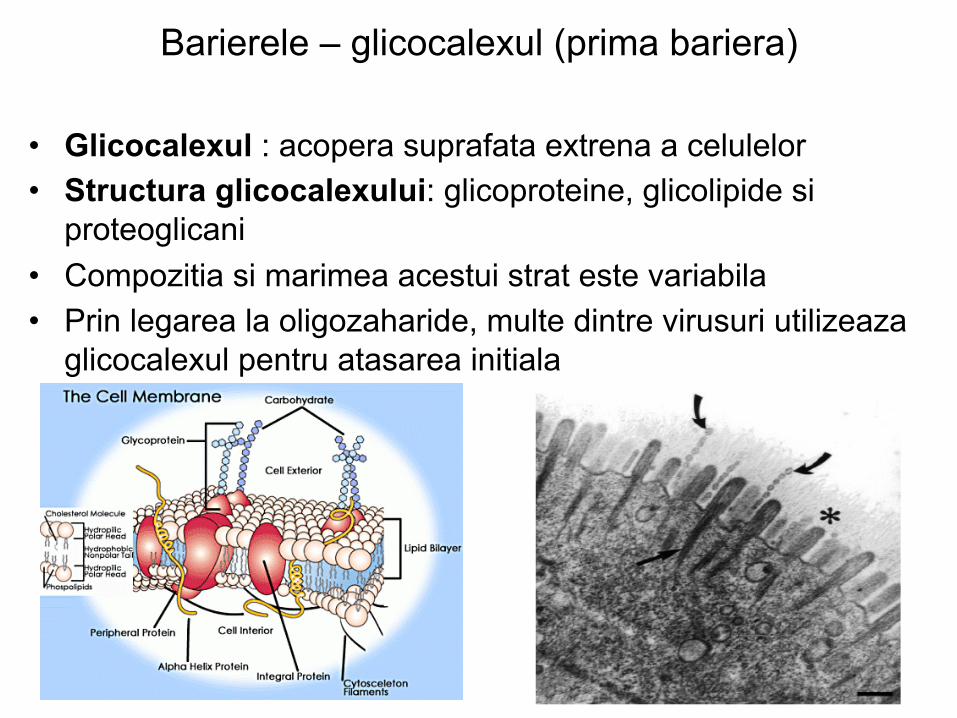
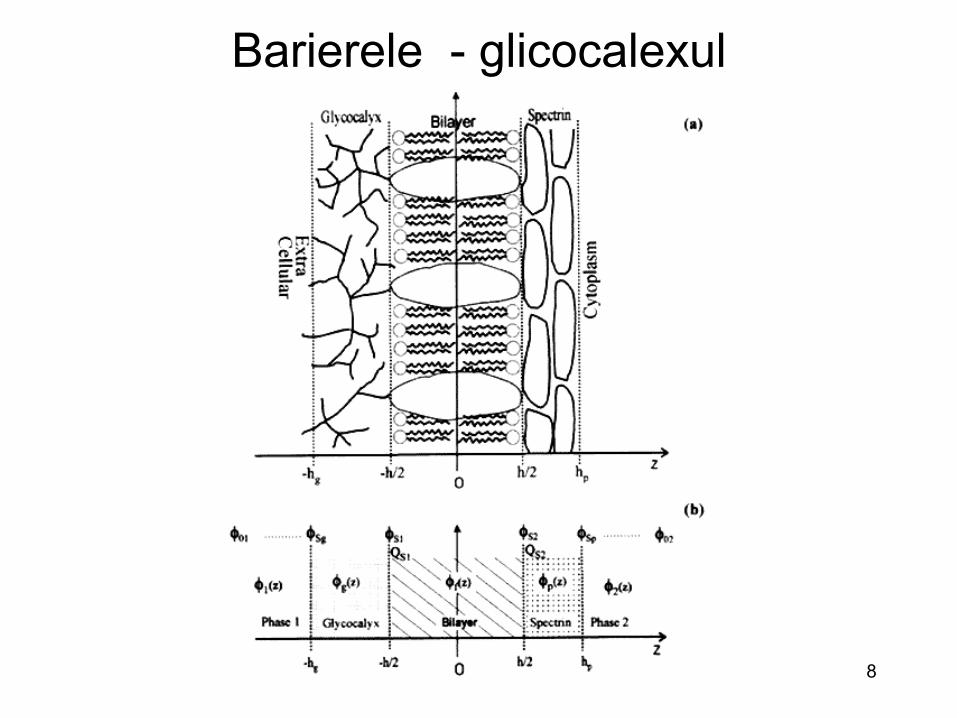
Barierele – glicocalexul (prima bariera)
• Glicocalexul : acopera suprafata extrena a celulelor • Structura glicocalexului: glicoproteine, glicolipide si
proteoglicani • Compozitia si marimea acestui strat este variabila • Prin legarea la oligozaharide, multe dintre virusuri utilizeaza
glicocalexul pentru atasarea initiala
8
Barierele - glicocalexul

9
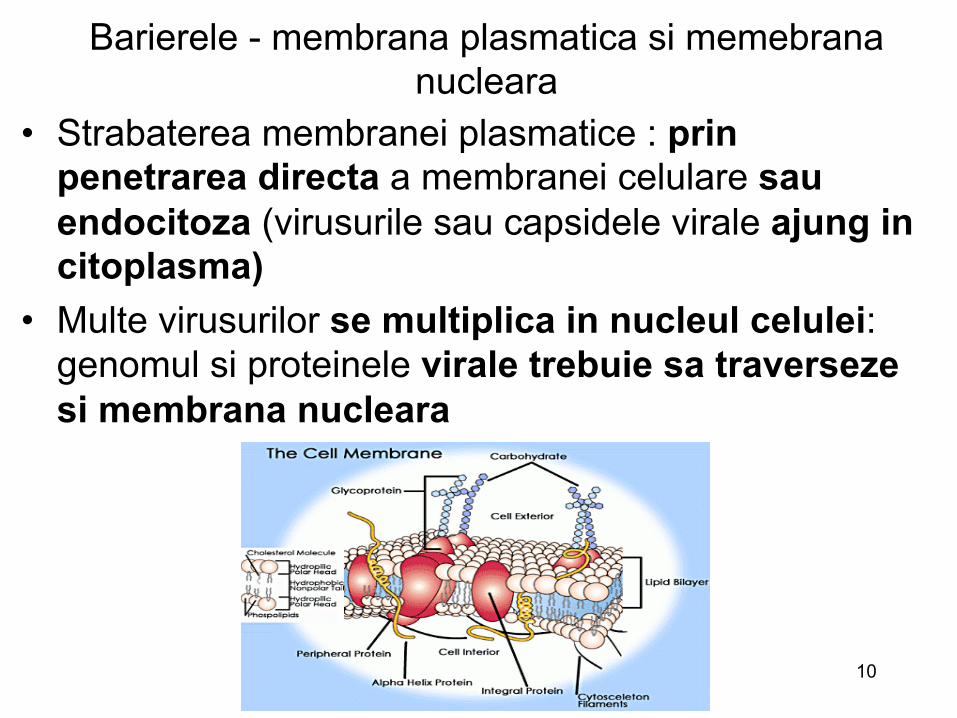
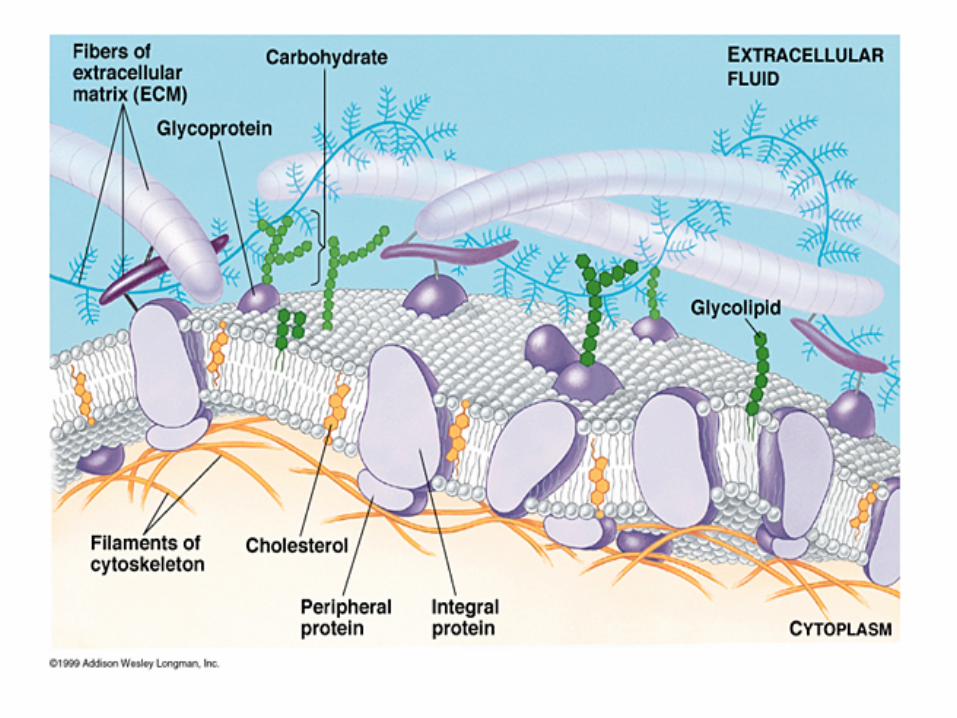
Barierele - membrana plasmatica (a doua bariera)
• Este responsabila de schimburile dintre celule si mediul extern
• Este partea cea mai complexa si mai dinamica din cadrul structurii unei celule
• Compozitia si proprietatile membranei plasmatice sunt reglate prin endocitoza si caile secretorii: prin asocieri si disocieri proteice continue
• Membrana plasmatica este un organ senzitiv : in recunoasterea si raspunsul la stimuli externi
• Virusurile utilizeaza aceste proprietati in timpul patrunderii in celule gazda
10
Barierele - membrana plasmatica si memebrana nucleara
• Strabaterea membranei plasmatice : prin penetrarea directa a membranei celulare sau endocitoza (virusurile sau capsidele virale ajung in citoplasma)
• Multe virusurilor se multiplica in nucleul celulei: genomul si proteinele virale trebuie sa traverseze si membrana nucleara
11
12

13
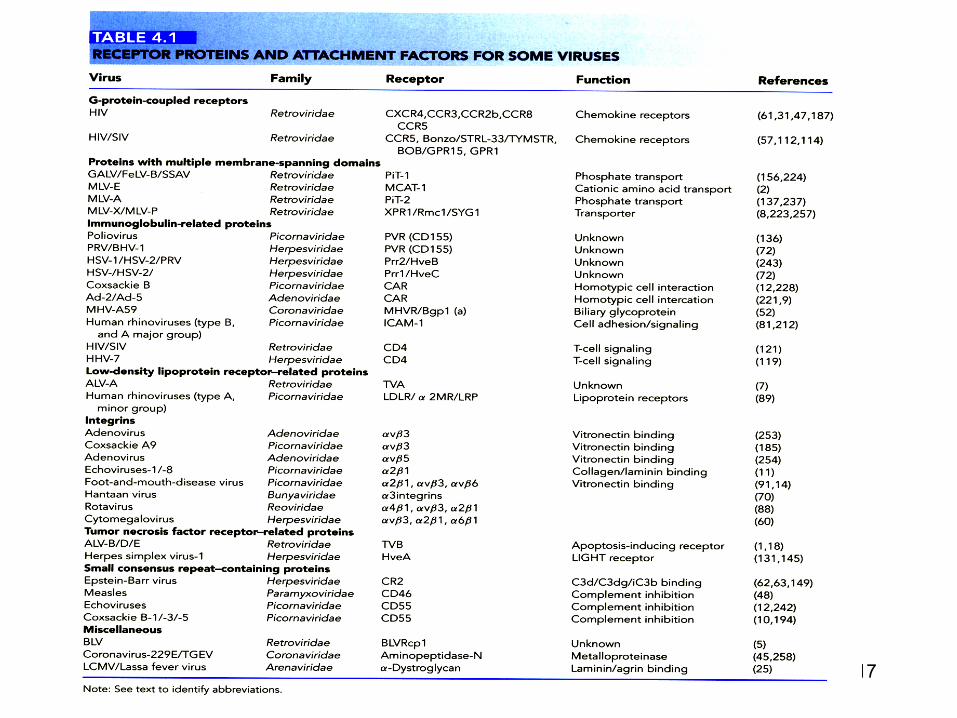
Atasarea virusului la suprafata celulara
• Virusurile infecteaza numai celulele la care se pot atasa
• Atasarea: prin interactiuni diverse dintre proteinele virale de suprafata si factori de atasare sau receptorii virali
• Identitatea si distributia acestor componente celulare determina celulele ce vor fi sau nu infectate
• Specificitatea si exprimarea receptorilor pot defini potentialul patogenic al unui virus dar si natura bolii pe care o cauzeaza

14
Atasarea virusului la suprafata celulara • Receptorii virusurilor : moleculele celulare de
suprafata ce ataseaza virusurile la o celula gazda si care favorizeaza intrarea virsului prin:
a) Modificari conformationale ale virusului ceea
ce determina pregatirea, asocierea cu alti receptori , fuziunea membranara si penetrarea
b) Transmiterea de semnale (de-a lungul membranei plasmatice) ce conduc la penetrarea virusului si la pregatirea celulei la invazia particulelor virale
c) Ghidarea particulelor virale catre diferitele cai de endocitoza

15
Atasarea virusului la suprafata celulara
• Factorii de atasare ajuta la concentrarea particulelor la suprafata celulara: favorizand intrarea si infectia
• In practica, distinctia dintre receptori si factori de atasare nu este simpla
• Receptorii si factorii de atasare : colectie de proteine, carbohidrati si lipide sau integrine
• Multe dintre virusurile cunoscute utilizeaza factori multiplii si receptori diferiti
• Virusurile interactioneaza cu acestia in paralel sau in serie
• Alte virusuri utilizeaza receptori diferiti pentru a infecta diferite celule

