
Referat - Hormoni Si Proteine
31
REFERAT Disciplină: Chimie Profesor: Buruiană Marlena Elev: Nicolaiev Claudiu Clasa: a XI-a B
-
Upload
claudiu-alexandru-nicolaiev -
Category
Documents
-
view
1.775 -
download
22
Transcript of Referat - Hormoni Si Proteine

REFERAT
Disciplină: ChimieProfesor: Buruiană MarlenaElev: Nicolaiev ClaudiuClasa: a XI-a B

CUPRINS
1. Hormoni1.1. Introducere1.2. Clasificare1.3. Efecte ale hormonilor1.4. Hormoni sintetizaţi chimic1.5. Cei mai importanţi hormoni şi rolurile acestora
2. Proteine2.1. Introducere2.2. Sinteza proteinelor2.3. Clasificare, proprietăţi chimice şi fizico-chimice2.4. Structura proteinelor2.5. Rolul proteinelor
3. Bibliografie

1. Hormoni
1.1. IntroducereHormonii sunt substanţe biochimice care transmit informaţii de la un organ sau ţesut la
altul, fiind secretaţi de glandele endocrine sau de alte ţesuturi, care stimulează şi coordonează activitatea anumitor organe sau a întregului organism.
Spre deosebire de sistemul nervos, unde informaţiile de la centru (creier sau măduva spinării) se propagă prin prelungirile axonice sau dendritice cu o viteză mare, de ordinul fracţiunilor de secundă, informaţiile transmise pe cale hormonală sunt mai lente (în cazul adrenalinei, durează secunde, pe când în cazul celorlalţi hormoni durează zile până când informaţiile ajung la organul sau ţesutul ţintă).
Hormonii au fost descoperiţi la începutul secolului XX, iar termenul a fost pentru prima oară folosit în anul 1905 de către psihologul englez Ernest Starling. Fiind substanţe care acţionează pe anumite organe ţintă ce au receptori speciali în membrana celulară care leagă moleculele hormonului respectiv, unde se declanşează o serie de reacţii biochimice, hormonii pătrund prin membrana celulară şi ajung în citoplasma şi nucleul celulei.
Hormonii din organismul animal sunt produşi şi de glandele endocrine: hipofiză, epifiză, tiroidă, paratiroidă, pancreas, glandele suprarenale cu corticosuprarenala şi insulele Langerhans, gonadele sau glandele genitale.
Denumirea de glande endocrine sau glande cu secreţie internă se datorează faptului că hormonii produşi de glandele respective se varsă direct în sânge, fiind transportaţi pe calea sanguină la organul ţintă.
Hormonii produşi de plante se numesc fitohormoni (au un nucleu format din o grupare fenolică şi steroidă) şi, asemănător hormonilor produşi de organismul animal, acţionează la distanţă, fiind suficientă o cantitate foarte mică, de ordinul nanogramelor.
Știinţa care se ocupă cu studiul glandelor endocrine şi hormonilor se numeşte endocrinologie.

1.2. Clasificare
Clasificarea hormonilor se face în funcţie de raza de acţiune, solubilitate, structura chimică şi locul de sinteză.
Clasificarea hormonilor în funcţie de raza de acţiune
În funcţie de raza de acţiune, hormonii sunt grupaţi în: Hormoni locali: autocrini şi paracrini Hormoni endocrini Neurohormoni
Hormoni endocrini Hormonii endocrini respectă definiţia clasică, fiind sintetizaţi şi secretaţi de celule
specializate, urmând să ajungă la celula ţintă prin intermediul sistemului circulator. Ei sunt legati de proteine transportoare, în cazul hormonilor hidrofobi, sau liberi în sânge, în cazul hormonilor hidrofili.
Hormonii paracrini Hormonii paracrini nu respectă definiţia clasică dată hormonilor, fiind sintetizaţi de o celulă
specializată, urmând să acţioneze asupra celulelor învecinate, fără a mai ajunge în sistemul circulator.
Hormonii autocrini Hormonii autocrini sunt sintetizaţi şi secretaţi de către o celulă în spaţiul extracelular,
acţionând ca mesager asupra aceleiaşi celule. În acest caz, celula ţintă şi celula secretoare reprezintă de fapt aceeaşi celulă.
Neurohormonii Neurohormonii sunt o categorie specială de substanţe chimice produse de către celulele
nervoase, eliberate în sânge prin terminaţiile axonale şi apoi vehiculate prin sistemul circulator până la nivelul celulei ţintă. Prin intermediul neurohormonilor se realizează interacţiunea dintre sistemul nervos şi sistemul endocrin.
Clasificare hormonilor în funcţie de solubilitate
În funcţie de solubilitate, hormonii se pot clasifica în hormonii hidrosolubili (hidrofili) şi hormoni liposolubili (hibrofobi). Hormonii liposolubili necesită proteine transportoare pentru a fi vehiculaţi prin sânge până la nivelul celulei ţintă. De asemenea, aceşti hormoni prezintă capacitatea de a traversa stratul dublu lipidic al membranei celulare, pătrunzând în interiorul celulei, unde se află receptori specifici fie în citoplasmă, fie legaţi de cromatină la nivelul nucleului. Hormonii hidrosolubili circulă liberi prin sistemul circulator, însă nu pot traversa membrana celulară. Receptorii pentru acest tip de hormoni se află încorporaţi în membrana celulară, determinând producţia de mesageri secunzi la nivelul spaţiului citoplasmatic, hormonul fiind considerat mesager primar.

