
curs5_neurobio
12
Cursul 5 1 Transducţia senzorială Senzaţia şi percepţia mediului înconjurător şi răspunsul organismului sunt realizate prin intermediul sistemelor senzoriale. Aceste sisteme au în comun 3 elemente: Stimulul; Transducţia stimulului în impuls nervos ce produce o experienţă subiectivă; Răspunsul. Stimulul prezintă mai multe proprietăţi, printre care: durată, localizare, intensitate, modul de acţiune, toate acestea fiind corelate cantitativ cu senzaţia trăită. Modalităţile de stimulare sunt următoarele: lumină, sunet, atingere, gust şi miros şi fiecare dintre acestea prezintă mai multe calităţi. Activarea mecanoreceptorilor Mecanoreceptorii transformă un stimul fizic în impuls electric astfel încât organismul este capabil să localizeze stimulul şi să perceapă intensitatea şi durata sa. La oameni, acest lucru este îndeplinit prin intermediul mecanoreceptorilor: corpusculii Meissner, Ruffini, Pacini. Mecanoreceptorii vertebratelor sunt relativ inaccesibili pentru studiu, dar s-au facut progrese prin studiul lui Caenorhabditis elegans. La C. elegans cuticula externă este cuplată la neuronii senzitivi printr-un înveliş. Acest înveliş este ataşat printr-un sistem de prindere la domeniul extracelular al unui canal ionic. Când este atinsă cuticula, învelişul este denaturant şi astfel se deschid canalele membranare cuplate la microtubulii intracelulari (fig. 5-1). Fig. 5-1 – Model molecular al acţiunii mecanoreceptorilor.
-
Upload
ralu-salajan -
Category
Documents
-
view
9 -
download
0
description
neuro
Transcript of curs5_neurobio

Cursul 5
1
Transducţia senzorială
Senzaţia şi percepţia mediului înconjurător şi răspunsul organismului sunt realizate prin intermediul sistemelor senzoriale. Aceste sisteme au în comun 3 elemente:
Stimulul; Transducţia stimulului în impuls nervos ce produce o experienţă
subiectivă; Răspunsul.
Stimulul prezintă mai multe proprietăţi, printre care: durată, localizare, intensitate, modul de acţiune, toate acestea fiind corelate cantitativ cu senzaţia trăită. Modalităţile de stimulare sunt următoarele: lumină, sunet, atingere, gust şi miros şi fiecare dintre acestea prezintă mai multe calităţi.
Activarea mecanoreceptorilor Mecanoreceptorii transformă un stimul fizic în impuls electric astfel încât
organismul este capabil să localizeze stimulul şi să perceapă intensitatea şi durata sa. La oameni, acest lucru este îndeplinit prin intermediul mecanoreceptorilor: corpusculii Meissner, Ruffini, Pacini. Mecanoreceptorii vertebratelor sunt relativ inaccesibili pentru studiu, dar s-au facut progrese prin studiul lui Caenorhabditis elegans. La C. elegans cuticula externă este cuplată la neuronii senzitivi printr-un înveliş. Acest înveliş este ataşat printr-un sistem de prindere la domeniul extracelular al unui canal ionic. Când este atinsă cuticula, învelişul este denaturant şi astfel se deschid canalele membranare cuplate la microtubulii intracelulari (fig. 5-1).
Fig. 5-1 – Model molecular al acţiunii mecanoreceptorilor.

Cursul 5
2
Mecanismul include un canal ionic ce permite un influx de Na+ şi K+ în celulă, producând depolarizarea membranei celulare şi apariţia potenţialului de receptor.
Odată cu apariţia potenţialului de acţiune, acesta se propagă de-a lungul fibrei nervoase către centrul sistemului nervos. Multe din aceste fibre sunt mielinizate, mărind astfel rata de transmisie a impulsului nervos către SNC. Corpul celular al neuronului se află în ganglionul din rădăcina posterioară a nervului spinal. Astfel, rădăcina posterioară a ganglionului prezintă 2 ramificaţii axonale, una care se proiectează la periferie, şi alta care se proiectează la nivelul SNC (fig. 5-2).
Fig. 5-2 – Morfologia unui ganglion al rădăcinii posterioare.
Potenţialul de receptor (potenţial generator) reprezintă modificarea potenţialului de membrană al receptorului determinată de interacţiunea dintre stimulul specific şi organul receptor;
Caracteristici: • se răspândeşte electrotonic în zonele învecinate, • răspunsul poate fi gradat, • poate rămâne localizat sau, când amplitudinea sa depăşeşte pragul poate iniţia un potenţial de acţiune, • cu cât potenţialul de receptor este mai mare decât nivelul prag, cu atât este mai mare frecvenţa de descărcare a potenţialelor de acţiune.
