
UNIVERSITATEA DE ŞTIINŢE AGRICOLE ŞIexplantele au fost recoltate din colţi de tuberculi,...
54
UNIVERSITATEA DE ŞTIINŢE AGRICOLE ŞI MEDICINǍ VETERINARĂ CLUJ-NAPOCA ŞCOALA DOCTORALǍ FACULTATEA DE AGRICULTURĂ Ing. ELIZA MARIA AGUD REZUMATUL TEZEI DE DOCTORAT Utilizarea culturilor in vitro în multiplicarea şi ameliorarea cartofului (Solanum tuberosum L.) Conducător ştiinţific: Prof. univ. dr. MIRCEA SAVATTI CLUJ-NAPOCA 2010
Transcript of UNIVERSITATEA DE ŞTIINŢE AGRICOLE ŞIexplantele au fost recoltate din colţi de tuberculi,...

UNIVERSITATEA DE ŞTIINŢE AGRICOLE ŞI MEDICINǍ VETERINARĂ CLUJ-NAPOCA
ŞCOALA DOCTORALǍ FA
CULTATEA DE AGRICULTURĂ
Ing. ELIZA MARIA AGUD
REZUMATUL TEZEI DE DOCTORAT
Utilizarea culturilor in vitro în multiplicarea şi ameliorarea cartofului (Solanum tuberosum L.)
Conducător ştiinţific:
Prof. univ. dr. MIRCEA SAVATTI
CLUJ-NAPOCA
2010

Utilizarea culturilor in vitro în multiplicarea şi ameliorarea cartofului
(Solanum tuberosum L.)
Rezumatul tezei de doctorat
Majoritatea studiilor efectuate timp de un secol în domeniul vitroculturilor
vegetale au servit la elucidarea mai multor aspecte privind regenerarea, creşterea,
organogeneza, embriogeneza somatică ş.a., a unor structuri tisulare sau a unor tipuri
celulare, în funcţie de compoziţia mediului de cultură şi a condiţiilor ecofiziologice
din recipientele de cultură şi a camerei de creştere. Există, însă, încă, numeroase
necunoscute cu privire la unii factori implicaţi în morfogeneză (în special la nivel
celular şi molecular), precum şi referitor la transmiterea şi a informaţiei genetice
nou dobândite.
Culturile in vitro au găsit într-un timp foarte scurt numeroase aplicaţii
practice în ameliorarea numeroaselor specii de cultură, implicit la cartof, privind
multiplicarea genotipurilor valoroase, propagarea stocurilor libere de viroze,
conservarea resurselor genetice ş.a. În plus, variaţia genetică manifestată de plantele
regenerate în cultura in vitro (variabilitatea somaclonală) a putut şi este folosită în
obţinerea de genotipuri valoroase.
Pornind de la aceste constatări ne-am străduit ca pe lângă sintetizarea
rezultatelor pro- şi contra, privind problema multiplicării şi creării de variaţii in
vitro la cartof, să încercăm să elaborăm propriile noastre păreri în cadrul tezei de
doctorat “Utilizarea culturilor in vitro în multiplicarea şi ameliorarea cartofului
(Solanum tuberosum L.)”.
În abordarea acestui subiect s-a urmărit elucidarea unor aspecte privind
posibilităţile de regenerare şi multiplicare in vitro la cartof, crearea de variabilitate
genetică, microtuberizarea şi determinarea diversitǎţii genetice.
În scopul celor menţionate, abordarea noastră se referă la unele aspecte
majore care să evidenţieze următoarele:
- îmbunǎtǎţirea tehnologiilor de micropropagare in vitro a cartofului;
2

- interacţiunea hormonilor de creştere asupra neoformǎrii plantulelor
(AIB, AIA, ANA, K, BAP);
- mediile de culturǎ în raport cu cerinţele nutritive ale explantelor de
cartof ;
- inducerea variaţiei somaclonale in vitro la cartof;
- testarea serologicǎ a plantelor de cartof regenerate in vitro;
- obţinerea de micro- şi minituberculi de cartof in vitro;
- determinarea diversitǎţii genetice a cultivarelor de cartof.
Pe parcursul elaborării tezei de doctorat ne-am străduit să dăm un răspuns cât
mai plauzibil elucidării aspectelor luate în studiu, corelând datele proprii cu
multiplele sugestii oferite de literatura de specialitate.
Prin natura sa biologică, plantă autogamă, dar cu posibilităţi de multiplicare
vegetativă, metodologia de lucru adoptată conferă posibilitatea decelării formelor
biologice ale multiplicării genotipurilor de cartof, in vitro şi adaptarea
neoplantulelor proceselor de tuberizare.
Regenerarea şi oragnogeneza in vitro la unele specii de plante constituie o
condiţie esenţială pentru reuşita multiplicării plantelor respective pe cale vegetativă.
Procesele de morfogeneză şi organogeneză se realizează sub influenţa factorilor
endogeni şi exogeni. Conţinutul endogen de hormoni, tipul acestora, precum şi
echipamentul enzimatic sunt factori cu rol de reglare a regenerării. Natura
fitohormonilor utilizaţi precum şi concentraţia lor, diferenţele balanţelor hormonale
au un rol important în procesele de organogeneză.
Materialul biologic utilizat, dupǎ cum s-a amintit, a fost constituit din opt
genotipuri de cartof, patru soiuri, Ostara, Desirée, Christian, Roclas şi patru
populaţii locale, Mǎdǎraş, Miercurea Ciuc, Tuşnad şi Lunca de Jos şi incidental
soiul Eba.
Cercetările efectuate în perioada 2006-2009, au fost realizate în Laboratorul
de biotehnologii a Universităţii de Ştiinţe Agricole şi Medicină Veterinară Cluj-
Napoca.
Metodele biotehnologice, prin aplicarea tehnicilor de culturǎ in vitro
reprezintǎ o cale sigurǎ pentru obţinerea unui material biologic liber de agenţi
3

patogeni şi calea cea mai rapidǎ pentru crearea şi multiplicarea genotipurilor
superioare de cartof.
Materialul biologic utilizat a fost constituit din opt cultivare de cartof
(Ostara, Desirée, Christian, Roclas, Mǎdǎraş, Miercurea Ciuc, Tuşnad, Lunca de Jos
şi incidental soiul Eba), cu caracteristici morfo-fiziologice distincte.
Pentru înfiinţarea culturilor primare, plantele mamǎ aparţinând cultivarelor
menţionate au fost alese cu grijǎ sub aspectul tipicitǎţii şi a autenticitǎţii, cât şi a
stǎrii fito-sanitare. Pentru ca prelevarea de meristeme şi inocularea acestora pe
mediu de culturǎ sǎ permitǎ iniţierea unei culturi in vitro s-a utilizat mediul de bazǎ
Murashige-Skoog, în condiţii aseptice, corectat cu fitohormoni, în concentraţii
diferite. Administrarea balanţei hormonale s-a fǎcut în funcţie de starea fiziologicǎ a
donorului din momentul explantǎrii ţesuturilor.
Capacitatea regenerativǎ a meristemelor de a forma plante sau calus, a depins
foarte mult de genotip, din mediul de culturǎ utilizat şi în special de regulatorii de
creştere prezenţi în substratul de culturǎ.
S-a constatat cǎ utilizarea explantelor recoltate de la diferite organe
influenţeazǎ procentul plantelor iniţiate. cele mai bune rezultate se obţin când
explantele au fost recoltate din colţi de tuberculi, fragmente de tulpinǎ şi rǎdǎcinǎ şi
mai puţin din fragmente de frunze şi mezofil folial (vezi tabelul 4.1).
Cultivarul Ostara realizeazǎ cele mai multe plante iniţiate când explantele
sunt recoltate de la organe în curs de evoluţie, urmat de cultivarul Desirée;
cultivarele populaţii locale au un comportament identic cu cel al soiurilor
omologate.
Tipul, concentraţia şi durata de acţiune a agenţilor chimici sterilizanţi asupra
organelor de la care se preleveazǎ explantele au efecte diferite. utilizarea
hipocloritului de calciu, hipocloritului de calciu + tween şi hipoclorit de sodiu în
concentraţie de 5-10%, au dat rezultate diferite, dovedindu-se eficiente hipocloritul
de calciu şi hipocloritul de calciu + tween în concentraţie de 5% cu o duratǎ a
tratamentului de 20 de minute (vezi tabelul 4.3).
4