Hormonii hidrosolubili Hormonii peptidici Hormonii proteici Hormonii derivati din aminoacizi
Hormoni liposolubili Hormonii steroizi Hormonii tiroidieni
Clasificarea hormonilor după structura chimică
Hormoni derivaţi din aminoacizi
Hormonul Aminoacid precursor Loc de sinteză
Dopamina Tyrozina Hipotalamus, medulosuprarenală
Adrenalina Tirozina Medulosuprarenală
Noradrenalina Tirozina Medulosuprarenală, nervi periferici
Histamina Histidina Celule bazofile, celule care se regenerează
Serotonina Triptofan Ficat, nervi simpatici
3,5,3'-triiodotironina Tirozina Glanda tiroidă
Hormoni polipeptidici, proteici şi glicoproteici
HormonNumăr de
resturi aminoacidiceLoc de sinteză
Hormonul eliberator al tireotropinei (TRH) 3 Hipotalamus
Met-encefalina 5 Hipotalamus
Leu-encefalina 5 Hipotalamus
Angiotensina II 8 Sânge
Vasopresina (ADH) 9 Neurohipofiză
Ocitocina 9 Neurohipofiză
Hormonul eliberator al gonadotropinelor (Gn-RH)
10 Hipotalamus
Somatostatina (GH-RIH) 14 Hipotalamus
Gastrina 17Mucoasă gastrică duodenală
Hormonul melanocitostimulator (MSH) 27 Hipofiză anterioară
Secretina 27 Duoden şi jejun
Peptidul intestinal vasoactiv (VIP) 28 Duoden şi jejun
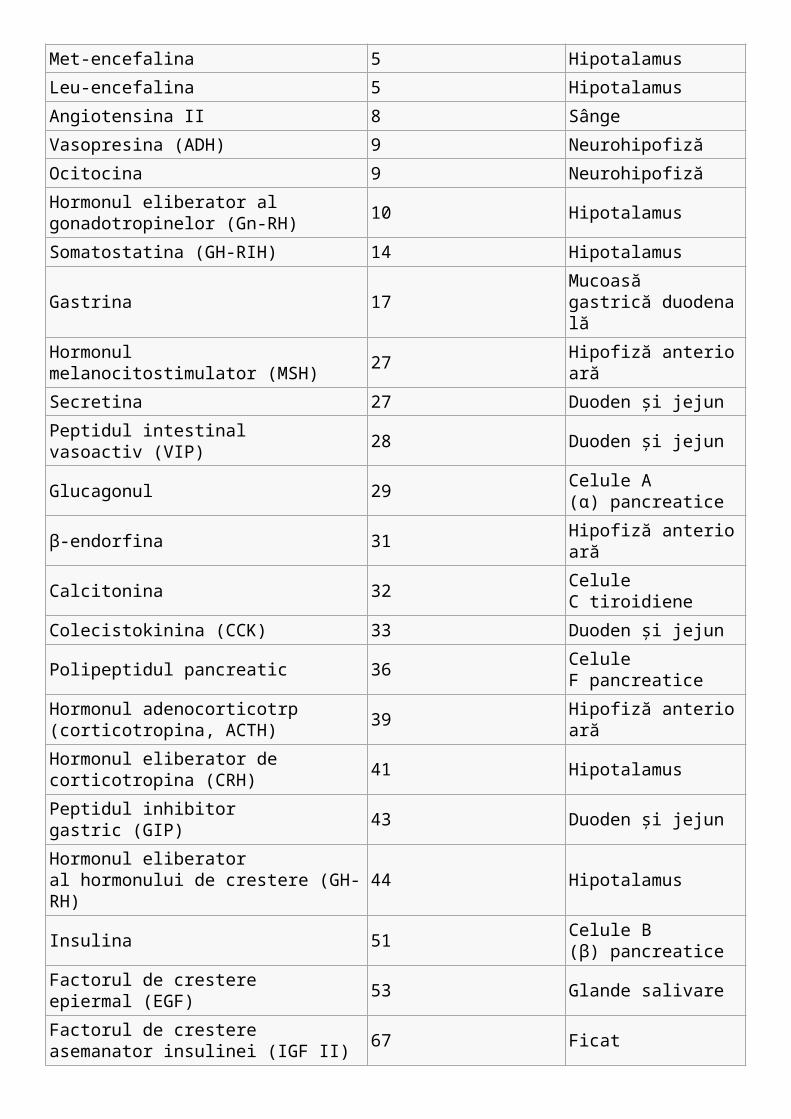
Glucagonul 29 Celule A (α) pancreatice
β-endorfina 31 Hipofiză anterioară
Calcitonina 32 Celule C tiroidiene
Colecistokinina (CCK) 33 Duoden şi jejun
Polipeptidul pancreatic 36 Celule F pancreatice
Hormonul adenocorticotrp (corticotropina, ACTH)
39 Hipofiză anterioară
Hormonul eliberator de corticotropina (CRH) 41 Hipotalamus
Peptidul inhibitor gastric (GIP) 43 Duoden şi jejun
Hormonul eliberator al hormonului de crestere (GH-RH)
44 Hipotalamus
Insulina 51 Celule B (β) pancreatice
Factorul de crestere epiermal (EGF) 53 Glande salivare
Factorul de crestere asemanator insulinei (IGF II)
67 Ficat
Somatomedina (IGF I) 70 Ficat
Hormonul parotidian 84 Celule parotidiene
Interleukine Variabil Celule albe din sânge
Prolactina (PRL) 198 Hipofiză anterioară
Hormonul tiroido-stimulator (TSH) 209 Hipofiză anterioară
Hormonul luteinizant (LH) 215 Hipofiză anterioară
Gonadotropina corionica umana (hCG) 231 Placentă
Hormonul foliculo-stimulator 236 Hipofiză anterioară
Hormoni steroizi Hormonii steroizi sunt hormoni ce au ca precursor comun colesterolul:
estradiol estronă testosteron 1,2 dihidroxicolecalciferol cortisol aldosteron
Hormonii eicosanoizi Hormoniii eicosanoizi reprezintă o serie de hormoni derivaţi din acidul arahidonic:
prostaglandinele primare (clasice, PG) endoperoxizii prostaglandinici (PGG2 şi PGH2) prostaciclina (PGI2) tromboxani (TX) leucotrienele (LT)
Clasificarea hormonilor în funcţie de locul de producere Hormonii sunt secretaţi la nivelul întregului organism, putând fi secretaţi de organe
specializate sau de celule individuale ce prezintă activitate secretorie.
Hormoni hipotalamici hormonul eliberator al tireotropinei (trh) hormonul eliberator al somatostatinei (gh-rih) hormonul eliberator al gonadotropinelor (gn-rh) hormonul eliberator al corticotropinei (crh) hormonul eliberator al somatotropinei (gh-rh)
Hormoni hipofizari somatotropina numita şi hormonul de crestere (gh) corticotropina numita şi hormonul adenocorticotrop (acth) gonadotropine (gn)
o hormonul luteinizant (lh)o hormonul foliculo-stimulator (fsh)
tireotropina (tsh) prolactina (prl)
Hormoni gastrointestinali gastrina colecistokinina (cck) secretina polipeptidul intestinal vasoactiv (vip) peptidul gastic inhibitor (gip) ghrelina peptidul yy parotina sialogastrona leptina motilina
Hormoni pancreatici insulina glucagonul somatostatina polipeptidul pancreatic
Hormoni suprarenalieni epinefrina (adrenalina) norepinefrina (noradrenalina) cortisolul aldosterona
Hormoni secretaţi de rinichi renina

1.3. Efecte ale hormonilorHormonii sunt compuşi cu acţiune directă sau indirectă asupra:
- glicemiei (concentraţia de zahăr din sânge), reglată prin insulină produs de pancreasul endocrin- lipemiei (concentraţia de grăsimi), reglată prin activitatea ficatului şi pancreasului- senzaţiei de foame- activităţii sexuale la bărbat sau femeie şi reglarea ciclului menstrual- dezvoltării sistemului osos- adaptării la stres.
Activitatea hormonală este dirijată prin:- axul hipotalamo-hipofizar printr-un sistem feedback- sistemul nervos vegetativ (parasimpatic şi ortosimpatic)- prin concentraţia în sânge a calciului, glucozei etc.

1.4. Hormoni sintetizaţi chimic
Esenţa fiziologiei şi patologiei glandelor endocrine este dependentă de hormonii produşi de glande. Aceste substanţe chimice au o structură chimică în general cunoscută. Majoritatea au fost sintetizaţi şi pot fi folosiţi ca medicamente. Pentru unii din ei, a căror sinteză chimică este dificilă, cum este hormonul de creştere (GH) al hipofizei sau insulina pancreasului, s-au pus la punct tehnici de inginerie genetică; sunt produşi de bacterii în care s-au introdus acizi dezoxiribonucleici recombinaţi, adică cu fragmente din acizii nucleici ai omului, care conţin programul genetic de sinteză a acestor hormoni.
Progresele chimiei moderne au permis să se sintetizeze substanţe înrudite cu hormonii naturali, dar mult mai puternici decât ei, numite analogi sintetici hormonali, şi care sunt folosiţi curent în terapeutică. Astfel, există dexametazonul şi prednisonul ca analogi ai cortizonului suprarenal, desmopresina ca analog al hormonului antidiuretic, dietilstilbestrolul ca analog al hormonilor estrogeni, buseralinul ca analog al neurohormonului gonadoliberină.
Administrarea hormonilor sau a analogilor lor sintetici ca medicamente trebuie făcută cu mare prudenţă, deoarece sunt substanţe foarte active, care influenţează puternic echilibrul organismului şi sunt supuşi unei autoreglări extrem de precise prin mecanisme cibernetice rapide de feedback. De exemplu, orice glandă endocrină produce hormonii ei specifici numai dacă concentraţia lor sangvină are tendinţa să scadă şi nu mai funcţionează dacă apare o creştere a nivelului sangvin a acestor hormoni. Dacă administrăm hormoni din afară, ca medicamente, fapt foarte răspândit mai ales când este vorba de hormonii cortizonici, glandele endocrine corespunzătoare sunt blocate şi se vor atrofia. În cazul suprarenalei, atrofia acesteia prin corticoterapia prelungită pune în pericol viaţa.

