
PAOLO DE ANGELIS CERCETĂRI PRIVIND INDICII BIOLOGICI … · Sericicultura este ştiinţa care se...
64
UNIVERSITATEA DE ŞTIINŢE AGRICOLE ŞI MEDICINĂ VETERINARĂ CLUJ-NAPOCA SCOALA DOCTORALĂ FACULTATEA DE ZOOTEHNIE ŞI BIOTEHNOLOGII DISCIPLINA DE TEHNOLOGIA PRODUCŢIILOR APICOLE ŞI SERICICOLE PAOLO DE ANGELIS CERCETĂRI PRIVIND INDICII BIOLOGICI ŞI TEHNOLOGICI LA UNII HIBRIZI AI VIERMILOR DE MĂTASE BOMBYX MORI L. CRESCUŢI ÎN TRANSILVANIA REZUMAT AL TEZEI DE DOCTORAT ÎNDRUMĂTOR Prof.Dr.Ing. Liviu Al. Mărghitaş CLUJ-NAPOCA 2009 1
Transcript of PAOLO DE ANGELIS CERCETĂRI PRIVIND INDICII BIOLOGICI … · Sericicultura este ştiinţa care se...

UNIVERSITATEA DE ŞTIINŢE AGRICOLE ŞI MEDICINĂ VETERINARĂ CLUJ-NAPOCA
SCOALA DOCTORALĂ FACULTATEA DE ZOOTEHNIE ŞI BIOTEHNOLOGII
DISCIPLINA DE TEHNOLOGIA PRODUCŢIILOR APICOLE ŞI SERICICOLE
PAOLO DE ANGELIS
CERCETĂRI PRIVIND INDICII BIOLOGICI ŞI TEHNOLOGICI LA UNII HIBRIZI AI
VIERMILOR DE MĂTASE BOMBYX MORI L. CRESCUŢI ÎN TRANSILVANIA
REZUMAT AL TEZEI DE DOCTORAT
ÎNDRUMĂTOR
Prof.Dr.Ing. Liviu Al. Mărghitaş
CLUJ-NAPOCA
2009
1

REZUMAT
SCOPUL ŞI OBIECTIVELE CERCETĂRILOR
Scopul cercetărilor întreprinse în cei doi ani experimentali a fost ierarhizarea raselor şi hibrizilor după principalii indici biologici ai larvelor, după indicii biologici ai gogoşii crude şi după indicii tehnologici ai gogoşii uscate şi ai fibrei de mătase.
Obiectivele cercetării au fost legate de următoarele aspecte: I. Studiul comparativ al raselor parentale şi al hibrizilor simpli; II. Studiul comparativ al raselor parentale şi al trihibrizilor III. Studiul comparativ al raselor parentale şi al tetrahibrizilor S-au studiat incubaţia şi ecloziunea, indicii biologici ai larvelor, ai gogoşilor
crude, precum şi indicii tehnologici ai gogoşii uscate şi a firului de mătase. Rezultatele obţinute în cei doi ani experimentali pentru cele patru rase experimentale, doisprezece hibrizi simpli, opt trihibrizi şi doi tetrahibrizi au fost utilizate pentru studiu comparativ între rasele parentale şi hibrizii amintiţi au fost urmărite diferenţele evolutive dintre aceştia în cei doi ani experimentali, în vederea întocmirii unei ierarhizări a raselor şi hibrizilor viermilor de mătase în funcţie de principalii indici biologici şi tehnologici amintiţi.
Indicii studiaţi pentru rasele parentale şi hibrizii rezultaţi din acestea sunt:
1. Incubaţia ouălor şi ecloziunea larvelor; 2. Indicii biologici ai larvelor:
♦ Masa larvelor ♦ Lungimea larvelor ♦ Durata stadiului larvar ♦ Masa glandelor sericigene ♦ Procentul de îngogoşare
3. Indicii biologici ai gogoşii crude: ♦ Masa gogoşii crude ♦ Axul longitudinal al gogoşilor ♦ Axul transversal al gogoşilor ♦ Masa incartamentului mătăsos ♦ Procentul de mătase din gogoaşă
4. Indicii tehnologici ai gogoşii uscate şi ai fibrei: ♦ Masa gogoşii uscate ♦ Lungimea firului de mătase ♦ Masa firului de mătase ♦ Greutatea deşeului din gogoaşă
2

REZUMAT
CAPITOLUL I STADIUL ACTUAL AL CERCETĂRILOR ŞI ISTORICUL
CREŞTERII VIERMILOR DE MĂTASE BOMBYX MORI L. ÎN ROMÂNIA ŞI PE MAPAMOND
Sericicultura este ştiinţa care se ocupă cu studiul biologiei şi tehnologiei
reproducerii, creşterii şi ameliorării tuturor speciilor sericicole care produc gogoşi în scopul obţinerii mătăsii naturale. Devidarea gogoşilor, prelucrarea firului, toarcerea şi executarea ţesăturilor formează obiectul industriei mătăsii.
Dintre toate speciile de viermi de mătase care ţes gogoşi din fir de mătase, o pondere de 90% este deţinută de viermele de mătase al dudului, urmată de viermele de mătase al ricinului şi al stejarului, aria de răspândire a acestor specii fiind legată de zona de răspândire a speciilor vegetale furnizoare de hrană. Aria de creştere a viermilor de mătase este în general mult mai restrânsă decât cea a speciilor care constituie baza furajeră a acestora.
Numele de sericicultură vine de la cuvântul latin “ser-seris” care înseamnă mătase, iar denumirea ştiinţifică a viermelui de mătase al dudului este Bombyx mori L.
Sericicultura reprezintă totodată şi o îndeletnicire rentabilă, uşoară, ce poate fi practicată atât de copii cât şi de vârstnici, creşterile se pot face în construcţii simple, amenajări sau alte spaţii gospodăreşti, ce nu se folosesc pe timpul verii, iar perioada de creştere a larvelor este scurtă, fiind în medie de cca. 30 de zile (Yokoyama T., 1973; Ifrim P., 1985; Mărgitaş L. Al., 1995; Savel I., 1998).
Ţara de origine a mătăsii o reprezintă China. Cercetările istorice şi arheologice realizate aici au pus în evidenţă faptul că, în jurul orăşelului chinez Schengze, din regiunea Wuxion, provincia Jiangsu, locuitorii acestor meleaguri cultivau duzi şi creşteau viermi de mătase; cunoşteau meşteşugul obţinerii firelor şi al ţesăturilor din mătase naturală.
Arta creşterii viermilor de mătase şi utilizarea mătăsii este atribuită tinerei prinţese chineze Si-Ling-Schi, principala dintre soţiile celebrului împărat Hoang-Ti sau Galben Impărat (2699- 2599 î.Hr.).
Pentru chinezii vechi, sericicultura a devenit o preocupare sacră, ce constituia un secret, protejat prin decrete imperiale. Divulgarea acestui secret se pedepsea cu moartea. Timp de aproximativ 3000 de ani ei au ştiut să păstreze acest secret biologic, cât şi producţia şi comerţul cu mătase naturală între graniţele Chinei. Chinezii aduceau stofele de mătase “pe drumul mătăsii” din China pe pieţele Turkistanului Oriental, unde negustorii mediteraneeni veneau să le cumpere.
Răspândirea secretului în lume privind producerea mătăsii a avut loc în jurul anului 200 î. Hr., când emigranţii chinezi au ajuns pentru prima dată în Coreea, ducând o dată cu ei şi secretul producerii mătăsii. De aici sericicultura s-a răspândit apoi în toată lumea asiatică, iar în anul 552 d.Hr. pe vremea împăratului Iustinian, doi călugări persani, după ce şi-au însuşit tehnica creşterii larvelor şi a prelucrării mătăsii au adus ascunse în bastoane de bambus ouăle fluturelui de mătase la Constantinopol, de unde s-a răspândit creşterea viermilor de mătase şi devidarea gogoşilor în tot Imperiul Bizantin. Apoi arabii au răspândit această îndeletnicire în Caucaz şi în Africa de Nord până în Spania, de unde a trecut apoi în Sicilia, în Italia
3

REZUMAT
de sud şi de nord, apoi în Franţa, unde primii duzi au fost cultivaţi în jurul anului 1260.
În perioada actuală sericicultura este răspândită în 36 de ţări, iar producţia mondială de gogoşi este de cca. 450.000 tone şi se realizează în principal în China (41%), Japonia (23%), India (9%), Coreea de Sud (9%), Comunitatea Statelor Independente (9%) etc. (Mǎrghitaş L. Al., şi col., 2003). Primul loc în producţia de mătase este deţinut de China (27 000 tone), urmată de Japonia (9 200 tone) şi India (9 000 tone), aceasta din urmă manifestând tendinţa de a deveni al doilea producător de mătase din lume. Datorită condiţiilor favorabile pentru cultura dudului, în ţara noastră sericicultura s-a dezvoltat foarte repede, iar sămânţa indigenă de viermi de mătase fiind indemnă de pebrină, a fost mult apreciată peste hotare (Mǎrghitaş L. Al. şi col., 2003). Astfel, din datele existente rezultă că în anul 1860 ţara noastră a exportat impresionanta cantitate de 6 064 kg sămânţă de viermi de mătase. În acest sens, în perioada 1840-1860 printre măsurile luate pentru dezvoltarea sericiculturii a fost şi înfiinţarea pepinierelor pentru producerea materialului destinat plantaţiilor de dud necesare asigurării bazei furajere. În ultimii 50 de ani, producţia de gogoşi a ţării noastre a crescut continuu, de la aproximativ 500 tone (1944) la circa 2000 tone (1989). După 1990, datorită crizei economice specifice perioadei de tranziţie înspre economia de piaţă producţia de gogoşi a scăzut an de an, ajungând în 1998 la o producţie de doar 3 tone gogoşi crude pe an. În momentul de faţă producţia se situează la nivele îngrijorătoare. În ceea ce priveşte producţia de mătase brută, aceasta a suferit o puternică regresie, reducându-se de la 120 tone (nivel atins în 1989) la 20 tone (1991); 6 tone (1995) şi 2 tone (2001). Această regresie s-a datorat pe de altă parte şi încetării activităţii filaturii de la Lugoj, zonă geografică cu un mare potenţial sericicol (Paşca I., 2004). În ultimii ani, în România s-au întreprins cercetări laborioase şi cu bune rezultate privind maladiile viermelui de mătase Bombyx mori L., încât au apărut o serie de publicaţii cu astfel de cercetări care urmăresc, în mod deosebit, poliedria, numită şi gălbinarea sau icterul viermelui de mătase, maladie de natură virală.
În momentul de faţă, sericicultura atât în România cât şi la nivel mondial, trece printr-o perioadă de criză, producţia de gogoşi de mătase fiind pe cale de dispariţie. La nivelul ţării noastre, sericicultura a decăzut mult o dată cu închiderea filaturii de la Lugoj. Deşi în ultimii ani, sericicultura a suferit această „decădere din drepturi”, la nivel de cercetare ea şi-a continuat activitatea, fiind chiar productivă din acest punct de vedere. Pe lângă munca de conservare a băncii de gene existente în prezent, s-au creat noi rase şi hibrizi de viermi de mătase folosind fondul genetic existent în prezent la S.C. SERICAROM S.A. Băneasa (peste 60 de rase de viermi de mătase).
4

REZUMAT
CAPITOLUL II CLASIFICAREA TAXONOMICĂ A VIERMILOR DE MĂTASE
ŞI CICLUL EVOLUTIV AL SPECIEI BOMBYX MORI L.
2.1. CLASIFICAREA TAXONOMICǍ A SPECIEI
Viermele de mătase al dudului face parte din: Încrengătura: Artropode Subâncrengătura: Tracheate Clasa: Insecta (Hexapoda) Subclasa: Pterigote Secţiunea: Neoptere Subsecţiunea: Metamorfe Ordinul: Lepidoptere superioare, Seria: Metaheterocere sau Lepidoptere nocturne superioare Familia: Bombycidae Genul: Bombyx Specia: Bombyx mori
2.2. SPECII DE VIERMI DE MĂTASE
Cele mai cunoscute specii de viermi de mătase, ale căror gogoşi pot fi utilizate pentru producerea mătăsii (fir sau vată) sunt: Bombyx mori L., Anthereae yammai Guer L., Anthereae assaamensis L., Anthereae pernyL., Theophyla mandarina L., Bombyx arrindia L., , Samia coecropia L., Saturnia pyri L., Saturnia pyretorum L., Attacus atlas L., Philosamia cynthia ricini L., Lasiocampa atus L., Bodecera madagascarensis L., etc. Dintre toate acestea, cea mai răspândită este specia Bombyx mori L. (care este considerată şi cea mai valoroasă dintre toate), apoi Anthereae perny (viermele de mătase al stejarului, ale cărui larve se hrănesc cu frunze de stejar) şi Philosamia cynthia ricini, (viermele de mătase al ricinului, ale cărui larve se hrănesc cu frunze de ricin) celelalte specii trăiesc în stare sălbatică, în China, India şi în alte ţări, unele fiind folosite pentru încrucişări cu diferite rase de viermi domestici (au rol în ameliorarea raselor).
2.3. CICLUL EVOLUTIV AL SPECIEI BOMBYX MORI L.
Viermele de mătase al dudului sau fluturele de mătase (Bombyx mori L.) face parte din grupa insectelor cu metamorfoză completă, în ciclul său evolutiv trecând succesiv prin 4 stadii diferite: ou (sau sămânţă), larvă (vierme sau omidă), crisalidă (nimfă sau pupă) şi fluture (sau adult), având una (rase monovoltine) sau mai multe generaţii (rase polivoltine) pe an. După fecundare au loc procese de embriogeneză timp de trei zile, apoi începând diapauza. Durata acesteia variază în funcţie de rasă, factorii genetici şi cei
5

REZUMAT
de mediu, la rasele noi formate fiind de 9-10 luni, durând în general din luna iulie până în aprilie. Ouăle menţinute în condiţii optime incubează timp de 10-14 zile, proces care se încheie cu ecloziunea larvelor. Stadiul larvar variază în funcţie de rasă, fiind cuprins între 25 şi 35 zile, larvele evoluând prin cinci vârste, delimitate de perioade de repaus, numite „somnuri”. Pe perioada acestor somnuri larvele nu se hrănesc şi are loc schimbarea tegumentului în urma năpârlirilor.
CAPITOLUL III
PROBLEMATICI ALE AMELIORĂRII, HIBRIDĂRII ŞI HETEROZISULUI LA VIERMII DE MĂTASE AI DUDULUI
3.1. SELECŢIA
Selecţia reprezintă prima şi cea mai importantă metodă de ameliorare, fiind
definită ca un complex de lucrări ce constau în alegerea şi împerecherea organismelor în scopul îmbunătăţirii raselor existente sau a creării de noi rase. Prin selecţie se reţin pentru reproducţie indivizii corespunzători pentru realizarea scopului propus. Aşadar, selecţia reprezintă o “discriminare reproductivă” (Bura M., 1992) realizată de către om prin alegerea unor genotipuri şi îndepărtarea altora. Prin lucrările de selecţie se urmăreşte obţinerea unor forme superioare celor existente la un moment dat, sub aspectul unuia sau mai multor caractere biologice, tehnologice sau economice. Specificul selecţiei este determinat de o serie de particularităţi ale viermelui de mătase, cum ar fi: prolificitatea mare, ciclul biologic scurt ce permite obţinerea mai multor generaţii pe an, durata scurtă a vieţii fluturelui care asigură o singură descendenţă şi imposibilitatea împerecherii descendenţei cu părinţii.
3.2. HIBRIDAREA
Hibridarea sau încrucişarea reprezintă o a doua etapă a procesului de ameliorare. În acest scop, indivizii selecţionaţi pentru reproducţie, aparţinând a două sau mai multe rase se împerechează dirijat între ei, în vederea asigurării unei combinaţii favorabile fixării caracterelor dorite.
Încă din secolul al XVIII-lea s-a observat că produşii obţinuţi în urma încrucişărilor (hibrizii) depăşesc de cele mai multe ori valoarea părinţilor, atât din punct de vedere al vitalităţii şi viabilităţii cât şi al însuşirilor productive.
3.3. CONSANGVINIZAREA
Consangvinizarea reprezintă procedeul prin care se împerechează între ei
indivizi înrudiţi ce aparţin aceleiaşi rase, în scopul fixării unor caractere valoroase sau formării de linii consangvine. Ea reprezintă una din metodele utilizate în procesul de creare şi ameliorare a unor tipuri şi rase de animale.
6