5

Tabelul 4.3.
Agenţi dezinfectanţi şi acţiunea lor asupra explantelor provenite de la donatori diferiţi
Desinfecting agents and their action on explants from different donors
Viabilitatea (%) Viability (%)
Grad de dezinfecţie (%) Desinfection degree (%)
Agent dezinfectant Desinfected
agent
Concentraţia Concentration
%
Durata tratament Treatment duration
(min)
Lǎstari tineri Young shoots
Muguri dorminzi
(tuberculi) Dormand
buds (tubers)
Colţi în creştere
(tuberculi)Growing shoots
(tubers)
Lǎstari tineri Young shoots
Muguri dorminzi
(tuberculi) Dormand
buds (tubers)
Colţi în creştere
(tuberculi)Growing shoots
(tubers) 5 20 88 81 99 66 75 80 7 20 79 74 87 73 78 93
Hipoclorit de calciu
Calcium hypochlorite 10 20 63 69 81 68 62 93
5 20 93 92 96 87 80 71 7 20 91 86 91 82 89 75
Hipoclorit de calciu + tween
Calcium hypochlorite
+ tween
10 20 76 70 73 70 76 84
5 20 85 80 98 82 80 80 7 20 71 70 90 67 70 81
Hipoclorit de sodiu
Sodium hypochlorite 10 20 73 70 80 68 67 78
Pe mǎsurǎ ce se mǎreşte concentraţia agentului dezinfectant, gradul de
dezinfecţie se amplificǎ dar viabilitatea explantei scade.
Pentru cultivarea celulelor şi ţesuturilor in vitro au fost utilizate pentru
îmbunǎtǎţirea mediilor nutritive de bazǎ, auxine, citochinine şi mai puţin gibereline.
Auxinele utilizate pentru inducerea diviziunilor celulare şi a rizogenezei au fost
acidul indolilacetic (AIA), acidul indolilbutiric (AIB) şi acidul alfa-naftilacetic (ANA)
în concentraţii de 0,5-2,0 mg/l. Citochininele de sintezǎ utilizate au fost chinetina (K)
(6-furfurilaminopurina) şi BAP (6-benzilaminopurina).
Interacţiunea dintre hormonii utilizaţi în mediul de culturǎ şi concentraţia
acestora la înmulţirea in vitro a cultivarelor de cartof determinǎ diferenţe
semnificative în ceea ce priveşte rata de neoformare a plantulelor.
Utilizarea acidului β-indolilbutiric (AIB) asupra formǎrii şi dezvoltǎrii
neoplantulelor de cartof, s-a evidenţiat, indiferent de genotip, asupra procesului de
rizogenezǎ, la nivelul concentraţiilor de 0,5-2,0 mg/l.
6
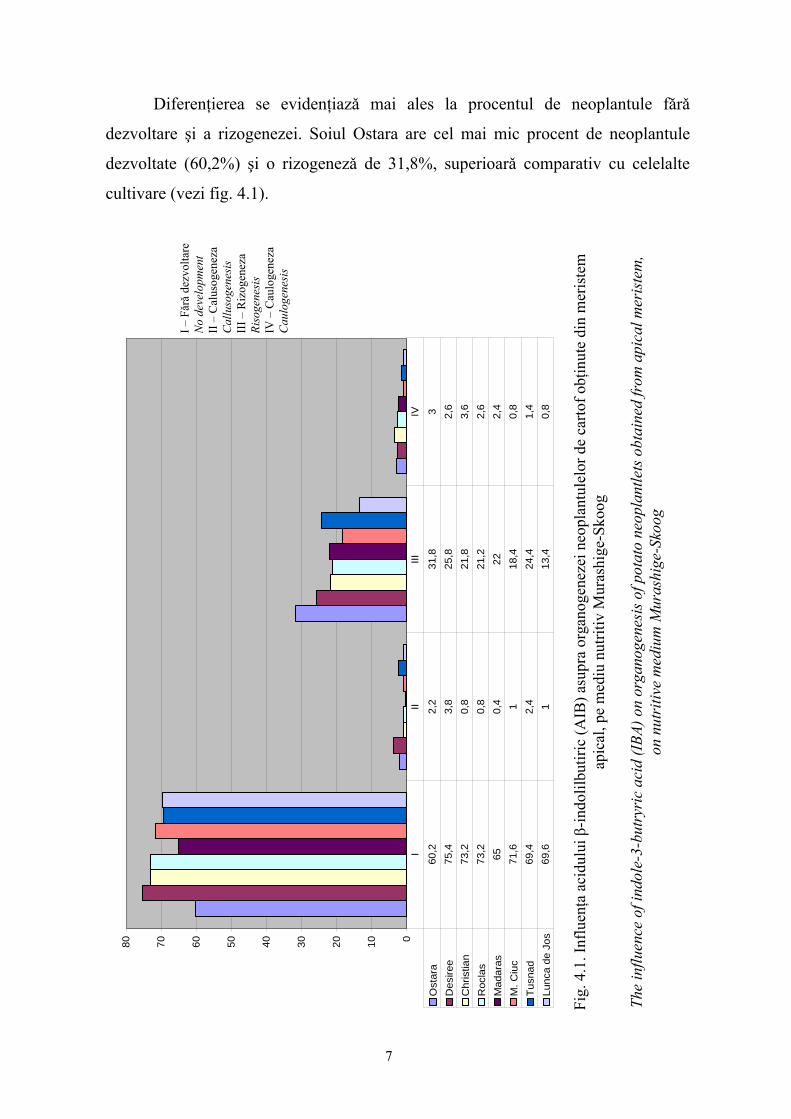
Diferenţierea se evidenţiazǎ mai ales la procentul de neoplantule fǎrǎ
dezvoltare şi a rizogenezei. Soiul Ostara are cel mai mic procent de neoplantule
dezvoltate (60,2%) şi o rizogenezǎ de 31,8%, superioarǎ comparativ cu celelalte
cultivare (vezi fig. 4.1).
01020304050607080
Ost
ara
Des
iree
Chr
istia
nR
ocla
sM
adar
asM
. Ciu
cTu
snad
Lunc
a de
Jos
Ost
ara
60,2
2,2
31,8
3
Des
iree
75,4
3,8
25,8
2,6
Chr
istia
n73
,20,
821
,83,
6
Roc
las
73,2
0,8
21,2
2,6
Mad
aras
650,
422
2,4
M. C
iuc
71,6
118
,40,
8
Tusn
ad69
,42,
424
,41,
4
Lunc
a de
Jos
69,6
113
,40,
8
III
IIIIV
I – Fǎrǎ
dezv
olta
re
No
deve
lopm
ent
II –
Cal
usog
enez
a C
allu
soge
nesi
s II
I – R
izog
enez
a Ri
soge
nesi
s IV
– C
aulo
gC
aulo
gene
sis en
eza
Fig.
4.1
. Inf
luenţa
aci
dulu
i β-in
dolil
butir
ic (A
IB) a
supr
a or
gano
gene
zei n
eopl
antu
lelo
r de
carto
f obţ
inut
e di
n m
eris
tem
ap
ical
, pe
med
iu n
utrit
iv M
uras
hige
-Sko
og
Th
e in
fluen
ce o
f ind
ole-
3-bu
tryr
ic a
cid
(IBA
) on
orga
noge
nesi
s of p
otat
o ne
opla
ntle
ts o
btai
ned
from
api
cal m
eris
tem
, on
nut
ritiv
e m
ediu
m M
uras
hige
-Sko
og
7
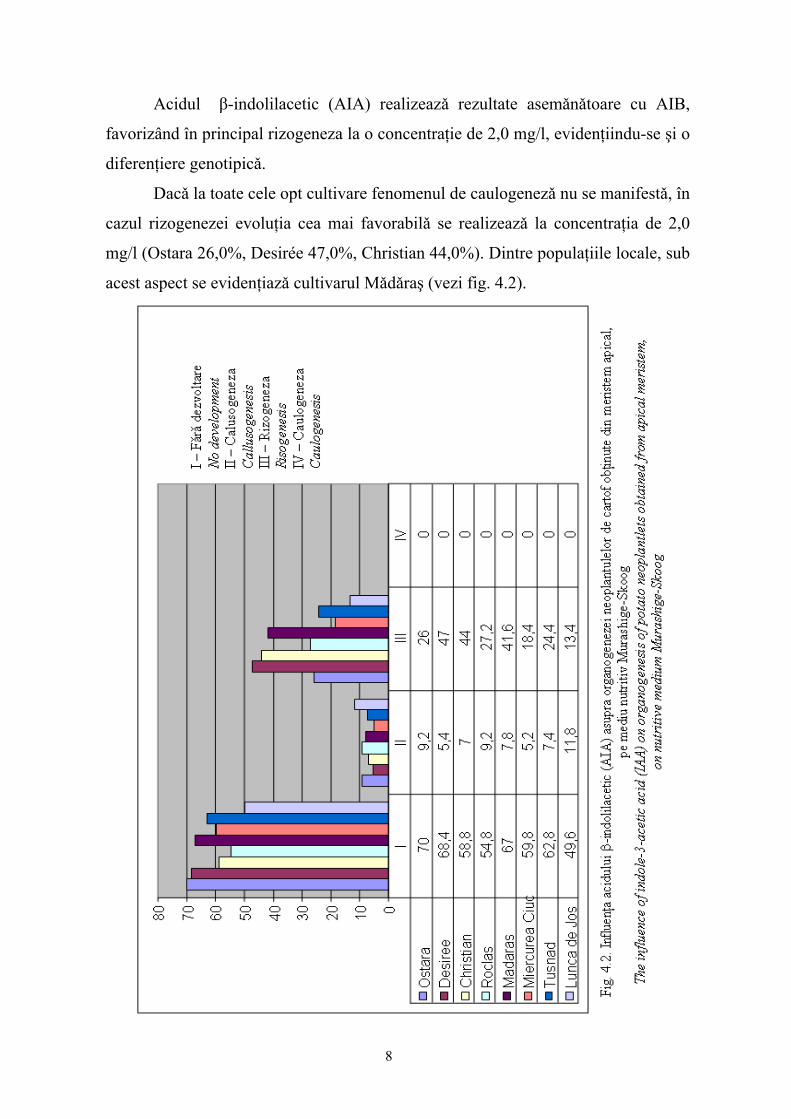
Acidul β-indolilacetic (AIA) realizeazǎ rezultate asemǎnǎtoare cu AIB,
favorizând în principal rizogeneza la o concentraţie de 2,0 mg/l, evidenţiindu-se şi o
diferenţiere genotipicǎ.
Dacǎ la toate cele opt cultivare fenomenul de caulogenezǎ nu se manifestǎ, în
cazul rizogenezei evoluţia cea mai favorabilǎ se realizeazǎ la concentraţia de 2,0
mg/l (Ostara 26,0%, Desirée 47,0%, Christian 44,0%). Dintre populaţiile locale, sub
acest aspect se evidenţiazǎ cultivarul Mǎdǎraş (vezi fig. 4.2).
8
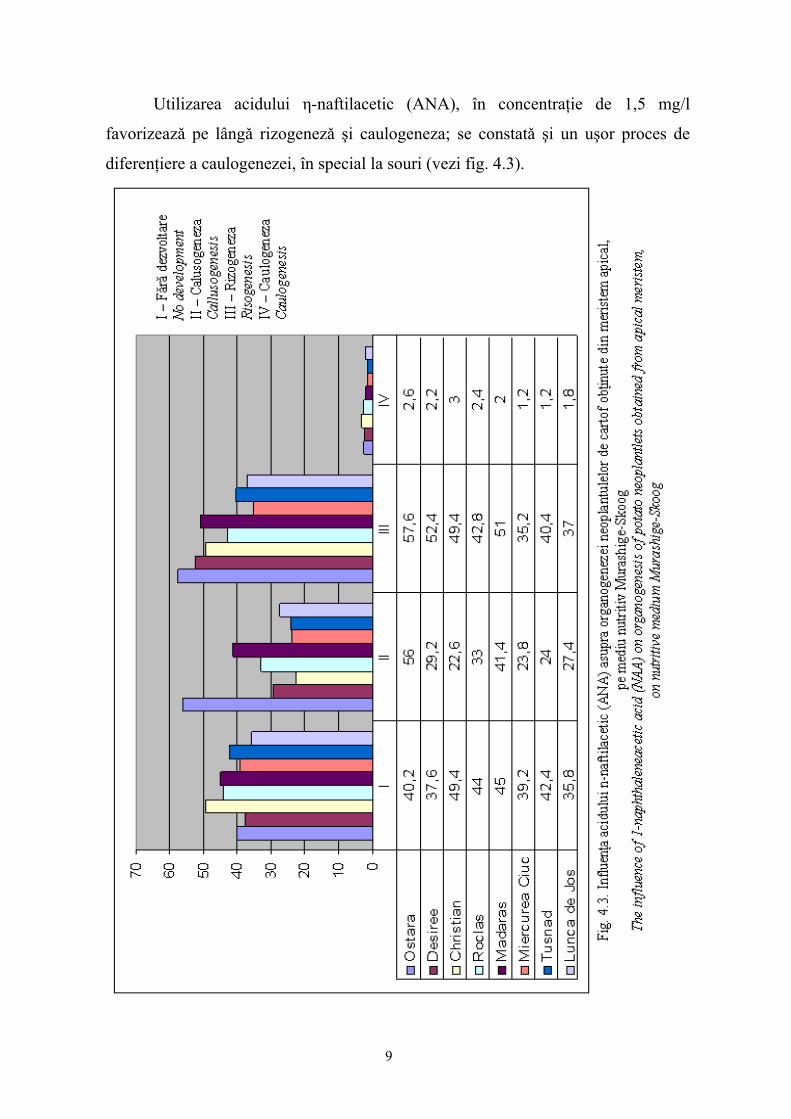
Utilizarea acidului η-naftilacetic (ANA), în concentraţie de 1,5 mg/l
favorizeazǎ pe lângǎ rizogenezǎ şi caulogeneza; se constatǎ şi un uşor proces de
diferenţiere a caulogenezei, în special la souri (vezi fig. 4.3).
9

