Biochimia Eritrocitului Matur
5
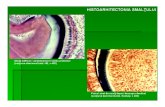
Barbu Oana Maria Biochimia eritrocitului matur Eritrocitele, numite si globule rosii, celule rosii sau hematii sunt elemente figurate circulante, lipsite de nucleu in faza adulta, de culoare rosie. Eritrocitele se formeaza in maduva rosie hematogena in urma eritropoezei, latura a procesului de hematopeza. In ultima faza a eritropoezei, nucleul este eliminate din citoplasma. Eritropoeza este un proces reglat hormonal de eritropoetina. Durata medie de viata a hematiilor este de 120 de zile, dupa care hematiile imbatranite sunt distruse in ficat, splina, maduva osoasa sau ganglioni limfatici prin hemoliza. Hematiile sunt maleabile si elastice, avand capacitatea de a se deforma reversibil la trecerea prin capilarele cu diametrul foarte ingust. Diametrul eritrocitelor este de 7.8 microni. Eritrocitul matur prezinta o forma lenticulara biconcava. El este alcatuit din stroma, membrana eritrocitara formata din mai multe straturi si citoplasma cu hemoglobina. Membrana hematiei este un complex lipo-glicoproteic, completat de citoschelet si reprezinta o veritabila bariera selectiva pentru schimburile cu mediul. Contine 52 % proteine, 40% lipide si 8% glucide asociate proteinelor. Membrana este alcatuita din 3 straturi: extern, mijlociu si intern. Stratul extern este de natura glicoproteica. Glicoproteinele prezente sunt predominant incarcate negativ datorita prezentei acidului salicilic. Acest strat se distinge prin prezenta a numerosi antigeni printre care si cei ai grupelor sangvine. Unele antigene sunt specifice eritrocitelor, in timp ce altele sunt intalnite si pe alte celule din organism. Stratul mijlociu este de natura lipidica. Contine spre exterior fosfatidilcolina si sfingomielina si spre interior fosfatidil serina si fosfatidiletanolamina. De asemenea mai contine si colesterol. Stratul intern este o retea de proteine fibrilare si globuloase, conferind hematiei rezistenta, forma, capacitate de deformabilitate si integrare in circulatie.
-
Upload
oana-barbu -
Category
Documents
-
view
20 -
download
1
description
Biochimia Eritrocitului Matur anul 1
Transcript of Biochimia Eritrocitului Matur
Barbu Oana Maria
Biochimia eritrocitului matur
Eritrocitele, numite si globule rosii, celule rosii sau hematii sunt elemente figurate circulante, lipsite de nucleu in faza adulta, de culoare rosie.Eritrocitele se formeaza in maduva rosie hematogena in urma eritropoezei, latura a procesului de hematopeza. In ultima faza a eritropoezei, nucleul este eliminate din citoplasma. Eritropoeza este un proces reglat hormonal de eritropoetina. Durata medie de viata a hematiilor este de 120 de zile, dupa care hematiile imbatranite sunt distruse in ficat, splina, maduva osoasa sau ganglioni limfatici prin hemoliza.Hematiile sunt maleabile si elastice, avand capacitatea de a se deforma reversibil la trecerea prin capilarele cu diametrul foarte ingust. Diametrul eritrocitelor este de 7.8 microni.Eritrocitul matur prezinta o forma lenticulara biconcava. El este alcatuit din stroma, membrana eritrocitara formata din mai multe straturi si citoplasma cu hemoglobina.Membrana hematiei este un complex lipo-glicoproteic, completat de citoschelet si reprezinta o veritabila bariera selectiva pentru schimburile cu mediul. Contine 52 % proteine, 40% lipide si 8% glucide asociate proteinelor. Membrana este alcatuita din 3 straturi: extern, mijlociu si intern. Stratul extern este de natura glicoproteica. Glicoproteinele prezente sunt predominant incarcate negativ datorita prezentei acidului salicilic. Acest strat se distinge prin prezenta a numerosi antigeni printre care si cei ai grupelor sangvine. Unele antigene sunt specifice eritrocitelor, in timp ce altele sunt intalnite si pe alte celule din organism. Stratul mijlociu este de natura lipidica. Contine spre exterior fosfatidilcolina si sfingomielina si spre interior fosfatidil serina si fosfatidiletanolamina. De asemenea mai contine si colesterol. Stratul intern este o retea de proteine fibrilare si globuloase, conferind hematiei rezistenta, forma, capacitate de deformabilitate si integrare in circulatie.Citoplasma (citosolul) contine 60% apa, 33-35% hemoglobina si 5-7% alte substante dintre care glucoza, glutation, ATP, compusi metalici (fosfor, zinc, fier).Principala functie a eritrocitelor este aceea de a vehicula hemoglobina, care la randul sau transporta oxigenul de la plamani catre testuturi. Hemoglobina este, din punct de vedere fiziologic, partea cea mai importanta a unei globule rosii. Hemoglobina este o cromoproteina colorata in rosu aprins.
