Curs Citoscheletul Martie 2009
55
CITOSCHELETUL
-
Upload
andreea-turcanu -
Category
Documents
-
view
494 -
download
10
Transcript of Curs Citoscheletul Martie 2009

CITOSCHELETUL

• Reţea complexă de filamente proteice răspândite în toată citoplasma: microfilamentele de actină (MFA), filamentele intermediare (FI) şi microtubulii (MT)
• Structură dinamică care răspunde mediului înconjurător celulei
• Oferă posibilitatea deplasării organitelor citoplasmatice
• Favorizează segregarea cromozomilor• Responsabil de contracţia musculară
CITOSCHELETUL
DEFINIŢIE

CELULA EUCARIOTĂ ŞI PROCARIOTĂ

CITOSCHELETUL ASPECT GENERAL

• Provin din polimerizarea actinei G • Există trei tipuri de actină: α în fibrele
musculare striate şi netede, β şi γ în celulele non-musculare
• Forma de stocaj este forma G, proteină cu GM 42 kDa, cu un acid aminat rar, 3-metil-histidina
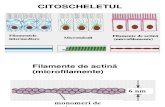
• Polimerizerea actinei G formează actina F, un filament cu diametrul de 7 nm
MICROFILAMENTELE DE ACTINĂ (MFA)

• Polimerizarea depinde de prezenţa şi concentraţia ionilor de Mg++ şi K+, de ATP şi de actina G
• Debutul se produce printr-o fază de nucleaţie, când trei molecule de actină se asamblează pentru a forma un trimer
• Urmează asocierea reversibilă a moleculelor de actină G
• Nucleaţia, polimerizarea, poziţia şi lungimea filamentelor de actină depind de o serie de proteine de legătură şi de hidroliza ATP-ului
Polimerizarea şi depolimerizarea actinei F numai în celulele non-musculare

• Citochalasinele, extracte de ciuperci microscopice, sunt inhibitori ai polimerizării actinei G
• Phaloidele (Amanita phalloides) sunt inhibitori ai depolimerizării şi provoacă intoxicaţii mortale
• Microfilamentele au o extremitate pozitivă a cărei creştere este mai rapidă decât extremitatea negativă.
• Diferenţa vitezei de creştere depinde de schimbarea conformaţiei moleculei de actină G
MICROFILAMENTELE DE ACTINĂ (MFA)

MFA – FORMA CELULARĂ ŞI FAGOCITOZĂ

MFA

• Instabilitate dinamică şi durată scurtă de viaţă• Actina G - polimerizare şi depolimerizare• Actina F - reticulare şi de dereticulare,
fasciculată şi defasciculată • Cea mai bine reprezentată proteină (5% din
totalul proteinelor) în celulele eucariote • Repartizată în întreaga citoplasmă, mai ales
perinuclear, dispoziţie sub formă de coş • La periferia celulei animale, sub membrana
plasmatică, dispunere în fascicole sau în reţele tridimensionale, formând cortexul celular
MFA CELULELOR NON-MUSCULARE

• ABP - actin binding protein - se asociază actinei G sau MFA
• Intervenind în polimerizarea/depolimerizarea MFA, în stabilizare, organizare în fascicole/reţea, fragmentare, fixare la faţa internă a membranei plasmatice
• În asociere cu MFA, constituie un citoschelet cortical (forma celulei )
PROTEINELE DE LEGĂTURĂ ALE ACTINEI

• Proteine de sechestrare de monomeri (profilină, depactină) reglează polimerizarea actinei G
• Proteine de blocaj ale extremităţilor - controlează lungimea MFA
• Proteine de stabilizare (tropomiozină) care menţin structura MFA
• Tropomiozina are numeroase situsuri de fixare la actină
PROTEINELE DE LEGĂTURĂ ALE ACTINEI

CITOSCHELETUL ŞI INTERACŢIUNILE SUBCELULARE

CITOSCHELETUL ŞI INTERACŢIUNILE SUBCELULARE

CITOSCHELETUL ŞI ADAPTĂRI ALE SUPRAFEŢEI CELULARE

CITOSCHELETUL ŞI FORMA CELULARĂ

FASCICULE CONTRACTILE• Sunt constituite din MFA asociate cu molecule de
miozină II • Se inseră de membrana plasmatică în puncte focale de
contact (placă de adezivitate, placă de aderenţă) conţin integrine
• Constituie inele contractile • Se asociază cu joncţiuni intermediare (centuri de
adezivitate)• În fasciculele contractile, miofilamentele de actină se
găsesc la distanţă de 30-60 nm unele de altele• Molecule de miozină II au o coadă şi două capete
globulare• Favorizează glisarea celor doua filamente de actină,
determinând contracţie fără modificarea lungimii