16
Atasarea virusului la suprafata celulara
• Factorii de atasare: – Prezenti la suprafata celulei – Permit « concentrarea » particulelor virale – Permit apropierea receptorilor celulari specifici – Exemple : carbohidrati – Membrii ai familiei lectinelor de tip C – Heparin-sulfati
17
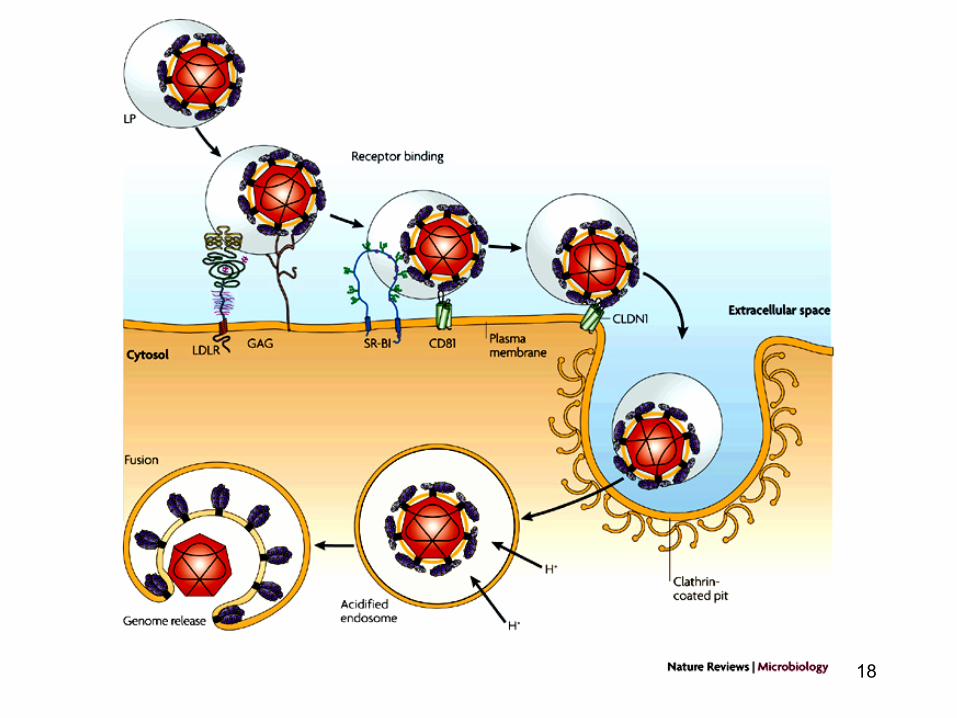
18

19
Atasarea virusului la suprafata celulara
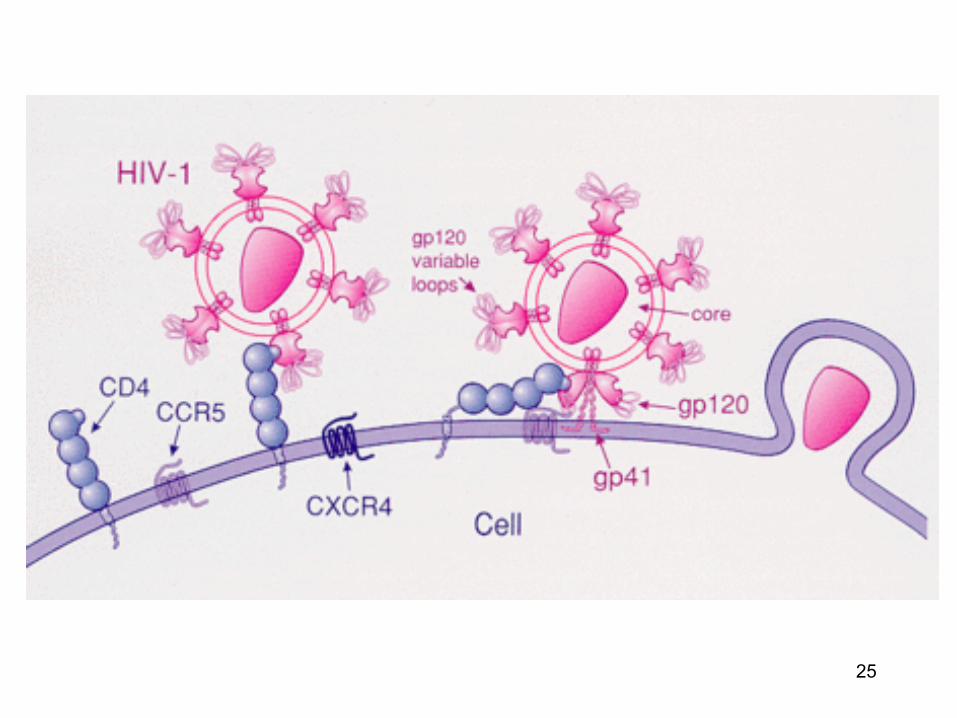
• Unele virusuri (pentru a patrunde intr-o celula gazda) necesita mai multi receptori: un receptor principal si unii coreceptori: de ex. HIV-1 (heparin sulfat proteoglicani - factori de atasare: CD4 - receptor; CXCR4, CCR5 - coreceptori)
• Aviditatea unui virus de atasare la o celula se datoreaza
prezentei a numeroase zone de atasare (la suprafata virusurilor) si mobilitatii laterale ale receptorilor moleculelor
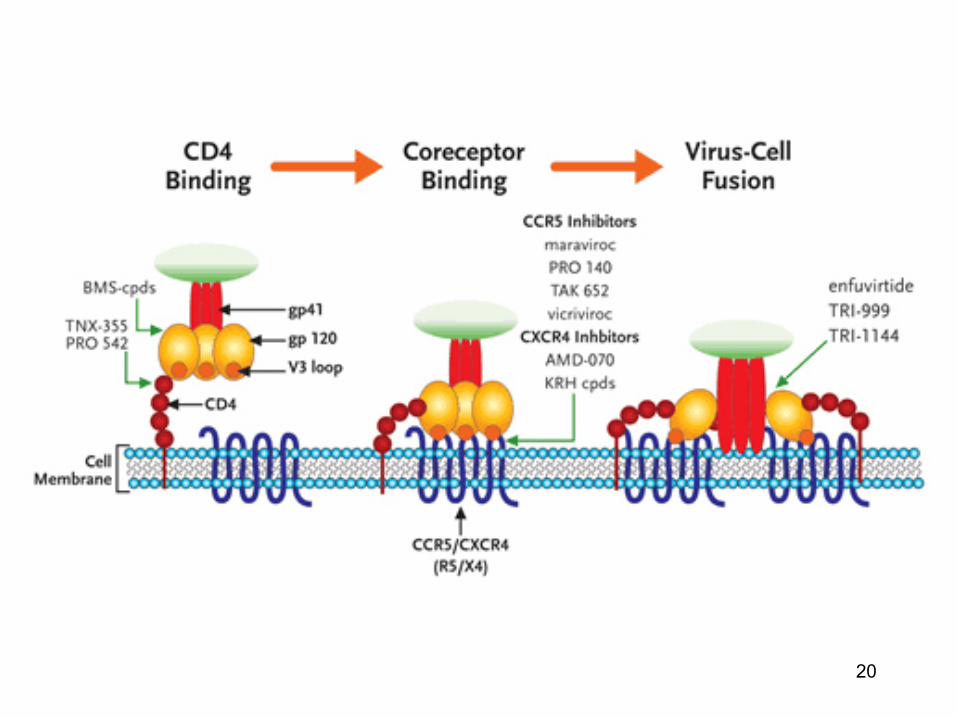
20
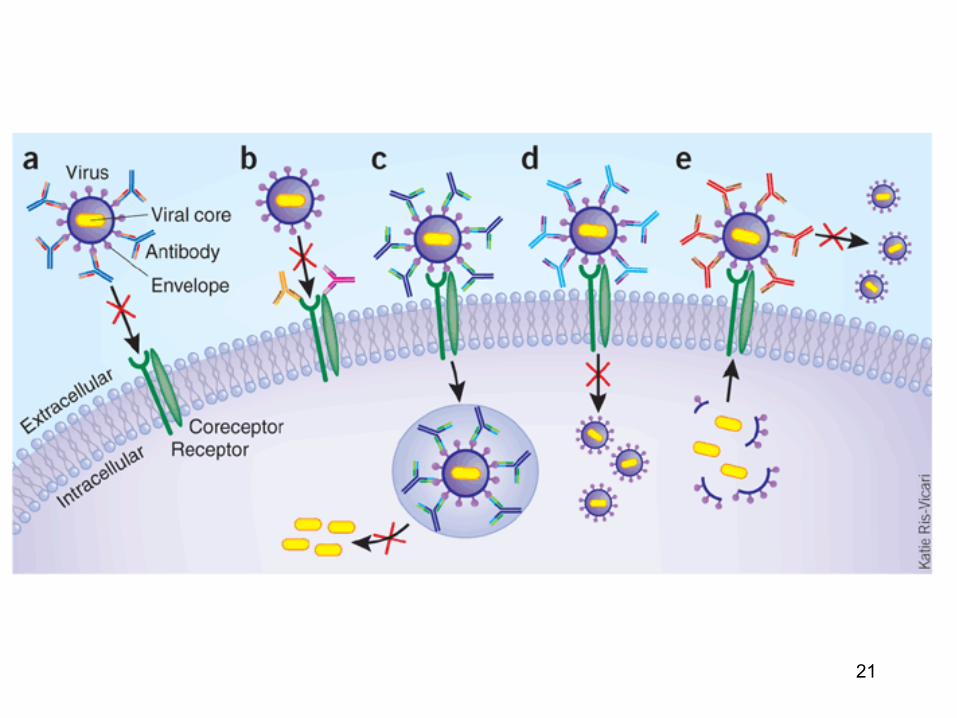
21

22
Atasarea virusului la suprafata celulara
• Receptorilor celulari sunt mobili: ataseaza virusurile • Consecintele prezentei acestor receptori: incluzia
virusurilor la nivelul straturilor lipidice sau la nivelul altor structuri specializate ale membranei (caveole etc.)
• In general interactiunea receptori-virus este directa:
uneori este necesara prezenta unei proteine de adaptare (infectia cu virusul dengue a macrofagelor ce prezinta receptorii Fc: ) – In acest caz particulele virale vor fi internalizate ca un complex imun

