
Recomandări de monitorizare pentru habitatul 6230* Pajişti ...LIFE05NAT/RO/000176 – “Habitate...
79
Marius Ioan Bărbos Dumitru Romulus Târziu PROIECT LIFE05 NAT/RO/000176: „Habitate prioritare alpine, subalpine şi forestiere din România!” Recomandări de monitorizare pentru habitatul 6230* Pajişti de Nardus stricta bogate în specii pe substraturi silicioase Braşov 2009
Transcript of Recomandări de monitorizare pentru habitatul 6230* Pajişti ...LIFE05NAT/RO/000176 – “Habitate...

Marius Ioan Bărbos Dumitru Romulus Târziu
PROIECT LIFE05 NAT/RO/000176: „Habitate prioritare alpine, subalpine şi forestiere din România!”
Recomandări de monitorizare pentru habitatul 6230* Pajişti de Nardus stricta bogate în specii pe
substraturi silicioase
Braşov 2009

2
© Facultatea de Silvicultură şi Exploatări Forestiere,
Universitatea „Transilvania“ din Braşov, 2009
Autori: Marius Ioan Bărbos şi Dumitru Romulus Târziu Referent ştiinŃific: Ioan Vasile Abrudan La baza acestui studiu au stat cercetările în teren desfăşurate în cadrul proiectului LIFE05NAT/RO/000176 cât şi informaŃii din alte lucrări de specialitate în domeniu. Lucrarea a fost realizată cu sprijin financiar din partea ComunităŃii Europene prin instrumentul financiar LIFE. Toate drepturile asupra acestei ediŃii aparŃin FacultăŃii de Silvicultură şi Exploatări Forestiere din Universitatea „Transilvania” Braşov. Orice reproducere parŃială sau totală fără acordul acestei instituŃii este interzisă. Studiul a fost elaborat în cadrul proiectului: LIFE05NAT/RO/000176 - "Habitate prioritare alpine, subalpine şi forestiere din România" BENEFICIAR Facultatea de Silvicultură şi Exploatări Forestiere Şirul Beethoven, Nr. 1 500123 – Braşov, România Tel: + 40 268 418 600 Fax: + 40 268 475 705 http://www.unitbv.ro/silvic/
PARTENERI: Ministerul Agriculturii, Pădurilor şi Dezvoltării Rurale B-dul. Carol I, nr. 24, Sector 3, 020921 – Bucureşti, România Tel: + 40 21 307 23 00 E-mail: [email protected] http://www.maap.ro/ Regia NaŃională a Pădurilor – ROMSILVA B-dul. Magheru, Nr 31 Sector 1, 010325 - Bucureşti, România Tel: + 40 21 317 10 05 Fax: + 40 21 316 84 28 E-mail: [email protected] http://www.rosilva.ro/ World Wide Fund for Nature – Danube Carpathian Programme Mariahilferstrasse, 88a/3/9 1070 – Viena, Austria Tel: + 43 1 524 54 70 Fax: + 43 1 524 54 70 70 http://www.panda.org

3
CUPRINS CUPRINS ....................................................................................................................................... 3 INTRODUCERE ........................................................................................................................... 5 I. MONITORIZAREA HABITATELOR ...................................................................................... 7 1.1. GeneralităŃi .......................................................................................................................... 7 1.2. Planificarea acŃiunilor de monitorizare ............................................................................... 7 1.2.1. Definirea obiectivelor .................................................................................................. 8 1.2.2. Evaluarea resurselor ..................................................................................................... 8 1.2.3. Alegerea atributelor şi a metodelor de lucru .............................................................. 10 1.2.4. Colectarea datelor ...................................................................................................... 10 1.2.4.1. Eşantionarea habitatelor ...................................................................................... 11 1.2.4.1.1. Tipul de eşantionaj ....................................................................................... 11 1.2.4.1.2. Mărimea suprafeŃelor de probă .................................................................... 12 1.2.4.1.3. Numărul şi forma suprafeŃelor de probă ...................................................... 13 1.2.4.1.4. SuprafeŃe de probă permanente vs. temporare ............................................. 13 1.2.4.1.5. Perioada optimă de colectare a datelor ........................................................ 14 1.2.4.1.6. FrecvenŃa colectării datelor .......................................................................... 15
1.2.4.2. Structura habitatelor ............................................................................................ 15 1.2.4.2.1. Structura calitativă ....................................................................................... 15 1.2.4.2.2. Structura spaŃială .......................................................................................... 17
1.2.4.3. Variabilele de mediu ........................................................................................... 17 1.2.5. Stocarea şi gestionarea datelor ................................................................................... 18 1.2.5. Analiza şi interpretarea rezultatelor ........................................................................... 19 1.2.6. Elaborarea raportului de monitorizare ....................................................................... 20
II. STAREA FAVORABILĂ PENTRU CONSERVAREA HABITATULUI 6230* ................ 22 2.1. Descrierea habitatului ....................................................................................................... 22 2.2. Starea favorabilă pentru conservare .................................................................................. 24 2.2.1. Atribute cantitative ..................................................................................................... 25 2.2.2. Structura calitativă ..................................................................................................... 30 2.2.3. Structura spaŃială ........................................................................................................ 31 2.2.4. Caracteristici edafice .................................................................................................. 33
III. PLANUL DE MONITORIZARE AL HABITATULUI 6230* ............................................. 35 3.1. Studii preliminare .............................................................................................................. 35 3.2. Colectarea datelor ............................................................................................................. 36 3.2.1. Eşantionarea ............................................................................................................... 36 3.2.2. Releveul fitosociologic .............................................................................................. 37 3.2.3. Structura calitativă a habitatelor (sinstructura) .......................................................... 37 3.2.4. Indicii fitopopulaŃionali ............................................................................................. 42 3.2.5. Structura spaŃială ........................................................................................................ 44 3.2.6. Variabilele de mediu .................................................................................................. 44 3.2.6.1. Factorii topografici .............................................................................................. 44

4
3.2.6.2. Factorii edafici .................................................................................................... 45 3.2.6.3. Factorii climatici ................................................................................................. 46 3.2.6.4. Factorii biotici ..................................................................................................... 46
3.3. Stocarea şi gestionarea datelor .......................................................................................... 47 3.4. Analiza datelor .................................................................................................................. 47 3.4.1. Transformarea datelor ................................................................................................ 48 3.4.2. Indici fitocenotici ....................................................................................................... 48 3.4.3. Clasificarea vegetaŃiei şi analiza cluster .................................................................... 56 3.4.4. Ordonarea vegetaŃiei .................................................................................................. 58 3.4.5. Analiza „arborilor” de regresie .................................................................................. 60 3.4.6. Răspunsul speciilor şi gradul de suprapunere al nişelor ............................................ 62
3.5. Elaborarea rapoartelor de monitorizare ............................................................................ 64 CONCLUZII ................................................................................................................................ 65 BIBLIOGRAFIE .......................................................................................................................... 66 ANEXA I ..................................................................................................................................... 78

5
INTRODUCERE
Monitorizarea „stării de sănătate” a habitatelor, şi în special a habitatelor Natura 2000 a devenit o cerinţă obligatorie odată cu aderarea României la Uniunea Europeană şi implementarea Reţelei Natura 2000 în ţara noastră. Dată fiind lipsa de experienţă în domeniul monitoringului ecologic, elaborarea unui ghid metodologic, chiar şi pentru un singur tip de habitat, devine nu numai necesară ci şi obligatorie.
Elaborarea materialului de faţă s-a dovedit a fi o adevărată provocare, atât prin gradul de noutate cât şi prin complexitatea temei abordate. Pentru atingerea obiectivelor, monitoringul ecologic apelează în principal la metode de lucru care au fost dezvoltate în cadrul altor ştiinţe, dar care au fost adaptate cerinţelor specifice monitorizării habitatelor.
Una dintre principalele piedici întâlnite a fost lipsa datelor ştiinţifice necesare alegerii indicatorilor (atributelor) care definesc starea favorabilă pentru conservare a habitatelor şi a valorilor pe care aceşti indicatori le pot lua. Deoarece habitatele sunt descrise prin prisma caracteristicilor structurale, dinamice şi funcţionale ale fitocenozelor constituente în relaţie cu ecotopul, şi mai puţin din prisma celorlaltor componente ale biocenozelor (animale, microorganisme etc), atributele care trebuie monitorizate, şi implicit şi metodele utilizate pentru măsurarea lor, au fost alese din domeniul ecologiei generale, ecologiei plantelor şi a fitosociologiei.
Datorită multidisciplinarităţii şi complexităţii metodologie de lucru (identificarea habitatelor pe teren, a speciilor, colectarea datelor, analiza datelor şi interpretarea rezultatelor), lucrarea de faţă nu este accesibilă publicului larg, monitorizarea habitatelor nu va putea fi făcută decât de către echipe de specialişti. Din aceste considerente, în realizarea lucrării s-a pus accent mai mult pe acele noţiuni şi metode care prezintă un anumit grad de noutate, metodele clasice, cu care cei mai mulţi sunt deja familiarizaţi, fiind doar amintite făcându-se şi trimiteri bibliografice.
Fără a avea pretenţia de a fi epuizat în întregime subiectul, lucrarea de faţă se doreşte un ghid util de iniţiere în tehnica monitorizării habitatelor Natura 2000 în general şi al habitatului 6230* (pajişti de Nardus stricta bogate în specii pe substrat silicios) în mod special.
Ajungând la finalizarea studiului privind recomandările de monitorizare a habitatului 6230*, dorim să aducem mulţumirile noastre tuturor acelora care ne-au ajutat pe parcursul elaborării lucrării de faţă. În primul rând mulţumim echipei de implementare a proiectului LIFE05NAT/RO/000176 – “Habitate prioritare alpine, subalpine şi forestiere în România”, proiect finanţat de Uniunea Europeană şi derulat şi coordonat de Facultatea de Silvicultură şi Exploatări Forestiere din cadrul Universităţii "Transilvania" – Braşov, în parteneriat cu Ministerul Agriculturii şi Dezvoltării Rurale, Regia Naţională a Pădurilor – ROMSILVA şi Fondul Mondial pentru Natură – Programul Dunăre Carpaţi (WWF – DCP), care ne-au oferit tot sprijinul logistic necesar finalizării studiului şi elaborării acestui material. Dorim să mulţumim şi colectivului Laboratorului de Pedologie al Facultăţii de Silvicultură şi Exploatări Forestiere din

6
cadrul Universităţii Transilvania din Braşov care ne-au ajutat cu analizele probelor de sol, analize care au permis identificarea principalelor atribute edafice care definesc starea favorabilă pentru conservare a habitatului studiat. De asemenea, adresăm mulţumirile noastre tuturor acelora care, sub o formă sau alta, ne-au sprijinit în munca noastră.

7
I. MONITORIZAREA HABITATELOR
1.1. Generalităţi
Monitorizarea reprezintă o măsurare sistematică de-a lungul timpului a unor variabile sau procese ecologice în concordanţă cu un scop bine precizat astfel încât să fie îndeplinite anumite standarde (ex. starea favorabilă pentru conservarea habitatelor, a speciilor etc) (Spellerberg 2005). O definiţie mai riguroasă a monitorizării ecologice este dată de către Hellawell (1991 ap Hill et al. 2005), monitorizarea fiind considerată „ ... observaţii intermitente
(regulate sau neregulate) realizate pentru a determina mărimea deviaţie faţă de un standard
predefinit sau gradul de deviaţie de la nişte rezultate aşteptate”. Monitorizarea speciilor sau habitatelor, pe lângă informaţiile pe care le oferă despre starea lor de conservare sau despre rezultatul diferitelor măsuri de management activ, face posibilă şi identificarea timpurie a unor tendinţe dinamice având un rol important în predicţia modificărilor structurale şi funcţionale, fapt ce permite luarea unor măsuri, în timp util, pentru conservarea acestora. Cunoaşterea bazată pe informaţiile obţinute în cadrul programelor de monitorizare asigură o mai bună înţelegere a problematicii analizate, fapt ce determină o îmbunătăţire a şanselor ca deciziile luate să fie bine documentate şi corecte (Artiola et al. 2004). Pe lângă aplicaţiile practice, monitorizarea ecologică are şi o importanţă teroretică, informaţiile obţinute având un rol deosebit de important în descifrarea legităţilor care determină structura, funcţiile şi dinamica ecosistemelor.
Monitorizarea habitatelor se face pe baza unor protocoale (planuri) de monitorizare care permit colectarea şi analiza datelor într-o formă standardizată, astfel încât datele colectate de persoane diferite la intervale de timp diferite să fie comparabile între ele şi să aibă aceaşi valoare informaţională.
1.2. Planificarea acţiunilor de monitorizare
Planificarea şi implementarea programelor de monitorizare necesită angajament şi viziune pe termen lung, pe termen scurt aceste programe reprezentând costuri adiţionale (Christensen et al. 1996).
Acţiunile de monitorizare a habitatelor nu se pot face la întâmplare, ci, aşa cum am arătat, trebuie să se realizeze într-o manieră standardizată. Planificarea acţiunilor de monitorizare este o etapă importantă şi obligatorie a oricărui program de monitorizare indiferent de scara la care acesta se realizează. Iniţierea şi dezvoltarea cu succes a unui

8
program de monitorizare depinde de claritatea cu care s-a definit scopul şi obiectivele acelei acţiuni. Monitorizarea poate avea o gamă largă de obiective, de la monitorizarea stării de conservare a unei singure populaţii sau a unui habitat, până la monitorizarea impactului pe care anumite activităţi umane le are asupra mediului sau a influenţelor exercitate de către schimbările climatice. Această primă etapă se finalizazează cu elaborarea protocoalelor de monitorizare care, aşa cum am amintit, permit identificarea clară şi pe termen lung a obiectivelor urmărite, alegerea variabilelor (atributelor) care vor fi măsurate şi a metodelor de colectare, stocare, gestionare şi analiză a datelor, precum şi a modului de elaborare şi prezentare a rapoartelor de monitorizare.
1.2.1. Definirea obiectivelor
Definirea obiectivelor constituie una dintre cele mai importante etape ale fiecărui program de monitorizare a habitatelor. Definirea eronată a obiectivelor urmărite iniţiază o reacţie în lanţ care poate determina eşecul întregului program de monitorizare, datele colectate neputând să ofere răspunsurile aşteptate la întrebările pe care noi am dorit să le adresăm, datele dovedindu-se astfel inutile. Obiectivele propuse sunt cele care condiţionează alegerea atât atributele care trebuie monitorizate, cât şi a metodelor utilizate pentru a măsura valorile acestora.
Procesul de definire a obiectivelor sprijină principiile de bune practici de management şi elaborarea planului de management în care monitorizarea trebuie să fie parte integrată (Hill et al. 2005). De aceea este important ca în definirea obiectivelor să răspundem la cel puţin două întrebări esenţiale, şi anume:
- „La ce scară se va realiza monitorizarea?” - „Ce anume trebuie să cunoaştem?”
Răspunsurile la aceste întrebări vor influenţa în mod direct complexitatea programului de monitorizare, costurile necesare realizării obiectivelor şi implicit şi metodologia de lucru aleasă pentru implementarea programului. Pentru definirea obiectivelor se vor analiza toate informaţiile publicate pentru tipul de habitat, situl şi regiunea care face obiectul programului de monitorizare (studii ecologice, inventar floristic, studii de vegetaţie, harţi de vegetaţie, topografice, geologice, edafice, planuri de management etc) astfel încât obiectivele propuse să fie cât mai clare şi realizabile. În cazul în care pentru un anumit habitat, sit sau regiune nu există publicate astfel de date, este imperios necesară efectuarea unor studii ecologice preliminare cât se poate de riguroase.
1.2.2. Evaluarea resurselor
Îndeplinirea obiectivelor propuse în programul de monitorizare depinde de resursele financiare, materiale şi umane disponibile. Imposibilitatea asigurării pe termen lung a acestor resurse duce la imposibilitatea realizării obiectivelor propuse şi implicit la eşecul întregului program. În condiţiile în care se constată că resursele existente nu sunt suficiente pentru îndeplinirea obiectivelor propuse, acestea se vor redefini astfel încât să se reducă costurile până la nivelul resurselor existente fără a se reduce valoarea informaţională a rezultatelor obţinute.

9
Definirea obiectivelorStudii ecologice
preliminare
Evaluarea
resurselor
Alegerea variabilelor de
măsurat şi a metodelor de
colectare şi analiză a datelor
Colectarea datelor
Stocarea şi
gestionarea
datelor
Analiza datelor şi
interpretarea
rezultatelor
Evaluarea
îndeplinirii
obiectivelor
Elaborarea
raportului de
monitorizare
Date
publicate
Figura 1. Planul conceptual al planificării programelor de monitorizare

10
1.2.3. Alegerea atributelor şi a metodelor de lucru
Obiectivele propuse în cadrul programului de monitorizare nu pot fi atinse fără o
alegerea riguroasă a atributelor care trebuie măsurate, a tipului de date colectate şi a metodelor utilizate pentru colectarea acestora. Alegerea variabilelor care vor fi monitorizate şi a metodelor de lucru se va face în acelaşi timp cu definirea obiectivelor, cele două procese fiind strâns corelate între ele. Pentru fiecare obiectiv propus se vor identifica acele caracteristici (atribute) ale habitatelor care reflectă proprietăţile acestora şi pot fi cuantificate (Hill et al. 2005). Atributele habitatelor pot fi cantitative (suprafaţă), structurale ( structura specifică, tipuri de comunităţi vegetale, bogăţia specifică, structura spaţială etc), funcţionale (productivitate, capacitate de suport, reglajul climatic etc.) sau dinamice (fluctuaţii, degenarare, regenerare, succesiuni etc). Fiecărui atribut, acolo unde informaţiile existente permit acest lucru, i se va defini un interval de valori pe care le poate lua, fără ca proprietăţile habitatelor analizate să fie alterate, uşurându-se astfel interpretarea rezultatelor.
Uneori, datorită complexităţii structurale şi funcţionale mari a habitatelor, dar şi a costurilor ridicate, vor fi monitorizate doar unele specii cheie, tipice, indicatoare şi/sau invazive, care pot să ofere informaţii valoroase asupra stării curente a habitatelor. În cazul monitorizării speciilor, atributele cantitative se referă la mărimea populaţiilor, densitate, acoperirea realizată de fiecare populaţie, cele calitative vor fi reprezentate de starea fenologică şi vitalitatea indivizilor, în timp ce atributele dinamice vor cuantifica informaţiile privind mortalitatea, natalitatea, emigrarea şi imigrarea fitoindivizilor.
În alegerea variabilelor care vor face obiectul activităţilor de monitorizare a habitatelor se va ţine cont de criteriile propuse de către Hunsaker et al. (1990) (Spellerberg 2005) şi anume:
1. Să se coreleze cu schimbările survenite în structura şi funcţiile habitatului; 2. Să fie adecvate pentru o monitorizare la nivel regional (naţional); 3. Să aibă efecte integrate în timp şi spaţiu; 4. Să nu fie ambigue; 5. Să poată fi cuantificate; 6. Să poată fi corelate cu structura şi funcţiile generale ale ecosistemului; 7. Să fie sensibile la acţiunile factorilor de stres sau la diferitele măsuri de
management; 8. Să poată fi măsurate într-o formă standardizată; 9. Să aibă o eroare de măsurare mică; 10. Să fie ieftine.
1.2.4. Colectarea datelor
Colectarea datelor pe teren se va face pe baza unui protocol stabilit în etapele anterioare. Natura, corectitudinea şi acurateţea datelor înregistrate pe teren vor influenţa în mod direct rezultatul tuturor celorlaltor analize şi prelucrări care vor fi efectuate şi implicit rezultatul final al programului de monitorizare. Înaintea începerii colectării datelor se recomandă organizarea unor sesiuni de training cu întreg personalul implicat în acest proces,

11
astfel încât fiecare persoană în parte să-şi însuşească cât mai temeinic metodologia de lucru şi modul de utilizare al aparaturii utilizate. Această acţiune este necesară în vederea minimalizării erorilor de măsurare, care pot să apară în special în cazul acelor variabile care se estimează vizual (abundenţa-dominanţa speciilor, acoperirea generală cu vegetaţie etc) şi care includ din start o doză mare de subiectivism.
1.2.4.1. Eşantionarea habitatelor
Datorită suprafeţei mari pe care o ocupă diversele habitate, nu se poate realiza un
studiu integral ale acestora, iar una dintre problemele delicate cu care se confruntă cercetătorul, este aceea a modului de amplasare a suprafeţelor de probă (eşantioanelor), mărimea, forma şi numărul lor.
1.2.4.1.1. Tipul de eşantionaj
Tipul de eşantionaj se referă la modul de alegere/amplasare a suprafeţelor de probă în cadrul cărora se vor înregistra valorile atributelor monitorizate.
Cele mai cunoscute metode de eşantionare sunt: Eşantionajul selectiv (subiectiv), care presupune ca suprafeţele de probă să fie alese în mod subiectiv, în funcţie de anumite criterii alese de cercetător (Kent et Coker 1992). Această metodă de eşantionare, deşi nu permite obţinerea unor rezultate semnificative din punct de vedere statistic, poate fi utilizată atunci când, scopul studiul efectuat este de clasificare cenotaxonomică a fragmentelor de vegetaţie, şi când vor fi alese suprafeţele de probă cele mai reprezentative. Eşantionajul aleatoriu, este cel mai comun şi uşor de utilizat şi care presupune ca fiecare punct din suprafaţa analizată să aibă şanse egale de a fi ales în cadrul unui studiu (Kent et Coker 1992, Krebs 1998, Artiola et al. 2004). Pentru aceasta, în cadrul suprafeţei analizate, se va stabili o reţea de coordonate, iar cu ajutorul tabelelor de numere aleatorii se vor alege eşantioanele care vor fi inventariate. Eşantionajul sistematic, în cadrul căruia, amplasarea eşantioanelor se face la intervale regulate, a căror lungime este fixă (Kent et Coker 1992, Krebs 1998). Cel mai comun mod de eşantionaj sistematic este cel în care suprafaţa de studiu este împărţită în pătrate egale iar eşantioanele sunt localizate, întotdeauna, în centrul acestor pătrate (Krebs 1998). Eşantionajul stratificat, presupune ca suprafaţa luată în studiu să fie împărţită în suprafeţe relativ omogene (tipuri de vegetaţie, condiţii pedoclimatice, orografice etc.), iar în cadrul fiecărei diviziuni, alegerea suprafeţelor de probă se va face în mod aleatoriu (Kent et Coker 1992, Krebs 1998). Eşantionajul stratificat este o metodă mult mai bună şi mai precisă decît celelate tipuri de eşantionaj, şi relativ uşor de utilizat pe teren (Krebs 1998). Acest tip de eşantionaj se recomandă pentru a fi utilizat în monitorizarea habitatelor cu o structură heterogenă, habitate în structura cărora se regăsesc mai multe tipuri de vegetaţie sau care se întâlnesc pe mai multe tipuri de substrat, pentru fiecare tip de comunitate vegetală/substrat efectuîndu-se ulterior un eşantionaj aleatoriu.
12
Eşantionajul adaptativ grupat, este utilizat mai ales în studiul populaţiilor rare sau în cazul când se urmăreşte studiul unor fragmente de habitate care sunt răspândite pe suprafeţe reduse. Când în cadrul unui eşantion apare o specie rară, sau o fitocenoză cu distribuţie spaţială redusă, următoarele eşantioane vor fi plasate în vecinătatea eşantionului original (Krebs 1998). În alegerea modului de amplasare (eşantionare) a habitatelor se va ţine cont şi de cerinţele pe care datele colectate trebuie să le îndeplinească astfel încât acestea să fie satisfăcute din punct de vedere statistic.
1.2.4.1.2. Mărimea suprafeţelor de probă
Un alt aspect destul de controversat, în studiile de monitorizare a habitatelor, este cel
legat de mărimea suprafeţelor de probă, mărime care este direct proporţională cu complexitatea structurii habitatului studiat. Alegerea suprafeţei eşantioanelor se bazează pe conceptul de areal minim care este definit ca suprafaţa minimă pe care trebuie efectuat releveul de vegetaţie pentru a surprinde majoritatea speciilor care intră în compoziţia fitocenozelor (Cristea et al. 2004). Metoda clasică de estimare a mărimii suprafeţei de probă presupune construirea curbei areal-specie, pe baza numărului de specii înregistrate în suprafeţe de probă a căror mărime creşte exponenţial (Braun-Blanquet 1932, Borza et Boşcaiu 1965, Moravec 1973, Cristea 1993, Cristea et al. 2004). O altă metodă este cea propusă de către Du Reitz, Fries, Osvald et Tengwall (1920), şi care se bazează pe construirea curbei areal-frecvenţă, curbă care prezintă grafic relaţia dintre mărimea suprafeţelor de probă şi numărul speciilor cu frecvenţă între 91-100% (Moravec 1973).
Dintre metodele care se bazează pe analiza statistică a vegetaţiei putem aminti curbele areal-similaritate floristică medie şi metoda propusă de Moravec (1973), curba areal-indice de omotonitate. Toate aceste metode au ca punct comun, faptul că, suprafaţa minimă a eşantionului este considerată suprafaţa la care curbele realizate tind să se aplatizeze.
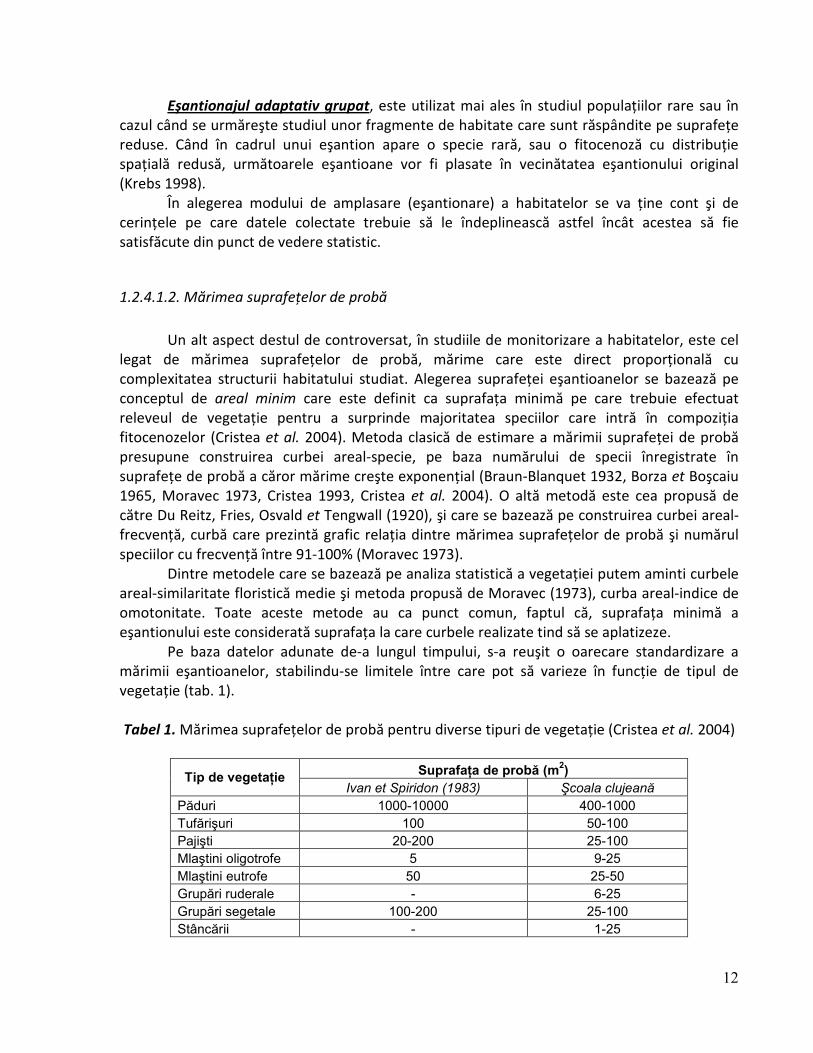
Pe baza datelor adunate de-a lungul timpului, s-a reuşit o oarecare standardizare a mărimii eşantioanelor, stabilindu-se limitele între care pot să varieze în funcţie de tipul de vegetaţie (tab. 1). Tabel 1. Mărimea suprafeţelor de probă pentru diverse tipuri de vegetaţie (Cristea et al. 2004)
Tip de vegetaŃie SuprafaŃa de probă (m2)
Ivan et Spiridon (1983) Şcoala clujeană
Păduri 1000-10000 400-1000
Tufărişuri 100 50-100
Pajişti 20-200 25-100
Mlaştini oligotrofe 5 9-25
Mlaştini eutrofe 50 25-50
Grupări ruderale - 6-25
Grupări segetale 100-200 25-100
Stâncării - 1-25

