
RAPORT ŞTIINŢIFIC Ş - automation.ucv.ro · pentru analiza problemelor biologice şi permite...
18
RAPORT ŞTIINŢIFIC ŞI TEHNIC Cuprins 1. Obiective generale 2. Obiectivele fazei de execuţie 3. Rezumatul fazei 4. Descrierea ştiinţifică 4.1. Descrierea tumorilor canceroase. Noţiuni generale. 4.2. Modelarea matematică a creşterii tumorilor nevascularizate 4.3. Cancerul gastric 4.4. Bibliografie 4.5. Analiza imagistică. Noţiuni generale 4.6. Bibliografie 4.7. Utilizarea imaginilor în diagnoză 4.8. Implementare program 4.9. Bibliografie 1. Obiective generale Elaborarea de modele conceptuale şi teorii. 2. Obiectivele fazei de execuţie Dezvoltarea de modele conceptuale. Studii cu privire la posibila utilizare a acestora în predicţia evoluţiei tumorilor canceroase. Identificarea formaţiunilor (celulelor) prin analiza automată a imaginilor. 3. Rezumatul fazei Modelarea matematică şi simularea sunt metode deosebit de eficiente, frecvent utilizate in toate domeniile. În ultimii ani s-au facut paşi concreţi în direcţia implicării teoriei sistemelor şi in studiul organismelor vii. Complexitatea sistemelor vii, chiar şi în cazul organismelor inferioare, este net superioară structurilor tehnice create de om. Acestea sunt constituite dintr-un mare număr de subsisteme, între care sunt stabilite multiple interconexiuni. Izolarea unui subsistem biologic, pentru studiul său, se face, inevitabil, cu preţul alterării propriei funcţii, cât şi a comportamentului sistemului în ansamblul său. Din acest motiv, datele obţinute experimental sunt diferite faţă de cele corespunzatoare unei evoluţii în condiţii normale. În concluzie, se recomandă studiul, pe cât posibil, al sistemului, în întregul său. Din punct de vedere sistemic, procesele ce se desfăşoară la nivel celular (inclusiv tumorile canceroase) au un pronunţat caracter neliniar. Deşi, în ceea ce priveşte studiul şi modelarea acestora s-au făcut progrese remarcabile, totuşi dezvoltarea şi aplicarea unor metode moderne este mai lentă în comparaţie cu alte domenii. Această întârziere este cauzată în principal de două caracteristici specifice ale bioproceselor. În cadrul acestei faze au fost trecute în revistă mai multe metode cu ajutorul cărora se pot simula evoluţiile populaţiilor de celule in diverse condiţii. De asemenea un accent deosebit s-a pus pe posibilitatea diagnosticării folosind prelucrarea automată a imaginilor. În acest sens sunt prezentate exemple de prelucrare a frotiurilor cu probe hematologice (leucemie limfatică acută şi mieloblastică).
Transcript of RAPORT ŞTIINŢIFIC Ş - automation.ucv.ro · pentru analiza problemelor biologice şi permite...

RAPORT ŞTIINŢIFIC ŞI TEHNIC
Cuprins
1. Obiective generale 2. Obiectivele fazei de execuţie 3. Rezumatul fazei 4. Descrierea ştiinţifică
4.1. Descrierea tumorilor canceroase. Noţiuni generale. 4.2. Modelarea matematică a creşterii tumorilor nevascularizate 4.3. Cancerul gastric 4.4. Bibliografie 4.5. Analiza imagistică. Noţiuni generale 4.6. Bibliografie 4.7. Utilizarea imaginilor în diagnoză 4.8. Implementare program 4.9. Bibliografie
1. Obiective generale
Elaborarea de modele conceptuale şi teorii.
2. Obiectivele fazei de execuţie
Dezvoltarea de modele conceptuale. Studii cu privire la posibila utilizare a acestora în predicţia evoluţiei tumorilor canceroase. Identificarea formaţiunilor (celulelor) prin analiza automată a imaginilor.
3. Rezumatul fazei
Modelarea matematică şi simularea sunt metode deosebit de eficiente, frecvent utilizate in toate domeniile. În ultimii ani s-au facut paşi concreţi în direcţia implicării teoriei sistemelor şi in studiul organismelor vii. Complexitatea sistemelor vii, chiar şi în cazul organismelor inferioare, este net superioară structurilor tehnice create de om. Acestea sunt constituite dintr-un mare număr de subsisteme, între care sunt stabilite multiple interconexiuni. Izolarea unui subsistem biologic, pentru studiul său, se face, inevitabil, cu preţul alterării propriei funcţii, cât şi a comportamentului sistemului în ansamblul său. Din acest motiv, datele obţinute experimental sunt diferite faţă de cele corespunzatoare unei evoluţii în condiţii normale. În concluzie, se recomandă studiul, pe cât posibil, al sistemului, în întregul său. Din punct de vedere sistemic, procesele ce se desfăşoară la nivel celular (inclusiv tumorile canceroase) au un pronunţat caracter neliniar. Deşi, în ceea ce priveşte studiul şi modelarea acestora s-au făcut progrese remarcabile, totuşi dezvoltarea şi aplicarea unor metode moderne este mai lentă în comparaţie cu alte domenii. Această întârziere este cauzată în principal de două caracteristici specifice ale bioproceselor. În cadrul acestei faze au fost trecute în revistă mai multe metode cu ajutorul cărora se pot simula evoluţiile populaţiilor de celule in diverse condiţii.
De asemenea un accent deosebit s-a pus pe posibilitatea diagnosticării folosind prelucrarea automată a imaginilor. În acest sens sunt prezentate exemple de prelucrare a frotiurilor cu probe hematologice (leucemie limfatică acută şi mieloblastică).

4. Descrierea ştiinţifică
4.1. Descrierea tumorilor canceroase. Noţiuni generale.
Una dintre problemele cu care se confruntă lumea medicală, de ani de zile este cancerul care este considerat ca fiind a doua cauză importantă a mortalităţii pe întregul glob. În România, în ultima decadă, s-a înregistrat o creştere rapidă a numărului tumorilor maligne, acestea reprezentând a doua cauză de deces, imediat dupa afecţiunile cardiovasculare. Puternicul impact al cancerului asupra societăţii româneşti se regăseşte nu numai în numărul mare de decese (circa 38.000 pe an, ceea ce reprezintă 13-14% din numărul total de decese) ci şi în numărul noilor cazuri înregistrate (circa 45.000 pe an) şi al bolnavilor în viaţă (circa 200.000). Pornind de la aceste date, tratarea cancerului a devenit una dintre priorităţile naţionale din domeniul medical. Nu există o definiţie simplă şi exactă a cancerului. Etimologia cuvântului provine din limba greacă “karkinos”, ceea ce înseamnă crustaceu, rac (cancer). În general termenul de cancer se referă la un grup de afecţiuni caracterizate prin creşterea anormală şi necontrolată a unei celule sau unui grup de celule, care invadează ţesuturile înconjuratoare şi care se pot împrăştia (metastază) la distanţă de ţesutul sau organul în care s-au format. Cancerul este produs prin apariţia unor mutaţii la nivelul celulelor normale prin care se modifică echilibrul dintre proliferarea şi moartea celulelor. Diviziunea celulelor devine rapidă, necontrolată şi generează tumori beningne sau maligne. Pierderea controlului proliferării celulare se produce prin mutaţii ale genelor care joacă un rol vital în diviziunea celulelor, în apoptoză (moartea celulelor) şi în repararea ADN-ului modificat. Sunt cunoscute peste 20 de mutaţii care iniţiază carcinogeneza şi sunt necesare mai multe mutaţii la aceeaşi celulă pentru a deveni o celulă modificată genetic cu potenţial neoplazic. Cancerul este în principal o boală a genelor care controlează creşterea celulară. Până în prezent, au fost lansate numai câteva modalităţi de tratare a cancerului (chirurgia, radioterapia şi chimioterapia), dar ele sunt considerate numai nişte ameliorante, mecanismele apariţiei, proliferării şi dezvoltării celulelor canceroase nefiind încă elucidate. Chirurgia este cea mai veche modalitate de tratament şi la un moment dat, reprezenta singura formă de terapie a cancerului. Această metodă este eficientă în cazurile tumorilor localizate la nivelul unui anumit organ. În cazul în care apar micrometastaze, chirurgia singură nu este eficientă ca modalitate de tratament şi se impune combinarea ei cu alte tehnici. Aceasta este indicată ca tratament pentru: tumori mamare, tumori de prostata, tumori orofaringiene, tumori ale pielii, tumori gastrointestinale, tumori pulmonare, etc. În situaţia în care tumora este greu accesibilă sau starea de sănătate generală a pacientului presupune un risc crescut al apariţiei complicaţiilor post-operatorii, radioterapia s-a dovedit a fi mai eficientă. Aceasta este indicată pentru limfom extranodular localizat, tumori nazale, tumori de creier, celule scamoase carcinomatoase, adenom perianal, etc. Ca şi chirurgia, radioterapia prezinta şi ea unele limitări. În cazul tumorilor voluminoase există riscul ca celulele transformate malign din centrul masei tumorale să nu fie afectate ceea ce conduce ulterior la recurenţa bolii. De aceea se recomandă utilizarea radioterapiei în combinaţie cu chirurgia şi/sau chimioterapia care implică folosirea medicamentelor. Cercetările privind metodele de terapie a cancerului continuă în întreaga lume din dorinţa de înţelegere a mecanismelor de evoluţie a celulelor canceroase în scopul elaborării unor metode de terapie eficiente. Toate acestea arată că şi în cazul unor modele simple, elementare, pot apărea numeroase aspecte interesante atât din punctul de vedere al medicului şi/sau biologului, cât şi din cel al matematicianului. Modelarea matematică realizează sinteza datelor obţinute experimental într-un sistem unitar, permite evidenţierea structurii interne şi a legăturilor cauzale dintre părţile componente şi măsoară ponderea cu care intervine fiecare subsistem la realizarea funcţiilor

