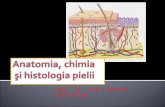
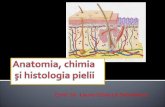
Funcia de Excreie a Pielii
28
Funcţia de excretie a pielii Funcţia de excreţie a pielii Rusu Alexandrina Maria Biologie (B2), anul 3 Universitatea “Babes Bolyai” 1
Transcript of Funcia de Excreie a Pielii

Funcţia de excretie a pielii
Funcţia de excreţie a pielii
Rusu Alexandrina MariaBiologie (B2), anul 3
Universitatea “Babes Bolyai”Facultatea de Biologie şi Geologie
Cluj-Napoca, 2010
1

Funcţia de excretie a pielii
Cuprins
1. Introducere.......................................................................................32. Sistemul tegumentar.........................................................................33. Funcţia secretoare.............................................................................44. Tegumentul e o barieră
osmotică......................................................85. Transport paracelular si transcelular.......................................126. Branhiile peştilor transportă ioni, în şi din apă.......................147. Reptilele şi păsările sunt înzestrate cu glande de
sare.............158. Glandele rectale ale elasmobranhiilor excretă Na+ şi Cl¯
reţinând ureea..........................................................................169. Bibliografie.............................................................................17
2

Funcţia de excretie a pielii
1.Introducere
În afara mecanismului secretor renal, la vertebrate ca şi la nevertebrate există o
serie de mecanisme extrarenale secundare. Unii din produşii rezultaţi în procesele de
catabolism ale organismului uman şi animal se elimină fie prin plămâni, fie prin
branhii, fie prin intestin, fie prin tegument.
Prin plămâni se elimină vapori de apă, bioxid de carbon şi în cazuri speciale,
anestezicii volatili; de asemenea, prin plămâni se poate elimina şi alcoolul.
Prin branhii se elimină bioxidul de carbon, sărurile monovalente, la teleosteenii
marini, uree şi mai ales amoniac, 70% din N amoniacal, la teleosteenii dulcicoli.
Prin intestin se pot elimina săruri minerale şi apă. (Roşca, 1977)
2. Sistemul tegumentar
Excreţia tegumentară este caracteristică tuturor vertebratelor. La batracieni,
tegumentul are rol în osmoreglare, se elimină la exterior apă şi săruri minerale în
cantitate mică. La reptile, tegumentul este permeabil pentru apă, iar năpârlirea
constituie un mecanism de excreţie, eliminându-se cataboliti acumulaţi în tegument.
La vertebratele superioare, la păsări şi la mamifere, pielea şi derivatele ei (unghii, păr,
pene, laână) acumulează o serie de produşi azotaţi, care sunt apoi eliminaţi la exterior
odată cu eliminarea acestora. (Strungaru şi colab., 1983)
Animalele își schimbă profilul fluidului extracelular, importând sau exportând ioni
și apă prin acele țesuturi epiteliale care comunică cu mediul exterior. Aici intră
suprafețele externe, ca pielea, branhiile, precum și suprafețele „externe” încorporate
ca lumenul din sistemul excretor și cel digestiv. Aceste țesuturi epiteliale formează
3