REZUMAT
Consangvinizarea permite fixarea ereditară şi intensificarea unor calităţi deosebite, eliminarea din cadrul rasei a unor indivizi nedoriţi, precum şi mărirea puterii de transmitere ereditară a reproducătorilor.
3.4. VIGOAREA HIBRIDĂ PENTRU DIFERITE CARACTERE
În timpul diferitelor studii efectuate pe viermele de mătase al dudului s-a observat că anumite caractere precum numărul de ouă/pontă, procentul de ecloziune, durata stadiului larvar, numărul de larve adulte viabile, numărul de gogoşi, greutatea gogoşii, masa incartamentului mătăsos sau lungimea firului de mătase nu manifestă fenomenul de heterozis cu aceeaşi intensitate.
3.5. HETEROZISUL ÎN FUNCŢIE DE DIFERITE SISTEME DE ÎNCRUCIŞARE
Hirobe T., (1985) observă faptul că valoarea heterozisului are cele mai
ridicate valori în cazul încrucişărilor simple dintre populaţii distincte genetic şi descreşte progresiv în următoarea ordine: F1>F2>F3>F4….În cazul viermelui de mătase, fenomenul de heterozis dispare complet în generaţia F14. Viermii de mătase “pot fi combinaţi” în urma câtorva tipuri de încrucişări dintre care amintim: încrucişările simple (AxB), încrucişări triple [(Ax B) x C] şi încrucişările cvadruple [(Ax B) x (Cx D)]. Tipurile de încrucişare practicate influenţează valoarea heterozisului, hibrizii rezultaţi în urma încrucişărilor simple prezentând de obicei o vigoare hibridă mai mare comparativ cu cei obţinuţi prin încrucişare triplă sau cvadruplă (Sohn K.W., 1983; Das S.K., 1994; Singh R. şi colab., 1998).
Studiile efectuate asupra variaţiei caracterelor cantitative la rasele parentale şi hibrizii obţinuţi prin diferite sisteme de încrucişare arată că variabilitatea pentru caracterele cantitative este mai mică la hibrizii obţinuţi prin încrucişări simple decât la rasele parentale şi decât la hibrizii obţinuţi prin încrucişări triple sau cvadruple (Watanabe 1961; Sohn, 1983; Yokoyama, 1973).
3.6. HETEROZISUL LA ÎNCRUCIŞĂRILE RECIPROCE
În cazul viermilor de mătase, o serie de caractere cantitative sunt strâns
legate de natura voltinismului şi de perioada de maturitate, caractere despre care se ştie că sunt legate de sex (Nagatomo, 1942; Morohoshi, 1949; Nakada, 1970, 1972; Tazima, 1988). Expresia heterozisului poate varia destul de mult în cazul încrucişărilor reciproce ale raselor viermilor de mătase.
Este cunoscut că dacă un hibrid prezintă un heterozis mai ridicat pentru un anumit caracter, heterozisul său poate să scadă sub valoarea hibridului său reciproc în cazul unui alt caracter diferit de primul.
3.7. HETEROZISUL ŞI MEDIUL DE VIAŢĂ
7

REZUMAT
Lerner (1954) în urma numeroaselor cercetări întreprinse observă că heterozigoţii sunt mai bine protejaţi decât homozigoţii împotriva variaţiilor mediului ambiant (variaţii de temperatură, umiditate, aerisire, spaţiu de creştere etc.). Aşadar, heterozigoţii ar prezenta mai puţine variaţii decât rasele parentale pentru un acelaşi mediu sau pentru medii diferite, ei fiind mult mai uniformi din acest punct de vedere. Gradul de manifestare al heterozisului obţinut pentru o încrucişare dată poate fi deci influenţat de mediu (Barlow 1981).
Conceptul după care interacţiunile genotip - mediu afectează amplitudinea heterozisului a fost abordat de către numeroşi cercetători la diferite specii de animale (Sang, 1964; Knight, 1973; Orozco, 1976; Griffing şi Zsiros, 1978; Ruban şi col., 1988; Ehiobu şi Goddard, 1989; Santiago şi col., 1989).
3.8. HETEROZISUL ÎN FUNCŢIE DE TIPUL DE VOLTINISM
Cele mai multe dintre studiile realizate până în prezent (Hirobe, 1985; Udupa şi Gowda, 1988; Nagaraju, 1990) arată că hibrizii obţinuţi din încrucişări triple şi cvadruple sunt inferiori hibrizilor obţinuţi din încrucişări simple. Rezultatele obţinute de Nagaraju (1990) pentru hibrizii obţinuţi din încrucişări triple (polivoltini x polivoltini) x bivoltini şi (polivoltini x bivoltini) x bivoltini se alătură şi ele acestor observaţii.
O astfel de diferenţă între hibrizii obţinuţi din încrucişări simple, triple şi cvadruple poate fi interpretată ca o consecinţă a faptului că unul din părinţii implicaţi în încrucişarea triplă şi ambii părinţi în încrucişarea cvadruplă sunt indivizi F1 ei înşişi. Mai mult, populaţia obţinută din hibrizii triplii sau cvadruplii este un amestec de genotipuri care se poate obţine prin încrucişări simple.
Hinajawa şi Ohtsuka (1975), au folosit pentru prima dată un sistem de anticipare a performanţei hibrizilor rezultaţi în urma încrucişărilor triple şi cvadruple din performanţa hibrizilor rezultaţi din încrucişările simple.
CAPITOLUL IV
RASE ŞI HIBRIZI DE VIERMI DE MĂTASE
4.1. DESCRIEREA GENERALĂ A RASELOR ŞI HIBRIZILOR SPECIEI BOMBYX MORI L.
Prin noţiunea de „rasă” de viermi de mătase se defineşte un grup de indivizi
izolaţi reproductiv, ce prezintă anumite cerinţe faţă de mediu. În cadrul raselor, se pot diferenţia linii, reprezentând subpopulaţii genetice, grupuri de indivizi izolate reproductiv, care evoluează sub influenţa selecţiei, eventual a derivei genetice şi care diferă între ele prin caractere morfologice sau productive. Ca provenienţă, rasele de viermi de mătase reprezintă entităţi biologice sau selecţionate din populaţii locale cu un grad mare de adaptabilitate la diferite condiţii de mediu.
Cele aproximativ 60 de rase de viermi de mătase ai dudului sunt clasificate în funcţie de o serie de criterii, cum ar fi: originea raselor, culoarea şi forma gogoşii,
8

REZUMAT
voltinismul etc. Astfel, în funcţie de voltinism (numărul de generaţii dintr-un an), se deosebesc:
- rase monovoltine (cu o singură generaţie pe an); - rase bivoltine (cu două generaţii pe an); - rase polivoltine (cu 5- 8 generaţii pe an).
După origine, rasele se clasifică în următoarele grupe: - rase japoneze; - rase chineze; - rase europene; - rase tropicale;
După forma gogoşilor, se deosebesc: - rase cu gogoşi ovale; - rase cu gogoşi cilindrice; - rase cu gogoşi sferice; - rase cu gogoşi fusiforme; - rase cu gogoşi centurate; După culoarea gogoşilor, se cunosc: - rase cu gogoşi albe; - rase cu gogoşi galbene; - rase cu gogoşi aurii; - rase cu gogoşi verzi; - rase cu gogoşi roz. În funcţie de gradul de hibridare, se disting următoarele tipuri de hibrizi: - hibrizi simpli - se obţin prin încrucişarea directă şi reciprocă a două rase
de viermi de mătase. - hibrizii tripli – se obţin prin încrucişarea directă şi reciprocă dintre un
hibrid simplu şi o rasă de viermi de mătase. - hibrizii cvadrupli - se obţin prin încrucişarea directă şi reciprocă dintre
doi hibrizi simpli de viermi de mătase.
4.2. CARACTERISTICILE PRODUCTIVE ALE RASELOR SPECIEI BOMBYX MORI L.
În cadrul acestui subcapitol sunt prezentate caracteristicile productive ale următoarelor rase: Alb Cislău, Băneasa 75, Alb Chinez 29, Alb Orşova, Alb Băneasa, S – 76, Alb Băneasa 1, Hebar 2/1, Hebar 1/18, EM5, SK2/F, N5, Saniş 8, Belokokonnaia 1, J90.
4.3. CARACTERISTICI PRODUCTIVE ALE HIBRIZILOR ŞI POLIHIBRIZILOR SPECIEI BOMBYX MORI L.
Datele referitoare la însuşirile biologice şi caracterele tehnologice ale hibrizilor şi polihibrizilor sunt extrem de limitate în literatura de specialitate, în acest capitol fiind descrişi următorii hibrizi consacraţi: hibridul simplu Băneasa 28, hibridul simplu Băneasa 29, hibridul simplu Cislău 8, hibridul simplu Super 1,
9

REZUMAT
tetrahibridul Băneasa 28 x Băneasa 29, tetrahibridul Băneasa 8 x Băneasa 9, tetrahibridul Băneasa 75 x Băneasa 9, tetrahibridul Băneasa 76 x Băneasa 4, tetrahibridul Cislău 8 x Cislău 1.
CAPITOLUL V TEHNOLOGII DE CREŞTERE A VIERMILOR DE MĂTASE AI
DUDULUI ÎN CONDIŢIILE PEDOCLIMATICE ALE TRANSILVANIEI
Tehnologiile de creştere a viermilor de mătase ai dudului cuprind cu
precădere aspecte ale incubaţiei ouălor, ecloziunii, condiţiilor de microclimat şi hrănirii acestora.
Incubaţia este un proces complex reprezentat de dezvoltarea embrionului din ou, şi influenţat de asigurarea unor condiţii optime de temperatură, umiditate, lumină şi aer. Ea poate fi naturală sau artificială.
Pregătirea spaţiilor se realizează în anotimpul de primăvară; sunt necesare o curăţire şi o dezinfecţie a spaţiului şi a inventarului utilizat. După curăţarea propriu-zisă a spaţiului inventarul de lemn se spală cu soluţie de sodă caustică 2-5%, după care acesta se clăteşte cu apă curată şi se usucă. Dezinfecţia se realizează cu formol 3%, după o etanşeizare a spaţiului destinat incubaţiei. În condiţii de temperatura de 23 – 250C, se aplică soluţia de formol cu ajutorul vermorelului, si se lasă închisă camera timp de 2 zile. După această dezinfecţie se execută văruirea, spălarea uşilor şi geamurilor, precum şi a pardoselii, continuându-se cu dezinfecţia în jurul clădirii.
Tehnologia propriu-zisă a incubaţie constă în introducerea ouălor în casetele de incubaţie în straturi de până la 2 mm, peste care se aşează o plasă de bumbac, o coală de hârtie perforată şi un pelur, acestea fiind fixate pe o ramă.
Temperatura are un rol deosebit de important, influenţând dezvoltarea embrionului (Ifrim, 1998; Reddy şi col., 2002), utilizându-se metoda temperaturii constante şi metoda ridicării treptate a temperaturii.
Perioada de incubaţie durează în mod normal 12-14 zile şi este strâns legată de condiţiile de microclimat arătate mai sus.
Ecloziunea este procesul prin care larva părăseşte oul prin micropil şi durează 2-3 zile. Cu 2-3 zile înainte de ecloziune larva consumă învelişul seros al oului, acesta se decolorează, iar în interiorul lui se aud pocnituri. Ecloziunea are loc între orele 5 şi 11. În prima zi apar aşa-numiţii „spioni”, care sunt primele larve eclozionate şi care nu se menţin pentru creştere. În prima zi eclozionează 43-44% din larve, în a doua zi 40-42%, iar restul în cea de-a treia zi. Larvele eclozionate după a treia zi se îndepărtează de la creştere, în vederea întocmirii unor loturi de creştere cât mai uniforme.
Un rol important în tehnologiile de creştere a viermilor de mătase îl ocupă asigurarea spaţiului de creştere. Valorificarea localurilor de creştere se realizează prin mărirea spaţiului util şi prin creşterea pe verticală a larvelor. Paturile de creştere utilizate prezintă o mare varietate de tipuri, determinată fiind de tradiţie şi de posibilităţile crescătorilor de viermi de mătase.
Larvele tinere sunt mai rezistente la temperaturi ridicate, o dezvoltare normală a larvei având loc numai în condiţiile respectării următoarelor temperaturi:
10

REZUMAT
26-270C pentru vârsta I şi II-a, 24-250C pentru vârsta a III-a, iar pentru vârsta a IV-a şi a V-a 23-240C. Depăşirea acestor valori duce la intensificarea proceselor fiziologice, având acţiune nefavorabilă asupra vitalităţii organismului (Rajasekhar R., 1993; Tsurumaki J. şi col., 1999).
Suprafaţa de creştere este un alt factor important al tehnologiei creşterii viermilor de mătase şi aceasta trebuie să asigure o hrănire uniformă larvelor. Pentru 1 gram de ouă este necesară o suprafaţă de 0.1 m2 în vârsta I, 0.2 m2 în vârsta a II-a, 0.5 m2 în vârsta a III-a, 1.5 m2 în vârsta a IV-a şi 3.0 m2 în vârsta a V-a.
Recoltarea frunzelor se face dimineaţa sau seara pe răcoare, evitându-se perioadele calde ale zilelor, întrucât frunzele au valoare nutritivă mai scăzută (Vlaic, B., 2007). Administrarea pe lăstari se face de 3-4 ori pe zi, frunzele ofilindu-se mai greu.
Mărghitaş L. Al., (1995) şi Morar R., (1999) recomandă pentru creşterea larvelor provenite dintr-un gram de ouă 35-40 kg de frunză de dud, administrată astfel: la vârsta I 0.2 kg, la vârsta a II-a 0.6 kg, la vârsta a III-a 2 kg, la vârsta a IV-a 5 kg, iar la vârsta a V-a 26-31 kg.
Nerespectarea acestor condiţii poate avea consecinţe grave, năpârlirea devenind imposibilă, producându-se moartea larvelor. Prima hrănire după năpârlire trebuie făcută cu grijă întrucât noul tegument al larvelor este foarte sensibil şi este de evitat rănirea acestuia.
CAPITOLUL VI
HRANA NATURALĂ ŞI DIFERITE ADAOSURI ADMINISTRATE VIERMILOR DE MĂTASE
Nutriţia este un factor important al creşterii viermilor de mătase. Cercetările privind nutriţia viermilor de mătase au cunoscut un adevărat progres ca urmare a apariţiei şi folosirii dietei artificiale, ceea ce a dus la obţinerea de importante beneficii în industria textilă. Singurul stadiu de dezvoltare în care viermii de mătase consumă hrană este cel larvar. În acest stadiu sunt acumulate rezerve de substanţe nutritive suficiente pentru faza de crisalidă şi fluture.
Principala sursă de hrană pentru specia Bombyx mori o reprezintă frunzele de dud, acestea conţinând cantităţi suficiente de proteine, glucide, grăsimi, săruri minerale, apă. Modul de asimilare a acestora depinde de condiţiile de microclimat asigurate larvelor, dar şi de calitatea frunzei de dud administrate.
CAPITOLUL VII
MATERIALE ŞI METODE
7.1. MATERIALE EXPERIMENTALE
7.1.1. Materialul biologic Materialul biologic folosit în cadru experimentelor în cei doi ani a fost constituit din 4 rase de viermi de mătase: Alb de Băneasa (AB), Băneasa 1 (B1), Alb de Cislău (AC), Alb chinezesc linia 29 (AC29), provenite din fondul genetic al
11