Efectul auxinelor artificiale asupra organogenezei evidenţiazǎ diferenţe
procentuale ale organogenezei, ceea ce face sǎ se creadǎ cǎ în procesul de
multiplicare in vitro a cartofului trebuie individualizat procesul, în sensul cǎ fiecare
cultivar îşi are o reacţie proprie faţǎ de activitatea hormonalǎ, fiecare genotip
trebuie sǎ fie individualizat pentru a anumitǎ structurǎ a mediilor de culturǎ.
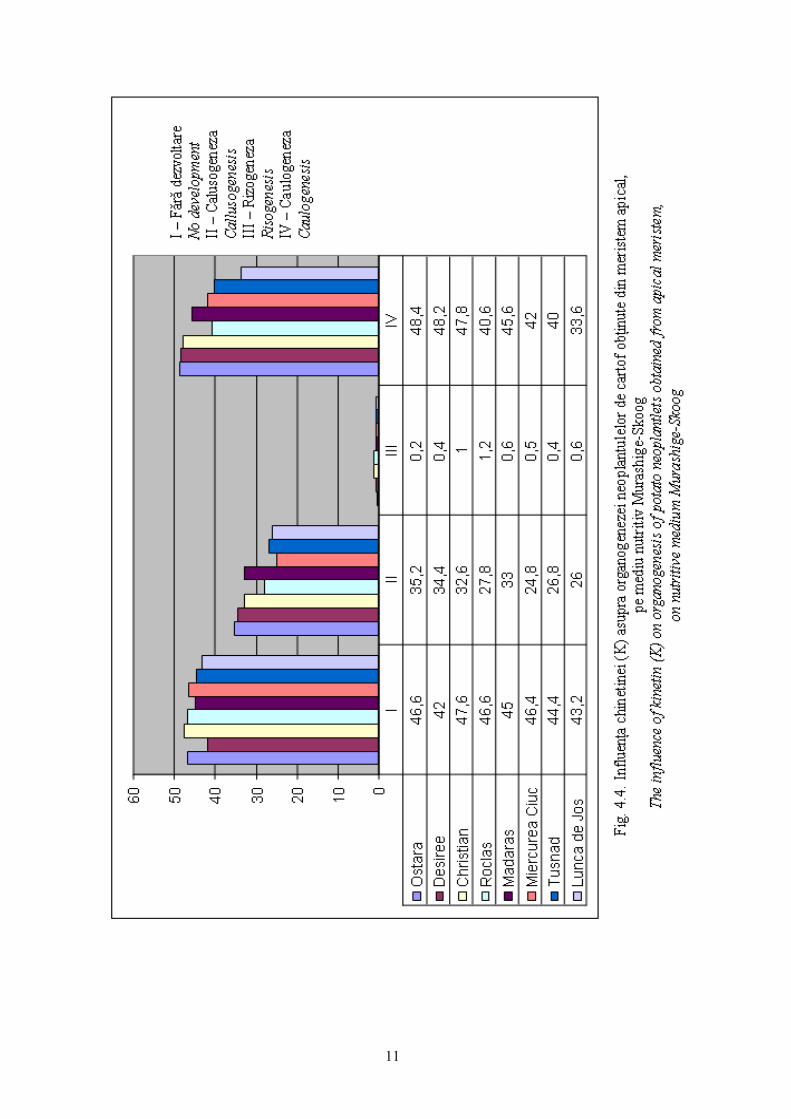
Influenţa chinetinei (K), are un efect pozitiv asupra caulogenezei, la
concentraţiile de 1,5-2,0 mg/l.
Rezultatele cele mai evidente sunt constatate la soiurile Ostara 56-66%,
Desirée 51-66% şi Christian 55-70%. Populaţia localǎ de Mǎdǎraş manifestǎ o
caulogenezǎ ridicatǎ de 52-62% (vezi fig. 4.4).
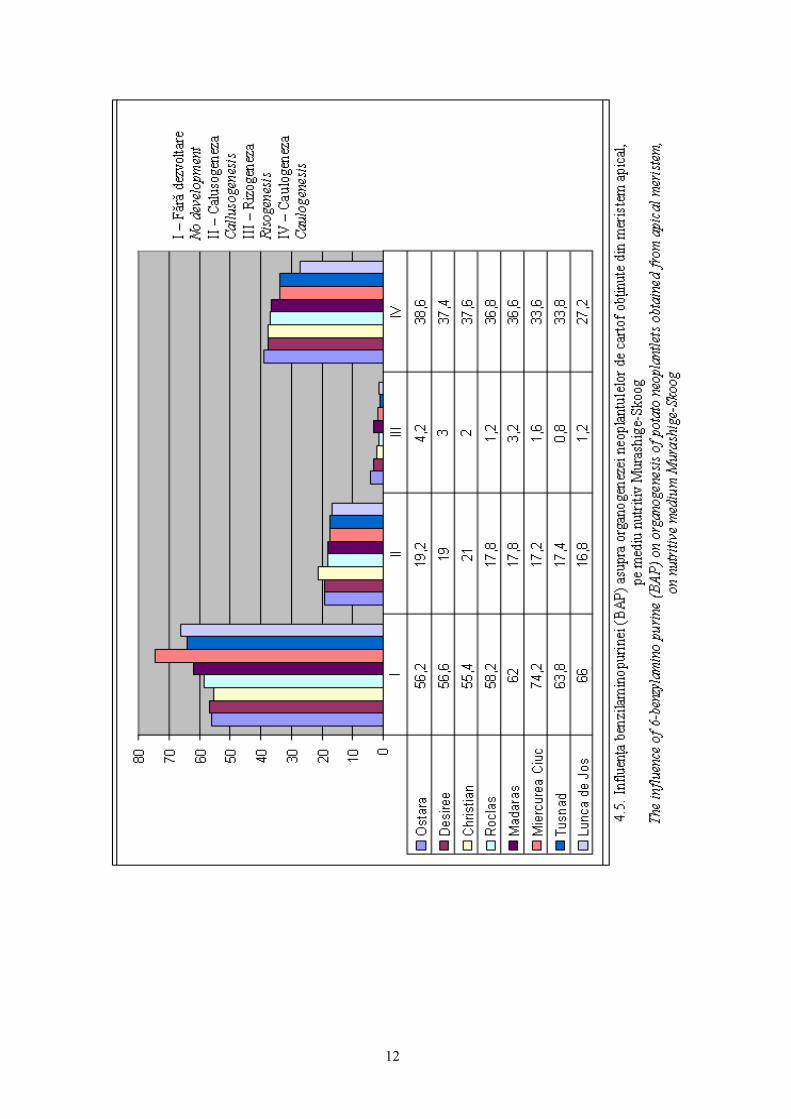
Benzilaminopurina (BAP) evidenţiazǎ practic acelaşi efect cu cel obţinut în
cazul utilizǎrii chinetinei (K), cu efecte majore asupra caulogenezei, apoi a
calusogenezei şi mai puţin asupra rizogenezei.
În cazul utilizǎrii BAP nu se evidenţiazǎ diferenţieri genotipice la nivelul
cultivarelor sub aspectul evoluţiei caulogenezei (38,6% Ostara, 37,4% Desirée,
37,6% Christian), (vezi fig. 4.5).
Fitohormonii orienteazǎ procesele de dezvoltare in vitro. O valoare ridicatǎ a
raportului auxinǎ/citochininǎ favorizeazǎ rizogeneza şi calusogeneza, iar o valoare
micǎ, duce la proliferarea mugurilor axilari şi a organogenezei.
În procesul de micromultiplicare in vitro s-a constatat cǎ mediul de culturǎ
Murashige-Skoog (1962) corespunde cerinţelor nutritive ale cartofului. Se constatǎ
cǎ apare necesitatea unei surse bogate în azot organic (glutamina) şi de fosfor
asigurat de fosfatul monosodic (NaH2PO4), la un pH de 5,8 - 6,0 - 6,5.
10
11
12
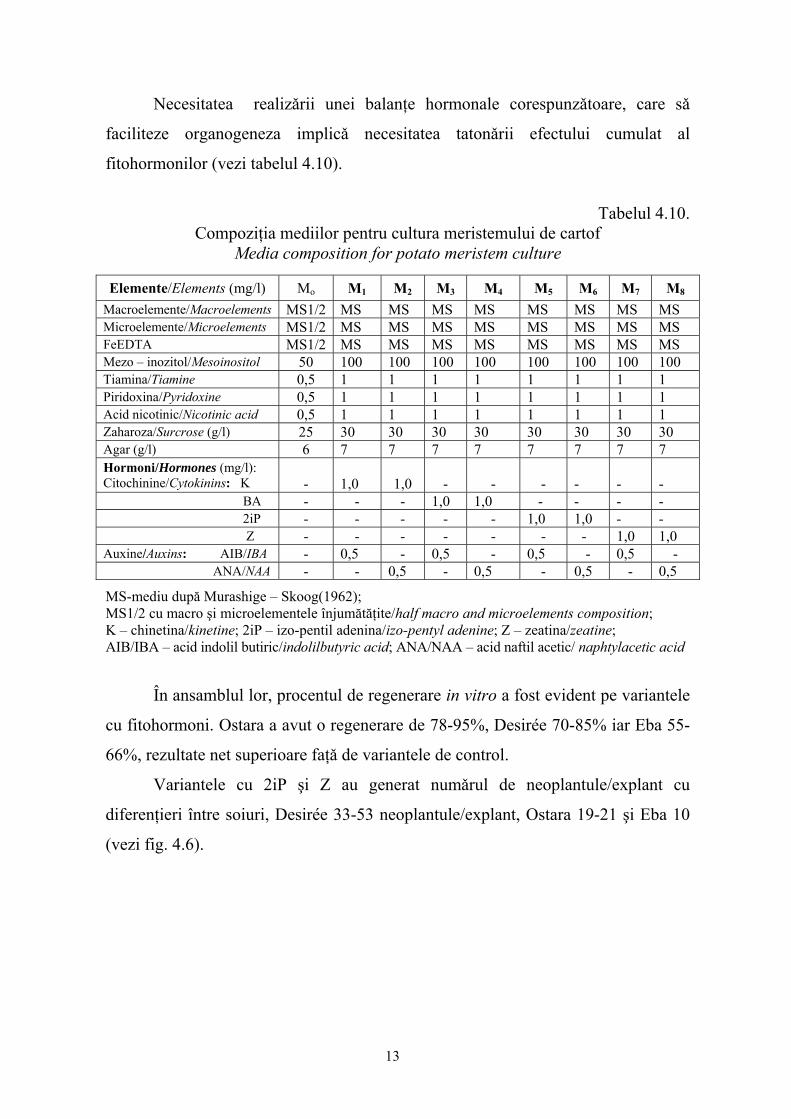
Necesitatea realizǎrii unei balanţe hormonale corespunzǎtoare, care sǎ
faciliteze organogeneza implicǎ necesitatea tatonǎrii efectului cumulat al
fitohormonilor (vezi tabelul 4.10).
Tabelul 4.10.
Compoziţia mediilor pentru cultura meristemului de cartof Media composition for potato meristem culture
Elemente/Elements (mg/l) Mo M1 M2 M3 M4 M5 M6 M7 M8
Macroelemente/Macroelements MS1/2 MS MS MS MS MS MS MS MS Microelemente/Microelements MS1/2 MS MS MS MS MS MS MS MS FeEDTA MS1/2 MS MS MS MS MS MS MS MS Mezo – inozitol/Mesoinositol 50 100 100 100 100 100 100 100 100 Tiamina/Tiamine 0,5 1 1 1 1 1 1 1 1 Piridoxina/Pyridoxine 0,5 1 1 1 1 1 1 1 1 Acid nicotinic/Nicotinic acid 0,5 1 1 1 1 1 1 1 1 Zaharoza/Surcrose (g/l) 25 30 30 30 30 30 30 30 30 Agar (g/l) 6 7 7 7 7 7 7 7 7 Hormoni/Hormones (mg/l): Citochinine/Cytokinins: K
-
1,0
1,0
-
-
-
-
-
-
BA - - - 1,0 1,0 - - - - 2iP - - - - - 1,0 1,0 - - Z - - - - - - - 1,0 1,0 Auxine/Auxins: AIB/IBA - 0,5 - 0,5 - 0,5 - 0,5 - ANA/NAA - - 0,5 - 0,5 - 0,5 - 0,5
MS-mediu după Murashige – Skoog(1962); MS1/2 cu macro şi microelementele înjumătăţite/half macro and microelements composition; K – chinetina/kinetine; 2iP – izo-pentil adenina/izo-pentyl adenine; Z – zeatina/zeatine; AIB/IBA – acid indolil butiric/indolilbutyric acid; ANA/NAA – acid naftil acetic/ naphtylacetic acid
În ansamblul lor, procentul de regenerare in vitro a fost evident pe variantele
cu fitohormoni. Ostara a avut o regenerare de 78-95%, Desirée 70-85% iar Eba 55-
66%, rezultate net superioare faţǎ de variantele de control.
Variantele cu 2iP şi Z au generat numǎrul de neoplantule/explant cu
diferenţieri între soiuri, Desirée 33-53 neoplantule/explant, Ostara 19-21 şi Eba 10
(vezi fig. 4.6).
13

Fig. 4.6. Procent de regenerare a soiurilor de cartof cultivate in vitro
Regeneration percentage of potato cultivars cultivated in vitro
Sistemul radicular este stimulat de prezenţa AIB, care în asociere cu Z şi 2iP
au generat numǎrul cel mai mare de rǎdǎcini/explant: 14 la soiul Desirée, 8-10 la
Ostara şi 7-10 la Eba (vezi fig. 4.7).
Fig. 4.7. Număr de tulpini regenerate in vitro din meristemul soiurilor de cartof
The number of in vitro regenerated stems from potato cultivars meristems
14

Se constatǎ un raport de invers proporţionalitate între perioada de vegetaţie
naturalǎ şi evoluţia in vitro a soiurilor de cartof, în favoarea celor cu perioade mai
scurte de vegetaţie (Ostara şi Desirée).
Efectuarea de subculturi se realizeazǎ în cazul cartofului prin transferarea
tulpiniţelor generate dintr-un inocul în micro- sau minibutaşi.
Pentru reuşita subculturilor in vitro, apare necesitatea cunoaşterii evoluţiei
cultivarelor la nivelul culturilor primare.
S-a evidenţiat capacitatea diferenţiatǎ a cultivarelor de a rǎspunde culturii in
vitro, cu precǎdere în favoarea soiurilor omologate.
Sub aspectul datelor procentuale medii a numǎrului de lǎstari formaţi, a
numǎrului total de internoduri soiurile Ostara, Desirée şi Christian manifestǎ o
capacitate regenerativǎ evidentǎ, cu diferenţieri faţǎ de media procentualǎ a
caraterelor menţionate, asigurate statistic.
Analiza caracterelor morfologice menţionate, la nivelul cultivarelor-populaţii
evidenţiazǎ diferenţieri marcante între acestea, dovedind heterogenitatea geneticǎ a
acestora (vezi tabelul 4.12).
15

Tabelul 4.12. Evoluţia explantelor cultivarelor de cartof in vitro The in vitro evolution of potato cultivars explants
Data obs. Ostara Desirée Christian Roclas Mǎdǎraş Tuşnad Miercurea
Ciuc Lunca de Jos
13.02 1,0 1,0 1,0 1,0 1,0 1,0 1,0 1,0 nr. lǎstarilor/ shoots number
6.03 3,2 4,4 4,8 2,8 3,2 1,2 1,4 1,0 DL 5,0 = 0,43
27.03 4,2 4,8 5,6 3,6 3,4 2,4 2,2 1,8 DL 1,0 = 1,05
17.04 4,4 4,8 5,2 3,4 5,0 3,6 2,2 2,0 DL 0,1 = 1,32
X 3,20x 3,75xx 4,15xxx 2,70 3,15x 2,050 1,700 1,4500
% 125.0 146.5 162.1 105.5 123.0 80.1 66.4 56.6 X = 2,56 = 100%
13.02 3.8 2.8 2.0 1.8 2.8 2.0 1.8 1.8 nr. noduri/lǎstar nodes no./shoot
6.03 6.7 4.9 4.8 5.7 2.7 6.1 5.3 6.3 DL 5,0 = 0,73
27.03 8.6 7.3 6.4 6.6 5.3 7.3 6.4 6.4 DL 1,0 = 1,18
17.04 8.1 7.4 7.1 6.4 4.1 8.0 6.7 6.5 DL 0,1 = 1,39
X 6.8xxx 5.60 4.75 5.12 3.72000 5.85x 5.05 5.85x
% 128.3 105.7 89.5 96.6 70.2 110.4 95.3 110.4 X = 5,30 = 100%
13.02 19.0 14.0 10.0 79.0 44.0 10.0 9.0 9.0 nr. total internoduri total no. of internodes
6.03 108.0 109.0 116.0 78.0 44.0 36.0 37.0 31.0 DL 5,0 = 20,0
27.03 180.0 174.0 180.0 118.0 90.0 88.0 70.0 57.0 DL 1,0 = 39,3
17.04 178.0 178.0 184.0 109.0 103.0 143.0 74.0 65.0 DL 0,1 = 46,4
X 121.5x 118.75x 120.0x 78.75 69.75 69.25 47.500 450.500
% 145.7 142.7 144.2 94.6 83.8 89.8 57.1 48.7 X = 83,21 = 100%
13.02 4.5 5.2 3.8 4.1 6.9 4.4 4.1 3.2 lungimea lǎstarilor shoots lenght (cm)
6.03 5.1 9.5 4.1 8.2 6.9 6.8 7.6 6.8 DL 5,0 = 0,32
27.03 7.7 12.4 7.1 8.2 6.9 7.9 7.3 6.8 DL 1,0 = 0,61
17.04 7.9 12.4 7.5 8.4 7.2 8.0 7.3 7.0 DL 0,1 = 1,93
X 6.30 9.9xxx 5.700 7.2 7.0 6.8 6.6 6.000
% 91.9 143.5 82.6 104.3 101.4 98.6 95.7 87.0 X = 6,9 = 100%
S-a remarcat existenţa unei corelaţii pozitive între perioada de vegetaţie şi
durata de timp în care meristemele s-au dezvoltat ca şi plantule. Soiul Ostara, soi
16

timpuriu, realizeazǎ o dezvoltare mai rapide a explantelor, în vitroplantule,
comparativ cu celelalte soiuri şi populaţii de cartof.
Pe parcursul a trei cicluri de subculturǎ apar diferenţieri genotipice diferite.
Cultivarul Ostara realizeazǎ cea mai bunǎ ratǎ de proliferare, de 51,0% (vezi tabelul
4.13).
17