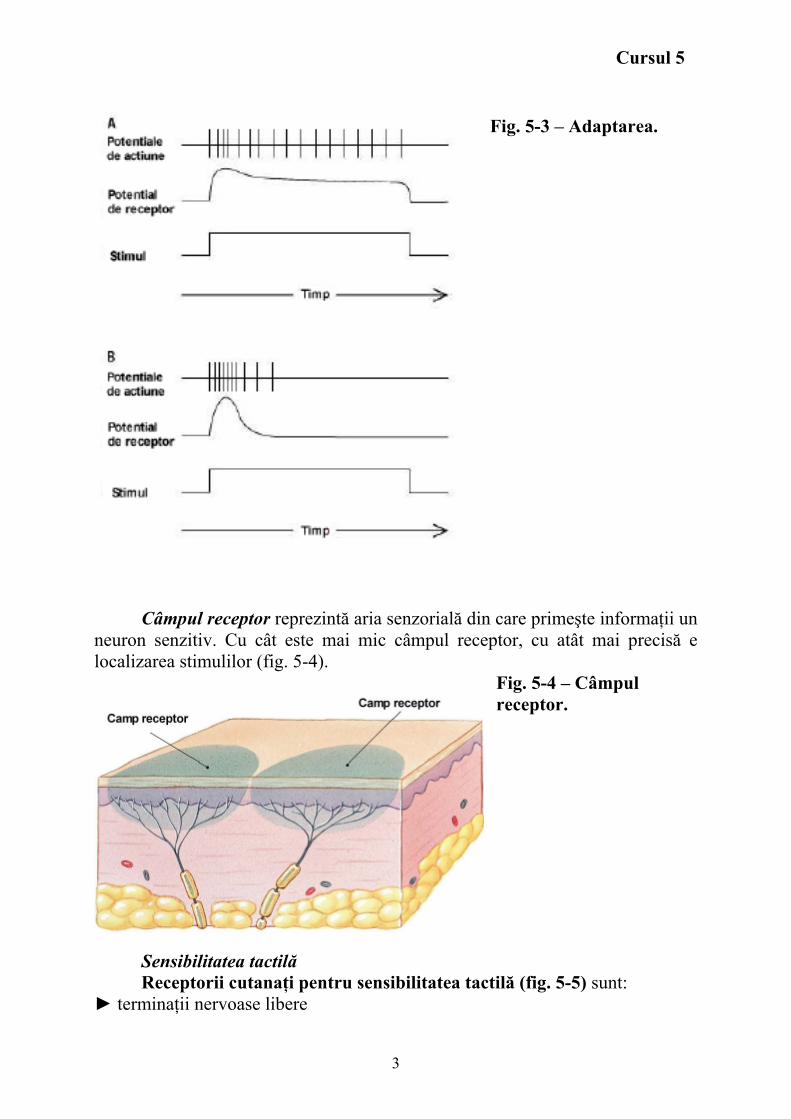
Adaptarea Aplicând asupra unui receptor un stimul stabil de intensitate constantă,
frecevenţa potenţialelor de acţiune în fibra nervoasă senzitivă scade cu timpul. A. receptori cu adaptare lentă (tonici) (fig. 5-3 A), B. receptori cu adaptare rapidă (fazici) (fig. 5-3 B).
Cursul 5
3
Fig. 5-3 – Adaptarea.
Câmpul receptor reprezintă aria senzorială din care primeşte informaţii un
neuron senzitiv. Cu cât este mai mic câmpul receptor, cu atât mai precisă e localizarea stimulilor (fig. 5-4).
Fig. 5-4 – Câmpul receptor.