REZUMAT
speciei Bombyx mori L. furnizate de S.C. Sericarom Băneasa S.A (Bucureşti). Au fost studiaţi de asemenea următorii hibrizi:
- 4 hibrizi simpli: ABxAC, ACxAB, B1xAC29 şi AC29xB1; - 8 trihibrizi: AC29x(AB x B1); (AB x B1)xAC29; (AB x B1)xAC; ACx(AB x
B1), (AC x AC29)xB1; B1x(AC x AC29); (AC29 x AC)xAB şi ABx(AC29 x AC);
- 2 tetrahibrizi: (AB x B1) x (AC x AC29); (AC x AC29) x (AB x B1); Creşterile raselor şi hibrizilor mai sus amintiţi au fost realizate în amândoi
anii experimentali (2007 şi 2008) în două repetiţii (C1 şi C2), utilizând 300 larve/repetiţie, iar datele redate în tabelele şi graficele din capitolele următoare reprezintă media valorilor a două repetiţii în acelaşi an experimental.
Numărul de indivizi studiaţi a fost diferit în funcţie de însuşirile analizate. Astfel pentru parametrii biologici ai pontelor (prolificitate, număr de ouă eclozionate şi procent de ecloziune) rezultatele exprimă media a 5 ponte diferite, atât la rasele cât şi la hibrizii studiaţi; masa larvelor a fost studiată pentru 20 de exemplare, lungimea larvelor pentru 10, masa glandei sericigene a fost determinată pe glandele extrase de la 15 larve, masa gogoşii crude pe câte 100 de larve de fiecare sex, masa incartamentului şi procentul de mătase din gogoaşă pe 15 gogoşi, iar axul longitudinal şi transversal precum şi masa gogoşii uscate, lungimea firului de mătase, masa firului şi a deşeului din gogoaşă pe câte 10 larve.
7.1.2. Materialele utilizate la organizarea seriilor de creştere
Creşterile viermilor de mătase au fost posibile datorită existenţei plantaţiei de duzi din specia Morus alba, soiul Ucraina 107 în cadrul Universităţii de Ştiinţe Agricole şi Medicină Veterinară Cluj-Napoca. Acest soi de origine ucraineană este rezistent la ger, pretându-se la exploatare în zona climaterică a Transilvaniei, fiind totodată foarte rezistent la bacterioze.
Din punct de vedere fizic şi calitativ frunza administrată a fost de calitate superioară. Recoltarea s-a făcut pe timp răcoros, dimineaţa şi seara, pentru că în aceste momente ale zilei frunza are o valoare nutritivă mai ridicată. Administrarea acesteia s-a făcut de 4 ori/zi.
Păstrarea frunzei fost făcută într-o încăpere răcoroasă (100C), iar înainte de administrare a fost introdusă în camera de pregătire a hranei la temperaturi de 250C. Pentru cele mai tinere larve frunza a fost tăiată înainte de administrare la o grosime de aproximativ 0.5cm. Pentru larvele din vârsta a II-a grosimea frunzelor a fost de 1-2 cm, iar pentru larvele din vârsta a III-a de 2-3 cm. Înaintea intrării în somn a larvelor frunzele nu au fost tăiate pentru a evita ofilirea prematură a acestora şi pentru a evita riscul îmbolnăvirii larvelor.
Pentru ca frunza să rămână proaspătă o perioadă mai lungă de timp, larvelor din vârstele înaintate (a IV-a şi a V-a) li s-au administrat frunze nedesprinse de pe ramuri.
Spaţiu destinat creşterii seriilor experimentale a avut o suprafaţă de 40 mp, cu amplasare nordică.
Înainte de începerea creşterii seriilor experimentale sala de creştere, modulul de creştere şi inventarul necesar au fost spălate mecanic şi apoi
12

REZUMAT
dezinfectate cu vapori de formaldehidă. Pentru o bună realizare a dezinfecţiei au fost izolate toate ferestrele şi uşile, iar pentru sporirea eficacităţii acesteia temperatura în interior a fost ridicată la 300C, iar umiditatea relativă a aerului la 85%. Vaporii au fost lăsaţi să acţioneze timp de 48 de ore, după care spaţiul astfel dezinfectat a fost lăsat să se aerisească pentru 3 zile. La intrarea în spaţiul de creştere a fost montat un dezinfector conţinând soluţie de hidroxid de sodiu 2%.
În spaţiul destinat creşterii larvelor viermilor de mătase au fost amplasate stelaje suprapuse pe patru nivele (Figura 1) cu picioare fixe şi paturi de creştere mobile. Distanţa dintre paturile de creştere este de 50 cm, iar cea dintre primul pat şi pardoseală este de 60 cm, pentru a echilibra diferenţele de temperatură dintre primul şi ultimul pat de creştere asigurând astfel condiţii uniforme de temperatură pentru toţi indivizii studiaţi. Pe paturile de creştere s-a pus hârtie albă, iar apoi s-au aşezat larvele eclozionate.
Figura 1. Modul sericicol familial (MSF)(original)
Schimbarea aşternutului s-a realizat cu ajutorul hârtiei perforate. Ochiurile
făcute în hârtia de aşternut prezintă diametre diferite, în funcţie de vârsta larvelor pentru care este utilizată hârtia perforată: 6 mm până a vârsta a III-a, 10 mm pentru vârsta a IV-a şi 13-15 mm pentru ultima vârstă a stadiului larvar. În perioada de somn larvele nu se mai hrănesc, ele revenind la comportamentul alimentar iniţial atunci când 90% din larve au ieşit din perioada de somn. Larvele care din diferite motive nu-şi pot schimba tegumentul vor muri, întrucât vechiul tegument, de dimensiuni mai mici decât cele necesare, nu le permite creşterea. Un important factor în perioada de creştere a larvelor îl constituie densitatea acestora. Astfel, pe perioada desfăşurării experimentului s-a asigurat un spaţiu între două larve egal cu grosimea acestora.
13

REZUMAT
7.2 METODA EXPERIMENTALĂ
Experienţele s-au desfăşurat în laboratorul Disciplinei de Tehnologia Producţiilor Apicole şi Sericicole din cadrul Universităţii de Ştiinţe Agricole şi Medicină Veterinară Cluj-Napoca şi la S.C. Sericarom Băneasa S.A. (Bucureşti), în perioada 2007 – 2008.
Larvele viermilor de mătase din variantele experimentale mai sus menţionate au fost hrănite cu frunză recoltată din plantaţia intensivă de dud cu trunchi înalt pe care o deţine Universitatea, plantaţie care se întinde pe o suprafaţă totală de 2 ha şi care este aflată în incinta Fermei Didactice Cluj-Napoca.
Datele cu privire la indicii urmăriţi (detaliaţi în Capitolul 7.1.) au fost înregistrate treptat pe perioada desfăşurării experimentelor în anii 2007 şi 2008, prin măsurători şi cântăriri electronice, iar interpretarea statistică a datelor a fost efectuată conform metodelor statistice recunoscute (Tacu, A, 1968; Petre A. şi col., 1989).
CAPITOLUL VIII
REZULTATE ŞI DISCUŢII
8.1. INCUBAŢIA ŞI ECLOZIUNEA VIRMILOR DE MĂTASE
8.1.1. Parametrii biologici ai pontelor şi ai ecloziunii la rasele şi hibrizii studiaţi
Pentru incubaţia ouălor raselor şi hibrizilor studiaţi s-a folosit metoda creşterii treptate a temperaturii, a cărei tehnică a fost descrisă de Mărghitaş, L. Al., (1995): de la temperatura de 150C menţinută timp de 3 zile, la 18 - 200C, timp de 2 zile, temperatura creşte apoi la 23-240C pentru 4 zile, iar la sfârşitul perioadei au fost menţinute 25-260C, pentru tot restul perioadei rămase până la ecloziune. Umiditatea relativă a aerului a variat între 75-85%, fiind mai ridicată în ultima parte a intervalului de incubaţie pentru a favoriza ecloziunea.
Un parametru deosebit de important îl reprezintă procentul de ecloziune al raselor, cel mai mic procent de ecloziune obţinându-se la rasa Alb chinezesc linia 29 (90.76%), valoare foarte apropiată de cea obţinută de Benţea (2006) (90.71%). Valoarea maximă pentru acest parametru s-a înregistrat la rasa Alb de Cislău (95.42%), valoare foarte apropiată de cea obţinută de alţi cercetători în domeniu (Paşca, 2004; Benţea, 2006). Valoarea medie a procentului de ecloziune a acestor rase a fost de 93,68%.
Numărul mediu de ouă/pontă la cei patru hibrizi simpli variază între 558 la hibridul AC29xB1 şi 694 la monohibridul ABxAC, ierarhie care se menţine şi pentru numărul de ouă eclozionate.
În ceea ce priveşte procentul de ecloziune, valoarea minimă a acestuia se înregistrează la hibridul ACxAB, acesta fiind de 94,78%, iar cel mai ridicat procent de 96,90% s-a înregistrat pentru hibrizii ABxAC şi B1xAC29. Media procentului de ecloziune pentru toţi cei patru monohobrizi studiaţi a fost de 96.36%. Din datele prezentate se observă o creştere cu 2.68% a mediei hibrizilor simplii faţă de rasele
14