23
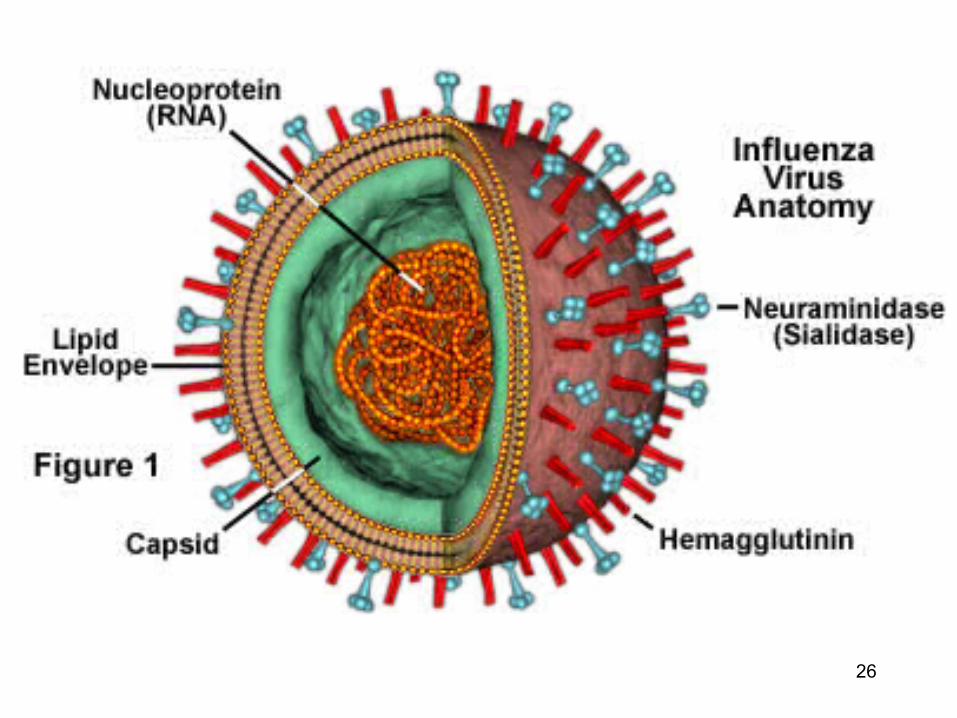
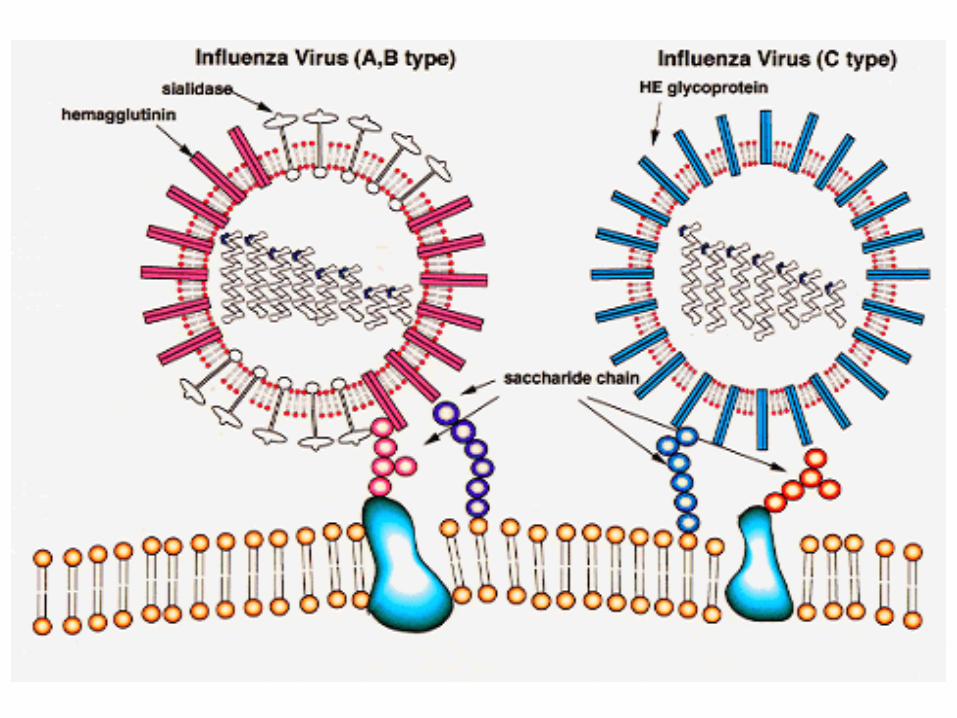
Proteinele virale responsabile de atasarea la celulele gazda (virusurile cu anvelopa)
• Virusurile cu anvelopa : prelungirile glicoproteice sunt responsabile de atasarea virusului la celula gazda (proteine de fuziune)
• Aceste proteine membranare sunt externalizate
la suprafata membranei virusului (un « receptor-binding domain » bine expus)
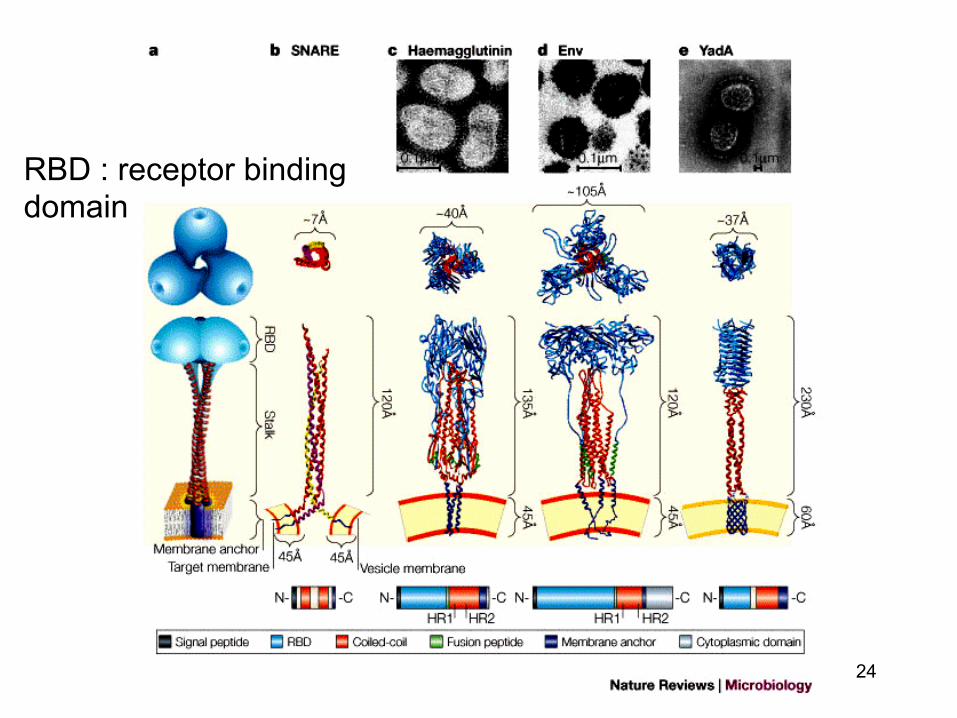
24
RBD : receptor binding domain
25
26
27

28
Proteinele virale responsabile de atasarea la celulele gazda (virusurile nude)
• La virusurile nude structurile ce se ataseaza la receptorii celulari sunt « prelungiri » de la nivelul capsidei virale (proteine de atasare)
• Locul de atasare a receptorului poate implica o singura proteina sau o suprafata compusa din mai multe lanturi polipeptidice
• Exemplu : « penton base protein » ale multor adenovirusuri contin secvente expuse (Arg-Gly-Asp) ce se asociaza cu integrine (receptori)
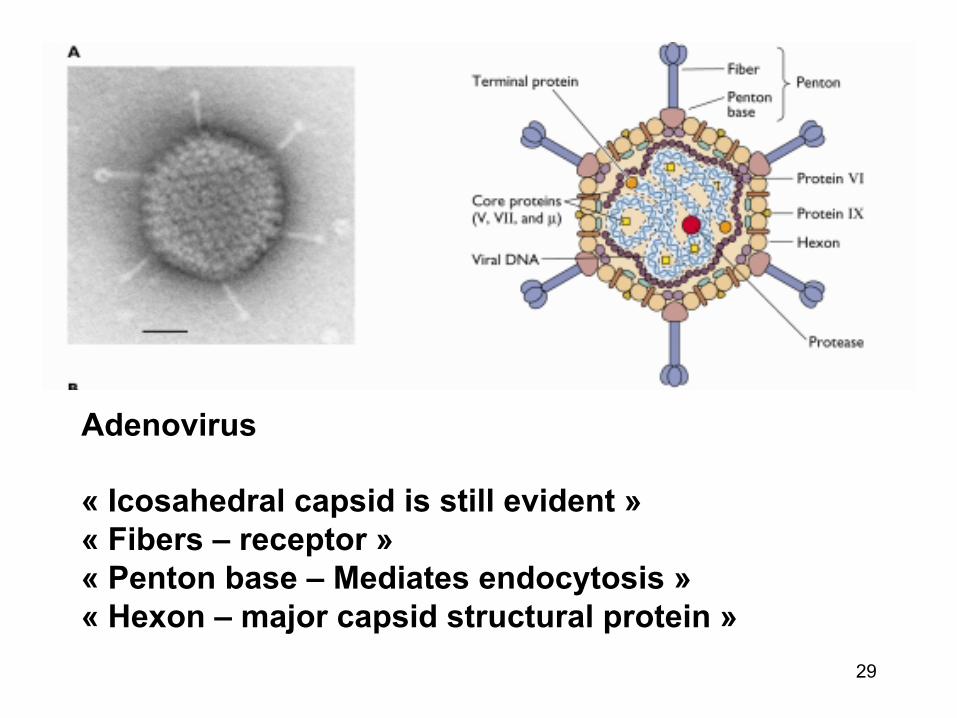
29
Adenovirus « Icosahedral capsid is still evident » « Fibers – receptor » « Penton base – Mediates endocytosis » « Hexon – major capsid structural protein »
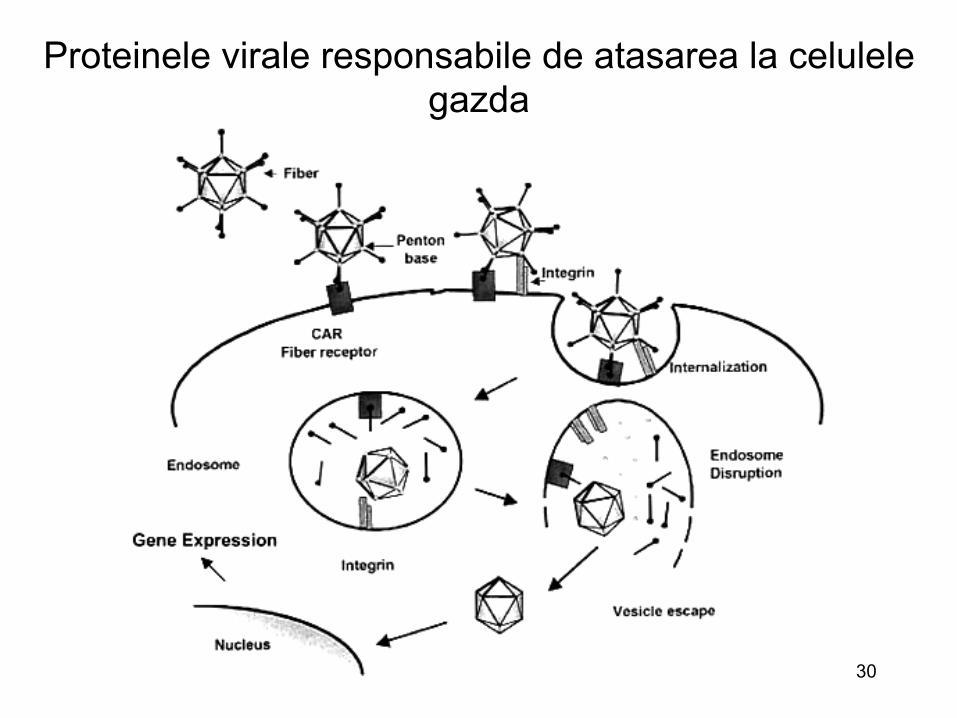
30
Proteinele virale responsabile de atasarea la celulele gazda

31
Rolul hidratilor de carbon : mono-, poli-zaharidele
• Rol important in timpul petrarii multor virusuri in celule
• Pentru unele virusuri, hidratii de carbon reprezinta insasi receptorii esentiali ai infectiei
• Pentru alte virusuri hidratii de carbon sunt factorii de atasare
• Implicarea hidratilor de carbon in infectiile virale: utilizarea enzimelor ce distrug glucidele determina o inhibitie a infectiei
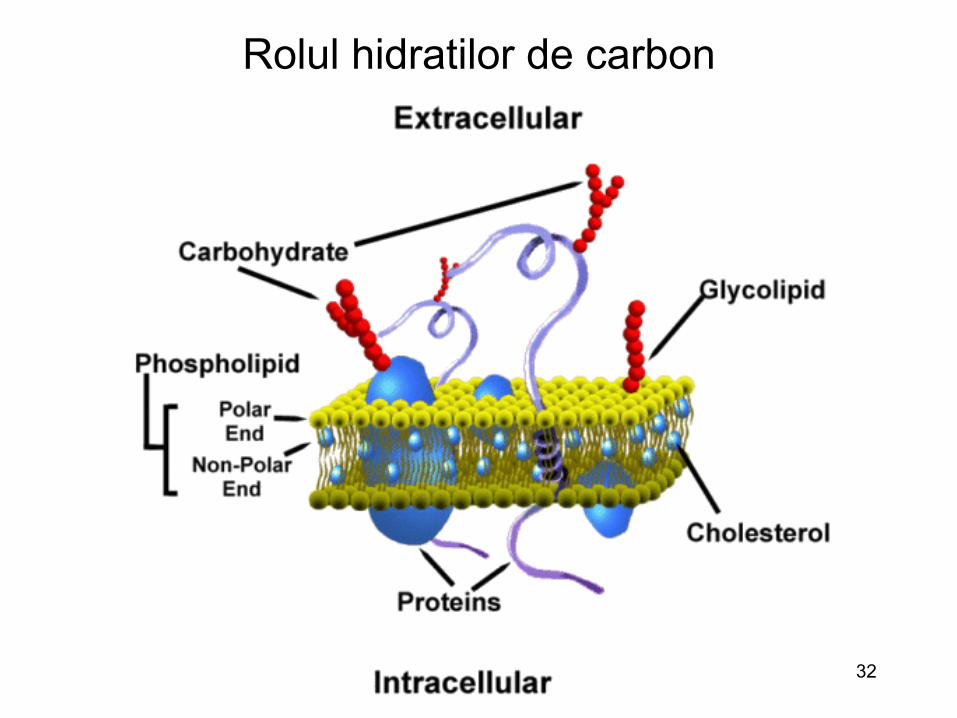
32
Rolul hidratilor de carbon

33
Rolul hidratilor de carbon • Glicoproteinele si glicolipidele (cu rezidurile terminale
de acid sialic) servesc drept receptori pentru multe virusuri
34

35
Rolul hidratilor de carbon
• Structura glicanilor exprimata in diferite specii prezinta o specificitate si joaca un rol esential in transmisia inter-specie
• Emergenta unei pandemii de gripa poate fi
asociata cu o rearanjare ale tulpinilor virale rezultand astfel o modificare a specificitatii hidratilor de carbon
36
Rolul hidratilor de carbon
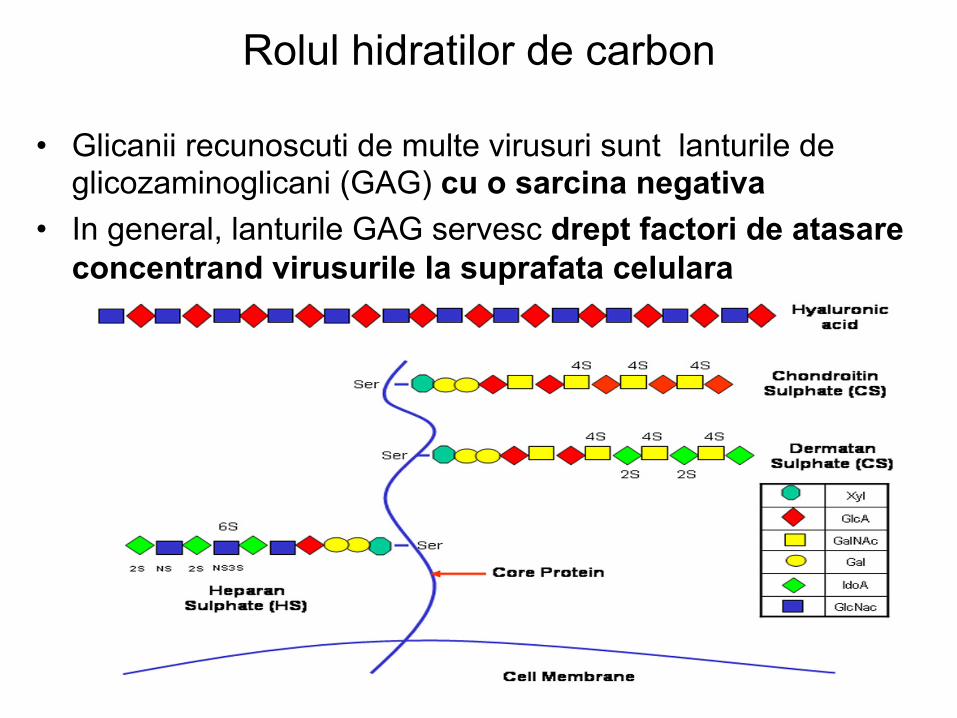
• Glicanii recunoscuti de multe virusuri sunt lanturile de glicozaminoglicani (GAG) cu o sarcina negativa
• In general, lanturile GAG servesc drept factori de atasare concentrand virusurile la suprafata celulara