1.5. Cei mai importanţi hormoni şi rolurile acestora
Cei mai importanţi hormoni sunt:a) hormonii hipotalamici:
hormonul eliberator ai somatotropinei (SRH sau GH-RH) hormonul eliberator ai tireotropinei (TRH), numit şi tireoliberină, este cel mai bine
cunoscut. Din punct de vedere chimic, este un tripeptid format din acid glutamic , histidină şi prolină. Structura chimică a acestui compus prezintă însă următoarele particularităţi: acidul glutamic este sub forma ciclică, parolina este încatenată prin hidrogenul iminic , iar gruparea carboxilică a parolinei se găseşte sub formă de amidă
hormonul eliberator al corticotropinei (CRH) se mai numeşte şi corticoliberină şi a fost pus în evidenţă prin cercetările lui Saffran şi Schally. S-a arătat mai târziu că acest polipeptid este alcătuit din 41 resturi de aminoacizi. Concomitent cu stimularea eliberării de corticotropină (ACTH), CRH-ul stimulează şi eliberarea de către adenohipofiză a unui alt compus biologic activ numit ß-endorfină. Aceasta are de asemenea structură polipeptidică şi are acţiunea opusă CRH-ului; este deci un inhibator al ACTH-ului.
hormonul eliberator al aigonadotropinelor (GN-RH) hormonul eliberator al somatostatinei (GH-RH)
b) hormonii hipofizari: somatotropina (GH) este esenţială în procesul de creştere, iar la adult are un rol important
în activităţile metabolice. Este sintetizată în celulele acidofile ale hipofizei anterioare, iar secreţia sa din granulele de depozit intracelulare este reglată de hormonii hipotalamici GHRH şi SRIF; sinteza lor depinde de neurotransmiţători, ca serotonina, dopamina, norepinefrina şi peptidele de eliberare a hormonului de creştere.
corticotropina (ACTH), hormonul adenocorcotrop sau corticotropina stimulează creşterea, dezvoltarea şi activitatea secretorie a glandelor corticosuprarenale. Hipersecreţia de ACTH determină hipertrofierea corticosuprarenalei şi hipersecreţie de hormoni ai acesteia, având ca urmare tulburări metabolice.
gonadotropina (Gn–LH şi FSH) controlează funcţiile gonadelor feminine şi masculine. De asemenea, controlează secreţia glandelor mamare la femeie.
tireotropina (TSH) stimuleaza creşerea, dezvoltarea şi secretia de hormoni ai glandei tiroide prolactina (PRL) este secretată de hipofiza anterioară şi eliberată episodic. Prolactina apare
în ser sub trei forme diferite. Predomină forma monomerică (80 %) care este activă din punct de vedere biologic şi imunologic; urmează în procent de 5-20 % forma dimerică care este biologic inactivă, iar în proporţie de 0.5-5 % forma tetramerică, cu activitate biologică scăzută. Organul ţintă pentru prolactină este glanda mamară unde îşi exercită funcţia lactogenică şi galactopoietică.
c) hormonii gastrointestinali: gastrina este un hormon secretat de celulele G (localizate în porţiunea distală a stomacului
şi în duoden) ca răspuns la stimularea vagală, ingestia de alimente/alcool/calciu sau prezenţa de secreţie gastrică alcalină. După eliberarea în circulaţie ajunge la nivelul stomacului unde stimulează secreţia de acizi, pepsină, factor intrinsec şi insulină.
colecistokinina este un hormon secretat de celulele duodenale, ca răspuns la prezenţa de alimente parţial digerate în duoden.

secretina este un hormon polipetidic secretat de mucoasa duodenală sub influenţa conţinutului gastric acid. Are rol stimulator asupra secreţiei pancreatice.
d) hormonii pancreatici: insulina este singurul hormon al organismului cu acţiune hipoglicemiantă. El face ca glucoza
din sânge să intre în celule, care o folosesc pentru producerea de energie. glucagonul este un hormon secretat de către pancreas şi care creşte concentraţia sangvină
a glucozei. Glucagonul este indicat pacienţilor diabetici în caz de hipoglicemie cauzată de o supradozare în insulină care a antrenat o stare de rău sau o comă.
e) hormonii trombocitari: tromboxani
f) hormonii limfocitari: leucotrienele sunt substanţe chimice sintetizate de celulele mastoide, bazofile, macrofage şi
alte ţesuturi din ţesutul arahidonic. Leucotrienele sunt implicate în reacţiile inflamatorii şi în răspunsul imun; ele cresc permeabilitatea vaselor sangvine mici. Produc contracţia muşchilor netezi şi atrag neutrofilele la locul infecţiei.
g) hormonii suprarenali: epinefrina sau adrenalina este un hormon produs de glanda suprarenală ce are un rol
primordial în funcţionarea sistemului nervos sinaptic. norepinefrina cortisolul este un hormon corticosteroid produs de către cortexul glandei suprarenale şi
care este implicat în răspunsul la stres. El creşte tensiunea arterială, glicemia, şi poate cauza infertilitate la femei.
aldosterona
h) hormonii renali: renina este o enzimă secretată de o zonă a rinichiului situată în apropierea glomerulilor şi
denumită aparat juxtaglomerular. Nivelul reninei în sânge creşte în cursul anumitor hipertensiuni, în cursul insuficienţelor renale sau cardiace. Din contră, nivelul său scade în cursul unor dereglări hormonale de origine suprarenaliană (hipercorticism, sindromul lui Conn).
i) hormonii din ţesutul miocardic atrial: polipeptidul natrinuretic.

2. Proteine
2.1. Introducere
Proteinele sunt substanţe organice macromoleculare formate din lanţuri simple sau complexe de aminoacizi; ele sunt prezente în celulele tuturor organismelor vii în proporţie de peste 50% din greutatea uscată. Toate proteinele sunt polimeri ai aminoacizilor, în care secvenţa acestora este codificată de către o genă. Fiecare proteină are secvenţa ei unică de aminoacizi, determinată de secvenţa nucleotidică a genei.
Proteinele sunt componente esenţiale, alături de apă, săruri anorganice, lipide, hidraţi de carbon, acizi nucleici, vitamine, enzime ale materialelor lichide şi gelificate din celule, în care se petrec interacţiunile chimice şi fizice intense cuprinse sub denumirea de viaţă.
De proteine depind multe funcţiuni importante ale organismelor vii. Sunt proteine unele substanţe cu puternică activitate biologică ale celulelor ca: enzimele, pigmenţii respiratori, mulţi hormoni şi anticorpii. Substanţa contractilă din fibrele musculare, din cilii şi din flagelele organismelor inferioare, care posedă proprietatea de a transforma energia chimică în energie mecanică este de asemenea o proteină.
Proteinele care asigură funcţiuni se bucură de proprietatea neîntâlnită la alte combinaţii, specificitatea. Proteinele diferitelor specii de animale şi vegetale sunt tipice pentru aceste specii şi se deosebesc de proteinele altor specii, în timp ce, de exemplu amidonul, glicogenul şi grăsimile nu diferă decât puţin de la o specie la alta. Deci, numărul proteinelor care se observă în natură este extrem de mare.
Pe baza solubilităţii se disting două clase de proteine: proteine insolubile şi solubile. Cele dintâi, numite proteine fibroase, se găsesc în organismul animal în stare solidă şi conferă ţesuturilor rezistenţa mecanică ( proteine de schelet) sau protecţie împotriva agenţilor exteriori. De exemplu, keratina din păr, unghii, copite, epiderma, colagenul din piele, oase şi tendoane, miozina din muşchi şi fibroina din mătase. În vegetale nu se găsesc proteine fibroase; funcţiunea lor este îndeplinită în plante de celuloză. Proteinele fibroase se dizolvă numai în acizi şi baze concentrate, la cald, dar aceasta dizolvare este însoţită de o degradare a macromoleculelor; din soluţiile obţinute nu se mai regenerează proteina iniţială.
Proteinele solubile sau globulare apar în celule în stare dizolvată sau sub formă de geluri hidratate. Albuminele sunt solubile în apă şi în soluţii diluate de electroliţi (acizi, baze, săruri); globulinele sunt solubile numai în soluţii de electroliţi. Din categoria aceasta fac parte toate proteinele cu proprietăţi fiziologice specifice: poteinele din serul sanguin, enzimele, hormonii proteici, anticorpii şi toxinele.
O categorie importantă sunt proteidele sau proteinele conjugate, combinaţii ale unei proteine cu o componentă neproteică.

