
unitatea 3 teoria cromosomiala.ppt
28
TEORIA CROMOSOMIALA A EREDITATII (MORGANISMUL) Thomas Hunt Morgan (1866-1945), în laboratorul său de la Universitatea Columbia
-
Upload
stamatin-doina -
Category
Documents
-
view
252 -
download
7
Transcript of unitatea 3 teoria cromosomiala.ppt

TEORIA CROMOSOMIALA AEREDITATII (MORGANISMUL)
Thomas Hunt Morgan (1866-1945), în laboratorul său de laUniversitatea Columbia

Mascul şi femelă de Drosophila melanogaster de tip sălbatic (Wild type)

Argumente care recomandă această specie pentru studiide genetică şi anume:a. Rapiditatea succesiunii generaţiilor (circa 10-15 zile pentru un ciclu
ontogenetic complet, la 24oC).b. Prolificitate crescută (circa 200 ouă/femelă).c. Pretenţii scăzute faţă de condiţiile de creştere (se dezvoltă bine în
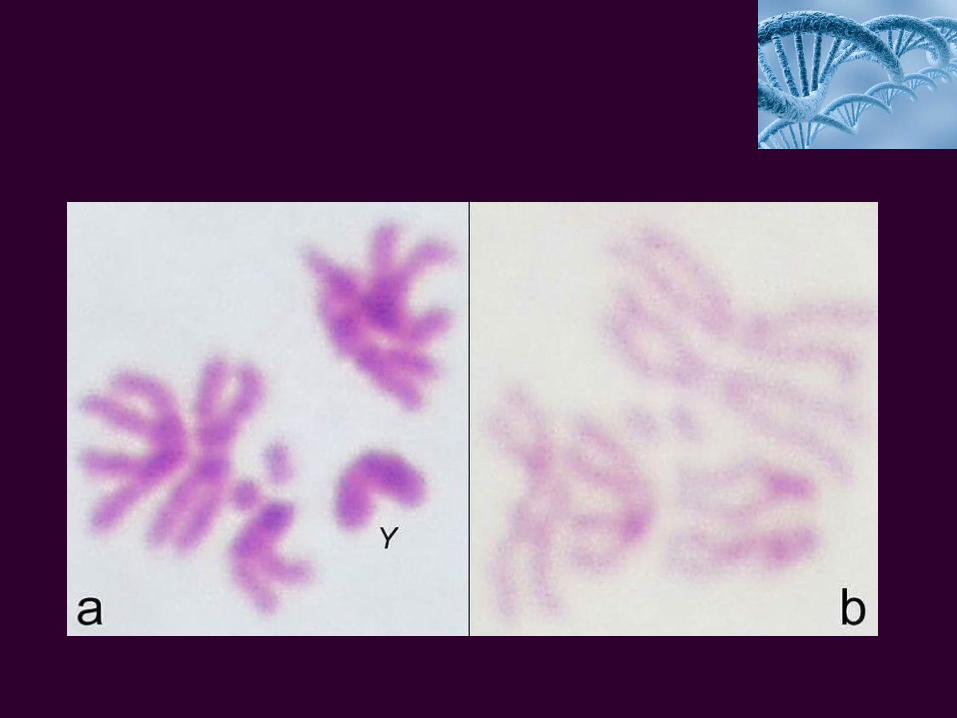
condiţii artificiale).d. Număr mic de cromosomi (2n=8) uşor identificabili. Astfel, perechea I
este alcătuită din cromosomi în formă de bastonaşe drepte la femelă şi unul în formă de bastonaş drept şi altul îndoit la un capăt, în cazul masculului. A se reţine acest aspect, ca argument pentru fundamentarea ipotezei plasării genelor pe cromosomi şi pentru considerarea respectivei perechi ca reprezentând cromosomii sexului (pentru femelă, sunt notaţi cu XX, iar pentru mascul, cu XY).
Cromosomii din perechile II şi III sunt de forma literei "V". iar cei ai perechii IV sunt punctiformi. Dacă cromosomii sexului mai poartă şi numele de heterosomi, restul sunt denumiţi autosomi.
Principala caracteristică a drosofilei care, pe lângă calităţile amintite anterior, o recomandă drept un material extrem de adecvat pentru investigaţii genetice, o constituie mutabilitatea ei ridicată (rata relativ înaltă de apariţie a mutaţiilor, atât în mod spontan, cât şi sub impactul anumitor tratamente).