sistemului. Simularea asigură validarea teoriilor concurente, înţelegerea modificărilor fiziopatologice şi sugerează experimente relevante. Ea reprezintă o unealtă foarte puternică pentru analiza problemelor biologice şi permite dezvoltarea şi testarea ipotezelor ce pot duce la o înţelegere mai bună a proceselor biologice. Principiile de bază ale unui model realist şi folositor sunt:
Înţelegerea şi aprecierea deplină a problemei biologice Reprezentarea matematică realistă a fenomenului biologic Găsirea unor soluţii folositoare, de preferat cantitative Interpretarea bilogică a rezultatelor matematice în termeni de înţelegere şi
predicţii. Modelarea matematică şi simularea sunt metode deosebit de eficiente, frecvent utilizate în toate domeniile. În ultimii ani s-au facut paşi concreţi în direcţia implicării teoriei sistemelor şi în studiul organismelor vii. Complexitatea sistemelor vii, chiar şi în cazul organismelor inferioare, este net superioară structurilor tehnice create de om. Acestea sunt constituite dintr-un mare număr de subsisteme, între care sunt stabilite multiple interconexiuni. Izolarea unui subsistem biologic, pentru studiul său, se face, inevitabil, cu preţul alterării propriei funcţii, cât şi a comportamentului sistemului în ansamblul său. Din acest motiv, datele obţinute experimental sunt diferite faţă de cele corespunzatoare unei evoluţii în condiţii normale. În concluzie, se recomandă studiul, pe cât posibil, al sistemului, în întregul său. Din punct de vedere sistemic, procesele ce se desfăşoară la nivel celular (inclusiv tumorile canceroase) au un pronunţat caracter neliniar. Deşi, în ceea ce priveşte studiul şi modelarea acestora s-au făcut progrese remarcabile, totuşi dezvoltarea şi aplicarea unor metode moderne este mai lentă în comparaţie cu alte domenii. Această întârziere este cauzată în principal de două caracteristici specifice ale bioproceselor. În primul rând, modelarea acestora este deosebit de dificilă. Aceste sisteme cuprind o multitudine de interacţiuni cu alte procese şi prin urmare funcţionarea acestora şi în special dinamicile de creştere, sunt, de multe ori, greu de înţeles, puternic neliniare şi nestaţionare. De asemenea, reproductibilitatea experimentelor este nesigură, iar lipsa de acurateţe a măsurătorilor poate conduce la o serie de probleme de identificare. În al doilea rând, aplicarea unor strategii de monitorizare se confruntă în majoritatea cazurilor cu absenţa unei instrumentaţii specifice, sigure şi ieftine, destinate măsurării directe şi/sau monitorizării în timp real a unor variabile biologice şi biochimice. Actualmente, piaţa oferă puţini senzori capabili să furnizeze astfel de măsurători, iar parametrii inaccesibili sau nemăsurabili trebuie determinaţi prin analize de laborator off-line. Costul şi durata acestor analize limitează frecvenţa cu care se efectuează aceste măsurători şi conduc la creşterea costurilor. Pentru surmontarea acestor probleme este necesară utilizarea tehnicilor avansate de modelare şi identificare care folosesc senzori software pentru reconstruirea stărilor şi/sau parametrilor nemăsurabili. Din cele prezentate, rezultă ca modelarea şi simularea sistemelor vii se înscrie în tendinţa modernă de integrare a cunoştiinţelor, fiind ele însele rezultatul unei colaborari interdisciplinare. Celulele canceroase se divid într-un mod necontrolat, numarul lor crescând până când formează o tumoră ce devine vizibilă clinic. Pe lângă înmulţirea extrem de rapidă, celulele canceroase sunt incapabile să se organizeze într-un mod adecvat, iar masa de ţesut care ia naştere nu are caracteristicile unui ţesut normal. În etapa următoare, celulele încep să se desprindă de tumora mamă, străbat membranele conjunctive, apoi pereţii vaselor limfatice sau sangvine pătrunzând în circulaţia limfatică celulele canceroase ajung iniţial în ganglionii limfatici, structuri cu rol imun. Sunt invadaţi iniţial ganglionii regionali, adică cei mai apropiaţi de tumoră. O mare parte din celule sunt omorâte la acest nivel; aceasta reprezintă etapa de invazie locoregională a cancerului. Cu timpul, celulele imune de la nivelul ganglionilor nu mai fac faţă afluxului de celule canceroase care continuă să se înmulţească. Celulele canceroase vor depaşi şi această barieră şi vor ajunge prin intermediul limfei în

circulaţia sangvină; o altă modalitate de a ajunge în sânge este prin străbaterea pereţilor vasculari încă de la început, la nivelul tumorii primare. După ce ajung în circulaţia sangvină, celulele canceroase se vor opri în diferite organe, acolo unde condiţiile favorizează dezvoltarea ulterioară: plămân, oase, creier, ficat, etc. În aceste organe o parte din celule vor muri, dar altele vor supravieţui şi se vor înmulti în continuare; aceasta reprezintă etapa metastatică, de boală generalizată. În fiecare etapă vor supravieţui cele mai agresive şi mai rezistente celule, astfel că şansele de vindecare sau de răspuns la tratament scad odata cu avansarea bolii. Capacitatea celulelor canceroase de a se răspândi în interiorul organismului este cea care cauzează gravitatea acestei boli. Cauza principală a cancerului este încă neclară. Factorii de mediu, cum ar fi poluarea chimică şi expunerea la radiaţii, cauzează unele forme de cancer; un număr mare de experimente ce au identificat substanţele chimice provocatoare de cancer (substanţe carcinogene) au fost efectuate pe animaleRadiaţiile ionizante, de asemenea, afectează materialul genetic al celulelor, astfel radiaţiile create în urma exploziei bombei atomice din Japonia, în 1945, a cauzat multe forme de cancer, unele declansându-se abia după 10-20 de ani de la expunerea la acestea. Există însă şi alţi factori care provoacă boala. De exemplu, unele cazuri de cancer pot fi produse de viruşi care afectează structura genetică a celulelor. Factori de risc:
Tutun 30% Infecţii 10% Alimentaţie nesănatoasă 33% Factori legaţi de sănătatea reproducerii 7% Profesie 4% Lumina solară şi radiaţiile ionizante 3% Poluare 2% Produşi industriali 1% Medicamente şi proceduri medicale 1% Aditivi alimentari 1% Alcool 2% Factori necunoscuţi 6%
În unele cazuri sistemul imunitar este de vină. S-ar părea că celulele canceroase sunt produse constant în interiorul organismului, însă acestea sunt distruse de obicei de către sistemul imunitar. Din unele motive, acest sistem poate să nu funcţioneze corect, permiţând înmulţirea celulelor canceroase. Diagnosticul de cancer trebuie confirmat. Suspiciunea de cancer o ridică anamneza, examenul clinic, examene imagistice şi de laborator. Examinarea clinică completă, acordă o mare atenţie verificării ganglionilor limfatici periferici dar mai ales a celor regionali tumorii. Confirmarea şi certitudinea diagnosticului de cancer este dată de examenul anatomopatologic. Această procedură poate fi efectuată fie prin biopsie, fie prin examinare citologică.
4.2. Modelarea matematică a creşterii tumorilor nevascularizate
Căutările în baza de date bibliografică PubMed relevă faptul că dintr-un număr de 1.5 milioane studii în domeniul cercetării cancerului, aproximativ 5% se axează pe modelarea matematică. În orice caz este evident faptul că algoritmii de modelare şi simulare specifici teoriei sistemelor, ar putea avea o contribuţie majoră în multe din domeniile de investigare