2.2. Sinteza proteinelor
Biosinteza Biosinteza proteinelor este un proces prin care fiecare celulă îşi sintetizează proteinele
proprii, prin intermediul unui proces care include multe etape, sinteza începând cu procesul de transcripţie ţi terminând cu procesul de translaţie. Procesul deşi similar, este diferit în funcţie de celulă: eucariotă sau procariotă.
Transcripţia Procesul de transcripţie necesită prezenţa unei singure molecule de ADN dublu catenar,
numit ADN "şablon", moleculă care intră în procesul de "iniţiere". Aici acţionează enzima ARN polimeraza, enzimă care se leagă de o anumită regiune din molecula de ADN, regiune (denumită promoter), de unde va începe transcripţia. Pe măsură ce ARN polimeraza se leagă de promoter, lanţurile de ADN vor începe sa se desfacă. Următorul proces în care intră ADN este procesul de elongaţie (alungire a catenei). Pe măsură ce ARN polimeraza se mişcă de-a lungul catenei de ADN, are loc sinteza ribonucleotidelor complementare (ARNm - ARN mesager). Acest ARN, după cum îi arată şi numele, se poate deplasa şi în alte părţi ale celulei cum ar fi reticulul endoplasmatic sau citoplasma.
Are loc adiţia unei grupări 5', grupare dinucleotidică care are rolul de a asigura stabilitatea ARN şi de a-l transforma în ARN matur. O secvenţă de aminoacizi este grefată în poziţia 3' terminală pentru protecţie, dar şi pentru a sluji drept şablon pentru procesele următoare. Mai departe are loc formarea ARN, care este apoi utilizat în ribozomi pentru sinteza proteinelor. La procariote legarea ARN de ribozomi are loc după ce acesta este îndepărtat de nucleoid; în contrast la procariote acest proces are loc chiar în membrana nucleară şi apoi translocat în citoplasmă. Rata sintezei proteică poate ajunge la circa 20 aminoacizi la procariote, mult mai puţin la eucariote.
Translaţia În timpul translaţiei ARNm transcris din ADN, acesta este decodat de ribozomi pentru
sinteza proteinelor. Procesul este divizat în trei etape: iniţierea elongarea faza terminală.
Ribozomul are situsuri de legare care permit altei molecule de ARNt (ARN de transfer), să se lege de o moleculă de ARNm, proces însoţit de prezenţa unui anticodon. Pe măsură ce ribozomul migrează de-a lungul moleculei de ARNm (un codon o dată) o altă moleculă de ARNt este ataşată la ARNm. Are loc eliberarea ARNt primar, iar aminoacidul care este ataşat de acesta este legat de ARNt secundar, care îl leagă de o altă moleculă de aminoacid. Translaţia continuă pe măsură ce lanţul de aminoacid este format. La un moment dat apare un codon de stop, o secvenţă formată din 3 nucleotide (UAG, UAA), care semnalează sfârşitul lanţului proteic. Chiar după terminarea translaţiei, lanţurile proteice pot suferi modificări post-translaţionale şi plierea lanţului proteic, responsabilă de structura secundară şi cea terţiară. Modificările post-translaţionale se referă la posibilitatea formării de legături disulfidice, sau de ataşarea la scheletul proteic a diferite grupări ca rol biochimic: acetat, fosfat etc.

Sinteza chimică Procesul de sinteză chimică poate avea loc în laborator, dar pentru lanţuri mici de proteine.
O serie de reacţii chimice cunoscute sub denumirea de sinteza peptidelor, permit producerea de cantităţi mari de proteine. Prin sinteza chimică se permite introducerea în lanţul proteic a aminoacizilor nenaturali, ataşarea de exemplu a unor grupări fluorescente. Metodele sunt utilizate în biochimie şi în biologia celulei. Sinteza are la bază cuplarea grupării carboxil -COOH (carbon terminus) cu gruparea amino (-NH2) (segmentul N terminus). Se cunosc 2 metode de sinteză pe cale chimică:
sinteza în fază lichidă, metoda clasică, care a fost înlocuită cu sinteza în fază solidă. sinteza în fază solidă (SPPS). Prin această metodă, se pot sintetiza proteine D, cu aminoacizi
D. În prima fază, s-a folosit metoda tBoc (terţ-butil-oxi-carbonil). Pentru înlăturarea acestuia din lanţul peptidic se foloseşte acidul fluorhidric, care este foarte nociv, periculos, iar din acest motiv, metoda nu se mai utilizează.

2.3. Clasificare, proprietăţi chimice şi fizico-chimice
Clasificare
În funcţie de compoziţia lor chimică ele pot fi clasificate în: Holoproteine cu următoarele clase de proteine:
o Proteine globulare (sferoproteine) sunt de regulă substanţe solubile în apă sau în soluţii saline: protaminele, histonele, prolaminele, gluteinele, globulinele, albuminele.
o Proteinele fibrilare (scleroproteinele) caracteristice regnului animal, cu rol de susţinere, protecţie şi rezistenţă mecanică: colagenul, cheratina şi elastina.
Heteroproteinele sunt proteine complexe care sunt constituite din o parte proteică şi o parte prostetică; în funcţie de această grupare se pot clasifica astfel:
o glicoproteineo lipoproteineo nucleoproteine
Proprietăţi chimice
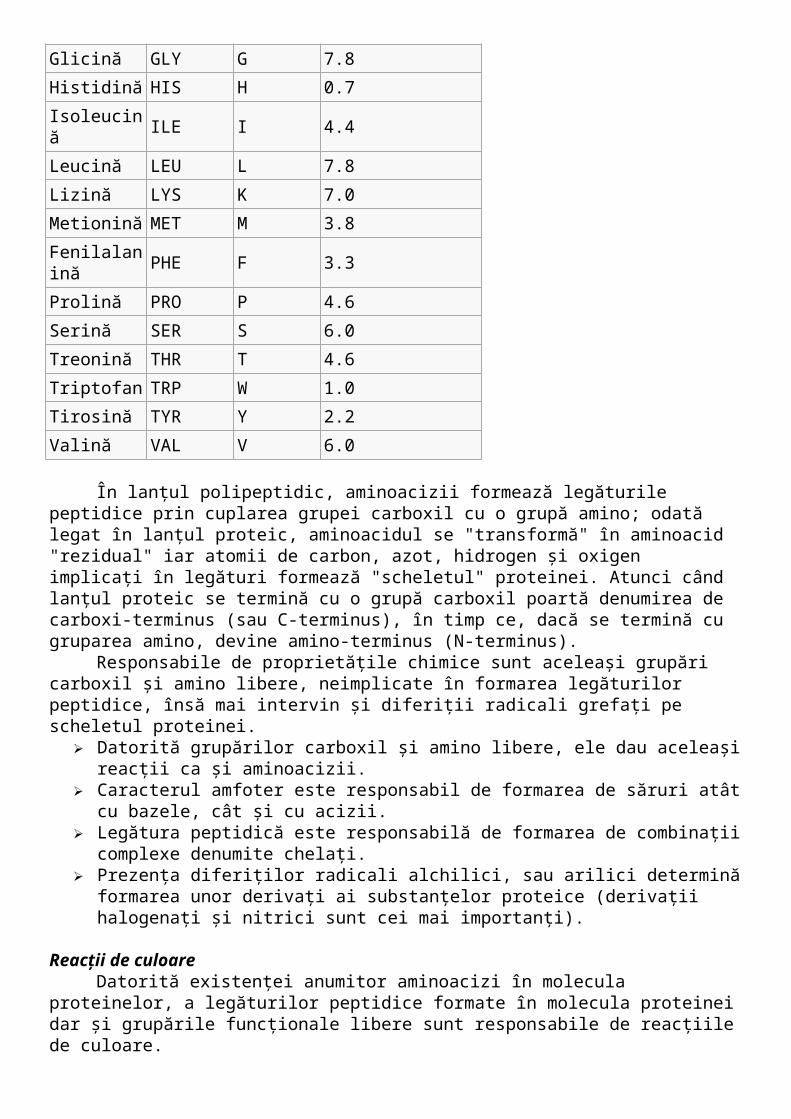
Aminoacizi standard Din punct de vedere chimic, proteinele sunt heteropolimeri constituiţi din 20 de L-α
aminoacizi (aşa numiţii aminoacizi standard), în care grupările carboxil se pot combina cu grupările amino formând legături peptidice şi rezultând lanţurile peptidice. Aminoacizii standard au proprietăţi variate, proprietăţi care sunt direct responsabile de structura tridimensională a proteinei, dar şi de proprietăţile acesteia.
Denumirea cod 3 literecod 1 literă
Abundenţă />(%) E.C.
Alanină ALA A 13.0
Arginină ARG R 5.3
Asparagină ASN N 9.9
Aspartat ASP D 9.9
Cisteină CYS C 1.8
Acid glutamic GLU E 10.8
Glutamină GLN Q 10.8
Glicină GLY G 7.8
Histidină HIS H 0.7
Isoleucină ILE I 4.4
Leucină LEU L 7.8
Lizină LYS K 7.0
Metionină MET M 3.8