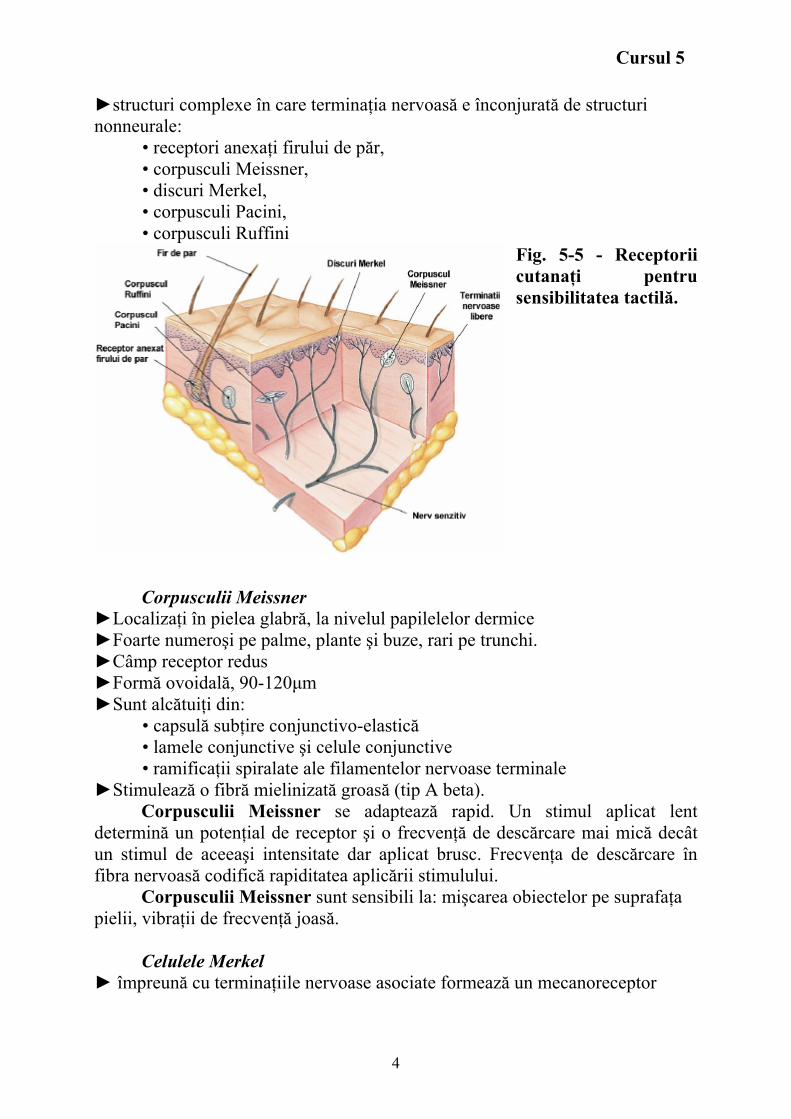
Sensibilitatea tactilă Receptorii cutanaţi pentru sensibilitatea tactilă (fig. 5-5) sunt:
► terminaţii nervoase libere
Cursul 5
4
►structuri complexe în care terminaţia nervoasă e înconjurată de structuri nonneurale:
• receptori anexaţi firului de păr, • corpusculi Meissner, • discuri Merkel, • corpusculi Pacini, • corpusculi Ruffini
Fig. 5-5 - Receptorii cutanaţi pentru sensibilitatea tactilă.
Corpusculii Meissner
►Localizaţi în pielea glabră, la nivelul papilelelor dermice ►Foarte numeroşi pe palme, plante şi buze, rari pe trunchi. ►Câmp receptor redus ►Formă ovoidală, 90-120μm ►Sunt alcătuiţi din:
• capsulă subţire conjunctivo-elastică • lamele conjunctive şi celule conjunctive • ramificaţii spiralate ale filamentelor nervoase terminale
►Stimulează o fibră mielinizată groasă (tip A beta). Corpusculii Meissner se adaptează rapid. Un stimul aplicat lent
determină un potenţial de receptor şi o frecvenţă de descărcare mai mică decât un stimul de aceeaşi intensitate dar aplicat brusc. Frecvenţa de descărcare în fibra nervoasă codifică rapiditatea aplicării stimulului.
Corpusculii Meissner sunt sensibili la: mişcarea obiectelor pe suprafaţa pielii, vibraţii de frecvenţă joasă.
Celulele Merkel
► împreună cu terminaţiile nervoase asociate formează un mecanoreceptor

Cursul 5
5
► prezintă markeri epiteliali dar şi anumite caracteristici de celulă nervoasă (proteina S 100, enolaza neuron-specifică, proteina neurofilamentului, cromogranină, sinaptolizină) ► prezintă vezicule neurosecretorii ce conţin neuropeptide, prezente în toată citoplasma, mai dense în regiunea din apropierea fibrelor nervoase ► sunt capabile să producă majoritatea neuromediatorilor identificaţi în piele ► exprimă receptori pentru numeroşi neuromediatori.
Deformarea mecanică a celulelor Merkel activează canale de Ca de tip ICa (calcium sustained current) generând un influx de Ca în celulă ce determină eliberarea de neurotransmiţători din celule ce acţionează asupra terminaţiilor nervoase.
Discurile Merkel transmit un impuls iniţial puternic, dar care se adaptează parţial şi apoi un impuls mai slab, continuu, ce se adaptează lent, fiind responsabile de furnizarea unor impulsuri stabile ce permit perceperea atingerii continue. Discurile Merkel sunt receptori cu câmp receptor îngust şi au un rol important în localizarea senzaţiilor tactile.