REZUMAT
parentale, lucru explicat cu prisosinţă de efectul heterozisului, manifestat încă din stadiul de ou.
Numărul mediu de ouă în cazul trihibrizilor variază între 619 la trihibridul B1x (ACx AC29) şi 749 la (ABx B1)xAC29. Numărul minim de ouă eclozionate s-a obţinut la hibridul (ABx B1)x AC, adică cu un număr mediu de 616 ouă, pe când cel maxim a fost la trihibridul (ABx B1)x AC29, unde acesta a fost de 730 ouă din care au rezultat larve. Procentul de ecloziune a prezentat variaţii între 94.06 pentru trihibridul (ACx AC29)x B1 şi 97.67% pentru AC29x (ABx B1). Media ecloziunii la trihibrizi a fost de 96,61%. În acest caz diferenţa de 2.93% înregistrată la procentul de ecloziune al trihibrizilor faţă de cel al raselor parentale poate fi considerat semnificativ.
Se poate observa o variaţie în ceea ce priveşte numărul mediu de ouă/ pontă cuprinsă între 672 la tetrahibridul (ACx AC29)x (ABx B1) şi 785 la hibridul (ABxB1)x (ACx AC29), diferenţă datorată probabil capacităţii combinative diferite. Numărul de ouă eclozionate variază în acelaşi mod la cei doi tetrahibrizi studiaţi, iar procentul de ecloziune este de 98.22 la (AB x B1) x (AC x AC29) şi de 97.76 la (AC x AC29) x (AB x B1).
Cel mai ridicat procent de ecloziune s-a înregistrat la tetrahibrizi, trihibrizii şi hibrizii simpli având procente de ecloziune relativ egale (Figura 2).
93.68
96.36 96.61
98.22
91
92
93
94
95
96
97
98
99
Proc
entu
l de
eclo
ziun
e m
ediu
/Ave
rage
ha
tchi
ng p
erce
nt (%
)
RASE/RACES HIBRIZI SIMPLI/SIMPLE HYBRIDS
TRIHIBRIZI/TRIHYBRIDS TETRAHIBRIZI/TETRAHYBRIDS
Figura 2. Procentul de ecloziune al raselor parentale, hibrizilor simpli, trihibrizilor
şi tetrahibrizilor viermilor de mătase
Ecloziunea larvelor s-a desfăşurat pe durata a trei zile, majoritatea
(aproximativ 65%) eclozionând în cea de-a doua zi; doar aceste larve au fost selecţionate pentru realizarea seriilor experimentale în vederea uniformizării loturilor şi minimizării riscului ca larvele să nu intre în mod grupat în somn.
15
REZUMAT
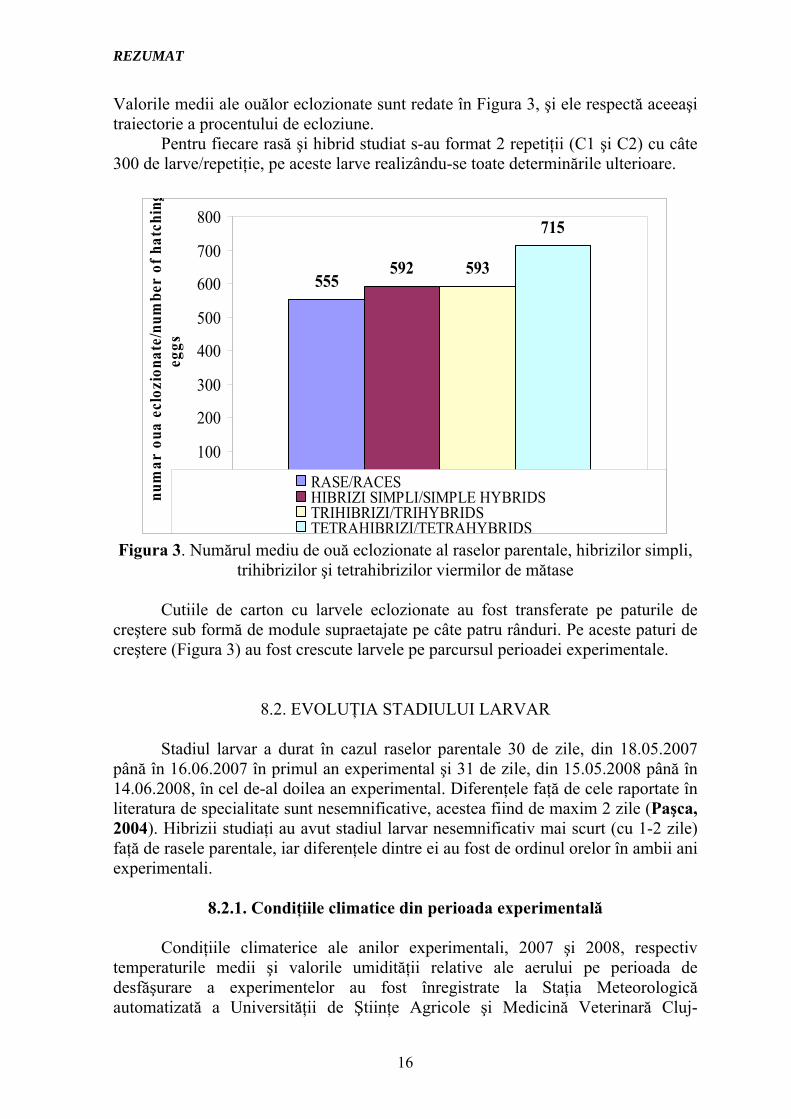
Valorile medii ale ouălor eclozionate sunt redate în Figura 3, şi ele respectă aceeaşi traiectorie a procentului de ecloziune.
Pentru fiecare rasă şi hibrid studiat s-au format 2 repetiţii (C1 şi C2) cu câte 300 de larve/repetiţie, pe aceste larve realizându-se toate determinările ulterioare.
555592 593
715
0
100
200
300
400
500
600
700
800
num
ar o
ua e
cloz
iona
te/n
umbe
r of
hat
chin
geg
gs
RASE/RACESHIBRIZI SIMPLI/SIMPLE HYBRIDSTRIHIBRIZI/TRIHYBRIDSTETRAHIBRIZI/TETRAHYBRIDS
Figura 3. Numărul mediu de ouă eclozionate al raselor parentale, hibrizilor simpli, trihibrizilor şi tetrahibrizilor viermilor de mătase
Cutiile de carton cu larvele eclozionate au fost transferate pe paturile de
creştere sub formă de module supraetajate pe câte patru rânduri. Pe aceste paturi de creştere (Figura 3) au fost crescute larvele pe parcursul perioadei experimentale.
8.2. EVOLUŢIA STADIULUI LARVAR
Stadiul larvar a durat în cazul raselor parentale 30 de zile, din 18.05.2007 până în 16.06.2007 în primul an experimental şi 31 de zile, din 15.05.2008 până în 14.06.2008, în cel de-al doilea an experimental. Diferenţele faţă de cele raportate în literatura de specialitate sunt nesemnificative, acestea fiind de maxim 2 zile (Paşca, 2004). Hibrizii studiaţi au avut stadiul larvar nesemnificativ mai scurt (cu 1-2 zile) faţă de rasele parentale, iar diferenţele dintre ei au fost de ordinul orelor în ambii ani experimentali.
8.2.1. Condiţiile climatice din perioada experimentală
Condiţiile climaterice ale anilor experimentali, 2007 şi 2008, respectiv temperaturile medii şi valorile umidităţii relative ale aerului pe perioada de desfăşurare a experimentelor au fost înregistrate la Staţia Meteorologică automatizată a Universităţii de Ştiinţe Agricole şi Medicină Veterinară Cluj-
16
REZUMAT
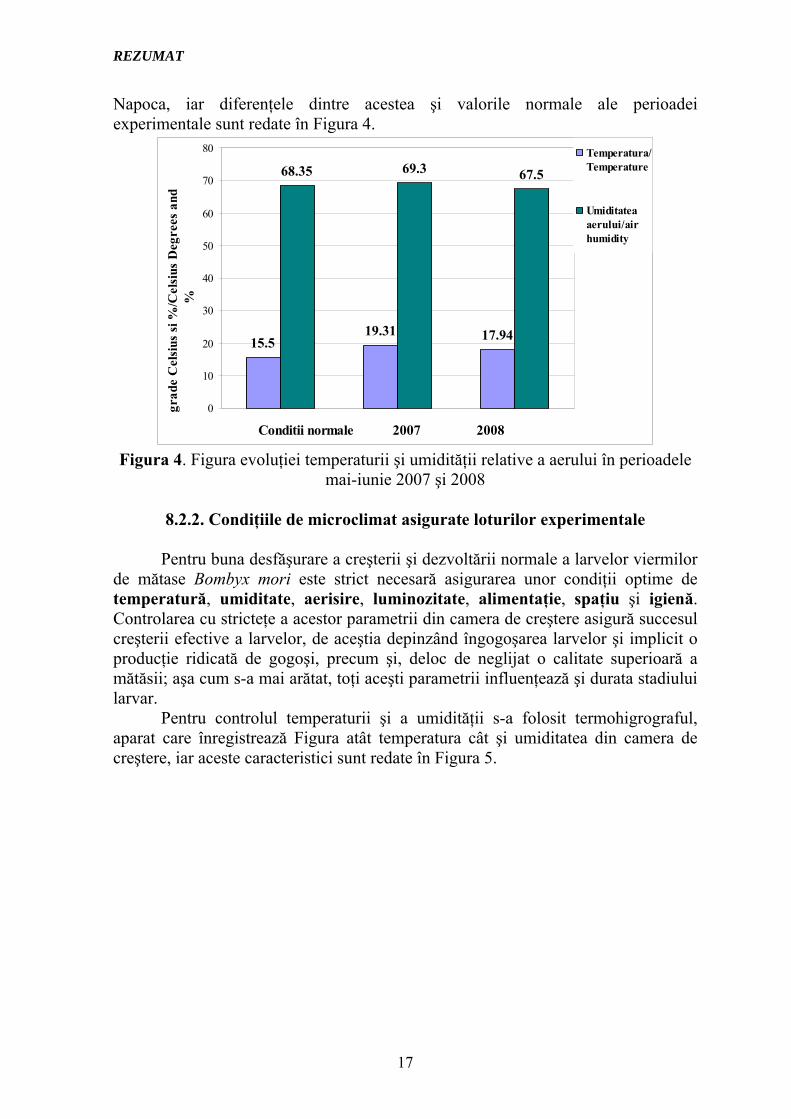
Napoca, iar diferenţele dintre acestea şi valorile normale ale perioadei experimentale sunt redate în Figura 4.
15.519.31 17.94
68.35 69.3 67.5
0
10
20
30
40
50
60
70
80
Conditii normale 2007 2008
grad
e C
elsi
us s
i %/C
elsi
us D
egre
es a
nd
%
Temperatura/Temperature
Umiditateaaerului/airhumidity
Figura 4. Figura evoluţiei temperaturii şi umidităţii relative a aerului în perioadele
mai-iunie 2007 şi 2008
8.2.2. Condiţiile de microclimat asigurate loturilor experimentale
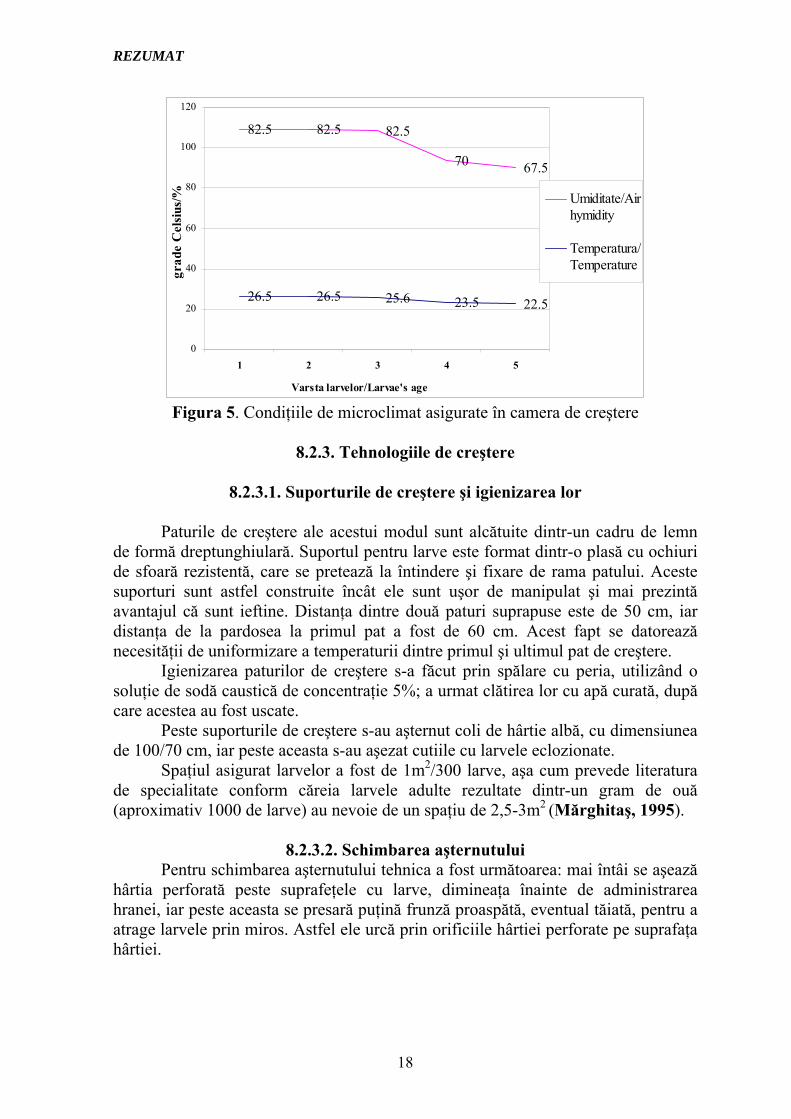
Pentru buna desfăşurare a creşterii şi dezvoltării normale a larvelor viermilor de mătase Bombyx mori este strict necesară asigurarea unor condiţii optime de temperatură, umiditate, aerisire, luminozitate, alimentaţie, spaţiu şi igienă. Controlarea cu stricteţe a acestor parametrii din camera de creştere asigură succesul creşterii efective a larvelor, de aceştia depinzând îngogoşarea larvelor şi implicit o producţie ridicată de gogoşi, precum şi, deloc de neglijat o calitate superioară a mătăsii; aşa cum s-a mai arătat, toţi aceşti parametrii influenţează şi durata stadiului larvar.
Pentru controlul temperaturii şi a umidităţii s-a folosit termohigrograful, aparat care înregistrează Figura atât temperatura cât şi umiditatea din camera de creştere, iar aceste caracteristici sunt redate în Figura 5.
17
REZUMAT
26.5 26.5 25.6 23.5 22.5
82.5 82.5 82.5
70 67.5
0
20
40
60
80
100
120
1 2 3 4 5
Varsta larvelor/Larvae's age
grad
e C
elsi
us/% Umiditate/Air
hymidity
Temperatura/Temperature
Figura 5. Condiţiile de microclimat asigurate în camera de creştere
8.2.3. Tehnologiile de creştere
8.2.3.1. Suporturile de creştere şi igienizarea lor
Paturile de creştere ale acestui modul sunt alcătuite dintr-un cadru de lemn
de formă dreptunghiulară. Suportul pentru larve este format dintr-o plasă cu ochiuri de sfoară rezistentă, care se pretează la întindere şi fixare de rama patului. Aceste suporturi sunt astfel construite încât ele sunt uşor de manipulat şi mai prezintă avantajul că sunt ieftine. Distanţa dintre două paturi suprapuse este de 50 cm, iar distanţa de la pardosea la primul pat a fost de 60 cm. Acest fapt se datorează necesităţii de uniformizare a temperaturii dintre primul şi ultimul pat de creştere.
Igienizarea paturilor de creştere s-a făcut prin spălare cu peria, utilizând o soluţie de sodă caustică de concentraţie 5%; a urmat clătirea lor cu apă curată, după care acestea au fost uscate.
Peste suporturile de creştere s-au aşternut coli de hârtie albă, cu dimensiunea de 100/70 cm, iar peste aceasta s-au aşezat cutiile cu larvele eclozionate.
Spaţiul asigurat larvelor a fost de 1m2/300 larve, aşa cum prevede literatura de specialitate conform căreia larvele adulte rezultate dintr-un gram de ouă (aproximativ 1000 de larve) au nevoie de un spaţiu de 2,5-3m2 (Mărghitaş, 1995).
8.2.3.2. Schimbarea aşternutului
Pentru schimbarea aşternutului tehnica a fost următoarea: mai întâi se aşează hârtia perforată peste suprafeţele cu larve, dimineaţa înainte de administrarea hranei, iar peste aceasta se presară puţină frunză proaspătă, eventual tăiată, pentru a atrage larvele prin miros. Astfel ele urcă prin orificiile hârtiei perforate pe suprafaţa hârtiei.
18