37
Rolul hidratilor de carbon
• Lista virusurilor care se ataseaza la lanturile GAG (de exemplu heparin sulfat) este in crestere (herpes-; flavi-; retro-; parvo-; parvo-; picorna- si papilomavirus)
• In majoritatea cazurilor, virusurile sunt cele care recunosc
glicanii celulelor gazda (este posibila si recunoasterea inversa)
• Unele lectine celulare de suprafata ataseaza glicanii
prezenti la nivelul proteinelor anvelopei unor virusuri • Exemple de astfel de lectine: DC-SIGN (« cell specific
intracellular adhesion molecule »)

38
Sistemul de semnalizare celulara (utilizat de catre virusuri)
• Multe virusuri utlizeaza sistemul de semnalizare celulara pentru a favoriza intrarea si a optimiza infectia celulei
• Semnalele sunt utilizate pentru :
– A declansa accesul la coreceptori, – A induce endocitoza – A reprograma calea endocitozei in favoarea virusurilor – A induce conditiile intracelulare favorabile infectiei
• Semnalizarea incepe la nivelul membranei plasmatice (dupa atasarea virusului la receptorii specifici) dar poate continua si la nivelul organelelor citoplasmatice ( endozomi sau caveozomi)

Sistemul de semnalizare celulara (utilizat de catre virusuri)
• Kinazele:
Sunt enzime ce catalizeaza reactii de fosforilare prin adaugarea unui ion fosfat la o molecula tinta Molecula tinta, numita substrat, poate fi : proteine, lipide, glucice etc.
39

Sistemul de semnalizare celulara (utilizat de catre virusuri)
• Mecanismele de fosforilare-defosforilare sunt utilizate pentru a regla activitatea celulei.
• Semnalizarea celulara utilizeaza PK (protein-kinaze) ce vor actiona asupra unor substraturi specifice pentru a induce un raspuns adecvat al celulei
40

41
Sistemul de semnalizare celulara (utilizat de catre virusuri)
• Atasarea virusului la receptorul specific duce la activarea tirozinei sau a altor kinaze ce declanseaza cascada de semnale : de la nivelul membranei plasmatice pana la nivelul citoplasmei si chiar a nucleului
• Semnalele induse de virusuri depind de o serie de
mesageri secundari (Ca, diacylglicerides etc), GTP (guanosine trifosfataze) ce controleaza traficul membranar

Sistemul de semnalizare celulara (utilizat de catre virusuri)
Concluzie : Sistemul de semnalizare celulara este utilizat de diferitele familii virale pentru atasarea, internalizarea si deci infectarea celulei gazda
42
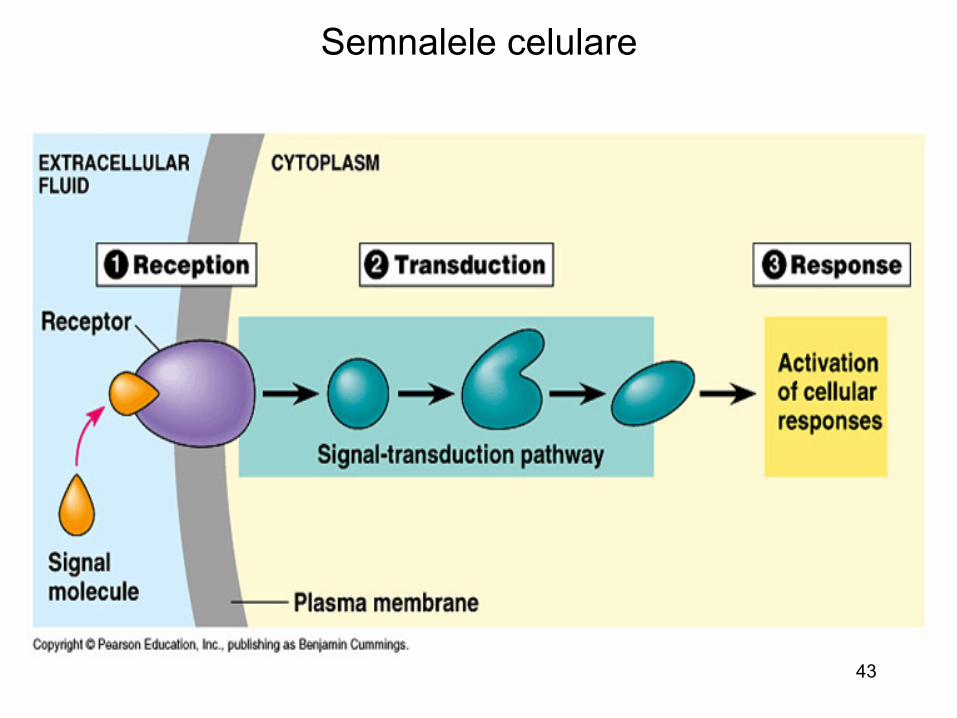
43
Semnalele celulare
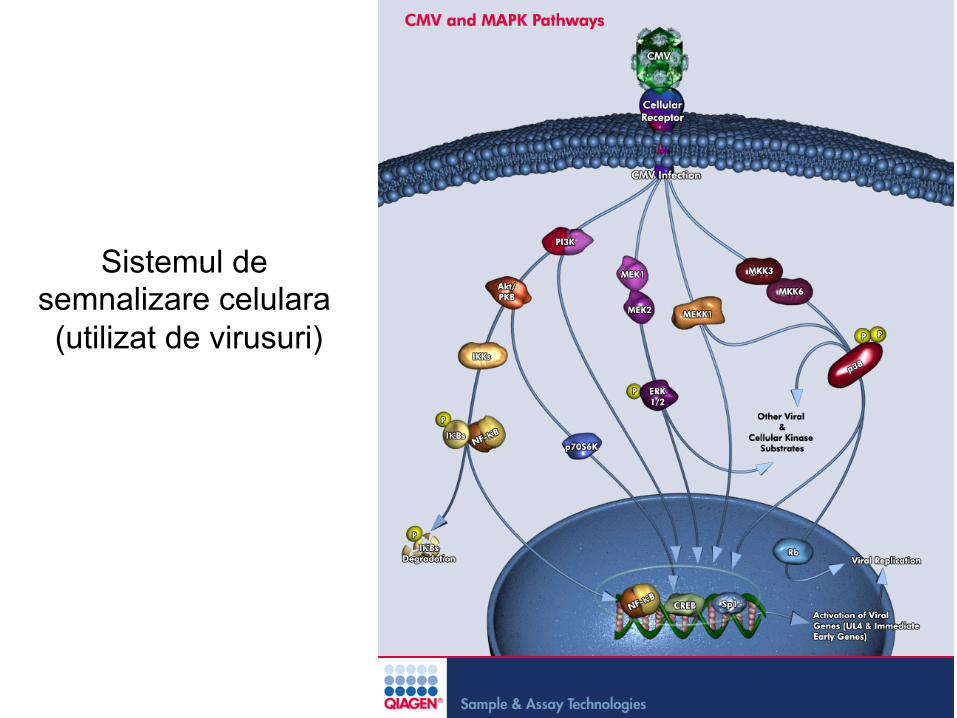
Sistemul de semnalizare celulara (utilizat de virusuri)
44

http://highered.mcgraw-hill.com/sites/0072556781/student_view0/chapter18/animation_quiz_1.html
45
Diferitele cai de patrundere a unui virus intr-o celula • Obiectiv: internalizarea genomului viral • Modurile de internalizare sunt comparabile cu internalizarea
macromoleculelor • Etape ce necesita un aport de energie
– Penetrare directa (rar) – Intrare prin fuziunea membranelor sub actiunea proteinelor
virale : virusurile cu anvelopa +++ – Intrarea prin endocitoza : formarea de vacuole
citoplasmatice – Cele doua mecanisme pot fi asociate : endocitoza si apoi
fuziune ( virusurile cu anvelopa)
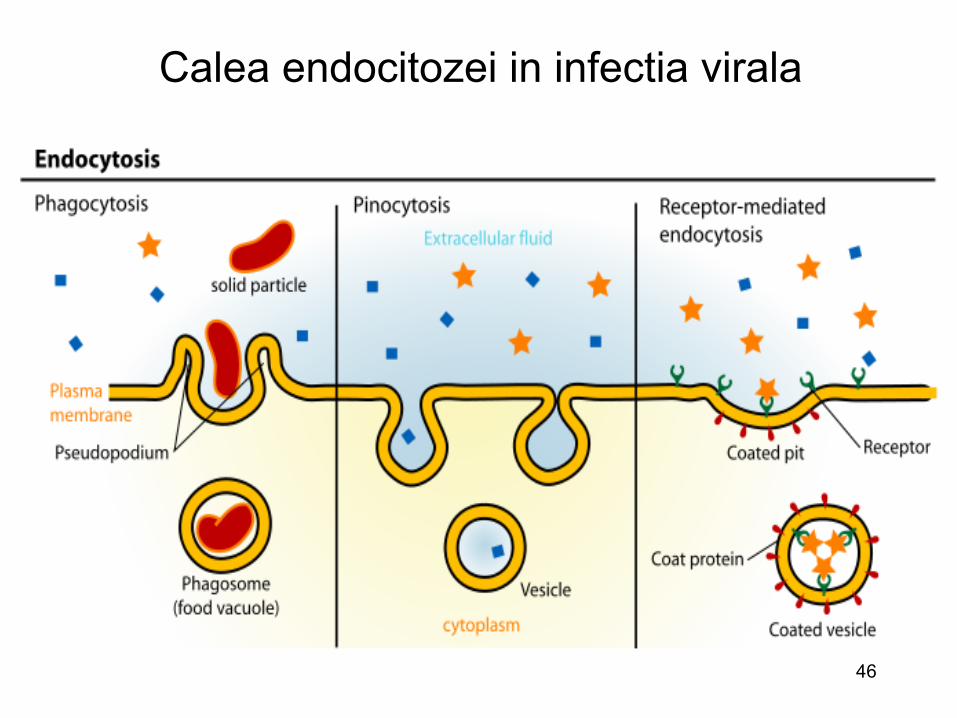
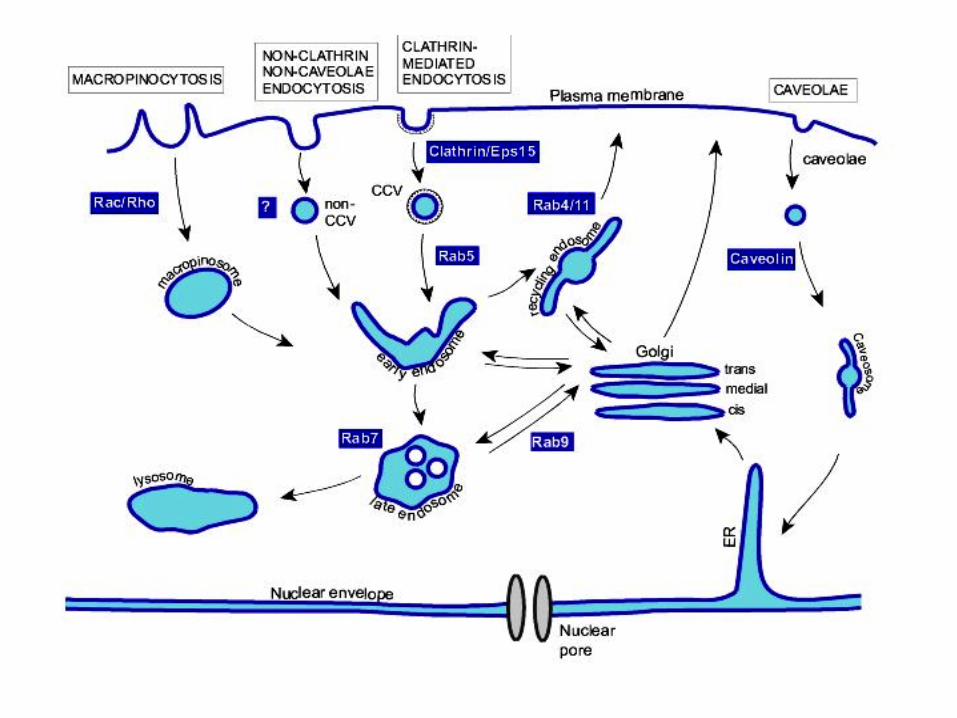
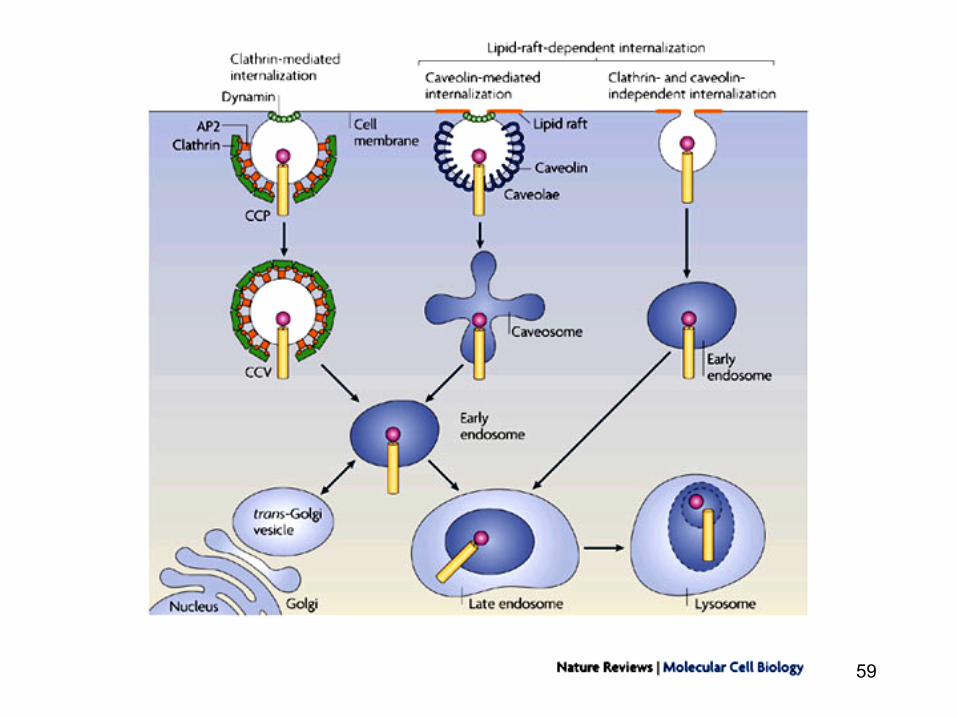
Calea endocitozei in infectia virala
46