MFA ŞI PUNCTELE FOCALE DE CONTACT
• - ancorarea celulelor la substrat• - controlul locomoţiei celulare• - controlul formei celulare• - stabilitatea legăturii intercelulare• - răspunsul citoplasmatic la factorii de creştere
sau la alţi stimuli.• Fasciculele contractile se pot organiza în inele
contractile sau centuri de adezivitate

FASCICULELE STRÂNSE
• MFA paralele, legate prin proteine de asociere (fimbrina sau vilina), la distanţă de 10-20 nm unele de altele
• Aceste fascicule nu sunt contractile• Expansiunile filiforme (filopode, lamelipode) -
în deplasarea fibroblaştilor sau în microvilozităţi

MFA ŞI MIOZINA I DEPLASAREA ORGANITELOR
• Miozina I care are un singur cap şi o coadă, este responsabilă de mişcarea de deplasare de-a lungul MFA
• Miozina se fixează prin coada sa pe organit şi prin capul său de un MFA care este ancorat la membrana plasmatică
• Migrarea de la extremitatea negativă la extremitatea pozitivă a MFA

REŢELE DE MICROFILAMENTE
• Microfilamentele controlează vâscozitatea citoplasmei organizându-se în reţele
• Filamina, un dimer din 2 subunităţi de 270 kDa, este un factor gelificator al citoplasmei
• Gelsolina, în prezenţa calciului, fragmentează microfilamentele şi transformă un gel citoplasmatic într-un sol (rol fundamental în exocitoză sau endocitoză)

MICROFILAMENTELE ŞI FORMA CELULARĂ Hematia - celulă rotundă cu 2 feţe biconcave
• Interacţiunea dintre spectrină şi microfilamente, proteinele de legătură sau proteinele membranare
• Proteina de banda 4.1 conectează complexele joncţionale cu domeniile transmembranare ale glicoforinelor
• Ankrina, care este principala proteină de legătură a eritrocitelor, leagă tetramerii spectrinei de proteina membranară de bandă 3

MFA ŞI MIOZINA CELULELOR MUSCULARE STRIATE
• În FMS actina F se găseşte sub formă stabilă în structuri contractile, miofibrilele
• Miofibrilele - structuri diferenţiate, asocierea MFA şi miozină
• Formate din filamente de titină care menţin miofilamentele de miozină pe loc şi din filamente de nebulină care reglează asamblarea miofilamentelor de actină

SARCOMERELE
• FMS are mai multe mii de miofibrile cu diametrul între 1 şi 2 μm, care parcurg longitudinal fibra musculară de la un capăt la altul, despărţite de o cantitate scăzută de sarcoplasmă
• Miofibrilele constituie partea esenţială a muşchiului, aproximativ 100 de milioane /cm² de muşchi uman

INTERACŢIUNEA MFA MIOZINĂ 1

SARCOMERELE• Structură heterogenă (alternanţe de zone sau discuri)• - discurile A (anizotrope în lumină polarizată) întunecate, cu o
lungime de 1,5 μm: ele sunt împărţite în două semidiscuri printr-o zonă clară H (Hensen) care prezintă în mijloc o linie întunecată, linia M (Mittelmembran)
• - discurile I (Isotrope), cu o grosime de 0,8 μm, prezentând pe mijloc o striaţie Z (Zwichenscheibe) - desmină
• Fiecare segment de miofibrilă cuprins între două striaţii Z constituie un sarcomer.
• Un sarcomer are două semidiscuri I şi un disc A. • Repetarea acestor unităţi dispuse cap la cap constituie o
miofibrilă. • Sarcomerele tuturor miofibrilelor unei fibre sunt situate la
acelaşi nivel -aspect striat miocitelor

ORGANIZAREA MIOFILAMENTELOR ÎN MIOFIBRILE
• Miofibrilele asocierea a două tipuri principale de microfilamente
• Filamentele de miozină (miozina II) au un diametru de 10 nm şi o lungime de 1,5 μm şi extremităţi conice
• MFA au un diametru de 5-7 nm şi în jur de 1μ lungime şi se inseră pe striaţiile Z
• Aceste două tipuri principale de miofilamente se asociază cu titina care este una dintre cele mai voluminoase molecule fibrilare cunoscute astăzi (3000 kDa) şi cu filamente de nebulină