experimentală a cancerului, întrucât există în acest moment un număr vast de date experimentale ce necesită o analiză sistematică. O formaţiune de celule tumorale cu evoluţie rapidă nu se poate dezvolta dincolo de o anumită dimensiune, întrucât există un echilibru între celulele din interiorul formaţiunii ce consumă nutrienţi şi dispersia nutrienţilor în formaţiune. Prin urmare, unul din paşii cei mai importanţi în creşterea tumorilor maligne este angiogenza, adică procesul prin care tumorile îşi dezvoltă propria reţea vasculară.
Fig. 4.1 Procesul de angiogeneză Andrea Hawkins-Mathematical Models of Tumor Growth (2007)
Din acest motiv, noi medicamente sunt create special pentru a distruge vasele de sânge tumorale. Imediat ce tumorile şi-au dezvoltat propria reţea vasculară, celulele tumorale se pot deplasa prin sistemul circulator (în metastază) şi pot forma tumori secundare oriunde în corp. După angiogeneză şi metastază, pacientul prezintă numeroase tumori în diferite părţi ale corpului, foarte dificil de detectat şi tratat. Întrucât există 3 stadii diferite (nevascularizat, vascularizat şi metastatic) în dezvoltarea cancerului, cercetătorii îşi concentrează eforturile către găsirea răspunsurilor la întrebări ce ţin de fiecare dintre aceste stadii. Ca prim pas am ales cunoaşterea stării curente a modelării matematice a creşterii tumorale nevascularizate (tumori fără vase de sânge). Asta nu înseamnă că acesta este aspectul cel mai important în creşterea tumorilor – din contră, din punct de vedere clinic, angiogeneza şi creşterea tumorilor vascularizate împreună cu metastaza, reprezintă cauza decesului pacientului, iar modelarea şi înţelegerea acestora sunt cruciale în terapia cancerului. Creşterea unei tumori nevascularizate e mult mai simplu de modelat matematic şi cu toate acestea, conţine mult mai multe elemente necesare unui model general de creştere tumorală vascularizată. Mai mult decât atât, uşurinţa şi repetabilitatea experimentelor cu tumori nevascularizate indică faptul că atât cantitatea cât şi calitatea probelor experimentale o depăşesc pe cea a tumorilor vascularizate, pentru care efectele sunt adesea dificil de izolat. În particular, întrucât unele celule tumorale dezvoltate in vitro formează agregate sferice, costurile relativ scăzute şi uşurinţa experimentelor in vitro în comparaţie cu cele pe animale au făcut ca analizele sferoidelor tumorale să devină foarte populare. Astfel, modelarea tumorilor nevascularizate e percepută ca un prim pas spre construirea unor modele pentru tumorile complet vascularizate. Mai mult decât atât există un număr de întrebări referitoare la tumorile nevascularizate, incluzând recenta controversă asupra ipotezei conform căreia toţi indivizii au tumori nevascularizate latente în corpurile lor. Aşadar, modelarea tumorilor nevascularizate poate fi folosită pentru experimetele pe tumori vascularizate şi metastatice, experimente care sunt mult mai greu de efectuat, întrucât trebuiesc făcute in vitro.
Principala descoperire a experimentelor in vitro pe tumori sferice a fost aceea că acestea cresc până ajung la dimensiuni critice, moment în care creşterea încetează. Această dimensiune critică este determinată de un echilibru între proliferarea celulelor şi moartea acestora în interiorul sferoidului. Pricipalele observaţii experimentale punctează faptul că atunci când celulele canceroase sunt într-un mediu hrănitor, ele proliferează, când sunt într-un mediu ostil, celulele canceroase declanşează moartea celulară (apoptoză), iar atunci când celulele sunt într-un mediu puţin hrănitor acestea rămân în stare latentă. Diferitele niveluri de nutriţie dintr-un sferoid tumoral sunt determinate de mişcarea şi de consumul de nutrienţi din interiorul tumorii. Faptul că experimentele in vitro au indicat clar că răspândirea nutrienţilor (în special a oxigenului) limitează creşterea sferoidelor tumorale a netezit drumul către ipoteza angiogenezei. Aceasta spune că, pentru a putea creşte, tumorile trebuie să obţină propriile lor vase de sânge, aşadar trebuie să “recruteze” vase de sânge de la vasculatura gazdă. Această ipoteză a condus la cercetări considerabile, chiar la un posibil tratament pentru cancer, prin identificarea factorilor care ar stopa angiogeneza. În prezent, câteva clinici evaluează eficacitatea şi siguranţa terapiilor antiangiogenetice şi comunitatea ştiinţifică aşteaptă rezultatele.
Procesul consumului şi răspândirii nutrienţilor în tumori a fost modelat începând cu mijlocul anilor 1960. Majoritatea modelelor pot fi grupate în doua categorii:
1) Modele matematice continue care constau în ecuaţii cu derivate parţiale. 2) Modele matematice discrete.
Majoritatea modelelor consideră proliferarea şi moartea celulelor tumorale a fi dependente de numai un nutrient generic (cel mai adesea oxigenul). Cu toate acestea unii iau în considerare efectele mai multor nutrienţi şi a pH-ului asupra populaţiei celulare. În timp ce detaliile oricărui model sunt diferite într-un context experimental specific, principiul fundamental ce stă la baza acestor modele poate fi înglobat în următoarea afirmaţie:
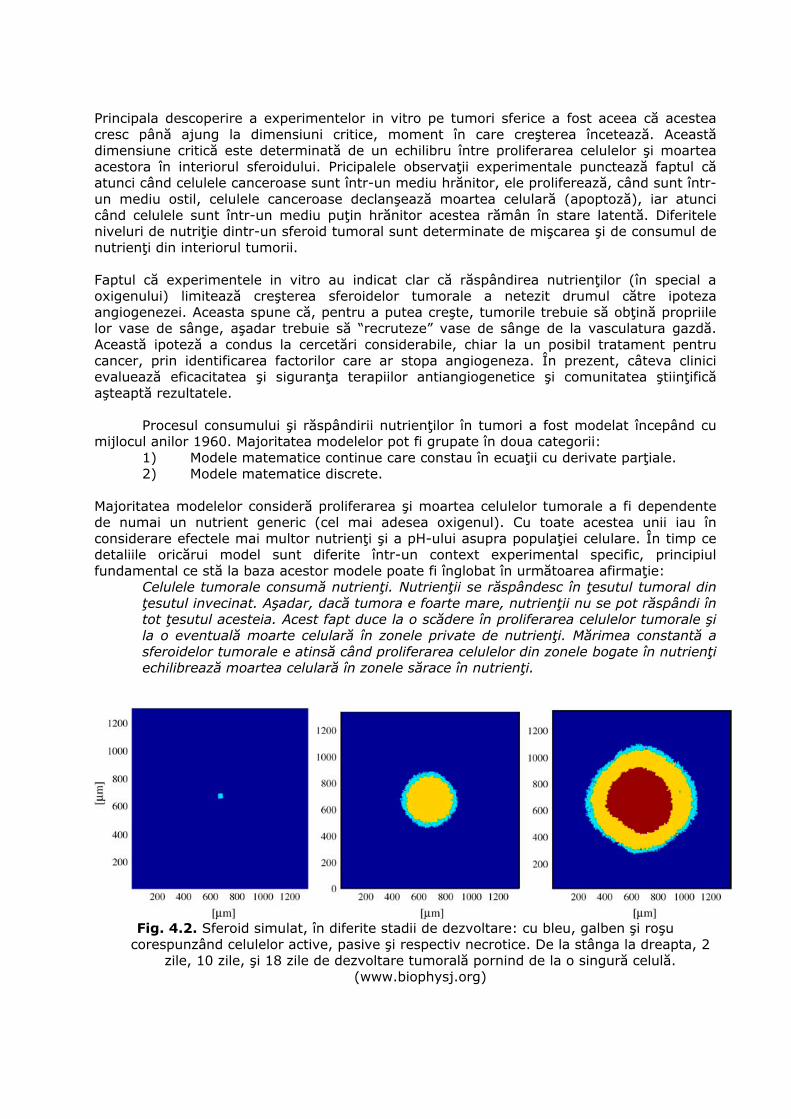
Celulele tumorale consumă nutrienţi. Nutrienţii se răspândesc în ţesutul tumoral din ţesutul invecinat. Aşadar, dacă tumora e foarte mare, nutrienţii nu se pot răspândi în tot ţesutul acesteia. Acest fapt duce la o scădere în proliferarea celulelor tumorale şi la o eventuală moarte celulară în zonele private de nutrienţi. Mărimea constantă a sferoidelor tumorale e atinsă când proliferarea celulelor din zonele bogate în nutrienţi echilibrează moartea celulară în zonele sărace în nutrienţi.
Fig. 4.2. Sferoid simulat, în diferite stadii de dezvoltare: cu bleu, galben şi roşu
corespunzând celulelor active, pasive şi respectiv necrotice. De la stânga la dreapta, 2 zile, 10 zile, şi 18 zile de dezvoltare tumorală pornind de la o singură celulă.
(www.biophysj.org)