Corpusculii Pacini
• foarte numeroşi în hipodermul palmelor şi plantelor • voluminoşi - lungime aprox 1 mm; câmp receptor larg • formă eliptică
Sunt alcătuiţi din:
►capsulă formată din 20-60 lamele concentrice cu aspect asemănător foiţelor de ceapă; ►fibră nervoasă cu diametru de aproximativ 2μm care se termină printr-o umflătură în contact cu celulele lamelare centrale; capătul fibrei e nemielinizat, dar cu puţin înainte de a părăsi corpusculul fibra devine mielinizată; primul nod Ranvier e situat de asemenea în interiorul corpusculului.
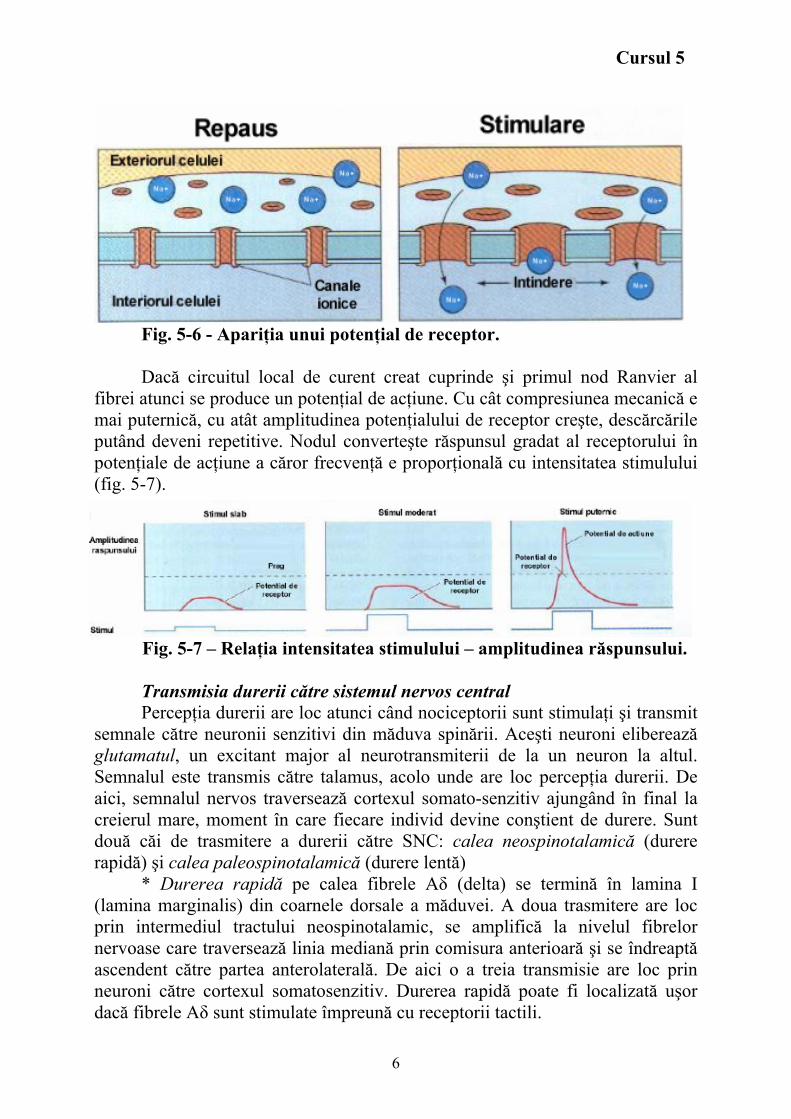
Stimul mecanic deformează lamela externă a capsulei => deformarea
transmisă la terminaţia nervoasă. Deformarea terminaţiei nervoase receptoare => deschiderea unor canale ionice şi creşterea conductanţei pentru Na+ (şi alţi ioni K +,Cl - sunt implicaţi) => modificare bruscă a potenţialului membranar, determinând apariţia unui potenţial de receptor (fig. 5-6).
Cursul 5
6
Fig. 5-6 - Apariţia unui potenţial de receptor. Dacă circuitul local de curent creat cuprinde şi primul nod Ranvier al
fibrei atunci se produce un potenţial de acţiune. Cu cât compresiunea mecanică e mai puternică, cu atât amplitudinea potenţialului de receptor creşte, descărcările putând deveni repetitive. Nodul converteşte răspunsul gradat al receptorului în potenţiale de acţiune a căror frecvenţă e proporţională cu intensitatea stimulului (fig. 5-7).