ORGANIZAREA MIOFILAMENTELOR ÎN MIOFIBRILE
• Filamentele de miozină şi MFA sunt paralele între ele şi longitudinale
• Filamentele de miozină ocupă în totalitate discul A• MFA din fiecare sarcomer pleacă de la striaţia Z, la marginea
discului H. • Semidiscurile I ale fiecărui sarcomer nu conţin decât MFA• Semidiscurile A situate de o parte şi de alta a discului H conţine
atât microfilamente de actină cât şi filamente de miozină. • Discul H are în partea mediană filament de miozină, uşor umflat
la mijloc. • Linia M corespunde ansamblului tuturor acestor umflături ale
miozinei.• Două specii moleculare ale miozinei: miozina de tip I, absentă în
celulele musculare şi care are rol în motilitatea celulară şi miozina II

FILAMENTELE DE MIOZINĂ II
• Sunt bipolare, GM de 450 kDa şi este alcătuită din 6 lanţuri polipeptidice : 2 lanţuri grele şi 2 perechi de lanţuri uşoare
• Lanţurile grele sunt identice – o regiune α helicoidală cu o lungime de 30 nm, numită coadă şi o regiune globulară carboxi-terminală, cu diametrul de 4 nm numită cap.
• Miozina poate fi scindată la nivelul punctelor de flexiune sub acţiunea unor enzime proteolitice
• Chimiotripsina scindează miozina în două domenii• Primul domeniu, meromiozina grea (HMM), conţine capul bilobat
şi un segment din coada lanţurilor grele, • Al doilea domeniu, meromiozina uşoară (LMM), cuprinde
segmentul aminoterminal al cozii• Prin tratarea domeniului HMM cu papaină, segmentul de coadă
este eliberat şi rezultă fragmentele S1, ce conţin câte un cap globular şi perechea de lanţuri asociate acestuia.
• Fragmentele S1 prezintă activitate ATP-azică

FILAMENTELE INTERMEDIARE (FI)
• Molecule fibrilare stabile, cu un diametru de 8-10 nm, intermediar între cel al MFA şi cel al MT, în nucleoplasma şi citoplasma tuturor celulelor animale
• În celulele epiteliale se dispun în jurul nucleului şi sub membrana plasmatică.
• Se inseră de proteinele membranare cu ajutorul proteinelor extrinseci citoplasmatice
• În celulele epiteliale se fixează de placa citoplasmatică a desmozomilor şi au rol mecanic
• În axonii celulelor nervoase formează o armătură• In FMS unifică faţa internă a membranei cu
miofibrilele - transmiterea forţei în cursul contracţiei

FILAMENTELE INTERMEDIARE (FI)
• FI (polimeri stabili) pot fi:• Homopolimeri : vimentina, desmina, GFA sau GFAP,
periferina• Heteropolimeri: citocheratinele, neurofilamentele.• Unitatea de bază - un monomer cu o parte centrală
(identică pentru mai multe tipuri) şi două extremităţi carboxy- şi amino-terminale diferite
• Partea centrală cuprinde 310 resturi de acizi aminaţi dispuşi în dublu helix cu trei regiuni în care dispoziţia acestor reziduuri nu este helicoidală

FILAMENTELE INTERMEDIARETipul I şi tipul II - cheratinele
• Tipul I - cheratinelor acide• Tipul II familiile de cheratine neutre şi bazice• Proteine fibrilare produse de cheratinocite sau celule
epiteliale• MARKER EPITELIAL CITOPLASMATIC• În cheratinocite sub formă de fascicule ancorate în plăcile
dense desmozomale• Citocheratinele CK5 şi CK14 - stratul bazal• CK1, CK6, CK10, CK16, - corpul mucos Malpighi• CK9 - stratul granular.• În epiderm CK unesc desmozomii între ei• CK se inseră de molecule de pacoglobulină - o placă densă
vecină şi paralelă cu membrana plasmatică• Moleculele de pacoglobulină se lipesc de caderinele
desmozomale transmembranare

FI – CITOCHERATINA IMUNOFLUORESCENŢĂ

FI – CITOCHERATINA IMUNOHISTOCHIMIE (IHC)

FILAMENTELE INTERMEDIARETipul III - Vimentina, Desmina, GFAP şi Periferina
• Vimentina (54 kDa) - MARKER al celulelor mezenchimatoase
• Seroasa pleurală, pericardică, peritoneală• În celulele endoteliale vasculare• În fibroblaşti, fibrocite, condrocite• În cursul embriogenezei, neuroblaştii (neuroni imaturi)
conţin VIM• Absenţa vimentinei în citoscheletul celulelor suşă
vimentină modifică morfologia nucleului• În adipocite VIM formează un fel de cuşcă în jurul
grăsimii• Menţine pe loc nucleul şi ansamblul organitelor celulare