Există puţine modele experimentale validate pentru dezvoltarea populaţiilor de celule şi interacţiunea nutrienţilor în acelaşi timp. Dintre acestea, l-am ales pe cel al lui Casciari, Sotirchos şi Sutherland, (Casciari, Sotirchos, Sutherland, 1992) deoarece prezintă exemple relevante ale clasei şi includ un nivel al complexităţii biochimice suficient pentru a fi relevante din punct de vedere experimental. Acest model se referă la tumorile sferice şi interacţiunea celulelor tumorale cu oxigenul, glucoza, lactaţi, dioxidul de carbon, bicarbonatul, clorul şi ionii de hidrogen. Ecuaţiile de conservare sunt (Casciari, Sotirchos, Sutherland, 1992):
iii PN
tC
=⋅∇+∂∂
(1)
în care iC reprezintă concentraţiile (subindicele a pentru oxigen, b pentru glucoză, c pentru ionul de lactat, d pentru dioxidul de carbon, e pentru ionul de bicarbonat, f pentru ionul de clor, g pentru concentraţia ionului de hidrogen), iN este fluxul fiecăreia din speciile chimice
din interiorul sferoidelor tumorale şi iP este gradul net de consum/producţie a speciilor chimice de către celulele tumorale şi în urma reacţiilor chimice cu alte specii. Pentru moleculele de glucoză, oxigen şi dioxid de carbon fluxul este dat de legea lui Fick:
iii CDN ∇−= (2) unde iD sunt coeficienţii dispersării constante. Fluxul de specii ionice (ionii de lactat, bicarbonat şi hidrogen) în soluţie diluată, trebuie să ia în considerare atât migraţia sarcinii induse de câmpul electric căt şi de dispersie şi este dat de:
iiiiii CDFCuzN ∇−Φ∇−= (3)
unde iz este sarcina ionică a speciei, iu este mobilitatea, F este constanta lui Faraday iar Φ este potenţialul electric. Pentru soluţii diluate Casciari, Sotirchos şi Sutherland l-au considerat pe iu ca fiind dat de ecuaţia Nernst-Einstein:
TRDu gii /= (4)
unde gR este constanta gazelor, T este temperatura absolută iar iD este constanta de
dispersie. Aplicând presupunerea conform căreia curentul electric net este zero astfel încât
∑ =k
kk Nz 0 atunci ecuaţia (3) poate fi rescrisă sub forma:
⎟⎟⎟
⎠
⎞
⎜⎜⎜
⎝
⎛ ∇−∇−=
∑∑
kkkk
kkkk
iiiii CDz
CDzCzCDN 2 (5)
Asfel, fluxul unei specii ionice depinde de gradienţii de concentraţie ai tuturor speciilor ionice prezente în ţesut. În final, luând în considerare căderea de glucoză prin glicoliză şi prin ciclul Krebs şi traseul metabolic detaliat pentru reglarea pH-ului la nivel unicelular, Casciari, Sotirchos şi Sutherland au ajuns la următorii termeni ce descriu consumul de oxigen şi glucoză de către

celulele tumorale şi interacţiunile dintre alte substanţe chimice şi specii ionice în următoarea înşiruire:
( )
,
,0,
,,3/2
,1
,
cagerdfg
f
agerdfe
gerdfd
abc
mbb
bnga
bbb
maa
amgb
aaa
PPCCkCkP
PPCCkCkP
CCkCkPPPP
KCC
CCBAP
KCC
CCBAP
+−−=
=
−−=
+−=−−=
⎟⎟⎠
⎞⎜⎜⎝
⎛+⎟
⎟⎠
⎞⎜⎜⎝
⎛⎟⎟⎠
⎞⎜⎜⎝
⎛+Ω−=
⎟⎟⎠
⎞⎜⎜⎝
⎛+⎟
⎟⎠
⎞⎜⎜⎝
⎛+Ω−=
(6)
unde Ω reprezintă numărul de celule dintr-un sferoid pe unitate volumică (presupus constant). În ecuaţiile (6) relaţiile dintre fedc PPPP ,,, şi gP au derivat din utilizarea
consideraţiilor detaliate ale ciclului Krebs, iar parametrii fK şi rK sunt determinaţi din
schemele reacţiilor chimice detaliate. Formele funcţionale ale consumului de oxigen aP şi
consumului de glucoză bP au derivat empiric din experimente. Au fost conduse în particular, masurători independente ale consumului de oxigen la diferite niveluri ale pH-ului şi glucozei şi consumul de glucoză la diferite niveluri ale pH-ului şi oxigenului iar formele funcţionale empirice ale sistemului (6) au fost corectate. Puţine modele au fost parametrizate şi testate experimental. În timp ce validarea experimentală este fără doar şi poate crucială pentru succesul oricărui model, modelele fenomenologice pot asigura un cadru în care ne putem gândi la ordinea fizică, permiţându-ne să cercetăm efectele diferitelor ipoteze, să înaintăm în gândirea conceptuală şi chiar să facem predicţii. Un exemplu pentru aceasta din urmă este oferit de Gatenby şi Gawlinski care descriu tranziţia dintre creşterea benignă şi cea malignă utilizând ipoteza mediului acid. Aceasta constă în 3 ecuaţii reacţie-dispersie cuplate pentru celulele canceroase, pentru celulele normale şi pentru aciditate şi cercetează ipoteza că, în urma unui metabolism anaerob, celulele canceroase crează un mediu acid care omoară celulele sănătoase. Modelul intuieşte existenţa unui gol interstiţial la interfaţa tumoră-gazdă, predicţia aceasta fiind ulterior confirmată experimental. Acest exemplu indică puterea unei abordări interdisciplinare şi explică potenţialul ei. Este regretabil că multe dintre prezicerile legate de modelare nu sunt testate experimental. Toate abordările principale menţionate mai sus consideră starea fiecărei celule sau populaţie de celule dinăuntrul tumorii a fi caracterizată prin vectorul variabil:
{ }uvxw ,,= (7)
unde x este poziţia celulei, v este viteza celulei şi u este un vector ce caracterizează starea biologică internă a celulei (poate conţine poziţia acesteia în ciclul celulei, interacţiunea ei cu mediul biochimic local, etc.). Interacţiunile mecanice dintre celule depind de poziţia x şi de viteza v. Acest tip de vector de stare îşi are originile în structurile Boltzmann şi în alte modele mecanice statistice, dar ne aşteptăm ca viteza celulei să joace un rol mai puţin important în biologia tumorii deoarece „efectele coliziunii” dintre celule nu se pot compara cu cele dintre moleculele de gaz, de exemplu.