47
Calea endocitozei in infectia virala
• Virusurile utilizeaza endocitoza drept cale de intrare in celula gazda: o cale sigura de deplasare a virusurilor spre citoscheletul cortical si alte bariere citoplasmatice
• Penetrarea intracelulara pe calea endocitozei determina
intarzierea imunorecunoasterii • Organelele intracelulare pot contine enzime (proteaze etc.)
necesare patrunderii si eliberarii genomului viral

48
Calea endocitozei in infectia virala (« clathrin coated pit » )
• Calea cea mai utilizata de catre virusuri pentru
patrunderea in celulele gazda este endocitoza mediata de « clathrin coated pit »
• Aceasta este calea utilizata de catre celule pentru
internalizarea de fluide, proteine membranare si lipide
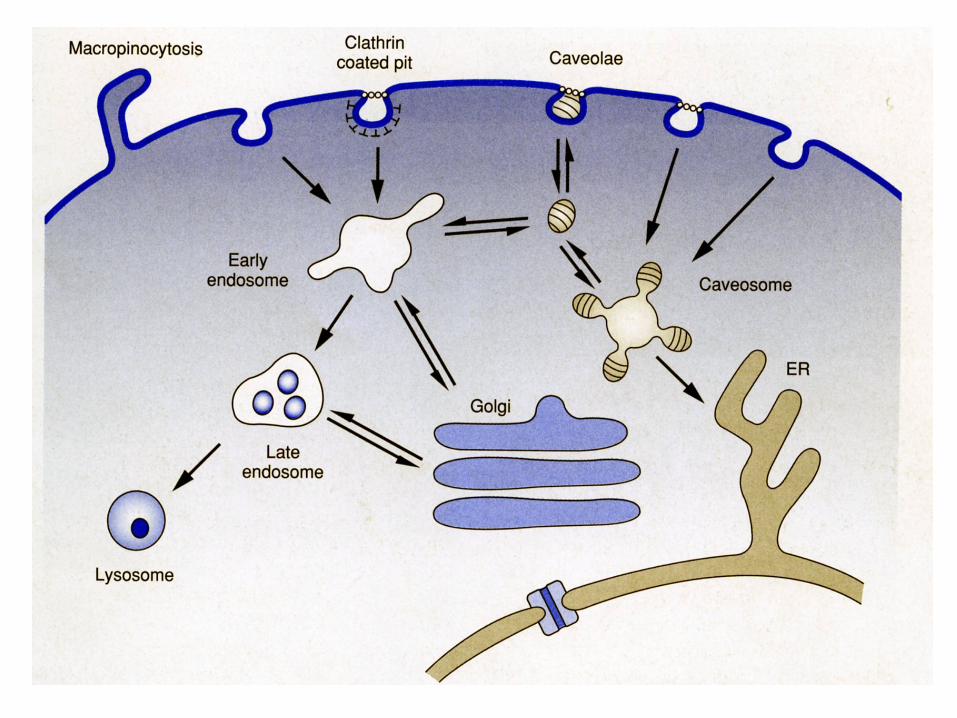
49

50
Calea endocitozei in infectia virala (« clathrin coated pit »)
• Virusurile se ataseaza la receptorii specifici ce prezinta semnalele necesare pentru internalizarea de catre fosele « clathrin-coated »
• Patrunderea se caracterizeaza printr-o cinetica
rapida: – Virusurile sunt internalizate cateva minute dupa
atasare si intr-o cantitate mare (≤ 3000 particule virale/min)
51

52
Calea endocitozei in infectia virala « clathrin coated pit »)
• Diametrul mare (peste 120 nm) al veziculelor este suficient pentru internalizarea (prin endocitoza) majoritatii virusurilor animale
• Endocitoza « clathrin-mediated » este un proces
continu si extrem de controlat • Calea « clathrin-mediaded » transporta virusurile
si receptorii asociati pana la nivelul endozomilor precoce unde sunt expusi la un pH moderat acid

53
Calea endocitozei in infectia virala « clathrin coated pit »
• Urmatoarea etapa este la nivelul endozomilor tarzii ce prezinta un pH mai scazut
• Urmatarea etapa (eventual) este la nivelul
lizozomilor unde particulele virale • In general lizozomii sunt rar implicati in
patrunderea particulelor virale

54
Calea endocitozei in infectia virala « clathrin coated pit »
• Dupa internalizare, virusurile dependente de un pH relativ ridicat patrund in endozomii precoce la cateva minute (5-10 minute) dupa internalizare
• Acesti endozomi sunt localizati la periferie si au un pH = 6 - 6,5
• Virusurile dependente de un pH mai scazut patrund mai
tarziu (10-50 minute dupa infectie) in endozomii tarzii (pH= 5-6)

55
Calea endocitozei « caveolae pathways » in infectia virala
• O alta cale de patrundere a particulelor virale este calea « caveolae pathways »
• Este dependenta de colesterol si de activarea unei
tirozin-kinaze • Veziculele ce transporta particulele virale (formata din
membrana plasmatica) sunt mici (70 nm diametru) • Dependenta de colesterol reflecta rolul central al lipidelor in
acesta cale de endocitoza

56
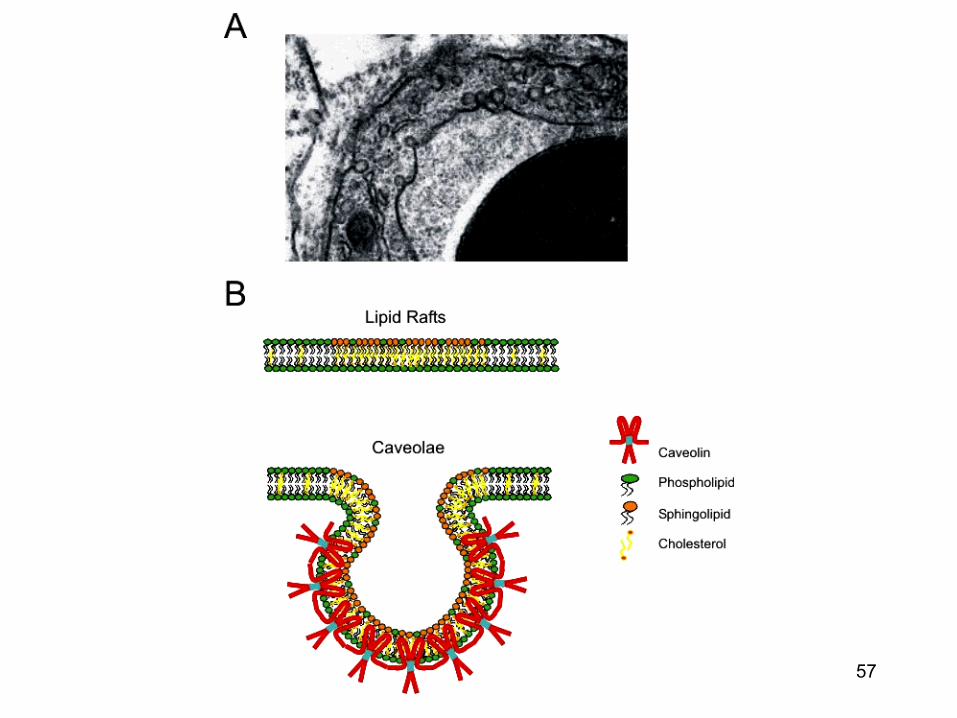
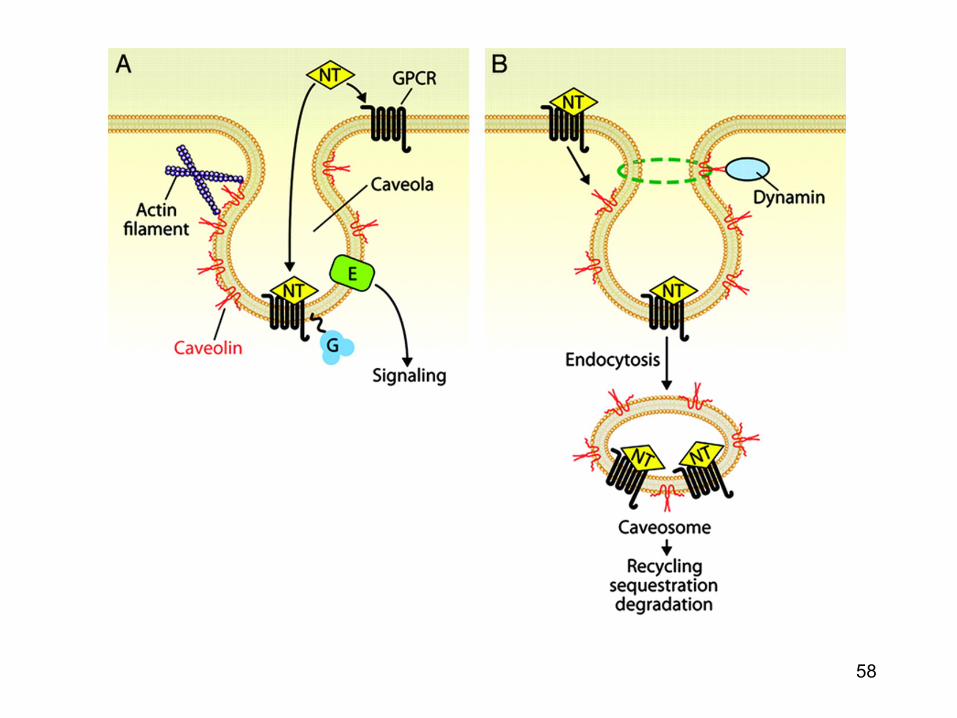
Calea endocitozei in infectia virala virala « caveolae pathways »
• Caveolele sunt formatiuni de 50-70 nm ce contin majoritar caveoline (proteine) iar membrana este bogata in colesterol si sfingolipide
• Suprafata majoritatii caveolelor sunt stabile • Odata cu fosforilarea tirozin-kinazei (de catre virus),
caveolele devin mai mobile si se deplaseaza in citoplasma unde fuzioneaza formand caveozomii.
57
58
59
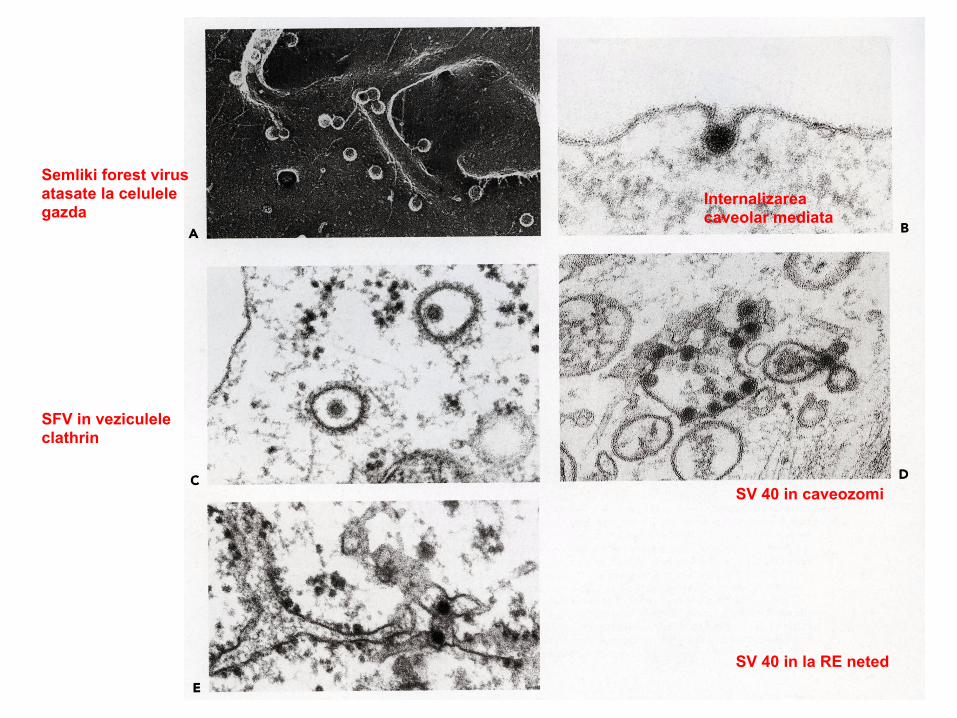
60
Semliki forest virus atasate la celulele gazda
Internalizarea caveolar mediata
SFV in veziculele clathrin
SV 40 in caveozomi
SV 40 in la RE neted