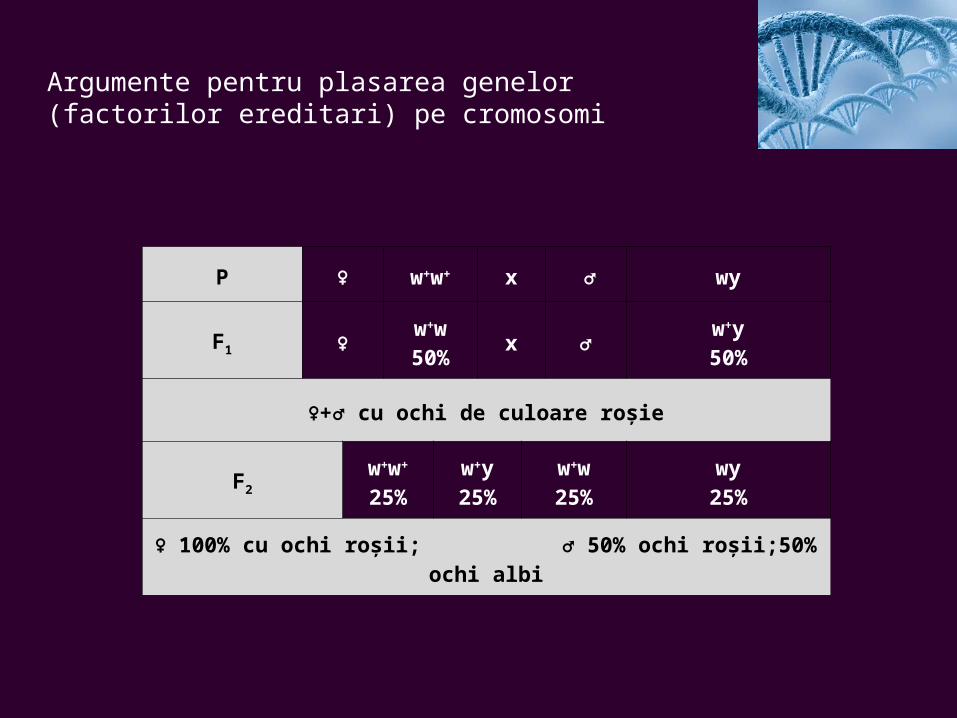
Argumente pentru plasarea genelor (factorilor ereditari) pe cromosomi
P ♀ w+w+ x ♂ wy
F1 ♀w+w50%
x ♂w+y50%
♀+♂ cu ochi de culoare roşie
F2
w+w+
25%w+y25%
w+w25%
wy25%
♀ 100% cu ochi roşii; ♂ 50% ochi roşii;50% ochi albi

Incrucisarea reciproca:
P ♀ ww x ♂ w+Y
F1 ♀ww+
50%x ♂
wY50%
♀ cu ochi roşii; ♂ cu ochi albi
F2
ww25%
w+w25%
wY25%
w+Y25%
♀50% cu ochi roşii;50% cu ochi albi♂ 50% cu ochi roşii;50% cu ochi albi

Întrucât atât masculii cât şi femelele au trei perechi identice de autosomi, diferenţele constând în prezenţa heterosomilor pereche (XX) la femele sau nepereche (XY) la masculi, rezultă că respectiva genă este plasată pe unul dintre cromosomii sexului şi anume pe cromosomul X, deoarece femelele heterozigote se comportă (fenotipic) asemenea celor homozigote dominante, în timp ce masculii (fiind hemizigoţi) se manifestă recesiv sau dominant, în funcţie de tipul cromosomului X pe care l-au primit (cu gena sălbatică sau cu alela mutantă plasată pe el).
Fenomenul, cunoscut şi sub numele de sex linkage, se constituie în cauză pentru abaterea de la tipul mendelian de segregare.
Plasarea liniară a genelor pe cromosomi
S-a efectuat o experienţă de hibridare (tip dihibridare), între indivizi de D. melanogaster ce difereau între ei prin culoarea ochilor şi forma aripilor.
Femela era fenotipic normală (pr+pr+vg+vg+), iar masculul dublu mutant - avea ochii purpurii (prpr) şi aripile vestigiale (vgvg).
În F1 s-au obţinut doar indivizi fenotipic normali.
Prin încrucişarea lor, în F2, în loc să se obţină segregarea de tip mendelian, de 9:3:3:1, s-au obţinut doar două categorii de fenotipuri - 75% de tip parental dominant şi 25% de tip parental recesiv. Deci, un raport tipic monohibridării (adică 3 : 1).