În principiu este posibil să se scrie un set de interacţiuni mecanice dintre celule utilizând mecanica statistică sau legile de mişcare ale lui Newton, dar acest lucru nu este făcut de obicei deoarece sistemul rezultat este mare, complicat, costisitor şi în acelaşi timp trebuie făcute multe presupuneri despre felul în care interacţionează celulele. Astfel, este foarte întâlnită înlocuirea legilor fizice de mişcare a celulei (raporturi de forţe sau mase) cu reguli de deplasare celulară. Câteva principii generale pentru aceste reguli de deplasare celulară sunt descrise mai jos. Principala dificultate constă în modelarea mişcării celulei. Primul pas în setarea regulilor mişcării celulei este partiţionarea spaţiului fizic în zone. Cel mai simplu mod de a face acest lucru este discretizarea sub forma unei grile regulate; grilele rectangulare sunt folosite de obicei pentru simplitate. O partiţionare alternativă folosită de Kansal distribuie aleator centrele celulelor şi apoi foloseşte un şablon Voronoi pentru a divide spaţiul în celule. A doua decizie în modelare este accea dacă grila este fixată în timp sau variază în timp ce elementele se mişcă. Este mult mai simplu să o considerăm fixă deoarece fiecare nod corespunde unei celule biologice sau unui spaţiu liber iar celulele se pot muta într-un nod apropiat din grilă care e liber. Este, bineînţeles, mult mai realist să avem celule care cresc şi se mişcă, cu o grilă care evoluează în timp ce tumora creşte, dar astfel de modele sunt mult mai greu de formulat şi astfel sunt mult mai rare. Pentru a găsi legea mişcării celulei într-un grilaj fix, cele mai multe modele consideră că interacţiunile dintre o celulă individuală şi celulele înconjurătoare sunt localizate în spaţiu şi apar doar în cazul celulelor dintr-o anumită vecinătate O (de obicei este reprezentată de vecinii cei mai apropiaţi ai celulei dar se pot extinde şi mai departe). O lege automată de acest fel spune că dacă o locaţie x este ocupată de o celulă canceroasă, aceasta se poate inmulţi cu probabilitatea k1, se poate lega de un efector chimic (cum ar fi: un factor de creştere sau un factor care poate determina celula să moară), poate deveni pasivă cu probabilitatea k2 sau poate migra într-un loc din apropiere cu probabilitatea k3. Înmulţirea celulelor depinde de nivelul de hrană şi reziduu într-un anumit punct şi ar trebui să provină din condiţiiile biochimice ce afectează ciclul celulei. Probabilităţile k1, k2, k3 sunt de obicei independente de geometria şi de mediul mecanic al celulei, dar în principiu pot fi şi ele încorporate. Din momentul în care s-a luat decizia de inmulţire, celula se va divide în două celule-fiică. Într-o grilă deformabilă această diviziune nu reprezintă o problemă deoarece celula ne face altceva decât să crească dublul mărimii originale după care se împarte în 2 celule, fiecare aceeaşi mărime ca şi originalul. Totuşi, într-o grilă fixă această creştere apare instantaneu, noile celule ocupând imediat 2 locuri. Una dinstre aceste celule se presupune că ocupă locul original al celulei-mamă dar cea de-a doua necesită un spaţiu în plus. Există două feluri de a determina unde se va pune această nouă celulă. De obicei se mută într-un loc învecinat dar trebuie decis dacă celulele înconjurătoare vor trebui împinse pentru a face loc (dacă da, unde ar trebui împinse?), sau dacă celula se poate muta doar într-un loc „vacant”. Determinarea acestui loc reprezintă una din principalele probleme ale abordării grilei fixe şi constituie principala dificultate a unei astfel de legi de mişcare. Pe lânga aceasta, structura tip grila aleasă poate influenţa artificial comportamentul global, într-o manieră similară cu efectele de grilaj ce pot fi observate în aproximarea numerică a soluţiilor ecuaţiilor diferenţiale parţiale. În al doilea caz se poate cuantifica acest efect prin analiză matematică, dar o astfel de cuantificare este foarte dificilă în ceea ce priveşte modelarea celulară automată. Mişcarea unei celule spre un loc învecinat ar putea depinde de:
a) numărul de locuri învecinate libere b) gradientul concentraţiei de nutrient c) mediul mecanic (forţe de interacţiune cu celulele din locurile învecinate).
Oricare dintre aceşti factori pot fi combinaţi cu o mobilitate aleatoare asignată fiecărei celule.

Diferite modele celulare încorporează diferite nivele de complexitate în mişcarea celulei, naşterea ei şi legi de distrugere. După această descriere a formulării generale vom descrie în continuare câteva exemple specifice de modele automate care au apărut în literatură. Unul dintre primele modele ce folosesc migraţia discretă a populaţiilor celulare utilizând un model complx al ciclului celular a fost condus de Duchting şi Vogelsaenger care au făcut o serie de simulări tridimensionale utilizând modelul pentru determinarea efectului pe care îl are radioterapia asupra tumorilor într-o manieră cantitativă şi bine parametrizată. Un alt model timpuriu care a părut să fie bine parametrizat este cel al lui Qi şi este unul dintre cele mai puţin complicate modele de acest tip şi descrie legile minime ce se utilizează pentru a reproduce legea creşterii lui Gompertz. Notă: Legea lui Gompertz afirmă că volumul V(t) al unei tumori la momentul t este dat de relaţia:
KKVtV
rt )exp(0)(
−
⎥⎦⎤
⎢⎣⎡= (8)
unde V0 este volumul iniţial al tumorii, K este valoarea de echilibru iar r este o constantă pozitivă.
4.3. Cancerul gastric Deşi incidenţa şi rata mortalităţii prin cancer gastric au scăzut în ultimii 60 de ani, neoplaziile de acest tip rămân în continuare o problemă importantă în patologia oncologică în SUA, Japonia, China, Chile şi Irlanda. Studiile epidemiologice au sugerat că riscul pentru cancer gastric este asociat cu nivelul socioeconomic scăzut al unor populaţii şi cu factori alimentari exogeni (contaminarea bacteriană a alimentelor, conservare inadecvată şi posibil, infecţia cu Helicobacter pylori) şi endogeni (scăderea acidităţii gastrice, gastrita trofică, anemie pernicioasă, intervenţii chirurgicale pe stomac, nitriţi alimentari, unele medicamente). La nivelul stomacului sunt întâlnite o multitudine de leziuni cu aspecte de multe ori asemănătoare, dar produse de cauze diferite şi evoluţie variabilă. Tumorile gastrice cele mai frecvent întâlnite sunt: leiomiom, hemangiom, lipom, neurofibrom, carcinom gastric intramural, leiomiosarcom. Dintre leziunile polipoide cele mai frecvent întâlnite sunt: polipi hiperplastic, hiperplazia focală fobeolară, eroziuni cronice, adenom, carcinoid, carcinom, limfom. Frecvent, caracteristicile maligne sau benigne ae acestor leziuni nu sunt specifice şi nu pot fi diferenţiate prin examene macroscopice morfopatologice. Există, de asemenea, leziuni precanceroase cu evoluţie îndelungată, care se pot transforma în timp în neoplasme gastrice. Cancerul gastric apare în 3 forme: adenocarcinomul gastric (90% din cazuri), limfomul gastric (7% din cazuri) şi sarcomul gastric (între 1 si 3% din cazuri). Adenocarcinomul gastric: Apare sub două forme:
Tipul difuz: celulele nu sunt legate între ele, infiltrează şi îngroaşă peretele gastric difuz, cuprinde întregul stomac şi reduce distensibilitatea şi elasticitatea peretelui gastric. Are prognostic sever.
Tipul intestinal: celulele aderă între ele, formează structuri tubulare cu aspect glandular, macroscopic are frecvent aspect ulcerativ, se dezvoltă mai ales în zona antrului şi micii curburi şi sunt precedate de leziuni precanceroase cu evoluţie prelungită.