Se constatǎ cǎ odatǎ cu avansarea în subculturǎ, apare fenomenul de
senectute, proliferǎrile având o marjǎ din ce în ce mai scǎzutǎ.
Inducerea variaţiei clonale in vitro la cartof prin culturǎ de calus depinde de
genotip, de explantul utilizat, de mediul de culturǎ şi în special de regulatorii de
creştere.
Regulatorul de creştere cu cele mai bune rezultate în inducerea calusului a
fost 2,4D (2,4-diclorofenoxiacetic), 4,0 mg/l, în combinaţia cu AIA (2,2 mg/l) şi
BAP (4 mg/l).
Explantele cu cea mai mare frecvenţǎ de calusare sunt cele provenind din
discuri de limb foliar, cultivate pe mediu de bazǎ MS (1962), fiind de 20,2% la soiul
Desirée, 18,1% la Ostara şi 7,5% la Christian. Regulatorul de creştere cu cele mai
bune rezultate în inducerea calusului a fost 2,4D (2,4-diclorofenoxiacetic), 4,0 mg/l,
în combinaţia cu ANA (2,2 mg/l) şi BAP (4 mg/l) (vezi tabelul 4.16).
Tabelul 4.16.
Frecvenţa inducerii calusului de cartof funcţie de genotip şi de tipul de inocul, dupǎ 6 sǎptǎmâni de la inoculare
Frequency of potato callus induction depending genotype and inoculum type, after 6 weeks after inoculation
Frecvenţa de calusare Callus frequency (%) Nr.
crt. Soiul
Cultivar discuri de limb foliar disk of leaf blade
segmente de peţiol petiole segments
1 Desirée 20,2 3,8
2 Ostara 18,1 2,1
3 Christian 7,5 3,0
Sub aspectul regenerǎrii de plantule la nivelul culturilor de calus, calusurile
iniţiate din diferite tipuri de explant şi subcultivate, manifestǎ o puternicǎ
interdependenţǎ între natura explantului, genotip, natura regulatorului de creştere
din substrat şi capacitatea morfogeneticǎ a calusului.
Cultivarul Desirée a manifestat cel mai activ comportament sub aspectul
regenerǎrii de plantule dupǎ subculturǎ, fenomen concretizat prin numǎrul cel mai
18
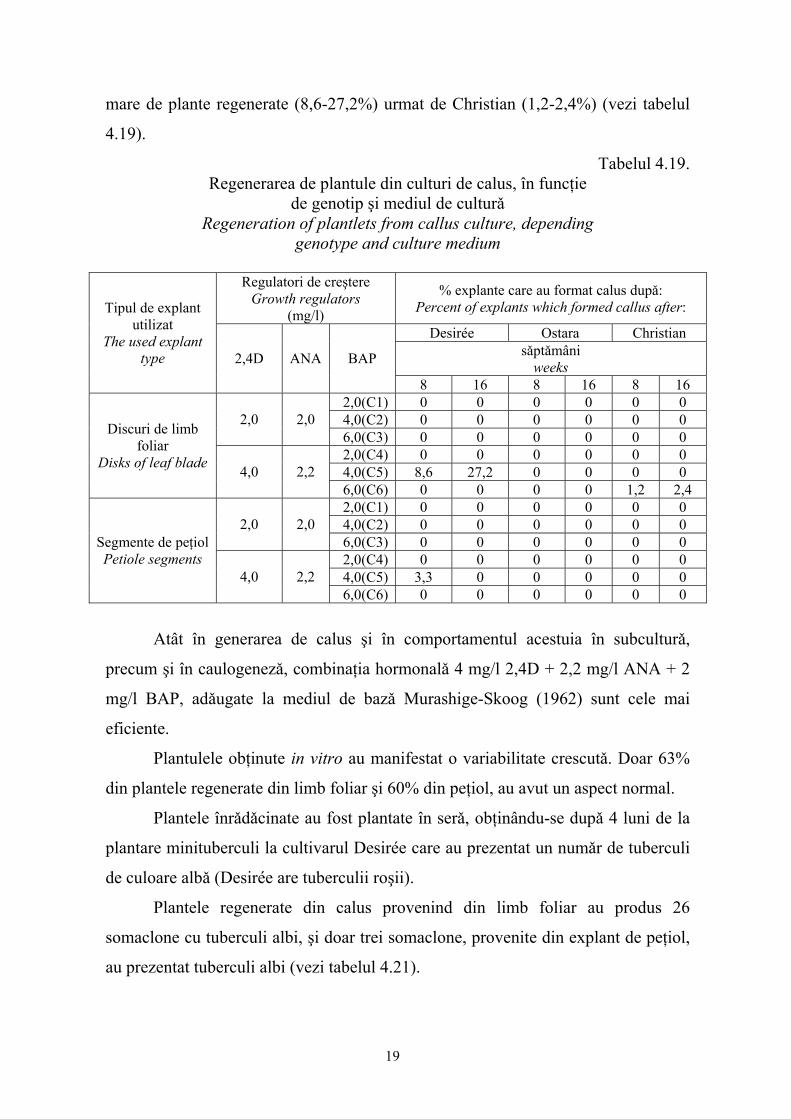
mare de plante regenerate (8,6-27,2%) urmat de Christian (1,2-2,4%) (vezi tabelul
4.19).
Tabelul 4.19. Regenerarea de plantule din culturi de calus, în funcţie
de genotip şi mediul de culturǎ Regeneration of plantlets from callus culture, depending
genotype and culture medium
Regulatori de creştere Growth regulators
(mg/l)
% explante care au format calus dupǎ: Percent of explants which formed callus after:
Desirée Ostara Christian sǎptǎmâni
weeks
Tipul de explant utilizat
The used explant type 2,4D ANA BAP
8 16 8 16 8 16 2,0(C1) 0 0 0 0 0 0 4,0(C2) 0 0 0 0 0 0 2,0 2,0 6,0(C3) 0 0 0 0 0 0 2,0(C4) 0 0 0 0 0 0 4,0(C5) 8,6 27,2 0 0 0 0
Discuri de limb foliar
Disks of leaf blade 4,0 2,2 6,0(C6) 0 0 0 0 1,2 2,4 2,0(C1) 0 0 0 0 0 0 4,0(C2) 0 0 0 0 0 0 2,0 2,0 6,0(C3) 0 0 0 0 0 0 2,0(C4) 0 0 0 0 0 0 4,0(C5) 3,3 0 0 0 0 0
Segmente de peţiol Petiole segments
4,0 2,2 6,0(C6) 0 0 0 0 0 0
Atât în generarea de calus şi în comportamentul acestuia în subculturǎ,
precum şi în caulogenezǎ, combinaţia hormonalǎ 4 mg/l 2,4D + 2,2 mg/l ANA + 2
mg/l BAP, adǎugate la mediul de bazǎ Murashige-Skoog (1962) sunt cele mai
eficiente.
Plantulele obţinute in vitro au manifestat o variabilitate crescutǎ. Doar 63%
din plantele regenerate din limb foliar şi 60% din peţiol, au avut un aspect normal.
Plantele înrǎdǎcinate au fost plantate în serǎ, obţinându-se dupǎ 4 luni de la
plantare minituberculi la cultivarul Desirée care au prezentat un numǎr de tuberculi
de culoare albǎ (Desirée are tuberculii roşii).
Plantele regenerate din calus provenind din limb foliar au produs 26
somaclone cu tuberculi albi, şi doar trei somaclone, provenite din explant de peţiol,
au prezentat tuberculi albi (vezi tabelul 4.21).
19

Tabelul 4.21.
Rezultate privind obţinerea de minituberculi din plantulele regenerate din culturi de calus de cartof, la cultivarul Desirée
Results regarding minitubers obtaining from plantlets regenerated from potato callus cultures, to Desirée cultivar
Numǎr plantule
derivate din calus şi plantate în serǎ Number of plantlets derivated from callus and planted in green house
Numǎr varianţi somaclonali cu tuberculi albi
Number of somaclona variants with white tubers
Numǎr varianţi somaclonali cu tuberculi roşii
Number of somaclona variants with white tubers
discuri de limb foliar = 60 disks of leaf blade = 60 26 34
segmente de peţiol = 60 petiole segments = 60 3 57
Pentru a releva existenţa unei variaţii somaclonale s-au efectuat o serie de
analize citologice, la somaclonele provenite de la cultivarul Desirée, rezultând o
variaţie de 42-45 de cromozomi, la plantele regenerate din calus, faţǎ de 48 de
cromozomi la formele tetraploide de cartof (vezi tabelul 4.22).
Tabelul 4.22.
Numǎrul de cromozomi la plantele diferenţiate din calus la cultivarul Desirée The number of chromosomes in callus differentiated plantlets of Desirée cultivar
Nr. crt.
Originea materialului Nature of material
Numǎr celule analizate The number of
amalysated cells
Numǎr cromozomi/celulǎ
Chromosomes number/cell
1 Plantǎ cu tuberculi albi, obţinutǎ din calus de limb foliar Plant with white tubers, obtained from callus of leaf blade 10 42/ cr./5
44/ cr./5
2 Plantǎ cu tuberculi albi, obţinutǎ din calus de peţiol Plant with white tubers, obtained from callus of petiole 10 44/ cr./10
3 Plantǎ cu tuberculi roşii, obţinutǎ din calus de limb foliar Plant with red tubers, obtained from callus of leaf blade 10 48/ cr./10
4 Plantǎ cu tuberculi albi, obţinutǎ din calus de peţiol Plant with white tubers, obtained from callus of petiole 10 44-45 cr./7
48 cr./3
5 Plantǎ cu tuberculi roşii, obţinutǎ din calus de peţiol Plant with red tubers, obtained from callus of petiole 10 48 cr./10
6 Plantǎ obţinutǎ din Desirée – planta mamǎ Plant obtained from Desirée – mother plant 10 48 cr./10
Datele obţinute impun observaţia cǎ în cazul culturii de calus la cartof şi
regenerarea de plante, genotipul reprezintǎ un factor esenţial în regenerarea in vitro,
iar în cadrul genotipului, sursa de explant.
20

Obţinerea de variaţii somaclonale cu unele caracteristici agronomice
favorabile pot fi utilizate în ameliorarea convenţionalǎ, constituind o sursǎ
potenţialǎ de creare de noi genotipuri de cartof.
Controlul fitosanitar efectuat prin testul ELISA, a materialului vegetal
obţinut a fost realizat în diferite stadii de creştere (plantule in vitro – vitroplantule –
şi plante în serǎ).
În urma testǎrilor efectuate la nivelul plantelor regenerate din meristeme,
privind prezenţa virusurilor X, Y, A, M, S şi VRF s-a constatat cǎ din totalul celor
80 de clone controlate, au fost lipsite de atacul virusului A şi VRF. Cultivarele
Desirée şi Eba nu au fost afectate nici de virusul Y. Un procent ridicat de plante
sǎnǎtoase (87,5%) au evidenţiat clonele faţǎ de virusul S şi 78,8% faţǎ de virusurile
X şi M (vezi tabelul 4.23).
Dupǎ circa o lunǎ de la plantarea în serǎ a materialului testat s-a efectuat al
doilea control ELISA, constatându-se o stare fitosanitarǎ corespunzǎtoare.
Microtuberizarea reprezintǎ faza tranzitorie între multiplicarea in vitro a
materialului sǎnǎtos şi înmulţirea in vivo.
Pentru microtuberizarea in vitro pot fi utilizaţi toţi microbutaşii rezultaţi la
faza de multiplicare, liberi de viroze, segmentaţi cu un nod.
Nivelul de prelevare a nodurilor din tulpiniţele vitroplantulelor, se poate
realiza pe etajele tulpinale superioare spre inferioare, constatându-se diferenţieri
privind numǎrul microtuberculilor/plantǎ la etajul superior.
Diferenţele între genotipuri, în procesul de tuberizare (Desirée, Ostara şi
Christian) în aceleaşi condiţii de culturǎ, evidenţiazǎ similitudinea comportamentalǎ
a soiurilor Desirée şi Ostara sub aspectul formǎrii numǎrului de
microtuberculi/plantǎ (1,5-1,6 inoculi pentru formarea in vitro a unui tubercul).
Soiul Christian are o capacitate mai redusǎ de tuberizare, fiind necesari 2,3 inoculi
pentru un minitubercul.
21