61
Penetrarea virusurilor prin fuziunea membranara (virusurile cu anvelopa)
• Virusurile cu anvelopa utilizeaza « vezicule de transport »
• Virusurile cu anvelopa transporta genomul viral
de la o celula la alta prin fuziunea membranara • Anvelopa virala este de fapt membrana veziculelor
de transport • Vezicula este formata (dupa « incarcarea »
capsidei virale in celulele infectate) prin fuziunea membranei plasmatice
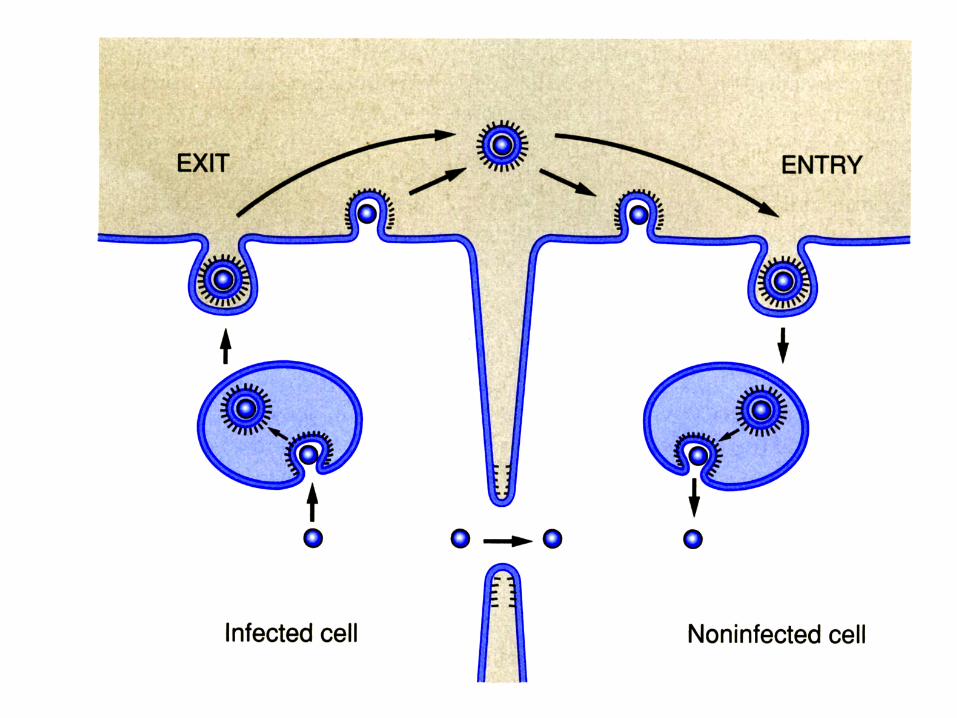
62

63
Patrunderea virusurilor prin fuziunea membranara
• In timpul patrunderii virusului, fuziunea are loc in general cu membrana endozomilor (nu intotdeauna)
• In acest context, infectia se poate produce printr-o fuziune directa dintre membrana plasmatica a unei celule infectate cu o celula vecina
• Fuziunea este mediata de proteinele virale exprimate la suprafata celulei infectate
• Rezultatul este adesea formarea de mari celule multinucleate (sincitii)

64
Patrunderea virusurilor prin fuziunea membranara
• Proteinele virale membranare implicate in fuziune sunt proteine membranare integrale
• Pentru a deveni competente (in fuziune) proteinele
virale (dupa asamblarea si maturarea lor in RE) sunt clivate (proteolitic) in cursul deplasarii lor in AG
• Exista doua tipuri de proteine virale de fuziune : de
tip I si tip II
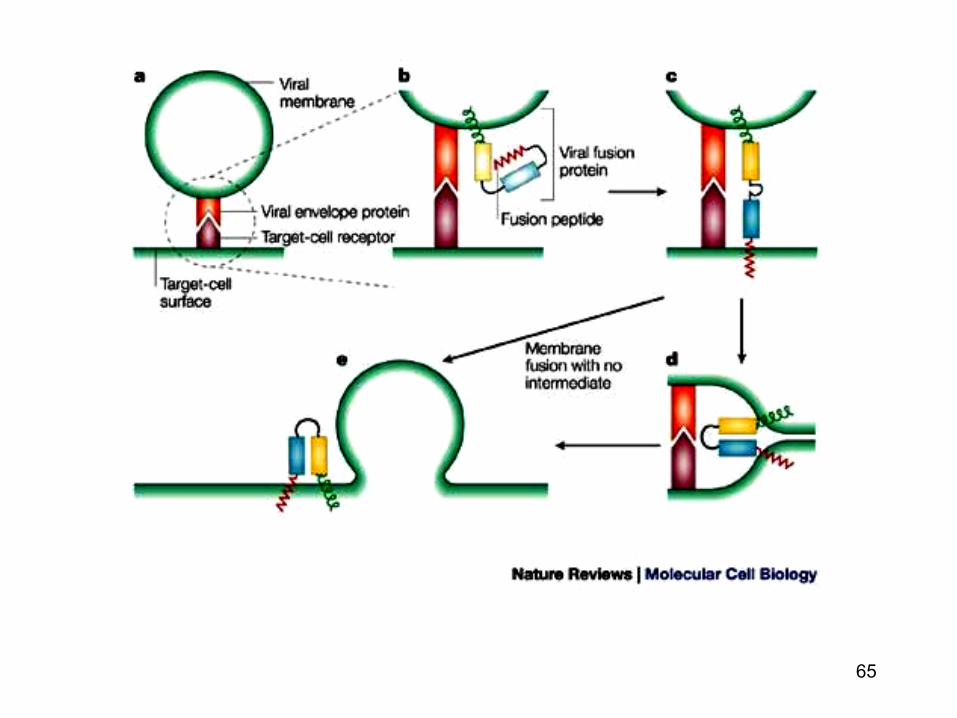
65
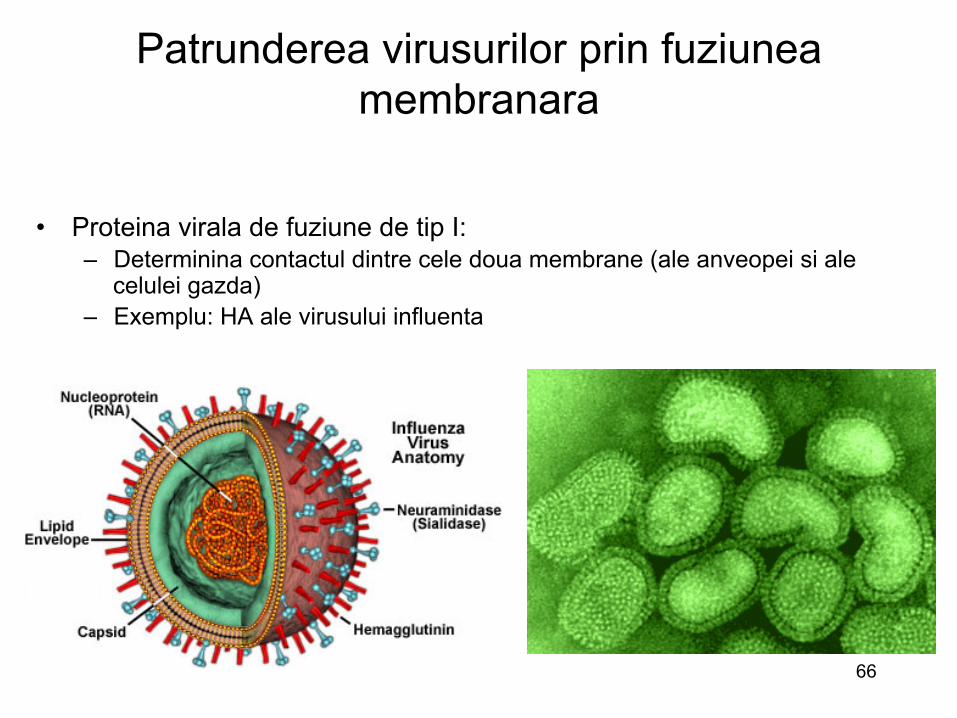
66
Patrunderea virusurilor prin fuziunea membranara
• Proteina virala de fuziune de tip I: – Determinina contactul dintre cele doua membrane (ale anveopei si ale
celulei gazda) – Exemplu: HA ale virusului influenta

67
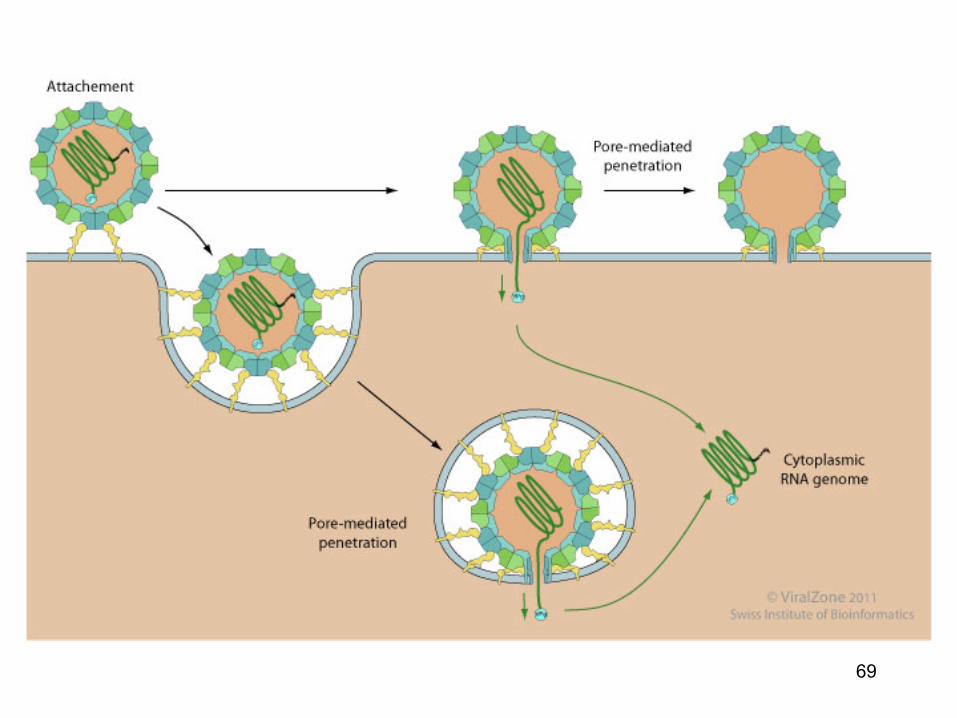
Patrunderea virusurilor nude
• Ca si virusurile cu anvelopa, particulele nude patrund in citosol de-a lungul membranei plasmatice sau de-a lungul membranelor diferitelor organele (endozomii, lizozomii, sau caveozomii)
• Semnalul ce declanseaza patrunderea virusului este dat de:
receptorii specifici, de un pH scazut etc • Un virus nud trebuie sa transfere capsida, genomul si
proteinele virale de-a lungul membranei celulare fara avantajul mecanismului de fuziune membranare

68
Patrunderea virusurilor nude • Se pare ca virusurile nude utilizeaza trei strategii
generale • Punctia membranei : particulele virale genereaza
un por la nivelul membranei (prin care genomul este eliberat in mod selectiv in citosol); capsida virala nu patrunde in citosol
• Perforarea : transferul intregii capside de-a lungul
membranei fara liza majora a membranei • Liza membranei: particulele virale determina liza
membranei organelelor citoplasmatice
69
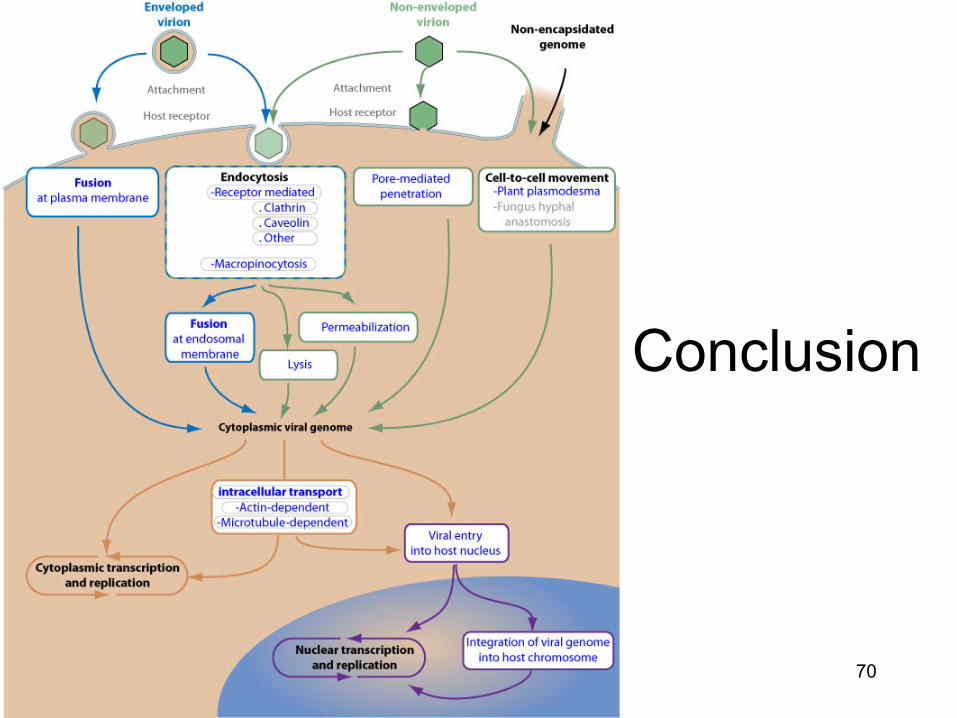
Conclusion
70