Prin urmare, se impune o primă concluzie. Cele două caractere nu segregă independent. De aici supoziţia că ele sunt plasate (genele ce le determină) pe acelaşi cromosom. Adică, pentru individul normal vom avea pr+ vg+, şi pr vg pentru cel mutant.
pr+ vg+ pr vg
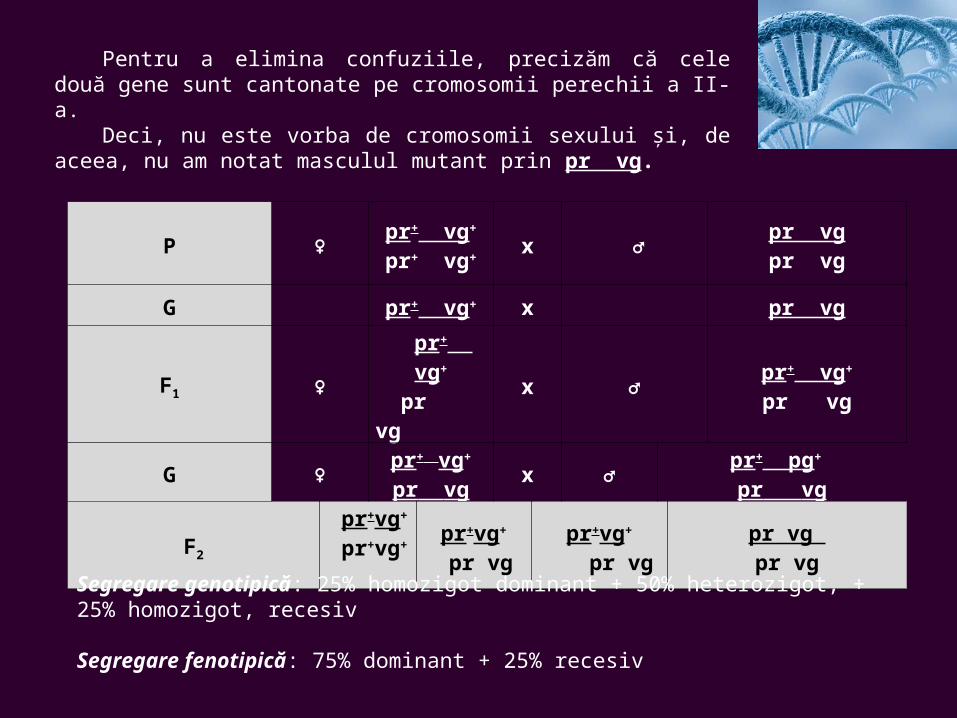
Pentru a elimina confuziile, precizăm că cele două gene sunt cantonate pe cromosomii perechii a II-a.
Deci, nu este vorba de cromosomii sexului şi, de aceea, nu am notat masculul mutant prin pr vg.
P ♀pr+ vg+
pr+ vg+x ♂
pr vgpr vg
G pr+ vg+ x pr vg
F1 ♀ pr+ vg+
pr vg x ♂
pr+ vg+
pr vg
G ♀pr+ vg+
pr vgx ♂
pr+ pg+ pr vg
F2
pr+vg+
pr+vg+
pr+vg+
pr vgpr+vg+
pr vgpr vg pr vg
Segregare genotipică: 25% homozigot dominant + 50% heterozigot, + 25% homozigot, recesiv
Segregare fenotipică: 75% dominant + 25% recesiv
Prin urmare cele două gene sunt localizate pe acelaşi cromosom şi se transmit împreună.
Fenomenul este cunoscut, în genetică, sub numele de linkage (înlănţuire, legare).
Este, în fond, nu numai o abatere, ci o completare a legii segregării independente a caracterelor, demonstrată de Mendel.
Transmiterea înlănţuită a tuturor genelor plasate pe un cromosom (linkage)
Autenticitatea linkage-ului a fost confirmată şi prin retroîncrucişarea de tipul Test-cross (FT).
Adică, un individ din F1 (un mascul) se încrucişează cu unul dublu mutant (o femelă). Se obţin două clase fenotipice şi anume: 50% indivizi normali şi 50% indivizi dublu mutanţi.
Precizăm că este obligatorie respectarea sexelor indicate, deoarece masculul nu prezintă fenomenul de crossing-over şi, în consecinţă, rezultatele experimentului sunt constante.