22

Sub aspectul greutǎţii microtuberculilor la soiul Desirée se realizeazǎ cea
mai mare greutate (0,70 g).
Varianta cea mai bunǎ s-a dovedit V4 (MS + AIB 1,6 mg/l + K 0,1 mg/l +
GA 0,1 mg/l), pe care 89% din inoculi au produs microtuberculii cu cea mai mare
greutate (0,86 g) (vezi tabelul 4.26).
Tabelul 4.26.
Influenţa compoziţiei mediilor de culturǎ asupra tuberizǎrii in vitro a cartofului, la 100 de zile de la inoculare a minibutaşilor pe mediu MS-62,
cu adaos de regulatori de creştere The influence of culture media on in vitro potato tuberisation, 100 days after
minicuttings inoculation on MS-62 medium, with growth regulators
Mediul de culturǎ MS (1962) favorizeazǎ microtuberizarea în prezenţa unei
concentraţii în zahǎr de 80 g/l şi dacǎ regulatorii de creştere prezenţi în mediul de
culturǎ sunt constituiţi din kinetinǎ 0,1 mg/l (vezi tabelul 4.28).
23
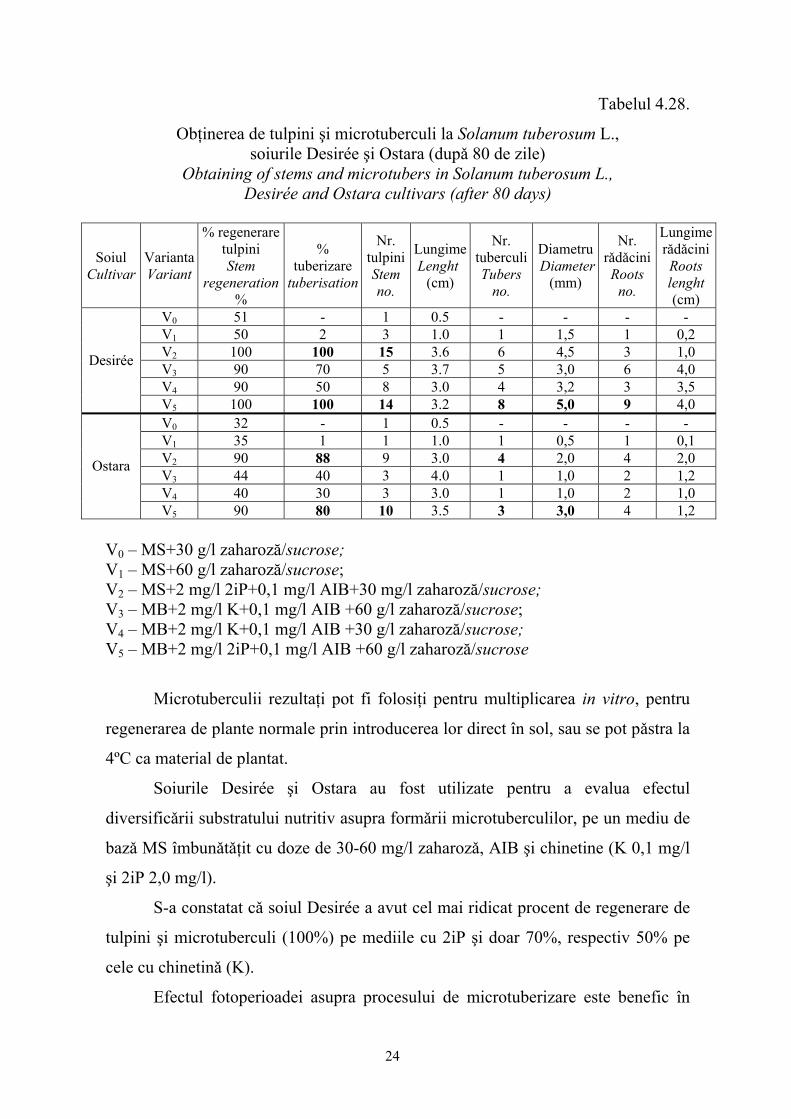
Tabelul 4.28.
Obţinerea de tulpini şi microtuberculi la Solanum tuberosum L., soiurile Desirée şi Ostara (dupǎ 80 de zile)
Obtaining of stems and microtubers in Solanum tuberosum L., Desirée and Ostara cultivars (after 80 days)
Soiul Cultivar
Varianta Variant
% regenerare tulpini Stem
regeneration %
% tuberizare
tuberisation
Nr. tulpiniStem no.
LungimeLenght
(cm)
Nr. tuberculiTubers
no.
Diametru Diameter
(mm)
Nr. rǎdǎcini Roots
no.
Lungime rǎdǎciniRoots lenght (cm)
V0 51 - 1 0.5 - - - - V1 50 2 3 1.0 1 1,5 1 0,2 V2 100 100 15 3.6 6 4,5 3 1,0 V3 90 70 5 3.7 5 3,0 6 4,0 V4 90 50 8 3.0 4 3,2 3 3,5
Desirée
V5 100 100 14 3.2 8 5,0 9 4,0 V0 32 - 1 0.5 - - - - V1 35 1 1 1.0 1 0,5 1 0,1 V2 90 88 9 3.0 4 2,0 4 2,0 V3 44 40 3 4.0 1 1,0 2 1,2 V4 40 30 3 3.0 1 1,0 2 1,0
Ostara
V5 90 80 10 3.5 3 3,0 4 1,2 V0 – MS+30 g/l zaharoză/sucrose; V1 – MS+60 g/l zaharoză/sucrose; V2 – MS+2 mg/l 2iP+0,1 mg/l AIB+30 mg/l zaharoză/sucrose; V3 – MB+2 mg/l K+0,1 mg/l AIB +60 g/l zaharoză/sucrose; V4 – MB+2 mg/l K+0,1 mg/l AIB +30 g/l zaharoză/sucrose; V5 – MB+2 mg/l 2iP+0,1 mg/l AIB +60 g/l zaharoză/sucrose
Microtuberculii rezultaţi pot fi folosiţi pentru multiplicarea in vitro, pentru
regenerarea de plante normale prin introducerea lor direct în sol, sau se pot pǎstra la
4ºC ca material de plantat.
Soiurile Desirée şi Ostara au fost utilizate pentru a evalua efectul
diversificǎrii substratului nutritiv asupra formǎrii microtuberculilor, pe un mediu de
bazǎ MS îmbunǎtǎţit cu doze de 30-60 mg/l zaharozǎ, AIB şi chinetine (K 0,1 mg/l
şi 2iP 2,0 mg/l).
S-a constatat cǎ soiul Desirée a avut cel mai ridicat procent de regenerare de
tulpini şi microtuberculi (100%) pe mediile cu 2iP şi doar 70%, respectiv 50% pe
cele cu chinetinǎ (K).
Efectul fotoperioadei asupra procesului de microtuberizare este benefic în
24

condiţiile unui mediu de culturǎ MS îmbogǎţit cu 6-10% zaharozǎ şi 1,0 mg/l
zeatinǎ, respectiv 0,5 mg/l AIB, în condiţiile unei fotoperioade de 16 ore/zi (vezi
tabelul 4.29).
Tabelul 4.29. Mediile de cultură cu diferite concentraţii de zaharoză experimentate la soiurile de cartof
Culture media with different concentration of sucrose, experimented in potato cultivars
Hormoni (mg/l) Hormones
Bonificare după Bonification after Var.
Mediul de bază Basal
medium
Concentraţia de zaharoză (%)
Sucrose concentration Z AIB/IAB 40 zile/days 80 zile/days
Vo MS Mediu/Medium : Murashige- Skoog (1962) x xx V1 MB 6 - - xx xxx V2 MB 8 - - xx xxx V3 MB 10 - - x xx V4 MB 6 1,0 0,5 xxx xxxx V5 MB 8 1,0 0,5 xxxx xxxxx V6 MB 10 1,0 0,5 xx xxx
MS - mediul Murashige-Skoog (1962); MB - mediu de bază după MS; Z - zeatine; AIB - acid indolilbutiric MS - Murashige-Skoog medium (1962); MB – basal medium after MS; Z – zeatine; IBA - indole-3-butryric acid Bonificarea: x-slab/low; xx-satisfǎcǎtor/satisfactory; xxx-bun/good; xxxx-foarte bun/very good; xxxxx-excepţional/exceptional
S-a constatat cǎ fotoperioada asociatǎ cu doza de 8% zaharozǎ şi 1,0
mg/l+0,5 mg/l AIB (V5) a stimulat tuberizarea cu 100% la soiul Ostara şi 35% la
soiul Eba (vezi fig. 4.13).
0
10
20
30
40
Vo V1 V2 V3 V4 V5 V6
Serie1
Fig. 4.13.(a) Rolul fotoperioadei şi a zaharozei asupra regenerării soiului de cartof Ostara in vitro (după 80
de zile): The role of photoperiod and sucrose on in vitro regeneration of Ostara cultivar
(after 80 days)
V0= 3%; V1=8%;V2=3,5%; V3= 6%; V4=29%; V5= 40%; V6= 21%
25

0
20
40
60
80
100
Vo V1 V2 V3 V4 V5 V6
Serie1
Fig. 4.13.(b) Rolul fotoperioadei şi a zaharozei asupra tuberizării soiul de cartof Ostara cultivat in vitro (după 80
de zile): The role of photoperiod and sucrose on in vitro tuberisation of Ostara cultivar
(after 80 days)
V0=0%; V1= 4%; V2= 10%; V3=3%; V4=80%; V5= 98%; V6= 72%
La o dozǎ de 10% zaharozǎ şi 1,0 mg/l Z+0,5 mg/l AIB tuberizarea se reduce
(18% dupǎ 80 de zile) apǎrând şi fenomenul de vitrificare.
Numǎrul de microtuberculi a oscilat în funcţie de densitatea minibutaşilor pe
vasul de culturǎ. numǎrul total de microtuberculi este mai redus la densitǎţile de 10
şi 15 plante/vas de culturǎ (301, respectiv 450), faţǎ de 5 plante/vas (520
microtuberculi).
Producerea de microtuberculi reprezintǎ o metodǎ eficace de obţinere, într-
un timp scurt, a unui material fitosanitar corespunzǎtor, metodǎ ce poate reduce
schema producerii de sǎmânţǎ cu 3-4 ani.
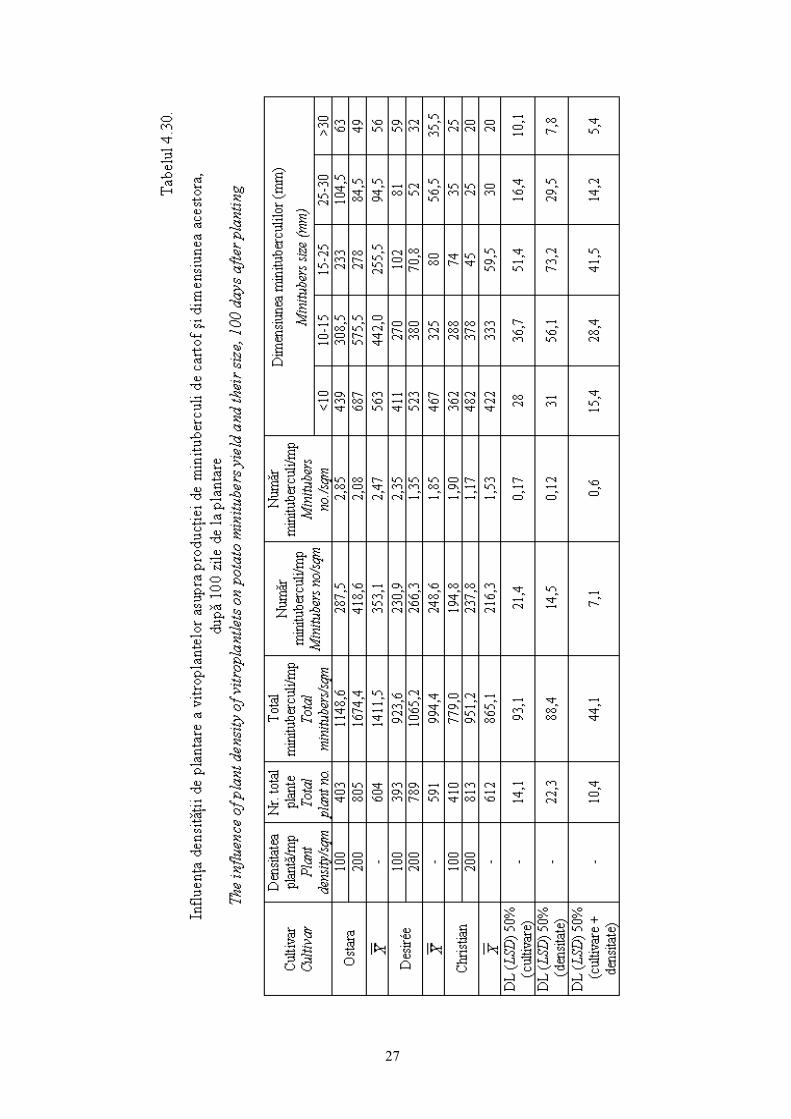
Producţia de minituberculi obţinuţi dupǎ 100 de zile de la plantarea în serǎ a
vitroplantulelor a fost influenţatǎ atât de densitatea de plantare cât şi de genotip.
În cazul densitǎţii de 200 plante/mp, numǎrul total de minituberculi a fost
mai mare decât în cazul densitǎţii de 100 plante/mp. în medie la 200 plante/mp se
obţin 307,6 minituberculi faţǎ de 237,7 la densitatea de 100 plante/mp.
Reacţia diferitǎ a genotipurilor este evidentǎ: cultivarul Ostara a produs cei
mai mulţi minituberculi/total (1411,5), comparativ cu Desirée (951,8) şi Christian
(353,1), (vezi tabelul 4.30).
26
27

Diferenţele sunt condiţionate de capacitatea de tuberizare, soiul Ostara având
o medie de 2,47 minituberculi/plantǎ, Desirée, 1,85 şi Christian 1,53.
Se pare cǎ, capacitatea de tuberizare se coreleazǎ cu caracterul de
precocitate.
Cele douǎ densitǎţi de plantare (100 plante/mp şi 200 plante/mp) au
influenţat evident nu numai numǎrul total de minituberculi obţinuţi per plantǎ, dar şi
procentul de minituberculi de diferite dimensiuni.
În cazul densitǎţii de 100 plante/mp procentul de minituberculi mai mari de
15 mm este de 34,1%, scǎzând la 24,5% la 200 plante/mp.
Minituberculii mai mici de 15 mm au un procent mai ridicat odatǎ cu
creşterea densitǎţii plantelor, 65% la densitatea de 100 plante/mp la 75,3% la 200
plante/mp.
Faptul cǎ la cele trei cultivare procentul de minituberculi >15 mm la 100 de
zile de la plantarea vitroplantulelor în sol a fost sub 40% indicǎ existenţa unor
resurse endogene, apǎrând necesitatea continuǎrii cercetǎrilor privind condiţiile de
nutriţie şi de minituberizare, condiţionate genotipic (vezi fig. 4.17-4.19).
28