13
1.2.4.1.3. Numărul şi forma suprafeţelor de probă
O altă problemă majoră a studiilor de monitorizare ecologică este aceea legată de numărul de eşantioane necesar pentru o estimare adecvată a parametrilor care caracterizează o variabilă cu o distribuţie normală (Artiola et al. 2004). Stabilirea numărului de eşantioane necesar pentru estimarea valorii medii a variabilelor măsurate trebuie să ţină cont de toleranţa sau eroare acceptată dar şi de intervalul de încredere dorit.
Numărul de suprafeţe de probă necesare pentru monitorizarea unui habitat depinde în primul rând de suprafaţa ocupată de acesta în zona de studiu dar şi de complexitatea structurală şi variabilitatea spaţială a acestuia. În alegerea suprafeţelor de probă, Kent et Coker (1992) recomandă utilizarea formelor pentru care efectul de margine (raportul perimetru/suprafaţă) este redus. Forma ideală este cea circulară, dar rezultate bune se obţin şi prin utilizarea eşantioanelor de formă pătrată sau dreptunghiulară cu dimensiunile laturilor în raport de 2:5, care permit inventarirea majorităţii speciilor componente, mai ales dacă dreptunghiul este orientat pe direcţia celui mai evident gradient ecologic (Cristea et al. 2004).
1.2.4.1.4. Suprafeţe de probă permanente vs. temporare
În funcţie de obiectivele propuse se va decide dacă suprafeţele de probă alese vor fi permanente (variabilele studiate vor măsurate de fiecare dată în aceleaşi suprafeţe de probă) sau vor fi temporare (de fiecare dată vor fi alese alte suprafeţe de probă în care se vor măsura variabilele studiate). Principalele avantaje şi dezavantaje ale suprafeţelor de probă permanente (Elzinga et al. 1998, Hill et al. 2005) sunt: I. Avantaje - o mai bună identificare a schimbărilor structurale şi a tendinţelor dinamice; - eroarea standard a mediei valorilor variabilelor măsurate este mai mică decât în cazul suprafeţelor de probă temporare; - variaţia bogaţiei specifice este mult mai bine evidenţiată în cazul suprafeţelor de probă permanente decât în cazul celor temporare; - sunt mult mai eficiente în monitorizarea populaţiilor speciilor rare. II. Dezavantaje - marcarea suprafeţelor de probăpermanente poate fi costisitoare şi mare consumatoare de timp; - efectuarea observaţiilor pe aceleaşi suprafeţe poate duce la alterarea sau deteriorarea atributelor care sunt monitorizate, pe lângă faptul că este inacceptabil din punct de vedere al conservării mai poate determina şi pierderea reprezentativităţii eşantionului pentru habitatul studiat; - suprafeţele de probă permanente pot fi afectate de evenimente neprevăzute. Alegerea tipurilor de suprafeţe de probă (permanente sau temporare) se va face ţinând cont de particularităţile fiecărui tip de habitat în parte.
14
1.2.4.1.5. Perioada optimă de colectare a datelor
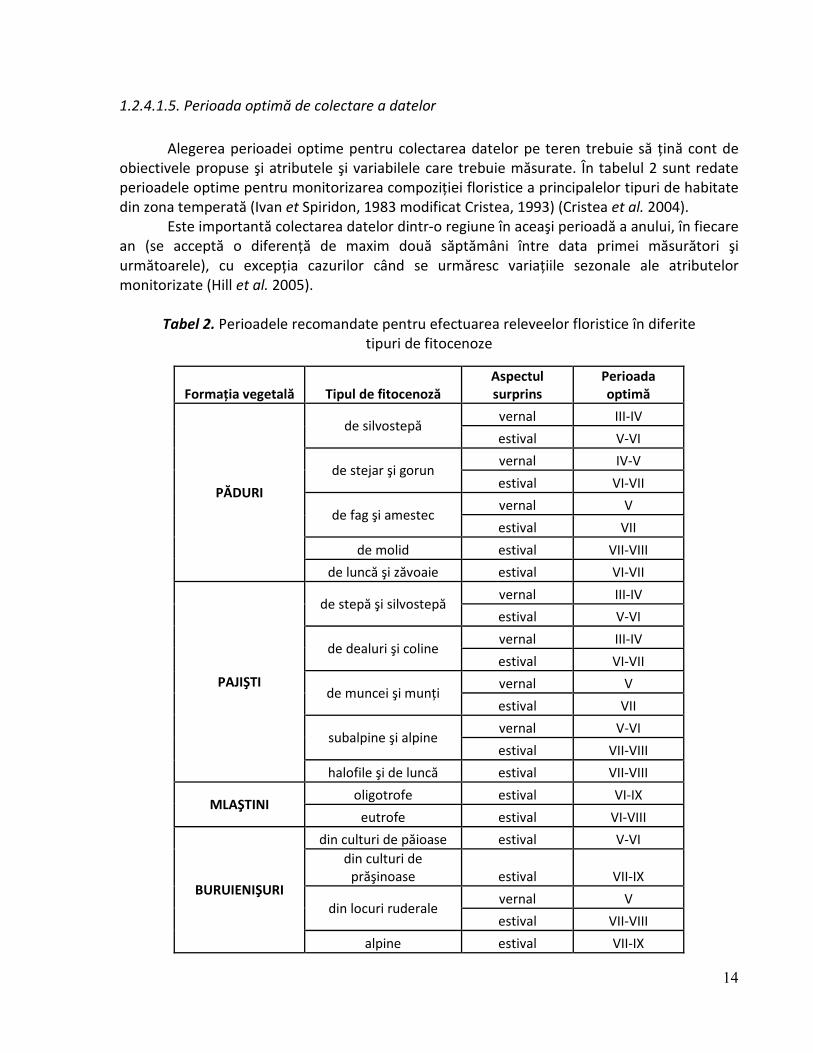
Alegerea perioadei optime pentru colectarea datelor pe teren trebuie să ţină cont de obiectivele propuse şi atributele şi variabilele care trebuie măsurate. În tabelul 2 sunt redate perioadele optime pentru monitorizarea compoziţiei floristice a principalelor tipuri de habitate din zona temperată (Ivan et Spiridon, 1983 modificat Cristea, 1993) (Cristea et al. 2004).
Este importantă colectarea datelor dintr-o regiune în aceaşi perioadă a anului, în fiecare an (se acceptă o diferenţă de maxim două săptămâni între data primei măsurători şi următoarele), cu excepţia cazurilor când se urmăresc variaţiile sezonale ale atributelor monitorizate (Hill et al. 2005).
Tabel 2. Perioadele recomandate pentru efectuarea releveelor floristice în diferite tipuri de fitocenoze
Formaţia vegetală Tipul de fitocenoză Aspectul surprins
Perioada optimă
PĂDURI
de silvostepă vernal III-IV
estival V-VI
de stejar şi gorun vernal IV-V
estival VI-VII
de fag şi amestec vernal V
estival VII
de molid estival VII-VIII
de luncă şi zăvoaie estival VI-VII
PAJIŞTI
de stepă şi silvostepă vernal III-IV
estival V-VI
de dealuri şi coline vernal III-IV
estival VI-VII
de muncei şi munţi vernal V
estival VII
subalpine şi alpine vernal V-VI
estival VII-VIII
halofile şi de luncă estival VII-VIII
MLAŞTINI oligotrofe estival VI-IX
eutrofe estival VI-VIII
BURUIENIŞURI
din culturi de păioase estival V-VI
din culturi de prăşinoase estival VII-IX
din locuri ruderale vernal V
estival VII-VIII
alpine estival VII-IX

15
1.2.4.1.6. Frecvenţa colectării datelor
Frecvenţa de eşantionare a habitatelor în vederea monitorizării lor depinde de o serie de factori cum ar fi: - obiectivele programului de monitorizare; - structura calitativă a habitatelor; - structura spaţială a habitatelor şi gradul lor de fragmentare; - intensitatea acţiunii factorilor perturbatori care pot determina procese dinamice reversibile sau succesionale etc. Pentru habitatele de pajişti, aupra cărora nu acţionează factori perturbatori de mare intensitate, monitorizarea se va face odată la 3-6 ani (Hill et al. 2005). Dat fiind faptul că în România nu s-au derulat astfel de programe de monitorizare, recomandăm, ca în primii ani de la demararea acţiunilor de monitorizare şi în funcţie de resursele disponibile, colectarea datelor să se facă anual, astfel încât stabilirea frecvenţei de monitorizare a fiecărui tip de habitat să se realizeze pe baza caracteristicilor structurale, funcţionale şi dinamice ale acestora.
1.2.4.2. Structura habitatelor
Caracterizarea structurii habitatelor se va face cu ajutorul releveului fitosociologic care poate fi definit ca „ metodă de bază ţn studiul calitativ şi cantitaiv al vegetaţiei, constând într-o succesiune de observaţii şi determinări (marea majoritate efectuate pe teren), finalizate prin transpunerea grafică a ambianţei eco-cenotice dintr-o suprafaţă de probă (fragment) delimitat în interiorul individului de asociaţie (fitocenozei)” (Cristea et al. 2004). Realizarea cu succes a acestei etape depinde de pregătirea teoretică şi practică a personalului care va efectua aceste ridicări pe teren.
1.2.4.2.1. Structura calitativă
În studiul şi monitorizarea structurii calitative a habitatelor se va urmări evidenţierea complexului de specii şi a diferitelor grupe funcţionale care caracterizează fiecare habitat în parte. Analiza structurii calitative se va face din prisma compoziţiei floristice, a grupelor cenotice, a structurii în diferite categorii de bioforme, de geoelemente, categorii ecologice şi economice. Compoziţia floristică a habitatelor cuprinde două componente de bază şi anume bogăţia specifică şi apartenenţa speciilor la anumite grupe cenotice. a. Bogăţia specifică sau diversitatea floristică a habitatelor analizate oferă informaţii importante asupra gradului de homeostazie a sistemului, stabilitatea sistemelor biologice fiind dependente de complexitatea structurală şi funcţională a acestora (Cristea et al. 2004). b. Grupele cenotice reunesc toate speciile caracteristice unei asociaţii vegetale şi unităţilor ierarhic superioare acesteia cuprind speciile edificatoare, caracteristice, şi uneori unele dintre speciile indicatoare şi diferenţiale (Cristea et al. 2004). Cunoaşterea acestor grupe

16
este importantă atât pentru încadrarea unui fragment de vegetaţie într-un anumit tip de habitat, dar şi pentru identificarea posibilelor scenarii privind dinamica acestora sau în formularea recomandărilor practice de management. Bioformele reunesc categoriile de plante care deşi aparţin la unităţi taxonomice diferite, ca rezultat al evoluţiei convergente în condiţii de mediu aproximativ identice, au dobândit o serie de caractere şi adaptări morfologice, anatomice şi fiziologice asemănătoare, care le oferă avantaje competitive în lupta interspecifică şi în valorificarea optimă a condiţiilor staţionale (Cristea 1993, Cristea et al. 2004). Cunoaşterea structurii habitatelor în bioforme este importantă în monitorizarea acestora deoarece (Cristea et al. 2004): - prin păstrarea în genofondul categoriilor de bioforme a sensului selecţie de biotop, oferă informaţii paleoecologice asupra florei şi vegetaţiei dintr-o anumită regiune; - oferă informaţii privind echilibrul vegetaţiei cu ansamblul condiţiilor de mediu şi cu particularităţile staţionale; - sugerează direcţiile posibile de evoluţie; - oferă informaţii privind intensitatea acţiunii factorilor de mediu; - contribuie la elucidarea structurii sinuziale a fitocenozelor analizate. Geoelementele reprezintă “ categorii de specii vegetale, mai mult sau mai puţin
îndepărtate filogenetic, care în decursul procesului de speciaţie au ocupat aceaşi regiune
geografică (mai mult sau mai puţin extinsă), urmând apoi căi specifice de migraţie şi integrare
cenotică înspre desăvârşirea arealelor actuale” (Cristea 1993). Analiza structurii habitatelor în categorii de geoelemente furnizează informaţii privind macro- şi microclimatele în care se dezvoltă, oferă repere în reconstituirea căilor de migraţie a unor specii. De asemenea, geoelementele stau la baza regionării biogeografice şi a raionării floristice, constituind un criteiu important în diagnosticarea habitatelor. Categoriile ecologice. Structura floristică a covorului vegetal, al unei anumite zone, este rezultatul unui lung şi complex proces de adaptare, al diferitelor specii de plante, la condiţiile de mediu date, fapt ilustrat în structura şi dinamica populaţiilor speciilor respective. Adaptarea este efectul variaţiilor cu caracter de regim, prin acţiunea selecţiei naturale, la variaţia periodică a diferiţilor factori ecologici între anumite limite, care devin limite de toleranţă ale populaţiei sau speciei respective (Botnariuc et Vădineanu 1982). Cu toate acestea, fiecare fitoindivid are un optim de dezvoltare la anumite valori ale factorilor ecologici. Cunoscând preferinţele unei specii faţă de factorii ecologici, valorile acestora pot fi apreciate după dezvoltarea indivizilor şi după structura populaţiei respective (Cristea 1993, Cristea et al. 2004). Structura habitatelor în diferite categorii ecologice se va aborda prin prisma indicilor ecologici şi a comportamentului social al plantelor. Indicii ecologici reprezintă codificarea preferinţelor speciilor faţă de un anumit factor de mediu. O abordare nouă a structurii în categorii ecologice este cea introdusă de către Borhidi (1995), care propune o clasificare a plantelor în funcţie de comportamentul social al acestora. Prin comportament social înţelegând modul prin care planta este legată de habitat, informaţia conţinută şi naturaleţea legăturii, bazându-se pe rolul pe care fiecare specie îl joacă în cadrul comunităţii vegetale (Borhidi 1995). Tipurile de comportament social al plantelor sunt parte integrantă a modelului C-S-R de strategie a plantelor propus de către Grime (1979) (Borhidi 1995). Categoriile economice. Plantele, singura sursă de energie autoregenerabilă de pe Terra, au jucat un rol deosebit de important, datorită multiplelor întrebuinţări pe care le-au avut de-a

17
lungul evoluţiei societăţii umane. Diferenţierea regională a florei şi vegetatiei a imprimat caractere specifice economiilor locale. În acest context, a apărut necesitatea estimării potenţialului economic al comunităţilor vegetale şi a gradului lor de exploatare.
1.2.4.2.2. Structura spaţială
Habitatele, ca orice sistem biologic, prezintă o structură tridimensională, structură care ” favorizează utilizarea la maximum de randament a condiţiilor ecotopului...”(Cristea 1993), determinată de distribuţia diferită a speciilor în spaţiu. În mod obişnuit, în studiile de vegetaţie se va analiza atât structura orizontală şi verticală a fitocenozelor studiate cât şi distribuţia lor locală.
Structura orizontală a fitocenozelor reprezintă modul de repartizare, a diferitelor populaţii vegetale, pe suprafaţa ocupată de fragmentul de vegetaţie analizat, astfel, putându-se evidenţia atât particularităţile microstaţionale ale biotopului cât şi relaţiile interspecifice (Cristea, 1993).
Structura verticală sau stratificarea comunităţilor vegetale, supraterană şi subterană, se datorează înălţimii sau adâncimii diferite la care speciile îşi etalează organele vegetative în funcţie de variaţia pe verticală a valorilor factorilor ecologici. Stratificarea vegetaţiei, care „ …
reprezintă o expresie directă a complexităţii ei” (Borza et Boşcaiu 1965) depinde şi de compoziţia specifică a fitocenozei, compoziţia în categorii de bioforme, stadiile ontogenetice de dezvoltare a fitoindivizilor. Cristea (1993) defineşte stratul ca fiind totalitatea fitopopulaţiilor dintr-o comunitate vegetală, „ … a căror părţi active sunt situate între
aproximativ aceleaşi înălţimi deasupra solului sau între aceleaşi adâncimi în sol, ca o adaptare
la valorificarea optimă a spaţiului biotopic”. Analiza structurii verticale a habitatelor oferă informaţii despre principalele specii din
structura acestora, modul de agregare a fitoindivizilor, gradul de închegare a vegetaţiei, complexitatea morfo-structurală a habitatului, strategiile adoptate de specii pentru valorificarea optimă a resurselor şi posibilele tendinţe evolutive (Cristea et al. 2004).
Distribuţia locală a fitocenozelor, un alt aspect important în studiul comunităţilor vegetale şi implicit al habitatelor, este reprezentată de modul în care se repartizează în spaţiu diversele fitocenoze în funcţie de distribuţia locală a factorilor orografici, geologici, edafici şi microclimatici care determină mozaicarea vegetaţiei (Cristea et al. 2004).
1.2.4.3. Variabilele de mediu
Plantele, ca orice organisme vii, nu trăiesc izolat ci în strânsă interdependenţă atât cu alte organisme (plante sau animale), cât şi cu materia nevie, formând împreună mediul. Variabilele de mediu luate în studiu au fost împărţite în patru mari categorii şi anume: factori topografici, factori climatici, factori edafici şi factori biotici care includ şi componenta antropică. Factorii topografici (relieful) influenţează în mod indirect structura şi distribuţia fitocenozelor prin modificarea regimului climatic şi a factorilor edafici. Chiar dacă factorii

18
topografici nu sunt susceptibili de modificări majore, înregistrarea acestora este importantă în vederea descifrării ecologiei habitatelor şi elaborarea măsurilor de management activ necesare pentru menţinerea acestora într-o stare favorabilă pentru conservare.
Factorii climatici. Prin climat se înţelege acţiunea complexă a factorilor meteorologici (lumina, temperatura, umiditatea, precipitaţiile, vântul) pe o anumită suprafaţă a Terrei (Pop 1977, 1979). În cadrul acestui climat general pot fi deosebite subunităţi mai mici cum ar fi macroclimatul sau climatul regional, mezoclimatul sau climatul local (topoclimatul) şi microclimatul sau climatul microreliefului care include şi fitoclimatul (Cristea 1991). Din punctul de vedere al studiilor de vegetaţie, un rol important îl are analiza macro- şi microclimatului, care condiţionează în mod direct structura şi raspândirea fitocenozelor.
Factorii edafici, prin proprietăţile lor, sunt factori limitativi care condiţionează creşterea şi dezvoltarea plantelor, constituirea şi dezvoltarea comunităţilor vegetale fiind în strânsă legătură cu aceştia.
Factorii biotici, alături de factorii abiotici contribuie în mod direct sau indirect la constituirea, structurarea şi distribuţia spaţială a fitocenozelor. Dintre factorii biotici cei mai importanţi din punct de vedere al monitorizării habitatelor amintim acoperirea generală a vegetaţiei, care oferă informaţii privind gradul de perturbare a fitocenozelor şi existenţa unor nişe libere care pot fi ocupate de alte specii şi acoperirea realizată de briofite, modul de folosinţă al habitatelor analizate şi grosimea stratului de litieră.
1.2.5. Stocarea şi gestionarea datelor
Datele colectate în etapa de teren a programului de monitorizare, indiferent de suportul pe care au fost acestea înregistrate (hârtie sau suport magnetic), vor fi introduse într-o bază de date care să permită atât stocarea lor în siguranţă, cât şi interogarea şi extragerea acestora pentru analiză. Deşi înregistrarea datelor direct pe suport magnetic (PDA) este mult mai rapidă şi tentantă (copierea datelor într-o bază de date fiind mult mai simplă şi rapidă), trebuie avut în vedere şi riscul ca aceste dispozitive să se defecteze putând duce la pierderea irevesibilă a datelor, dovedindu-se până la urmă o metodă mult mai costisitoare. De aceea, se recomandă fie utilizarea unor fişe standard pentru înregistrarea datelor pe teren, fie realizarea periodică a unor copii de siguranţă a datelor colectate. Deşi nu există o structură standard a bazelor de date, aceasta rămânând la latitudinea fiecărui coordonator de program de monitorizare, totuşi, se recomandă ca acestea să îndeplinească câteva criterii: - să fie simple, clare, intuitive şi uşor de utilizat; - să conţină câmpuri pentru fiecare dintre variabilele măsurate; - să conţină informaţii complete despre localizarea fiecărei suprafeţe de probă, tipul de eşantionaj, orice alte informaţii care pot influenţa interpretarea datelor; - pentru datele estimate pe baza unor scări valorice, trebuie să fie precizată clar metoda folosită; - trebuie menţionate metodele prin care au fost obţinute datele şi unitatea lor de măsură acolo unde este cazul (concentraţia cationilor din sol, grosimea stratului de litieră etc.);

19
Deoarece colectarea datelor presupune o muncă laborioasă fiind constisitoare şi greu de repetat, achiziţionarea unor computere performante, deşi mai scumpe, trebuie privită ca o investiţie pe termen lung. Pentru protejarea datelor se vor realiza, periodic, copii de siguranţă a tuturor datelor obţinute, copii care se vor păstra pe alte computere decât cele pe care lucrează. De asemenea, fişele de teren, hărţile, înregistrările originale vor fi arhivate şi păstrate în siguranţă, putându-se dovedi deosebit de valoroase pe viitor. Tot pentru o mai bună protecţie a datelor, accesul cu drepturi depline (editare şi modificare date) la bazele de date trebuie limitat la un număr cât mai mic de utilizatori, ceilalţi având acces la date, fără a le putea însă modifica (Hill et al. 2005).
1.2.5. Analiza şi interpretarea rezultatelor
Alegerea metodelor de analiză a datelor colectate şi interpretare a rezultatelor trebuie să ţină cont, în primul, de natura acestora. Variabilele înregistrate, pe teren prin estimări şi măsurători directe, sau obţinute în laborator prin analize specifice pot fi clasificate în (Zar 1999) :
- variabile de interval – sunt variabilele a căror scală de măsurare are intervale de mărime constantă, dar nu au o valoare zero adevărată;
- variabile de raport – sunt variabile de interval dar care au o valoare zero adevărată, numită valoare de raport;
- variabile ordinale – sunt variabilele a căror scală de măsurare este ordonată dar nu are intervale definite;
- variabile nominale – sunt variabile calitative care pot lua un număr fiit de valori neordonate;
În funcţie de valorile pe care le pot lua, variabilele mai pot fi clasificate în (Witmer 1999, Zar 1999):
- variabile discrete – valorile pe care le pot lua sunt numere întregi (ex. numărul de flori al unui fitoindivid);
- variabile continue - valorile lor pot lua orice număr (ex. concentraţia cationilor din sol).
Variabilele de interval, de raport şi cele ordinale pot fi continue sau discrete în timp ce variabilele nominale nu pot fi decât discrete (Zar 1999).
Pentru analiza datelor colecate în cadrul programelor de monitorizare se recomandă parcurgerea următoarelor etape:
a. Transformarea şi codificarea datelor. Deoarece majoritatea metodelor statistice sunt aplicabile doar pentru datele cu o distribuţie normală, iar datele ecologice nu respectă această distribuţie este nevoie tranformarea acestora. În funcţie de natura variabilelor măsurate, acestea pot fi transformate prin logaritmare sau cu ajutorul funcţiei arcsin. În acelaşi timp, datorită limitărilor impuse de majoritatea programelor de analiză statistică, denumirile variabilelor analizate vor fi codificate astfel încât lungimea lor să nu depăşească 8 caractere.
b. Eliminarea variabilelor outliers. Datorită erorilor de eşantionare, sau datorită altor factori (perturbări puternice, heterogenitatea comunităţilor vegetale etc), unele variabile au valori mult îndepărtate faţă de restul valorilor înregistrate. Aceste valori extreme trebuie

20
eliminate din setul de date care vor fi analizete deoarece majoritatea prelucrărilor statistice sunt influenţate în mod negativ de prezenţa valorilor outliers (Gauch Jr 1982).
c. Analiza datelor. În alegerea metodelor de analiză a datelor ecologice trebuie să se ţină cont de cîteva condiţii pe care acestea trebuie să le întrunească (Gauch Jr 1982), dintre care amintim următoarele:
- să fie precise în reprezentarea structurii datelor analizate; - să fie eficiente; - să fie robuste în sensul de a nu fi influenţate de variabilitatea proprietăţilor diferitelor
date; - să fie obiective, cu excepţia cazurilor în care se acceptă sau este necesară (din diverse
motive) o anumită doză de subiectivitate. De asemenea, metodele de prelucrare a datelor, în special cele statistice, trebuie să
poată fi utilizate cu tipul de date avute la dispoziţie. De exemplu, dacă dorim să analiză răspunsul unei variabile dependente faţă de variaţia mai multor variabile independente acest lucru se va face prin regresie. Dacă atât variabila dependentă cât şi cele independente sunt variabile cantitative continue, se va alege regresia multiplă, dar, dacă unele, sau toate variabilele independente sunt nominale se va alege ca metodă de analiză regresia logistică. Metodele de analiză a datelor pot să fie grupate în metode statistice de bază şi metode statistice complexe sau multivariate. Metodele multivariate de analiză contribuie în mod special la elaborarea ipotezelor de lucru, în timp ce metodele statistice clasice sunt extrem de utile în testarea acestor ipoteze. d. Interpretarea rezultatelor se va face ţinând cont de natura datelor, metodele de analiză folosite pentru prelucrarea lor şi semnificaţia ecologică a acestora.
1.2.6. Elaborarea raportului de monitorizare
Pentru elaborarea raportului de monitorizare trebuie să se ia în calcul mesajul principal pe care vrea să-l transmită şi cui i se adresează (Hill et al. 2005). Publicul ţintă căruia i se va adresa raportul va fi cel care va decide nivelul de detaliere tehnică a problematicilor expuse şi nivelul ştiinţific al limbajului utilizat. Indiferent de gradul de detaliere, raportul de monitorizare trebuie să cuprindă următoarele aspecte:
- importanţa realizării studiului de monitorizare; - prezentarea obiectivelor propuse; - prezentarea generală a gradului de cunoaştere a problematicii studiate; - prezentarea metodelor de lucru utilizate pentru colectarea şi analiza datelor
precum şi motivarea alegeri acestor metode de lucru; - prezentarea rezultatelor obţinute; - discutarea rezultatelor şi a implicaţiilor teoretice şi practice ale acestora; - concluziile privind importanţa studiului şi elaborarea de recomandări pentru viitor; - sursele bibliografice care au fost consultate.

21
Se va acorda mare atenţie modului de editare a informaţiilor prezentate sub formă de tabele sau grafice, acestea trebuind să fie cât mai clare şi complete (evitarea aglomerării graficului prin utilizarea unor adnotări care nu sunt necesare pentru interpretarea acestuia, includerea legendei cu explicarea simbolurilor utilizate, explicarea prescurtărilor folosite în grafice sau tabele etc.).