Funcţia de excretie a pielii
suprafața de separație dintre animal și mediu și, de obicei, au alte responsabilități
fiziologice, precum respirația și digestia. Totuși, proprietățile care fac ca un țesut să
fie bun la schimbul de gaze sau absorbția nutrienților – suprafață mare și nivel ridicat
de permeabilitate- îl fac mai vulnerabil la mișcarea ionilor și a apei. Un țesut epitelial
are proprietăți care reflectă un echilibru între diferitele sale roluri fiziologice. Fiecare
animal se bazeză pe șiruri de țesuturi pentru a controla balanța osmotică și ionică.
(Moyes si Schulte, 2006)
3. Funcţia secretoare
Funcţia secretoare a tegumentului este bine dezvoltată la om şi la mamiferele la
care există un sistem de glande sudoripare, care sunt capabile să secrete la suprafaţa
tegumentului o sudoare cu o compoziţie complexă, în care intră şi produşii de
excreţie.
3.1. Sudoarea
O funcţie sudoripară a tegumentului întâlnim la om, la cal, la bou, la pisica şi la
căţei; ea lipseşte complet la câinele adult, la iepure, la capră, la liliac, la cetacee, la
care lipsesc şi glandele sudoripare.
Glandele sudoripare şi regiunile sudoripare sunt răspândite neuniform: la om sunt
răspândite inegal pe podul palmei, pe talpa piciorului, la axile, pe frunte, pe partea
anterioară a pieptului; la bou şi la cal sunt răspândite pe toată suprafaţa corpului, la
pisica şi la căţei se afla răspândite numai în pulpa degetelor, acolo unde tegumentul nu
este acoperit de păr. (Roşca, 1977)
Încă din 1917, Schiefferdecker a denumit cele 2 tipuri de glande sudoripare
existente la om şi la alte mamifere, glande ecrine şi respectiv apocrine. Aceste
denumiri exprima de fapt modul de secreţie al acestor glande, dedus din aspectul lor
histologic; în primul caz, secreţia se produce fără o pierdere aparenţa de substanţă
celulară, pe când în cazul glandelor apocrine, secreţia reprezintă aparent un derivat al
citoplasmei însăşi. Principala funcţie e glandelor sudoripare o reprezintă
termoreglarea. Termoreglarea şi în mod implicit activitatea acestor glande este
controlată de structuri nervoase centrale – hipotalamusul anterior via sistemul nervos
4

Funcţia de excretie a pielii
vegetativ simpatic. (Cârsteanu şi Dumitru, 1976)
3.1.1G landele ecrine
Sunt formate dintr-un glomerul secretor şi un duct care se deschide la suprafaţa
pielii. Porţiunea secretorie constă dintr-un rând de celule, spre lumen, printre care se
găsesc celule mioepiteliale aplicate pe o membrană bazală. Celulele secretorii sunt de
două tipuri: “clare” şi “întunecate”. Majoritatea celulelor sunt mari, clare, cu
mitocondrii în mare măsură vacuolizate, glicogen şi granule de secreţie, ele secretând
material apos şi glicogen. Celulele cu aspect întunecat conţin granule secretorii
electron-dense cu substanţa mucoida. Ele se dizolvă în citoplasma apicală, spre
ectoplasma, fiind exocitate în lumen odată cu veziculele mici. În lumen nu sunt
observate granule de secreţie.
Ductul ecrin intradermic este format din două straturi de celule, bazal şi lumenal.
Stratul de celule bazale conţin în mod deosebit mitocondrii şi microvili, iar celulele
lumenale conţin numeroase tonofilamente su au joncţiuni desmozomale. Aceşti tubi se
deschid la suprafaţa pielii printr-un canal helicoidal. (Păiş, 1983)
Secrertia ecrina reprezintă o soluţie apoasă, cu o compoziţie variabilă în funcţie de
cantitatea secreţiei, de durata pasajului prin conduct. Cu toate că prezintă similitudini
fundamentale cu plasma din care derivă, aceasta secreţie este hipotona, conţine mult
mai puţini electroliţi şi foarte puţină glucoză. În schimb, oferă o mare concentraţie de
lactat. Secreţia glandelor ecrine este reprezentată de apă şi electroliţi: Na Cl şi K din
care o parte se reabsorb la nivelul ductului glandei. Mai secretă uree, calciu,
aminoacizi, imunoglobuline, bicarbonat. Este stimulată de temperatură şi de stimulii
emoţionali via simpaticul colinergic.
Sunt aprox 3 milioane (2-5 mil) densitatea lor variind întere 80-600/cm2, fiind
singurele glande sudoripare al căror rol este clar: termoreglarea.Se activează de la
naştere spre deosebire de cele apocrine care se activează la pubertate. Prezenţa lor este
pe toată suprafaţa corpului, cu excepţia zonelor genitale, buzelor şi canalului
auricular, fiind mai dense la nivelul feţei palmare a mâinii şi la nivelul frunţii.
(Cârsteanu şi Dumitru, 1976)
5