100 plante/mp >15 mm = 34.1% <15 mm = 65.0%
38.2
26.8
20.3
8.35.5
<10 mm10-15 mm15-25 mm25-30 mm>30 mm
200 plante/mp >15 mm = 24.5% <15 mm = 75.3%
41
34.3
16.6
5 2.9
<10 mm10-15 mm15-25 mm25-30 mm>30 mm
Fig. 4.17. Influenţa densitǎţii de plantare asupra procentului de minituberculi de diferite mǎrimi, la soiul Ostara
The influence of planting density on diffrent size minitubers percentage, Ostara cultivar
29
100 plante/mp >15 mm = 26.2% <15 mm = 73.6%
445.5
29.2
11
8.8
6.4
<10 mm10-15 mm15-25 mm25-30 mm>30 mm
200 plante/mp >15 mm = 14.5% <15 mm = 84.8%
49.1
35.7
6.64.9 3
<10 mm10-15 mm15-25 mm25-30 mm>30 mm
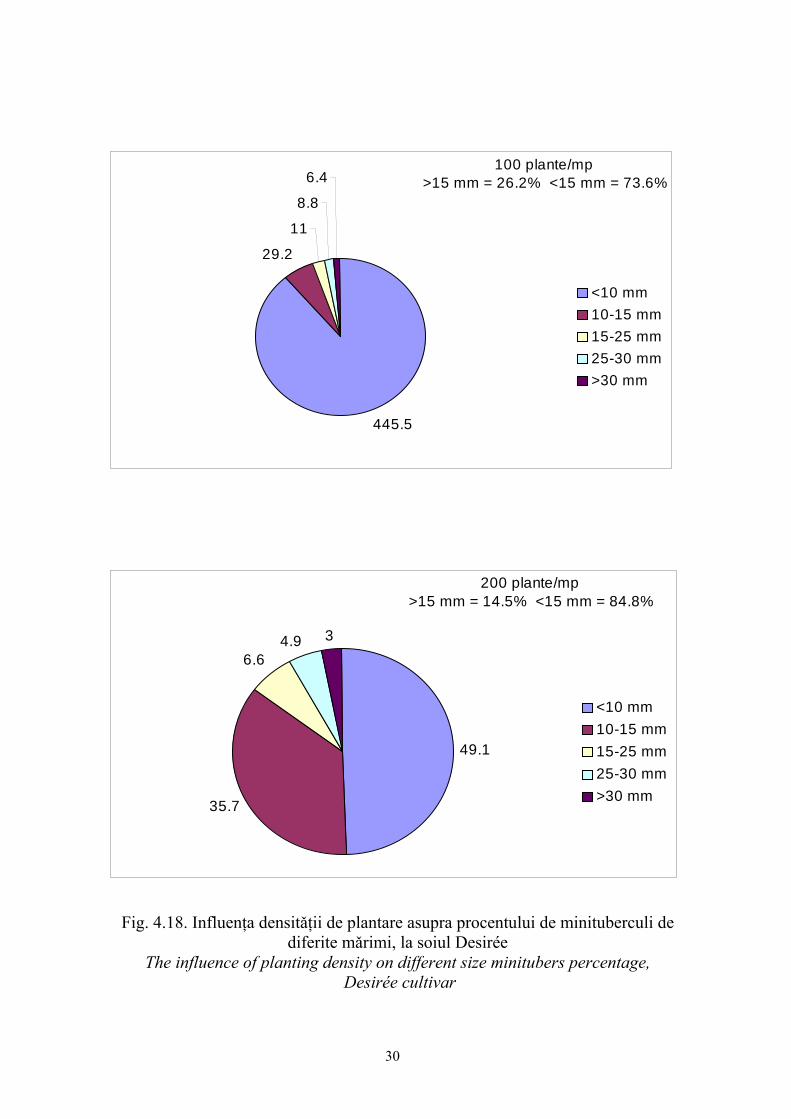
Fig. 4.18. Influenţa densitǎţii de plantare asupra procentului de minituberculi de diferite mǎrimi, la soiul Desirée
The influence of planting density on different size minitubers percentage, Desirée cultivar
30

100 plante/mp >15 mm = 16.6% <15 mm = 83.5%
46.5
37
9.54.5 2.6
<10 mm10-15 mm15-25 mm25-30 mm>30 mm
200 plante/mp >15 mm 9.4% <15 mm = 90.4%
50.939.7
4.7
2.6
2.1
<10 mm10-15 mm15-25 mm25-30 mm>30 mm
Fig. 4.19. Influenţa densitǎţii de plantare asupra procentului de minituberculi de diferite mǎrimi, la soiul Christian
The influence of planting density on different size minitubers percentage, Christian cultivar
31

Determinarea diversitǎţii genetice a cultivarelor de cartof s-a realizat cu
ajutorul markerilor RAPD urmǎrindu-se identificarea variabilitǎţii clonale şi a
distanţelor genetice la cele opt cultivare de cartof utilizate în studiile noastre.
Pentru realizarea dezideratului de mai sus au fost utilizaţi 26 de primeri
RAPD. Cele mai multe benzi polimorfice au fost obţinute cu primerul OPH12 (14
benzi), iar cele mai puţine cu primerii OPB17, OPF02 şi OPC08 (o bandǎ).
Numǎrul total de benzi obţinute cu cei 26 de primeri utilizaţi pentru analiza
RAPD a celor opt probe a fost de 269, dintre care 111 au fost monomorfice şi 158
polimorfice, media fiind de 6 benzi polimorfice/primer (vezi tabelul 5.2).
Tabelul 5.2. Rezultatele obţinute în urma amplificării celor 8 probe din specia Solanum
tuberosum cu 26 de primeri RAPD Results obtained after amplification of 8 samples from Solanum tuberosum
with 26 RAPD primers
Nr. crt. Primer Primer
Benzi polimorfice Pholymorphic bands
1 OPC 08 10 2 OPC 02 0 3 OPA 01 0 4 OPH 15 9 5 OPF 02 1 6 OPA 02 4 7 OPF 13 3 8 OPA 20 8 9 OPD 19 4
10 OPD 20 3 11 OPC 14 8 12 OPA 09 7 13 OPAL 20 7 14 OPF 20 13 15 OPA 03 5 16 OPB 12 6 17 OPAB 18 6 18 OPAB 11 1 19 OPB 17 4 20 OPH 02 12 21 OPB 09 9 22 OPG 07 6 23 OPB 10 5 24 OPB 11 7 25 OPF 16 6 26 OPH 12 14
32
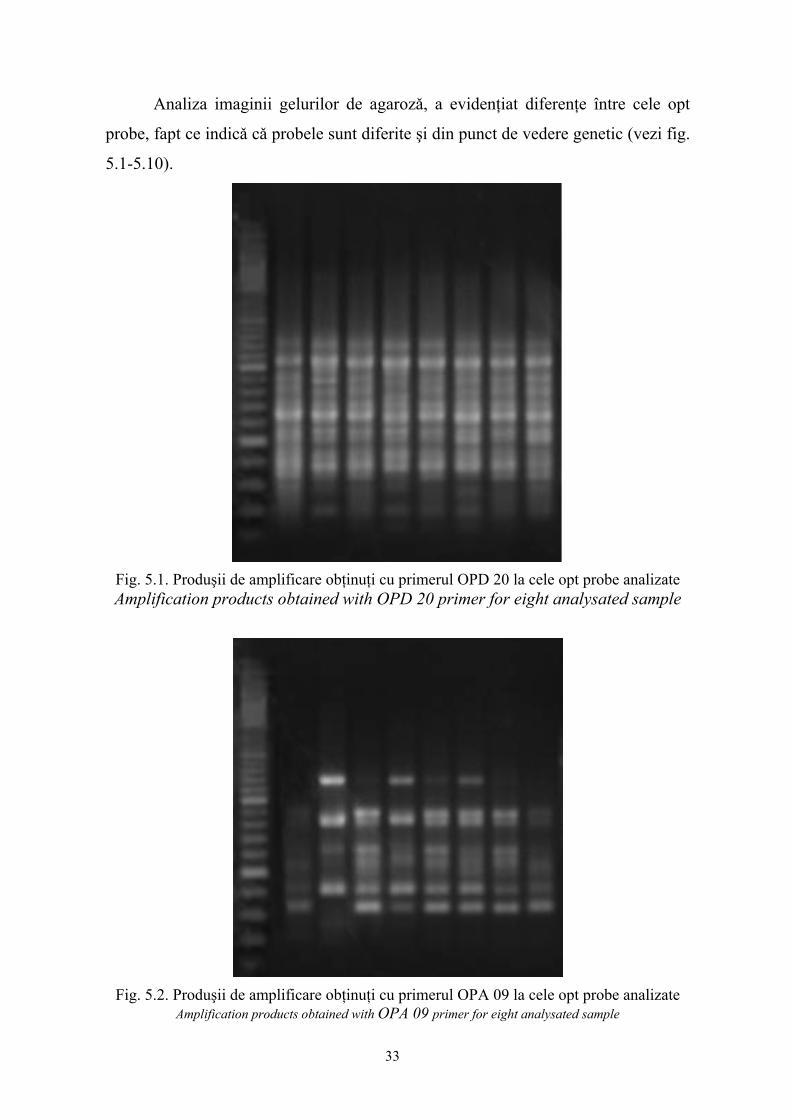
Analiza imaginii gelurilor de agarozǎ, a evidenţiat diferenţe între cele opt
probe, fapt ce indicǎ cǎ probele sunt diferite şi din punct de vedere genetic (vezi fig.
5.1-5.10).
Fig. 5.1. Produşii de amplificare obţinuţi cu primerul OPD 20 la cele opt probe analizate Amplification products obtained with OPD 20 primer for eight analysated sample
Fig. 5.2. Produşii de amplificare obţinuţi cu primerul OPA 09 la cele opt probe analizate Amplification products obtained with OPA 09 primer for eight analysated sample
33
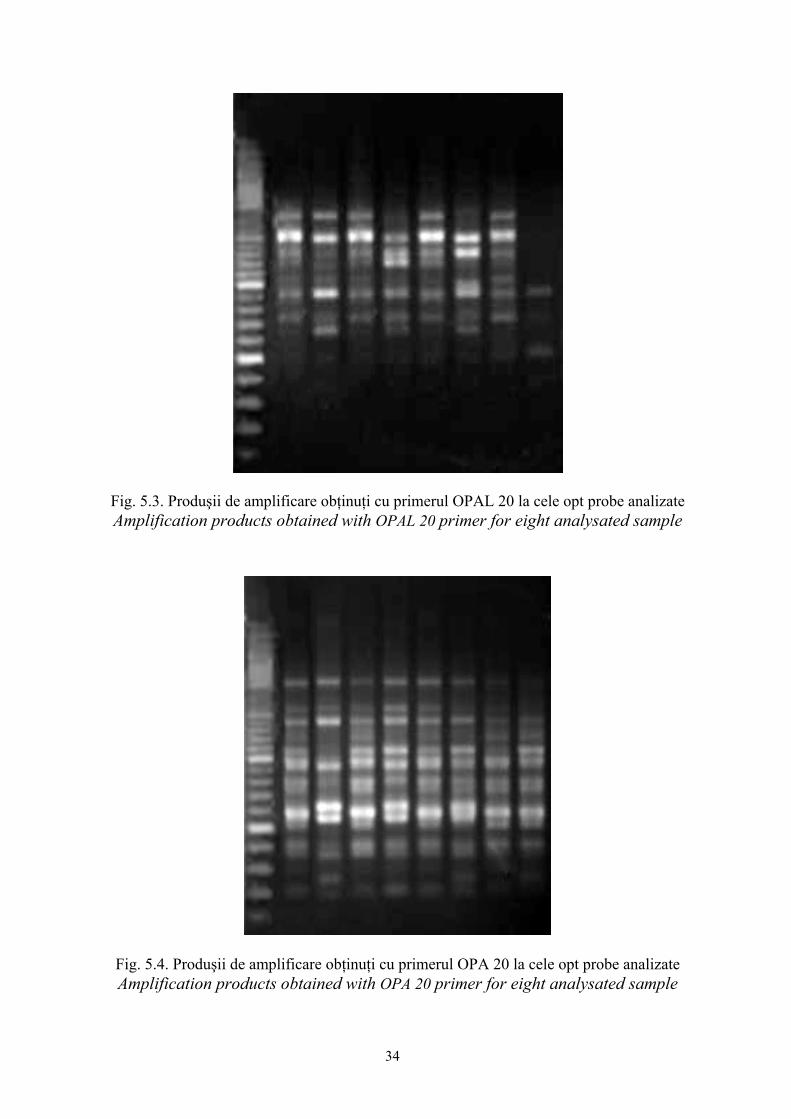
Fig. 5.3. Produşii de amplificare obţinuţi cu primerul OPAL 20 la cele opt probe analizate Amplification products obtained with OPAL 20 primer for eight analysated sample
Fig. 5.4. Produşii de amplificare obţinuţi cu primerul OPA 20 la cele opt probe analizate Amplification products obtained with OPA 20 primer for eight analysated sample
34
Fig. 5.5. Produşii de amplificare obţinuţi cu primerul OPF 20 la cele opt probe analizate Amplification products obtained with OPF 20 primer for eight analysated sample
Fig. 5.6. Produşii de amplificare obţinuţi cu primerul OPF 13 la cele opt probe analizate Amplification products obtained with OPF 13 primer for eight analysated sample
35
Fig. 5.7. Produşii de amplificare obţinuţi cu primerul OPB 11 la cele opt probe analizate Amplification products obtained with OPB 11 primer for eight analysated sample
Fig. 5.8. Produşii de amplificare obţinuţi cu primerul OPB 17 la cele opt probe analizate Amplification products obtained with OPB 17 primer for eight analysated sample
36
Fig. 5.9. Produşii de amplificare obţinuţi cu primerul OPA 03 la cele opt probe analizate Amplification products obtained with OPA 03 primer for eight analysated sample
Fig. 5.10. Produşii de amplificare obţinuţi cu primerul OPC 08 la cele opt probe analizate Amplification products obtained with OPC 08 primer for eight analysated sample
37