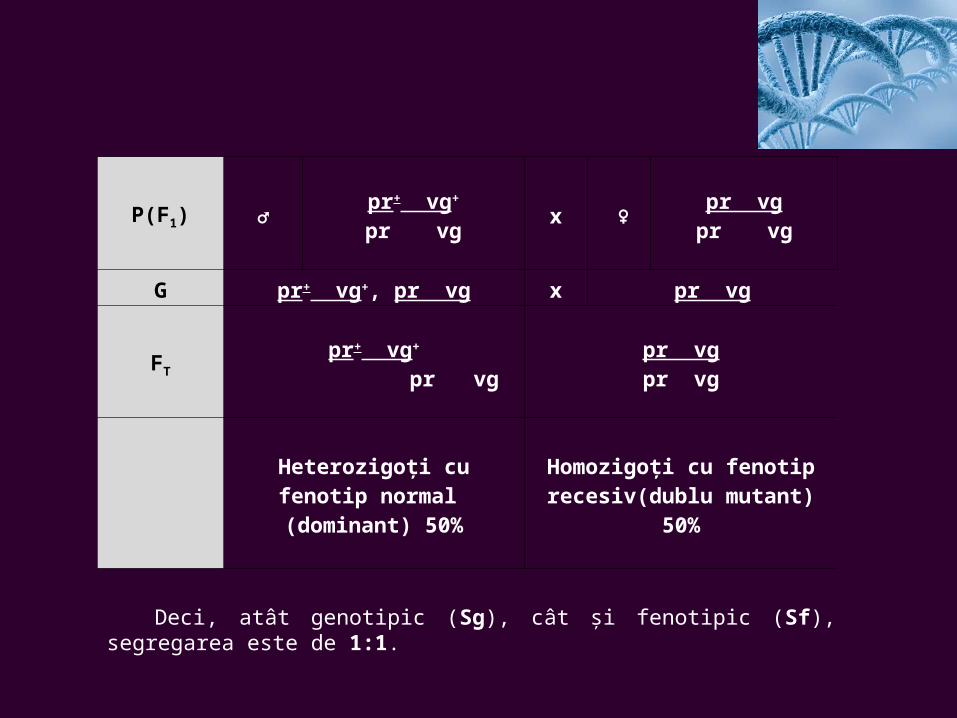
P(F1) ♂pr+ vg+
pr vgx ♀
pr vgpr vg
G pr+ vg+, pr vg x pr vg
FT
pr+ vg+
pr vgpr vgpr vg
Heterozigoţi cu fenotip normal (dominant)
50%
Homozigoţi cu fenotip recesiv(dublu mutant)
50%
Deci, atât genotipic (Sg), cât şi fenotipic (Sf), segregarea este de 1:1.
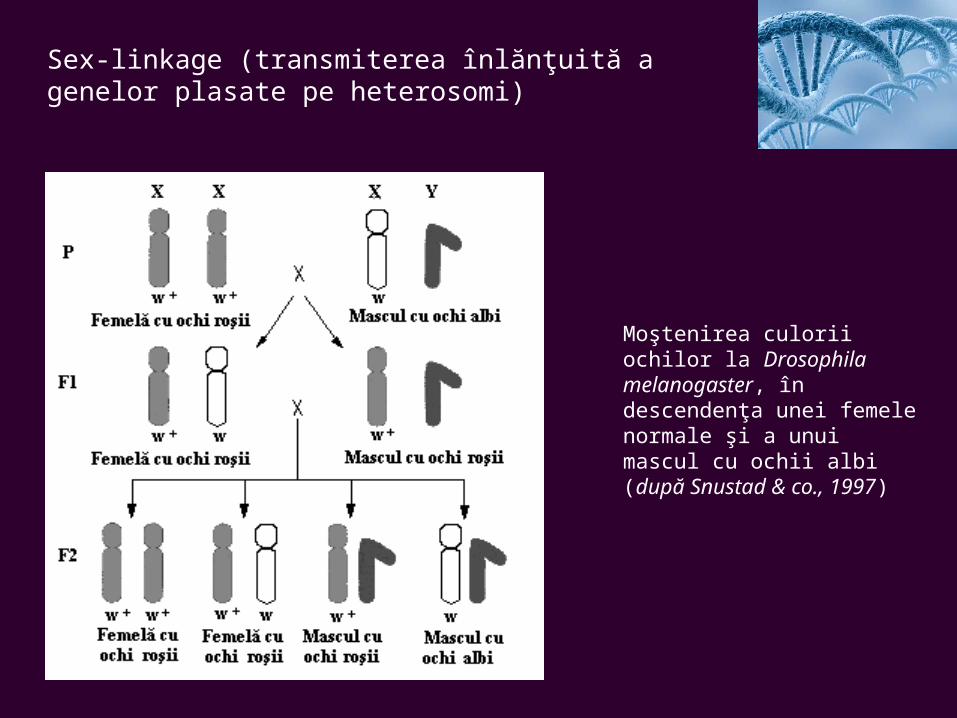
Sex-linkage (transmiterea înlănţuită a genelor plasate pe heterosomi)
Moştenirea culorii ochilor la Drosophila melanogaster, în descendenţa unei femele normale şi a unui mascul cu ochii albi (după Snustad & co., 1997)
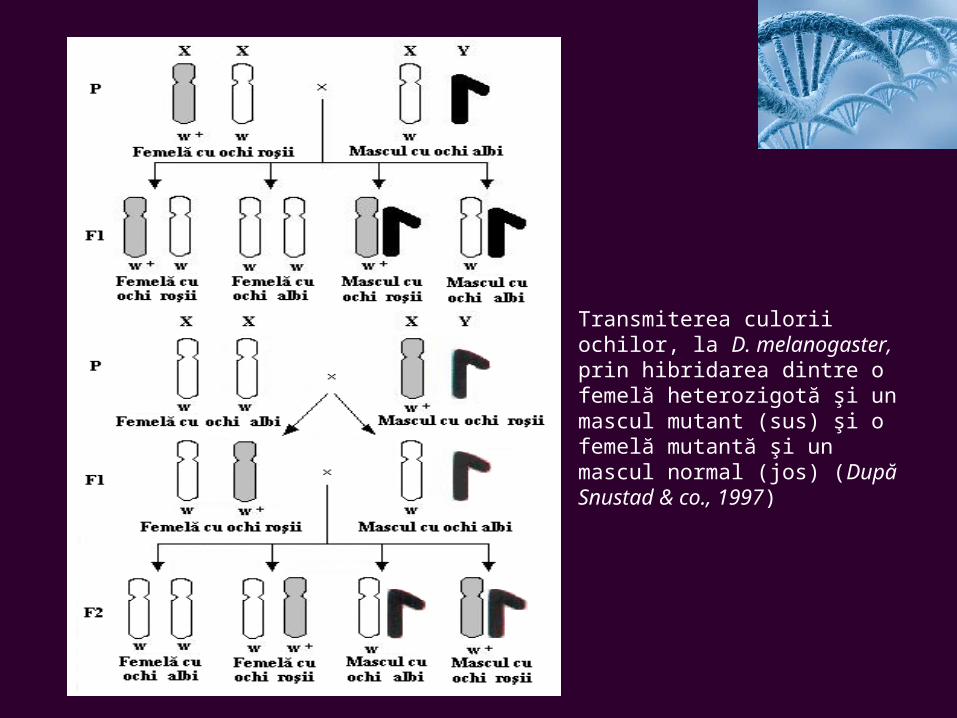
Transmiterea culorii ochilor, la D. melanogaster, prin hibridarea dintre o femelă heterozigotă şi un mascul mutant (sus) şi o femelă mutantă şi un mascul normal (jos) (După Snustad & co., 1997)

Genele care fac parte din aceeaşi grupă de linkage, sunt gene linkate.
Cromosomii reprezentând substratul (suportul) material al genelor, rezultă că în meioză asortarea întâmplătoare a cromosomilor în gameţi este, în fapt, asortarea întâmplătoare a grupelor de linkage. Altfel spus, o genă şi chiar o grupă de linkage, au comportamente identice cromosomilor.
În cazul sex-linkage-ului, are importanţă sexul indivizilor pe care-i folosim în încrucişare. Evident, când genele sunt localizate pe autosomi, sexul indivizilor utilizaţi pentru încrucişare nu ar trebui să aibă importanţă. Dar, aşa cum precizam mai sus, la Drosophila melanogaster, între autosomii masculului nu are loc fenomenul de crossing-over. Prin urmare, când se aşteaptă o constanţă a rezultatelor, este bine să se ţină cont de acest lucru.