Funcţia de excretie a pielii
3.1.2 G landele apocrine
Glandele apocrine sunt considerate ca fiind mai primitive decât cele ecrine. La om
glandele apocrine produc mirosul pielii specific individului. Sunt localizate la nivelul
axilei şi zonelor genitale şi devin active la pubertate. În viaţa fetală sunt răspândite că
şi glandele eccrine pe toată suprafaţa corpului însă din luna a 5 a de sarcină dispar, cu
excepţia unui număr mic, care sunt înâlnite în zonele în care dezvoltarea lor este
maximă în viaţa adultă (Cârsteanu si Dumitru, 1976).
Sunt formate dintr-o porţiune secretorie şi un duct, care se deschide în foliculul
pilos. Porţiunea glandulară constă dintr-un rând de celule voluminoase, cuboidale,
sprijinite pe o membrană bazală, printre ele fiind celule mioepiteliale asemănătoare
celor din glanda ecrină. Celulele secretorii conţin un aparat Golgi dezvoltat, reticul
endoplasmatic granular dispus frecvent în jurul mitocondriilor, ribozomi, granule de
secreţie şi o cantitate de glicogen mai mică decât în celulele secretorii ecrine.
Granulele sunt de două tipuri: întunecate şi clare. Granulele de secreţie sunt cele
întunecate, cu conţinut electron-dens lamelar şi granular şi sunt mai mari decât cele
din celulele ecrine întunecate. Granulele clare sunt uniforme ca dimensiune, cu puţin
material şi au în partea bazală a celulei resturi de criste mitocondriale.
Celulele conductului glandelor apocrine conţin mai puţine mitocondrii decât în
celulele ductului ecrin. Conductul glandelor apocrine se deschide la nivelul foliculului
pilos.
Secreţia apocrină este compusă din lipide. Secreţia apocrină inodoră este
descompusa rapid la suprafaţa pielii de flora bacteriană, dând naştere unor produşi
odoranţi. (Păiş, 1983)
Spre deosebire de urină, sudoarea este un produs de secreţie şi nu ultrafiltrat al
plasmei sangvine. La nivelului tubului excretor are loc un proces de reabsorbţie a Na
şi Cl, ajungându-se în final la un lichid hipoton faţă de plasmă. Prin sudoare se
excreta mari cantităţi de apă (92,2%) şi diferite substanţe anorganice şi organice
(cloruri, bicarbonaţi, fosfaţi, amoniac, uree, aminoacizi, acizi graşi volatili, acidi
lactic, unele vitamine). Activitatea glandelor sudoripare e controlată prin mecanisme
nervoase şi umorale. Stimularea lor este adrenergică atât prin adrenalină şi
noradrenalina circulantă cât şi prin mediatorii adrenergici ai sistemului nervos
6

Funcţia de excretie a pielii
ganglionar simpatic. (Strungaru şi colab., 1983)
Nr.1 Distribuţia glandelor sudoripare
3.2 Sebumul
Glandele sebacee sunt ataşate foliculului pilos, avand funcţia de secreţie a
sebumului.
La periferia unui acin glandular se găsesc celule nediferenţiate dispuse pe o
membrană bazală. Ele conţin un nucleu heterocromatinic sau eucromatinic şi în
citoplasma se găsesc numeroşi ribozomi, reticul endoplasmatic neted şi granular,
complex Golgi mic, mitocondrii pleomorfe, adeseroi cu criste perpendiculare pe axa
lungă , mănunchiuri de filamente similare celor din celulele epidermice nediferenţiate,
granule de glicogen. Celulele sunt conectate prin desmozomi şi joncţiuni înguste. La
începutul maturării, celulele care se diferenţiază au complex Golgi dezvoltat,
numeroşi ribozomi, reticul endoplasmatic neted, particule de glicogen şi un număr
mic de vezicule cu sebum de mărime diferită. Tubulii reticulului endoplamatic sunt
adesea conectaţi cu membranele Golgi în jurul nucleului. Odată cu acumularea
vacuolelor lipidice în citoplasma, celulele se măresc în volum, zonele Golgi ca şi
reticulul endoplasmatic neted, apar clar legate de veziculele cu conţinut lipidic
moderat electron-dens. Lipidele se formează în reticulul enoplasmatic neted şi agregă
ca vacuole în zona Golgi.
7