Fig. 5-7 – Relaţia intensitatea stimulului – amplitudinea răspunsului. Transmisia durerii către sistemul nervos central Percepţia durerii are loc atunci când nociceptorii sunt stimulaţi şi transmit
semnale către neuronii senzitivi din măduva spinării. Aceşti neuroni eliberează glutamatul, un excitant major al neurotransmiterii de la un neuron la altul. Semnalul este transmis către talamus, acolo unde are loc percepţia durerii. De aici, semnalul nervos traversează cortexul somato-senzitiv ajungând în final la creierul mare, moment în care fiecare individ devine conştient de durere. Sunt două căi de trasmitere a durerii către SNC: calea neospinotalamică (durere rapidă) şi calea paleospinotalamică (durere lentă)
* Durerea rapidă pe calea fibrele Aδ (delta) se termină în lamina I (lamina marginalis) din coarnele dorsale a măduvei. A doua trasmitere are loc prin intermediul tractului neospinotalamic, se amplifică la nivelul fibrelor nervoase care traversează linia mediană prin comisura anterioară şi se îndreaptă ascendent către partea anterolaterală. De aici o a treia transmisie are loc prin neuroni către cortexul somatosenzitiv. Durerea rapidă poate fi localizată uşor dacă fibrele Aδ sunt stimulate împreună cu receptorii tactili.

Cursul 5
7
* Durerea lentă este transmisă pe calea fibrele C către lamina II şi III (substanţa gelatinoasă), de aici neuronii transmiţând-o către lamina V, situată în cornul dorsal al măduvei spinării. De aici se intersectează cu fibrele durerii rapide, traversează în partea opusă via comisura anterioară, apoi traversează ascendent până în calea anterolaterală. Aceşti neuroni se termină în creier: aproximativ o zecime de fibre oprindu-se în talamus, iar restul în măduvă şi mezencefal.
Căile aferente: fibre senzitive nociceptive, somatice (fibre slab mielinizate), şi vegetative (fibre nemielinizate C), incluse în nervii cranieni.
căile ascendente ale sensibilităţii dureroase formează fasciculul spinotalamic lateral. Centrii de integrare şi percepere a durerii:
centrii talamici ( percepere inconştientă, nelocalizată însoţită de alerta generala)
sistemul limbic care coordonează componenta afectivă. centrii corticali aflaţi in lobii parietali.
Căile descendente de suprimare a durerii: substanţa cenuşie, nucleii rafeului median, substanţa reticulată a mezencefalului. Din punct de vedere biochimic în farmacologia durerii sînt implicate foarte multe substanţe: serotonina, histamina, bradikinina (întilnite la nivelul nociceptorilor mediatori algogeni), endorfine, dinorfine, enkefaline, endomorfine, (neuromediatori peptidergici).
Sensibilitatea termică
► Permite aprecierea diferenţelor relative de temperatură ► Intensitatea senzaţiei termice este proporţională cu viteza de schimb termic între obiect şi organism. ► Conductibilitatea termică a unui obiect este implicată în senzaţia produsă. ► Senzaţia termică prezintă diferite gradaţii: îngheţ, frig, răcoros, indiferent, călduţ, cald, fierbinte. ► Sunt implicate mai multe tipuri de receptori: receptori pentru cald şi pentru rece (răspund la stimuli termici inofensivi) şi receptori pentru durere (stimulaţi numai de nivelurile extreme de temperatură). ► Numărul termoreceptorilor variază de la o regiune a pielii la alta: • buze 15-20 puncte de rece pe cm2, • degete 3-5 puncte de rece pe cm2, • trunchi 1 punct de rece pe cm2.
Există de 3-10 ori mai mulţi receptori pentru rece decât pentru cald. ► Termoreceptorii răspund mult mai intens la modificările de temperatură decât la temperaturile staţionare.

Cursul 5
8
Receptorii pentru cald ► Detectează variaţii termice superioare temperaturii cutanate. ► Sunt terminaţii nervoase libere, cea mai mare parte amielinice şi în mică măsură fibre mielinizate subţiri. ► Transmit impulsurile în principal prin fibre nervoase tip C cu viteze de transmitere de 0,4-2 m/s. ► Încep să descarce impulsuri la temperaturi cutanate de peste 30oC.
Receptorii pentru rece
► Detectează variaţii termice inferioare temperaturii cutanate ► Sunt aproape în exclusivitate fibre subţiri mielinizate ► Ca receptor pentru rece a fost identificată o terminaţie nervosă subţire mielinizată de tip A delta, care se ramifică de câteva ori, vârfurile ramificaţiilor protruzionând în suprafaţa bazală a celulelor epidermale bazale. ► Transmit impulsurile prin fibre nervoase A delta cu viteze de aproximativ 20 m/s, dar senzaţia de rece poate fi transmisă şi prin fibre de tip C. ► Descarcă impulsuri la temperaturi între 15-35 oC. Receptorii pentru rece încep să descarce paradoxal la valori ale temperaturii cutanate de 43-47 oC.