22
II. STAREA FAVORABILĂ PENTRU CONSERVAREA HABITATULUI 6230*
2.1. Descrierea habitatului
În accepţiunea manualului european (EUR27) dar şi al celui românesc de interpretare al
habitatelor Natura 2000 (Gafta et Owen 2008), habitatul 6230* (pajişti de Nardus bogate în specii pe substraturi silicatice din zone montane şi submontane în Europa continentală) este reprezentat de pajişti permanente secundare de Nardus stricta, ale căror fitocenoze vegetează în diferite condiţii staţionale şi care au o plasticitate ecologică mare, ceea ce imprimă habitatului o mare heterogenitate structurală. În acest tip de habitat au fost încadrate doar pajiştile în care abundenţa-dominanţa speciei Nardus stricta este de cel putin 2 pe scara Braun-Blanquet (metoda fitosociologică) ceea ce corespunde unei acoperiri medii minime de 17,5%. Au fost excluse din acest habitat pajiştile puternic perturbate ca urmare a supraexploatării lor, în special prin păşunat excesiv, fapt ce a dus la o puternică degradare a acestora.
Din punct de vedere fitosociologic, acestui habitat îi corespund următorii cenotaxoni: Scorzonero roseae-Festucetum nigricantis (Puşcaru et al. 1956) Coldea 1978 (syn.: Festucetum
rubrae fallax Puşcaru et al. 1956, Festucetum rubrae montanum Csűrös et Resmeriţă 1960); Violo declinatae-Nardetum Simon 1966 (syn.: Nardetum strictae montanum Resmeriţă et Csűrös 1963, Nardetum strictae alpinum Buia et al. 1962, Nardetum alpigenum carpaticum
Borza 1959); Nardo-Festucetum tenuifoliae Buiculescu 1971; Festuco rubrae-Agrostietum
capillaris Horvat 1951 subas. nardetosum strictae Pop 1976, Poetum medie-Nardetum strictae Resmeriţă 1987.
Dintre speciile caracteristice acestui habitat putem enumera: Agrostis capillaris, Festuca
rubra, Nardus stricta, Deschampsia flexuosa, Potentilla erecta, Viola declinata, Antennaria
dioica, Danthonia decumbens, Genista tinctoria ssp. tinctoria, Veronica officinalis, Polygala
vulgaris, Cerastium fontanum ssp. vulgare, Hieracium pilosella, Viola canina, Plantago
lanceolata, Luzula campestris, Carex pallescens, Campanula serrata, Scabiosa lucida, Carlina
vulgaris ssp. vulgaris, Luzula luzuloides ssp. cuprina, Gentiana acaulis, Festuca tenuifolia, Cerastium fontanum ssp. triviale, Alchemilla glaucescens, Trifolium dubium, Potentilla aurea ssp. chrysocraspeda, Festuca airoides, Festuca nigrescens, Plantago atrata, Ranunculus
serpens, Luzula multiflora, Geum montanum etc. Fitocenozele acestui habitat prezintă o plasticitate ecologică relativ mare, fapt
demonstrat şi de ecartul altitudinal, acestea putând fi găsite la altitudini cuprinse între 500 şi 2300 m.s.m., ocupând staţiuni xero-mezofile sau mezofile cu soluri oligotrofe acide. Pajiştile încadrate în acest tip de habitat se găsesc pe platouri sau versanţi cu pante de până la 50-54o fără a fi puternic condiţionate de expoziţia staţiunilor pe care le ocupă.
Din punct de vedere sindinamic, nardetele din etajul montan (Violo declinatae-
Nardetum strictae) s-au dezvoltat din pajiştile edificate de păiuşul roşu (Festuca rubra, F.

23
nigrescens) şi iarba câmpului (Agrostis capillaris), pe care le-a invadat ca urmare a suprapăşunării acestora. Suprapăşunatul a dus la acidifierea solului, tasarea lui şi la eliminarea speciilor bune furajere, al căror loc a fost luat de către ţepoşică (părul porcului) (Pop et al. 2002). Lipsa lucrărilor curente de întreţinere şi fertilizare, precum şi păşunatul abuziv, neraţional cu oile, au creat în timp condiţii nefavorabile pentru creşterea şi dezvoltarea plantelor autotrofe, valoroase şi condiţii optime pentru instalarea şi extinderea speciei Nardus
stricta. De asemenea, nardetele, prin târlire, în funcţie de intensitatea ei, vor evolua fie spre pajişti edificate de Agrostis capillaris, Festuca rubra şi Festuca nigrescens, prin reducerea abundenţei ţepoşicii, fie spre pajişti nitrofile edificate de Rumex alpinus şi Urtica dioica prin eliminarea completă a lui Nardus stricta, urmând ca în decurs de câţiva ani ţepoşica să se reinstaleze în staţiunile târlite. Prin modificarea regimului hidric al solului pajiştile de ţepoşică pot evolua spre pajişti edificate de Molinia caerulea sau specii de Juncus.
Variaţia puternică a valorilor unor factori ecologici pot duce la modificări structural majore care determină dinamica covorului vegetal şi al habitatelor ducând la înlocuirea unor tipuri de habitate cu altele.
În cele aproximativ 600 de relevee analizate în cadrul proiectului LIFE05 NAT/RO/000176 (din literatură şi efectuate pe teren în cadrul proiectului), au fost identificaţi 390 de taxoni vegetali. Importanţa sozologică a habitatului 6230* se datorează faptului că în structura pajiştilor de Nardus stricta, se regăsesc 40 de taxoni vulnerabili, rari sau periclitaţi (tab. 3 dintre cei înscrişi în listele roşii naţionale iar dintre aceştia doi taxoni sunt rari la nivel european (Dianthus glacialis s. gelidus şi Anthemis macrantha). Astfel, speciile vulnerabile, rare sau periclitate reprezintă peste 10% din totalul taxonilor identificaţi în structura acestui tip de habitat la nivel naţional.
Tabel 3. Lista taxonilor din listele roşii naţionale şi europene care se regasesc în structura habitatului 6230*
(LRN – liste roşii naţionale, LRE – liste roşii europene, V-vulnerabil, R-rar, NT-ameninţat)
LRN LRE Specia
V Arnica montana L.
V Rhododendron myrtifolium Schott & Kotschy
V Thymus serpyllum L.
R Scorzonera purpurea L. s. rosea (Waldst. & Kit.) Nyman
R Melampyrum saxosum Baumg.
R Gymnadenia conopsea (L.) R.Br.
R Scorzonera humilis L.
R Phyteuma spicatum L.
R Dactylorhiza maculata (L.) Soó
R Coeloglossum viride (L.) Hartm.
R Listera ovata (L.) R.Br.
R Platanthera bifolia (L.) Rich.
R Veronica alpina L.
R Gentiana acaulis L.

24
LRN LRE Specia
R Festuca tenuifolia Sibth.
R Cerastium transsilvanicum Schur
R Sagina saginoides (L.) H.Karst.
R Centaurea uniflora Turra s. nervosa (Willd.) Bonnier&Lay
R Phyteuma confusum A.Kern.
R Plantago gentianoides Sibth. & Sm.
R Vaccinium uliginosum L. s. microphyllum Lange
R Dianthus barbatus L. s. compactus (Kit.) Heuff.
R R Dianthus glacialis Haenke s. gelidus (Schott; Nyman&K)Tutin
R Silene nutans L. s. dubia (Herbich) Zapal.
R Plantago atrata Hoppe
R Leucanthemopsis alpina (L.) Heywood s. alpina
R Viola dacica Borbás
R Diphasiastrum alpinum (L.) Holub
R Galium pumilum Murray
R R Anthemis macrantha Heuff.
R Crepis conyzifolia (Gouan) A.Kern.
R Pinguicula vulgaris L.
R Carex brunnescens (Pers.) Poir.
R Phyteuma vagneri A.Kern.
R Sesleria coerulans Friv.
R Ranunculus crenatus Waldst. & Kit.
R Gymnadenia odoratissima (L.) Rich.
NT Cirsium furiens Griseb. & Schenk
NT Galanthus nivalis L.
NT Thymus comosus Heuff. ex Griseb.
Prezenţa în număr mare a speciilor cu valoare sozologică mare constituie un argument în plus în favoarea adoptării unor măsuri specific de conservare a acestui tip de habitat.
2.2. Starea favorabilă pentru conservare
Directiva Habitate cuprinde o serie de cerinţe pentru Statele Membre cu privire la
implementarea măsurilor de conservare pentru habitatele şi speciile de interes Comunitar. Obiectivul general al acestor măsuri ar fi atingerea scopului general al acestei Directive, menţionat în Articolul 2(1) “de a contribui la asigurarea biodiversităţii prin conservarea
habitatelor naturale precum şi a faunei şi florei sălbatice pe teritoriul european al Statelor
Membre la care Tratatul se aplică”. Articolul 2(2) menţionează “Măsurile luate în baza
prezentei Directive vizează menţinerea sau restabilirea, într-o stare favorabilă de conservare, a
habitatelor naturale şi a speciilor din fauna şi flora sălbatică de interes comunitar”, iar la

25
punctul 3 al aceluiaşi articol se arată că “Măsurile luate în baza prezentei Directive ţin seama de
exigenţele economice, sociale şi culturale ca şi de particularităţile regionale şi locale.”
Articolul 1(e) al Directivei Habiate defineşte starea favorabilă de conservare ca fiind “starea de conservare a unui habitat natural înseamnă suma influenţelor ce acţionează asupra
unui habitat natural şi a speciilor tipice pe care le adăposteşte, care pot afecta pe termen lung
repartiţia sa naturală, structura şi funcţiile sale, ca şi supravieţuirea pe termen lung a speciilor
sale tipice, pe teritoriul vizat în Articolul 2”. În conformitate cu Directiva Habitate, Articolul 1(e), un habitat va fi în stare favorabilă de conservare atunci când:
- aria sa de răspândire naturală ca şi suprafeţele pe care le acoperă în cadrul acestei arii sunt stabilite sau în extindere;
- există structura şi funcţiile specifice necesare pentru menţinere pe termen lung şi este posibil să existe în viitorul previzibil;
- stadiul de conservare a speciilor care îi sunt tipice este favorabil conform punctului (i) (datele relative la dinamica populaţiei speciei în cauză arată că această specie continuă si este posibil să continue, pe termen lung, să fie o componentă viabilă a habitatului său natura; aria de repartiţie naturală a speciei nu se reduce şi nu riscă să se reducă într-un viitor previzibil şi există şi probabil va exista un habitat destul de întins pentru ca populaţiile sale să se menţină pe termen lung). Evaluarea stării favorabile de conservare se va face pe baza unor atribute care descriu
(direct sau indirect) starea acestuia şi care trebuie să îndeplinească căteva cerinţe minime, şi anume:
- să fie măsurabile, astfel încât să se poate cuantifica şi monitoriza limitele acestuia; - să fie descrie starea caracteristicii şi nu factorii care o influenţează.
Limitele acceptabile între care pot varia valorile unui atribut astfel încât habitatul să fie într-o stare favorabilă de conservare nu trebuie să reprezinte nivelul optim sau ţintă ci să ia în considerare o variaţie acceptabilă.
Pe baza atributelor care definesc starea favorabilă de conservare şi a limitelor între care pot varia valorile acestora se va stabili atât un meniu de măsuri de management cât şi un plan de monitorizare.
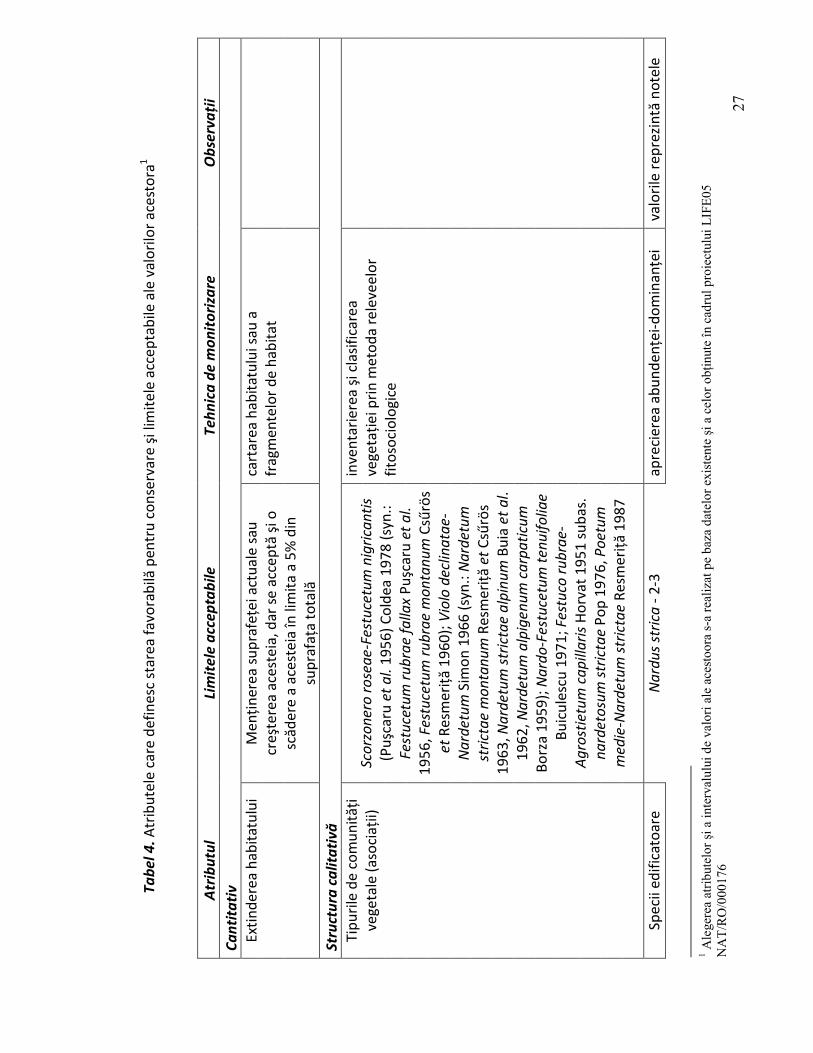
2.2.1. Atribute cantitative
Extinderea habitatului. Suprafaţa habitatului sau a fragmentelor de habitat analizate trebuie să se menţină în limitele actuale sau să crească. Analiza suprafeţei ocupată de habitat într-un sit se va face prin cartarea limitelor fiecărui tip de comunitate vegetală (asociaţie). În funcţie de suprafaţa ocupată şi de particularităţile structurale şi funcţionale ale fragmentelor de habitat, se poate accepta în unele cazuri şi o descreştere a suprafeţei cu maximum 5%. Chiar dacă această descreştere nu pune în pericol existenţa şi conservarea habitatului, totuşi indică prezenţa unor factori perturbatori care, dacă nu se iau măsuri de eliminare sau contracarare a efectelor lor, în timp pot să ducă ladispariţia habitatului. De aceea, se recomandă, ca în cazul unor descreşteri ale suprafeţelor ocupate de fragmentele de habitat monitorizate, să se

26
realizeze studii mai aprofundate pentru identificarea factorilor care determină aceste perturbări.
27
Ta
bel
4. A
trib
ute
le c
are
def
ines
c st
area
fav
ora
bilă
pen
tru
co
nse
rvar
e şi
lim
itel
e ac
cep
tab
ile a
le v
alo
rilo
r ac
esto
ra1
Atr
ibu
tul
Lim
itel
e a
ccep
tab
ile
Teh
nic
a d
e m
on
ito
riza
re
Ob
serv
aţi
i
Ca
nti
tati
v
Exti
nd
erea
hab
itat
ulu
i M
enţi
ner
ea s
up
rafe
ţei a
ctu
ale
sau
cr
eşte
rea
aces
teia
, dar
se
acce
ptă
şi o
sc
ăder
e a
aces
teia
în li
mit
a a
5%
din
su
pra
faţa
to
tală
cart
area
hab
itat
ulu
i sau
a
frag
men
telo
r d
e h
abit
at
Stru
ctu
ra c
alit
ati
vă
Tip
uri
le d
e co
mu
nit
ăţi
vege
tale
(as
oci
aţii)
Sco
rzo
ne
ro r
ose
ae
-Fe
stu
cetu
m n
igri
can
tis
(Pu
şcar
u e
t a
l. 1
95
6)
Co
ldea
19
78
(syn
.:
Fe
stu
cetu
m r
ub
rae
fa
lla
x P
uşc
aru
et
al.
1
95
6, F
est
uce
tum
ru
bra
e m
on
tan
um
Csű
rös
et
Res
mer
iţă
19
60);
Vio
lo d
ecl
ina
tae
-
Na
rde
tum
Sim
on
19
66 (
syn
.: N
ard
etu
m
stri
cta
e m
on
tan
um
Res
mer
iţă
et
Csű
rös
19
63
, Na
rde
tum
str
icta
e a
lpin
um
Bu
ia e
t a
l. 1
96
2, N
ard
etu
m a
lpig
en
um
ca
rpa
ticu
m
Bo
rza
195
9);
Na
rdo
-Fe
stu
cetu
m t
en
uif
oli
ae
B
uic
ule
scu
19
71
; Fe
stu
co r
ub
rae
-
Ag
rost
ietu
m c
ap
illa
ris
Ho
rvat
19
51
sub
as.
na
rde
tosu
m s
tric
tae
Po
p 1
97
6, P
oe
tum
me
die
-Na
rde
tum
str
icta
e R
esm
eriţ
ă 1
987
inve
nta
rier
ea ş
i cla
sifi
care
a ve
geta
ţiei
pri
n m
eto
da
rele
veel
or
fito
soci
olo
gice
Spec
ii ed
ific
ato
are
Na
rdu
s st
rica
- 2
-3
apre
cier
ea a
bu
nd
enţe
i-d
om
inan
ţei
valo
rile
rep
rezi
ntă
no
tele
1 Alegerea atributelor şi a intervalului de valori ale acestoora s-a realizat pe baza datelor existente şi a celor obŃinute în cadrul proiectului LIFE05
NAT/RO/000176
28
Atr
ibu
tul
Lim
itel
e a
ccep
tab
ile
Teh
nic
a d
e m
on
ito
riza
re
Ob
serv
aţi
i
Ag
rost
is c
ap
illa
ris
- 1
-4
spec
iilo
r p
e sc
ara
Bra
un
-Bla
nq
uet
cu
aj
uto
rul r
amei
met
rice
p
e sc
ara
Bra
un
-Bla
qu
et
Fe
stu
ca r
ub
ra -
2-4
Fe
stu
ca n
igre
sce
ns
- 2-
4
Fe
stu
ca t
en
uif
oli
a -
2-4
Po
a m
ed
ia -
2-3
Spec
ii ca
ract
eris
tice
M
inim
um
10
sp
ecii/
rele
veu
din
tre
urm
ăto
arel
e:
Sco
rzo
ne
ra p
urp
ure
a s
sp r
ose
a,
Ca
mp
an
ula
serr
ata
, V
iola
de
clin
ata
, V
iola
da
cica
, H
iera
ciu
m
pil
ose
lla
, A
nte
nn
ari
a d
ioic
a,
Da
nth
on
ia
de
cum
be
ns,
Ve
ron
ica
off
icin
ali
s, P
ote
nti
lla
ere
cta
, A
chil
lea
str
icta
, C
are
x p
all
esc
en
s, L
uzu
la
sud
eti
ca,
Po
ten
till
a a
ure
a s
sp.
au
rea
, A
rnic
a
mo
nta
na
, G
ymn
ad
en
ia c
on
op
sea
,
Co
elo
glo
ssu
m v
irid
e,
Po
lyg
ala
vu
lga
ris,
Ge
nti
an
a p
ne
um
on
an
the
, G
en
ista
tin
cto
ria
,
Ca
mp
an
ula
pa
tula
ssp
ab
ieti
na
, Po
ten
till
a
au
rea
ssp
. Ch
ryso
cra
spe
da
, Fe
stu
ca a
iro
ide
s
etc
.
rele
veu
l fit
oso
cio
logi
c, s
tud
ii p
op
ula
ţio
nal
e
Spec
ii in
vazi
ve/c
olo
nia
liste
De
sch
am
psi
a c
esp
ito
sa,
Cir
siu
m v
ulg
are
,
Ca
rdu
us
aca
nth
oid
es,
Po
lyg
on
um
avic
ula
re,
Ru
me
x a
lpin
us,
Ca
pse
lla
bu
rsa
-pa
sto
ris,
Pte
rid
ium
aq
uil
inu
m,
Ta
raxa
cum
off
icin
ale
,
Cra
tae
gu
s m
on
og
yn
a,
Va
ccin
ium
myrt
illu
s, V
.
vit
is-i
da
ea
etc
. şi s
ă re
aliz
eze
o a
cop
erir
e d
e su
b
5%
din
su
pra
faţa
to
tală
rele
veu
l fit
oso
cio
logi
c, s
tud
ii p
op
ula
ţio
nal
e
Bo
găţi
a sp
ecif
ică
> 2
5 sp
ecii/
10
0 m
2 re
leve
ul f
ito
soci
olo
gic
Dis
trib
uţi
a ab
un
den
ţelo
r re
lati
ve a
sp
eciil
or
Log-
no
rmal
ă, Z
ipf,
Zip
f-M
and
elb
rot
anal
iza
dis
trib
uţi
ei a
bu
nd
enţe
lor
rela
tive
pri
n r
ealiz
area
cu
rbel
or
ran
g-ab
un
den
ţă

29
Atr
ibu
tul
Lim
itel
e a
ccep
tab
ile
Teh
nic
a d
e m
on
ito
riza
re
Ob
serv
aţi
i
Stru
ctu
ra s
pa
ţia
lă
Înăl
ţim
ea m
edie
a
vege
taţi
ei
25
-45
cm
m
ăsu
răto
ri p
e te
ren
Stra
tifi
care
a ve
geta
ţiei
>
3-4
str
atu
ri
met
od
a p
rofi
lelo
r d
e ve
geta
ţie
Sup
rafa
ţă f
ără
vege
taţi
e să
rep
rezi
nte
max
imu
m 5
% d
in s
up
rafa
ţa
tota
lă a
hab
itat
ulu
i, d
ar s
ă n
u e
xist
e su
pra
feţe
mai
mar
i de
100
cm
2
met
od
a p
rofi
lelo
r d
e ve
geta
ţie
Gro
sim
ea m
edie
a
stra
tulu
i de
litie
ră
5-3
5 m
m
măs
ură
tori
pe
tere
n, m
inim
um
5
pen
tru
fie
care
rel
eveu
Ca
ract
eris
tici
ed
afi
ce
Rea
cţia
so
lulu
i (p
H)
4,5
-6
det
erm
inar
e d
irec
tă p
e te
ren
sau
în
lab
ora
tor
Hu
mu
s <
15
%
det
erm
inar
e în
lab
ora
tor
Co
nce
ntr
aţia
fo
sfo
rulu
i so
lub
il >
0,2
mg/
100
g s
ol
det
erm
inar
e în
lab
ora
tor
dat
ori
tă li
pse
i de
info
rmaţ
ii va
lide
nu
a f
ost
p
osi
bilă
fix
area
lim
itel
or
sup
erio
are
pen
tru
co
nce
traţ
ia e
lem
ente
lor
anal
izat
e as
tfel
încâ
t,
pen
tru
fie
care
sit
e în
par
te
se v
or
det
erm
ina
aces
te
limit
e
Co
nce
ntr
aţia
azo
tulu
i am
on
iaca
l >
1,9
5 m
g/1
00
g so
l d
eter
min
are
în la
bo
rato
r
Co
nce
ntr
aţia
ion
ilor
de
po
tasi
u
> 2
5 m
g/ 1
00
g so
l d
eter
min
are
în la
bo
rato
r
Co
nce
ntr
aţia
ion
ilor
de
calc
iu
> 1
0 m
g/10
0 g
so
l d
eter
min
are
în la
bo
rato
r

30
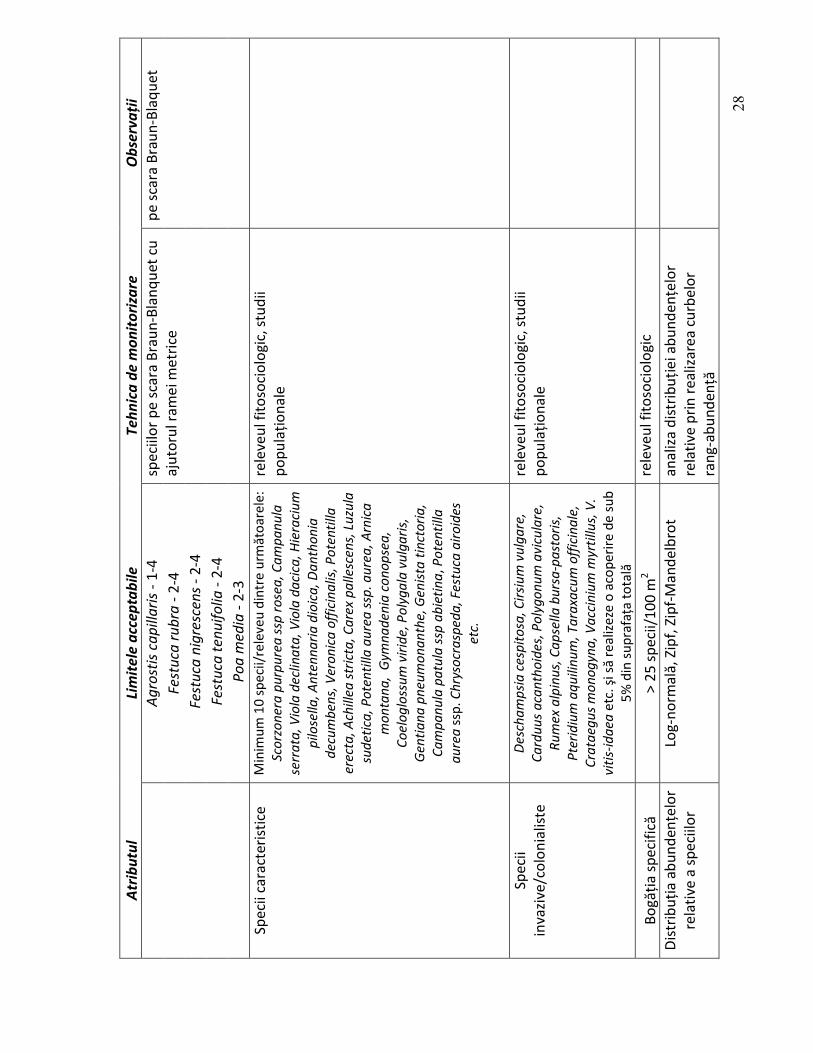
2.2.2. Structura calitativă
În alegerea atributelor structurale s-a pornit de la însăşi definiţia habitatului, definiţie
care subliniază două caracteristici foarte importante şi anume: - să fie pajişti de Nardus stricta, ceea ce presupune ca ţepoşica să fie o specie
edificatoare (dominantă sau codominantă); - să fie pajişti bogate în specii.
Tipurile de comunităţi vegetale (asociaţii). Deoarece nu toate fitocenozele în care
Nardus stricta este specie edificatoare aparţin acestui tip de habitat, este importantă încadrarea corectă a fitocenozelor analizate în asociaţiile vegetale care aparţin habitatului 6230*. Fiecare dintre asociaţiile vegetale enumerate în descrierea habitatului de pajişti de Nardus stricta bogate în specii pe soluri silicioase, are nişte caracteristici structurale, funcţionale şi dinamice care le individualizează şi valorizează din punct de vedere conservativ.
Speciile edificatoare sunt speciile care realizează cea mai mare cantitate de fitomasă şi a căror influenţă asupra celorlalte specii se manifestă diferit în funcţie de stadiul lor de dezvoltare (Grime 2001). Fără a minimaliza rolul celorlaltor specii în cadrul fiotcenozelor, speciile edificatoare sunt cele care determină în mare măsură structura, stabilitatea, funcţiile şi dinamica acestora.
Speciile caracteristice sunt speciile care au o răspândire limitată aproape exculsiv la unităţile cenotaxonomice ierarhic superioare asociaţiilor vegetale care intră în structura habitatului. Prezenţa sau absenţa lor reflectă „starea de sănătate” a habitatului.
Speciile invazive/colonialiste sunt specii care, ca şi precedentele, oferă informaţii preţioase despre influenţa şi intensitatea factorilor ecologici în general şi a celor antropo-zoogeni în special. Numărul acestora alături de abundenţa lor şi de modul de distribuţie în cadrul fitocenozelor reflectă gradul de degradare al habitatului.
Bogăţia specifică. Diversitatea biologică poate fi definită ca varietatea şi abundenţa speciilor într-o anumită unitate de studiu şi poate fi împărţită în două componente şi anume: bogăţia specifică şi echitabilitatea (Magurran 2004). Bogăţia specifică reprezintă numărul de specii, sau densitatea speciilor pe unitatea de suprafaţă, în timp ce echitabilitatea oferă o măsură a cât de asemănătoare sunt speciile din punct de vedere al abundenţei lor.
Bogăţia specifică a fragmentelor de vegetaţie analizate oferă informaţii preţioase asupra asupra gradului de homeostazie a fitocenozelor, particularităţilor ecotopului (inclusive presiunea antropo-zoogenă), stadiului de evoluţie al fitocenozelor şi chiar asupra corectitudinii cu care fitocenozele au fost analizate (Cristea 1993). Numărul speciilor variază în funcţie de o serie de factori cum ar fi: latitudinea şi altitudinea zonei în care se efectuează studiile, gradul de antropizare, stadiu de evoluţie al fitocenozei, tipul de vegetaţie etc.
Curbele rang-abundenţă reprezintă o modalitate grafică de sinteză a diversităţii specifice şi echitabilităţii prezentând în spaţiul semi-logaritmic distribuţia acoperirii relative (frecvenţei) a speciilor dintr-o fitocenoză, în funcţie de rangul acestora (Cristea et al. 2004). Curbele rang frecvenţă pot prezenta câteva modele în funcţie de caracteristicele ecologice ale fitocenozelor analizate. Acestea se pot încadra în cinci modele de bază: seria geometrică, modelul “bastonului rupt”, modelul log-normal, modelul Zipf şi modelul Zipf-Mandelbrot