Funcţia de excretie a pielii
Tonofilamentele, mai numeroase în celulele periferice nediferenţiate, scad
cantitativ în celulele parţial diferenţiate şi sunt greu de identificat în celulele mature
pline cu sebum spre interiorul acinului glandular. În toate stadiile maturării, celulele
conţin lizozomi cu matrice omogenă. Celulele mature sunt mari şi au numeroase
vacuole de sebum cu dimensiuni relativ uniforme. Vacuolele de sebum conţin o
substanţă densă, scualen şi lipide săturate. Când celulele suferă ruptură şi îşi varsă
conţinutul, nucleul degenerează.
Glandele sebacee sunt conectate la foliculul polisebaceu printr-un duct format de
celule epiteliale spinoase. Porţiunea ductului din apropierea acinilor produce atât
keratina cât şi sebum.
Secreţia sebacee este controlată hormonal în special de către hormonii androgeni.
Ea conţine trigliceride, colesterol, acizi graşi liberi, scualen, ceride, etc. Sebumul
protejează pielea împotriva agenţilor infecţioşi şi realizează un control asupra
pierderilor de apă. De asemenea, atunci când muşchiul ecrector al părului se contracta
şi părul devine vertical, la frig în special, se elimină o mică cantitate de sebum care
împiedică pierderea căldurii. (Păiş, 1983)
Nr.2 Folicul pilos şi glandele sale
4. Tegumentul e o barieră osmotică
Animelele reduc fluxul apei prin suprafața corpului, controlând numărul proteinelor
8

Funcţia de excretie a pielii
acvaporine în membrana plasmei. Fiecare acvaporină permite pătrunderea a peste un
miliard de molecule de apă pe secundă. O celulă epitelială cu acvaporine poate fi de
100 de ori mai permeabilă la apă decât o celulă fără acvaporine. Nivelul de
acvaporine în membrana plasmei depinde de exprimarea genelor acvaporine și de
căile de trafic intracelular care controlează transferul de acvaporine dintre vezicule și
membrana plasmei.
Unele animale atenuează pierderea apei prin acoperirea suprafețelor externe cu un
strat subțire de celule hidrofobice. Mucusul – o secreție extracelulară de
mucopolizaharide, lipide și proteine – este un exemplu de barieră hidrofobică.
Straturile de mucus de pe suprafața plamânilor și a tractului gastrointestinal
diminuează pierderea apei prin epiteliu. O varietate de animale semiacvatice, precum
broaștele folosesc mucusul pentru prevenirea pierderii apei și pentru a menține pielea
hidratată. Stratul gros de mucus al unui pește hibernant cu respirație aeriană se uscă
pentru a forma un strat impermeabil care previne deshidratarea animalelor în sezonul
estival, adică în perioada somnului de vară. Mucusul de suprafață reduce de asemenea
dificultatea osmoreglării prin captarea unui strat de apă între animal și mediu. Acest
strat de apă este un micro-compartiment care acționeaă ca o zonă tampon osmotică și
ionică.
Animalele de uscat folosesc adaptări mai elaborate în structura epitelială pentru a
preveni pierderea apei prin piele. Keratinocitele pielii amfibienilor tereștri și
amniotelor secretă proteine și lipide modificate pentru a forma o bază extracelulară
densă și hidrofobică. Amniotele, dar nu și amfibienii, au un strat suplimentar peste
keratocite. Acest strat, numit stratum corneum, este format din keratinocite, care se
diferenţiază pentru a forma corneocita. Apoi celula produce un strat complex de
proteine, numit înveliș cornos, care până la urmă înlocuiește membrana plasmei
corneocitei. În timpul cornificării, celulele corneocite sunt „programate” să moară,
ceea ce rămâne fiind rețeaua de keratină înconjurată de învelișul cornos. Proteinele
extracelulare de bază colectează aceste resturi de celule în grămezi de molecule de
lipide numite membrană lamelară. După ce s-a format, această mixtură de proteine și
lipide este supusă unei serii de procese enzimatice și chimice care o transformă în
stratum corneum. Deși țesutul este mort, acesta rămâne sensibil la schimbări fizice,
9