71
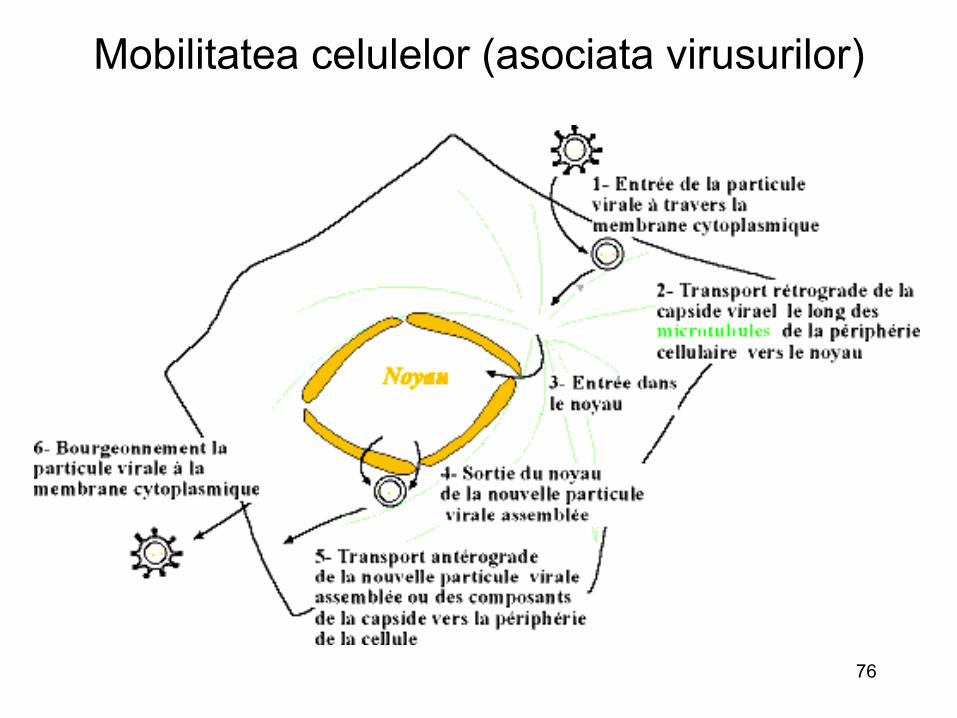
Traficul intracelular si virusurile
• Inainte de replicarea genomului viral, particulele virale si capsidele trebuie transportate in locuri specifice : in nucleu ( localizari specifice) sau in citosol (asociate RE sau RE-Golgi)
• Virusurile utilizeaza sistemul de transport citoplasmatic celular
• Pentru distantele mari, particulele virale utilizeaza mecanismele mediate de reteaua de microtubuli
• Virusurile parcurg distante diferite: virusurile neurotrope se deplaseaza de-a lungul axonilor (retrograd)
• Alte virusuri, se multiplica la nivelul suprafetei citosolice ale endozomilor si lizozomilor (particulele virale nu trebuie sa paraseaza locul de patrundere)

72
Traficul intracelular si virusurile
• Pentru a se deplasa in citoplasma, virusurile au doua posibilitati: – Dupa ce patrund, sunt transportate de veziculele procesului de
endocitoza (utilizeaza astfel miscarea veziculelor) – Dupa ce patrund in citosol, virusurile sau capsida lor se asociaza
cu molecule de tipul dineina
• Majoritatea virusurilor utilizeaza ambele cai de deplasare in citoplasma : traficul citoplasmatic sau vezicular

73
Mobilitatea celulelor (asociata virusurilor)
• Marcarea proteinelor celulare GFP permite (prin
microscopie in timp real) analiza etapelor de patrundere a virusurilor
• Prima etapa este atasarea virusului la un
receptor celular
• Etapele urmatoare depind de natura virusului si a receptorului : endocitoza, integrarea in caveole etc.

74
Mobilitatea celulara (asociata virusurilor)
• Mobilitatea celulara joaca un rol activ in transportul particulelor virale in celule gazda
• Mobilitatea celulara: formata din extensii mobile si fine
ale membranei plasmatice stabilizate de o retea de filamente de actina
• Miscarile au loc cu o frecventa de 1-2 µm/min • Dupa endocitoza si penetrare, citoscheletul format din
actina-microtubuli favorizeaza deplasarea intracelulara a particulelor virale
75
Mobilitatea celulara (asociata virusurilor)
76
Mobilitatea celulelor (asociata virusurilor)
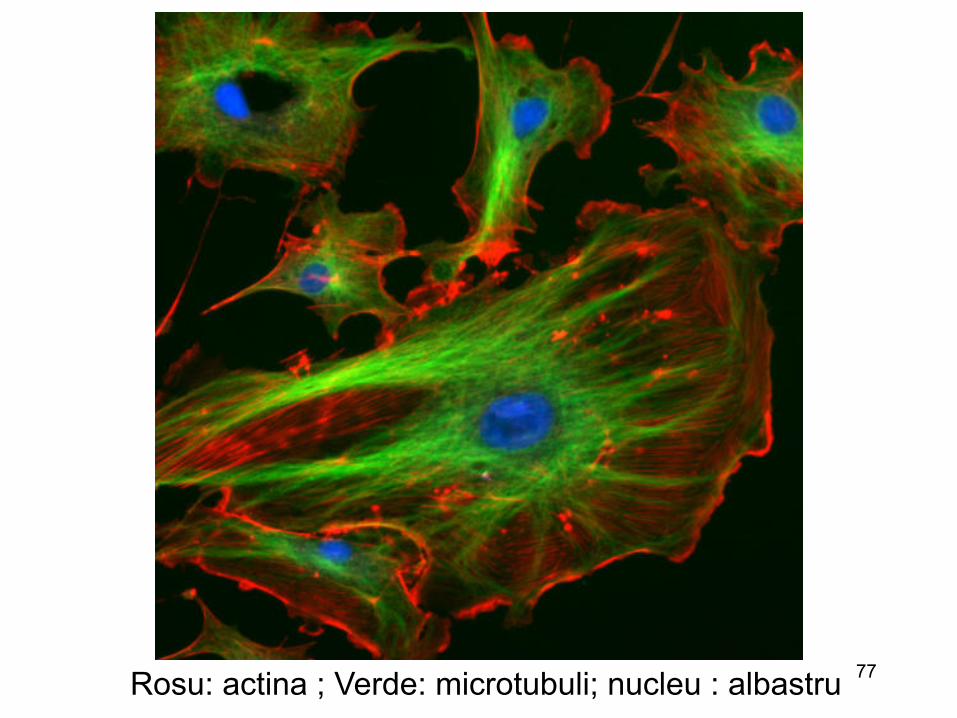
77 Rosu: actina ; Verde: microtubuli; nucleu : albastru

Dezasamblarea virusurilor
• Intrarea majoritatii virusurilor include o dezasamblare totala sau partiala
• Pentru virusurile cu anvelopa : pierderea
anvelopei in timpul fuziunii membranare
78

Dezasamblarea (virusurile nude)
• Procesul de dezasamblare include: – Modificari conformationale – Pierderea progresiva a proteinelor structurale – Clivaje proteolotice – Interactiuni intermoleculare
• Exemplu : adenovirus 2 :
– Dezasamblarea incepe la suprafata celulara prin pierderea fibrelor – Activarea proteazelor virale (L3/p23) – Clivajul proteolitic al proteinelor capsidei – Eliminarea ADN-ului prin NPC (« nuclear pore complex »)
79
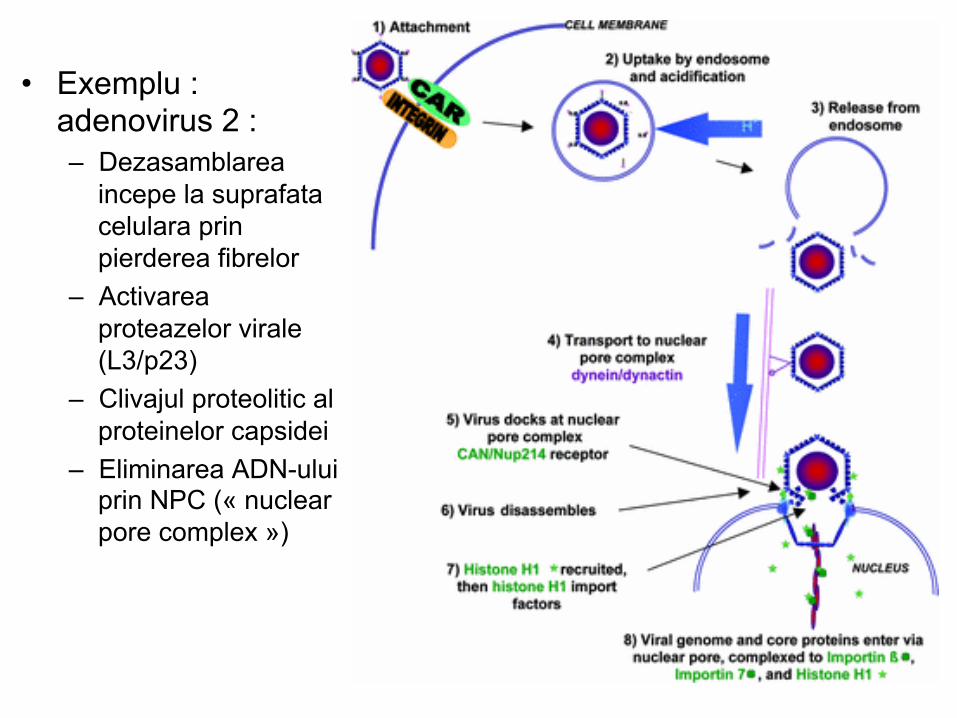
• Exemplu : adenovirus 2 : – Dezasamblarea
incepe la suprafata celulara prin pierderea fibrelor
– Activarea proteazelor virale (L3/p23)
– Clivajul proteolitic al proteinelor capsidei
– Eliminarea ADN-ului prin NPC (« nuclear pore complex »)
80