REZUMAT
8.2.4. Succesiunea vârstelor larvare delimitate de perioadele de somn
Prima vârstă larvară a durat 3-4 zile şi a fost urmată de o zi de somn; a doua
vârstă a durat 2-3 zile şi a fost şi ea urmată de o zi de somn; vârsta a 3-a larvară a durat 3-4 zile, încheindu-se cu o zi şi jumătate de somn. Vârsta a 4-a a durat 4-5 zile, iar la sfârşitul ei larvele au trecut timp de două zile printr-o perioadă de somn. Ultima vârstă (a-5-a) a durat 8-9 zile, iar la sfârşitul ei a avut loc îngogoşarea.
8.2.5. Hrana naturală – recoltare, pregătire şi administrare
Hrana utilizată în cadrul experimentelor pentru toate vârstele larvare, atât la rasele parentale cât şi la hibrizi a fost frunza de dud proaspăt recoltată de la soiul Ucraina 107 din plantaţia USAMV Cluj-Napoca, ea fiind administrată în acelaşi număr de tainuri la toate variantele. Pentru larvele aflate în primele trei vârste, frunzele s-au administrat tocate (Figura 2), iar apoi întregi sau chiar nedetaşate de pe ramuri. Recoltarea frunzei s-a realizat dimineaţa şi seara, în aceste momente ale zilei frunza având cel mai ridicat conţinut în principii nutritivi, putându-se asigura larvelor necesarul de proteine şi săruri minerale.
Figura 2. Administrarea frunzelor tocate (original)
Frunza administrată a fost curată, lipsită de agenţi poluanţi, iar transportul acesteia spre locaţia de creştere s-a realizat cu mijloace auto, în cel mai scurt timp.
8.3. INDICII BIOLOGICI AI LARVELOR
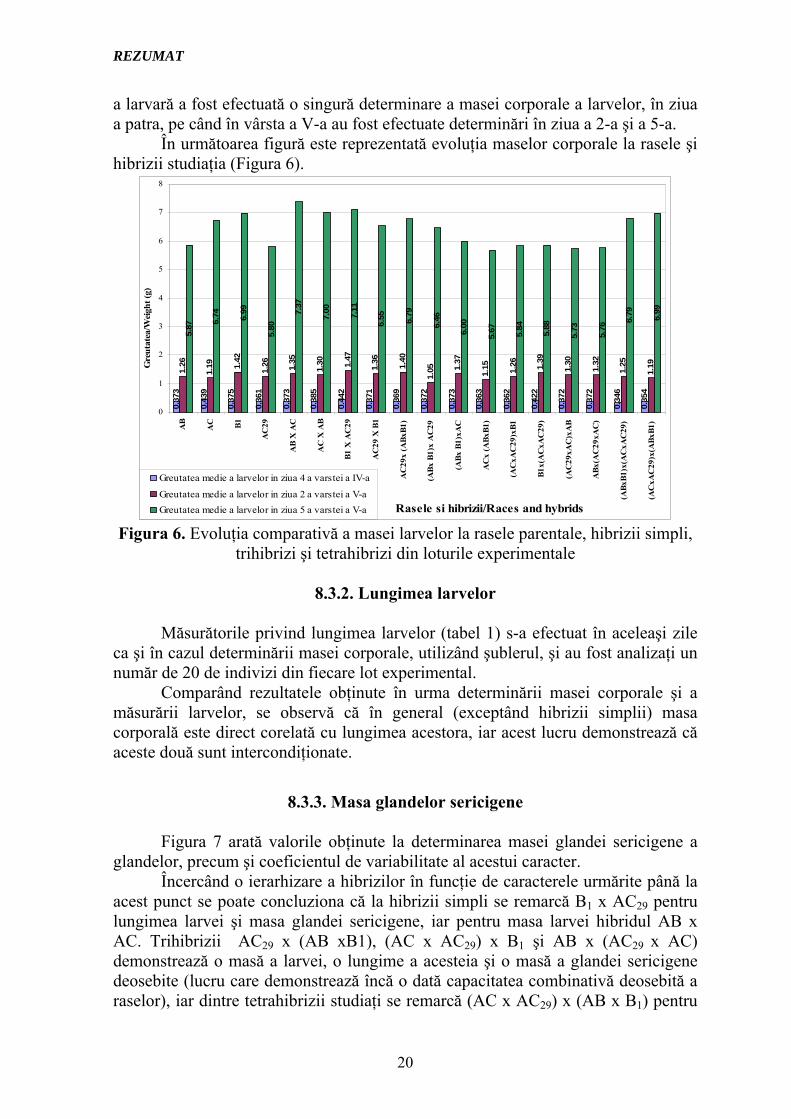
8.3.1. Masa larvelor Pentru determinarea acestor măsurători morfologice din fiecare lot
experimental au fost luaţi aleatoriu un număr de 20 de indivizi, iar determinarea masei larvelor s-a efectuat cu o balanţă electronică de mare precizie. În vârsta a IV-
19
REZUMAT
a larvară a fost efectuată o singură determinare a masei corporale a larvelor, în ziua a patra, pe când în vârsta a V-a au fost efectuate determinări în ziua a 2-a şi a 5-a.
În următoarea figură este reprezentată evoluţia maselor corporale la rasele şi hibrizii studiaţia (Figura 6).
0.37
3
0.43
9
0.37
5
0.36
1
0.37
3
0.38
5
0.44
2
0.37
1
0.36
9
0.37
2
0.37
3
0.36
3
0.36
2
0.42
2
0.37
2
0.37
2
0.34
6
0.35
4
1.26
1.19 1.
42
1.26 1.35
1.30 1.47
1.36 1.40
1.05 1.
37
1.15 1.26 1.39
1.30
1.32
1.25
1.19
5.87 6.
74 6.99
5.80
7.37
7.00 7.11
6.55 6.79
6.46
6.00
5.67 5.84
5.88
5.73
5.76
6.79 6.99
0
1
2
3
4
5
6
7
8A
B
AC B1
AC
29
AB
X A
C
AC
X A
B
B1 X
AC
29
AC
29 X
B1
AC
29x
(ABx
B1)
(ABx
B1)
x A
C29
(ABx
B1)
xAC
AC
x (A
BxB1
)
(AC
xAC
29)x
B1
B1x(
AC
xAC
29)
(AC
29xA
C)x
AB
ABx
(AC
29xA
C)
(ABx
B1)x
(AC
xAC
29)
(AC
xAC
29)x
(ABx
B1)
Rasele si hibrizii/Races and hybrids
Gre
utat
ea/W
eigh
t (g)
Greutatea medie a larvelor in ziua 4 a varstei a IV-a
Greutatea medie a larvelor in ziua 2 a varstei a V-aGreutatea medie a larvelor in ziua 5 a varstei a V-a
Figura 6. Evoluţia comparativă a masei larvelor la rasele parentale, hibrizii simpli, trihibrizi şi tetrahibrizi din loturile experimentale
8.3.2. Lungimea larvelor
Măsurătorile privind lungimea larvelor (tabel 1) s-a efectuat în aceleaşi zile
ca şi în cazul determinării masei corporale, utilizând şublerul, şi au fost analizaţi un număr de 20 de indivizi din fiecare lot experimental.
Comparând rezultatele obţinute în urma determinării masei corporale şi a măsurării larvelor, se observă că în general (exceptând hibrizii simplii) masa corporală este direct corelată cu lungimea acestora, iar acest lucru demonstrează că aceste două sunt intercondiţionate.
8.3.3. Masa glandelor sericigene
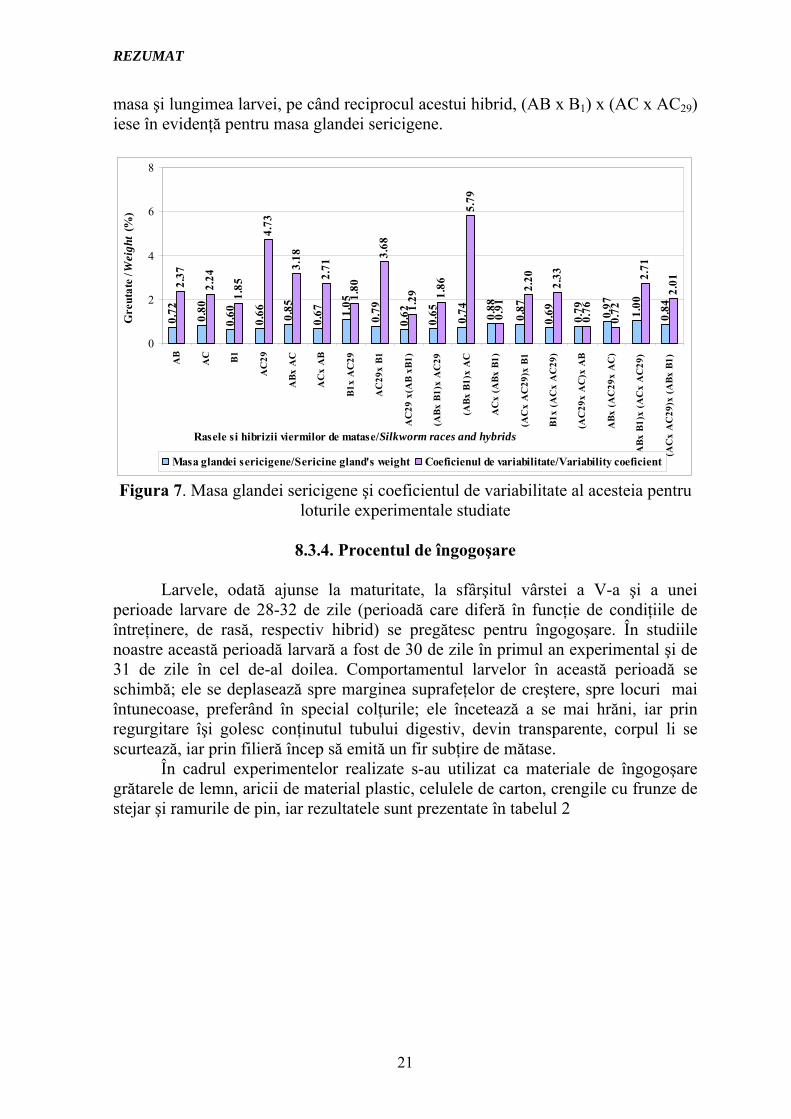
Figura 7 arată valorile obţinute la determinarea masei glandei sericigene a glandelor, precum şi coeficientul de variabilitate al acestui caracter.
Încercând o ierarhizare a hibrizilor în funcţie de caracterele urmărite până la acest punct se poate concluziona că la hibrizii simpli se remarcă B1 x AC29 pentru lungimea larvei şi masa glandei sericigene, iar pentru masa larvei hibridul AB x AC. Trihibrizii AC29 x (AB xB1), (AC x AC29) x B1 şi AB x (AC29 x AC) demonstrează o masă a larvei, o lungime a acesteia şi o masă a glandei sericigene deosebite (lucru care demonstrează încă o dată capacitatea combinativă deosebită a raselor), iar dintre tetrahibrizii studiaţi se remarcă (AC x AC29) x (AB x B1) pentru
20
REZUMAT
masa şi lungimea larvei, pe când reciprocul acestui hibrid, (AB x B1) x (AC x AC29) iese în evidenţă pentru masa glandei sericigene.
0.66 0.85
0.67 0.79
0.62
0.65 0.74 0.87
0.69 0.79 1.00
2.24
4.73
3.18
2.71
3.68
5.79
2.33 2.
71
2.01
0.80 1.05
0.72
0.60 0.840.97
0.88
2.20
1.86
1.29
2.37
1.85
1.80
0.72
0.760.91
0
2
4
6
8A
B
AC B1
AC
29
ABx
AC
AC
x A
B
B1x
AC
29
AC
29x
B1
AC
29 x
(AB
xB1)
(ABx
B1)
x A
C29
(ABx
B1)
x A
C
AC
x (A
Bx B
1)
(AC
x A
C29
)x B
1
B1x
(AC
x A
C29
)
(AC
29x
AC
)x A
B
ABx
(AC
29x
AC
)
(ABx
B1)
x (A
Cx
AC
29)
(AC
x A
C29
)x (A
Bx B
1)
Rasele si hibrizii viermilor de matase/Silkworm races and hybrids
Gre
utat
e / W
eigh
t (%
)
Masa glandei sericigene/Sericine gland's weight Coeficienul de variabilitate/Variability coeficient
Figura 7. Masa glandei sericigene şi coeficientul de variabilitate al acesteia pentru loturile experimentale studiate
8.3.4. Procentul de îngogoşare
Larvele, odată ajunse la maturitate, la sfârşitul vârstei a V-a şi a unei
perioade larvare de 28-32 de zile (perioadă care diferă în funcţie de condiţiile de întreţinere, de rasă, respectiv hibrid) se pregătesc pentru îngogoşare. În studiile noastre această perioadă larvară a fost de 30 de zile în primul an experimental şi de 31 de zile în cel de-al doilea. Comportamentul larvelor în această perioadă se schimbă; ele se deplasează spre marginea suprafeţelor de creştere, spre locuri mai întunecoase, preferând în special colţurile; ele încetează a se mai hrăni, iar prin regurgitare îşi golesc conţinutul tubului digestiv, devin transparente, corpul li se scurtează, iar prin filieră încep să emită un fir subţire de mătase.
În cadrul experimentelor realizate s-au utilizat ca materiale de îngogoşare grătarele de lemn, aricii de material plastic, celulele de carton, crengile cu frunze de stejar şi ramurile de pin, iar rezultatele sunt prezentate în tabelul 2
21
REZUMAT
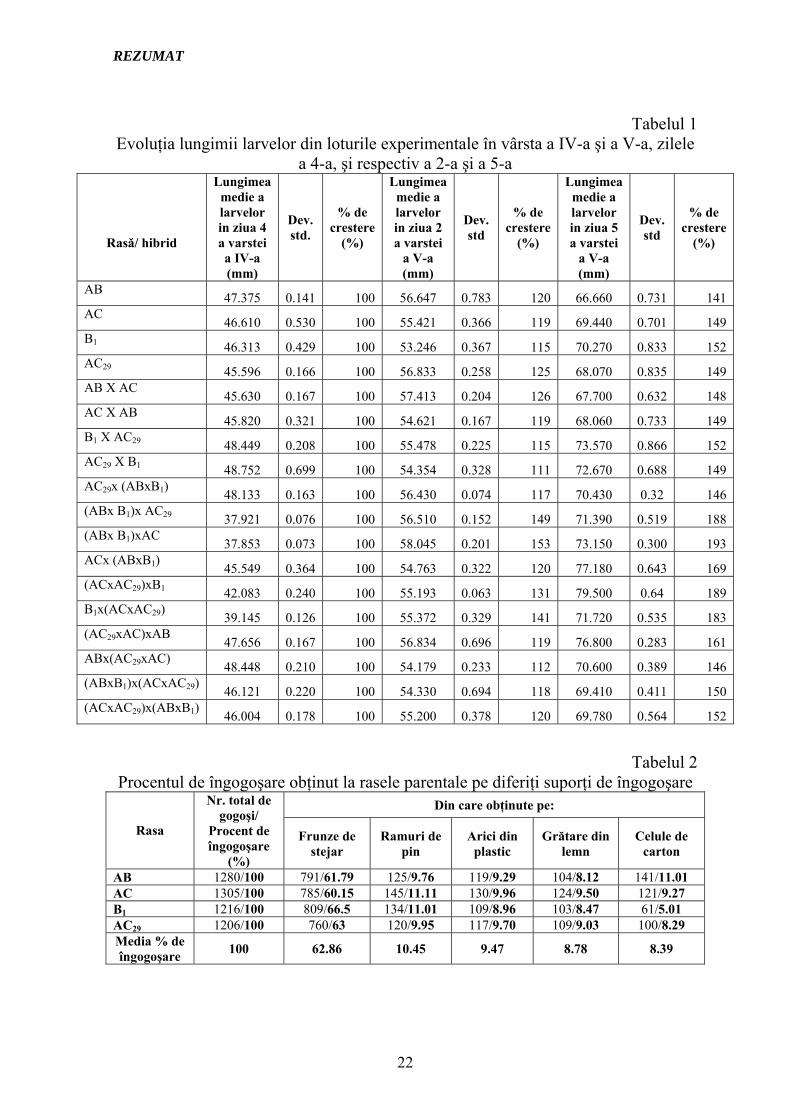
Tabelul 1
Evoluţia lungimii larvelor din loturile experimentale în vârsta a IV-a şi a V-a, zilele a 4-a, şi respectiv a 2-a şi a 5-a
Rasǎ/ hibrid
Lungimea medie a larvelor in ziua 4 a varstei a IV-a (mm)
Dev. std.
% de crestere
(%)
Lungimea medie a larvelor in ziua 2 a varstei
a V-a (mm)
Dev. std
% de crestere
(%)
Lungimea medie a larvelor in ziua 5 a varstei
a V-a (mm)
Dev. std
% de crestere
(%)
AB 47.375 0.141 100 56.647 0.783 120 66.660 0.731 141 AC 46.610 0.530 100 55.421 0.366 119 69.440 0.701 149 B1 46.313 0.429 100 53.246 0.367 115 70.270 0.833 152 AC29 45.596 0.166 100 56.833 0.258 125 68.070 0.835 149 AB X AC 45.630 0.167 100 57.413 0.204 126 67.700 0.632 148 AC X AB 45.820 0.321 100 54.621 0.167 119 68.060 0.733 149 B1 X AC29 48.449 0.208 100 55.478 0.225 115 73.570 0.866 152 AC29 X B1 48.752 0.699 100 54.354 0.328 111 72.670 0.688 149 AC29x (ABxB1) 48.133 0.163 100 56.430 0.074 117 70.430 0.32 146 (ABx B1)x AC29 37.921 0.076 100 56.510 0.152 149 71.390 0.519 188 (ABx B1)xAC 37.853 0.073 100 58.045 0.201 153 73.150 0.300 193 ACx (ABxB1) 45.549 0.364 100 54.763 0.322 120 77.180 0.643 169 (ACxAC29)xB1 42.083 0.240 100 55.193 0.063 131 79.500 0.64 189 B1x(ACxAC29) 39.145 0.126 100 55.372 0.329 141 71.720 0.535 183 (AC29xAC)xAB 47.656 0.167 100 56.834 0.696 119 76.800 0.283 161 ABx(AC29xAC) 48.448 0.210 100 54.179 0.233 112 70.600 0.389 146 (ABxB1)x(ACxAC29) 46.121 0.220 100 54.330 0.694 118 69.410 0.411 150 (ACxAC29)x(ABxB1) 46.004 0.178 100 55.200 0.378 120 69.780 0.564 152
Tabelul 2 Procentul de îngogoşare obţinut la rasele parentale pe diferiţi suporţi de îngogoşare
Din care obţinute pe:
Rasa
Nr. total de gogoşi/
Procent de îngogoşare
(%)
Frunze de stejar
Ramuri de pin
Arici din plastic
Grătare din lemn
Celule de carton
AB 1280/100 791/61.79 125/9.76 119/9.29 104/8.12 141/11.01 AC 1305/100 785/60.15 145/11.11 130/9.96 124/9.50 121/9.27 B1 1216/100 809/66.5 134/11.01 109/8.96 103/8.47 61/5.01 AC29 1206/100 760/63 120/9.95 117/9.70 109/9.03 100/8.29 Media % de îngogoşare 100 62.86 10.45 9.47 8.78 8.39
22