Funcţia de excretie a pielii
declanșând secreția de proteine, lipide și factori semnalizatori din keratinocitele din
stratul inferior.
Nr.3 Stratum corneum
Solzii reptilelor și ai păsărilor sunt formate din bucăți de stratum corneum
interconectate (în mare parte keratină). Pielea mamiferelor este de asemenea
keratinizată, deși puține mamifere mai au „slozii” vechi, precum învelișul cozii unei
rozătoare sau cochilia unui tatu. Modificările la nivelul stratum corneum cheratinizat
oferă vertebratelor terestre alte tipuri de structuri, cum ar fi solzii de pe burta șerpilor
folosiți pentru locomoție sau țepii protectori ai șopârlelor de deșert. Totuși, toate
tetrapodele depind de stratum corneum cheratinizat pentru a preveni deshidratarea.
Cuticula insectelor este o rețea complexă de molecule hidrofobe care acoperă toate
suprafețele externe ale insectelor, inclusiv pe cele ale traheei și ale intestinelor.
Componenta structurală principală a cuticulei este polizaharidul chitin. Acesta e
sintetizat în celulele epidermelor și apoi transportat în spațiul extracelular unde este
modificat chimic, cristalizat și combinat cu alte proteine și polizaharide pentru a
obține proprietățile fizice potrivite. Cuticula matură are foarte puțină permeabilitate la
apă, aceasta fiind suficient de rigidă pentru a se comporta ca un schelet extern pentru
animal.
Atât tecile tetrapodelor cât și cele ale insectelor terestre sunt înzestrate cu un strat
10

Funcţia de excretie a pielii
adițional de lipide care reduce pierderea apei prin evaporare. Celulele epidermelor
secretă aceste lipide care apoi formează un strat continuu cu rol etanș. Păsările și
mamiferele posedă un strat subțire de glicolipide care acoperă stratum corneum și
umple golurile dintre celule. Exoscheletul insectelor are de asemenea o suprafață care
acoperă în lanț acizii grași și esterii de ceară. De fapt, acest strat subțire de lipide oferă
rezistență exoscheletului insectei la permeabilitatea apei. Capacitatea straturilor de
lipide de a limita mișcarea apei depinde de interacțiunea dintre moleculele de lipide,
creând o barieră hidrofobică care oprește apa.
Aceleași procese au loc și la nivelul altui țesut epitelial: suprafața respiratoare.
Cantitatea de apă pierdută la nivel respirator depinde de trăsăturile structurale ale tecii
exterioare dar și de alți factori. De exemplu, un animal care respiră prin plămâni, cu o
rată ridicată a metabolismului va avea o rată a ventilației mai ridicată și prin urmare o
pierdere mai mare de apă (prin respirație) decât un animal cu o rată mai mică a
metabolismului. Multe animale, în special cele de deșert, sunt adaptate anatomic
pentru a reduce pierderea apei prin respirație.
Țesuturile epiteliale au în comun patru proprietăți specializate care afectează
mișcarea ionilor. Proprietățile țesuturilor epiteliale depind atât de proprietățile
transportului celulelor epiteliale individuale, cât și de modului în care celulele sunt
interconectate pentru a forma țesutul.
A) Funcția celulei epiteliale depinde de distribuția asimetrică a transportorilor
prin celulă. Membrana apicală a celulei, expusă mediului extern, are un profil
de proteine diferit de cel al membranei bazilaterale a celulei, care este
orientată spre interior. Această topografie a celulei ia naștere deoarece celulele
introduc proteine în locurile corespunzătoare și limitează mișcarea lor în
stratul dublu de lipide.O dată ce au ajuns în membrană, acestea sunt ancorate
în poziție fiind atașate de schelet.
B) Celulele epiteliale sunt interconectate prin legături de proteine care transformă
grupul de celule într-un strat de țesut impermeabil. Astfel, se formează
încrucișări strânse atunci când proteine din membrana unei celule se
conectează la proteine specifice dintr-o celulă adiacentă. Interacțiunea dintre
celulele adiacente limitează mișcarea substanței dizolvate și a apei în jurul
11