Fenilalanină PHE F 3.3
Prolină PRO P 4.6
Serină SER S 6.0
Treonină THR T 4.6
Triptofan TRP W 1.0
Tirosină TYR Y 2.2
Valină VAL V 6.0
În lanţul polipeptidic, aminoacizii formează legăturile peptidice prin cuplarea grupei carboxil cu o grupă amino; odată legat în lanţul proteic, aminoacidul se "transformă" în aminoacid "rezidual" iar atomii de carbon, azot, hidrogen şi oxigen implicaţi în legături formează "scheletul" proteinei. Atunci când lanţul proteic se termină cu o grupă carboxil poartă denumirea de carboxi-terminus (sau C-terminus), în timp ce, dacă se termină cu gruparea amino, devine amino-terminus (N-terminus).
Responsabile de proprietăţile chimice sunt aceleaşi grupări carboxil şi amino libere, neimplicate în formarea legăturilor peptidice, însă mai intervin şi diferiţii radicali grefaţi pe scheletul proteinei.
Datorită grupărilor carboxil şi amino libere, ele dau aceleaşi reacţii ca şi aminoacizii. Caracterul amfoter este responsabil de formarea de săruri atât cu bazele, cât şi cu acizii. Legătura peptidică este responsabilă de formarea de combinaţii complexe denumite
chelaţi. Prezenţa diferiţilor radicali alchilici, sau arilici determină formarea unor derivaţi ai
substanţelor proteice (derivaţii halogenaţi şi nitrici sunt cei mai importanţi).
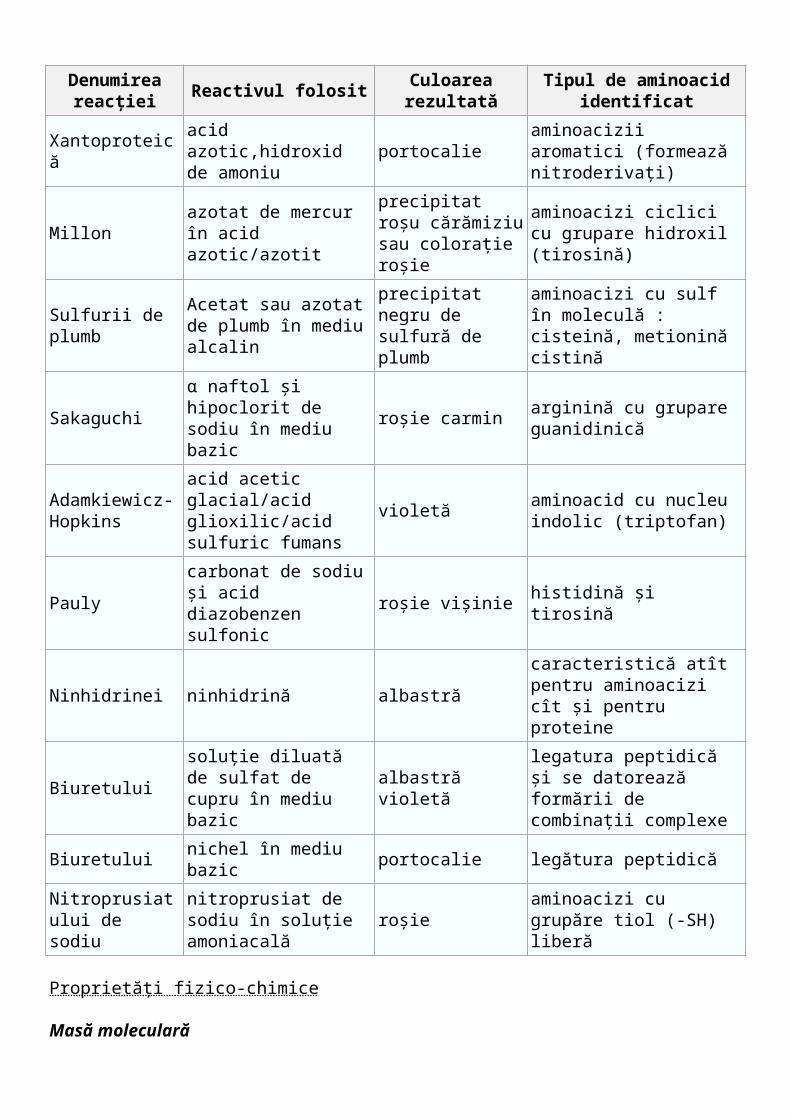
Reacţii de culoare Datorită existenţei anumitor aminoacizi în molecula proteinelor, a legăturilor peptidice
formate în molecula proteinei dar şi grupările funcţionale libere sunt responsabile de reacţiile de culoare.
Denumirea reacţiei
Reactivul folosit Culoarea rezultatăTipul de aminoacid
identificat
Xantoproteicăacid azotic,hidroxid de amoniu
portocalieaminoacizii aromatici (formează nitroderivaţi)
Millonazotat de mercur în acid azotic/azotit
precipitat roşu cărămiziu sau coloraţie roşie
aminoacizi ciclici cu grupare hidroxil (tirosină)
Sulfurii de plumbAcetat sau azotat de plumb în mediu alcalin
precipitat negru de sulfură de plumb
aminoacizi cu sulf în moleculă : cisteină, metionină cistină
Sakaguchiα naftol şi hipoclorit de sodiu în mediu bazic
roşie carminarginină cu grupare guanidinică
Adamkiewicz-Hopkins
acid acetic glacial/acid glioxilic/acid sulfuric
violetă aminoacid cu nucleu indolic (triptofan)