Crossing-over (schimbul reciproc de gene)
Nu întotdeauna genele aparţinând aceluiaşi grup de linkage, se transmit împreună. S-au constatat nenumărate excepţii. În tentativa de a da explicaţii adecvate abaterilor de la regulă, Morgan şi echipa sa au constatat că între cromosomii omologi poate avea loc un schimb reciproc de gene. Fenomenul a primit numele de crossing-over.
Crossing-over între cromosomii sexului
Prin încrucişarea unei femele de Drosophila melanogaster, dublu mutantă (cu două mutaţii recesive, ale genelor localizate pe cromosomul X, determinând culoarea galbenă a corpului şi culoarea albă a ochilor), cu un mascul normal, în F1 s-au obţinut doar indivizi de două tipuri - deci, o segregare de 1:1. Femelele erau cu fenotip normal, dar heterozigote şi masculii cu fenotipizare mutantă, dar hemizigoţi genotipic. Fenomenul a primit denumirea de ereditate în cruciş (criss-cross), deoarece fenotipul mamei se manifestă la fii, iar cel al tatălui la fiice.
Prin încrucişarea indivizilor femeli şi masculi din F1, s-a constatat apariţia, în F2, a patru clase de indivizi - două clase conforme rezultatelor determinate de linkage şi alte două clase cu recombinare de caractere. Proporţia era de 49,25% pentru fiecare dintre primele două clase şi de 0,75% pentru fiecare din clasele cu recombinare.