Funcţia de excretie a pielii
celulelor. Aceste conexiuni intercelulare creează de asemenea un fel de centură
de proteine în jurul circumferinței celulei epiteliale, restricționând mișcarea
liberă a proteinelor din membrană între regiunile apicale și bazilaterale ale
membranei pentru a menține topografia celulară.
C) Țesuturile epiteliale sunt formate din multe tipuri de celule. Această diversitate
este mai accentuată în sistemul digestiv. Totuși, chiar și țesuturile relativ
simple, de exemplu cel al branhiilor la pești, sunt compuse din mai multe
tipuri de celule, fiecare cu roluri importante, cum ar fi furnizarea unor
capacități de transport specifice sau sprijin structual.
D) Transportul de ioni necesită multă energie. Majoritatea celulelor epiteliale cu
rol major în transport posedă mitocondrii în abundență pentru a produce ATP.
În unele cazuri, mitocondriile sunt foarte apropiate de regiunile membranei
plasmei care dirijează procesele de transport. În alte cazuri, proteinele motor și
citoscheletul transportă activ mitocondriile spre aceste regiuni când
metabolismul cere o creștere. Eforturile energetice ale transportului ionilor pot
fi răspunzătoare pentru aproape jumătate din rata metabolică a țesutului.
(Moyes si Schulte, 2006)
5. Transport paracelular şi transcelular
Deși țesuturile epiteliale transportă anumite substanțe dizolvate în scopuri proprii,
cele mai multe procese de transport servesc transferului substanțelor dizolvate dintr-o
parte a țesutului în cealaltă. Țesuturile epiteliale folosesc două căi principale de
transport prin celulă. Tansportul transcelular este mișcarea substanțelor dizolvate (sau
a apei) prin celulele epiteliale. De exemplu, substanțele dizolvate se pot dispersa din
ECF (fluidul extracelular) care îmbăiază celulele, mișcându-se spre membrana
bazilaterală, prin citoplasm și prin membrana apicală ajungând în mediul extern (fie
apa liberă, sau lumenul unui organ care comunică cu meiul extern). Invers, mișcarea
substanțelor dizolvate (sau a apei) între celule adiacente se numește transport
paracelular. De exemplu, moleculele se împrăștie din sânge, prin ECF și intră în
12