fumans
Paulycarbonat de sodiu şi acid diazobenzen sulfonic
roşie vişinie histidină şi tirosină
Ninhidrinei ninhidrină albastrăcaracteristică atît pentru aminoacizi cît şi pentru proteine
Biuretuluisoluţie diluată de sulfat de cupru în mediu bazic
albastră violetălegatura peptidică şi se datorează formării de combinaţii complexe
Biuretului nichel în mediu bazic portocalie legătura peptidică
Nitroprusiatului de sodiu
nitroprusiat de sodiu în soluţie amoniacală
roşieaminoacizi cu grupăre tiol (-SH) liberă
Proprietăţi fizico-chimice
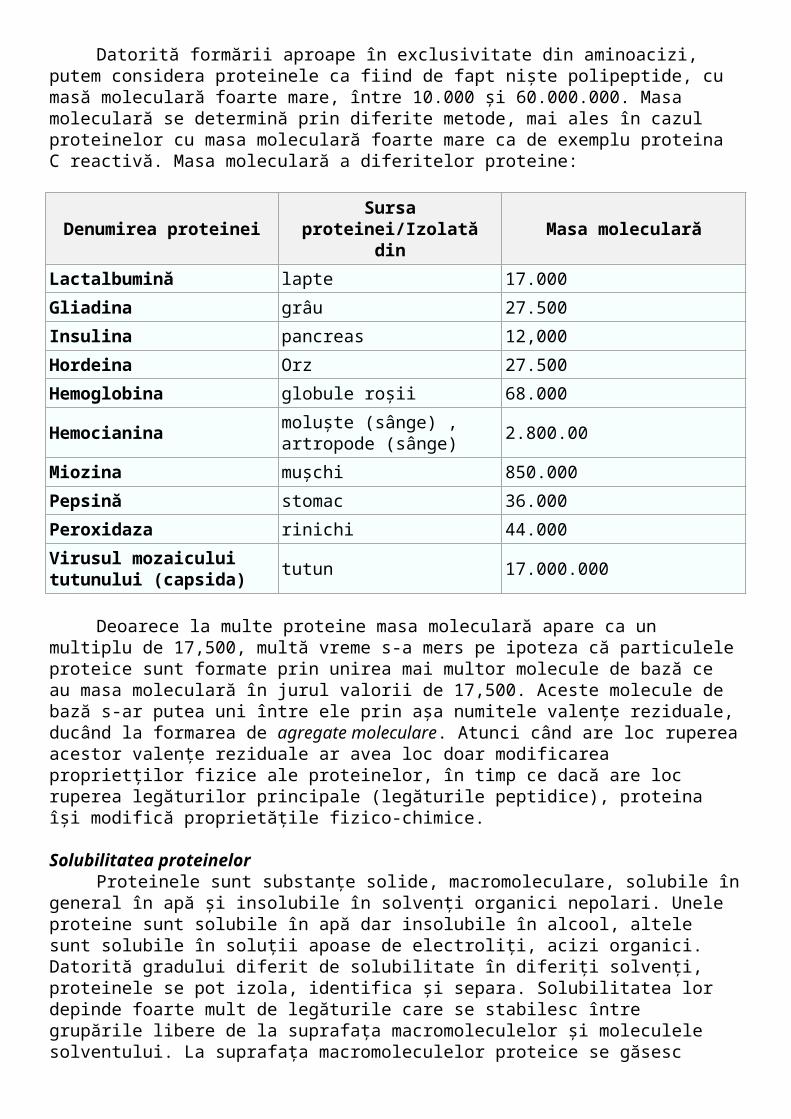
Masă moleculară Datorită formării aproape în exclusivitate din aminoacizi, putem considera proteinele ca
fiind de fapt nişte polipeptide, cu masă moleculară foarte mare, între 10.000 şi 60.000.000. Masa moleculară se determină prin diferite metode, mai ales în cazul proteinelor cu masa moleculară foarte mare ca de exemplu proteina C reactivă. Masa moleculară a diferitelor proteine:
Denumirea proteinei Sursa proteinei/Izolată din Masa moleculară
Lactalbumină lapte 17.000
Gliadina grâu 27.500
Insulina pancreas 12,000
Hordeina Orz 27.500
Hemoglobina globule roşii 68.000
Hemocianinamoluşte (sânge) , artropode (sânge)
2.800.00
Miozina muşchi 850.000
Pepsină stomac 36.000
Peroxidaza rinichi 44.000
Virusul mozaicului tutunului (capsida)
tutun 17.000.000
Deoarece la multe proteine masa moleculară apare ca un multiplu de 17,500, multă vreme s-a mers pe ipoteza că particulele proteice sunt formate prin unirea mai multor molecule de bază ce au masa moleculară în jurul valorii de 17,500. Aceste molecule de bază s-ar putea uni între ele prin aşa numitele valenţe reziduale, ducând la formarea de agregate moleculare. Atunci când are loc ruperea acestor valenţe reziduale ar avea loc doar modificarea proprietţilor fizice ale proteinelor, în timp ce dacă are loc ruperea legăturilor principale (legăturile peptidice), proteina îşi modifică proprietăţile fizico-chimice.