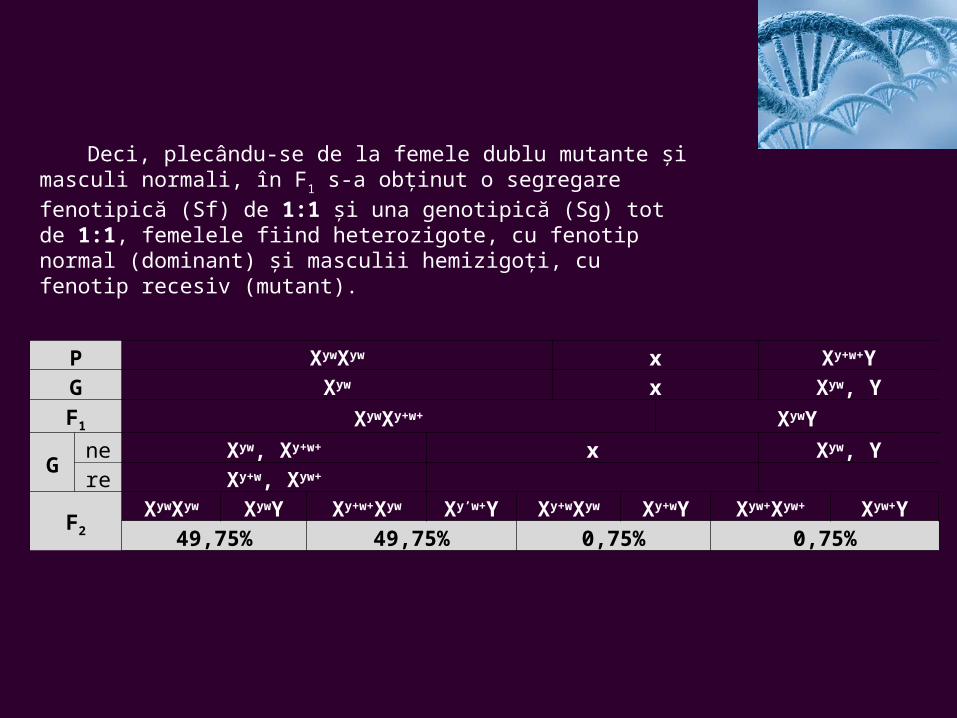
Deci, plecându-se de la femele dublu mutante şi masculi normali, în F1 s-a obţinut o segregare fenotipică (Sf) de 1:1 şi una genotipică (Sg) tot de 1:1, femelele fiind heterozigote, cu fenotip normal (dominant) şi masculii hemizigoţi, cu fenotip recesiv (mutant).
P XywXyw x Xy+w+YG Xyw x Xyw, YF1 XywXy+w+ XywY
Gne Xyw, Xy+w+ x Xyw, Yre Xy+w, Xyw+
F2
XywXyw XywY Xy+w+Xyw Xy’w+Y Xy+wXyw Xy+wY Xyw+Xyw+ Xyw+Y49,75% 49,75% 0,75% 0,75%

Teoretic, gameţii indivizilor de F1 trebuiau să fie doar de două tipuri: la femele Xyw şi Xy+w+, iar la masculi Xyw şi Y.
Dar rezultatele încrucişării indivizilor din F1, dând un cu totul alt raport de segregare în F2 şi, mai ales, dând o categorie de indivizi cu recombinări de caractere, s-a ajuns la concluzia că a avut loc crossing-overul între cromosomii omologi şi apariţia a două noi tipuri de gameţi în cazul femelelor: Xy+w şi Xyw+. Prezenţa acestor gameţi explică apariţia indivizilor cu fenotipuri recombinate, în F2. Aceştia sunt atât masculi, cât şi femele, cu corp gri şi ochi albi sau cu ochi roşii şi corp galben.
Deci, între genele y+ şi w+ şi alelele lor, situate pe cromosomii X omologi, au avut loc schimburi, rezultând noi grupe de linkage. Adică, din y+w+ şi yw, au rezultat y+w şi yw+, alături de tipurile parentale care s-au menţinut.
Crossing-over între autosomi
Încrucişând o femelă de Drosophila melanogaster cu corpul negru (b - de la black) şi aripi normale (vg+) cu un mascul cu corpul gri (b+) şi aripi vestigiale (vg), în F1 s-au obţinut indivizi heterozigoţi b+b vg+vg, cu fenotipul normal.
Încrucişându-se indivizii din F1 cu indivizi dublu mutanţi (încrucişare de analiză genetică), s-a constatat că în FT, fenotipul indivizilor diferă în funcţie de sexul lor.
Astfel, dacă se efectuează o retroîncrucişare între un mascul din F1, heterozigot (b+bvg+vg) cu o femelă mutantă (bbvgvg), în FT se obţine segregarea de 1:1, ca în experimentele pentru demonstrarea fenomenului linkage (adică reapar indivizii cu fenotip parental - corp negru şi aripi normale şi corp gri cu aripi vestigiale). Deci, linkage-ul este total.