REZUMAT
8.4. INDICII BIOLOGICI AI GOGOŞILOR CRUDE
8.4.1. Masa gogoşilor crude
Masa gogoşii crude a fost apreciată prin cântărirea individuală a unui număr de 200 de gogoşi (100 au fost femele şi 100 mascule), pentru fiecare rasă parentală şi hibrid studiat, şi ele au rezultat în urma sexării crisalidelor, obţinute din cele două repetiţii ale fiecărui lot experimental.
Rezultatele obţinute pentru această caracteristică sunt prezentate în tabelul 3.
Tabelul 3 Masa gogoşilor crude mascule şi femele
Rasa/ Hibridul
Media raselor/ hibrizilor
Masa gogosii mascule crude
(g)
Masa gogosii crude femele (g)
Media gogosilor de ambele sexe
AB 1.404 1.777 1.591 AC 1.349 1.801 1.575 B1 1.439 1.752 1.596 AC29
1.630
1.531 1.989 1.760 AB x AC 1.502 1.859 1.681 AC x AB 1.491 1.821 1.656 B1 x AC29 1.570 1.789 1.680 AC29 x B1
1.668
1.431 1.879 1.655
AC29 x (ABxB1) 1.701 1.986 1.844
(ABxB1) x AC29 1.299 1.673 1.486 (ABxB1) x AC 1.469 1.856 1.663 AC x (ABxB1) 1.693 1.982 1.838 (ACxAC29) x B1 1.899 2.400 2.150 B1 x (ACxAC29) 1.383 1.678 1.531 (AC29xAC) x AB 1.415 1.802 1.609 AB x (AC29xAC)
1.714
1.419 1.766 1.593 (ABxB1) x (ACxAC29) 1.382 1.704 1.543 (ACxAC29) x (ABxB1)
1.496 1.220 1.677 1.449
8.4.2. Masa incartamentului mătăsos
Pentru determinarea masa incartamentului mătăsos, gogoaşa a fost
secţionată, apoi s-a îndepărtat crisalida şi tegumentul de năpârlire din gogoaşă. Cântărirea incartamentului mătăsos s-a realizat cu ajutorul unei balanţe electronice de mare precizie. Determinarea masei incartamentului s-a realizat diferenţiat pe sexe, aşa cum s-a procedat şi în cazul masei gogoşii crude, numărul gogoşilor studiate pentru acest caracter fiind de 15 gogoşi pentru fiecare sex.
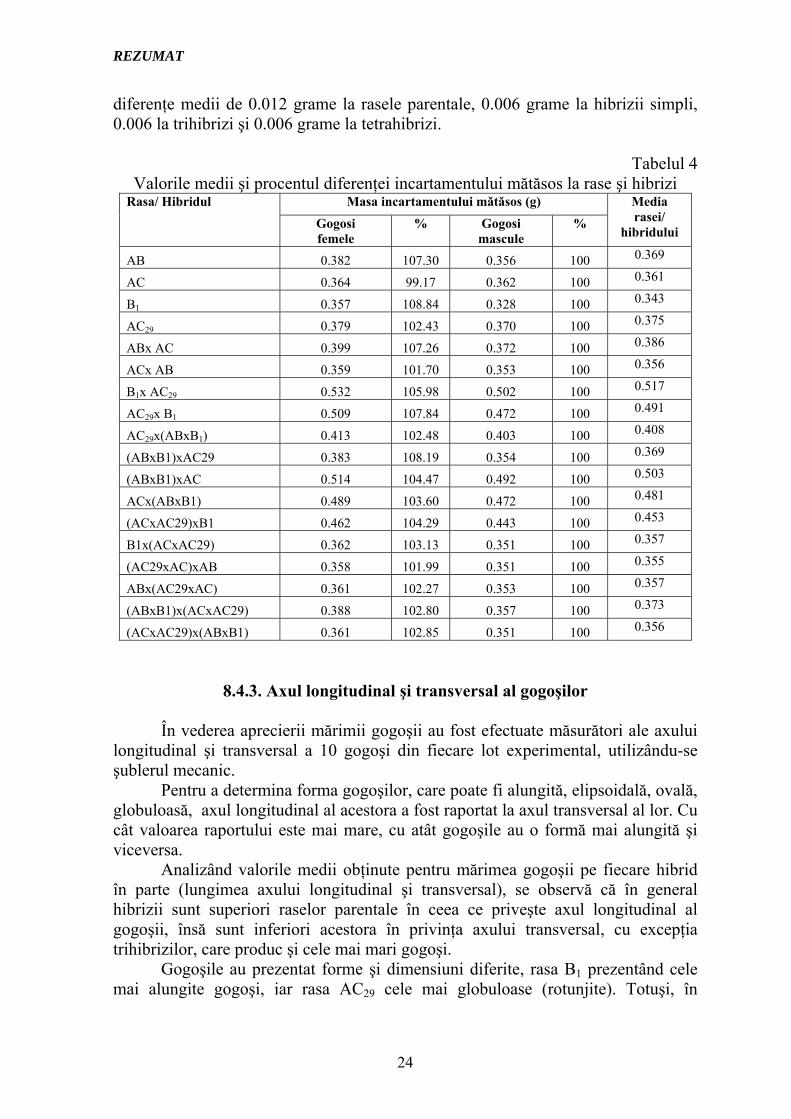
În tabelul 4 sunt prezentate valorile medii şi procentul diferenţei incartamentului mătăsos la rasele şi hibrizii studiaţi, diferenţiat pe sexe. Şi în acest caz se verifică superioritatea gogoşilor femele faţă de cele mascule, obţinându-se
23
REZUMAT
diferenţe medii de 0.012 grame la rasele parentale, 0.006 grame la hibrizii simpli, 0.006 la trihibrizi şi 0.006 grame la tetrahibrizi.
Tabelul 4
Valorile medii şi procentul diferenţei incartamentului mătăsos la rase şi hibrizi Masa incartamentului mǎtǎsos (g) Rasa/ Hibridul
Gogosi femele
% Gogosi mascule
%
Media rasei/
hibridului
AB 0.382 107.30 0.356 100 0.369
AC 0.364 99.17 0.362 100 0.361
B1 0.357 108.84 0.328 100 0.343
AC29 0.379 102.43 0.370 100 0.375
ABx AC 0.399 107.26 0.372 100 0.386
ACx AB 0.359 101.70 0.353 100 0.356
B1x AC29 0.532 105.98 0.502 100 0.517
AC29x B1 0.509 107.84 0.472 100 0.491
AC29x(ABxB1) 0.413 102.48 0.403 100 0.408
(ABxB1)xAC29 0.383 108.19 0.354 100 0.369
(ABxB1)xAC 0.514 104.47 0.492 100 0.503
ACx(ABxB1) 0.489 103.60 0.472 100 0.481
(ACxAC29)xB1 0.462 104.29 0.443 100 0.453
B1x(ACxAC29) 0.362 103.13 0.351 100 0.357
(AC29xAC)xAB 0.358 101.99 0.351 100 0.355
ABx(AC29xAC) 0.361 102.27 0.353 100 0.357
(ABxB1)x(ACxAC29) 0.388 102.80 0.357 100 0.373
(ACxAC29)x(ABxB1) 0.361 102.85 0.351 100 0.356
8.4.3. Axul longitudinal şi transversal al gogoşilor
În vederea aprecierii mărimii gogoşii au fost efectuate măsurători ale axului longitudinal şi transversal a 10 gogoşi din fiecare lot experimental, utilizându-se şublerul mecanic.
Pentru a determina forma gogoşilor, care poate fi alungită, elipsoidală, ovală, globuloasă, axul longitudinal al acestora a fost raportat la axul transversal al lor. Cu cât valoarea raportului este mai mare, cu atât gogoşile au o formă mai alungită şi viceversa.
Analizând valorile medii obţinute pentru mărimea gogoşii pe fiecare hibrid în parte (lungimea axului longitudinal şi transversal), se observă că în general hibrizii sunt superiori raselor parentale în ceea ce priveşte axul longitudinal al gogoşii, însă sunt inferiori acestora în privinţa axului transversal, cu excepţia trihibrizilor, care produc şi cele mai mari gogoşi.
Gogoşile au prezentat forme şi dimensiuni diferite, rasa B1 prezentând cele mai alungite gogoşi, iar rasa AC29 cele mai globuloase (rotunjite). Totuşi, în
24

REZUMAT
general, gogoşile produse de toate categoriile de hibrizi sunt uniforme în privinţa formei.
8.4.4. Procentul de mătase din gogoaşă
Însuşirile gogoşilor crude studiate anterior (masa gogoşii crude şi masa incartamentului mǎtǎsos), au demonstrat superioritatea gogoşilor femele faţă de cele mascule; au fost analizate un număr de 15 gogoşi din fiecare lot experimental. În cazul procentului de mătase din gogoaşă se verifică în schimb superioritatea gogoşilor mascule faţă de cele femele, aşa cum reiese din tabelul 5.
Tabelul 5 Conţinutul gogoşilor mascule şi femele în mǎtase crudǎ (%)
Procentul de mǎtase din gogoaşǎ(%)
Rasa/ Hibridul Gogosi femele
(%)
Dev. std. %
Gogosi mascule
(%)
Dev. std %
Media rasei/
hibridului (%)
Dev. std.
AB 21.654 0.299 100 24.677 0.341 113.96 23.166 0.320
AC 19.941 0.276 100 25.401 0.351 127.38 22.671 0.314
B1 20.241 0.280 100 23.171 0.320 114.48 21.706 0.300
AC29 19.305 0.267 100 24.123 0.333 124.96 21.714 0.300
Media raselor 22.289 0.311 100 26.257 0.362 117.80 24.273 0.337 ABx AC 20.873 0.288 100 24.353 0.336 116.67 22.613 0.312
ACx AB 19.710 0.272 100 23.664 0.327 120.06 21.687 0.300
B1x AC29 20.488 0.283 100 24.506 0.339 119.61 22.497 0.311
AC29x B1 20.578 0.284 100 23.9 0.330 116.14 22.239 0.307 Media hibrizilor simpli 20.412 0.282 100 24.106 0.333 118.09 22.259 0.307
AC29x(ABxB1) 20.755 0.287 100 24.075 0.333 116.00 22.415 0.310
(ABxB1)xAC29 21.847 0.302 100 27.611 0.381 126.38 24.729 0.342
(ABxB1)xAC 22.524 0.311 100 26.259 0.363 116.58 24.392 0.337
ACx(ABxB1) 20.583 0.284 100 25.487 0.352 123.83 23.035 0.318
(ACxAC29)xB1 19.287 0.266 100 24.034 0.332 124.61 21.661 0.299
B1x(ACxAC29) 21.235 0.293 100 25.424 0.351 119.73 23.330 0.322
(AC29xAC)xAB 20.337 0.281 100 24.715 0.341 121.53 22.526 0.311
ABx(AC29xAC) 20.284 0.280 100 24.325 0.336 119.92 22.305 0.308
Media trihibrizilor 20.857 0.288 100 25.241 0.349 121.02 23.049 0.318 (ABxB1)x (ACxAC29) 19.736 0.273 100 26.138 0.361 132.44 22.937 0.317
(ACxAC29)x (ABxB1) 21.644 0.299 100 28.97 0.400 133.85 25.307 0.350 Media tetrahibrizilor 20.690 0.282 100 27.554 0.381 133.18 24.122 0.333
8.5. INDICII TEHNOLOGICI AI GOGOŞII USCATE ŞI AI FIBREI
În urma determinǎrilor realizate pe gogoşile crude la 10 zile de la îngogoşarea în masă, acestea au fost „etufate” (crisalidele din interior au fost omorâte). Metoda de etufare aleasǎ a fost cea cu aer cald utilizându-se etuva
25
REZUMAT
încălzită la temperatura de etufare de circa 800C, timp de mai multe ore, aceastǎ metodǎ nedegradând prea mult firul de mǎtase.
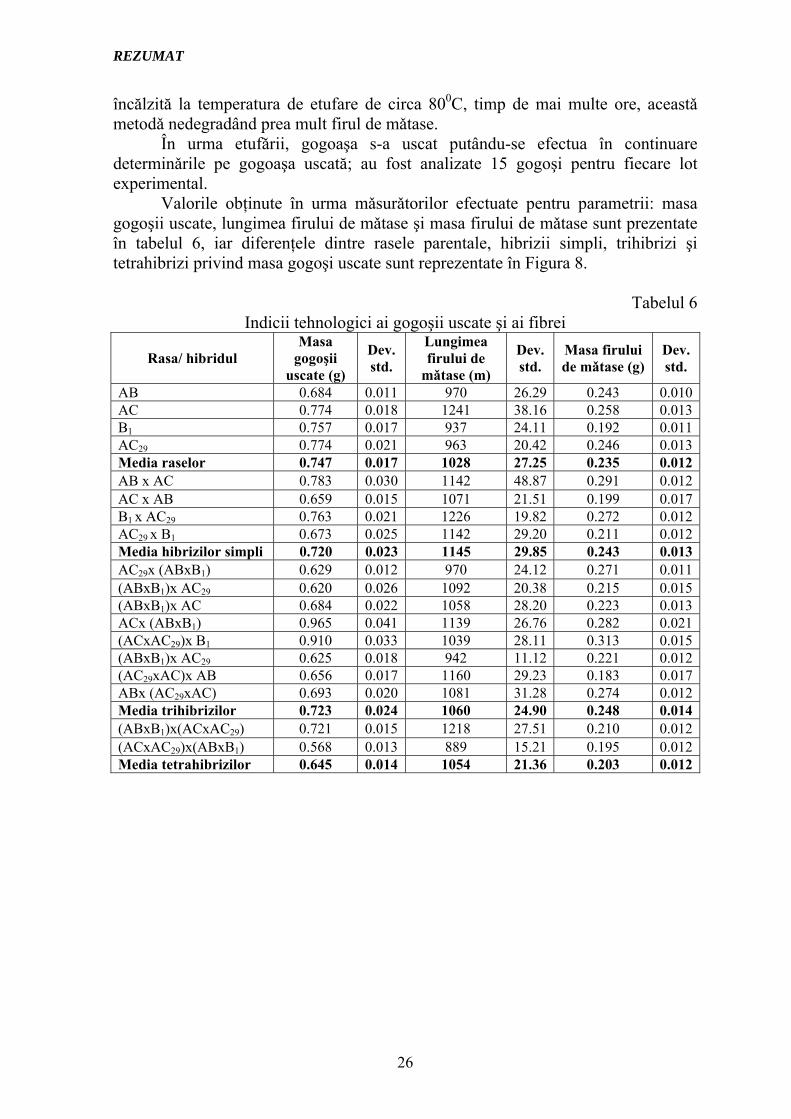
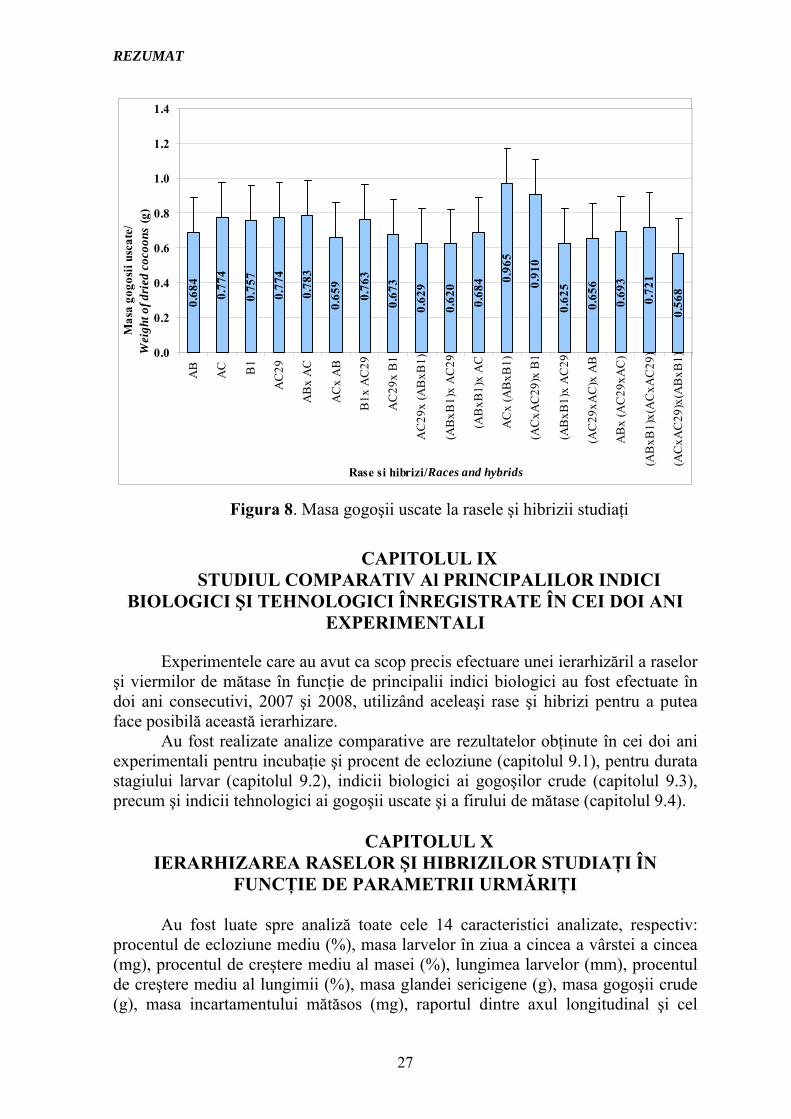
În urma etufǎrii, gogoaşa s-a uscat putându-se efectua în continuare determinǎrile pe gogoaşa uscată; au fost analizate 15 gogoşi pentru fiecare lot experimental. Valorile obţinute în urma mǎsurǎtorilor efectuate pentru parametrii: masa gogoşii uscate, lungimea firului de mǎtase şi masa firului de mǎtase sunt prezentate în tabelul 6, iar diferenţele dintre rasele parentale, hibrizii simpli, trihibrizi şi tetrahibrizi privind masa gogoşi uscate sunt reprezentate în Figura 8.
Tabelul 6
Indicii tehnologici ai gogoşii uscate şi ai fibrei
Rasa/ hibridul Masa
gogoşii uscate (g)
Dev. std.
Lungimea firului de
mǎtase (m)
Dev. std.
Masa firului de mǎtase (g)
Dev. std.
AB 0.684 0.011 970 26.29 0.243 0.010AC 0.774 0.018 1241 38.16 0.258 0.013B1 0.757 0.017 937 24.11 0.192 0.011AC29 0.774 0.021 963 20.42 0.246 0.013Media raselor 0.747 0.017 1028 27.25 0.235 0.012AB x AC 0.783 0.030 1142 48.87 0.291 0.012AC x AB 0.659 0.015 1071 21.51 0.199 0.017B1 x AC29 0.763 0.021 1226 19.82 0.272 0.012AC29 x B1 0.673 0.025 1142 29.20 0.211 0.012Media hibrizilor simpli 0.720 0.023 1145 29.85 0.243 0.013AC29x (ABxB1) 0.629 0.012 970 24.12 0.271 0.011(ABxB1)x AC29 0.620 0.026 1092 20.38 0.215 0.015(ABxB1)x AC 0.684 0.022 1058 28.20 0.223 0.013ACx (ABxB1) 0.965 0.041 1139 26.76 0.282 0.021(ACxAC29)x B1 0.910 0.033 1039 28.11 0.313 0.015(ABxB1)x AC29 0.625 0.018 942 11.12 0.221 0.012(AC29xAC)x AB 0.656 0.017 1160 29.23 0.183 0.017ABx (AC29xAC) 0.693 0.020 1081 31.28 0.274 0.012Media trihibrizilor 0.723 0.024 1060 24.90 0.248 0.014(ABxB1)x(ACxAC29) 0.721 0.015 1218 27.51 0.210 0.012(ACxAC29)x(ABxB1) 0.568 0.013 889 15.21 0.195 0.012Media tetrahibrizilor 0.645 0.014 1054 21.36 0.203 0.012
26
REZUMAT
0.68
4
0.77
4
0.75
7
0.77
4
0.78
3
0.65
9
0.76
3
0.67
3
0.62
9
0.62
0
0.68
4 0.96
5
0.91
0
0.62
5
0.65
6
0.69
3
0.72
1
0.56
8
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
AB
AC B1
AC
29
AB
x A
C
AC
x A
B
B1x
AC
29
AC
29x
B1
AC
29x
(AB
xB1)
(AB
xB1)
x A
C29
(AB
xB1)
x A
C
AC
x (A
BxB
1)
(AC
xAC
29)x
B1
(AB
xB1)
x A
C29
(AC
29xA
C)x
AB
AB
x (A
C29
xAC
)
(AB
xB1)
x(A
CxA
C29
)
(AC
xAC
29)x
(AB
xB1)
Rase si hibrizi/Races and hybrids
Mas
a go
gosi
i usc
ate/
W
eigh
t of d
ried
coc
oons
(g)
Figura 8. Masa gogoşii uscate la rasele şi hibrizii studiaţi
CAPITOLUL IX
STUDIUL COMPARATIV Al PRINCIPALILOR INDICI BIOLOGICI ŞI TEHNOLOGICI ÎNREGISTRATE ÎN CEI DOI ANI
EXPERIMENTALI
Experimentele care au avut ca scop precis efectuare unei ierarhizăril a raselor şi viermilor de mătase în funcţie de principalii indici biologici au fost efectuate în doi ani consecutivi, 2007 şi 2008, utilizând aceleaşi rase şi hibrizi pentru a putea face posibilă această ierarhizare.
Au fost realizate analize comparative are rezultatelor obţinute în cei doi ani experimentali pentru incubaţie şi procent de ecloziune (capitolul 9.1), pentru durata stagiului larvar (capitolul 9.2), indicii biologici ai gogoşilor crude (capitolul 9.3), precum şi indicii tehnologici ai gogoşii uscate şi a firului de mătase (capitolul 9.4).
CAPITOLUL X
IERARHIZAREA RASELOR ŞI HIBRIZILOR STUDIAŢI ÎN FUNCŢIE DE PARAMETRII URMĂRIŢI
Au fost luate spre analiză toate cele 14 caracteristici analizate, respectiv:
procentul de ecloziune mediu (%), masa larvelor în ziua a cincea a vârstei a cincea (mg), procentul de creştere mediu al masei (%), lungimea larvelor (mm), procentul de creştere mediu al lungimii (%), masa glandei sericigene (g), masa gogoşii crude (g), masa incartamentului mătăsos (mg), raportul dintre axul longitudinal şi cel
27