Adenocarcinoamele gastrice diseminează prin invazie directă în ţesuturile din jurul stomacului pe cale limfatică şi în peritoneu. Limfomul gastric primitiv: Aspect macroscopic asemănător cu adenocarcinomul. Microscopic este limfom nehodgkinian cu celule de tip B. Tratamentul şi evoluţia sunt mai favorabile decât în cazul adenocarcinomului. Foarte rar este format din celule de tip limfom hodgkinian.
Sarcomul gastric: Este de tipul leiomiosarcom. Microscopic, aspectul este de tip leziune benignă dar evoluţia este malignă, invadează rar organele din apropierea stomacului şi nu produce metastaze în ganglionii limfatici. Clasificare Borrmann a leziunilor neoplazice gastrice: Formele morfologice ale cancerului gastric au fost clasificate de către Borrmann în 4 tipuri:
Tipul I: Tumoră de formă polipoidă, apare în 30% din cazuri, nu permite diferenţierea între leziunile benigne şi maligne.
Tipul II: Tumoră de formă polipoidă cu ulceraţie. Apare în 30% din cazuri, are aspect de formaţiune exofitică cu un crater central ulcerativ prin necroză.
Tipul III: tumoră infiltrativă cu ulceraţie. Apare în 10% din cazuri. Este produsă de cancer infiltrativ în mucoasă, slab delimitat de ţesutul sănătos din jur, cu ulceraţie superficială.
Tipul IV: Tumoră difuză infiltrativă. Apare în 30% din cazuri. Constă în infiltraţie sub mucoasă extinsă, mucoasa este neregulată, peretele stomacului este indurat nedistensibil.
Metode de evidenţiere imagistică a neoplazmelor gastrice: În procesul de investigare a neoplaziilor stomacului se folosesc următoarele metode imagistice:
examinare radiologică simplă, cu dublu contrast şi cu distensie gastrică examinare tomografică computerizată examinare endoscopică examinare ultrasonografică endoscopică biopsie gastroendoscopică examen citologic al lichidului gastric spontan sau prin periaj metoda vizualizării celulelor neoplazice cu ajutorul proteinelor fosforescente
Metodele exploratorii folosite pentru investigarea cancerului gastric incipient nu pot stabili cu certitudine caracterul neoplazic al unei leziuni gastrice. Pentru leziunile precanceroase benigne sau maligne metodele imagistice prezentate nu pot asigura un diagnostic de certitudine. Aplicarea metodelor imagistice prezentate în practica oncologică a pus în evidenţă unele dificultăţi în diagnosticarea cancerului gastric care explică erorile de diagnostic şi consecutiv, de tratament, ale cancerelor gastrice.
Localizarea anatomică a cancerului şi ulcerului gastric nu prezintă specificitate Diferenţierea între adenocarcinom şi limfom este frecvent incertă la nivel
morfopatologic macroscopic, iar biopsia necesită recoltarea probelor dintr-un număr mare de puncte şi din profunzmea mucoasei.
Diferenţierea radiologică între leziunile de ulcer gastric şi neoplazm ulcerat nu este precisă
Biopsia negativă pentru neoplazm nu exclude diagnosticul de cancer gastric Aspectul microscopic benign nu exclude o evoluţie malignă a leziunilor Leziunile precanceroase pot deveni neoplazme în intervale de timp variabile.
Metoda vizualizării celulelor canceroase prin proteine fosforescente:

Recent a fost introdusă în practica oncologică metoda proteinelor fosforescente pentru vizualizarea neoplasmelor „in vivo”. Proteinele fosforescente sunt produse naturale ale celulelor, dar exprimarea efectului fosforescent poate fi crescută prin introducerea în celulă a unei trans-gene purtată de un vector viral. Metoda constă în folosirea unui virus purtător al acestei gene, care infectează specific şi stabil numai celulele neoplazice, nu şi celulele sănătoase din ţesutul gazdă. Replicarea intra-celulară a virusului vector produce creşterea sintezei proeinelor fosforescente, astfel incât celulele neoplazice devin vizibile şi pot fi detectate vizual începând de la aglomerări celulare cu diametrul de 1 mm în organele gazdă, în sânge sau la distanţă în organe în care s-au produs metastaze. Au fost elaborate protocoale ale acestei metode pentru vizualizarea tumorilor din plămâni, esofag, stomac, intestin, rect şi pancreas. Sunt folosite proteine fosforescente în special de culoare verde dar şi proteine cu alte culori. La nivel celular au fost colorate nucleul în verde şi citoplasma în roşu pentru vizualizarea modificărlor în cursul diviziunilor celulare. Metoda este considerată o „microscopie intravitală” şi asigură vizualizarea directă în timp real pentru: creşterea tumorilor primare, mobilizarea celulelor neoplazice prin sânge şi limfă, invazia altor ţesuturi cu formare de metastaze, angiogeneză, interacţiunea tumoră-mediu înconjurător şi modificările tumorii sub influenţa tratamentului. Pentru cancerele gastrice au fost folosite tulpini de virus herpes simplex cu colorare în verde fosforescent a celulelor neoplazice în stomac şi metastaze. (vezi exemplele din fig. 4.3).
Imagine preluată din art. Jian-Hua Zhang şi colab., Construction and identification of recombinant vectors carrying herpes simplex virus thymidine kinase and cytokine genes expressed in gastric carcinoma cell line SGC 7901
Imagine preluate din art. Adusumilli şi colab., Real-time diagnostic imaging of tumors and metastases by use of a replication-competent herpes vector to facilitate minimally invasive oncological surgery
Fig. 4.3. Transformarea celulelor canceroase în celule fosforescente.
4.4. Bibliografie
1. R. A. Anderson. A hybrid mathematical model of solid tumour invasion: the importance of cell adhesion. Mathematical Medicine and Biology, 22, 2005.
2. R. P. Araujo and D. L. S. McElwain. A mixture theory for the genesis of residual stresses in growing tissues I: A general formulation. SIAM J. Appl. Math., 65:1261-1284, 2005.

3. H. B. Frieboes, J. S. Lowengrub, S.Wise, X. Zheng, P. Macklin, E. L. Bearer, and V. Cristini. Computer simulation of glioma growth and morphology. NeuroImage, Article in Press, 2007.
4. D. Hanahan and R. A. Weinburg. The hallmarks of cancer. Cell, 100:57-70, January 2000. 5. S. Sun, M. F. Wheeler, M. Obeyesekere, and C. Patrick, Jr. Multiscale angiogenesis modeling
using mixed finite element methods. Multiscale Modeling and Simulations 4(4):1137-1167,2005.
6. Tiina Roose, S. Jonathan Chapman, Philip K. Maini, Mathematical Models of Avascular Tumor Growth, SIAM REVIEW Society for Industrial and Applied Mathematics, Vol. 49,No . 2, pp . 179–208, 2007.
7. J.J. Casciari, S.V. Sotirchos, and R.M. Sutherland, Mathematical modelling of microenvironment and growth in EMT6/Ro multicellular tumour spheroids, Cell Proliferation, 25 (1992), pp. 1–22.
8. R.A. Gatenby and E.T. Gawlinski, A reaction-diffusion model of cancer invasion, Cancer Res., 56 (1996), pp. 5745–5753.
9. W. Duchting and T. Vogelsaenger, Recent progress in modelling and simulation of threedimensional tumor growth and treatment, Biosystems, 18 (1985), pp. 493–412.
10. A.-S. Qi, X. Zheng, C.Y. Du, and B.S. An, A cellular automaton model of cancerous growth, J. Theoret. Biol., 161 (1993), pp. 1–12.
11. R.A. Gatenby and P.K. Maini, Mathematical oncology: Cancer summed up, Nature, 421 (2003), p. 321.
12. F. Michor, Y. Iwasa, and M.A. Nowak, Dynamics of cancer progression, Nature Rev. Cancer, 4 (2004), pp. 197–205.
13. R.M. Sutherland, Cell and environment interactions in tumor microregions: The multicell spheroid model, Science, 240 (1988), pp. 177–184.
14. T. Udagawa, A. Fernandez, E.-G. Achilles, J. Folkman, and R.J. D’Amato, Persistence of microscopic human cancers in mice: Alterations in the angiogenic balance accompanies loss of tumor dormancy, Faseb J., 16 (2002), pp. 1361–1370.
15. P. Yu, M. Mustata, L. Peng, J. J. Turek, M. R. Melloch, P. M. W. French, and D. D. Nolte, Holographic optical coherence imaging of rat osteogenic sarcoma tumor spheroids, 43(2004), pp. 4862–4873.
16. P. Carmeliet and R.K. Jain, Angiogenesis in cancer and other diseases, Nature, 407 (2000), pp. 249–257.
17. M.V. Blagosklonny, Antiangiogenic therapy and tumor progression, Cancer Cell, 5 (2004), pp. 13–17.
18. N. Mantzaris, S. Webb, and H.G. Othmer, Mathematical modeling of tumor-induced angiogenesis, J. Math. Biol., 95 (2004), pp. 111–187.
19. N. Bellomo and L. Preziosi, Modelling and mathematical problems related to tumor evolution and its interaction with the immune system, Math. Comput. Modelling, 32 (2000), pp. 413–452.
20. S.A. Frank, Somatic mosaicism and cancer: Inference based on a conditional Luria-Delbruck distribution, J. Theoret. Biol., 223(2003 ), pp. 405–412.
21. R.A. Gatenby and B.R. Frieden, Application of information theory and extreme physical information to carcinogenesis, Cancer Res., 62 (2002), pp. 3675–3684.
22. H. Haschke, A biokinetic model to describe consequences of inhibition/stimulation in DNAproofreading and repair–1. Development of the model, J. Theoret. Biol., 212 (2001), pp. 425–456.
23. Y. Iwasa, F. Michor, and M.A. Nowak, Evolutionary dynamics of invasion and escape, J. Theoret. Biol., 226 (2004), pp. 205–214.
24. T.V. Karpinets and B.D. Foy, Model of the developing tumorigenic phenotype in mammalian cells and the roles of sustained stress and replicative senescence, J. Theoret. Biol., 227 (2004), pp. 253–264.
25. N.L. Komarova, A. Sengupta, and M.A. Nowak, Mutation-selection networks of cancer initiation: Tumor suppressor genes and chromosomal instability, J. Theoret. Biol., 223 (2003), pp. 433–450.
26. F. Michor, S.A. Frank, R.M. May, Y. Iwasa, and M.A. Nowak, Somatic selection for and against cancer, J. Theoret. Biol., 225 (2003), pp. 377–382.
27. J.B. Plotkin and M.A. Nowak, The different effects of apoptosis and DNA repair on tumorigenesis, J. Theoret. Biol., 214 (2002), pp. 453–467.
28. R.V. Sole and T.S. Deisboeck, An error catastrophe in cancer?, J. Theoret. Biol., 228 (2004), pp. 47–54.
29. R.K. Jain, Delivery of molecular and cellular medicine to solid tumors, Advances in Drug Delivery Reviews, 46 (2001), pp. 149–168.
30. R.P. Araujo and D.L.S. McElwain, A history of the study of solid tumor growth: The contribution of mathematical modelling, Bull. Math. Biol., 66 (2004), pp. 1039–1091.