Baza moleculară a termorecepţiei Termoreceptorii prezintă canale ionice din familia TRP (Transient
Receptor Potential), ce răspund la diferite praguri de temperatură. ►Canal implicat în senzaţia de rece - CMR-1 (TRPM8)
• este activat la o temperatură de aproximativ 28oC, determinând un influx de Ca2+. Acest influx creşte în magnitudine pe măsură ce temperatura scade spre 8oC. • mentolul stimulează de asemenea acest receptor.
►Canale importante pentru perceperea senzaţiei de durere-cald ♦TRPV1
• canal cationic neselectiv activat de temperaturi din categoria fierbinte. • activat de asemenea de capsaicină (aplicată pe piele determină o senzaţie de arsură).
♦TRPV2 • un omolog al acestuia care răspunde la temperaturi mai ridicate.
Transducţia vizuală
Studiul biochimic al proceselor moleculare intracelulare de la nivelul fotoreceptorilor comporta 2 nivele:
etapa nucleară – de sinteză proteică intracelulară; etapa membranoasă – de schimburi cu structurile învecinate.
La nivelul celulelor vizuale şi a celulelor epiteliului pigmentar, transformarea energiei luminoase în energie chimică este determinată de

Cursul 5
9
fotopigmenţi. Pentru a asigura vederea continuă, aceştia sunt în mod permanent degradaţi şi resintetizaţi (fig. 5-8). Sunt cunoscute 4 tipuri de pigmenţi vizuali: rodopsina, porfirodopsina, iodopsina, si cianiodopsina.
Fig. 5-8 – Fotodisocierea rodopsinei. Transducţia este procesul biologic prin care semnalul detectat de un
receptor este transmis unor proteine intracelulare şi este apoi tradus pentru a declanşa răspunsul celular. În transducţia vizuală, fotonul (semnalul) este absorbit de către rodopsină (proteina receptoare).
Membrana celulelor fotoreceptoare prezintă ca majoritatea celulelor din organism o polarizare electrică, fiind încărcată cu sarcini electrice pozitive la exterior şi negative la interior. Diferenţa de potenţial între exteriorul şi interiorul retinei poartă numele de potenţial membranar de repaus şi are valoarea de 30-40mV. Având în vedere că toate celulele excitabile din organism, inclusiv neuronii, prezintă potenţiale de membrană care variază între 50-90mV, rezultă că celulele fotoreceptoare retiniene se află într-o stare permanentă de depolarizare. Aceasta se datorează unei permeabilităţi crescute în repaus a segmentului extern al bastonaşelor pentru ionii de Na+, permeabilitate explicată prin existenţa unui număr mare de canale de Na+ deschise permanent în starea de repaus. Astfel la întuneric (repaus) există un intens flux transmembranar de Na+ către interiorul celulei; excesul de ioni de Na+ pătrunşi în interiorul bastonaşului este eliminat în mediul extracelular datorită activităţii unei pompe de Na+.
ATP-aza Na-K dependentă din membrana segmentului intern stabileşte şi menţine gradientele de Na+ şi K+. Ieşirea K+ din celulă va determina un potenţial de 20mV, negativ în interior. Stimularea luminoasă a celulei cu bastonaş determină închiderea rapidă a canalelor pentru Na+, care face ca potenţialul electric al celulei cu bastonaş să devină mai electronegativ, determinând

Cursul 5
10
hiperpolarizarea membranei. Ca rezultat se eliberează neurotransmiţătorul în fanta sinaptică dintre celula cu bastonaş şi celula bipolară, declanşând impulsul nervos. Astfel, paradoxal, la întuneric celulele cu bastonaş sunt depolarizate şi eliberează neurotransmiţătorul, iar la lumină are loc o hiperpolarizare a membranei acestuia şi inhibarea neurotransmiţătorului la nivelul sinapsei.
A fost destul de dificil de explicat mecanismul cuplării fotochimice şi electrice prin care se ajunge de la modificările conformaţionale suferite de rodopsină sub acţiunea luminii, la închiderea canalelor de Na+ şi astfel la hiperpolarizarea relativă. Aceasta se realizează prin menţinerea canalelor de Na+ deschise în repaus datorită unei concentraţii intracelulare mari de GMPc. Prin activarea fosfodiesterazei specifice (PDE), GMPc este transformat în 5’-GMP; scăderea concentraţiei intracelulare a GMPc are drept consecinţă închiderea canalelor de Na+ de la nivelul segmentului extern. Astfel, în repaus (la întuneric), concentraţia intracelulară a GMPc este neobişnuit de mare, menţinând deschise majoritatea canalelor de Na+.