Funcţia de excretie a pielii
spațiile înguste ale fluidului interstițial dintre celulele adiacente. De aici, moleculele
trec prin îmbinările înguste care conectează celulele epiteliale în fâșii. Celulele vecine
pot secreta molecule în spațiul interstițial pentru a-i controla caracterul, astfel
creându-se gradienți care direcționează mișcările paracelulare. În timp ce îmbinările
înguste pot împiedica moleculele mari să treacă, moleculele mici (apă, ioni) pot trece
prin legăturile de proteine. Țesuturile care permit transportul paracelular sunt
frecvent numite legături lejere. Țesuturile care dirijează transport paracelular minim
se numesc legături strânse. Țesuturile epiteliale conțin șiruri de transportori, inclusiv
următorii transportori implicați frecvent în echilibrul apei și al ionilor.
Na+/K+ ATP-ază este esențial în mișcarea ionilor, folosind energia hidrolizei
ATP pentru a exporta 3Na+ în schimbul importului a 2K+. Unele țesuturi
folosesc o H+ ATP-ază pentru a pompa protoni pentru a schimba pH-ul, o
forță conducătoare pentru alte procese de transport.
Diferite canale de ioni (Cl¯, K+ și Na+) se pot deschide sau închide ca reacție
la semnale mecanice, electrice sau chimice, pentru a permite ionilor specifici
să curgă pe gradienți electrochimici.
Co-transportorii electroneutri duc atât anioni cât și cationi în aceași direcție ca
reacție la gradienții electrochimici. Există cotransportori Na+ -K+ -2Cl¯
(NKCC) și cotransportori K+ -Cl¯.
Diferiți schimbători electroneutri sunt transportori reversibili conduși de gradienții
electrochimici, inclusiv de pH. Există schimbători Na+/H+ (NHE), NH4+/H+, și
Cl¯/HCO3¯. (Moyes si Schulte, 2006)
13

Funcţia de excretie a pielii
Nr.4 Tipuri de transport
6. Branhiile peştilor transportă ioni în şi din apă
Lamelele branhiilor sunt compuse din celule de clorură bogate în mitocondrii
dispersate între două tipuri de celule, una bogată în mitocondrii, iar cealaltă cu
conținut scăzut în mitocondrii.
Direcția transportului ionilor și a apei depinde de salinitatea apei. Branhiile unui
pește de apă dulce are nevoie de Na+, Ca+ și alți ioni din apă, adesea împotriva
gradienților electrochimici înclinați. Spre deosebire de peștii de apă dulce, peștii
marini trebuie să evite acumularea excesivă de ioni și să limiteze pierderea apei.
Branhiile sunt foarte importante în echilibrarea ionilor, iar celulele de clorură în
special, sunt decisive în excreția ionilor.
Responsabilitățile celulelor care pompează ioni din branhiile peștilor se schimbă
în funcție de condițiile exterioare. Unele specii sunt diadromi, migrând între ape
sărate și ape dulci. Peștii catadromi precum țiparii, migrează spre ape sărate pentru a
se înmulți. Pe de altă parte, peștii anadromi, precum somonul, migrează din apa sărată
spre apa dulce pentru a se reproduce. Somonii tineri cresc în apa dulce apoi migrează
spre mare. Înaintea migrării, branhiile acestor pești suferă o reorganizare celulară
impresionantă deoarece proprietățile celulelor branhiilor care pompează ioni se
pregătesc pentru noul mediu. În mod bizar, procesul de remodelare, numit
„smoltification”(proces metabolic intern, prin care peştele se adaptează de la apă
dulce la apă sărată pentru a evita stresul) este declanșat de numeroși hormoni.
Schimbările osmoregulatoare sunt în mare parte mediate de hormonii de creștere,
cortisol, și într-o mai mică măsură de hormonul tiroidian. „Smoltification” duce de
asemenea la remodelarea altor țesuturi implicate în echilibrarea ionilor și apei,
inclusiv tractul gastro-intestinal și probabil rinichii. Ca urmare a schimbărilor de la
nivelul celulelor, somonii suferă schimbări osmotice și ionice minore la intrarea în apa
de mare. (Moyes si Schulte, 2006)
14