Analiza distanţelor genetice, a evidenţiat faptul cǎ cele mai apropiate probe
sub aspect genetic sunt Miercurea Ciuc şi Tuşnad, şi cele mai îndepǎrtate sunt cele
înregistrate între grupul de referinţǎ (outgroup) şi restul probelor (vezi tabelul 5.3).
38

Relaţiile dintre probele analizate evidenţiazǎ în dendrograma apǎrutǎ în urma
analizei cu primerii RAPD, formarea a douǎ grupuri, unul constituit din Desirée,
Mǎdǎraş, Lunca de Jos, Miercurea Ciuc şi Tuşnad, iar celǎlalt din Roclas, Christian
şi Ostara (vezi fig. 5.11).
Fig. 5.11. Dendrograma, generată de programul TreeView (PAGE, 1996), alcătuită pe baza distribuţiei genetice, după metoda UPGMA, utilizând coeficientul Nei-Li/Dice cu ajutorul
programului FreeTree (HAMPl şi colab., 2001) Dendogram, generated by TreeView program (PAGE, 1996), formed on base of genetic
distribution, after UPGMA method, using Nei-Li/Dice coefficient with the aid of FreeTree program
39

Apropierea cultivarului Desirée de populaţiile locale de cartof, nu indicǎ
obligatoriu cǎ Desirée s-ar constitui într-un genitor al populaţiilor ci mai degrabǎ,
acest fapt se datoreşte impurificǎrii biologice a populaţiilor cu soiul cultivat.
40

B I B L I O G R A F I E S E L E C T I VĂ
1. AGUD, ELIZA, M. SAVATTI, MARIA ZǍPÂRŢAN, 2008, The Growth Hormones Involved in the In Vitro Tuberisation of Some Potato Cultivars, În: Analele Universitǎţii din Oradea, Fascicula: Protecţia Mediului, vol. XIII, Ed. Universitǎţii din Oradea
2. AGUD, ELIZA, MARIA ZǍPÂRŢAN, M. SAVATTI, 2009, The in vitro Regenerative Capacity of the Potato Cultivars Ostara, Desirée and Eba Meristem, În: Agricultura – revistǎ de ştiinţǎ şi practicǎ agricolǎ, anul XVIII, nr. 1-2, Ed. AcademicPres Cluj-Napoca
3. AGUD, ELIZA, 2009, The in vitro multiplication of Eba potato cultivar, În: Agricultura – revistǎ de ştiinţǎ şi practicǎ agricolǎ, anul XVIII, nr. 3-4, Ed. AcademicPres Cluj-Napoca
4. AGUD, ELIZA, MARIA ZǍPÂRŢAN, M. SAVATTI, Z. CAP, 2009, The Effect of the Photoperiod and of the Dose of Sucrose from the Environment Over some Potatoes Varieties Cultivated In Vitro, În: Analele Universitǎţii din Oradea, Fascicula: Protecţia Mediului, vol. XIV, Ed. Universitǎţii din Oradea
5. AGUD, ELIZA, M. SAVATTI, E. PANTEA, MARIA ZǍPÂRŢAN, Hormonii de creştere implicaţi în tuberizarea in vitro la unele soiuri de cartof, sub tipar la Universitatea din Oradea
6. ARDELEAN, M., R. SESTRAŞ, MIRELA CORDEA, 2002, Tehnicǎ experimentalǎ horticolǎ, Ed. AcademicPres, Cluj-Napoca
7. ARDELEAN, M., C. BOTEZ, M. SAVATTI, ELENA TĂMAŞ, M. SAVATTI jr., R. SESTRAŞ, MIRELA MÜLLER, 1995, Impactul biotehnologiilor moderne asupra creǎrii de noi soiuri de plante de culturǎ, Bul. USAMV, Cluj-Napoca, A-H, 49/2
8. BUTIUC, A.L, MARIA ZǍPÂRŢAN şi T. BORZA, 1996, Rolul unor citochinine în inducerea şi creşterea minituberculilor obţinuţi in vitro la soiul de cartoof Desirée, în: Analele Universitǎţii din Oradea, Fascola biologie, Tom III
9. BUTIUC-KEUL, A., C. MUNTEANU-DELIU, E. SZABO, S. MOCAN, C. DELIU, 1997-1998. In vitro induction and development of micro tubers in potato (Solanum tuberosum L.) I. Effects of growth regulators and sucrose concentration, În: Contribuţii Botanice, II, Grǎdina Botanicǎ, Cluj-Napoca
10. BUTIUC-KEUL, ANCA, 2006. Markeri moleculari utilizaţi în geneticǎ şi biotehnologia vegetalǎ, Ed. Mega Cluj-Napoca
41

11. CACHIŢǍ-COSMA, DORINA, MARIA ZǍPÂRŢAN, 1991. Potato tuberogenesis using in vitro bi-layer technique, In: In vitro explants cultures – present and perspective, The IVth National Symposium on Plant Cell land and Tissue Culture, Cluj-Napoca
12. CACHIŢǍ-COSMA, DORINA, C. DELIU, LENUŢA RAKOSY-TICAN, A. ARDELEAN, 2004. Tratat de biotehnologie vegetalǎ, vol. 1, Ed. Dacia Cluj-Napoca
13. CACHIŢǍ-COSMA, DORINA, 2007. Micropropagarea speciilor de interes economic prin utilizarea de dispozitive automate sau de roboţi, În: Micropropagarea speciilor vegetale, Lucrǎrile celui de al XV-lea Simpozion Naţional de Culturi de Ţesuturi şi Celule Vegetale, Iaşi, iunie 2006, Ed. Risoprint Cluj-Napoca
14. CACHIŢǍ-COSMA, DORINA, A. ARDELEAN, 2009. Tratat de biotehnologie vegetalǎ, vol. 2, Ed. Dacia Cluj-Napoca
15. CACHIŢĂ-COSMA, DORINA, 1987, Metode in vitro la plantele de culturǎ, Ed. Ceres, Bucureşti
16. CACHIŢĂ-COSMA, DORINA, A. ARDELEAN, C. CRĂCIUN, 1997, Actualitate şi pespective în biotehnologiile vegetale, Ed. V. Goldiş, Arad
17. CACHIŢĂ-COSMA, DORINA, CAMELIA SAND, 2000, Biotehnologie vegetalǎ, vol. I, Ed. Mira Design, Sibiu
18. PAMFIL, D., 1985, Regenerarea de garoafe din cultura de calus, Lucrǎrile celui de al III-lea Simpozion Naţional de culturi de celule şi ţesuturi vegetale, Bucureşti
19. PANFIL, D., 1980, Cultura de meristeme in vitro, Horticultura 3
20. SAVATTI, M., MARIA ZĂPÂRŢAN, 1991, Capacitatea de regenerare in vitro a unor tipuri de explante de vârste diferite la sparcetǎ (O viciifolia Scop.), Bul. IACN, A-H, 45/1
21. SAVATTI, M., M. SAVATTI jr., L. MUNTEAN jr., 2003, Ameliorarea plantelor – teorie şi practicǎ, Ed. AcademicPres, Cluj-Napoca
22. SAVATTI, M., G. NEDELEA, M. ARDELEAN, 2004, Tratat de ameliorarea plantelor, Ed. Marineasa, Timişoara
23. ZǍPÂRŢAN, MARIA, DORINA CACHIŢǍ-COSMA, C. DRAICA, 1991. Potato shoot behaviour on media containing zeatin, coumarin or cicocel (CCC), În: In vitro explant cultures – present and perspective, The IVth National Symposium on Plant Cell land and Tissue Culture, Cluj-Napoca
24. ZǍPÂRŢAN, MARIA, 1992. In vitro tuberizationin some potato cultivars, În: Studia Univ. Babeş-Bolyai, Biologia, XXXVII, 2.
42

UNIVERSITATEA DE ŞTIINŢE AGRICOLE ŞI MEDICINǍ VETERINARĂ CLUJ-NAPOCA
ŞCOALA DOCTORALǍ FACULTATEA DE AGRICULTURĂ
Ing. ELIZA MARIA AGUD
Summary of the PhD thesis
The use of in vitro cultures in potato (Solanum tuberosum L.) multiplication and breeding
Conducător ştiinţific:
Prof. univ. dr. MIRCEA SAVATTI
CLUJ-NAPOCA
2010
43

The use of in vitro cultures in potato (Solanum tuberosum L.) multiplication and breeding
Summary of the PhD thesis
Most of the research that has been carried out in the field of vegetal vitro-
cultures over a century has revealed essential aspects related to the regeneration, the
growth, the organogenesis, the somatic embryogenesis and other phenomena
manifested in tissue structures or cell types, in relation with the composition of the
culture medium and the eco-physiological conditions in the culture containers or in
the growth chambers. However, there are still numerous aspects that have remained
unknown; some refer to the factors involved in morphogenesis (especially at cell
and molecular level), some others refer to the transmission of the newly acquired
genetic information.
In vitro cultures have found, in a relatively short time, several practical
applications in breeding a large number of crop species, including potato, regarding
the multiplication of the valuable genotypes, the propagation of virus-free stocks,
the preservation of genetical resources and so on. Moreover, the genetical variation
showed by in vitro regenerated plants (somaclonal variability) could and are used to
obtain valuable genotypes.
Starting from these statements, we tried, besides the summing up of the pros
and cons concerning the multiplication and the creation of in vitro genetic mutations
in potato, to elaborate our own views within the PhD thesis entitled: The use of in
vitro cultures in potato (Solanum tuberosum L.) multiplication and breeding.
In the present PhD thesis we tried to clarify several aspects regarding the in
vitro regeneration and multiplication possibilities of potato, the creation of genetic
variability, microtuberisation and determination of genetic diversity.
44

In order to achieve all these, our approach refers to several major aspects
which can point out the following:
- improving technologies for in vitro micropropagation of potato;
- interaction of growth hormones (IBA, IAA, NAA, K, BAP) upon
plantlets neoformation
- de culture media compared with the nutritive demands of potato
explants ;
- induction of in vitro somaclonal variation in potato
- serological testing of in vitro regenerated potato plantlets;
- in vitro obtaining of potato micro- and minitubers;
- determination of potato cultivars genetic diversity
During the elaboration of the PhD thesis we tried to elucidate the aspects
taken into study, correlating our own data with the numerous suggestions offered
by the scientific literature.
As potato is an autogamous plant with possibilities of vegetative
multiplication, the methodology we applied offers the possibility of in vitro
determination of the biological forms of potato genotype multiplication, and also
of adaptation of the neoplantlets to tuberisations processes.
In vitro regeneration and organogenesis in some species of plants is an
essential condition to accomplish vegetative multiplication. The morphogenesis and
organogenesis processes are achieved under the influence of the endogenous and
exogenous factors. The endogenous content of hormones, their type, and also the
enzymatic equipment are factors with an important role in controlling the
regeneration process. The nature of the phytohormones used, and also their
concentration, the differences of the hormonal balances have an important role in
the organogenesis processes.
The biological material used was represented by eight potato genotypes, four
cultivars Ostara, Desirée, Christian, Roclas and four local populations Mǎdǎraş,
Miercurea Ciuc, Tuşnad and Lunca de Jos , and, incidentally, the Eba cultivar .
45

The research was carried out between 2006-2009, in the Biotechnology
Laboratory of the University of Agricultural Sciences and Veterinary Medicine of
Cluj-Napoca.
The biotechnological methods, by applications of the in vitro techniques, are
a safe way for obtaining pathogen-free biological material and the quickest way for
the creation and multiplication of superior potato genotypes.
The biological material taken into consideration consisted of eight potato
cultivars with distinct morpho-physiological features (Ostara, Desirée, Christian,
Roclas, Mǎdǎraş, Miercurea Ciuc, Tuşnad, Lunca de Jos and, incidentally, Eba).
In order to create primary cultures, the mother-plants belonging to the
previously mentioned cultivars were carefully selected from the point of view of
their type, authenticity and phyto-sanitary condition. For the prelevation of
meristems and their inoculation on the culture medium to allow for the initiation of
an in vitro culture, the Murashige-Skoog basic medium was used, under aseptic
conditions and corrected with phyto-hormones in different concentrations.
The regenerative capacity of meristems to form plants or callus depended a
lot on genotype, culture medium and especially on the growth regulators.
One of our observations was that the percentage of initiated plants was
influenced by the organs from which the explants had been harvested. The best
results were obtained with explants originating from tuber shots, stem and root
fragments, while not so good percentages were obtained with leaf fragments and
leaf mesophile (Table 4.1).
The Ostara cultivar provided the highest number of initiated plants when the
explants were prelevated from growing organs, being followed by the Desiree
cultivar; the local population cultivars behaved identically to the certified cultivars.
The type, concentration and duration of the sterilizing chemical agents action
upon the organs from which the explants are prelevated have different effects. The
use of calcium hypochlorite, calcium hypochlorite + tween and sodium hypochlorite
in concentrations of 5-10% gave different results; the most efficient proved to be the
calcium hypochlorite and the calcium hypochlorite + tween with a concentration of
5% and treatment duration of 20 minutes (Table 4.3).
46