VIMENTINA - IHC

FILAMENTELE INTERMEDIARETipul III - Vimentina, Desmina, GFAP şi Periferina
• Desmina (53 kDa) MARKER al FMN sau FMS• FMS cursul miogenezei sau în cursul regenerării conţin o
mare cantitate de DES• În FMS adulte DES leagă striaţiile Z vecine unele de
altele şi de membrana plasmatică• GFAP (50 kDa) MARKER al celulelor de origine
neuroectodermică -nevroglia sistemului nervos central (astrocite şi microglie) şi a sistemului nervos periferic (celule Schwann ale fibrelor amielinice)
• Periferina (57 kDa) MARKER al neuronilor în cursul dezvoltării embrionare şi în cursul regenerării
• PER este indispensabilă regenerării neuronale

FI – DESMINA – IMUNOFLUORESCENŢĂ ŞI IHC

GFAP - IMUNOFLURESCENŢĂ

GFAP - IHC

FILAMENTELE INTERMEDIARE Tipul IV - neurofilamentele
• NF - localizate în neuroni şi se asociază MT axonilor• 3 tipuri: NF-L (light), NF-M (medium), NF-H (heavy)• Reprezintă 25% dintre proteinele axonului şi intervin
împreună cu neurotubulii în transportul intra-axonal (flux axonal)
• Menţin rigidităţii şi rezistenţei axonului precum • Prin fosforilarea NF-H şi a NF-M se adaugă la
microfilamente grupări fosfat (sarcina negativă)• Forţele de respingere, generate prin creşterea
sarcinilor negative, asigură un diametru axonal constant.

NF - IMUNOFLUORESCENŢĂ

NF – Schemă şi IHC

FILAMENTELE INTERMEDIARETipul V - laminine nucleare
• Lamininele constituie citoscheletul nucleului • Se cunosc 2 tipuri: laminine de tip I (cu subtipurile A şi C),
laminine de tip II (cu subtipurile B1, B2)• Formează o reţea tridimensională care acoperă complet, cu
excepţia porilor, faţa internă a învelişului nuclear• Microfilamentele de tip A şi C menţin forma nucleului şi
servesc drept loc de ancorare pentru moleculele de ADN • Laminina B nu are afinitate pentru nucleotide• Intervin în mitoză - lamininele A şi C se fixează pe siturile
specifice de legătură ale cromozomilor mitotici• Lamininele intervin în momentul reconstrucţiei nucleului şi al
învelişului nuclear

MICROTUBULII (MT)
• MT sunt polimeri instabili lungi şi rigizi de heterodimeri α-β de tubulină
• Au o extremitate pozitivă distală (apropiată de membrana celulară) şi o extremitate negativă aproape de centrul celular
• Au forma unor tubi cu diametrul în jur de 25 nm, a căror polimerizare şi depolimerizare depind de hidroliza GTP
• Se găsesc la vertebratele superioare, în toate tipurile celulare (cu excepţia hematiei)
• Sunt mai numeroşi în neuronii cerebrali unde reprezintă în jur de 20 % dintre proteinele solubile.

MICROTUBULI

Microtubuli - Ultrastructura cilului

CENTROZOMUL - MTOC
• MT se dispun radiar în jurul centrosomului - MTOC (microtubule organizing center)
• MTOC - doi centrioli şi o masă granulo-filamentoasă pericentriolară
• MT cresc plecând din centrozom - extremitatea lor negativă pătrunde în masa pericentriolară, dar nu intră niciodată în contact cu centriolii .
• Organizare moleculară a MT• MT au un diametru extern uniform, perete dens, îngroşat, de
5 nm şi o cavitate axială de 14 nm lărgime• Lungimea de mai mulţi microni • MT nu se ramifică• Peretele are 13 protofilamente, alternanţă de tubuline α şi β
de 5 nm diametru, paralele şi având aceeaşi polaritate, legate lateral unele de altele

CICLUL CELULAR – MTOC – FUSUL DE DIVIZIUNE

FUNCŢIILE MICROTUBULILOR
• Dinamica mediului extracelular• Deplasarea celulelor• Morfogeneza celulei şi menţinerea formei celulare• Polaritatea celulară• Menţinerea structurii membranei celulare• Migrarea vacuolelor de endocitoză• Eliberarea granulelor de secreţie• Integritatea aparatului Golgi• Mişcarea cromozomilor în cursul mitozei şi a meiozei• Transportul dirijat al ARN-m în citoplasmă• Transportul substanţelor şi al materialului intracelular

CITOSCHELETUL – IMUNOFLUORESCENŢĂ


CITOSCHELETUL ŞI MEMBRANA PLASMATICĂ

CITOSCHELETUL ŞI TRANSPORTUL TRANSMEMBRANAR

CITOSCHELETUL


