Funcţia de excretie a pielii
7.Reptilele şi păsările sunt înzestrate cu glande de sare
Apa dulce are o concentrație foarte mică de substanțe dizolvate, de aceea creează o
presiune interioară care ajută la direcționarea absorbției apei. Totuși, puține specii de
animale beau apă de mare. Asta înseamnă două lucruri: în primul rând, moleculele de
apă trebuie să fie transportate selectiv prin intestin contra gradientului osmotic.
Probabil transportul transcelular prin epiteliul compact/strâns este important la aceste
animale. În al doilea rând, animalele trebuie să fie capabile să elimine sarea conținută
în apa de mare consumată cu hrana. Multe reptile și păsări au o glandă de sare care
ajută echilibrarea ionilor și a apei excretând soluții de Na+ și Cl¯ hiper osmotice. Fie
că trăiesc în ocean sau în deșert, speciile care au această glandă pot face față lipsei de
apă dulce, consumând apă de mare hipertonică sau chiar numai din mâncare. La
păsări, glanda de sare se găsește într-o adâncitură de la baza ciocului, iar secrețiile
acesteia se scurg printr-un canal care străbate ciocul și are deschidere la nări. Secreția
glandei de sare nazală poate fi de până la trei ori mai concentrată decât plasma. Prin
urmare, dacă o pasăre bea 30 ml de apă de mare, aceasta poate excreta toată sarea în
10 ml de secreție a glandei de sare, câștigând 20 de ml de apă pură. Glandele de sare
pot să facă asta forțând energia metabolică să pompeze ioni, ajutată de un sistem de
contra-curent asemănător cu cel al rinichiilor.
Glanda de sare e formată dintr-o serie de tubulețe secretoare care extrag sarea din
sânge. Fiecare tubuleț secretor este un orificiu inchis care se deplasează în paralel
spre o rețea capilară, sângele curgând de la baza tubului până la celălalt capăt inchis.
Secrețiile curg de la capătul închis până la cel deschis, constituind un schimbător
contra-curent care permite tubului să concentreze soluțiile mult mai ușor. Multe
tuburi sunt adunate pentru a forma lobi care se scurg în canalul central care transportă
secreția spre nări. Celulele epiteliale care căptușesc tubul secretor extrag sarea din
fluidul intestinal aflat între celulele tuburilor și sânge. (Moyes si Schulte, 2006)
15

Funcţia de excretie a pielii
8.Glandele rectale ale elasmobranhiilor excretă Na+ și Cl¯
reţinând ureea
Ca păsările de mare, elasmobranhiile au un organ excretor suplimentar care ajută la
excreția sării. Glanda rectală este compusă din mai multe tuburi înconjurate de
capilare. Celulele din fiecare tubuleț transportă activ Na+ și Cl din sânge, apoi secretă
acești ioni în lumenul tubular. Invaginarea bazilaterală în aceste celule, semănătoare
cu microviliile, măresc suprafața pentru schimbul de ioni cu sângele.
Secreția de sare e stimulată de o creștere indusă hormonal în activitatea canalului
Cl¯. (Moyes si Schulte, 2006)
16

Funcţia de excretie a pielii
9.Bibliografie
Moyes,Christopher., Schulte, Patricia.: Principles of Animal Physiology , Editura
Pearson, 2006.
Strungaru, Gr., Pop, M., Hefco, V.: Fiziologie animală, Editura Didactica si
Pedagogica, Bucuresti, 1983.
Păiş, V.: Ultrastructura pielii umane, Editura Medicală,. Bucureşti, 1983.
Roşca, D.: Fiziologie animală, Editura Didactică şi Pedagogică, Bucureşti, 1977.
Cârsteanu, M., Dumitru, R.: Biochimia pielii, Editura Medicală, Bucureşti, 1976.
Nr.1 http://hiperhidroza.com/wp-content/uploads/2008/09/distributie-glande.jpg
Nr.2 http://hiperhidroza.com/wp-content/uploads/2008/09/Glande-sudoriparestructur
%C4%83-simplu.jpg
Nr.3 http://img.medscape.com/fullsize/migrated/564/078/bjd564078.fig1.gif
Nr.4http://www.nanoanalytics.com/en/hardwareproducts/cellzscope/howitworks/
chapter01/cellzscope0301.gif
17