Pb vg+
b vg+x
b+ vgb+ vg
G b vg+ x b+ vg
F1
b vg+
b+ vgx
b vgb vg
G b vg+, b+ vg x b vg
FT
b vg+
b vgb+ vgb vg
Sf 1 1
Sg 1(bbvg+vg) 1(b+bvgvg)

În încrucişarea reciprocă (retroîncrucişare) dintre o femelă hibridă de F1 (b+bvg+vg) şi un mascul dublu mutant (bbvgvg), în FT, pe lângă tipurile parentale (corp negru şi aripi normale; corp gri şi aripi vestigiale), mai apar încă două clase fenotipice (care, în mod normal, nu trebuiau să apară: corp gri şi aripi normale; corp negru şi aripi vestigiale). Într-o populaţie suficient de mare, proporţia indivizilor este de 41,5% pentru fiecare dintre tipurile parentale şi câte 8,5% pentru fiecare dintre tipurile recombinante. Deci, linkage-ul genelor b+ şi vg+ se dereglează şi respectivele gene "segregă", datorită schimbului reciproc de material cromatic între cromosomii omologi.
Rezultă că femelele din F1 pot forma patru tipuri de gameţi - două normale şi două recombinate, primii fiind numiţi gameţi crossing-overi, ceilalţi fiind gameţi necrossing-overi.

Pb vg+
b vg+x
b+ vgb+ vg
G b vg+ x b+ vg
F1
b vg+
b+ vgx
b vgb vg
GF1
b vg+, b+ vgşi b+ vg+, b vg
x b vg
FT
b vg+
b vgb+ vgb vg
b+ vg+
b vgb vgb vg
segregare
41,5% 41,5% 8,5% 8,5%
83,0 % indivizi necrossing-overi
17,0%indivizi crossing-
overi

Crossing-over simplu la plante
C. Hutchinson a utilizat două linii homozigote de Zea mays - una cu boabe conţinând aleuronă colorată şi endosperm neted (c+c+sh+sh+), iar cealaltă cu aleuronă incoloră şi endosperm zbârcit (ccshsh).
Hibridarea între cele două linii a permis obţinerea, în F1, heterozigoţilor c+csh+sh, cu expresie fenotipică normală.
Prin retroîncrucişarea unui astfel de hibrid, din F1, cu un părinte dublu mutant, în FT nu s-a obţinut o segregare de 1:1, cum ar fi fost normal în linkage, ci s-au obţinut patru clase fenotipice - două asemănătoare cu părinţii (adică fără crossing-over) şi două recombinante (cu crossing-over), care nu erau aşteptate în mod normal.
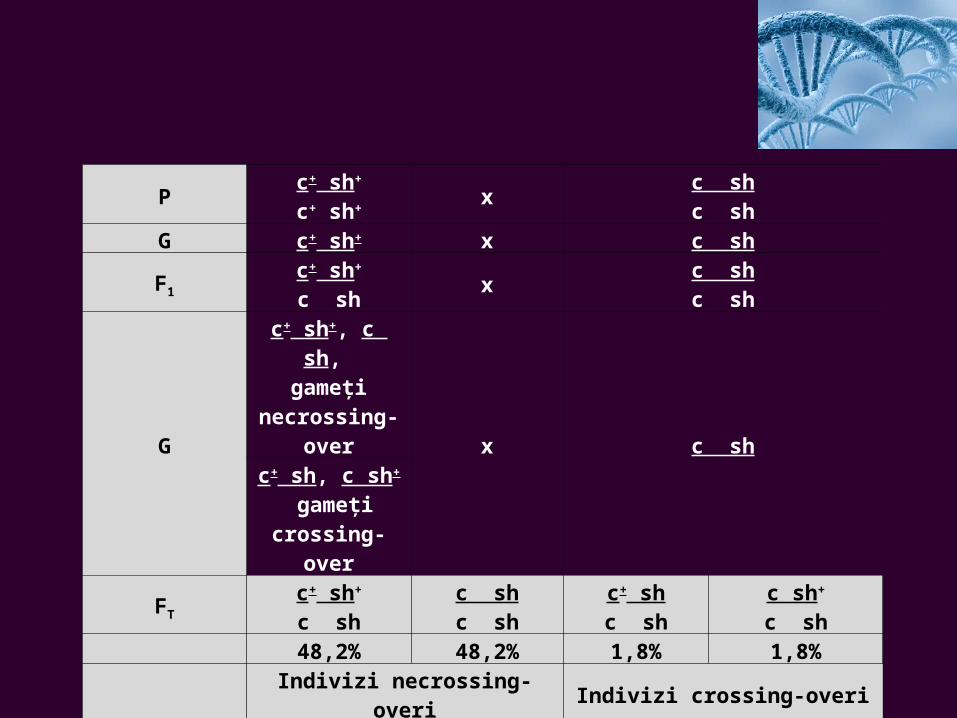
Pc+ sh+
c+ sh+x
c shc sh
G c+ sh+ x c sh
F1
c+ sh+
c shx
c shc sh
G
c+ sh+, c sh, gameţi
necrossing-over
x c shc+ sh, c sh+
gameţi crossing-
over
FT
c+ sh+
c shc shc sh
c+ shc sh
c sh+
c sh48,2% 48,2% 1,8% 1,8%
Indivizi necrossing-overi Indivizi crossing-overi

Crossing-over dublu la plante
M.M. Rhoades a descoperit un crossing-over dublu, tot la Zea mays.