Prin această cascadă de reacţii, canalele de Na+ din membrana plasmatică, menţinute deschise iniţial prin legarea GMPc, se vor închide rapid ca urmare a scăderii nivelului intracelular al acestuia. Amplificarea aestei cascade este potenţată de către rodopsina activată, care la rândul ei activează câteva sute de molecule de transducină şi de PDE, hidrolizând mii de molecule de GMPc pe secundă. Complexul transducină activă-αGTP se fixează pe o PDE specifică pentru GMPc. Această PDE are la rândul ei 3 subunităţi proteice, lanţurile α, β, şi γ, inegale ca mărime.
Canalele de Na+ sunt de asemenea permeabile şi pentru ionii de Ca2+, astfel încât la închiderea lor influxul de Ca2+ este inhibat, determinând scăderea acestuia la nivel intracitoplasmatic. Scăderea Ca2+ stimulează guanilatciclaza să producă GMPc, aducând rapid celulele în starea în care se aflau înainte de excitaţia luminoasă.
PDE rămâne activă atâta timp cât subunităţile gama rămân disociate de subunităţile alfa şi beta. Dar însăşi transducina alfa este o GTP-ază ce hidrolizează lent GTP-ul care o activează. Hidroliza GTP la GDP eliberează subunitatea gama, care se poate lega ulterior inactivând PDE.
Pe de altă parte se cunoaşte existenţa unor mecanisme compensatoare de blocare a sistemului de hidroliză al GMPc, cel mai important fiind legat de acţiunea combinată a două protein-enzime specifice, rodopsinkinaza şi arestina.
Cursul 5
11
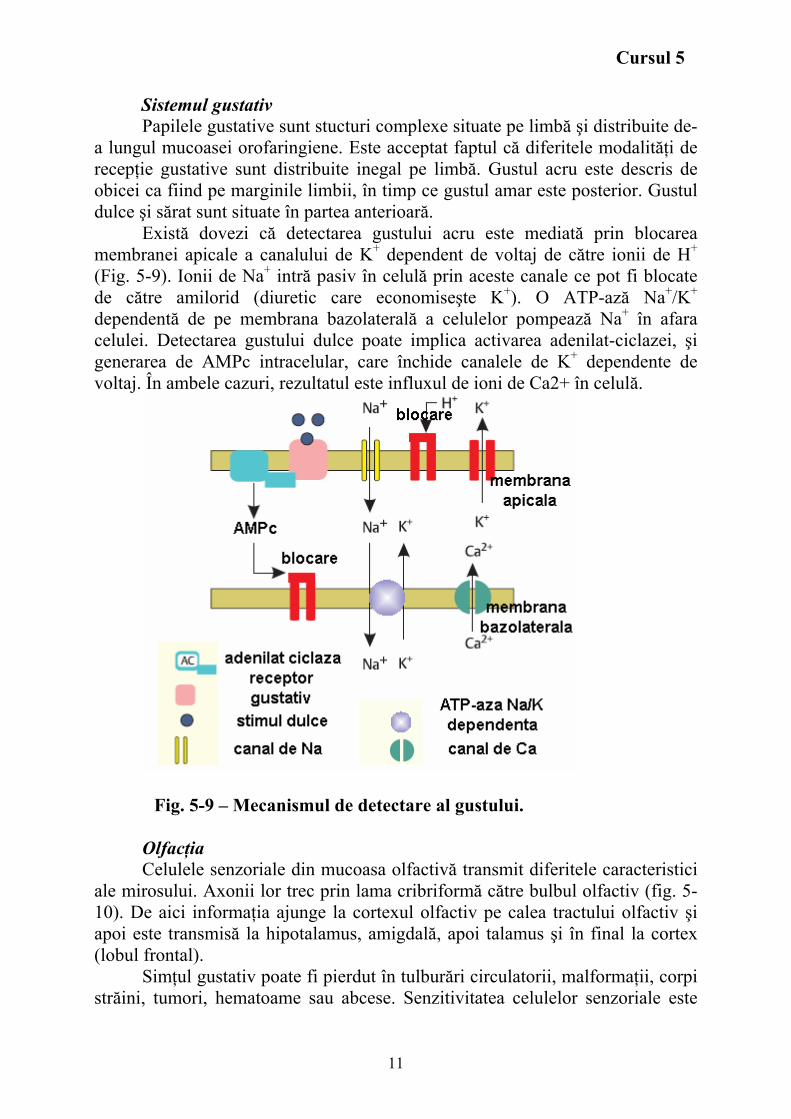
Sistemul gustativ Papilele gustative sunt stucturi complexe situate pe limbă şi distribuite de-
a lungul mucoasei orofaringiene. Este acceptat faptul că diferitele modalităţi de recepţie gustative sunt distribuite inegal pe limbă. Gustul acru este descris de obicei ca fiind pe marginile limbii, în timp ce gustul amar este posterior. Gustul dulce şi sărat sunt situate în partea anterioară.