31. J.A. Adam, Mathematical models of perivascular spheroid development and catastrophetheoretic description of rapid metastatic growth/tumor remission, Invastion and Metastasis, 16 (1996), pp. 247–267.
32. H.M. Byrne, The role of mathematics in solid tumour growth, Math. Today, 35 (1999), pp. 59–89.
33. A.C. Burton, Rate of growth of solid tumours as a problem of diffusion, Growth, 30 (1966), pp. 157–176.
34. J.A. Adam, A simplified mathematical model of tumor growth, Math. Biosci., 81 (1986), pp. 229–244.
35. J.A. Adam, A mathematical model of tumor growth. II. Effects of geometry and spatial uniformity on stability, Math. Biosci., 86 (1987), pp. 183–211.
36. J.A. Adam, A mathematical model of tumor growth. III. Comparison with experiments, Math. Biosci., 86 (1987), pp. 213–227.
37. J.A. Adam and S.A. Maggelakis, Mathematical models of tumor growth. IV. Effects of a necrotic core, Math. Biosci., 97 (1989), pp. 121–136.
38. J.A. Adam and S.A. Maggelakis, Diffusion regulated characteristics of a spherical perivascular carcinoma, Bull. Math. Biol., 52 (1990), pp. 549–582.
39. Harrison Principiile Medicinei Interne vol. 2,Editura Teora ,1998,pag.1532-1534 40. Prasad S. Adusumilli, Brendon M. Stiles, Mei-Ki Chan, David P. Eisenberg, Zhenkun Yu,
Stephen F. Stanziale, Rumana Huq, Richard J. Wong, Valerie W. Rusch, and Yuman Fong, Real-time diagnostic imaging of tumors and metastases by use of a replication-competent herpes vector to facilitate minimally invasive oncological surgery, The FASEB Journal express article 10.1096/fj.05-5316fje. Published online February 8, 2006.
41. Berthold Block,Guido Schachschd,Hartmut Schmidt Endoscopy of the Upper GI Tract, Edit.Thieme, New York,2004,pag.115-120.
42. Jian-Hua Zhang şi colab., Construction and identification of recombinant vectors carrying herpes simplex virus thymidine kinase and cytokine genes expressed in gastric carcinoma cell line SGC 7901, World Journal Gastroenterology, 2004, 10(1), pag. 26-30.

4.5. Analiza imagistică. Noţiuni generale Prelucrarea digitala a imaginilor reprezinta un domeniu ştiinţific de sine statator, ce are la baza o teorie matematica riguroasa, bine pusa la punct. Prin imagine se întelege o codificare sub forma de hartă a unei suprafete dreptunghiulare ce contine o imagine sub forma discreta, memorata prin valorile intensitatii luminoase (0 sau 1 pentru alb/negru, 0-valoare maximă pentru tonuri de gri şi în format RGB pentru color) a fiecarui pixel din imagine. În concluzie, o imagine se caracterizeaza prin rezolutia sa, prin tipul de codificare a intensitatii luminoase si prin matricea de intensitati, I(x,y). Mai trebuie mentionat ca originea se afla întotdeauna în coltul din stânga-sus si sensurile pozitive ale axelor sunt spre dreapta pentru axa Ox si în jos pentru axa Oy. Filtrarea Operatia de filtrare este utilizata pentru eliminarea zgomotelor si la evidentierea muchilor. In general, se folosesc urmatoarele trei tipuri de filtre:
filtru trece-jos – utilizat pentru eliminarea zgomotelor, spectrul imaginii este uniformizat;
filtru trece-banda – folosit de obicei pentru prelucrarea imaginilor provenite din teledectie;
filtru trece-sus – este utilizat pentru evidentierea contururilor datorita comportarii de tip derivator.
Filtrarea culorilor Pentru transformarea intensitatii codificata în format RGB în nivele de gri se pot folosi urmatoarele formule:
( )BGRgri IIII ++= - medie aritmetica
222BGRgri IIII ++= - norma culorii
Din considerente de conservare a energiei luminoase radiate de fiecare pixel am dezvoltat prezenta metoda. Daca dorim sa conservam energia unui pixel, trebuie sa tinem cont de energia fiecarei componente. Cunoastem ca λR= 632,8 nm, λG = 546,1 nm si λB = 435,8 nm. Presupunând ca R, G si B iau valori în intervalul 0....255, putem calcula energia totala a unui pixel:
⎟⎟⎠
⎞⎜⎜⎝
⎛λ
+λ
+λ
=BGR
totBGRhcE
Aceasta energie trebuie distribuita egal între cele trei componente. Fie K valoarea componentelor. Atunci putem scrie:
⎟⎟⎠
⎞⎜⎜⎝
⎛λ
+λ
+λ
=BGR
tot KhcE 111
Din egalitatea ultimelor doua relatii obtinem:
RGBRBG
GRBRBG BGRK
λλ+λλ+λλλλ+λλ+λλ
=
Dupa obtinerea imaginii în tonuri (nivele) de gri se poate face transformarea urmatoare spre imagine alb-negru. Si aici exista mai multe metode ce difera între ele prin modalitatea de stabilire a nivelului de prag între alb si negru.