31
(Wilson 1991). Fiecare dintre aceste modele are o anumită semnificaţie ecologică. Astfel, seria geometrică corespunde teoriei nişei pre-ocupate şi este caracteristică fitocenozelor pioniere, modelul “bastonului rupt” reflectă ocuparea aleatoare a nişelor, iar modelul log-normal este caracteristic fitocenozelor din staţiuni fertile dar şi în pajişti secundare (Cristea et al. 2004). Modelele Zipf şi Zipf-Mandelbrot, utilizate mai frecvent în lingvistică (Aoki 1995) au pătruns relativ recent în studiile de ecologie (Mouillot et Lepretre 2000). Din punct de vedere al interpretării ecologice a acestor modele, în cazul comunităţilor vegetale, prezenţa unei specii este foarte probabilă doar atunci când o serie de condiţii sunt îndeplinite (Wilson 1991). Dacă modelul log-normal poate fi privit ca rezultatul acţiunii simultane a mai multor factori, modelul Zipf şi Zipf-Mandelbrot poate fi privit ca rezultatul acţiunii secvenţiale a mai multor factori (Wilson 1991). Din punct de vedere al interpretării ecologice a curbelor rang-frecvenţă, succesiunea poate fi privită ca un progres de la seria geometrică spre modelul Zipf şi Zipf-Mandelbrot şi reîntoarcerea la seria geometrică în cazul senescenţei comunităţilor vegetale (Aoki 1995).
Analiza curbelor rang-frecvenţă permite compararea datelor colectate în perioade de timp diferite, oferind informaţii importante privind procesele dinamice care au loc la nivelul habitatului.
2.2.3. Structura spaţială
Înălţimea medie a vegetaţiei reprezintă un factor important care influenţează în mod direct şi indirect structura şi dinamica fitocenozelor. Înălţimea vegetaţiei influenţează în mod direct distanţa de dispersie a seminţelor (Soons et al. 2004), iar indirect, prin reglarea cantităţii de lumină care ajunge pe sol, germinaţia seminţelor (Kelly 1989) şi procesul de înfrăţire al gramineelor (Grant et al. 1996). Stratificarea vegetaţiei este considerată ca fiind „... o expresie directă a complexităţii
vegetaţiei” (Borza et Boşcaiu 1965). Stratificarea asigură o utilizare optimă a resurselor influenţând în acest mod atât compoziţia specifică cât şi funcţia de producţie şi mediogenă, stabilitatea şi dinamica fitocenozelor. Cu cât o fitocenoză este constituită din mai multe straturi, cu atât este mai stabilă. Suprafaţa de sol fără vegetaţie reprezintă o măsură indirectă a intensităţii acţiunii factorilor perturbatori. Suprafeţele mici de teren nud pot să aibă un efect benefic asupra fitodiversităţii habitatului prin faptul că permite pătrunderea şi stabilirea unor specii de plante. Grosimea stratului de litieră. Litiera, este un factor de mediu cu un rol foarte complex, având o importanţă deosebită în determinarea structurii şi dinamicii fitocenozelor. În funcţie de modul în care influenţează plantele şi comunităţile vegetale, efectele litierei pot fi: - directe – constituind o barieră fizică pentru creşterea şi dezvoltarea plantulelor (Hector et al. 2000), pentru disiparea vaporilor de apă din sol (McAlpine et Drake 2003), împiedică contactul direct dintre seminţe şi sol (Rotundo et Aguiar 2005), împiedică eroziunea solului datorată apelor pluviale şi protejează solul împotriva îngheţului (Walsh et Voigt 1977), oferă protecţie seminţelor şi plantulelor tinere împotriva granivorelor şi ierbivorelor (Quested et Eriksson 2006);

32
- indirecte – determinând schimbări microclimatice care afectează temperatura şi umiditatea solului dar şi cantitatea şi calitatea radiaţiei solare (Facelli et Carson 1991, Facelli et Pickett 1991, Bosy et Reader 1995, Nakamura 1996, Eckstein et Donath 2005), determinând o scădere a variaţiilor temperaturii (Bosy et Reader 1995), ameliorează condiţiile de umiditate a solului, mai ales în perioadele de stres hidric (Maret et Wilson 2005) şi reduce gradul de iradiere al solului cu până la 99% (Bosy et Reader 1995). Proprietăţile de infiltrare, structura şi textura, cel puţin a orizontului superior al solului sunt determinate, parţial, de cantitatea şi calitatea litierei (Walsh et Voigt 1977). De asemenea, litiera are un rol important în circuitul biogeochimic al substanţelor nutritive (Walsh et Voigt 1977, Hector et al. 2000, Aerts et al. 2003, Eckstein et Donath 2005, Quested et Eriksson 2006). În funcţie de mecanismele care stau la baza influenţelor exercitate de gosimea stratului de litieră asupra comunităţilor vegetale putem deosebi următoarele efecte: - efecte fizice – sunt rezultatul proceselor care duc la modificarea condiţiilor microclimatice, determină interacţiunile plantelor cu mediul fizic (Nakamura 1996, Rotundo et Aguiar 2005). Acumularea de litieră prezintă o heterogenitate spaţială datorată heterogenităţii vegetaţiei, a redistribuirii litierei şi descompunerii ei inegale, ceea ce determină o heterogenitate a condiţiilor microclimatice (Facelli et Pickett 1991); - efecte chimice – care se datorează în primul rând proceselor de descompunere la care este supusă litiera, acesta fiind un determinant major al înmagazinării carbonului şi al altor nutrienţi şi al fluxului de materie în ecosisteme (Quested et Eriksson 2006). Descompunerea litierei şi eliberarea nutrienţilor este controlată de o mare varietate de proprietăţi chimice ale acesteia, dintre care putem aminti concentraţia de N, raportul C:N, concentraţia de P, raportul C:P, raportul N:P, cantitatea de lignină, concentraţia compuşilor fenolici (Aerts et al. 2003). Proprietăţile chimice ale litierei sunt determinate de compoziţia calitativă a acestei care se datorează, în mare parte, speciilor dominante (Hoorens et al. 2002, 2003b, a). Litiera speciilor de dicotiledonate poate avea un efect chimic mult mai puternic decât cea a gramineelor datorită fitotoxinelor şi a eliberării de nutrienţi într-o perioadă de timp mai scurtă, viteza de descompunere a litierei produsă de graminee fiind mai mică (Xiong et al. 2003). De asemenea, descompunerea litierei afectează pH-ul şi conţinutul chimic al apelor de percolare şi conduce la formarea acizilor prehumici, humici şi fulvici (Walsh et Voigt 1977), având un efect de fertilizare a solului (Monk et Gabrielson 1985); - efecte biologice – dintre care menţionăm reducerea densităţii şi a numărului de specii (Monk et Gabrielson 1985, Carson et Peterson 1990, Foster et Gross 1998), influenţează germinaţia seminţelor, creşterea şi dezvoltarea plantulelor, incidenţa bolilor fungice, prezenţa şi intensitatea fenomenelor alelopatice (Xiong et al. 2003, Rotundo et Aguiar 2005) şi dinamica populaţiilor de nevertebrate din sol (Xiong et Nilsson 1999). De asemenea, afectează şi relaţiile intra- şi interspecifice ale indivizilor populaţiilor de plante (Nakamura 1996), cu rol important în structurarea fitocenozelor. Dacă analizăm rolul litierei asupra populaţiilor de plante şi a comunităţilor vegetale din punct de vedere calitativ, efectele acesteia pot fi: - efecte pozitive – care se pot datora îmbunătăţirii condiţiilor hidrice şi reducerea stresului termic, aportului de substanţe nutritive şi a diseminării seminţelor ca urmare a redistribuirii ei (Foster et Gross 1998, Xiong et Nilsson 1999, Eckstein et Donath 2005, Maret et

33
Wilson 2005, Quested et Eriksson 2006), iar în stadiile iniţiale ale succesiunii, acumularea de litieră promovează dezvoltarea plantulelor (Nakamura 1996); - efecte negative – determinate în general de creşterea cantitatăţii de litieră ceea ce duce la o reducere a intensităţii luminii la nivelul solului (Facelli et Pickett 1991), creşterea numărului de bacterii şi ciuperci patogene ca urmare a creşterii umidităţii şi a temperaturii (Bosy et Reader 1995, Maret et Wilson 2005), inhibă germinaţia seminţelor, în special a celor mici, a căror germinaţie este redusă cu până 100%, şi creşterea şi dezvoltarea plantulelor (Bosy et Reader 1995, Foster et Gross 1998, McAlpine et Drake 2003). Toate aceste modificări cauzate de acumularea de litieră determină o scădere a bogăţiei specifice, a biodiversităţii şi a productivităţii fitocenozelor (Monk et Gabrielson 1985, Eckstein et Donath 2005). Litiera este un factor fundamental care controlează structura, stabilitatea şi dinamica comunităţilor vegetale (Carson et Peterson 1990, Grime 2001, Bărbos 2007), având un rol mai important în organizarea fitocenozelor decât în circuitul biogeochimic al nutrienţilor (Facelli et Carson 1991, Bărbos 2007).
2.2.4. Caracteristici edafice
Reacţia (aciditatea) solului este un factor care poate influenţa în mod direct sau indirect structura şi dinamica covorului vegetal. Astfel, variaţia pH-ului are un rol important în determinarea structurii fitocenozelor, influenţând în mod direct atât germinarea seminţelor (Berendse et al. 1998), cât şi a creşterii şi dezvoltării sistemului radicular al plantelor (Balsberg-Pahlsson 1995) şi a capacităţii de absorbţie a substanţelor nutritive de către acestea (Fageria et
al. 1989, Tyler et Olsson 2001) având astfel, un rol important în determinarea vitalităţii plantelor. Influenţa indirectă a acidităţii solului asupra fitocenozelor se manifestă prin intermediul proceselor de descompunere şi mineralizare a materiei organice. Aciditatea solului determină compoziţia şi activitatea florei microbiene, prin aceasta determinând atât viteza de descompunere şi mineralizare a substanţei organice, cât şi tipul de humus rezultat în urma acestor procese (Hopkins et al. 1990, Greszta et al. 1992, Raubuch et Beese 2005) (Hopkins et
al. 1990, Greszta et al. 1992, Raubuch et Beese 2005). Humusul din sol, prin proprietăţile şi compoziţia chimică, este o componentă esenţială a solului care îi conferă acestuia un anumit grad de fertilitate (Blaga et al. 2005). Humusul reprezintă rezerva permanentă de elemente minerale nutritive a solului pe care le eliberează treptat pentru a fi folosite de către plante în nutriţie (Chiriţă 1955, 1974, Blaga et al. 2005). Fertilitatea nu este singura caracteristică a solului care este influenţată în mod direct de către cantitatea de humus. Humusul, împreună cu argila atenuează levigarea unor cationi şi influenţează în mod favorabil formarea structurii glomerulare a solului influenţând astfel porozitatea şi permeabilitatea solului. De asemenea, humusul îmbunătăţeşte însuşirile fizice ale solurilor prin acizii humici care imprimă solurilor culori închise determinând astfel o creştere a gradului de absorbţie a radiaţilor calorice şi implicit o creştere a gradului de încălzire (Blaga et al. 2005). Fosforul solubil din sol împreună cu azotul sunt pricipalii nutrienţi necesari pentru creşterea şi dezvoltarea plantelor (Parton et al. 1988, Niinemets et Kull 2005), raportul dintre cele două elemente din ţesutul plantelor indicând care dintre ele are rol de factor limitativ.

34
Astfel, un raport N:P < 10 (Gusewell et Koerselman 2002, Gusewell 2004, 2005) sau < 14 (Mamolos et al. 2005) indică faptul că azotul are rol limitativ, iar un raport N:P >16 (Mamolos et al. 2005) sau > 20 (Gusewell 2004) indică faptul că fosforul are rol limitativ. Dacă din punct de vedere cantitativ, o mare parte a variabilităţii fosforului din sol poate fi atribuită texturii solului (Stewart et Cole 1989), din punct de vedere calitativ un rol important îl are troficitatea solului şi tipul de vegetaţie. Astfel, în solurile oligotrofe, fosforul se găseşte predominant sub formă organică, disponibilitatea sa pentru plante fiind condiţionată de mineralizarea acesteia (Williams et al. 1999). Din punct de vedere al vegetaţiei, aceasta afectează distribuţia formelor de fosfor din sol, pajiştile conţinând fosfor în combinaţii organice labile sau moderat-labile, care poate fi potenţial utilizabil de către plante (Magid 1993). Influenţa fosforului în viaţa plantelor se manifestă atât direct, prin rolul nutritiv, cât şi indirect, fiind unul dintre factorii care condiţionează mineralizarea azotului, fertilizarea solurilor cu fosfor având un impact pozitiv asupra circuitului azotului (Purchase 1974). Efectul fosforului asupra mineralizării azotului este amplificat de prezenţa vegetaţiei, datorită stimulării creşterii rădăcinilor, fapt ce determină o creştere a volumului rizosferei şi, implicit, a numărului de microorganisme nitrificatoare (Cornish et Raison 1977). Azotul amoniacal este unul dintre principalii factori limitativi care determină atât creşterea şi dezvoltarea plantelor cât şi bogăţia specifică a fitocenozelor. Resursele de azot din sol sunt atât minerale sub formă de azot nitric şi amoniacal, cât şi organic sub formă de aminoacizi simpli care pot fi absorbiţi de către plante (Näsholm et al. 2000). Solurile acide din zona temperată şi arctic-alpină, sunt mai bogate în azot amoniacal decât în azot nitric (Gigon et Rorison 1972, Atkin et Cummins 1994). Preferinţa plantelor pentru azotul amoniacal (NH4
+ ) putând fi pusă pe seama mecanismelor diferite de absorbţie a azotului, absorbţia azotului amoniacal necesitând un consum mai mic de energie decât al azotului nitric (Atkin et Cummins 1994). Concentraţia ionilor de Ca++ din sol influenţează compoziţia specifică a solului prin modificarea pH-ului acestuia şi dezvoltarea rădăcinilor plantelor. Multe dintre speciile acidofile sunt afectate în mod direct de o creştere a concentraţiei ionilor de calciu din sol atât ca urmare a scăderii acidităţii solului cât şi indirect datorită efectului de inhibiţie pe care îl au speciile competitoare asupra acestora (Grime et al. 1996, Schellberg et al. 1999). Unele specii ca Festuca rubra, Agrostis capillaries etc sunt favorizate de creşteri mici ale nivelului de calciu din sol în timp ce alte specii ca Nardus stricta sunt inhibate (Bradshaw et al. 1958, Bărbos 2007). De asemenea, concentraţia ionilor de calciu din sol, prin modificarea acidităţii acestuia, influenţează şi bogăţia specifică a fitocenozelor (Common et al. 1991, Bărbos 2007, Hejcman et
al. 2007).

35
III. PLANUL DE MONITORIZARE AL HABITATULUI 6230*
3.1. Studii preliminare
Înaintea demarării oricărui program de monitorizare a habitatelor Natura 2000, în
general, şi a pajiştilor de Nardus stricta bogate în specii pe substrat silicatic (habitatul 6230*), în mod special, este necesară efectuarea unor studii preliminare care să ofere date privind corologia habitatului dar şi a particularităţiilor structurale, funcţionale şi dinamice în situl în care se vor efectua activităţile de monitorizare. Într-o primă etapă este obligatorie studierea tuturor datelor publicate privind tipurile de comunităţi vegetale identificate în zona de interes, structura floristică a acestora, ecologia, dinamica şi corologia asociaţilor vegetale existente precum şi date privind variabilele de mediu cum ar fi: suprafaţa eşantioanelor şi numărul acestora, elementele de mezo- şi microclimă, de relief, structura geologică a substratului, tipurile de sol, modul de utilizare etc. Pentru realizarea acestui prim pas se vor consulta articolele ştiinţifice publicate, baze de date, hărţi geologice, topografice, harta solurilor etc. În condiţiile în care astfel de date nu există, sau sunt insuficiente se vor efectua studii ecologice preliminare a căror principal scop este acela de a oferi informaţii privind distribuţia spaţială a tipurile de comunităţi vegetale care intră în componenţa habitatului 6230* şi structura acestora, variabilitatea acestora, existenţa şi intensitatea factorilor perturbatori etc. În cadrul studiilor preliminare pe teren, în cazul în care nu există astfel de date, se va efectua cartarea habitatului în situl sau regiunea zonei de interes. În cadrul habitatului se vor carta separat fiecare dintre asociaţiile care intră în structura acestuia. Pentru realizarea cartării se vor utiliza hărţi topografice la scara 1:5000, 1:10000, 1:25000, în funcţie de mărimea zonei ţintă, sisteme GPS cu o acurateţe mare şi software GIS. Pentru fiecare fragment de habitat (poligon), pe teren se va delimita conturul cu ajutorul harţilor şi a GPS-ului, urmând ca în laborator, aceste date să fie prelucrate cu un soft de tip GIS. Una dintre principalele probleme în cartarea habitatului o constituie identificarea limitelor acestuia. Dacă în unele cazuri(când se învecinează cu o pădure, zonă umedă etc) limitele habitatului sunt evidente, de cele mai multe ori aceste limite sunt difuze, mai ales când se învecinează tot cu un habitat de pajişte. În aceste cazuri, pentru identificarea limitelor se foloseşte metoda taxaţiei lineare (Borza et Boşcaiu 1965). Pentru aceasta, de-a lungul transectului se vor efectua ridicări succesive pe suprafeţe de probă cu suprafaţa de 1 m2 în care se notează atât prezenţa speciilor cât şi dominanţa lor. Prin compararea datelor astfel obţinute, se pot delimita, relativ uşor, limitele dintre diversele tipuri de fitocenoze, limite oglindite în variaţia compoziţie floristice şi a indicelui de dominanţă al

36
speciilor. De asemenea, limitele dintre două grupări vegetale se pot stabili şi prin calcularea unor indici fitocenotici (omotonitatea, indicele de similaritate) pe baza datelor înregistrate pe teren. Cartarea este necesară atât pentru stabilirea numărului de eşantioane necesare monitorizării habitatului cât şi pentru monitorizarea de-a lungul timpului a extinderii acestuia, aceste date constituind punctul de referinţă pentru viitor.
3.2. Colectarea datelor
Aşa cum am arătat şi în primul capitol, în programele de monitorizare a habitatelor trebuie acordată o importanţă deosebită etapei de colectare a datelor. De natura şi corectirudinea datelor colectate depinde succesul întregului program de monitorizare.
3.2.1. Eşantionarea
Tipul de eşantionaj. Pentru monitorizarea habitatului de pajişti de Nardus
stricta bogate în specii pe substrat silicios se va efectua un eşantionaj stratificat. Astfel, în cadrul habitatului 6230* se va efectua un eşantionaj aleatoriu în fiecare tip de comunitate vegetală identificată şi cartată, evitându-se zonele de ecoton. Pentru aceasta, se va realiza un grid de 10x10 m (100 m2) care se va suprapune peste poligoanele care reprezintă pajiştile care trebuie monitorizate. Fiecare celulă a gridului reprezintă câte o potenţială suprafaţă de probă şi va primi un număr unic de identificare. Alegerea eşantioanelor care vor fi analizate se va face prin metoda numerelor aleatoare. Această metodă, presupune ca, din numărul total de eşantioane posibile, să se extragă, în mod aleatoriu, numerele celor n suprafeţe de probă necesare.
De exemplu, în cazul unei suprafeţe de 100 ha care conţine 10000 de posibile suprafeţe de probă din care vor fi analizate doar 95, aplicând metoda numerelor aleatoare, eşantioanele care vor avea următoarele numere de identificare:
4, 4353, 1745, 9299, 7439, 39, 3038, 830, 3810, 8337, 930, 2104, 1094, 7373, 4120, 2129, 400,
5566, 9140, 6287, 3551, 5719, 5183, 7732, 4571, 6220, 2982, 2214, 2412, 4430, 1022, 7442,
4936, 9815, 6803, 7637, 6848, 3433, 4377, 4946, 1060, 5946, 1888, 2073, 3772, 8514, 4500,
4605, 8487, 9640, 8745, 7319, 3083, 6307, 4060, 6045, 7138, 9620, 6100, 5534, 3725, 3542,
4256, 4234, 1099, 310, 2732, 8859, 4835, 7636, 539, 5236, 314, 898, 790, 6985, 8724, 8185,
3808, 346, 7174, 7279, 1722, 1648, 4960, 1437, 4473, 3797, 3529, 5194, 8442, 9474, 7302,
2303, 9268
În cazul în care eşantioanele, astfel alese, sunt situate în zone de ecoton sau, din diverse
motive, nu pot fi utilizate pentru scopul propus, acestea vor fi înlocuite cu altele alese prin aceaşi metodă. Forma şi mărimea eşantioanelor. Pentru monitorizarea pajiştilor de Nardus stricta se vor utiliza suprafeţe de probă de formă pătrată şi 100 m2 (10x10 m).