REZUMAT
transversal, procentul de mătase din gogoaşă, masa gogoşii uscate (g), lungimea firului de mătase (m) şi masa firului de mătase (g).
Analizând rasele parentale se observă că cea mai valoroasă rasă parentală este AC, care a înregistrat valori maxime pentru 6 din caracteristicile analizate urmată de rasele parentale B1 şi AC29, care au înregistrat fiecare valori maxime pentru câte 4 din caracteristicile urmărite.
Dintre hibrizii simpli se remarcă AB x AC, la care 8 caracteristici au prezentat valorile maxime; acesta este urmat de hibridul B1 X AC29, cu 6 valori maxime.
Trihibrizii studiaţi sunt mult mai neuniformi, privind procentul de ecloziune şi indicii biologici ale larvelor, ieşind în evidenţă (AC x AC29) x B1 şi (ABx B1) x AC, cu câte 3 valori maxime înregistrate la caracteristicile urmărite.
Tetrahibridul cele mai valoros după ierarhizare este (ABxB1)x(ACxAC29).
28

REZUMAT
CONCLUZII
1. În cadrul prezentei lucrări sunt expuse date cu privire la incubaţia şi ecloziunea ouălor viermilor de mătase şi date cu privire la principalii indici biologici ai larvelor. Studiul efectuat a permis o ierarhizare a raselor parentale studiate şi a diferitelor tipuri de hibrizi, luând în considerare caracterele biologice urmărite, acest lucru corespunzând întru totul cu scopul şi obiectivele cercetărilor. 2. Condiţiile de microclimat din sala de creştere, calitatea frunzelor administrate, igienizarea spaţiului destinat creşterii şi tehnologia de creştere aplicată, reprezintă factori deosebit de importanţi în buna desfăşurare a experimentelor, motiv pentru care au fost asigurate toate aceste condiţii în parametrii optimi pentru specia Bombyx mori. 3. Procentul de ecloziune (%) cel mai ridicat s-a observat în cazul tetrahibrizilor (98.22%), un procent de doar 93.68% fiind înregistrat de rasele parentale. Implicit această tendinţă se menţine şi pentru numărul de ouă eclozionate. 4. Uniformitatea materialului biologic este demonstrată de durata stadiului larvar; acesta a durat la rasele parentale 30 de zile în anul 2007 şi 31 de zile în anul 2008, iar diferenţele obţinute în cercetările noastre faţă de cele obţinute de alţi cercetători s-au dovedit a fi nesemnificative. Un alt aspect important în demonstrarea uniformităţii materialului biologic este faptul că hibrizii studiaţi au avut stadiul larvar nesemnificativ mai scurt (cu 1-2 zile) faţă de rasele parentale, iar diferenţele dintre ei au fost de ordinul orelor în ambii ani experimentali. 5. Masa larvelor (g) a prezentat valori medii care se încadrau în valorile obţinute de alţi cercetători, evidenţiindu-se rasa B1, hibridul simplu AB x AC, trihibridul (AB x B1) x AC şi tetrahibridul (AB x B1) x (AC x AC29). 6. În ceea ce priveşte lungimea larvelor (mm) se evidenţiază rasa AC, hibridul B1 x AC29, trihibridul (AC x AC29) x B1 şi tetrahibridul (AC x AC29) x (AB x B1). 7. Masa glandelor sericigene (g) a evidenţiat ca superioară rasa AC, hibridul simplu B1 x AC29, trihibridul (AB x B1) x AC şi tetrahibridul (AB x B1) x (AC x AC29). 8. Studiu masei gogoşilor crude (g) a arătat superioritatea rasei parentale AC29, a hibridului simplu AB x AC, a trihibridului (AC x AC29) x B1, şi a tetrahibridului (AB x B1) x (AC x AC29). 9. Rasa AC29, hibridul simplu B1 x AC29, trihibridul (AB x B1) x AC, şi reciprocul acestuia AC x (AB x B1), precum şi tetrahibridul (AB x B1) x (AC x AC29) au prezentat cele mai mari valori în ceea ce priveşte masa incartamentului mătăsos. 10. Analiza valorilor medii obţinute pentru mărimea gogoşii pentru fiecare hibrid în parte (lungimea axului longitudinal şi transversal), arată că în general hibrizii sunt superiori raselor parentale în ceea ce priveşte axul longitudinal al gogoşii, însă sunt inferiori acestora în privinţa axului transversal, cu excepţia trihibrizilor, care produc şi cele mai mari gogoşi. 11. Cel mai ridicat procent mediu de mătase este întâlnit la rasele parentale (26.257%), urmat de tetrahibrizi (27.554%) şi trihibrizi, care prezintă valori foarte
29

REZUMAT
apropiate (25.241%), pe ultimul loc situându-se hibrizii simpli cu un procent mediu de 24.106%. 12. Între indicii biologici ai gogoşilor crude (masa gogoşilor crude, masa incartamentului mătăsos, raportul dintre axul longitudinal şi cel transversal al gogoşii, procentul de mătase din gogoaşă) nu există corelaţii pozitive. 13. Indicii tehnologici ai gogoşii uscate şi ai fibrei studiaţi au fost masa gogoşii uscate, lungimea firului de mătase, masa firului de mătase şi greutatea deşeului din gogoaşă. 14. Masa gogoşii uscate este superioară la rasele parentale (0.747 grame), şi este urmată de hibrizii simpli (0.720 grame), care sunt foarte apropiaţi de media trihibrizilor (0.723 grame). Tetrahibrizii se situează pe ultimul loc cu o valoare medie de 0.645 grame. 15. Rasele parentale studiate prezintă o medie a lungimii firului de mătase de 1028 m, fiind inferioare hibrizilor simpli, care prezintă o medie de 1145 m, trihibrizilor (1060 m) şi tetrahibrizilor (1054 m); aceste valori concordă cu cele declarate de alţi cercetători români şi din străinătate. 16. Pentru masa firului de mătase se evidenţiază rasa AC (0.258 g), hibridul simplu (0.291 g), trihibridul (AC x AC29) x B1, şi tetrahibridul (AB x B1) x (AC x AC29). Per ansamblu, cea mai ridicată medie a masei firului de mătase se înregistrează la trihibrizi (0.248 grame), urmată de media hibrizilor simpli (0.243 grame), a raselor parentale (0.235 grame) şi a tetrahibrizilor (0.203 grame). 17. Greutatea deşeului din gogoaşă arată superioritatea tetrahibrizilor, care prezintă media cea mai scăzută (0.031 grame), urmaţi de trihibrizi, monohibrizi şi rasele parentale. Pentru acest caracter coeficientul de variabilitate este ridicat, demonstrând neomogenitatea grupurilor experimentale. 18. În urma ierarhizării finale a raselor parentale, hibrizilor, trihibrizilor şi tetrahibrizilor studiaţi se poate concluziona că, luând în considerare parametrii urmăriţi, tetrahibridul (ABxB1)x(ACxAC29) este superior tuturor celorlalţi hibrizi, acesta înregistrând pentru 9 din cele 14 caracteristici urmărite valoarea maximă. El este îndeaproape urmat de hibridul simplu AB X AC, care a înregistrat valori maxime la 8 dintre caracteristicile urmărite. Pentru 6 caractere urmărite rasa parentală AC şi hibridul B1 X AC29 au prezentat cele mai ridicate valori; pentru aceste considerente putem recomanda aceşti hibrizi şi rase parentale pentru creşterile din zona Transilvaniei.
30

REZUMAT
BIBLIOGRAFIE SELECTIVĂ 1. BARLOW R., 1981, Experimental evidence for interaction between heterosis and environment in
animals, Animal Breeding Abst. 49, p.715- 739. 2. BENŢEA, M., 2006, Teză de Doctorat. USAMV Cluj-Napoca. 3. BURA M., 1992, Curs de tehnologia producţiei sericicole. Universitatea de Ştiinţe Agricole a
Banatului Timişoara. 4. DAS S.K., PATTNAIK S., GHOSH B., SINGH T., NAIR B.P., SEN S.K. AND SUBBA RAO G.,
1994, Heterosis analysis in some three way crosses of Bombyx mori L., Sericologia, 34 (1) p. 51-61. 5. EHIOBU N. G., GODDARD M. E., 1989, Heterosis in cross between lines of Drosophila
melanogaster selected for adaption to different environments. Theo. Appl. Genet., 77, p.253- 259. 6. GRIFFING B., ZSIROS E., 1971, Heterosis associated with genotype – environment interactions,
Genetics, 68, p. 443- 445. 7. HINAJAWA C., OHTSUKA M., 1975, Heterosis value in three- way and tetra- way crosses of
Bombyx mori L. hybrids, Theo. Appl. Genet., 43, p. 275- 278. 8. HIROBE T., 1985, On the recent advancement of silkworm breeding in Japan. Tamogawa Univ.
Japan, p. 157- 161. 9. IFRIM S., 1985, Ce ştim despre mǎtase, Editura Tehnicǎ Bucureşti,. 10. KNIGHT R., 1973, The relation between hybrid vigour and genotype and environment interaction.
Theo. Appl. Genet., 43, p.311- 318. 11. LERNER I. M., 1954, Genetic homeostasis, John Wiley and Sons Inc., New York. 12. MĂRGHITAŞ LIVIU AL., 1995, Creşterea viermilor de mătase, Ed. Ceres, Bucureşti. 13. MǍRGHITAŞ LIVIU AL., DEZMIREAN D., PAŞCA I., 2003, Sericicultura, Ed. Mediamira, Cluj-
Napoca. 14. MORAR R., DANA PUSTA, 1999, Zootehnie specială, III, Ed. Relief, p. 88. 15. MOROHOSHI S., 1949, Developmental mechanism in Bombyx mori. Meibundo, Tokyo. 16. NAGATOMO T., 1942 On the inheritance of the voltinism in the silkworm, J. Sericult. Sci. Japan,
Vol. 13, p. 114- 115. 17. NAKADA T., 1970, Researches on the sex- linked inheritance of the cocoon weight of reciprocal
crossings, J. Fac. Agric., Hokkaido University, 56, p. 348- 358. 18. OROZCO, 1976, Heterosis and genotype environment interaction. Theoretical and experimental
aspects. Bulletin technique, Departament de Genetique Animale, INRA, 24, p. 43- 52. 19. PAŞCA I., 2004, Teză de Doctorat. USAMV Cluj-Napoca. 20. PETRE, A., A. VLAIC, MĂRIOARA POP, 1989, Lucrări practice de Genetică animală, Tipo
Agronomia, Cluj-Napoca. 21. RAJASEKHAR, R., 1993, Studies on the temperature induced modulations on the growth and
reproductive physiology of silkworm, Bombyx mori L, PhD Thesis, S. V. University Tirupathi, India.
22. REDDY P. L., S.S. NAIK, P. M. MOHAN, 1998, Hatching pattern of silkworm, Bombyx mori L., as influenced by light intensity, Indian Journal of Sericiculture, 37 (2), p. 116- 122.
23. RUBAN P. S., CUNNINGHAM E. P., SHARP P. M., 1988, Heterosis and nutrition interaction in Drosophila. Theo. Appl. Genet., 76, p.136- 142.
24. SANG J. H., 1964, Nutritional requirements of inbred lines and crosses of Drosophila melanogaster. Chart. Res., 5, p. 50- 67.
25. SANTIAGO E., DIMINGUEZ A., ALBORNOZ J. PINEIRO R., IZOUIERDO J. I., 1989, Environmental sensivity and heterosis for egg laying in Drosophila melanogaster. Theor. Appl. Genet., 78, p. 243- 248.
26. SAVEL I., 1998, Mǎtasea naturalǎ, Ed. Ceres, Bucureşti. 27. SINGH R., SUDHAKOVA R. P., DATTA R.K., 1998, Studies on hybrid vigor in different crosses
of the silkworm Bombyx mori L., Sericologia, 38 (1), p. 155- 158. 28. SOHN K.W., 1983, Studies on variability in single, three- way and double crosses of silkworm,
Bombyx mori. Dissertation submitted to CSRTI, Mysore, India. 29. TACU, A., 1968, Metode statistice în zootehnie şi medicină veterinară, Ed. Agrosilvică, Bucureşti. 30. TAZIMA T., 1988, A view on the improvement of Mysore breeds. Proc. Intern. Cong. Trop.
Sericult. Bangalore, IV, 1- 5. 31. TSURUMAKI J., J. ISHIGURO, A. YAMANAKA, K. ENDO, 1999, Effects of photoperiod and
temperature on seasonal morph development and diapause egg oviposition in bivoltine race (Daizo) of the silkmoth, Bombyx mori L., Journal of Insect Physiology, 45, p. 101 – 106.
32. VLAIC, B., 2007, Teză de doctorat, USAMV Cluj-Napoca.
31