Solubilitatea proteinelor Proteinele sunt substanţe solide, macromoleculare, solubile în general în apă şi insolubile în
solvenţi organici nepolari. Unele proteine sunt solubile în apă dar insolubile în alcool, altele sunt solubile în soluţii apoase de electroliţi, acizi organici. Datorită gradului diferit de solubilitate în diferiţi solvenţi, proteinele se pot izola, identifica şi separa. Solubilitatea lor depinde foarte mult de legăturile care se stabilesc între grupările libere de la suprafaţa macromoleculelor şi moleculele solventului. La suprafaţa macromoleculelor proteice se găsesc grupări libere de tip polar,-COOH, -NH2, -OH, -SH, -NH, grupări cu caracter hidrofil care favorizează dizolvarea proteinelor în apă. De asemenea există grupări de tip apolar, hidrofobe, de regulă radicali de hidrocarburi -CH3, -C6H5, -C2H5, care favorizează dizolvarea proteinelor în alcool. Însă în marea lor majoritate predomină grupările polare, determinante pentru caracterul hidrofil. În contact cu apa proteinele greu solubile manifestă fenomenul de gonflare, datorită tendinţei de hidratare datorată grupărilor polare. Gelatina de exemplu se îmbibă foarte puternic cu apa dând naştere prin răcire la geluri. La dizolvarea proteinelor în apă, are loc fenomenul de formare a coloizilor hidrofili. S-a constatat că în soluţii diluate se găsesc macromolecule proteice izolate, iar în cazul soluţiilor concentrate se formează agregate de macromolecule proteice. Soluţiile coloidale ale proteinelor, coagulează prin încălzire, prezintă efectul Tyndall (dispersia fasciculului de lumină).
Caracterul amfoter Proteinele, la fel ca şi aminoacizii, sunt substanţe amfotere şi formează în soluţii apoase
amfioni. În mediu acid proteinele se comportă ca baze slabe, ele primind protoni şi formând cationi proteici. Reacţia stă la baza electroforezei proteinelor, datorită încărcării pozitive cationii migrează spre catod, fenomen numit cataforeză, proteina fiind în acest caz electropozitivă.În mediu bazic proteinele se comportă ca acizii slabi, ele cedând protoni, se formează astfel anioni proteici, care migrează spre anod fenomenul fiind denumit anaforeză, proteina având încărcare electronegativă.
Datorită caracterului amfoter proteinele pot neutraliza cantităţi mici de substanţă acidă sau bazică, având în acest fel rol de soluţie tampon, prin acest lucru contribuind la menţinerea echilibrului acido-bazic al organismului. În general caracterul amfoter este imprimat de cele grupările -NH2 şi -COOH libere care nu sunt implicate în legăturile peptidice. Dacă în molecula proteinei există mai mulţi aminoacizi dicarboxilici atunci molecula se va comporta ca un acid slab, iar în cele în care predomină aminoacizii diaminaţi se comportă ca baze slabe. Chiar dacă într-o moleculă există un număr egal de grupări amino şi carboxil, deci teoretic molecula ar trebui sa fie neutră, în realitate datorită gradului de ionizare mult mai mare a grupării carboxil faţă de gruparea amino, molecula proteinei va avea un caracter slab acid, în soluţia ei întâlnindu-se amfionii proteici, anioni proteici şi protoni (H+).
Punctul izoelectric Prin acidulare, echilibrul reacţiei se deplasează spre formarea de cationi proteici. La o
anumită concentraţie a H+, proteina devine neutră deoarece gruparea aminică şi cea carboxilică sunt la fel de disociate şi deci molecula este neutră din punct de vedere electric. În acel moment se vor găsi în soluţie amfioni, H+, ioni hidroxil HO-; pH-ul la care soluţia unei proteine conţine anioni şi cationi în proporţie egală poarta denumirea de punct izoelectric, se notează cu pHi, fiind o constantă foarte importantă a proteinelor. Fiecare proteină la punctul izoelectric are un comportament specific, având o solubilitate şi reactivitate chimică minimă; de asemenea hidratarea particulelor coloidale, vâscozitatea şi presiunea osmotică sunt de asemenea minime. Precipitarea proteinei la punctul izoelectric este în schimb maximă, dar nu se deplasează sub
influenţa curentului electric. De obicei valorile punctului izoelectric variază între 2,9 şi 12,5 şi se determină prin diferite metode: potenţiometrice, electroforetice.
Precipitarea proteinelor Sub acţiunea diferiţilor factori fizici (ultrasunete, radiaţii cu diferite lungimi de undă,
căldură), factori chimici (acizi, baze, diferiţi solvenţi organici), sau mecanici (agitare), are loc fenomenul de precipitare a proteinelor, precipitarea care poate fi reversibilă sau ireversibilă.
Precipitare reversibilă Precipitarea reversibilă se poate produce sub acţiunea soluţiilor concentrate ale sărurilor
alcaline dar şi în prezenţa unor dizolvanţi organici miscibili cu apa în orice proporţie, cum sunt de exemplu acetona şi alcoolul. În cadrul acestei precipitări molecula proteinei suferă unele modificări fizico-chimice, dar nu are loc afectarea structurii moleculare. Puterea de precipitare a proteinelor de către diferiţi ioni este data de seria liofilă a lui Hofmeister. Dacă anionul rămîne acelaşi, puterea de precipitare a cationilor scade în următoarea ordine: Li+>Na+>NH4
+> cănd cationul rămâne acelaşi anionii se comportă astfel: SO4
2->PO43->CH3COO->Citrat->tartrat->Cl->NO3
-
>ClO3->Br->I->SCN-. Solvenţii de tipul alcoolului sau acetonei, în funcţie de concentraţia lor, pot
forma fie precipitate reversibile, fie ireversibile. Sărurile alcaline au un comportament diferit faţă de proteine, în soluţii diluate mărind solubilitatea proteinelor, iar în soluţii concentrate determinând precipitarea lor reversibilă. De altfel soluţiile sărurilor alcaline de diferite concentraţii se folosesc pentru precipitarea fracţionată a proteinelor din amestecuri.
Precipitare ireversibilă În cursul acestei precipitări molecula proteinei suferă modificări fizico-chimice ireversibile
având loc şi modificarea structurii moleculare. De regulă se produce la adăugarea de soluţii ale metalelor grele (Cu,Pb, Hg, Fe), a acizilor minerali tari (HNO3, H2SO4) acidul tricloracetic, a soluţiilor concentrate de alcool sau acetonă, sau, în cazul anumitor proteine, în prezenţa căldurii. Prin precipitare ireversibilă proteinele îşi pierd activitatea biologică (enzimatică, hormonală, etc.), are loc o descreştere a solubilităţii, modificarea activităţii optice şi, de asemenea, sunt mai uşor de degradat sub acţiunea unor enzime proteolitice. Prin îndepărtarea factorilor care au dus la precipitare, proteinele nu revin la forma lor iniţială şi nu îşi pot reface structura moleculară. Proteinele precipitate îşi pierd din proprietăţile hidrofile "obţinând" proprietăţi hidrofobe.
2.4. Rolul proteinelorDatorită compoziţiei, fiind formate exclusiv din aminoacizi, se întâlnesc alături de alţi
compuşi importanţi de tipul polizaharidelor, lipidelor şi acizilor nucleici începând cu structura virusurilor, a organismelor procariote, eucariote şi terminând cu omul. Practic, nu se concepe viaţă fără proteine. Proteinele pot fi enzime care catalizează diferite reacţii biochimice în organism, altele pot juca un rol important în menţinerea integrităţii celulare (proteinele din peretele celular), în răspunsul imun şi autoimun al organismului.
Majoritatea microorganismelor şi plantelor pot sintetiza toţi cei 20 aminoacizi standard, în timp ce organismele animale obţin anumiţi aminoacizi din dietă (aminoacizii esenţiali). Enzime cheie, cum ar fi de exemplu aspartat kinaza, enzimă care catalizează prima etapă în sinteza aminoacizilor lisină, metionină şi treonină din acidul aspartic, nu sunt prezente în organismele de tip animal. La aceste organisme aminoacizii se obţin prin consumul hranei conţinând proteine. Proteinele ingerate sunt supuse acţiunii acidului clorhidric din stomac şi acţiunii enzimelor numite proteaze, proces în urma căruia lanţurile proteice sunt scindate (denaturate). Ingestia aminoacizilor esenţiali este foarte importantă pentru sănătatea organismului, deoarece fără aceşti aminoacizi nu se poate desfăşura sinteza proteinelor necesare organismului. De asemenea, aminoacizii sunt o sursă importantă de azot; unii aminoacizi nu sunt utilizaţi direct în sinteza proteică, ci sunt introduşi în procesul de gluconeogeneză, proces prin care organismul asigură necesarul de glucoză în perioadele de înfometare (mai ales proteinele aflate în muşchi).
2.5. Structura proteinelor
După cum s-a văzut anterior, lanţurile peptidice sunt formate de grupările carboxil şi aminice a aminoacizilor; există de fapt 2 forme pentru fiecare proteină, numite forme de rezonanţă:
una datorată dublei legături care asigură rigiditatea şi nu permite rotaţia în jurul axei sale; a doua formă de rezonanţă este dată de unghiul diedru φ(planul atomilor C'-N-Cα-C'), ψ
(planul atomilor N-Cα-C'-N), ω (planul atomilor Cα-C'-N-Cα), unghiurile φ şi ψ pot avea diferite valori fiind responsabile de gradul de libertate a proteinelor, controlînd structura tridimensională a lanţului proteic.
Structura substanţelor proteice este încă insuficient cunoscută datorită dinamicităţii structurii proteinelor, deoarece ele sunt în permanenţă supuse unor procese de sinteză şi de degradare.
Structura primară Structura primară este dată de aminoacizii care intră în lanţul proteic prin formarea
legăturilor peptidice. În proteinele naturale, legătura peptidică se stabileşte între gruparea carboxilică de la C1 şi gruparea aminică de la C2, încât lanţul peptidic va fi format dintr-o succesiune de unităţi CO-NH-CH, legate cap la cap.
La unul din capetele lanţului peptidic se găseşte o grupare -NH2 liberă, iar la celălalt capăt se află o grupare -COOH liberă. Legătura peptidică -CO-NH- se găseşte în acelaşi plan, iar carbonul -CH- se poate roti, putând să apară în planuri diferite. Datorită lungimii relativ mici a catenelor laterale, ele se pot aranja de o parte şi de alta a lanţului proteic, astfel că lanţul proteic nu este ramificat.
Datorită deplasării alternative a unui electron de la gruparea -NH la C=O, se produce oscilarea dublei legături de la atomul de carbon şi oxigen la atomul de azot, formându-se astfel cele două forme mezomere.
Datorită numărului relativ mic de aminoacizi care intră în structura proteinelor, teoretic ar trebui să se formeze proteine cu masa moleculară în jur de 4200. Însă în realitate masele moleculare ale proteinelor au valori de peste 10,000 ceea ce a dus la concluzia că cel puţin o parte de aminoacizi se repetă de mai multe ori în cadrul unei molecule. Ipoteza că proteinele sunt formate din lanţuri lineare de aminoacizi a fost fomulată pentru prima dată în anul 1902, la a 74-a reuniune a Societăţii Oamenilor de Știinţă din Germania, ţinută în oraşul Karlsbad, de către Franz Hofmeister (ţinând cont de reacţia biuretului) şi Emil Fischer (care aduce clarificări asupra scheletului proteic). Ipoteza că în molecula proteinelor există legături amidice fusese elaborată de chimistul francez Grimaux încă din anul 1882. În ciuda evidenţelor care demonstrau faptul că proteinele supuse acţiunii proteolitice se scindează în oligopeptide, ideea că lanţul proteic este liniar, au fost idei greu de "digerat". În perioada respectivă, numeroşi savanţi (William Astbury, Hermann Staudinger), punând la îndoială acest lucru, prin argumentarea că legăturile amidice nu sunt îndeajuns de puternice pentru a susţine o moleculă proteică lungă.Cu timpul au apărut diverse ipoteze:
Ipoteza coloidală care susţinea ca proteinele sunt ansambluri moleculare coloidal formate din molecule mai mici - ipoteză contrazisă de măsurarea ultracentrifugării de către Svedberg care arată faptul că proteinele sunt molecule bine definite, au greutate moleculară, iar prin electroforeză Arne Tiselius demonstrează că proteinele sunt molecule unice.
Ipoteza a 2-a, numită ipoteza ciclol, avansată de Dorothy Wrinch, are la bază 3 elemente:

o Ciclol reaction în care gruparea carbonil şi gruparea amino a 2 peptide se incrucişează C=O + HN → C(OH)-N (aşa numita legătură în cruce); aceste legături sunt de tip covalent, similare cu legăturile covalente de hidrogen propuse de William Astbury, pentru a explica stabilitatea structurii proteice.
o Lanţurile beta vecine au la bază o serie de reacţii de tip ciclolo Structura proteinelor mici corespund aşa numitelor "solid de tip Platon", fără să
existe colţuri libere.
Alte ipoteze au fost lansate de către Emil Abderhalden (modelul dicetopiperazinic), sau Troesengaard în anul 1942 (modelul pirol/piperidină). Toate aceste modele au fost infirmate de Frederick Sanger care reuşeşte să identifice secvenţa aminoacizilor din insulină, dar şi de determinările cristalografice efectuate de Max Perutz şi John Kendrew asupra mioglobinei şi hemoglobinei.
Structura secundară Structura secundară se referă la forma şi la lungimea lanţurilor polipeptidice, proprietăţi
induse de legăturile de hidrogen. Cele mai întâlnite tipuri de structura secundară sunt alpha helixul şi lanţurile beta.
Alte helix-uri sunt, din punct de vedere energetic, favorabile formării legăturilor de hidrogen, dar sunt rareori observat în proteinele naturale exceptînd părţile terminale ale helixului α în timpul formării scheletului proteic (de obicei centrul helixului). Aminoacizii au un comportament diferit privind posibilitatea formării structurii secundare. Prolina şi glicina sunt cunoscuţi ca aşa numiţii "spărgători de helix", deoarece afectează configuraţia scheletului proteic; ambii aminoacizi au abilităţi conformaţionale neobişnuite şi de regulă se găsesc în colţurile scheletului proteic. Aminoacizii care preferă să adopte conformaţia helixului proteic fac parte din aşa numita serie MALEK (codurile formate din 1 literă a aminoacizilor: metionină, alanină, leucină, acid glutamic şi lizina); prin contrast aminoacizii aromatici (triptofanul, tirosina şi fenilalanina, dar şi aminoacizii cu legare prin carbonul beta (izoleucina, valina şi treonina, adoptă configuraţia β.
Structura secundară cunoaşte cîteva ipoteze privind formarea ei: Teoria polipeptidică formulată de către E. Hoffmeister în 1902 şi dezvoltată ulterioe de
către E.Fischer, are la bază conceptul conform căruia moleculele proteice sunt formate din lanţuri polipeptidice foarte lungi. Teoria are cîteva dezavantaje:
o nu explică diferenţierea biologică a anumitor proteineo unele proteine sunt rezistente la acţiunea enzimelor proteolitice (deşi datorită
lungimii lanţului nu ar trebui) Teoria plierii şi răsucirii lanţului polipeptidice a fost elaborată de către Corey şi Pauling în
1943 şi a fost confirmată prin spectrele de difracţie cu raze X, microscopului electronic , prin măsurarea unghiurilor de valenţă, a distanţelor interatomice, au confirmat faptul că lanţul polipeptidic se găseşte sub formă pliată.
o Structura în foaie pliantă. Plierea catenei are loc prin formarea legăturilor de hidrogen între gruparea carboxilică a unui aminoacid şi gruparea aminică a aminoacidului vecin. Lanţul polipetidic pliat se prezintză ca o panglică îndoită alternativ la dreapta şi la stînga, plierea avînd loc în dreptul carbonilor metinici. Mai multe lanţuri pliate polipeptidice pliate dau naştere unei reţele, între aceste lanţuri pliate putîndu-se de asemenea forma legături de hidrogen, acestea fiind în număr mai mare când grupările terminale a 2 lanţuri sunt aranjate diferit (-NH2 şi COOH, sau HOOC-şi -NH2). Catenele polipeptidice pliate predomină în proteinele fibrilare şi mai puţin în cele globulare. După valoarea perioadei de identitate se cunosc mai multe

tipuri de proteine cu structură pliată. Prin perioada de identitate se înţelege distanţa cea mai mică la care se repetă aminoacizii identici din moleculă.
o Structura α elicoidală, ipoteză lansată de Corey şi Pauling, ipoteză conform căreia lanţul polipeptidic se poate prezenta şi înfăşurat sub formă de spirală. În acest model, fiecare spiră conţine de obicei 27 aminoacizi, iar distanţa între spire este de 5,44 A0. Fiecare aminoacid măreşte spira cu 1,47 A0. În faţa fiecărei grupări -CO- va apare la o distanţă de 2,8A0. o grupare NH de la al treilea aminoacid. Între aceste grupări se stabilesc punţile de hidrogen care asigură stabilitatea α helix-ului. În acest model lanţul polipeptidic se prezintă sub forma unui surub cu pasul fie spre dreapta, fie spre stînga. În cazul proteinelor naturale, acestea datorită conţinutului în L-aminoacizi, pasul helixului va fi spre dreapta, catenele laterale ies în afara corpului propriu-zis putînd reacţiona fie cu moleculele solventului fie cu alte catene polipeptidice. Canalul format în interiorul helixului este foarte îngust, în el nu poate pătrunde molecula solventului. Legăturile peptidice sunt plane, iar 2 planuri consecutive -CO-NH- formează un unghi de 1800, rotirea lanţului se face la carbonul α(metinic).
Structura terţiară Prin intermediul cristalografiei cu raze X s-a dovedit faptul că macromoleculele proteice au
o conformaţie tridrimensională, realizată de obicei prin intermediul cuplării mai multor lanţuri polipeptidice scurte între ele, cuplare care duce la formarea fibrelor proteice;legăturile intercatenare pot fi principale sau secundare:
Legături de hidrogen, sunt legături coordinativ heteropolare care se stabilesc cu uşurinţă între gruparea carbonil C=O (electronegativă) şi gruparea NH- (electropozitivă), din 2 lanţuri polipeptidice alăturate, sau în cazul formelor lactam-lactimă între gruparea -OH şi azotul iminic =NH.
Legăturile de hidrogen au lungimea cuprinsă între 2,7-3,1A şi energia de 3-7Kcal/mol la peptide, iar la apă 2-3Kcal/mol. Legăturile de hidrogen se pot stabili şi între catenele lateralecare au grupări carboxil, hidroxil, amino sau tiolice. Din punct de vedere energetic[17]legătura de hidrogen nu este puternică dar datorită răspîndirii relativ uniforme de-a lungul scheletului proteic oferă proteinei stabilitatea necesară.În afară de legăturile disulfidice, se mai pot stabili alte tipuri de legături: legături ionice (stabilite de obicei între grupările aminice şi cele carboxilice ionizate), legături de tip van der Waals (legături electrostatice slabe care se stabilesc între radicalii hidrofobi), legături fosfodiesterice (între 2 resturi de serină şi acid fosforic), legături eterice (stabilite la nivelul aminoacizilor cu grupări hidroxilice).
Structura cuaternară Structura cuaternară se referă la modul în care se unesc subunităţile proteice. Enzimele
care catalizează asamblarea acestor subunităţi poartă denumirea de holoenzime, în care o parte poartă denumirea de subunităţi reglatoare şi subunităţi catalitice.Proteine care au structura cuaternară: hemoglobina, ADN polimeraza şi canalele ionice, dar şi nucleozomi şi nanotubuli, care sunt complexe multiproteice. Fragmentele proteice pot suferi transformări în structura cuaternară, transformări care se reflectă fie în structurile individuale fie în reorientările fiecărei subunităţi proteice. Numărul subunităţilor din oligomerice sunt denumite prin adăugarea sufixului -mer (grecescul pentru subunitate), precedat de numele subunităţii.

3. Bibliografie
www.wikipedia.org www.referat.ro www.referat.unica.ro Vlădescu, Badea, Doicin - Chimie C1. Manual pentru clasa a XI-a, Grup
Editorial Art, 2006 Alexandrescu, Dănciulescu - Chimie organică pentru liceu, Editura LVS
Crepuscul, 2009 Arsene, Popescu - Chimie şi probleme de chimie organică, Editura Tehnică,
1979 Alexandrescu, Zaharia, Nedelcu - Chimie X, Editura LVS Crepuscul, 2005 Vlădescu, Tăbărăşanu-Mihăilă, Doicin - Chimie. Manual pentru clasa a X-a,
Grup Editorial Art, 2005 Loloiu, Baciu, Bogdan - Chimie C1. Manual pentru clasa a XI-a, Editura ALL,
2001


