Segmentarea Segmentarea este un proces de partitionare a imaginii digitizate in submultimi, prin atribuirea pixelilor indiividuali la aceste submultimi (denumite si clase), rezultand obiecte distincte din scena. Algoritmii de segmentare au la baza, in general, 2 principii:
Dicontinuitate, avand ca principala metoda detectia conturului; Similitudine, cu metoda pragului si metoda regiunilor.
Detecţia contururilor Un operator de contur este un operator matematic, cu extindere spaţială mică, construit pentru a determina existenta unui contur local în funcţia imagine. Exista multe tipuri de operatori de contur, principiul care-i uneşte fiind faptul că ei determină modulul şi uneori direcţia schimbării intensităţii nivelului de gri într-o imagine digitală. Modulul furnizat de aceşti operatori caracterizează severitatea schimbării nivelului de gri de la o vecinatate la alta a imaginii. Cei mai simpli şi cei mai rapizi asemenea operatori de detecţie a conturului se bazează pe utilizarea măştilor de convoluţie spaţială care baleiază întreaga imagine pixel cu pixel, calculând o imagine proporţională cu discontinuitatea existentă în porţiunea de imagine corespunzătoare poziţiei curente a măştii. Muchia este linia ce delimiteaza doua zone ale imaginii cu diferenta mare de contrast. Deoarece caracterizeaza limitele unui obiect, muchiile sunt utile în procesul de segmentare si identificare a obiectelor dintr-o imagine. Ideal, o muchie este reprezentata printr-o discontinuitate a parametrului intensitate, adica o dreapta cu panta infinita. Metoda pragului Pentru multe aplicaţii, datorită diferenţelor semnificative între nivelele de gri ale pixelilor aferenţi obiectului şi respectiv fondului, criteriul de segmentare care poate fi folosit este valoarea nivelului de gri. Metoda care se utilizează în acest caz, foarte rapidă, este denumită segmentare cu prag şi ea implementează o transformare punctuală simpla. Pixelul din punctul de coordonate (i,j) este etichetat ca fiind pixel obiect dacă valoarea sa f(i,j) este mai mare decât un prag. Obţinerea unor bune rezultate cu acesta metodă depinde de modalitatea de alegere a pragului, care poate fi o valoare pentru o imagine data sau o funcţie netedă dependentă de poziţia pixelului curent. A doua metodă, mai complicata, stabileste nivelul intensitatii de prag dintre alb si negru prin analiza locala a imaginii si stabilirea în functie de histograma intensitatilor a nivelului de prag. A treia metoda foloseste un filtru coplanar ce exploateaza coplanaritatea nivelelor de gri ale pixelilor învecinati. Avantajele metodei rezida în înlaturarea zgomotului, netezirea zonala a imaginii si pastrarea muchiilor ascutite. Metoda regiunilor Segmentarea iterativa sau segmentarea prin tehnici de relaxare este o metoda care foloseste procedee probabilistice de clasificare. Acest tip de procesare este fundamental in analiza automata a scenelor si in recunoasterea de modele, unde scopul principal este de a extrage in mod automat date detaliate despre continutul imaginii la nivel obiect. Obiectivul tehnicii de segmentare este de a partitiona o imagine data in regiuni sau componente. Se remarca din analiza precedenta faptul ca putem considera segmentarea fie ca o problema punctuala fie ca una regionala. In prima categorie intra metode care se bazeaza pe examinarea imaginilor pixel cu pixel. In a doua categorie, intra metodele care se bazeaza pe utilizarea informatiilor din imagine in prescrierea vecinatatilor. In ambele cazuri, putem vedea problema ca o luare de decizie in procesul de recunoastere a modelelor, ale carui obiective sunt stabilite in limitele unor regiuni.

Prima operatie este de a localiza granitele sau marginile regiunilor. A doua operatie este de a grupa punctele in regiuni similare, cu alte cuvinte determinarea hotarelor. Histograma. Operaţii Histograma unei imagini o funcţie care indică câţi pixeli au un anume nivel de gri. De obicei, numărul de nivele de gri este 255 (un pixel este reprezentat pe 1 byte). f(g) = p, unde: g = nivel de gri, g între 0 şi 255; p = numărul de pixeli ce au valoarea g. La o analiză a histogramei diverselor tipuri de imagini, se constată că, de obieci, o imagine are mai multe nivele de gri şi prezintă două maxime locale. Folosind aceasta caracteristică se poate face o segmentare cu prag a imaginii iniţiale, alegând pragul ca fiind, de regulă, minimul local dintre cele două puncte de maxim. Astfel, pixelii cu nivel de gri mai mic decât pragul ales pot fi consideraţi ca fiind, de exemplu, fundalul (li se atribuie valoarea corespunzătoare negrului), iar cei pentru care nivelul de gri este mai mare decât pragul sunt pixelii obiectului. Metrici Masurarea distantelor se foloseste, de obicei, pentru extragerea de caracteristici din simboluri, ajutand la impartirea acestora in clase de echivalenta. Matematic, numim d
distanta pe multimea X orice aplicatie d : X×X→ +ℜ care are proprietatile:
d (x,y) = 0 ⇔ x = y, reflexivitate d (x,y) = d(y,x), ∀x,y ∈X, simetrie d(x,y) < d(x,z) + d(z,y), ∀x,y,z ∈X, axioma triunghiului
O metrica generala, din care pot fi extrase prin particularizare multe altele, este metrica Minkowski. Distanta dintre doua puncte a si b, in spatiul euclidian n-dimensional, se obtine cu formula:
( ) ∑−
−=n
i
sbiai
nM xxbad
1,
Cea mai uzuala metrica este cea euclidiana care are forma urmatoare pentru punctele a si b. Se obtine di metrica Minkovski alegand s=2.
( ) ( )∑=
−=n
ibiai
ne xxbad
1
2,
Distanta Manhattan se defineste cu formula urmatoare, alegand s=1 in metrica Minkovski.
( ) ∑=
−=n
ibiai
nm xxbad
1,
Recunoasterea formelor Recunoaşterea formelor reprezintă o modalitate des folosită de a extrage informaţiile din imaginile achiziţionate. Recunoaşterea formelor constă într-o clasificare şi/sau o descriere a conţinutului imagini. Clasificarea constă în atribuirea unei forme necunoscute din imaginea preluată la o clasa dintr-un set predefinit de clase; operaţia de clasificare va produce la ieşire o nouă imagine care reprezintă o hartă a obiectelor aflate în scenă. În noua imagine, valorile pixelilor reprezintă de fapt codurile asociate claselor corespunzătoare. Clasificarea foloseşte metode matematice numite metode de recunoaştere teoretic-decizionale sau statistice, metode ce se bazează pe elemente din teoria deciziilor statistice. Algoritmii de clasificare se bazează în extragerea caracteristicilor (features) pe o măsură a similarităţii (o distanţă, de exemplu). Un pas important în proiectarea sistemelor de

clasificare automată este selecţia caracteristicilor, aceasta deoarece componentele vectorului de caracteristici presupune prezenţa unei cantităţi destul de mari de informaţie. Acestă selecţie este o problemă dependentă de numărul de clase şi de formele analizate. Un clasificator conţine, în general, trei module: modulul de clasificare propriu-zis, un modul de învăţare (presupune prezenţa unui set de eşantioane/imagini de antrenare) şi un modul de selecţie şi extragere a caracteristicilor. Setul de antrenare poate conţine de exemplu o bază de date cu diferite caracteristici ale obiectelor ce pot apare în scenă (semnături spectrale de exemplu sau semnături geometrice). Învăţarea presupune existenţa acestui set de antrenare, set ce cuprinde eşantioane pentru care se ştie apartenenţa claselor; prezenţa setuluide antrenare este necesară în cadrul clasificării supervizate. 4.6. Bibliografie
1. Abeloff and colab.(2004) Clinical Oncology, Churchill Livingstone, Elsevier, 3rd edition;
2. Guyton A., Hall J. (1996) Textbook of Medical Physiology, W. B. Saunders Company, Philadelphia, 9th edition;
3. Iancu I., Iancu E. (2003) Modelare şi simulare în fiziologie, Editura Universitaria, Craiova;
4. Ioan Ispas, Modelarea si modele matematice in recunoasterea obiectelor si clasificarea automata a imaginilor, Catedra de Matematica-Informatica, Universitatea Petru-Maior, Targu Mures;
5. Stefan Holban, Dan Ciresan, (2004) Analiza si prelucrarea imaginii in vederea procesului de recunoastere cu retele neuronale artificiale, Universitatea ”Politehnica” Timisoara, Facultatea de Automatica si Calculatoare, Timisoara;
6. Formatul de imagine bitmap, descriere disponibila online la adresa http://en.wikipedia.org/wiki/Windows_bitmap
7. Formate digitale pentru imagini, descriere disponibila online la adresa http://en.wikipedia.org/wiki/Image_file_formats
8. Modelul aditiv de culori RGB – rosu verde albastru, descriere disponibila online la adresa http://en.wikipedia.org/wiki/RGB
9. International Organization for Standardization (ISO), Limbajul de programare C, standard disponibil online la adresa http://www.iso.org/iso/iso_catalogue/catalogue_tc/catalogue_detail.htm?csnumber=29237
10. Allegro, librarie grafica open-source disponibila online la adresa http://www.talula.demon.co.uk/allegro/


