Există dovezi că detectarea gustului acru este mediată prin blocarea membranei apicale a canalului de K+ dependent de voltaj de către ionii de H+ (Fig. 5-9). Ionii de Na+ intră pasiv în celulă prin aceste canale ce pot fi blocate de către amilorid (diuretic care economiseşte K+). O ATP-ază Na+/K+ dependentă de pe membrana bazolaterală a celulelor pompează Na+ în afara celulei. Detectarea gustului dulce poate implica activarea adenilat-ciclazei, şi generarea de AMPc intracelular, care închide canalele de K+ dependente de voltaj. În ambele cazuri, rezultatul este influxul de ioni de Ca2+ în celulă.
Fig. 5-9 – Mecanismul de detectare al gustului.
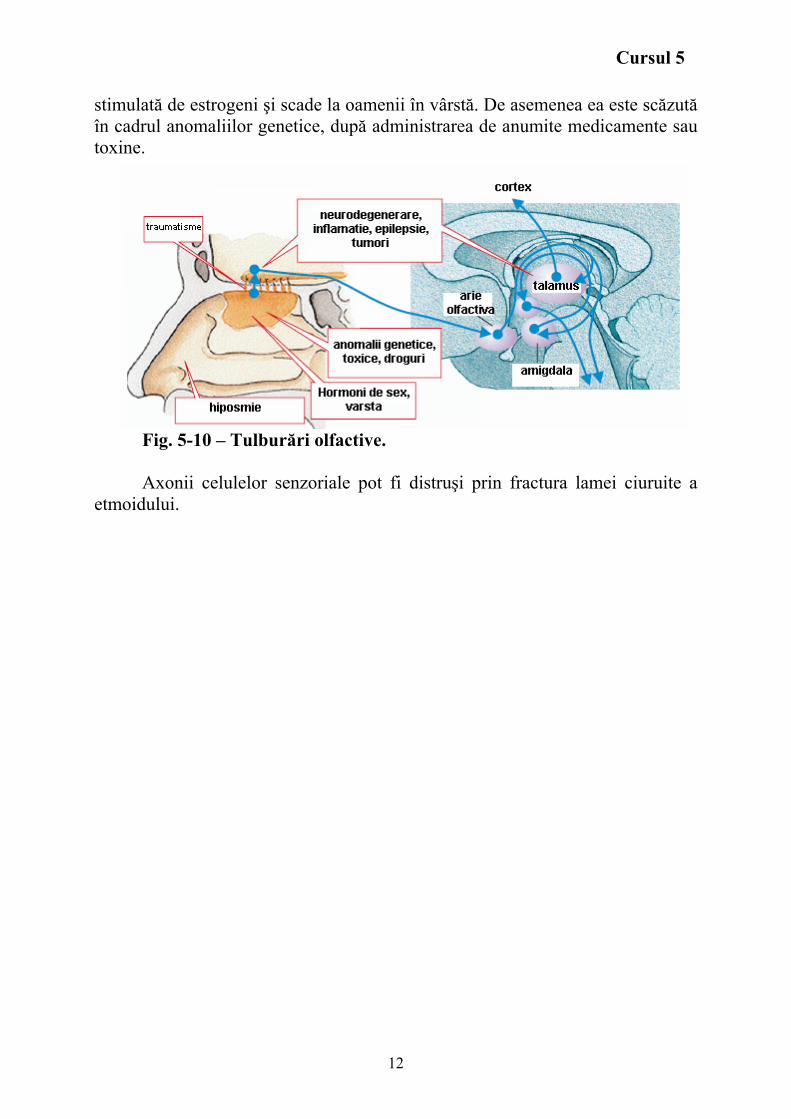
Olfacţia Celulele senzoriale din mucoasa olfactivă transmit diferitele caracteristici
ale mirosului. Axonii lor trec prin lama cribriformă către bulbul olfactiv (fig. 5-10). De aici informaţia ajunge la cortexul olfactiv pe calea tractului olfactiv şi apoi este transmisă la hipotalamus, amigdală, apoi talamus şi în final la cortex (lobul frontal).
Simţul gustativ poate fi pierdut în tulburări circulatorii, malformaţii, corpi străini, tumori, hematoame sau abcese. Senzitivitatea celulelor senzoriale este
Cursul 5
12
stimulată de estrogeni şi scade la oamenii în vârstă. De asemenea ea este scăzută în cadrul anomaliilor genetice, după administrarea de anumite medicamente sau toxine.
Fig. 5-10 – Tulburări olfactive. Axonii celulelor senzoriale pot fi distruşi prin fractura lamei ciuruite a
etmoidului.


