As the concentration of the disinfectant agent rises, the disinfection level
rises too but the explant viability decreases.
Auxins, cytokinins and gibberellins (less than the others) were applied for
improving the basic culture media used for the in vitro cultivation of cells and
tissues.
The auxins used for inducing cell division and risogenesis were the indole-3-
acetic acid (IAA), the indole-3-butryric acid (IBA) and the 1-naphthaleneacetic acid
(NAA), with concentrations of 0.5-2.0 mg/l. The synthesis cytokinins we used were
kinetin (K) and 6-benzylamino purine (BAP).
The interaction between the hormones used in the culture medium and their
concentration in the in vitro multiplication of potato cultivars determines significant
differences from the point of view of the plantlet neo-formation rate.
The influence of the indole-3-butryric acid (IBA) on the formation and
development of potato neoplantlets manifested itself, irrespective of genotype, upon
the risogenesis process, for concentrations of 0.5- 2.0 mg/l.
The differentiation is obvious in what regards the percentage of neoplantlets
with no development and risogenesis. The Ostara cultivar provided the lowest
percentage of developed neoplantlets (60.2%) and a risogenesis percentage of
31.8%, superior to the other cultivars (Fig. 4.1).
The indole-3-acetic acid (IAA) gave results that were similar to the AIB,
being in favour mainly to risogenesis, in concentrations of 2.0mg/l; genotypical
differentiations were also noticed in this case.
While the caulogenesis phenomenon did not occur in any of the eight
cultivars, in the case of risogenesis the most obvious evolution was obtained with
concentrations of 2.0 mg/l (Ostara 26.0%, Desirée 47.0%, Christian 44.0%). Of all
the local populations, this aspect was present in the Mǎdǎraş cultivar. (Fig. 4.2)
The use of the 1-naphthaleneacetic acid (NAA), in concentrations of 1.5
mg/l, favours both risogenesis and caulogenesis; also, a slight differentiation of
caulogenesis was noticed, mainly in the certified cultivars (Fig. 4.3).
The effect of artificial auxins on organogenesis results in percentage
differences of organogenesis, which draws to the conclusion that the in vitro
47

multiplication process must be individualised, as each cultivar has a specific
reaction to the different hormones and therefore each genotype must be
individualised for a certain structure of the culture medium.
Kinetin (K) has a positive effect on caulogenesis, when in concentrations of
1.5-2.0 mg/l. The most relevant results were obtained in Ostara 56-66%, Desirée
51-66% and Christian 55-70%.The local population of Mǎdǎraş also produced a
high rate of caulo-genesis, of 52-62% (Fig. 4.4).
In the case of 6-benzylamino purine (BAP), the effects were similar to those
noticed for kinetin (K), with a major impact on caulogenesis, callusogenesis and
then, less than on the previous ones, on risogenesis. The BAP does not produce
genotypical differentiations among the cultivars from the point of view of the
caulogenesis evolution (38.6% Ostara, 37.4% Desirée, 37.6% Christian) (Fig. 4.5).
Phyto-hormones direct the in vitro development processes. A high value of
the ratio auxin/cytokinin favours risogenesis and callusogenesis, whereas a low
value of this ratio leads to proliferation of axillary buds and organogenesis.
The evolution of the in vitro micro-multiplication process showed the fact
that the Murashige-Skoog (1962) medium suits the nutritive needs of the potato. We
noticed a need for a rich source of organic nitrogen (glutamine) and phosphorus
contained in the monosodic phosphate (NaH2PO4), with a pH of 5.8 - 6.0 - 6.5.
The need to identify the right hormonal balance, able to facilitate
organogenesis, implies the necessity to study the summed-up effect of the phyto-
hormones (Table 4.10).
On the whole, the in vitro regeneration percentage was relevant on the
variants containing phyto-hormones. Ostara had a regeneration percentage of 78-
95%, Desirée 70-85% , and Eba 55-66%, all these results being superior to those of
the control.
The variants with 2iP and Z generated differentiated numbers of
neoplantlets/explant, with Desirée -33-53 neoplantlets/explant, Ostara -19-21, and
Eba- 10 (Fig. 4.6).
48

The root system is stimulated by the presence of IBA, which, in combination
with Z and 2iP, generated the highest number of roots/explant: 14 in Desiree, 8-10
in Ostara and 7-10 in Eba (Fig. 4.7).
There is indirect correlation between the natural vegetation period and the in
vitro evolution of the potato cultivars, in favour of those with shorter vegetation
periods (Ostara şi Desirée).
Subcultures can be created in potato by transfering the generated stems from
an inocul to micro- or mini-cuttings. In order to create successful in vitro
subcultures, it is necessary to study the cultivar evolution in primary cultures.
We noticed a differentiated capacity of the cultivars to respond to in vitro
cultivation, especially in the case of the certified cultivars.
If we take into consideration the mean percentages of formed shots and the
total number of internodes, the regenerative capacity of the Ostara, Desirée and
Christian cultivars is obvious, with differentiations as compared to the mean
percentages, statistically assured, of the mentioned traits.
An analysis of the mentioned morphological traits, at the level of the local
population cultivars, shows significant differentiations among them, which proves
their genetic heterogenity (Table 4.12).
There are positive correlations between the vegetation period and the period
taken by meristems to become plantlets. The Ostara cultivar, which is an early
cultivar, displayed quicker development of the explants into vitro-plantlets, as
compared to the other potato cultivars and populations.
During the three subculture cycles there were different genotypical
differentiations. The Ostara cultivar had the best proliferation rate, of 51.0% (Table
4.13).
While going further into subculture, an aging phenomenon occurs, the
proliferation rate being lower and lower.
Inducing in vitro clonal variation in potato through callus culture depends on
genotype, type of explant, culture medium and mainly growth regulators.
49

The growth regulator with the best results in inducing callus was 2,4 D (2,4-
dichlorophenoxyacetic acid), 4.0 mg/l, in combination with IAA (2.2 mg/l) and
BAP (4 mg/l).
The explants with the most callus formation frequency are those originating
from the leaf blade discs, cultivated on MS (1962): 20.2% for Desirée, 18.1% for
Ostara and 7.5% for Christian.The growth regulator with the best results in callus
induction was 2,4D (2,4-diclorofenoxiacetic), 4.0 mg/l, in combination with NAA
(2.2 mg/l) and BAP (4 mg/l) (Table 4.16).
Calluses initiated from different types of explant and subcultivated displayed
a strong interdependence between the nature of the explant, genotype, the nature of
the growth regulator and the morpho-genetic capacity of the callus.
The Desiree cultivar was the most active from the point of view of plantlet
regeneration after subculture, providing the highest number of regenerated plants:
(8.6-27.2%), followed by Christian (1.2-2.4%) (Table 4.19).
In generating callus and using it in subcultures, as well as in caulogenesis,
the hormonal combination 4 mg/l 2,4D + 2.2 mg/l NAA + 2 mg/l BAP, added to the
basic medium Murashige-Skoog (1962) was the most effective.
The in vitro obtained plants displayed high variability. Only 63% of the
plants regenerated from leaf blade and 60% of those regenerated from petiole had a
normal aspect.
The rooted plantlets were planted in the greenhouse; four months after the
plantation, mini-tubers were obtained in the Desiree cultivar, some of them being
white in colour (unlike the usual colour for the Desiree cultivar, which is red).
The plants regenerated from callus originating from leaf blade produced 26
somaclones with white tubers, whereas only three somaclones originating from
petiole explants gave white tubers (Table 4.21).
In order to prove the existence of somaclonal variation, a number of
cytological analyses were carried out on the somaclones belonging to the Desiree
cultivar, resulting in a variation of 42-45 chromosomes in plants regenerated from
callus, as compared to 48 chromosomes in the tetraploid forms (Table 4.22).
50

The data obtained this way lead to the observation that, for potato callus
cultures and plant regeneration, the genotype represents an essential factor for the in
vitro regeneration, and, within the genotype, especially the explant source.
Obtaining somaclonal variations with cetain agronomical favourable traits is
of great importance for conventional breeding, becoming a potential source for
creating new potato genotypes.
The phyto-sanitary control of the vegetal material obtained this way, which
was carried out through the ELISA test, followed the growth stages (in vitro
plantlets- vitroplantlets – plants in the greenhouse).
As a result of the tests undertaken on plants regenerated from meristems,
regarding the presence of the X, Y, A, M, S and VRF viruses, it was noticed that all
the 80 tested clones were free of the A and VRF viruses. The Desiree and Eba
cultivars were also free of the Y virus. A high percentage of healthy plants (87.5%)
was obtained when tested for the S virus, similar to the results for the X and M
viruses- 78.8% (Table 4.23).
The second ELISA test was applied about one month after the planting in the
greenhouse and the phyto-sanitary condition was also good.
Micro-tuberisation is a transitory phase between the in vitro multiplication of
the healthy material and the in vivo multiplication.
For the in vitro micro-tuberisation, all the virus-free, one-noded mini-
cuttings resulting from the multiplication phase can be used.
The node prelevation from the little stems of the vitroplantlets can be done
starting with the superior parts of the stems, as we noticed differentiations in the
number of mini-tubers/plant in the superior part.
A comparison between the genotypes in the tuberisation process (Desirée,
Ostara şi Christian), under the same growing conditions, reveals a similar behaviour
of Desirée and Ostara in what regards the number and formation of the minitubers
(1.5-1.6 inoculs for the in vitro formation of one tuber). On the other hand, the
Christian cultivar had a reduced tuberisation capacity; 2.3 inoculs were necessary
for one mini-tuber.
51

As far as the weight of the tubers is concerned, this was the highest for the
minitubers belonging to the Desiree cultivar (0.70 g).
The best variant proved to be V4 (MS + IBA 1.6 mg/l + K 0.1 mg/l + GA 0.1
mg/l), with 89% of the inoculs producing the minitubers with the highest weight
(o.86 g) (Table 4.26).
The MS (1962) medium favours micro-tuberisation in the presence of sugar,
in concentrations of 80 g/l and if the growth regulators consist of kinetin 0.1 mg/l.
(Table 4.28)
The resulted micro-tubers can be further used for in vitro multiplication, for
the regeneration of normal plants by planting them directly into the soil, or they can
be stored at 4ºC as material to be planted.
The Desirée and Ostara cultivars were used for evaluating the effect of
diversification of the nutritive content upon micro-tuber formation, on a basic MS
medium enriched with doses of 30-60 mg/l sucrose, IBA and kinetin (K 0.1 mg/l
and 2iP 2.0 mg/l).
The Desiree cultivar provided the highest stem and micro-tuber regeneration
percentage (100%) on the 2iP media, and only 70%, respectively 50% on those with
kinetin.
The efffect of the photo-period on the micro-tuberisation process is
beneficial on an MS medium enriched with 6-10% sucrose and 0.1mg/l Z,
respectively 0.5 mg/l IBA for photo-periods of 16 hours/day. (Table 4.29)
Photo-period, in combination with 8% sucrose and 1.0 mg/l Z+0.5 mg/l IBA
(V5) stimulated tuberisation, with 100% in Ostara and 35% in Eba (Fig. 4.13).
With a dose of 10% sucrose and 1.0 mg/l Z+0.5 mg/l IBA, tuberisation is
much reduced (18% after 80 days).
The number of micro-tubers oscillated with the mini-cuttings density/pot.The
total number of micro-tubers is lower for densities of 10 and 15 plants/pot (301,
respectively 450), as compared to densities of 5 plants/pot (520 micro-tubers).
The production of micro-tubers is an effective method for obtaining, in a
short time, phyto-sanitarily suitable material, thus making it possible to reduce the
seed production scheme with 3-4 years.
52

The minituber yield, 100 days after having planted the vitro-plantlets in the
greenhouse,was influenced both by the planting density and genotype.
For 200 plants/sqm density, the total number of minitubers was higher than
with 100 plants/sqm density. As average values, we obtained 307.6 minitubers for
200 plants/sqm density, respectively 237.7 minitubers for 100 plants/sqm density.
The different response of cultivars is obvious: the Ostara cultivar produced
the most minitubers (1411.5), as compared with Desiree (951.8) and Christian
(353.1) (Table 4.30).
The differences were conditioned by the tuberisation capacity; the Ostara
cultivar produced 2.47 minitubers/plant, Desirée, 1.85 and Christian 1.53.
It seems that the tuberisation capacity is correlated with earliness traits.
The two planting densities (100 plants/sqm and 200 plants/sqm) clearly
influenced not only the total minituber number per plant, but also the percentage of
different size minitubers.
For a density of 100 plants/sqm, the percentage of minitubers bigger than 15
mm is 34.1%, respectively 24.5% for 200 plants/sqm.
The percentage of minitubers smaller than 15 mm increased with plant
density, 65% for 100 plants/sqm, respectively 75.3% for 200 plante/sqm.
The fact that the three cultivars produced only 40% minitubers bigger than
15 mm, 100 days after having planted the vitroplantlets in the soil, denotes the
existence of endogenous resources. This fact is an interesting subject for furure
research, to prove the genetic influence on minituberisation and nutrition conditions
(Fig. 4.17 - 4.19).
RAPD markers were used to identify the genetic diversity of potato cultivars,
analysing the clone variability and the genetic distances of the eight cultivars used
in our research.
A number of 26 RAPD primers were used for studying genetic diversity. The
most polymorphic bands were obtained for primer OPH12 (14 bands), and the least
for OPB17, OPF02 and OPC08 primers (one band).
53

54
The total number obtained with 26 RAPD primers was 269, of which 111
were monomorphic and 158 were polymorphic; the average was 6 polymorphic
bands/primer (Table 5.2).
The image analysis of agarose gels showed obvious differences between the
eigth samples, indicating that the samples are different genetically as well (Fig. 5.1-
5.10).
The genetic distances analysis showed the fact that the genetically closest
samples are Miercurea Ciuc and Tuşnad and the farthest are between outgroup and
the rest of the samples (Table 5.3).
The relations between samples identify two groups in the RAPD primers
dendogram, one consisted in Desirée, Mǎdǎraş, Lunca de Jos, Miercurea Ciuc and
Tuşnad and the other with Roclas, Christian and Ostara (Fig. 5.11).
The close relationship between Desirée and the potato local population, not
necessarily indicate that Desirée is a population genitor, but rather the fact that
biological contamination of population with this cultivar is present.


