81
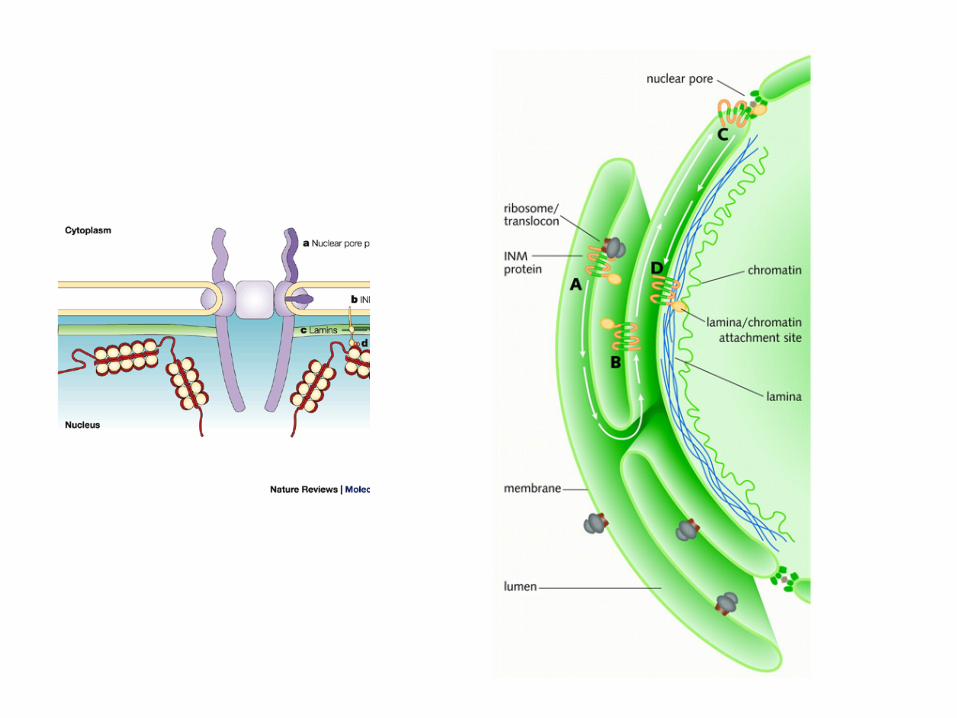
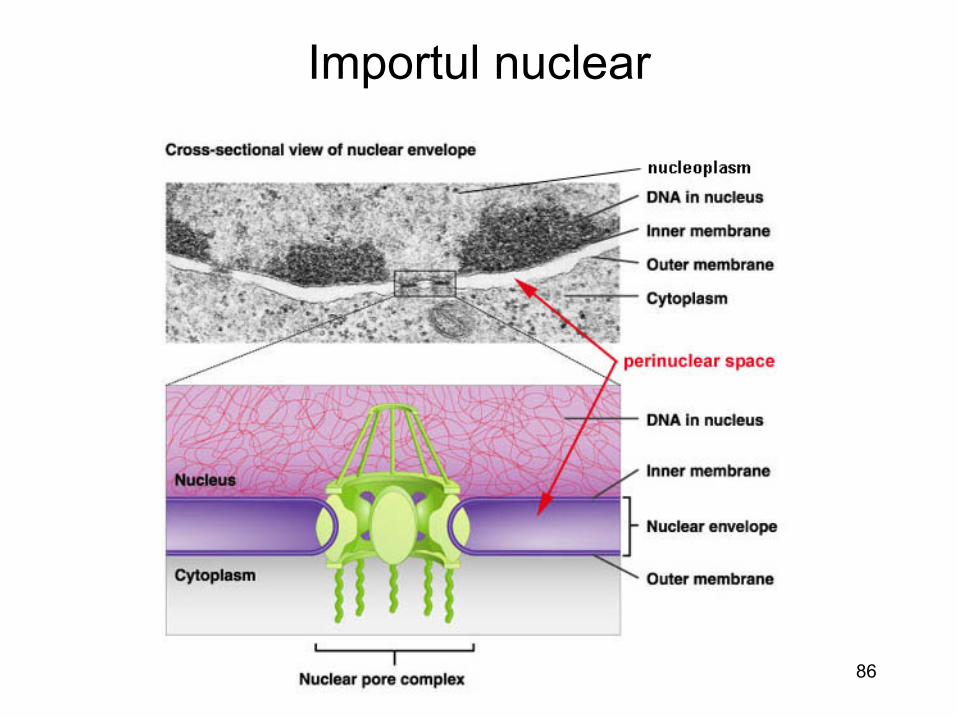
Importul nuclear
• Majoritatea virusurilor ADN si cateva virusuri ARN (-) se multiplica in nucleu
• Pentru a patrunde (genomul si proteinele virale accesorii) in
nucleu virusurile au doua posibilitati: – pot utiliza NPC (« nuclear pore complex ») sau – Virusurile si capsidele virale pot sa astepte in citosol ruperea
membranei nucleare (in dimpul diviziunii celulare) : retrovirusurile (cu exceptia lentivirusurile)
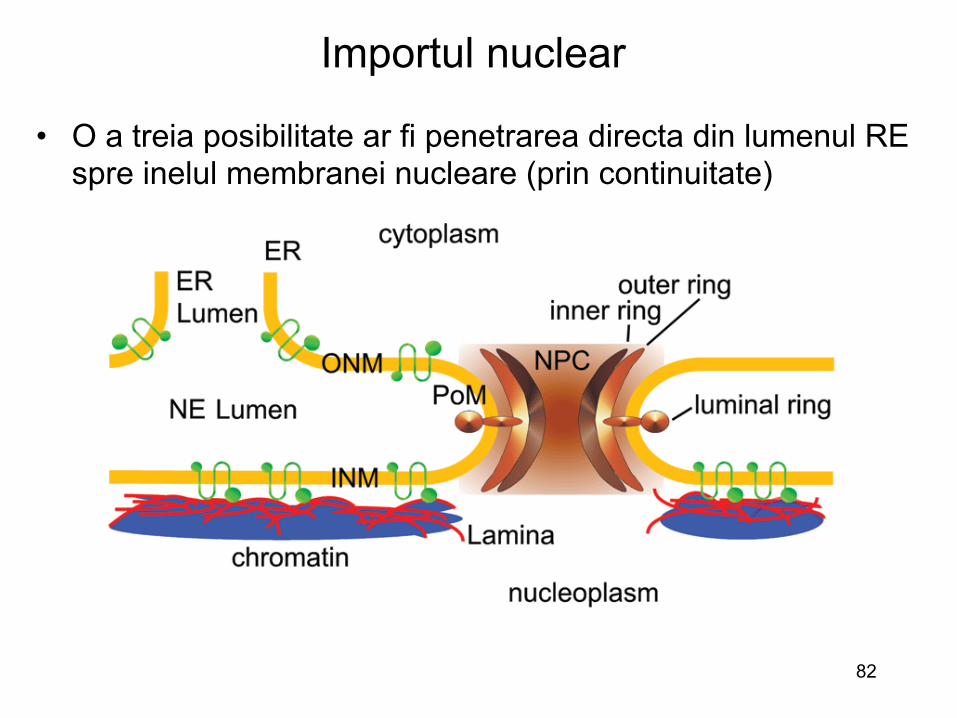
Importul nuclear
• O a treia posibilitate ar fi penetrarea directa din lumenul RE spre inelul membranei nucleare (prin continuitate)
82
83
Importul nuclear
Adenovirus
84
Importul nuclear
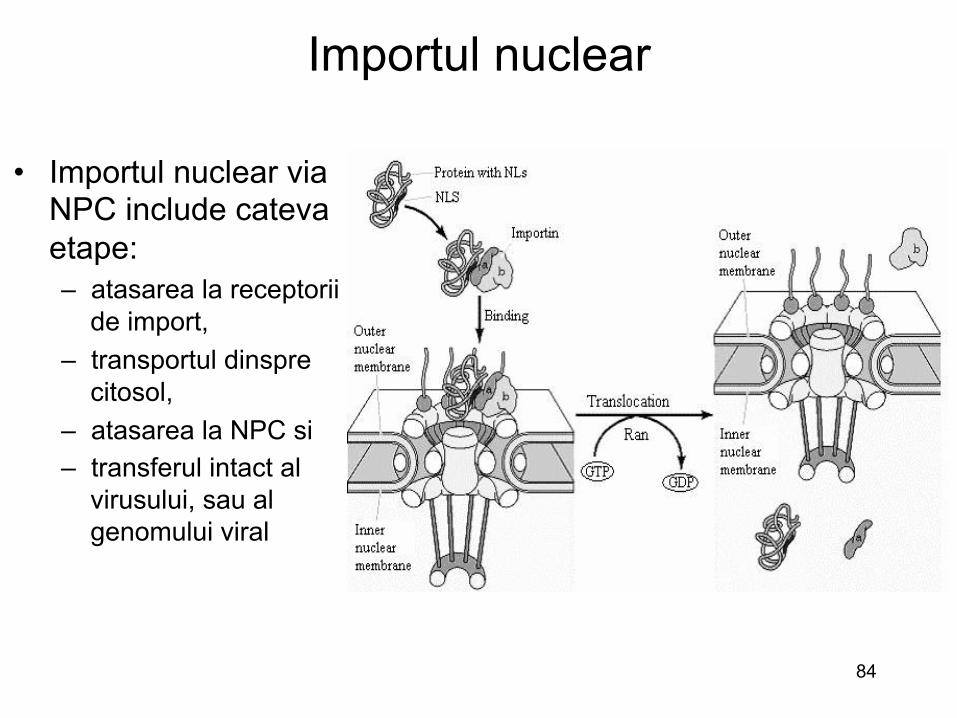
• Importul nuclear via NPC include cateva etape: – atasarea la receptorii
de import, – transportul dinspre
citosol, – atasarea la NPC si – transferul intact al
virusului, sau al genomului viral
Importul nuclear
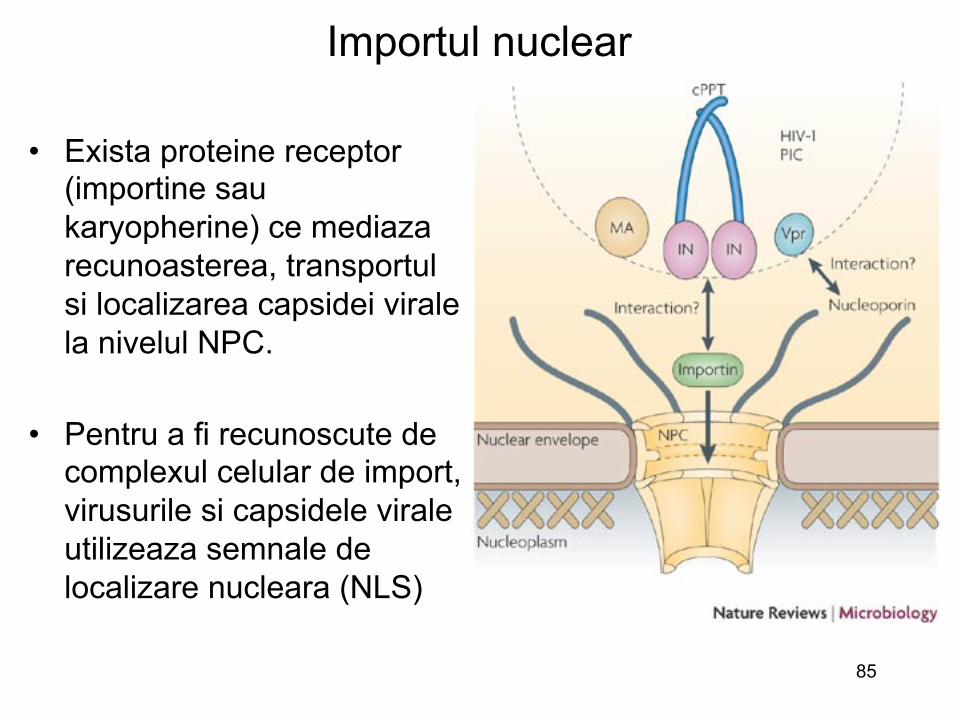
• Exista proteine receptor (importine sau karyopherine) ce mediaza recunoasterea, transportul si localizarea capsidei virale la nivelul NPC.
• Pentru a fi recunoscute de
complexul celular de import, virusurile si capsidele virale utilizeaza semnale de localizare nucleara (NLS)
85
86
Importul nuclear
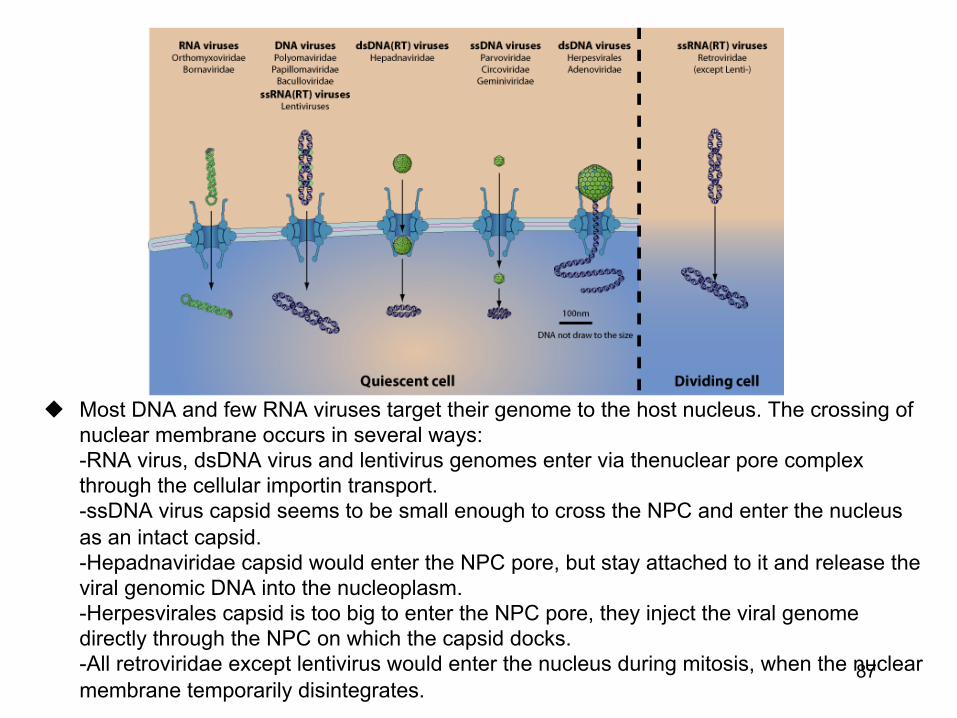
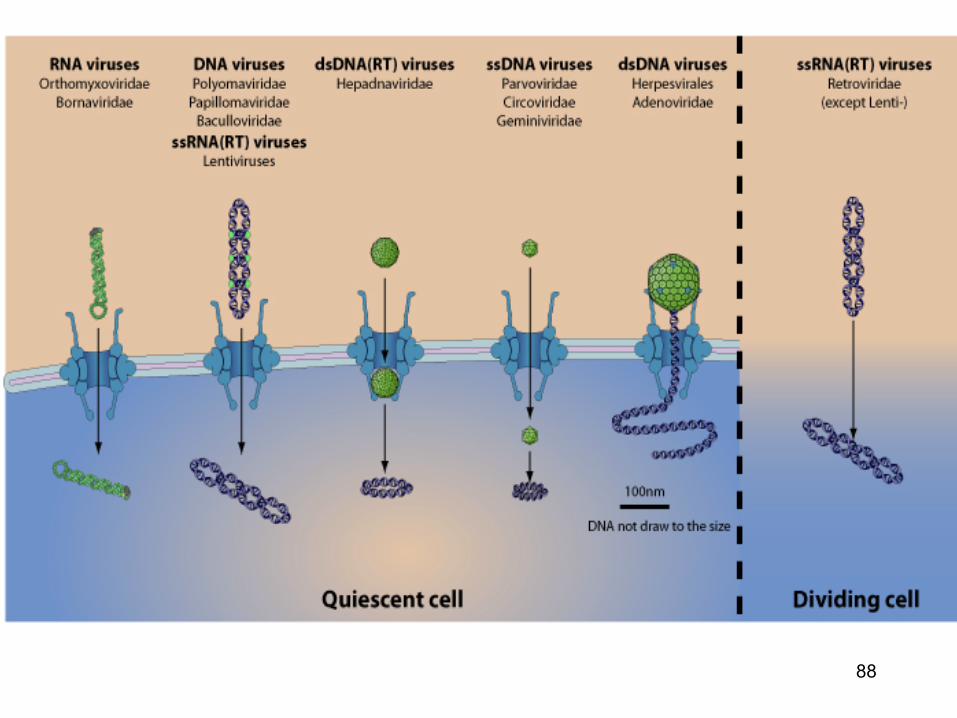
u Most DNA and few RNA viruses target their genome to the host nucleus. The crossing of nuclear membrane occurs in several ways: -RNA virus, dsDNA virus and lentivirus genomes enter via thenuclear pore complex through the cellular importin transport. -ssDNA virus capsid seems to be small enough to cross the NPC and enter the nucleus as an intact capsid. -Hepadnaviridae capsid would enter the NPC pore, but stay attached to it and release the viral genomic DNA into the nucleoplasm. -Herpesvirales capsid is too big to enter the NPC pore, they inject the viral genome directly through the NPC on which the capsid docks. -All retroviridae except lentivirus would enter the nucleus during mitosis, when the nuclear membrane temporarily disintegrates.
87
88


