Încrucişând plante cu fenotip normal verde (v+), având nervura centrală a frunzei de culoare verde (bw+), boabele cu grăuncioare de aleuronă incoloră (pr+) şi plante mutante pentru cele trei caractere, adică având fenotipul verde deschis (v, de la virescent), nervura centrală de culoare brună (bw, de la brown) şi aleurona purpurie (pr), în F1 a obţinut plante heterozigote cu fenotipul normal. Precizăm că cele trei gene sunt localizate pe cromosomii din perechea a V-a.
Hibrizii din F1, retroîncrucişaţi cu indivizi triplu mutanţi, nu s-au comportat în FT conform legilor mendeliene ale segregării, adică nu au dat 50% indivizi cu fenotip normal şi 50% cu fenotip recesiv (mutant). Rezultatul a fost neaşteptat, obţinându-se opt tipuri de indivizi: două tipuri parentale şi şase tipuri recombinate.


Mecanismele crossing-overului
Pentru explicarea modalităţilor concrete prin care se realizează crossing-overul s-au emis mai multe ipoteze, care pot fi grupate în două mari categorii, şi anume:
•Cele ce consideră că fenomenul se datorează ruperii cromatidelor şi schimbului reciproc de segmente (breakage theory) (Fig.1).
•Cele ce consideră drept cauză a fenomenului replicaţia selectivă a cromatidelor în timpul dublării materialului cromosomial (copy-choice theory) (Fig.2).
Schematic, situaţia se redă astfel:
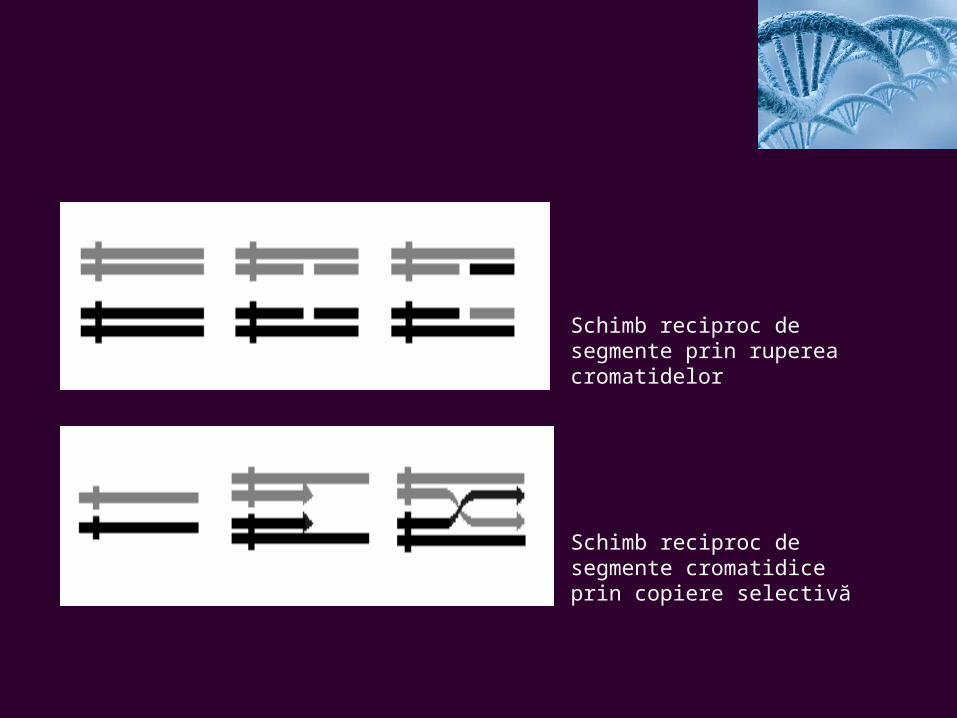
Schimb reciproc de segmente prin ruperea cromatidelor
Schimb reciproc de segmente cromatidice prin copiere selectivă

INTREBARI DE VERIFICARE
1.Care au fost avantajele folosirii Drosophilei melanogaster in cercetarile de genetica
2. Ce reprezinta linkage-ul
3. Ce reprezinta si cand are loc crossing-overul
4. Intocmiti schemele hibridarilor care sa ilustreze fenomenele de linkage si crossing-over
5. Care sunt mecanismele moleculare care conduc la aparitia crossing-overului


