37
Numărul de eşantioane se va stabili în funcţie de mărimea fragmentelor de habitat. Pentru determinarea numărului de eşantioane necesare, se va alege valoarea probabilităţii de transgresiune (de obicei se utilizează o valoare p = 0,05), iar pe baza datelor obţinute din studiile preliminare se va stabili intervalul de încredere (toleranţă) dorit (Krebs 1998, Krebs et
Kenney 2000). De exemplu, pentru un poligon cu o suprafaţă de 100 ha, care conţine 10000 de posibile suprafeţe de probă, pentru ca variabilele măsurate să aibă o probabilitate de transgresiune p=0,05 şi o toleranţă de ±10% numărul de eşantioane necesar este de 95. Marcarea suprafeţelor de probă se va face atât prin notarea coordonatelor geografice, cu ajutorul GPS-ului, a colţului de jos din partea stânga a fiecărui eşantion. Datorită erorilor relativ mari pe care le au măsurătorile făcute cu GPS-ul (în funcţie de marcă şi model) se recomandă montarea unor markeri. Frecvenţa eşantionării. Dată fiind stabilitatea relativ mare a fitocenozelor care intră în componenţa habitatului 6230*, şi având în vedere şi costurile mari, colectarea datelor pe teren se poate efectua odată la 3 ani. Cu toate acestea, se recomandă vizitarea suprafeţelor de probă anual astfel încât să se poată surprinde eventualele modificări datorate acţiunii unor factori imprevizibili. În condiţiile în care programul de monitorizare urmăreşte efectele măsurilor de management adaptativ care au ca scop readucerea habitatului într-o stare favorabilă pentru conservare, monitorizarea suprafeţelor de probă se va realiza anual.
3.2.2. Releveul fitosociologic
Releveul fitosociologic reprezintă o modalitate standardizată (vezi Anexa I) de colectare a datelor pe teren utilizată în studiile de vegetaţie. Se recomandă utilizarea acestor fişe standard pentru a se evita omiterea înregistrării unor date care se pot dovedi a fi foarte importante. Este obligatorie completarea pe teren a tuturor informaţiilor disponibile. Câmpurile corespunzătoare variabilelor ecologice care nu se pot măsura pe teren (concentraţia cationilor din sol, textura solului etc.) vor fi completate ulterior după analiza probelor de sol. În funcţie de complexitatea studiului, la fiecare specie, pe lângă abundenţa-dominanţa ei, se vor nota şi date privind starea fenologică, stratul din care fac parte şi numărul de indivizi din suprafaţa de probă (densitatea). În cazul gramineelor cespitoase (care formează tufe) se vor nota fie numărul de tufe, fie se va determina numărul mediu de tulpini/tufă şi se va extrapola la nivelul întregii suprafeţe de probă. În aprecierea abundenţei-dominanţei speciilor este recomandată utilizarea ramelor metrice pentru reducerea erorilor datorate subiectivităţii fiecărui observator în estimarea acesteia.
3.2.3. Structura calitativă a habitatelor (sinstructura)
Compoziţia floristică este determinată de totalitatea speciilor din fitocenoza analizată şi
reprezintă “ … un element important de diferenţiere, delimitare şi tipizare a fitocenozelor” (Ivan 1979). Din acest considerent, o importanţă deosebită o are corecta identificare şi determinare a taxonilor vegetali. Pentru aceasta se va utiliza Flora R.P.R – R.S.R., vol. I-XIII (Săvulescu 1952-1976) şi Flora mică ilustrată a României, (Ciocârlan 1988, 1990, 2000). Datorită deselor modificări nomenclaturale a speciilor, pentru standardizarea denumirilor ştiinţifice se va utiliza

38
Flora Europaea (Tutin et al. 1968, 1972, 1976, 1980, Tutin et al. 1993). În cazul în care determinarea corectă a speciilor un poate fi făcută pe teren, se vor colecta 2-3 exemplare care vor fi determinate în laborator, sau vor fi trimise spre determinare unor specialişti.
Pentru încadrarea speciilor care alcătuiesc fitocenozele analizate în diferite grupe cenotice, categorii de bioforme, geoelemente, ecologice şi economice, se recomandă consultarea mongrafiilor publicate pentru flora şi vegetaţia României (Pop 1978, 1982, Coldea 1990, 1991, Sanda et al. 1998, Sanda et al. 2001, Pop et al. 2002, Sanda 2002, Sanda et al. 2003). Deoarece aceste aspecte sunt tratate pe larg în toate studiile de vegetaţie, nu vom insista asupra lor.
Pentru tipurile de comportament social al plantelor se va consulta lucrarea monografică a lui Borhidi (1995). Prin comportament social se înţelege modul prin care planta este legată de habitat, informaţia conţinută şi naturaleţea legăturii, bazându-se pe rolul pe care fiecare specie îl joacă în cadrul comunităţii vegetale.
Abordarea studiilor de vegetaţie prin prisma tipurilor de comportament social al plantelor este puţin întâlnită, deşi oferă informaţii preţioase pentru înţelegerea mecanismelor care stau la baza formării şi evoluţiei diferitelor tipuri de habitate, vom prezenta sistemul de clasificare a speciilor şi caracteristicile fiecărui tip de comportament social. În funcţie de comportamentul social, plantele se împart în: I. Plante competitoare ( C ) II. Plante tolerante la stres (ST) A. Plante tolerante la stres, cu amplitudine ecologică mică: specialiste (S)
B. Plante tolerante la stres, cu amplitudine ecologică mare: generaliste sau
companioane (G)
III. Plante ruderale (R) A. Plante de habitate perturbate de factorii naturali: pionieri naturali (NP)
B. Plante de habitate perturbate de factorii umani:
1. Plante tolerante la perturbări ale habitatelor naturale (DT) 2. Elemente antropofile din flora nativă: buruieni native (W) 3. Elemente antropogene străine de regiune:
a. Plante introduse în cultură şi sălbaticite (I) b. Buruieni adventive (A)
4. Plante competitoare ale habitatelor secundare a. Competitoare ruderale ale florei naturale (RC) b. Competitoare străine, invadatoare agresive (AC)
Deoarece, în opinia autorului, tipurile de comportament social exprimă variatele stadii
ale naturaleţei sau perturbării relaţiei plantă-habitat, acestea pot fi înlocuite cu indici relativi ca „valoarea naturaleţei”, în caracterizarea fiecărui tip comportamental vom prezenta şi indicii propuşi de către acesta.
Plantele specialiste (S); valoare +6 – grupează plantele stenotolerante, majoritatea având o competitivitate redusă, dar care sunt indicatori sensibili a factorilor ecologici ai habitatului şi/sau specii caracteristice a unei comunităţi sau grup de comunităţi (alianţă), fiind purtătoarele unor importante informaţii ecologice şi fitosociologice. Ca urmare a competivităţii lor scăzute într-un mediu dat, ele sunt primele specii care răspund tuturor schimbărilor care au

39
loc în cadrul habitatului. Chiar dacă nu au un rol important în stabilitatea sau rezilienţa comunităţii vegetale, absenţa sau dispariţia lor este un semn al perturbării, iar reapariţia lor este dovada indubitabilă a reabilitării comunităţii respective.
Plantele competitoare (C); valoare +5 – reprezintă speciile dominante ale comunităţilor naturale sau cel puţin în unele dintre straturile lor, incluzând şi speciile competitoare tolerante la stres. Majoritatea speciilor din această categorie sunt perene sau lemnoase, au o producţie de biomasă mare, iar prin activitatea lor reglează consumul de materie şi energie, distribuţia şi fluxul în cadrul comunităţii, selectează şi controlează posibilitatea coexistenşei grupurilor de specii însoţitoare, iar prin biomasa produsă, influenţează procesele de dezvoltare a solului. Ele sunt capabile să stabilizeze compoziţia şi funcţionarea comunităţii pe o perioadă lungă de timp. De asemenea, fiind relativ rezistente la influenţele perturbatoare, protejează structura şi caracteristicile de bază ale comunităţilor de efectele acestor influenţe, pentru un timp îndelungat.
Plantele generaliste (însoţitoare, companioane) (G); valoare +4 – sunt plante cu o largă amplitudine ecologică din comunităţile vegetale naturale, care supravieţuiesc într-o mare diversitate de habitate deşi nu sunt foarte tolerante faţă de perturbările antropogene. Majoritatea sunt plante perene, care, prin marea lor adaptabilitate se integrează foarte bine în lanţurile trofice şi energetice ale comunităţilor din care fac parte, iar prin umplerea golurilor existente joacă un rol important în balanţa, stabilitatea şi dinamica internă a fitocenozelor. De asemnea, iau parte la amortizarea perturbărilor minore şi au importanţă în menşinerea diversităţii genetice şi a funţionării homeostatice.
Pionierii naturali (NP); valoare +3 – sunt speciile care intră în componenţa stadiilor iniţiale ale seriilor succesionale dezvoltate pe substraturi puternic perturbate, dar, care, deşi sunt tolerante faţă de acţiunea factorilor abiotici, cerinţele lor faţă de nutrienţi şi competitivitate sunt scăzute, majoritatea fiind plante anuale. În consecinţă, capacitatea lor de a menţine în timp stabilitatea comunităţilor vegetale din care fac parte, este de asemenea scăzută., dar, au un rol important în reabilitarea fitocenozelor.
Plantele tolerante la perturbări (DT); valoare +2 – această categorie include elementele pioniere ale succesiunilor secundare, care apar după distrugerea comunităţilor permanente şi sunt capabile să folosească surplusul de nutrienţinaturali realizat în timpul perturbărilor. Aici pot fi încadrate plantele perene care iau parte la popularea substratelor naturale sau seminaturale, la origini fiind specii însoşitoare (generaliste) care pot utiliza habitate perturbate prezentând o dinamică populaţională ridicată.
Buruienile (W); valoare +1 – grupează speciile de plante care trăiesc în habitate artificiale sau puternic perturbate de influenţe antropice repetate pe perioade lungi de timp. Majoritatea lor sunt plante anuale sau efemere capabilă să dea naştere la 3-4 generaţii în cursul unei perioade de vegetaţie. Ele sunt specii de plante care cresc pe marginea drumurilor, pe soluri fertilizate sau detritice, semănături etc., care de-a lungul timpului s-au naturalizat în flora spontană.
Plantele introduse (I); valoare –1 – cuprinde speciile de plante străine într-o regiune sau floră şi care au fost introduse intenţionat şi aclimatizate ca potenţiale culturi utilizate în diferite scopuri economice. Speciile de plante introduse, care prin agresivitatea lor depăşeşsc limitele suprafeţelor cultivate şi ocupă habitate naturale crează tipuri de comunităţi vegetale străine

40
unei regiuni vor fi tratate ca o categorie aparte. Numărul speciilor introduse se modifică constant şi în majoritatea cazurilor se obsrvă o tendinţă de creştere a numărului lor.
Plantele adventive (A); valoare –1 – grupează speciile de plante străine pentru o regiune sau floră, care au fost introduse accidental, ca urmare a activităţilor umane, şi care se aclimatizează sub formă de buruieni. În habitatele lor originale, majoritatea sunt specii ale comunităţilor naturale. Aceste specii se adaptează singure la vegetaţia habitatelor secundare, dar niciodată nu invadează masiv comunităţile vegetale naturale.
Competitorii ruderali (RC); valoare –2 – reprezintă buruienile dominante care formează comunităţi şi care datorită strategiei eficiente de propagare şi a mediilor cu competitivitate scăzută devin edificatori secundari şi sunt capabile să transforme habitatul şi să modifice tendinţa succesională a fitocenozelor. Majoritate produc cantităţi mari de seminţe care îşi conservă capacitatea germinativă timp îndelungat, chiar şi în condiţii nefavorabile. Multe geofite, ca urmare a utilizării pe scară largă a ierbicidelor, devin buruieni agresive.
Plantele invazive competitoare (AC); valoare –3 – sunt acele specii de plante străine pentru o regiune şi floră, introduse intenţionat sau accidental, care prin strategia lor agresivă de propagare şi datorită mediului cu o competitivitate scăzută şi independenţa lor faţă lanţurile trofice locale, sunt capabile să invadeze spaţiile libere din comunităţile naturale şi seminaturale şi să devină dominante. Deseori se manifestă ca edificatori secundari, iar prin eliminarea treptată a competitorilor locali devin un obstacol al proceselor naturale de succesiune. Ca urmare, ele pot distruge stabilitatea fitocenozelor naturale şi deviază succesiunea naturală şi reabilitarea lor de la tendinţa progresivă, producând deficienţe majore, determinând apariţia de ecosisteme străine, punând în pericol supravieţuirea şi regenerarea habitatelor şi comunităţilor vegetale naturale.
Pentru o mai bună vizualizare a structurii fitocenozelor monitorizate, se pot realiza spectrele diferitelor categorii de elemente structurale (bioforme, geoelemente, categorii ecologice etc.), care nu sunt altceva decât transpunerea grafică a ponderii acestora în cadrul eşantioanelor analizate. Pentru realizarea spectrelor se poate lua în calcul fie numărul de specii din fiecare categorie în parte raportat la numărul total de specii (ponderea procentuală), fie abundenţa-dominanţa speciilor din fiecare categorie raportată la acoperirea generală cu vegetaţie a eşantionului/eşantioanelor analizate.
Realizarea spectrelor diferitelor categorii de specii este o metodă ieftină şi rapidă de evidenţiere a anumitor modificări a structurii calitative a habitatelor analizate, modificări care sunt rezultatul acţiunii diferiţilor factori ecologici, care pot fi astfel identificaţi. Cunoscând autoecologia speciilor care alcătuiesc o fitocenoză, analiza spectrelor permite o caracterizare a staţiunilor în care aceste fragmente de vegetaţie se întâlnesc. Astfel, în cadrul programelor de monitorizare, prin compararea spectrelor realizate pe baza datelor colectate în perioade diferite de timp, se pot pune în evidenţă anumite tendinţe dinamice ale fitocenozelor care intră în structura habitatului/habitatelor analizate. Cu toate acestea, pentru studiile de monitorizare, nu este suficientă simpla reprezentare grafică a structurii eşantioanelor analizate, eventualele diferenţe, care se pot datora şi întâmplării nu numai acţiunii anumitor factori de mediu, trebuie testate cu ajutorul testelor statistice parametrice sau neparametrice (în funcţie de datele colectate), astfel încât, rezultatele obţinute să aibă acoperire statistică. Spectrele, alături de alte metode de analiză, au un rol important şi elaborarea diverselor ipoteze de lucru, care
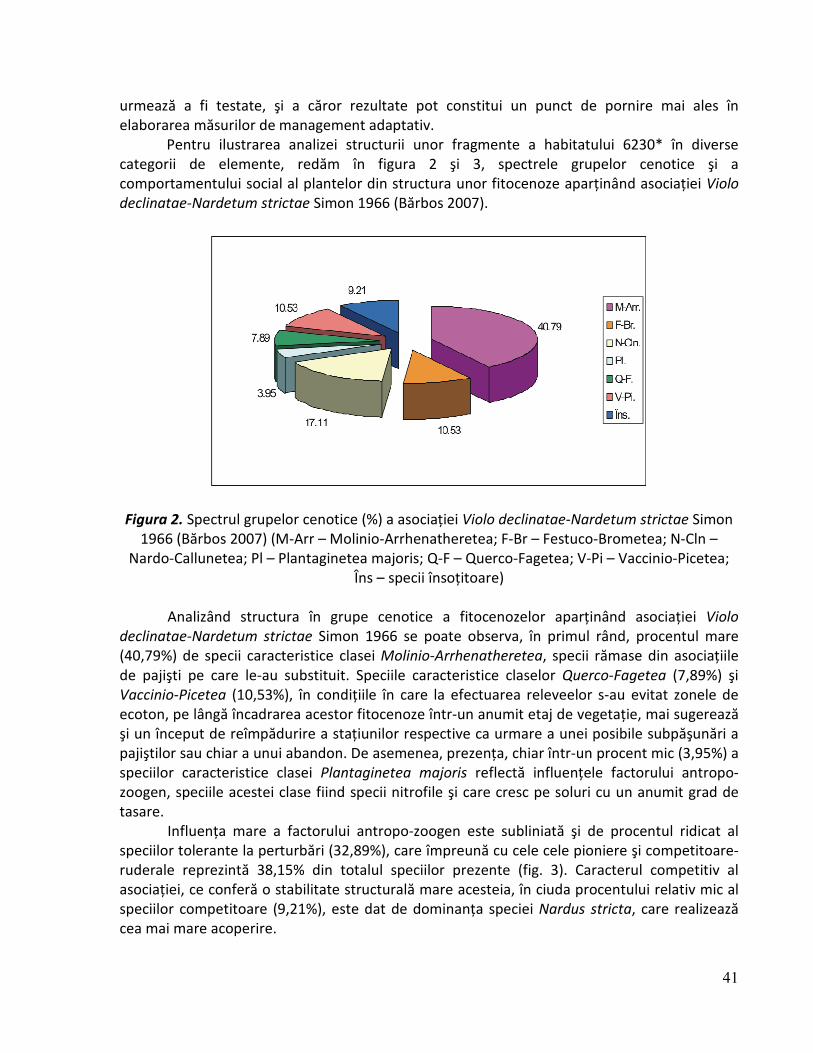
41
urmează a fi testate, şi a căror rezultate pot constitui un punct de pornire mai ales în elaborarea măsurilor de management adaptativ.
Pentru ilustrarea analizei structurii unor fragmente a habitatului 6230* în diverse categorii de elemente, redăm în figura 2 şi 3, spectrele grupelor cenotice şi a comportamentului social al plantelor din structura unor fitocenoze aparţinând asociaţiei Violo
declinatae-Nardetum strictae Simon 1966 (Bărbos 2007).
Figura 2. Spectrul grupelor cenotice (%) a asociaţiei Violo declinatae-Nardetum strictae Simon 1966 (Bărbos 2007) (M-Arr – Molinio-Arrhenatheretea; F-Br – Festuco-Brometea; N-Cln –
Nardo-Callunetea; Pl – Plantaginetea majoris; Q-F – Querco-Fagetea; V-Pi – Vaccinio-Picetea; Îns – specii însoţitoare)
Analizând structura în grupe cenotice a fitocenozelor aparţinând asociaţiei Violo
declinatae-Nardetum strictae Simon 1966 se poate observa, în primul rând, procentul mare (40,79%) de specii caracteristice clasei Molinio-Arrhenatheretea, specii rămase din asociaţiile de pajişti pe care le-au substituit. Speciile caracteristice claselor Querco-Fagetea (7,89%) şi Vaccinio-Picetea (10,53%), în condiţiile în care la efectuarea releveelor s-au evitat zonele de ecoton, pe lângă încadrarea acestor fitocenoze într-un anumit etaj de vegetaţie, mai sugerează şi un început de reîmpădurire a staţiunilor respective ca urmare a unei posibile subpăşunări a pajiştilor sau chiar a unui abandon. De asemenea, prezenţa, chiar într-un procent mic (3,95%) a speciilor caracteristice clasei Plantaginetea majoris reflectă influenţele factorului antropo-zoogen, speciile acestei clase fiind specii nitrofile şi care cresc pe soluri cu un anumit grad de tasare. Influenţa mare a factorului antropo-zoogen este subliniată şi de procentul ridicat al speciilor tolerante la perturbări (32,89%), care împreună cu cele cele pioniere şi competitoare-ruderale reprezintă 38,15% din totalul speciilor prezente (fig. 3). Caracterul competitiv al asociaţiei, ce conferă o stabilitate structurală mare acesteia, în ciuda procentului relativ mic al speciilor competitoare (9,21%), este dat de dominanţa speciei Nardus stricta, care realizează cea mai mare acoperire.

42
Figura 3. Spectrul tipurilor de comportament social al plantelor (%) asociaţiei Violo declinatae-
Nardetum strictae Simon 1966 (Bărbos 2007) (C – competitoare; G – generaliste; DT – tolerante la perturbări; NP – pionieri naturali; RC – competitoare ruderale; S – specialiste; Ned –
comportament nedeterminat)
3.2.4. Indicii fitopopulaţionali
Abundenţa-Dominanţa. Pe suprafeţe mici, estimarea acoperirii poate fi deseori
suficientă, dar “… pe suprafeţe mari, în special pe suprafeţe mai puţin omogene, estimarea
combinată poate fi un avantaj” (Braun-Blanquet 1932). Astfel, Braun-Blanquet (1932) propune utilizarea unei scări alfa-numerice cu şapte trepte pentru evaluarea abundenţei-dominanţei (AD). Corespondenţa dintre treptele scării şi acoperirea în procente este redată în tabelul 5.
Tabel 5. Scara de apreciere a abundenţei-dominanţei, în sistemul Braun-Blanquet, completat de Tuxen şi Ellenberg (Cristea, 1993)
Braun-Blanquet orig. Intervalul de acoperire % ADm, Tuxen-Ellenberg, 1937
r 0.01-0,1 0,1
+ 0,1-1 0,5
1 1-10 5,0
2 10-25 17,5
3 25-50 37,5
4 50-75 62,5
5 75-100 87,5
Aprecierea abundenţei-dominanţei speciilor se va realiza cu ajutorul ramei metrice.
Este o metodă destule de expeditivă dar şi exactă în acelaşi timp, care presupune înregistrarea grafică la scară a suprafeţei ocupate de fiecare specie în parte sau numai a speciilor

43
edificatoare iar media tuturor repetiţiilor reprezintă valoarea acoperirii fiecărei specii şi se exprimă procentual (Weaver et Clements 1938, Greig-Smith 1964, Cristea 1993, Cristea et al. 2004). Pentru studiul formaţiunilor vegetale de pajişti se poate utiliza o scară de 1 :10 sau 1 :5 (Weaver et Clements, 1938). Utilizarea acestei metode permite şi evidenţierea structurii orizontale a fitocenozelor studiate precum şi a raporturilor spaţiale existente între specii.(Cristea, 1993).
O altă metodă de estimare a abundenţei-dominanţei speciilor este cea a fotografiei. Această metodă presupune fotografierea unor suprafeţe clar delimitate, iar pe baza fotografiilor astfel realizate se poate calcula raportul dintre suprafaţa acoperită şi suprafaţa nudă precum şi suprafaţa acoperită de speciile edificatoare (Weaver et Clements, 1938, Cristea, 1993). Dezavantajul acestei metodă constaă în faptul ca un se vor putea identifica toate speciile de plante din cadrul eşantionului analizat. Totuşi, fotografia poate fi utilizată pentru a compara structura orizontală a eşantionului la diferite intervale de timp.
În cazul studiilor populaţionale sau a studiilor mai detaliate se poate utiliza ca indice populaţional calitativ şi starea fenologică a plantelor. Starea fenologică sau periodicitatea se referă la fenomenele ritmice care apar în viaţa socială a plantelor (Braun-Blanquet, 1932; Borza et Boşcaiu, 1965; Cristea, 1993). Succesiunea diferitelor stadii fenologice determină aspectul unei fitocenoze la un moment dat din perioada de vegetaţie (Cristea, 1993). Înflorirea şi fructificarea au o importanţă mai redusă decât creşterea tulpinilor, durata cât plantele au frunze, cădrea frunzelor şi reînoirea formaţiunilor de absorbţie a rădăcinilor (Braun-Blanquet, 1932). Pentru aprecierea fenofazelor vegetaţiei ierboase se poate utiliza sistemul propus de Ivan et Spiridon (1983) şi care este redat mai jos.
Fazele şi subfazele fenologice pentru vegetaţie ierboasă (Ivan et Spiridon 1983)
I. Faza vegetativă
1. apariŃia plantulei 2. formarea rozetei 3. formarea tulpinii şi frunzelor
II. Butonizarea
1. apariŃia primilor butoni 2. butonizarea în masă
III. Înflorirea
1. începutul înfloririi 2. înflorirea în masă 3. sfârşitul înfloririi
IV. Fructificarea
1. apariŃia primelor fructe

44
2. fructificarea în masă 3. maturarea primelor fructe 4. maturarea în masă a fructelor 5. scuturarea fructelor şi diseminarea
V. Uscarea 1. începutul uscării 2. uscarea totală
Ca şi în cazul structurii calitative a fitocenozelor, rezultatele analizei stării fenologice a plantelor se pot realiza reprezentări grafice – fenospectre – prin utilizarea unui set de simboluri pentru fiecare fază sau subfază fenologică. Analiza fenologică a fragmentelor de habitat studiate poate oferi informaţii deosebit de importante pentru realizarea planurilor de management. Astfel, pe baza acestor studii se pot face recomandări practice privind perioada de începere/terminare a păşunatului, a perioadei optime de cosire etc.
3.2.5. Structura spaţială
Pentru analiza structurii orizontale a fiotcenozelor se vor utiliza metoda transectelor liniare şi metoda proiecţiilor orizontale (Cristea et al. 2004). Metoda transectelor liniare presupune notarea succesiunii fitoindivizilor de-a lungul unei linii a cărei lungime se stabileşte în funcţie de complexitatea vegetaţiei. Metoda proiecţiilor orizontale se bazează pe metodele de apreciere a abundenţei-dominanţei speciilor şi reprezintă transpunerea grafică a dispoziţiei şi relaţiilor spaţiale a fitoindivizilor. Evidenţierea structurii verticale se realizează cu ajutorul profilelor de vegetaţie care nu sunt altceva decât reprezentarea habitusului fiecărei specii de-a lungul unui transect liniar.
3.2.6. Variabilele de mediu
Plantele, ca orice organisme vii, nu trăiesc izolat ci în strânsă interdependenţă atât cu alte organisme (plante sau animale), cât şi cu materia nevie, formând împreună mediul. Variabilele de mediu luate în studiu vor fi împărţite în patru mari categorii şi anume: factori topografici, factori climatici, factori edafici şi factori biotici care includ şi componenta antropică.
3.2.6.1. Factorii topografici
Dintre factorii topografici, în studiul de monitorizare a habitatului 6230* vor fi înregistrate următoarele componente: a) coordonatele geografice (Latitudine/Longitudine) vor fi determinate cu ajutorul GPS-ului, coordonatele fiind înregistrate în sistem de proiecţie geografică cu datum WGS84.

45
b) forma de relief - componentă a factorilor topografici se va înregistra pe baza următoarei scări: 1 - vale; 2 - versant; 3 - creastă; 4 - platou. c) poziţia pe pantă a fitocenozelor, a cărei înregistrare se va face utilizând următoarea scară (Parker 1982): 1 - baza pantei; 2 – treimea inferioară a pantei; 3 - treimea mijlocie a pantei; 4 – treimea superioară a pantei; 5 - vârful pantei. d) forma pantei influenţează regimul climatic, în principal prin modificarea regimului termic şi hidric. Pentru cuantificarea acesteia se recomandă utilizarea unei scări cu 5 trepte şi anume (Parker 1982): 1 - concavă; 2 - concav-dreaptă; 3 - dreaptă; 4 - convexă; 5 - convex-dreaptă. e) panta sau înclinarea va fi măsurată cu ajutorul clinometrului sau pe baza hărţilor topografice. f) altitudinea se va determinata cu ajutorul altimetrului iar valorile ei sunt exprimate în m.s.m. (metrii deasupra nivelului mării). g) expoziţia se va determina cu ajutorul busolei şi se va exprima în grade.
3.2.6.2. Factorii edafici
Pentru determinarea caracteristicilor solurilor pe care vegetează comunităţile vegetale
eşantionate, din fiecare releveu vor fi recoltate câte 5 probe de sol din primii 20 cm de sol. Cele 5 probe vor fi amestecate rezultând o probă de sol compozită pentru fiecare releveu de vegetaţie. Colectarea probelor de sol se va realiza printr-un eşantionaj aleatoriu cu ajutorul sondei de sol. Fiecare probă de sol va fi ambalată într-o pungă de plastic şi etichetată corespunzător, astfel încât fiecărei probe să-i corespundă un singur cod de identificare, în principiu acelaşi caşi pentru releveu floristic.
Analiza probelor de sol se va face într-un laborator specializat. Pentru fiecare probă compozită de sol se vor face următoarele determinări: conţinutul de humus, de azot total, azot amoniacal, carbon organic, pH-ul, aciditatea totală, bazele de schimb, concentraţia cationilor bazici schimbabili (Ca2+, Mg2+, K+, Na+), capacitatea totală de schimb cationic, gradul de saturaţie în baze şifosforul mobil. Pentru fiecare determinare se va nota metoda şi apartura utilizată.

46
Pentru a se evita modificările caracteristicilor chimice ale solului, probele de sol recoltate, până la predarea lor la laborator pentru analize, se vor păstra la rece în lăzi frigorifice la temperaturi cât mai scăzute.
3.2.6.3. Factorii climatici
Pentru monitorizarea habitatului 6230*, acolo unde este posibil se vor procura datele climatice înregistrate de staţiile meteo cele mai apropiate. O altă metodă, deşi destul de costisitoare, presupune instalarea unor dispozitive automate de înregistrare (logger) care vor fi instalate, la o adâncime de 20 cm în sol, în suprafeţele de probă permanente. Aceste dispozitive permit înregistrarea pe o durată lungă de timp (2-3 ani) şi cu o frecvenţă ridicată, a datelor privind umiditatea relativă şi temperatura din sol.
3.2.6.4. Factorii biotici
Dintre factorii biotici luaţi în studiu amintim acoperirea generală a vegetaţiei, care oferă informaţii privind gradul de perturbare a fitocenozelor şi existenţa unor nişe libere care pot fi ocupate de alte specii. De asemenea, vor fi înregistrate date referitoare la modul de folosinţă a pajiştilor şi grosimea stratului de litieră. Modul de folosinţă a pajiştilor a fost codificat astfel încât să reflecte şi intensitatea acţiunii factorului antropo-zoogen, treptele acestei scări fiind (Bărbos 2007): 1 – fâneaţă; 2 – fâneaţă păşunată primăvara şi toamna; 3 – pajişte păşunată de vite; 4 – pajişte păşunată de vite şi oi; 5 – pajişte păşunată de oi; 6 – pajişte târlită. În cadrul acestor trepte, pentru o mai bună apreciere a impactului antropo-zoogen asupra comunităţilor vegetale, pentru păşuni, au fost introduse următoarele subdiviziuni: abandonat, subpăşunat, păşunat moderat şi suprapăşunat. Încadrarea într-una din aceste categorii se va face pe baza raportului dintre încărcătura de animale de pe păşune şi capacitatea de păşunat a fitocenozei respective. Toate datele vor fi transformate în unităţi vită mare (UVM). Datele privind încărcătura de animale se vor obţine de la proprietari, custozi sau autorităţi locale.
Grosimea stratului de litieră se va măsura în minimum 5 puncte diferite alese aleatoriu în cadrul fiecărui releveu, după care se va calcula media aritmetică, rezultatele fiind exprimate în mm.

47
3.3. Stocarea şi gestionarea datelor
Pentru stocarea şi gestionarea datelor colectate se vor utiliza fie baze de date special concepute şi create pentru studiile de vegetaţie, ca de exemplu Turboveg (Hennekens 1998-2009) (fig. 4), fie baze de date create în Access sau Excel. În mod obligatoriu, în bazele de date se vor introduce toate datele colectate împreună cu informaţiile necesare pentru identificarea fiecărui eşantion şi alte informaţii necesare prelucrării şi interpretării datelor (unitatea de măsură, metoda de măsurare etc.).
Figura 4. Stocarea datelor în Turboveg
3.4. Analiza datelor
Scopul principal al utilizării analizei numerice a datelor eco-cenotice este acela de a
descrie şi a interpreta structura unui set de date, utilizând o serie de metode numerice. Aceasta se datorează în primul rând exigenţei tot mai crescute în ceea ce priveşte credibilitatea rezultatelor obţinute în studiile de vegetaţie şi oferă o bază ştiinţifică solidă, care permite evidenţierea unor “patterns” care stau la baza originii, dezvoltării şi funcţionării comunităţilor vegetale.