Formarea hemoglobineiIn cadrul ciclului Krebs, se produce succinil CoA care se va lega de glicina formand o molecula de pirol. La randul lor, 4 molecule de pirol se unesc formand protoporfirina IX, iar aceasta leaga apoi fierul si formeaza molecula de hem. In final, fiecare molecula de hem se leaga de un lant polipeptidic lung, care este o globina sintetizata de ribozomi, formand o subunitate a hemoglobinei numita lant hemoglobinic. Patru dintre aceste lanturi se unesc prin legaturi labile si formeaza o molecula completa de hemoglobina. Exista unele mici deosebiri intre diferitele lanturi hemoglobinice, in functie de compozitia in aminoacizi a regiunii polipeptidice. Aceste tipuri de lanturi sunt denumite alfa, beta, gamma si delta. Cea mai frecventa forma de hemoglobina la adult, hemoglobina A este o combinatie dintre 2 lanturi alfa si 2 lanturi beta.Fiecare lant hemoglobinic are o gruprare prostetica hem ce contine un atom de fier iar in fiecare molecula de hemoglobina sunt 4 lanturi hemoglobine, prin urmare, vor exista 4 atomi de fier in fiecare molecula de hemoglobina.
Datorita prezentei hemoglobinei, eritrocitele joaca cel mai important rol in cadrul respiratiei. La nivelul plamanilor, hemoglobina fixeaza reversibil oxigenul molecular din aer transformandu-se in oxihemoglobina. O molecula de hemoglobina poate fixa 4 molecule de oxigen gazos. Acest lucru este explicat de faptul ca in structura hemoglobinei intalnit 4 atomi de fier ce vor lega 4 molecule de oxigen. Oxigenul nu se combina cu cele doua valente pozitive ale fierului din molecula de hemoglobina, ci se leaga slab de atomul de fier in asa-numita legatura coordinativa. Aceasta legatura exte extrem de labila, de aceea reactia este foarte usor reversibila. In plus, oxigenul nu se ionizeaza, fiind transportat catre tesuturi ca oxigen molecular, unde, datorita legaturilor labile, usor revesibile, este eliberat in lochidele tisulare tot sub forma de oxigen molecular si nu de oxigen ionic.La nivelul tesuturilor, hemoglobina cedeaza oxigenul si preia dioxidul de carbon rezultat din procesele catabolice ale celulelor dand nastere carbohemoglobinei.Daca in aer exista monoxid de carbon, hemoglobina se combina preferential cu acesta (Hb are o afinitate de 300 de ori mai mare fata de CO comparativ cu O2) formandu-se carboxihemoglobina, Aceasta este o reactie ireversibila iar functia de aprovizionare cu oxigen a celulelor se diminueaza sau inceteaza, instalandu-se hipoxia.Eritrocitele au si alte functii in afara transportului hemoglobinei. De exemplu, ele contin o cantitate mare de anhidraza carbonica, enzima ce catalizeaza reactia revesibila dintre dioxidul de carbon si apa cu formarea de acid carbonic, amplificand de cateva mii de ori rata acestei reactii. Viteza de desfasurare a acestei reactii, permite ca apa din componenta sangelui sa transporte, de la tesuturi la plamani, cantitati enorme de dioxid de carbon sub forma de ion carbonat, in plamani acesta fiind reconvertit in dioxid de carbon si eliminat in atmosfera ca un produs rezidual.O particularitate metabolica a eritrocitului este aceea ca eritrocitele au un consum metabolic foarte scazut participand cu doar 0.1% la metabolismul general (2cal/24h). Nu au mitocondrii, prin urmare fac doar glicoliza anaeroba. Consuma foarte putin din oxigenul pe care il transporta. Hematiile necesita energie pentru mentinerea formei si a deformabilitatii, asigurarea concentratiei electrolitilor de o parte si de alta a membranei celulare, mentinerea fierului in stare feroasa, redusa, pentru fixarea O si cedarea CO . Aceasra energie o procura prin metabolizarea glucozei. Glucoza din plasma patrunde in eritrocit prin difuziune facilitata (mecanism activ insulino-independent) si este metabolizata pe calea glicolitica cu ajutorul enzimelor cheie HK, FF si piruvatkinaza (90%) sau pe cale oxidativa a suntului pentozofosfatilor (8-10%). Hemoglobina intracelulara este un excelent tampon acido-bazic si de aceea, eritrocitele, sunt responsabile pentru cea mai mare parte a capacitatii de tamponare acido-bazica a sangelui. Principalele sisteme tampon din cadrul eritrocitelor sunt Hb redusa/ HbK, O2Hb/O2HbK si KH2PO4. La disocierea Hb, disociaza nu doar gruparea carboxil si amino, ci si gruparile imidazolice ale His. In refacerea cantitatii de bicarbonat de Na, un rol important revine unui fenomen: FENOMENUL DE MEMBRAN HAMBURGER (fenomen de migrare a ionului de Cl).
Metabolismul fieruluiDupa ce este absorbit din intestinul subtire, fierul se leaga imediat de o beta-globulina plasmatica, apotransferina, formand transferina, care este transportata apoi in plasma. Fierul este legat slab in transferina, de aceea poate fi eliberat catre orice celula din organism.In citoplasma celulara, fierul se leaga in principal de o proteina numita apoferitina, formand feritina. Avand o greutate moleculara mare, feritina poate contine atat concentratii mici de fier cat si concentratii mari. Mici cantitati de fier sunt depozitate sub o forma extrem de insolubila, numita hemosiderina. Aceasta este produsa in special atunci cand cantitatea totala de fier din organism depaseste capacitatea de depozitare a apoferitinei.La persoanele care nu au cantitati suficiente de transferina in sange apare o incapacitate de a transporta fierul in eritroblastii ceea ce determina o anemie hipocroma severa, cu alte cuvinte hematiile contin o cantitate mult mai mica de hemoglobina decat in mod normal.