48
3.4.1. Transformarea datelor
Nu toate datele brute pot fi utilizate în forma în care au fost culese ci necesită anumite transformări în vederea prelucrărilor statistice. Astfel, un prim pas, necesar, îl constituie transformarea codurilor de abundenţă-dominanţă în valori de acoperire procentuală, după scara Braun-Blanquet completată de Tüxen şi Ellenberg (Cristea et al. 2004). În analiza răspunsului speciilor, pentru normalizarea distribuţiei valorilor de abundenţă-dominanţă medie, deoarece acestea exprimă proporţii, au fost transformate cu ajutorul funcţiei arcsin cu ajutorul formulei (Zar 1999):
pp arcsin'= ,
unde: - p’ – valoarea transformată a abundenţei-dominanţei; - p – valoarea abundenţei-dominanţei exprimată în procente. În cazul variabilelor nominale calitative (forma de relief, poziţia pe pantă, forma pantei), acestea au fost recodificate binar în aşa-numitele variabile dummy (Legendre et Legendre 1998, ter Braak et Šmilauer 2002b, Leps et Smilauer 2003). Este recomandat ca Palmer (1993) recomandă ca variabilele care măsoară chimismul solului să fie logaritmate înainte de utilizarea lor în analizele multivariate (Palmer 1993b). Pentru a putea fi folosite în analize, denumirea speciilor va fi codificată astfel încât să nu depăşească 8 caractere, primele 4 reprezentând genul, iar următoarele 4 specia. În cazul subspeciilor, primele 8 caractere reprezintă genul, următoarele două, specia şi ultimele două, subspecia.
3.4.2. Indici fitocenotici
În etapa de prelucrare în laborator, pentru a putea evidenţia atât caracteristicile
generale ale cenotaxonilor, cât şi particularităţile fiecăruia, pe baza datelor înregistrate pe teren, se vor calcula o serie de indici sintetici (fitocenotici), cum ar fi: constanţa speciilor, abundenţa-dominanţa medie, indici de similaritate, omotonitate, diversitate, echitabilitate. Constanţa speciilor (K) oferă informaţii despre fidelitatea unei specii faţă de o anumită asociaţie vegetală, şi se bazează pe raportul dintre numărul eşantioane în care apare specia respectivă şi numărul total de eşantioane (Braun-Blanquet, 1932, Borza et Boşcaiu, 1965, Cristea, 1993). În calcularea constanţei se va lua în considerare numai prezenţa indivizilor unei populaţii în cadrul suprafeţei de probă fără a ţine cont şi de ceilalţi indici fitopopulaţionali. Abundenţa-Dominanţa medie (ADm) reprezintă acoperirea medie procentuală realizată de o anumită specie în cadrul fitocenozelor studiate (Cristea, 1993). ADm este un indice utilizat atât pentru reprezentarea grafică a compoziţiei în diferite categorii de elemente prin metoda Diemont, cât şi pentru calcularea altor indici sintetici sau a prelucrărilor statistice.

49
Indicii de asociere a speciilor estimează tendinţa a două sau mai multe specii de a coabita împreună în anumite fitocenoze. Două specii pot fi corelate pozitiv, ca urmare a dependenţei uneia de cealaltă sau atunci când sunt afectate în acelaşi mod de către factorii ecologici, sau pot fi corelate negativ, atunci când se exclud reciproc ca urmare a competiţiei, efectelor alelopatice sau datorită faptului că au aceleaşi cerinţe faţă de factorii de mediu (Goodall 1973). Corelaţia dintre specii poate fi studiată prin înregistrarea prezenţei sau absenţei lor într-un număr de eşantioane, prin estimarea cantitativă a abundenţei lor în fiecare eşantion sau prin măsurarea distanţei dintre indivizii diferitelor specii. Indicii care exprimă asocierea dintre două specii pot lua valori cuprinse între –1 şi +1, iar indicii care măsoară doar apariţia împreună a două specii iau valori între 0 şi 1 (Jackson et al. 1989). Indiferent de metodele utilizate pentru estimarea asocierii a două specii, calcularea indicilor respectivi are la bază termenii rezultaţi prin întocmirea tabelului de contingenţă (Goodall 1973):
Specia A
Specia B
Prezenţă Absenţă
Prezenţă a b a+b
Absenţă c d c+d
a+c b+c N=a+b+c+d
unde:
- a – numărul releveelor (eşantioanelor) în care sunt prezente ambele specii; - b – numărul releveelor în care este prezentă numai specia B; - c – numărul releveelor în care este prezentă numai specia A; - d – numărul releveelor în care nici una dintre specii nu este prezentă; - N – numărul total de relevee.
Dintre indicii de asociere ai speciilor, indicele propus de către Sokal et Michener (1958, ap. Goodall 1973) tratează în mod egal, atât prezenţa cât şi absenţa speciilor şi poate fi calculat după formula:
(a+d)/N Alţi indici, cum ar fi indicele derivat din teoria informaţiei propus de către Gordon (1966, Goodall 1973)calculat cu formula:
!!!!log2
dcba
N;
indicele Yule (1900, ap. Jackson et al., 1989):

50
bcad
bcadYule
+−
= ;
şi indicele Phi propus tot de către Yule (1912, ap. Jackson et al., 1989):
( )( )( )( )[ ] 21
dcdbcaba
bcadPhi
++++
−=
introduc în calculul asocierii speciilor şi cazurile când cele două specii nu sunt prezente în eşantioanele analizate.
Una dintre cele mai utilizate metode de estimare a asocierii speciilor este testul 2χ care
se bazează pe ipoteza că probabilitatea de a găsi fiecare specie într-un set de eşantioane, este uniformă (Goodall 1973, Kent et Coker, 1992). Asocierea dintre două specii poate fi calculată după următoarea formulă, utilizând factorul de corecţie pentru continuitate al lui Yates (1934):
( )( )( )( )( )dbcadcba
NbcadN
++++
−−=
2
2 2/χ
Pentru o mai mare acurateţe se poate utiliza în calcule o formulă alternativă (Gilbert et Wells 1966):
( )( )( )( ) ( ) ( ) ( ) ( )!!!!
!!!!!ln2
2ln
32
dcdbcaba
dcbaN
dbcadcba
N
+++++
++++=
πχ
şi compararea cu distribuţia teoretică χ2 cu un grad de libertate (Goodall, 1973). Dacă, pe lângă prezenţa sau absenţa speciilor, informaţiile despre eşantioanele studiate conţin şi unele date cantitative, se poate estima asocierea dintre două specii şi pe baza acestor date. Astfel, Ellenberg (1956) propune măsurarea cantitativă a asocierii dintre două specii pe baza formulei (Goodall 1973) :
( ) ( )
++++ ∑∑∑∑
iV
i
iU
i
iT
ii
iT
ii yxyxyx 2/
unde:
- xi şi yi reprezintă biomasa speciilor A şi B; - T – numărul de eşantioane în care sunt prezente cele două specii; - U – numărul de eşantioane în care este prezentă numai specia A; - V – numărul de eşantioane în care este prezentă numai specia B.
Primul care a studiat corelaţia dintre specii pe baza distanţei dintre indivizi a fost Dice (1952), care a luat două specii mai puţin abundente şi a măsurat distanţa dintre fiecare individ

51
al unei specii până la primul individ al altei specii (Goodall 1973). Pielou (1961) a înregistrat indivizi întâmplători împreună cu cei mai apropiaţi indivizi vecini, după care a întocmit un tabel de contingenţă pentru fiecare două specii, în care a înregistrat dacă specia A a fost sau nu un individ întâmplător şi dacă specia B a fost sau nu cel mai apropiat vecin (Goodall 1973). Pe baza datelor obţinute din tabelul de contingenţă, se calculează semnificaţia relaţiei dintre specii, cu ajutorul formulei
( )( )( ) ( )( )
1b c M
Sa b b d a c c d
+= −
+ + + + +
şi este utilizată ca un indice de segregare, care arată gradul în care indivizii unei specii tind să aibă ca cel mai apropiat vecin, mai degrabă alţi indivizi din aceeaşi specie decât din specii diferite. Indicii de asociere a speciilor permit identificarea unor specii ţintă a căror monitorizare oferă informaţii importante privind “starea de sănătate” a habitatului şi eventualele tendinţe dinamice. Indicii de similaritate oferă informaţii despre asemănarea floristică a două sau mai multe eşantioane. Gradul de similaritate dintre relevee şi gradul de asociere al speciilor sunt “două feţe ale aceleiaşi monede”(Goodall 1973), ambele aspecte având un rol important în clasificarea, ordonarea şi recunoaşterea comunităţilor vegetale. Similaritatea floristică a releveelor poate fi calculată pe baza prezenţei sau absenţei speciilor ori a datelor cantitative şi ia valori între 0 şi 1 (Jackson et al. 1989). Punctul de pornire pentru calcularea indicilor de similaritate, îl constituie, ca şi în cazul asocierii speciilor, construirea tabelului de contingenţă.
În analiza noastră am utilizat indicele de similaritate Bray-Curtis, care este un indice cantitativ semimetric, deseori folosit pentru estimarea similarităţii eşantioanelor, şi care poate lua valori cuprinse între 0 şi 1. Formula de calcul a acestui indice este (Krebs 1998, Legendre et Legendre 1998, Podani 2001, Oksanen et al. 2008):
( )∑
∑
=
=
+
−=
n
i
ikij
n
i
ikij
XX
XX
B
1
1
unde: - Xij, Xik – abundenţa speciei i în eşantioanele j şi k;
- n – numărul total de specii din eşantioane. Valoarea indicatoare a speciilor (Dufrêne et Legendre 1997) constituie un alt aspect cu
importanţă deosebită în clasificarea şi analiza vegetaţiei. Valoarea indicatoare a speciilor oferă atât o bază statistică solidă pentru stabilirea speciilor diferenţiale pentru fiecare unitate cenotaxonomică, cât şi un plus de informaţie necesar pentru interpretarea din punct de vedere ecologic a rezultatelor, prin prisma teoriei speciilor indicatoare. Valoarea indicatoare a speciilor poate fi calculată după cum urmează:

52
iijij NNA /=
jijij NreleveeNreleveeB /=
100××= ijijij BAIndVal
unde: - Aij – abundenţa medie a speciei i în fiecare cenotaxon j; - Bij – frecvenţa relativă de apariţie a speciei i în cenotaxonul j; - Nij –abundenţa medie a speciei i în cenotaxonul j; - Nreleveeij – numărul de relevee din cenotaxonul j în care apare specia i; - Ni –abundenţa speciei i în releveele analizate; - Nreleveej – numărul de relevee din cenotaxonul j; - IndVal – valoarea indicatoare a speciei i pentru cenotaxonul j.
Valoarea indicatoare a speciilor, fiind calculată independent de celelalte specii, permite compararea valorii indicatoare în comunităţi diferite, metoda nefiind influenţată de diferenţa de număr de relevee dintre unitaţile cenotaxonomice analizate sau de diferenţa de abundenţă a unei specii în diferiţi cenotaxoni (Legendre et Legendre 1998). IndVal poate lua valori cuprinse între 0 şi 100%. Speciile cu IndVal ≥ 25% pot fi considerate ca specii bune indicatoare pentru un anumit cenotaxon.
Pentru exemplificare a fost analizat un set de 59 de relevee efectuate în anul 2008 în cadrul proiectului LIFE05NAT/RO/000176 – “Habitate prioritare alpine, subalpine şi forestiere în România”, fragmentele de vegetaŃie luate în studiu aparŃinând la trei asociaŃii diferite din structura habitatului 6230* şi anume: Violo declinatae-Nardetum Simon 1966 (syn.: Nardetum strictae montanum Resmeriţă et Csűrös 1963, Nardetum strictae alpinum Buia et al. 1962, Nardetum alpigenum carpaticum Borza 1959); Festuco rubrae-Agrostietum capillaris Horvat 1951 subas. nardetosum strictae Pop 1976 şi Scorzonero roseae-Festucetum nigricantis
(Puşcaru et al. 1956) Coldea 1978 (syn.: Festucetum rubrae fallax Puşcaru et al. 1956, Festucetum rubrae montanum Csűrös et Resmeriţă 1960). Rezultatele sunt prezentate parţial în tabelul 5. Calcularea IndVal s-a realizat cu ajutorul “bibliotecii” labdsv versiunea 1.0-2 (Roberts 2006) calculându-se şi probabilitatea asociată fiecărei valori pe baza a 9999 permutari. Au fost considerate specii cu o bună valoare indicatoare cele cu IndVal ≥ 25% şi probabilitatea de transgresiune p<0.05. Analiza rezultatelor obţinute relevă faptul că cele mai multe specii cu valoare discriminantă mare sunt prezente în fitocenozele edificate de Festuca rubra şi Agrostis
capillaris şi în cele dominate de Festuca nigrescens. Pajiştile aparţinând asociaţiei Violo
declinatae-Nardetum Simon 1966 nu au decât o singură specie cu valoare discriminantă şi anume ţepoşica, care este şi specia dominantă. Acest fapt sugerează că aceste pajişti sunt degradate, ele provenind din celelate două printr-o supraexploatare a acestora. Dorim să subliniem importanţa calculării valorii indicatoare a speciilor, speciilor cu valoare indicatoare mare putând fi utilizate ca specii ţintă a căror monitorizare poate oferi informaţii preţioase privind „starea de sănătate” a habitatului.
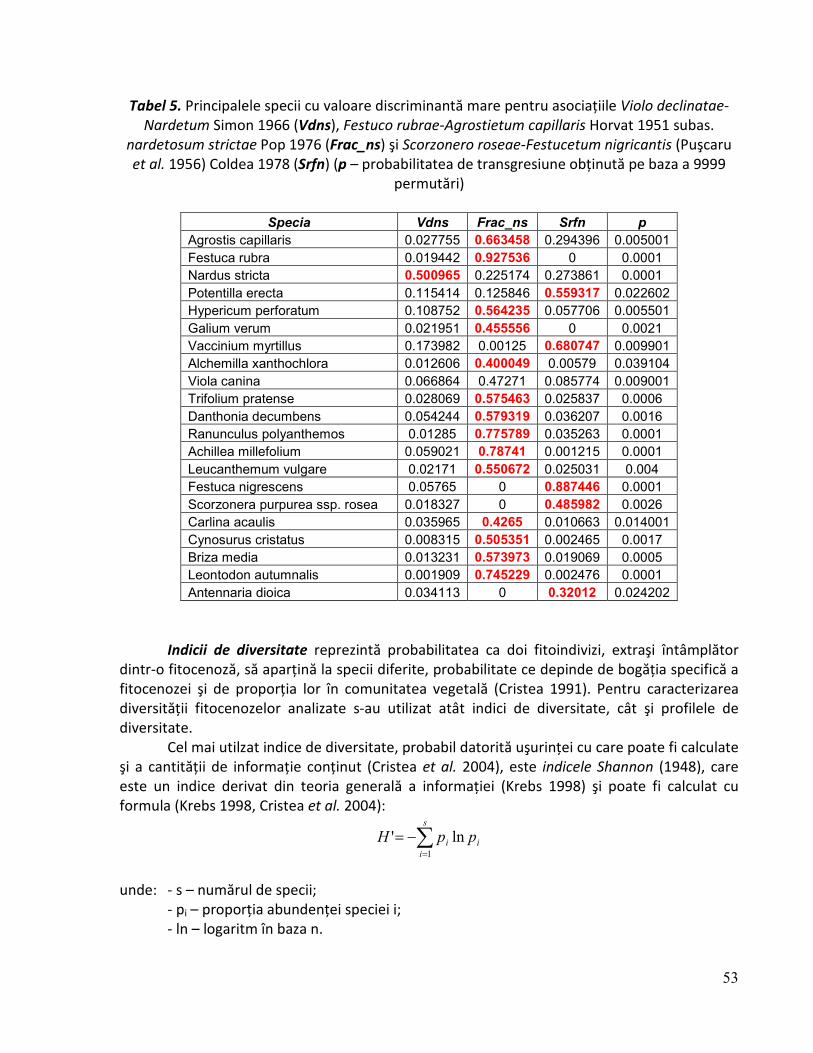
53
Tabel 5. Principalele specii cu valoare discriminantă mare pentru asociaţiile Violo declinatae-
Nardetum Simon 1966 (Vdns), Festuco rubrae-Agrostietum capillaris Horvat 1951 subas. nardetosum strictae Pop 1976 (Frac_ns) şi Scorzonero roseae-Festucetum nigricantis (Puşcaru et al. 1956) Coldea 1978 (Srfn) (p – probabilitatea de transgresiune obţinută pe baza a 9999
permutări)
Specia Vdns Frac_ns Srfn p
Agrostis capillaris 0.027755 0.663458 0.294396 0.005001
Festuca rubra 0.019442 0.927536 0 0.0001
Nardus stricta 0.500965 0.225174 0.273861 0.0001
Potentilla erecta 0.115414 0.125846 0.559317 0.022602
Hypericum perforatum 0.108752 0.564235 0.057706 0.005501
Galium verum 0.021951 0.455556 0 0.0021
Vaccinium myrtillus 0.173982 0.00125 0.680747 0.009901
Alchemilla xanthochlora 0.012606 0.400049 0.00579 0.039104
Viola canina 0.066864 0.47271 0.085774 0.009001
Trifolium pratense 0.028069 0.575463 0.025837 0.0006
Danthonia decumbens 0.054244 0.579319 0.036207 0.0016
Ranunculus polyanthemos 0.01285 0.775789 0.035263 0.0001
Achillea millefolium 0.059021 0.78741 0.001215 0.0001
Leucanthemum vulgare 0.02171 0.550672 0.025031 0.004
Festuca nigrescens 0.05765 0 0.887446 0.0001
Scorzonera purpurea ssp. rosea 0.018327 0 0.485982 0.0026
Carlina acaulis 0.035965 0.4265 0.010663 0.014001
Cynosurus cristatus 0.008315 0.505351 0.002465 0.0017
Briza media 0.013231 0.573973 0.019069 0.0005
Leontodon autumnalis 0.001909 0.745229 0.002476 0.0001
Antennaria dioica 0.034113 0 0.32012 0.024202
Indicii de diversitate reprezintă probabilitatea ca doi fitoindivizi, extraşi întâmplător dintr-o fitocenoză, să aparţină la specii diferite, probabilitate ce depinde de bogăţia specifică a fitocenozei şi de proporţia lor în comunitatea vegetală (Cristea 1991). Pentru caracterizarea diversităţii fitocenozelor analizate s-au utilizat atât indici de diversitate, cât şi profilele de diversitate.
Cel mai utilzat indice de diversitate, probabil datorită uşurinţei cu care poate fi calculate şi a cantităţii de informaţie conţinut (Cristea et al. 2004), este indicele Shannon (1948), care este un indice derivat din teoria generală a informaţiei (Krebs 1998) şi poate fi calculat cu formula (Krebs 1998, Cristea et al. 2004):
∑=
−=s
i
ii ppH1
ln'
unde: - s – numărul de specii;
- pi – proporţia abundenţei speciei i; - ln – logaritm în baza n.

54
Valorile pe care le poate lua indicele Shannon au următoarele semnificaţii (Cristea,
1991): - 0,1-1 – diversitate foarte scăzută; - 1-2,5 – diversitate scăzută; - 2,5-4 – diversitate medie; - 4-7 – diversitate ridicată; - peste 7 – diversitate foarte ridicată. Indicele Shannon poate fi exprimat şi într-o formă exponenţială, în unităţi de număr de
specii după formula (Krebs 1998): '
1HeN = ,
unde: e – baza logaritmului natural (e=2.71828); H’ – indicele Shannon calculat cu logaritm în baza e; N1 – numărul de specii egal comune care produc aceaşi diversitate ca şi H’. O serie de autori (Hill 1973, Peet 1974, Krebs 1998) recomandă utilizarea indicelui Shannon în forma exponenţială, deoarece unităţile (numărul de specii) reprezintă un concept mult mai uşor de înţeles, dar şi deoarece este sensibil la abundenţa speciilor rare din comunităţile analizate. Pentru calcularea celor doi indici de diversitate se poate utiliza programul Ecological Methodology versiunea 5.2 (Krebs et Kenney 2000). Deoarece diferiţi indici de diversitate pot caracteriza în diferite moduri aceaşi comunitate (Hurlbert 1971), o posibilă alternativă o constituie utilizarea profilului de diversitate în locul indicilor de diversitate cu valoare numerică (Tóthmérész 1995, Cristea et al. 2004). Cea mai cunoscută modalitate de a obţine profile de diversitate este cea a estimării entropiei generalizate (HR) de ordinul α cu ajutorul funcţiei Rényi (1961), după cum urmează (Cristea et al. 2004):
( )α
α
α
−=
∑=
1
ln1
S
i
ip
HR
unde semnificaţiile termenilor sunt aceleaşi ca în cazul formulei lui Shannon (α ≥ 0, α ≠ 1), formula lui Shannon fiind un caz particular al funcţiei Rényi (Oksanen et al. 2008). Pentru compararea diversităţii fitocenozelor analizate, se vor realiza profilele de diversitate pentru cele mai reprezentative relevee (releveele tip) din fiecare sintaxon identificat. Realizarea profilelor de diversitate se poate face utilizând “biblioteca” vegan (Oksanen et al. 2008) pentru R (R Development Core Team 2008). Neajunsul acestei metode de analiză a diversităţii fitocenozelor constă în faptul că diversitatea releveelor a căror curbe se intersectează nu poate fi comparată.
Curbele rang-abundenţă reprezintă curbele care aproximează cel mai bine distribuţia abundenţelelor relative ale speciilor în funcţie de rangul acestora (fig. 6). Acestea se pot încadra în cinci modele de bază: seria geometrică, modelul “bastonului rupt”, modelul log-normal, modelul Zipf şi modelul Zipf-Mandelbrot (Wilson 1991), fiecare dintre aceste modele având o anumită semnificaţie ecologică (vezi capitolul II). Analiza curbelor rang-frecvenţă
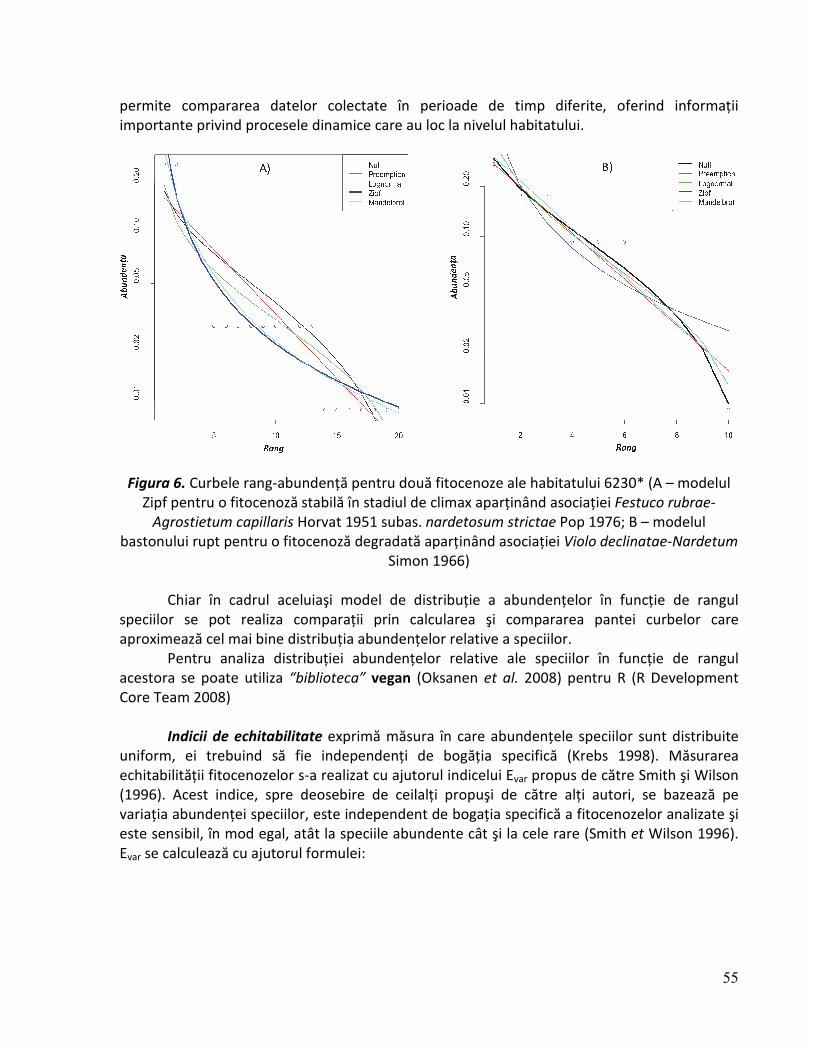
55
permite compararea datelor colectate în perioade de timp diferite, oferind informaţii importante privind procesele dinamice care au loc la nivelul habitatului.
Figura 6. Curbele rang-abundenţă pentru două fitocenoze ale habitatului 6230* (A – modelul Zipf pentru o fitocenoză stabilă în stadiul de climax aparţinând asociaţiei Festuco rubrae-
Agrostietum capillaris Horvat 1951 subas. nardetosum strictae Pop 1976; B – modelul bastonului rupt pentru o fitocenoză degradată aparţinând asociaţiei Violo declinatae-Nardetum
Simon 1966) Chiar în cadrul aceluiaşi model de distribuţie a abundenţelor în funcţie de rangul
speciilor se pot realiza comparaţii prin calcularea şi compararea pantei curbelor care aproximează cel mai bine distribuţia abundenţelor relative a speciilor.
Pentru analiza distribuţiei abundenţelor relative ale speciilor în funcţie de rangul acestora se poate utiliza “biblioteca” vegan (Oksanen et al. 2008) pentru R (R Development Core Team 2008) Indicii de echitabilitate exprimă măsura în care abundenţele speciilor sunt distribuite uniform, ei trebuind să fie independenţi de bogăţia specifică (Krebs 1998). Măsurarea echitabilităţii fitocenozelor s-a realizat cu ajutorul indicelui Evar propus de către Smith şi Wilson (1996). Acest indice, spre deosebire de ceilalţi propuşi de către alţi autori, se bazează pe variaţia abundenţei speciilor, este independent de bogaţia specifică a fitocenozelor analizate şi este sensibil, în mod egal, atât la speciile abundente cât şi la cele rare (Smith et Wilson 1996). Evar se calculează cu ajutorul formulei:

56
( ) ( )
−
−=
∑ ∑= =
s
i
s
j
jeie ssnn
E
1
2
1
var
/loglogarctan
21
π
,
unde: - ni – abundenţa speciei i;
- nj – abundenţa speciei j; - s – acoperirea generală.
Calcularea indicelui Evar se poate realiza cu ajutorul programului Ecological Methodology versiunea 5.2 (Krebs et Kenney 2000).
3.4.3. Clasificarea vegetaţiei şi analiza cluster
Unul dintre obiectivele majore ale studiilor de vegetaţie este acela de a surprinde discontinuităţile covorului vegetal şi de a clasifica diferitele tipuri de vegetaţie, pe baza unor caracteristici structurale general valabile, într-un sistem ierarhic a cărui unitate de bază o constituie asociaţia vegetală (tipul de comunitate vegetală). În monitorizarea habitatului 6230*, analizele cluster sunt utile pentru stabilirea apartenenţei fitocenozelor analizate la una dintre unităţile sintaxonomice care intră în compunerea acestui habitat, dar şi pentru evidenţierea modificărilor structurale de-a lungul timpului şi identificarea posibilelor tendinţe evolutive. Una dintre cele mai utilizate metode de analiză cluster este cea ierahică, având la bază metoda UPGMA (Unweighted Pair-Group Method using Arithmetic average) (Legendre et Legendre 1998), a matricei de disimilaritate pentru obţinerea căreia se recomandă utilizarea indicelui de similaritate Bray-Curtis. Rezultatul analizei cluster poate fi reprezentat grafic sub forma unei dendrograme (fig. 7). Gradul de distorsiune al dendrogramei poate fi apreciat cu ajutorul indicelui cofenetic, care măsoară gradul de similaritate între matricea iniţială de de similaritate şi matricea pe baza căreia a fost construită dendrograma. Pentru analiza cluster a datelor de vegetaţie se va utiliza “biblioteca” vegan (Oksanen et al. 2008) pentru R (R Development Core Team 2008).
Una dintre problemele majore ale clasificărilor numerice este aceea a stabilirii numărului optim de grupe, a nivelului la care dendrograma va fi “tăiată” (Popma et al. 1983, Everitt 1993). Stabilirea numărului optim de grupe s-a făcut după metoda propusă de către Fraley şi Raftery (2000) având la bază metoda EM (Expectation-Maximization), care utilizează BIC (Bayesian Information Criterion) pentru a determina numărul de grupe existente în datele analizate, metodă implementată în “biblioteca” mclust (Fraley et Raftery 2006) pentru R. Pentru a determina releveul cel mai reprezentativ pentru fiecare cenotaxon se recomandă efectuarea şi unei analize fuzzy cluster, care presupune ca, fiecărui releveu să-i fie atribuită o anumită probabilitatea de a aparţine unui grup sau altuia, releveele cu probabilitatea cea mai mare de a aparţine unui anumit cenotaxon fiind considerate ca fiind releveele tip. Analiza fuzzy
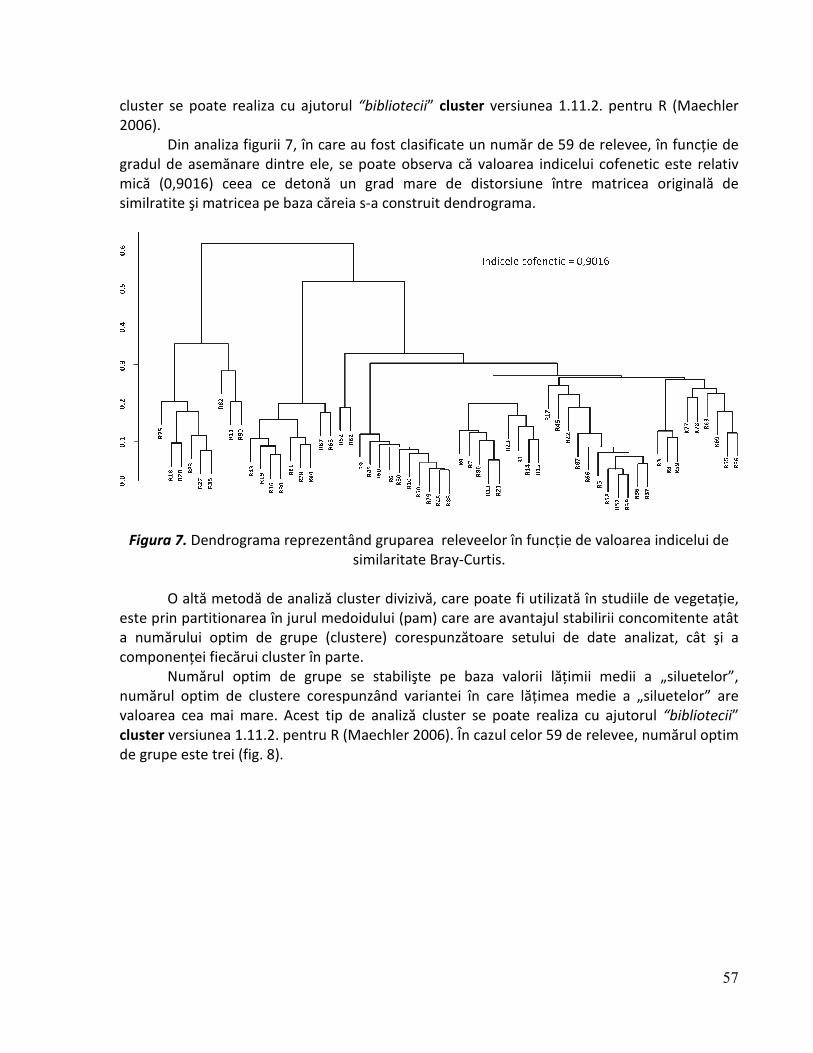
57
cluster se poate realiza cu ajutorul “bibliotecii” cluster versiunea 1.11.2. pentru R (Maechler 2006).
Din analiza figurii 7, în care au fost clasificate un număr de 59 de relevee, în funcţie de gradul de asemănare dintre ele, se poate observa că valoarea indicelui cofenetic este relativ mică (0,9016) ceea ce detonă un grad mare de distorsiune între matricea originală de similratite şi matricea pe baza căreia s-a construit dendrograma.
Figura 7. Dendrograma reprezentând gruparea releveelor în funcţie de valoarea indicelui de similaritate Bray-Curtis.
O altă metodă de analiză cluster divizivă, care poate fi utilizată în studiile de vegetaţie,
este prin partitionarea în jurul medoidului (pam) care are avantajul stabilirii concomitente atât a numărului optim de grupe (clustere) corespunzătoare setului de date analizat, cât şi a componenţei fiecărui cluster în parte.
Numărul optim de grupe se stabilişte pe baza valorii lăţimii medii a „siluetelor”, numărul optim de clustere corespunzând variantei în care lăţimea medie a „siluetelor” are valoarea cea mai mare. Acest tip de analiză cluster se poate realiza cu ajutorul “bibliotecii” cluster versiunea 1.11.2. pentru R (Maechler 2006). În cazul celor 59 de relevee, numărul optim de grupe este trei (fig. 8).

58
Silhouette width si
0.0 0.2 0.4 0.6 0.8 1.0
Silhouette plot of pam(x = dist, k = 3, diss = T, stand = F)
Average silhouette width : 0.51
n = 59 3 clusters Cj
j : nj | avei∈Cj s
1 : 41 | 0.46
2 : 9 | 0.55
3 : 9 | 0.69
Figura 8. Reprezentarea grafică sub formă de “siluete” a rezultatului analizei cluster prin partiţionarea în jurul medoidului (pam)
Nomenclatura fitosociologică a pajiştilor analizate se va face în conformitate cu prevederile Codului International de Nomenclatură Fitosociologică (Weber et al. 2000) după sistemele propuse de către Coldea (1991) şi Pop et al. (2002).
3.4.4. Ordonarea vegetaţiei
Ordonarea datelor de vegetaţie este un termen colectiv pentru tehnicile multivariate
care au ca scop aranjarea eşantioanelor în funcţie de structura specifică a acestora (ter Braak 1995b), fiind un proces de reprezentare a relaţiilor existente între eşantioanele de vegetaţie, specii şi factorii de mediu în spaţiul dimensional redus al câtorva axe (în general primele 2-3 axe), într-un mod cât mai fidel posibil. Produsul final îl reprezintă un grafic (de obicei bidimensional), în care distanţa dintre puncte (specii, eşantioane) reflectă gradul de asemănare dintre acestea în cea ce priveşte structura sau cerinţele ecologice ale acestora (Gauch Jr. 1982). Din punct de vedere statistic, ordonarea este un proces de regresii şi calibrări succesive care se opresc atunci când poziţia relativă a speciilor sau releveelor nu mai poate fi imbunătăţită, axele fiind considerate variabile latente sau variabile de mediu ipotetice (ter Braak et Prentince 1988, ter Braak 1995a, b, ter Braak et Looman 1995, ter Braak et Šmilauer 2002b, Cristea et al. 2004). Poziţia relativă a unei specii sau a unui releveu poate fi descrisă ca fiind o funcţie a factorilor de mediu înregistraţi, în cazul analizei directe a gradienţilor, sau acestea (specii/relevee) pot fi poziţionate în lungul unor axe imaginare care pot fi interpretate ca şi gradienţi ecologici, în cazul analizei indirecte a gradienţilor.

59
În studiile de monitorizare vor fi utilizate două dintre cele mai uzuale metode de ordonare, şi anume: analiza „destinsă” a corespondenţelor (DCA) (Hill et Gauch Jr 1980) şi analiza canonică a corespondenţelor (CCA) (ter Braak 1986), amândouă fiind metode care se bazează pe răspunsul unimodal al speciilor în lungul gradienţilor ecologici. Analiza „destinsă” a corespondeţelor (DCA) este o metodă de ordonare indirectă care porneşte de la premisa că fiecare dintre axele extrase reprezintă un gradient ecologic care, de cele mai multe ori poate fi o combinaţie de mai mulţi factori de mediu. Lungimea axelor este exprimată în unităţi standard de deviaţie (SD) care sunt o măsură a β-diversităţii, a ratei de înlocuire a speciilor în lungul gradienţilor ecologici (Hill et Gauch Jr 1980, Digby et Kempton 1987). DCA realizează o ordonare a matricei relevee-specii bazându-se doar pe variaţia compoziţiei floristice dintre relevee, poziţia factorilor de mediu fiind ajustată ulterior. Analiza DCA este utilizată şi pentru determinarea tipului de răspuns al speciilor şi implicit pentru alegerea tipului de ordonare a datelor. Astfel, dacă lungimea gradientului celui mai mare are o valoare mai mare de 4 SD, atunci răspunsul speciilor este unul puternic unimodal (ter Braak et Šmilauer 2002a). Dacă cel mai lung gradient are o valoare sub 3SD, atunci alegerea unei metode liniare de analiză poate fi cea mai bună soluţie. Pentru gradienţi cu o lungime cuprinsă între 3 şi 4 SD atât metodele liniare cât şi cele unimodale vor avea rezultate asemănătoare (Lepš et Šmilauer 2003). În ciuda limitărilor evidenţiate de o serie de autori (Digby et Kempton 1987, Wartenberg et al. 1987, ter Braak et Prentince 1988, Kent et Coker 1992, Palmer 1993b, Legendre et Legendre 1998, ter Braak et Šmilauer 2002b) DCA continuă să fie o metodă, des utilizată, de analiză indirectă a gradienţilor, metodă care îşi dovedeşte utilitatea mai ales atunci când în analiza multivariată a vegetaţiei nu se dispune de valori ale unor factori ecologici (Whittaker 1987, Peet et al. 1988, Palmer 1993a, ter Braak et Šmilauer 2002b, Cristea et al. 2004); când se urmăreşte doar evidenţierea structurii vegetaţiei analizate (McCune 1997), dar şi ca metodă de validare a clasificărilor efectuate cu alte metode (ter Braak 1995b, van Tongeren 1995, Legendre et Legendre 1998, ter Braak et Šmilauer 2002b). Analiza DCA a vegetaţiei se poate realiza cu ajutorul „bibliotecii” vegan implentată în R versiunea 2.8.1 şi a programului CANOCO versiunea 4.5 (ter Braak et Šmilauer 2002a), având ca scop atât determinarea tipului de răspuns a speciilor (liniar sau unimodal) în vederea alegerii metodelor de analiză cât şi pentru evidenţierea structurii fitocenozelor şi validarea clasificării ierarhice a vegetaţiei. Analiza canonică a corespondenţelor (CCA) este una dintre cele mai răspândite metode de analiză multivariată a datelor ecologice. CCA este o metodă de analiză directă a gradienţilor care produce o ajustare liniară optimă a distribuţiei abundenţelor speciilor în funcţie de variabilele de mediu măsurate, axele extrase fiind combinaţii liniare ale factorilor ecologici care explică cel mai bine variaţia distribuţiei speciilor (ter Braak 1986, ter Braak et Prentince 1988, ter Braak 1995b, ter Braak et Šmilauer 2002b). Pentru analiza CCA datele de vegetaţie sunt grupate în două matrici, matricea relevee-specii şi matricea relevee-variabile de mediu. Una dintre condiţiile de baza ale analizei canonice a corespondenţelor este dimensiunea matricii relevee-variabile de mediu, numărul releveelor trebuind să fie mult mai mare decât numărul variabilelor de mediu (Cristea et al. 2004), în caz contrar analiza se reduce la o simplă analiză a corespondenţelor (Oksanen et al. 2008).

60
Analiza CCA a datelor de vegetaţie şi a factorilor de mediu luaţi în studiu se poate realiza cu ajutorul programului CANOCO 4.5 (ter Braak et Šmilauer 2002a) şi a “bibliotecii” vegan (Oksanen et al. 2008) pentru R. Pentru analiza CCA a fost aplicat principiul parcimoniei (Briciul lui Occam) conform căruia un model al oricărui sistem trebuie să fie cât mai simplu posibil prin eliminarea oricărei informaţii cu caracter redundant sau inutile (Crawley 2002, 2005). Din matricea relevee-specii vor fi eliminate speciile cu o prezenţă sub 2%, specii care pot distorsiona rezultatul ordonării. Gradul în care factorii ecologici rămaşi în studiu explică o parte din varianţa totală a setului de date va fi testat cu ajutorul testului F, probabilitatea fiind calculată printr-un test Monte Carlo cu un număr de 9999 permutări, pentru analizele ulterioare fiind aleşi doar factorii puternic corelaţi cu o probabilitate de transgresiune de p < 0.05. De asemenea, se va calcula valoarea factorului de inflaţie (VIF), care oferă informaţii despre existenţa fenomenului de colinearitate între factorii de mediu studiaţi. O valoare mare > 10 (Oksanen et al. 2008) sau > 20 (ter Braak et Šmilauer 2002b) indică faptul că unul sau mai mulţi dintre factorii analizaţi prezintă fenomenul de colinearitate. Variabilele de mediu rămase în studiu, se vor utiliza pentru a „construi” modelul care explică cel mai bine varianţa asociată primelor două axe CCA. Pentru aceasta, toate variabilele de mediu vor fi introduse pe rând, în analiza CCA, cel mai bun model fiind determinat pe baza valorii minime Akaike’s Information Criterion (AIC). Factorii ecologici, cu excepţia celor nominali (modul de utilizare al pajiştilor) sunt reprezentaţi grafic sub formă de vectori, a căror lungime este direct proporţională cu amplitudinea factorului respectiv, aceştia putând fi interpretaţi şi în conjuncţie cu speciile sau cu releveele. Interpretarea rezultatelor se poate face atât pe baza reprezentării grafice, cât şi be baza output-ului analizei CCA, care oferă date despre lungimea vectorilor, gardul de corelaţie al acestora cu variabilele dependente şi cu axele canonice, varianţa explicată de primele patru axe extrase, varianţa totală a setului de date şi coordonatele punctelor de proiecţie ale speciilor şi releveelor.
3.4.5. Analiza „arborilor” de regresie
„Arborii” de regresie reprezintă o metodă utilă de analiză a datelor ecologice complexe, reprezentând un complement sau o alternativă la o serie de metode statistice tradiţionale cum ar fi regresiile multiple, analiza varianţei, regresiile logistice, modelele log-lineare etc. Utilizarea acestei metode permite explicarea variaţei unei variabile dependente, prin divizări repetate a setului de date în mai multe grupuri omogene, pe baza combinaţiilor dintre variabilele independente, care pot fi nominale şi/sau numerice (De'Ath et Fabricius 2000, Lawrence et Ripple 2000, Crawley 2002). Această metodă de analiză a datelor ecologice se bazează pe divizarea recursivă binară a variabilelor independente în lungul unei axe de coordonate, astfel încât fiecare nod are valoarea maximă a puterii de discriminare a fiecărei variabile în cele două ramuri (Crawley 2002). Rolul variabilelor continue este determinat pe baza testelor ANOVA, în timp ce analiza variabilelor nominale nu este altceva decât o clasificare (Crawley 2002, Therneau et Atkinson 2006, Therneau et al. 2006). Pentru realizarea „arborilor” de regresie, dintre toate variabilele independente este selectată cea care explică cea mai mare proporţie din devianţă, devianţa fiind calculată pe baza unei valori prag a variabilei, această valoare determinând două valori medii a variabilei dependente pentru fiecare dintre cele două ramuri

61
rezultate. Ca valoare prag se alege valoare care determină cea mai mică abatere de la medie, fiind utilizată pentru divizarea datelor în două seturi. În cazul fiecărui subset se reia întregul algoritm de calcul, procesul contiunănd până nu mai există posibilitatea de a reduce valoarea devianţei. „Arborii” de regresie şi clasificare sunt reprezentaţi grafic, cu „rădăcina”, care reperezintă datele nedivizate, în partea superioară, de la care pornesc „ramurile” şi „frunzele”. Lungimea „ramurilor” oferă informaţii despre importanţa factorilor în explicarea varianţei unui set de date. Cu cât lungimea lor este mai mare, cu atât este mai mare şi importanţa lor. „Frunzele” reprezintă grupurile finale, care nu mai pot fi divizate. Fiecărui nod i se poate asocia informaţii privind devianţa, valoarea medie a variabilei dependente, valorile prag ale variabilelor independente, varianţa etc. Pe baza varianţei existente la nivelul „rădăcinii” şi a nodurilor terminale („frunzelor”) se poate calcula procentul din varianţă care este explicat de „arbore” (fig. 4). Una dintre principalele probleme cu care se confruntă aplicarea practică a acestei metode, este aceea a nivelului optim la care să se oprească divizările datelor. Astfel, „creşterea
arborilor” poate fi controlată atât prin stabilirea numărului minim de cazuri (eşantioane) care pot forma un grup sau a parametrului de complexitate, cât şi prin validări încrucişate, alegerea celui mai bun model făcându-se astfel încât, eroarea estimată asociată modelului să aibă o distribuţie în intervalul 1SE (eroarea standard a celei mai mici valori), aşa-numita regula 1-SE (De'Ath et Fabricius 2000, De'Ath 2002).
Deoarece axele extrase în urma ordonărilor directe sau indirecte, reprezintă, de cele mai multe ori, un gradient complex (o combinaţie de mai mulţi factori), utilizarea „arborilor” de regresie reprezintă o soluţie simplă pentru descifrarea rolului pe care fiecare variabilă ambientală o are în explicarea unei anumite structuri. Această metodă poate fi utilizată şi pentru a identifica variabilele de mediu care influenţează abundenţa-dominţa unei specii.

62
Figura 9. Analiza arborilor de regresie pentru identificarea principalilor factori care determină
variaţia numărului de specii/releveu (past_c – utilizarea pajiştilor ca păşuni pentru vite; past_cs – utilizarea pajiştilor ca păşuni mixte; past_s – utilizarea pajiştilor ca păşuni pentru oi; stbl –
pajişti târlite; medw – fâneţe; mdw_p – fâneţe păşunate primăvara şi toamna; n – numărul de relevee prezente în fiecare nod terminal; r2 – coeficientul de determinare) (Bărbos 2007)
Pentru descifrarea structurii gardienţilor asociaţi principalelor axe de ordonare, în
analiza „arborilor” de regresie vor fi folosite, ca variabile dependente valorile proiecţiei releveelor pe axe, iar ca variabile independente factorii ecologici luaţi în studiu. De asemenea, această metodă poate fi aplicată şi pentru a identifica principalii factori care influenţează acoperirea medie procentuală a speciilor edificatoare. În acest caz, pentru a identifica şi eventualul rol pe care relaţiile interspecifice îl pot avea în abundenţa-dominţa unei specii, pe lângă factorii de mediu vor mai fi introduse ca variabile independente şi speciile edificatoare. Analiza „arborilor” de regresie s-e poate efectua cu ajutorul „bibliotecilor” rpart versiunea 3.1-29 (Therneau et Atkinson 2006), tree versiunea 1.0-24 (Ripley 2006), party versiunea 0.9-995 (Hothorn et al. 2009) şi mvpart versiunea 1.2-4 (Therneau et al. 2006) pentru R.
3.4.6. Răspunsul speciilor şi gradul de suprapunere al nişelor
Variaţia abundenţei speciilor în spaţiu şi timp poate fi privită ca fiind reacţia speciilor respective la variaţiile factorilor ecologici care condiţionează existenţa lor. Unul dintre obiectivele oamenilor de ştiinţă a fost şi acela de a găsi un model care să aproximeze cât mai bine posibil modalitatea în care o specie oarecare răspunde la acţiunea unor factori de mediu. În analiza răspunsului speciilor vegetale, în mod obişnuit, se iau în studiu valorile de acoperire procentuală sau frecvenţă ale speciilor respective, indici populaţionali care impun şi anumite
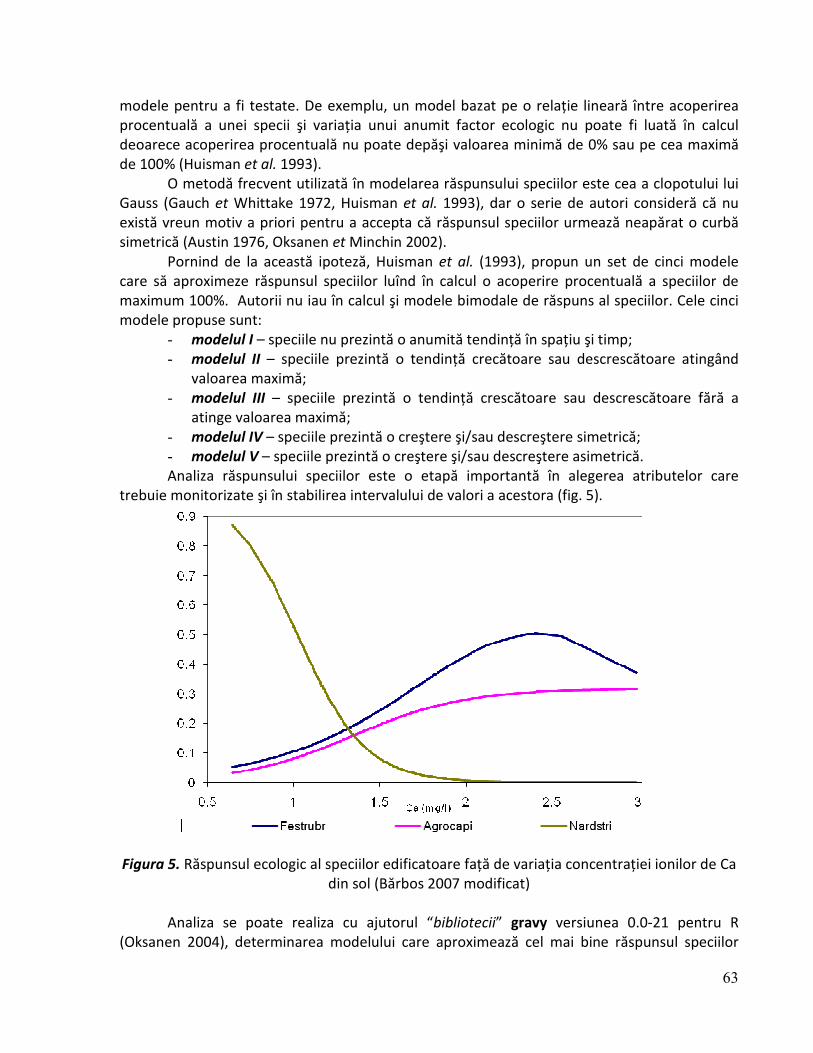
63
modele pentru a fi testate. De exemplu, un model bazat pe o relaţie lineară între acoperirea procentuală a unei specii şi variaţia unui anumit factor ecologic nu poate fi luată în calcul deoarece acoperirea procentuală nu poate depăşi valoarea minimă de 0% sau pe cea maximă de 100% (Huisman et al. 1993). O metodă frecvent utilizată în modelarea răspunsului speciilor este cea a clopotului lui Gauss (Gauch et Whittake 1972, Huisman et al. 1993), dar o serie de autori consideră că nu există vreun motiv a priori pentru a accepta că răspunsul speciilor urmează neapărat o curbă simetrică (Austin 1976, Oksanen et Minchin 2002). Pornind de la această ipoteză, Huisman et al. (1993), propun un set de cinci modele care să aproximeze răspunsul speciilor luînd în calcul o acoperire procentuală a speciilor de maximum 100%. Autorii nu iau în calcul şi modele bimodale de răspuns al speciilor. Cele cinci modele propuse sunt:
- modelul I – speciile nu prezintă o anumită tendinţă în spaţiu şi timp; - modelul II – speciile prezintă o tendinţă crecătoare sau descrescătoare atingând
valoarea maximă; - modelul III – speciile prezintă o tendinţă crescătoare sau descrescătoare fără a
atinge valoarea maximă; - modelul IV – speciile prezintă o creştere şi/sau descreştere simetrică; - modelul V – speciile prezintă o creştere şi/sau descreştere asimetrică. Analiza răspunsului speciilor este o etapă importantă în alegerea atributelor care
trebuie monitorizate şi în stabilirea intervalului de valori a acestora (fig. 5).
Figura 5. Răspunsul ecologic al speciilor edificatoare faţă de variaţia concentraţiei ionilor de Ca din sol (Bărbos 2007 modificat)
Analiza se poate realiza cu ajutorul “bibliotecii” gravy versiunea 0.0-21 pentru R
(Oksanen 2004), determinarea modelului care aproximează cel mai bine răspunsul speciilor

64
făcându-se pe baza valorii BIC (Bayesian Criterion Information) la o probabilitate de transgresiune p = 0,05. În cazul modelelor IV şi V se va determina şi valoarea optimă a variabilei de mediu care determină o amplitudine maximă a răspunsului speciilor faţă de acea variabilă.
3.5. Elaborarea rapoartelor de monitorizare
În funcţie de scopul urmărit, raportul de monitorizare, aşa cum am arătat şi în primul capitol trebuie să cuprindă următoarele aspecte:
- importanţa realizării studiului de monitorizare; - prezentarea obiectivelor propuse; - prezentarea metodelor de lucru utilizate pentru colectarea şi analiza datelor
precum şi motivarea alegeri acestor metode de lucru; - prezentarea rezultatelor obţinute; - discutarea rezultatelor şi a implicaţiilor practice ale acestora; - concluziile privind importanţa studiului şi elaborarea de recomandări pentru viitor; - sursele bibliografice care au fost consultate.
Rapoartele de monitorizare trebuie să fie clare, concise şi să folosească un limbaj
accesibil. În elaborarea rapoartelor de monitorizare se va ţine cont de faptul că acestea vor reprezenta argumentele ştiinţifice pe baza cărora, factorii de decizie, vor lua deciziile adecvate privind măsurile de management necesare pentru conservarea habitatului.

65
CONCLUZII Din analiza datelor prezentate dorim să subliniem câteva aspecte cu carcater general dar deosebit de importante pentru succesul programelor de monitorizare, şi anume: - monitorizarea habitatelor, în toate etapele realizării ei (design, colectarea datelor, analiza datelor şi interpretarea rezultatelor etc) este o activitate care necesită o bună cunoaştere atât a problemelor teoretice cât şi a metodologiei de lucru; - pentru elaborarea planurilor de monitorizare este recomandată realizarea unor studii preliminare pentru detreminarea variabilităţii habitatului, a numărului minim de eşantioane, suprafaţa minimă a eşantioanelor etc.; - definirea obiectivelor trebuie să ţină cont, pe lângă caracteristicile şi particularităţile fiecărui habitat şi a fiecărei staţiuni, şi de resursele materiale, umane şi financiare; - alegerea variabilelor de măsurat trebuie făcută concomitent cu alegerea metodelor de prelucrare a datelor permiţând astfel alegerea optimă a metodelor de colectare; - în planurile de monitorizare trebuie să fie incluse şi variabile care, chiar dacă nu influenţează în mod evident starea actuală a habitatului, este posibil ca pe viitor rolul lor să crească; - alegerea metodelor de colectare a datelor pe teren trebuie făcută astfel încât să se minimalizeze probabilitatea apariţiei erorilor care pot compromite întregul proces de monitorizare; - monitorizarea nu presupune doar înregistrarea periodică a unor variabile ci, mai mult decât atât, reprezintă un proces complex de colectare şi analiză a datelor, interpretare a rezultatelor şi integrarea acestora într-un proces decizional; - programele de monitorizare oferă argumentele ştiinţifice necesare luării deciziilor privind măsurile de management necesare pentru conservarea habitatelor, decizii care sunt influenţate de factori politici, economici şi sociali.

66
BIBLIOGRAFIE
Aerts, R., de Caluwe H., Beltman B. 2003. Plant community mediated vs. nutritional controls on litter decomposition rates in grasslands. Ecology 84:3198-3208.
Aoki, I. 1995. Diversity and rank-abundance relationship concerning biotic compartments. Ecological Modelling 82:21-26.
Artiola, J. F., Pepper I. L., Brusseau M. L., editors. 2004. Environmental monitoring and characterization. Elsevier Academic Press, Burlington, San Diego, London.
Atkin, O. K., Cummins W. R. 1994. The effect of nitrogen source on growth, nitrogen economy and respiration of two high arctic plant species differing in relative growth rate. Functional Ecology 8:389-399.
Austin, M. P. 1976. On non-linear species response models in ordination. Vegetatio 33:33-41.
Balsberg-Pahlsson, A. M. 1995. Growth, radicle and root hair development of Deschampsia flexuosa (L) Trin seedlings in relation to soil acidity. Plant and Soil 175:125-132.
Bărbos, M. I. 2007. Studii privind cenologia şi ecologia pajiştilor montane din judeţul Maramureş. Universitatea "Babeş-Bolyai", Cluj Napoca.
Berendse, F., Lammerts E. J., Olff H. 1998. Soil organic matter accumulation and its implications for nitrogen mineralization and plant species composition during succesion in coastal dune slacks. Plant Ecology 137:71-78.
Blaga, G., Filipov F., Udrescu S., Rusu I., Vasile D. 2005. Pedologie. Editura AcademicPres, Cluj Napoca.
Borhidi, A. 1995. Social behaviour types, the naturalness and relative ecological indicator values of the higher plants in the Hungarian Flora. Acta Botanica Hungarica 39:97-181.
Borza, A., Boşcaiu N. 1965. Introducere în studiul covorului vegetal. Editura Academiei R.P.R., Bucureşti.

67
Bosy, J. L. et Reader R. J. 1995. Mechanisms underlying the suppression of forb seedling emergence by grass ( Poa pratensis ) litter. Functional Ecology 9:635-639.
Botnariuc, N., Vădineanu A. 1982. Ecologie. Editura Didactică şi Pedagogică, Bucureşti.
Bradshaw, A. D., Lodge R. W., Jowett D., Chadwick M. J. 1958. Experimental Investigations Into the Mineral-Nutrition of Several Grass Species .1. Calcium Level. Journal of Ecology 46:749-757.
Braun-Blanquet, J. 1932. Plant sociology; the study of plant communities. McGraw-Hill Book Company, Inc.
Carson, W. P., Peterson C. J. 1990. The role of litter in an old-field community - impact of litter quantity in different seasons on plant-species richness and abundance. Oecologia 85:8-13.
Chiriţă, C. D. 1955. Pedologie generală. Editura Agro-Silvică de Stat, Bucuresti.
Chiriţă, C. D. 1974. Ecopedologie cu baze de pedologie generală. Editura Ceres, Bucureşti.
Christensen, N. L., Bartuska A. M., Brown J. H., Carpenter S., D'Antonio C., Francis R., Franklin J. F., MacMahon J. A., Noss R. F., Parsons D. J., Peterson C. H., Turner M. G., Woodmansee R. G. 1996. The report of the Ecological Society of America Committee on scientific basis for ecosystem management. Ecological Applications 6:665-691.
Ciocârlan, V. 1988. Flora ilustrată a României. Editura Ceres, Bucuresti.
Ciocârlan, V. 1990. Flora ilustrată a României. Editura Ceres, Bucureşti.
Ciocârlan, V. 2000. Flora ilustrată a României. Pteridophyta et Spermatophyta. Editura Ceres, Bucureşti.
Coldea, G. 1990. Muntii Rodnei. Studiu geobotanic. Editura Academiei Romane, Bucuresti.
Coldea, G. 1991. Prodrome des associations vegetales des Carpates de sud-est (Carpates Roumaines). Documents Phytosociologiques 13:317-539.
Common, T. G., Hunter E. A., Floate M. J. S., Eadie J., Hodgson J. 1991. The long-term effects of a range of pasture treatments applied to three semi-natural hill grassland communities. 1. Pasture production and botanical composition. Grass and Forage Science 46:239-251.
Cornish, P. S., Raison R. J. 1977. Effects of phosphorus and plants on nitrogen mineralization in three grassland soils. Plant and Soil 47:289-295.

68
Crawley, M. J. 2002. Statistical computing. An introduction to data analysis using S-Plus. John Wiley & Sons, Ltd., Chichester.
Crawley, M. J. 2005. Statistics. An introduction using R. John Wiley & Sons, Ltd, Chichester.
Cristea, V. 1991. Fitocenologie şi vegetaţia României; Îndrumător de lucrări practice. Universitatea Babeş-Bolyai, Cluj Napoca.
Cristea, V. 1993. Fitosociologie şi vegetaţia României. Universitatea Babeş-Bolyai, Cluj Napoca.
Cristea, V., Gafta D., Pedrotti F. 2004. Fitosociologie. Editura Presa Universitară Clujeană, Cluj Napoca.
De'Ath, G. 2002. Multivariate regression trees: a new technique for modeling species-environment relationships. Ecology 83:1105-1117.
De'Ath, G., Fabricius K. E. 2000. Classification and regression trees: A powerful yet simple technique for ecological data analysis. Ecology 81:3178-3192.
Digby, P. G. N., Kempton R. A. 1987. Multivariate analysis of ecological communities. Chapman and Hall.
Dufrêne, M., Legendre P. 1997. Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs 67:345-366.
Eckstein, R. L., Donath T. W. 2005. Interactions between litter and water availability affect seedling emergence in four familial pairs of floodplain species. Journal of Ecology 93:807-816.
Elzinga, C. L., Salzer D. W., Willoughby J. W. 1998. Mesuring & monitoring plant populations. BLM/RS/ST-98/005+1730, Bureau of Land Management.
European Commission, 1992, Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and wild fauna and flora
European Commission, 2007, Interpretation Manual of European Union Habitats - EUR 27, DG Environment - Nature and Biodiversity
Everitt, B. S. 1993. Cluster analysis. Cambridge University Press.
Facelli, J. M., Carson W. P. 1991. Heterogeneity of plant litter accumulation in successional communities. Bulletin of the Torrey Botanical Club 118:62-66.

69
Facelli, J. M., Pickett S. T. A. 1991. Plant litter - light interception and effects on an old-field plant community. Ecology 72:1024-1031.
Fageria, N. K., Baligar V. C., Wright R. J. 1989. Growth and nutrient concentrations of alfalfa and common bean as influenced by soil acidity. Plant and Soil 119:331-333.
Foster, B. L., Gross K. L. 1998. Species richness in a successional grassland: effects of nitrogen enrichment and plant litter. Ecology 79:2593-2602.
Fraley, C., Raftery A. E. 2006. mclust: Model-based cluster analysis. R package version 2.1-12. Department of Statistics and University of Washington.
Gafta, D., Owen M., editors. 2008. Manual de interpretare a habitatelor Natura 2000 din România. Editura Risoprint, Cluj Napoca.
Gauch, H. G., Whittake R. H. 1972. Coenocline simulation. Ecology 53:446-&.
Gauch Jr, H. G. 1982. Multivariate analysis in community ecology. Cambridge University Press, Cambridge, London, New York, Melbourne, Sydney.
Gigon, A., Rorison I. H. 1972. Response of some ecologically distinct plant species to nitrate-nitrogen and to ammonium-nitrogen. Journal of Ecology 60:93-&.
Gilbert, N., Wells T. C. E. 1966. Analysis of Quadrat Data. Journal of Ecology 54:675-&.
Goodall, D. W. 1973. Sample similarity and species correlation. Pages 105-156 in R. H. Whittaker, editor. Ordination and classification of communities. Dr. W. Junk, The Hague.
Grant, S. A., Torvell L., Sim E. M., Small J. L., Armstrong R. H. 1996. Controlled grazing studies on Nardus stricta grassland: effects of between-tussock sward height and species of grazer on Nardus utilization and floristic composition in two fields in Scotland. Journal of Applied Ecology 33:1053-1064.
Greig-Smith, P. 1964. Quantitative plant ecology. Butterworths Publications, London.
Greszta, J., Gruszka A., Wachalewski T. 1992. Humus degradation under the influence of simulated "acid rain". Water, Air, and Soil Pollution 63:51-66.
Grime, J. P. 2001. Plant strategies, vegetation processes, and ecosystem properties. John Wiley & Sons, Ltd.

70
Grime, J. P., Hodgson J. G., Hunt R. 1996. Comparative plant ecology. A functional approach to common British species. Chapman and Hall, London.
Gusewell, S. 2004. N : P ratios in terrestrial plants: variation and functional significance. New Phytologist 164:243-266.
Gusewell, S. 2005. High nitrogen: phosphorus ratios reduce nutrient retention and second-year growth of wetland sedges. New Phytologist 166:537-550.
Gusewell, S., Koerselman M. 2002. Variation in nitrogen and phosphorus concentrations of wetland plants. Perspectives in Plant Ecology Evolution and Systematics 5:37-61.
Hector, A., Beale A. J., Minns A., Otway S. J., Lawton J. H. 2000. Consequences of the reduction of plant diversity for litter decomposition: effects through litter quality and microenvironment. Oikos 90:357-371.
Hejcman, M., Klaudisova M., Schellberg J., Honsova D. 2007. The regen grassland experiment: Plant species composition after 64 years of fertilizer application Agriculture Ecosystems & Environment 122:259-266.
Hennekens, S. M. 1998-2009. Turboveg for Windows.International single user version.
Hill, D., Fasham M., Tucker G., Shewry M., Shaw D., editors. 2005. Handbook of biodiversity methods. Survey, evaluation and monitoring. Cambridge University Press, New York.
Hill, M. O. 1973. Diversity and evenness - unifying notation and its consequences. Ecology 54:427-432.
Hill, M. O. et Gauch Jr H. G. 1980. Detrended correspondence analysis: an improved ordination technique. Vegetatio 42:47-58.
Hoorens, B., Aerts R., Stroetenga M. 2002. Litter quality and interactive effects in litter mixtures: more negative interactions under elevated CO2? Journal of Ecology 90:1009-1016.
Hoorens, B., Aerts R., Stroetenga M. 2003a. Does initial litter chemistry explain litter mixture effects on decomposition? Oecologia 137:578-586.
Hoorens, B., Aerts R., Stroetenga M. 2003b. Is there a trade-off between the plant's growth response to elevated CO2 and subsequent litter decomposability? Oikos 103:17-30.

71
Hopkins, D. W., Ibrahim D. M., Odonnell A. G., Shiel R. S. 1990. Decomposition of cellulose, soil organic-matter and plant litter in a temperate grassland soil. Plant and Soil 124:79-85.
Hothorn, T., Hornik K., Strobl C., Zeileis A. 2009. party: A laboratory for Recursive Partytioning.
Huisman, J., Olaff H., Fresco L. F. M. 1993. A hierarchical set of models for species response analysis. Journal of Vegetation Science 4:37-46.
Hurlbert, S. H. 1971. Nonconcept of species diversity - critique and alternative parameters. Ecology 52:577.
Ivan, D. 1979. Fitocenologie si vegetatia RSR. Editura Didactica si Pedagogica, Bucuresti.
Jackson, D. A., Somers K. M., Harvey H. H. 1989. Similarity coefficients: measures of cooccurence and association or simply measures of occurence. The American Naturalist 133:436-453.
Kelly, D. 1989. Demography of short-lived plants in chalk grassland .2. Control of mortality and fecundity. Journal of Ecology 77:770-784.
Kent, M., Coker P. 1992. Vegetation description and analysis. A practical approach. Belhaven Press, London.
Krebs, C. J. 1998. Ecological methodology. Addison Wesley Longman, Inc., Menlo Park.
Krebs, C. J. et Kenney A. J. 2000. Ecological methodology.
Lawrence, R. L., Ripple W. J. 2000. Fifteen years of revegetation of Mount St. Helens: A landscape-scale analysis. Ecology 81:2742-2752.
Legendre, P., Legendre L. 1998. Numerical ecology. Elsevier Science, Amsterdam.
Leps, J., Smilauer P. 2003. Multivariate analysis of ecological data using Canoco. Cambridge University Press.
Maechler, M. 2006. cluster: Cluster analysis.
Magid, J. 1993. Vegetation effects on phosphorus fractions in set-aside soils. Plant and Soil 149:111-119.

72
Magurran, A. E. 2004. Measuring biological diversity. Blackwell Science Ltd, Oxford.
Mamolos, A. P., Vasilikos C. V., Veresoglou D. S. 2005. Vegetation in contrasting soil water sites of upland herbaceous grasslands and N : P ratios as indicators of nutrient limitation. Plant and Soil 270:355-369.
Maret, M. P., Wilson M. V. 2005. Fire and litter effects on seedling establishment in western Oregon upland prairies. Restoration Ecology 13:562-568.
McAlpine, K. G., Drake D. R. 2003. The effects of small-scale environmental heterogeneity on seed germination in experimental treefall gaps in New Zealand. Plant Ecology 165:207-215.
McCune, B. 1997. Influence of noisy environmental data on canonical correspondence analysis. Ecology 78:2617-2623.
Monk, C. D., Gabrielson F. C. 1985. Effects of shade, litter and root competition on old-field vegetation in South-Carolina. Bulletin of the Torrey Botanical Club 112:383-392.
Moravec, J. 1973. The determination of the minimal area of phytocoenosis. Folia Geobotanica & Phytotaxonomica 8:23-47.
Mouillot, D., Lepretre A. 2000. Introduction of relative abundance distribution (RAD) indices, estimated from the rank-frequency diagrams (RFD), to assess changes in community diversity. Environmental Monitoring and Assessment 63:279-295.
Nakamura, T. 1996. Ecological relationships between seedling emergence and litter cover in the earliest stage of plant succession on sandy soil. Ecological Research 11:105-110.
Näsholm, T., Huss-Danell K., Högberg P. 2000. Uptake of organic nitrogen in the field by four agriculturally important plant species. Ecology 81:1155-1161.
Niinemets, U., Kull K. 2005. Co-limitation of plant primary productivity by nitrogen and phosphorus in a species-rich wooded meadow on calcareous soils. Acta Oecologica-International Journal of Ecology 28:345-356.
Oksanen, J. 2004. gravy: Gradient Analysis of Vegetation. R package version 0.0-21.
Oksanen, J., Kindt R., Legendre P., O'Hara B., Simpson G. L., Solymos P., Stevens M. H. H., Wagner H. 2008. vegan: Community Ecology Package.

73
Oksanen, J., Minchin P. R. 2002. Continuum theory revisited: what shape are species responses along ecological gradients? Ecological Modelling 157:119-129.
Palmer, M. W. 1993a. Putting Things in Even Better Order - the Advantages of Canonical Correspondence-Analysis. Ecology 74:2215-2230.
Palmer, M. W. 1993b. Putting things in even better order: the advantages of canonical correspondence analysis. Ecology 74:2215-2230.
Parker, A. J. 1982. The topographic relative moisture index: an approach to moisture assessment in mountain terrain. Physical Geography 3:160-168.
Parton, W. J., Stewart J. W. B., Cole C. V. 1988. Dynamics of C, N, P and S in grassland soils - a model. Biogeochemistry 5:109-131.
Peet, R. K. 1974. The measurementof species diversity. Annual Review of Ecology and Systematics 5:285-307.
Peet, R. K., Knox R. G., Case J. S., Allen R. B. 1988. Putting things in order: the advantages of detrended correspondence analysis. The American Naturalist 131:924-934.
Podani, J. 2001. SYN-TAX 2000. Computer program for data analysis in ecology and systematics. User's manual. Scientia Publishing, Budapest.
Pop, I. 1977. Biogeografie ecologică. Editura Dacia, Cluj Napoca.
Pop, I. 1978. Flora si vegetatia Muntilor Zarandului. Contribuţii Botanice 18:7-214.
Pop, I. 1979. Biogeografie ecologică. Editura Dacia, Cluj Napoca.
Pop, I. 1982. Plante spontane si subspontane cu valoare economica din flora RSR. Contribuţii Botanice XXII:131-142.
Pop, I., Cristea V., Hodisan I. 2002. Vegetatia judetului Cluj (Studiu fitocenologic, ecologic, bioeconomic si eco-protectiv). Contribuţii Botanice 35:5-254.
Popma, J., Mucina L., van Tongeren O. F. R., van der Maarel E. 1983. On the determination of optimal levels in phytosociological classification. Vegetatio 52:65-75.
Purchase, B. S. 1974. Influence of phosphate deficiency on nitrification. Plant and Soil 41:541-547.

74
Quested, H., Eriksson O. 2006. Litter species composition influences the performance of seedlings of grassland herbs. Functional Ecology 20:522-532.
R Development Core Team, 2008, R: A language and environment for statistical computing, version 2.8.1, Vienna, R Foundation for Statistical Computing
Raubuch, M., Beese F. 2005. Influence of soil acidity on depth gradients of microbial biomass in beech fores soils. European Journal of Forest Research 124:87-93.
Ripley, B. 2006. tree. Classification and regression trees, version 1.0-24.
Rotundo, J. L., Aguiar M. R. 2005. Litter effects on plant regeneration in arid lands: a complex balance between seed retention, seed longevity and soil-seed contact. Journal of Ecology 93:829-838.
Sanda, V. 2002. Vademecum ceno-structural privind covorul vegetal din România. Editura Vergiliu, Bucureşti.
Sanda, V., Biţă-Nicolae C. D., Barabaş N. 2003. Flora cormofitelor spontane şi cultivate din România. Editura Ion Borcea, Bacău.
Sanda, V., Popescu A., Barabaş N. 1998. Cenotaxonomia şi caracterizarea gruparilor vegetale din România. Studii si Comunicari - Biologie Vegetala 14:9-366.
Sanda, V., Popescu A., Stancu D. I. 2001. Structura cenotică şi caracterizarea ecologică a fitocenozelor din România. Editura Conphis, Piteşti.
Săvulescu, T., editor. 1952-1976. Flora R.P.R. - R.S.R. Editura Academiei, Bucureşti.
Schellberg, J., Moseler B. M., Kuhbauch W., Rademacher I. F. 1999. Long-term effect of fertilizer on soil nutrient concentration, yeld, forage quality and floristic composition of a hay meadow in the Eifel mountains, Germany Grass and Forage Science 54:195-207.
Smith, B., Wilson J. B. 1996. A consumer's guide to evenness indices. Oikos 76:70-82.
Soons, M. B., Heil G. W., Nathan R., Katul G. G. 2004. Determinants of long-distance seed dispersal by wind in grasslands. Ecology 85:3056-3068.
Spellerberg, I. 2005. Monitoring ecological change. Second edition. Cambridge University Press, Cambridge.

75
Stewart, J. W. B., Cole C. V. 1989. Influences of elemental interactions and pedogenic processes in organic-matter dynamics. Plant and Soil 115:199-209.
ter Braak, C. J. F. 1986. Canonical correspondence analysis - A new eigenvector technique for multivariate direct gradient analysis. Ecology 67:1167-1179.
ter Braak, C. J. F. 1995a. Calibration. Pages 78-90 in R. H. G. Jongman, C. J. F. ter Braak, and O. F. R. van Tongeren, editors. Data analysis in community and landscape ecology. Cambridge University Press.
ter Braak, C. J. F. 1995b. Ordination. Pages 91-173 in R. H. G. Jongman, C. J. F. ter Braak, and O. F. R. van Tongeren, editors. Data analysis in community and landscape ecology. Cambridge University Press.
ter Braak, C. J. F., Looman C. W. N. 1995. Regression. Pages 29-77 in R. H. G. Jongman, C. J. F. ter Braak, and O. F. R. van Tongeren, editors. Data analysis in community and landscape ecology. Cambridge University Press.
ter Braak, C. J. F., Prentince I. C. 1988. A theory of gradient analysis. Advances in Ecological Research 18:271-317.
ter Braak, C. J. F., Šmilauer P. 2002a. Canoco. Biometris - Plant Research International, Wageningen.
ter Braak, C. J. F., Šmilauer P. 2002b. CANOCO Reference Manual and CanoDraw for Windows User's guide. Software for canonical community ordination (version 4.5). Microcomputer Power, Ithaca, New York, USA.
Therneau, T. M., Atkinson B. 2006. rpart: Recursive Partitioning. R package version 3.1-29.
Therneau, T. M., Atkinson B., Ripley B., Oksanen J., De'Ath G. 2006. mvpart. Multivariate regression trees, version 1.2-4.
Tóthmérész, B. 1995. Comparison of different methods for diversity ordering. Journal of Vegetation Science 6:283-290.
Tutin, T. G., Burges N. A., Chater A. O., Edmonson J. R., Heywood V. H., Moore D. M., Valentine D. H., Walters S. M., Webb D. A. 1993. Flora Europaea. Second edition. Cambridge University Press.

76
Tutin, T. G., Heywood V. H., Burges N. A., Moore D. M., Valentine D. H., Walters S. M., Webb D. A. 1968. Flora Europaea. Cambridge University Press.
Tutin, T. G., Heywood V. H., Burges N. A., Moore D. M., Valentine D. H., Walters S. M., Webb D. A. 1972. Flora Europaea. Cambridge University Press.
Tutin, T. G., Heywood V. H., Burges N. A., Moore D. M., Valentine D. H., Walters S. M., Webb D. A. 1976. Flora Europaea. Cambridge University Press.
Tutin, T. G., Heywood V. H., Burges N. A., Moore D. M., Valentine D. H., Walters S. M., Webb D. A. 1980. Flora Europaea. Cambridge University Press.
Tyler, G. et Olsson T. 2001. Plant uptake of major and minor mineral elements as influenced by soil acidity and liming. Plant and Soil 230:307-321.
van Tongeren, O. F. R. 1995. Cluster analysis. Pages 174-212 in R. H. G. Jongman, C. J. F. ter Braak, and O. F. R. van Tongeren, editors. Data analysis in community and landscape ecology. Cambridge University Press, Cambridge.
Walsh, R. P. D., Voigt P. J. 1977. Vegetation litter - underestimated variable in hydrology and geomorphology. Journal of Biogeography 4:253-274.
Wartenberg, D., Ferson S., Rohlf J. F. 1987. Putting things in order: a critique of detrended correspondence analysis. The American Naturalist 129:434-448.
Weaver, J. E., Clements F. E. 1938. Plant ecology. McGraw-Hill Book Company, Inc., New York, London.
Weber, H. E., Moravec J., Theurillat J. P. 2000. International Code of phytosociological nomenclature. 3rd edition. Journal of Vegetation Science 11:739-768.
Whittaker, R. J. 1987. An application of detrended correspondence analysis and non-metric multidimensional scaling to the identification and analysis of environmental factor complex and vegetation structures. The Journal of Ecology 75:363-376.
Williams, B. L., Shand C. A., Sellers S., Young M. E. 1999. Impact of synthetic sheep urine on N and P in two pastures in the Scottish uplands. Plant and Soil 214:93-103.
Wilson, J. B. 1991. Methods for fitting dominance diversity curves. Journal of Vegetation Science 2:35-46.

77
Witmer, S. 1999. Statistics for life sciences. Prentice Hall.
Xiong, S. J., Johansson M. E., Hughes F. M. R., Hayes A., Richards K. S., Nilsson C. 2003. Interactive effects of soil moisture, vegetation canopy, plant litter and seed addition on plant diversity in a wetland community. Journal of Ecology 91:976-986.
Xiong, S. J., Nilsson C. 1999. The effects of plant litter on vegetation: a meta-analysis. Journal of Ecology 87:984-994.
Zar, J. H. 1999. Biostatistical analysis. Prentice-Hall International, Inc.

78
ANEXA I Data………………..…. Cod releveu.......………......……….
Cod foto...................... Cod probă de sol............................
FIŞĂ PENTRU DESCRIEREA VEGETAłIEI Observator: AsociaŃia…………………………..............................………………………………..S……...………m2
I. Aşezare: jud…………………………....…localitatea……..............….......……………..................................
punct (toponimia)…............................................................................................................................ latitudine N.................................longitudine E........................... cod hartă........................................ II. Factori de mediu:
FACTORI TOPOGRAFICI Forma relief Forma pantei
1. vale 1. concavă 2. versant 2. concavă/dreaptă 3. creastă 3. dreaptă 4. platou 4. convex/dreaptă
PoziŃia pe pantă 5. convexă 1 baza pantei Pantă 2 pantă inferioară ExpoziŃie 3 pantă mijlocie Altitudine 4 pantă superioară Obs:
5 vîrful pantei FACTORI EDAFICI
Schelet (%) Schelet (tip)
0. fără schelet pietriş mărunt ( 2-5 mm) 1. slab scheletice < 5% pietriş mijlociu (5-10 mm) 2. moderat scheletice 5-25% pietriş mare (10-20 mm) 3. semischeletice 25-50% pietre mici (20-50 mm) 4. scheletice 50-75% pietre mijlocii (50-100 mm) 5. excesiv scheletice >75% pietre mari (100-200 mm) 6. excesiv scheletic de sol stâncos bolovani sau blocuri ( >200 mm)
Textura 1. nisip pH 2. nisip lutos Obs: 3. lut 4. lut argilos 5. argilă lutoasă 6. argilă 7. lut prăfos 8. argilă prăfoasă
FACTORI BIOTICI Acoperire generală(%) Acoperire muşchi (%)
H max vegetaŃie (cm) H max edificatori (cm)
Grosimea stratului de litieră (mm) Intensitatea exploatării
Managementul ariei Abandon Subpăşunat Moderat Suprapăşunat
1 fâneaŃă 2 fâneaŃă păşunată 3 păşune vite 4 păşune vite şi oi 5 păşune oi 6 păşune târlită Număr animale/ha Nr zile de păşunat Începutul perioadei de păşunat Sfârşitul perioadei de păşunat
Stratificare I II III IV V VI VII VIII IX X
ÎnălŃime (cm) 0 – 5 5-25 25-50 50-100 1-2 m 2-4 m 4-8 m 8-16 m 16-32 m > 32 m
Acoperire (%)

79
Nr. crt
Specia Stratul AD2 Faze şi
subfaze fenologice
Densitatea (nr/m2)
min max medie
1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. 27. 28. 29. 30. 31. 32. 33. 34. 35. 36. 37. 38. 39. 40. 41. 42. 43. 44. 45. 46.
2 Valori AD: r-0.1%; +-0.5%; 1-5.0%; 2-17.5%; 3-37.5%; 4-62.5%; 5-87.5%


