REZUMAT
32
33. WATANABE H., 1961, Studies on difference in the variability of larval body and cocoon weights between single cross and three- way cross or double cross hybrids in the silkworm, Bombyx mori., J. Sericult. Sci. Japan, 30, p. 463 – 467.
34. YOKOYAMA T., 1973, The history of sericulture science in relation to industry. In History of Entomology. R. F. Smith, T. E. Mittler and C. N. Smith (Eds.), Ann. Rev. Palo Alto, California, p. 267- 284.

UNIVERITY OF AGRICULTURAL SCIENCES AND VETERINARY MEDICINE CLUJ-NAPOCA
FACULTY OF ANIMAL BREEDING AND BIOTECHNOLOGY DEPARTMENT OF APICULTURAL AND SERICULTURAL
TECHONOLOGIES
PAOLO DE ANGELIS
RESEARCH REGARDING THE MAIN BIOLOGICAL AND
TECHNOLOGICAL INDEXES OF SILKWORMS HYBRIDS
BRED IN TRANSYLVANIA AREA
SUMMARY OF PhD THESIS
SCIENTIFIC COORDINATOR
Prof. Liviu Al. Mărghitaş, PhD
Cluj-Napoca
2009
33

SUMMARY
THE AIM AND THE OBJECTIVES OF RESEARCH
The aim of researches in two experimental years was the races and hybrids hierarchy function of the main biological index of larvae, the biological index of crude cocoons and technological index of dry cocoons and silk thread.
The research objectives were related at the following aspects: 1. The compared study of parental races and simple hybrids; 2. The compared study of parental races and trihybrids 3. The compared study of parental races and tetrahybrids.
It was study the incubation and hatching, the biological index of larvae, the biological index of crude cocoons and the technological index of dry cocoons and silk thread.
The results obtained in two experimental years for the four experimental races, twelve simple hybrids, eight trihybrids and two tetrahybrids were used for a comparative study between parental races and these hybrids; the evolutionary differences between these were study in two experimental years, making possible the hierarchy function of the main biological and technological index.
The index studied for parental races and hybrids resulted from those are:
1. Eggs incubation and hatching of silkworms 2. Larvae’s biological indexes ♦ Larvae’s weight ♦ Larvae’s length ♦ The larval period ♦ The weight of the serigene gland ♦ The cocoon making percent 3. The biological indexes of the crude cocoon ♦ The weight of crude cocoons ♦ The longitudinal axis of cocoons ♦ The transversal axis of cocoons ♦ The weight of the silk incartament ♦ The silk percent in cocoons 4. The technological indexes of the dry cocoon and fiber’s ♦ The weight of dry cocoon ♦ The length of the silk wire ♦ The weight of the silk wire
34

SUMMARY
CHAPTER I THE PRESENT STAGE OF RESEARCH AND HISTORY OF
SILKWORM BOMBYX MORI L. BREEDING IN ROMANIA AND WORLDWIDE
Sericulture is a science which studies the biology and reproduction and the
breeding of all species which produce cocoons whit the aim of natural silk production. The cocoons and thread processing, the purring and the fabric execution all form the object of silk industry.
Between all species which create cocoons from silk thread 90% is occupied by mulberry silkworm, followed by castor-oil plant and oak silkworms; the spread of these species is give by the areas where the vegetal species are growing. The area in which the silkworms are breeding is generally more limited than that of vegetal species can be found.
The name of sericulture came from Latin word “ser-seris”, which means „silk”, and the scientifically name of silkworm is Bombyx mori L.
Sericulture represent a gainful and a easy pursuit, which can be practiced by children and old persons, and the breeding can bee made in simple rooms which are not used during summer, and the growing period is very short, being around of 30 days (Yokoyama T., 1973; Ifrim P., 1985; Mărgitaş L. Al., 1995; Savel I., 1998).
The originally country of silk is China. The historical and archeological researches realized here highlight that around the little city Schengze, from Wuxion region, Jiangsu province, the citizens cultivated mulberry trees and they growing silkworms; they knew the pursuit of thread and fabric making.
The art of silkworm breeding and their use is assigned to the Si-Ling-Schi Chinese princess, who was the principle wife of famous king Hoang-Ti, called the Yellow-King (2699- 2599 b.c.).
For the old Chinese, the sericulture became a sacred pursuit, which was a secret, protected by imperial decrees. The disclosure of this secret was the death punish. For 3000 years they knew to keep this biological secret and the production and selling between the China borders. The Chinese brought the silk fabrics on the “silk way” in the Oriental Turkistan markets, where the mediterranean merchants came to buy its.
The secret spearing worldwide regarding the silk production took place around the year 200 b.c., when the Chinese emigrants arrived for the first time in Korea. From here the sericulture spread in all Asian world, and in 552 AD, on the time of King Justinian, two Persians monks, after they learn the art of larvae’s breeding and thread obtaining, they bring in bamboo’s sticks the eggs of silk butterfly at Constantinople. From here the silkworm breeding and cocoons processing was spread in all Byzantine Empire. Then the arabs spread this pursuit in Caucasus and North Africa throw Spain, and from here pass in Sicily, South and North Italy, then in France, where the first mulberry trees were cultivate around the year 1260.
Nowadays, sericulture is spread in 36 countries, and the world cocoons production is about 450.000 tone and is realized principally by China (41%), Japan (23%), India (9%), South Korea (9%), Russia (9%) etc. (Mǎrghitaş L. Al., et al,
35

SUMMARY
2003). On the first place in silkworm production is China (27 000 tone), followed by Japan (9 200 tone) and India (9 000 tone); the last one show the tendency to become the second silk producer from entire world. Due to the favorable conditions for mulberry exploitations, in our country the sericulture developed very fast, and the indigene eggs are free of disease, being very appreciated by other countries (Mǎrghitaş L. Al. et al, 2003). From the existing data results that in 1860s our country exported the impressive quantity of 6064 kg of silkworms eggs. In this sense in 1840-1860s between the measurement adopted of sericulture development was the founded of nurseries for mulberry productions.
In the last 50 years the cocoons production in our country increased permanently at about 500 tone (1944s), arriving at 2000 tone (1989s). After 1990s, due to the economic crisis, the crude cocoons production fell year after year, in arriving in 1998s at a production of 3 tones of crude cocoons on year. Nowadays the production is at a worrying level. Regarding the gross silk production, this suffered a powerful regression, from 120 tones (the level in 1989s) at 20 tones (1991s), 6 tones (1995s) and 2 tones (2001s). This regression toke place due to the stops of spinning activity from Lugoj; this area has a huge sericulture potential (Paşca I., 2004). In the last years, in Romania were done laborious researches, which given good results regarding the silkworm’s disease, and a lot of publications were made regarding the polyhedron, named the yellow-disease or silkworm jaundice, being a disease produced by a virus.
Now the sericulture in Romania and worldwide pass a crisis period and the cocoons production is in extinction. In our country the sericulture degenerated when the spinning from Lugoj was closed. Even in the last years the sericulture suffered this „right debasement”, at the research level it continue the activity, being very productive from this point of view. The gene bank is maintained and new races and hybrids of silkworms were created, using the gene bank existing at S.C. SERICAROM S.A. Băneasa (over 60 races of silkworms).
CHAPTER II ZOOLOGICAL SYSTEMATIC OF SILKWORM AND EVOLUTIVE
CYCLE OF BOMBYX MORI L. SPECIE
2.1. ZOOLOGICAL SYSTEMATIC OF SPECIE
The mulberry silkworm take parts of: Phylum: Artropode Underphylum: Tracheate Classa: Insecta (Hexapoda) Underclass: Pterigote Section: Neoptere Undersection: Metamorfe Order: Lepidoptere superioare,
36

SUMMARY
Series: Metaheterocere sau Lepidoptere nocturne superioare Family: Bombycidae Gens: Bombyx Species: Bombyx mori
2.2. SILKWORM’S SPECIES
The most know species of silkworms whose cocoons can be used for silk production are: Bombyx mori L., Anthereae yammai Guer L., Anthereae assaamensis L., Anthereae pernyL., Theophyla mandarina L., Bombyx arrindia L., , Samia coecropia L., Saturnia pyri L., Saturnia pyretorum L., Attacus atlas L., Philosamia cynthia ricini L., Lasiocampa atus L., Bodecera madagascarensis L., etc. Frome those, the most spread one is Bombyx mori L. (which is considered the most valuable from all of them), followed by Anthereae perny (the oak silkworm, feed whit oak leaves) and Philosamia cynthia ricini, (the castor-oil silkworm, feed whit castor-oil leaves). The other species leaves wildly in China, India and other countries, some of them being used for crossing whit different domestic’s silkworm’s races.
2.3. EVOLUTIVE CYCLE OF BOMBYX MORI L. SPECIE
The mulberry silkworm or silkworm butterfly (Bombyx mori L.) takes part from the group of insects which has a complete metamorphosis, in its evolutionary cycle passing through 4 different stages: egg (or seed), larvae (worm or caterpillar), chrysalides (nymph or pupae) and butterfly (or adult), having one (monovoltine race) or more generations (polivoltine races) per year. After fertilization the embryogenesis process take place for three days, then diapauses started. These are during function of race, and function of genetics’ and environmental factors, and at the new formed races this take place for 9-10 months, generally from July to April. The eggs maintained in optimal conditions are incubated for 10-14 days, and this process is closed whit the larva’s hatching. The larval stage varies function of race, being between 25 and 35 days, and the larvae’s are passing thought five instars, delimitated by resting periods, called „sleep”. During these sleeping periods the larvae doesn’t eat and the skin changing has place.
CHAPTER III
SILKWORM BREEDING, CROSS-BREEDING AND HETEROSIS
3.1. SELECTION The selection represent the first and the most important method of breeding,
being defined like a complex of works consisting in choosing and mating the organisms whit the aim of improving the existing races or creating the new ones. By
37

SUMMARY
selection are maintained for mating the corresponding samples for proposed aim. In this way, the selection represents a „reproductive discrimination” (Bura M., 1992) realized by man through choosing of some genotypes and the remove of others. By selection is pursuing the obtaining of superior’s forms for one or more biological, technological or economical characteristics. The selections specify is determined by a series of silkworms feature, such as: huge prolificacy, short biological cycle which permits to obtain of more generation per year, and the impossibility of mating the descendents whit the parents.
3.2.CROSS-BREEDING
The cross-breeding represent the second step of breeding process. For this aim the selected samples for reproduction, appertained of two or more races are mating directed between them, for realize a favorable combination for followed characteristics.
From the 18th century was observed that the products obtained after cross-breeding (hybrids) overcome the values of genitors, for viability, vitality and productive characters.
3.3. CONSANGUINITY
The consanguinity represents the proceeding by which the related samples
appertaining at the same race are mating between them, whit the aim of fixing some valuable characters or forming consanguine lines. It represents one of the methods used in creation process and breeding of some types or animal’s races. The consanguinity permits the hereditary fixing of valuable qualities and the increase of some qualities, the elimination from a race at the undesired samples and the increase of hereditary transmission.
3.4.HYBRID VIGOUR FOR DIFFERENT CHARACTERS
During the different studies on mulberry silkworm was observed that some characters like the eggs number/clock, the hatching percent, the period of larval stage, the viable adult larvae’s number, the cocoon’s weight, the silk incartament weight or the silk length doesn’t manifest the heterosis phenomenon whit the same intensity.
3.5. DIFFERENT CROSS-BREEDING SYSTEMS AND HETEROSIS
Hirobe T., (1985) observe that the highest heterosis value is obtained after
simple crossing between genetically distinctive populations and this decrease progressively in the next order: F1>F2>F3>F4….In the case of silkworm the heterosis phenomenon disappearing completely in F14 generation. The silkworms “can be combined” after some crossing like: simple crossing (AxB), triple crossing [(Ax B) x C] and quadruplicate crossing [(Ax B) x (Cx D)] crossing. The type of practiced crossing influence the heterosis value, the hybrids
38

SUMMARY
resulted after simple crossing representing usually a hybrid vigour bigger comparatively whit those obtained by triple or quadruplicate crossing (Sohn K.W., 1983; Das S.K., 1994; Singh R. et al., 1998).
The study made on quantitative characters at parental races and hybrids obtained by different crossing systems shows that the variability for quantitative characters is smaller at the hybrids obtained by simple crossing comparative whit parental races and hybrids obtained by triple or quadruplicate (Watanabe 1961; Sohn, 1983; Yokoyama, 1973).
3.6. HETEROSIS AT RECIPROCAL CROSS-BREEDINGS
In the case of silkworm a lot of quantitative characters are straight connected
whit the nature of voltinism and the maturating period, and these characters are sex connected (Nagatomo, 1942; Morohoshi, 1949; Nakada, 1970, 1972; Tazima, 1988). The expression of heterosis can vary a lot in the case of reciprocal crossing at the silkworm.
It is well know that if a hybrid presents a high heterosis for a character, its heterosis can decrease under the value of its reciprocal hybrid for another character.
3.7. HETEROSIS AND ENVIRONMENT
Lerner (1954) after numerous researches observed that the heterozygous are
much more protected than homozygous against the environmental changes (variation of temperature, humidity, air, growing space, etc.). So, heterozygous should presented fewer variations than the parental races for the same or different environment, they being much uniforms from this point of view. The degree of manifestation of heterosis obtained for a crossing can be influence by the environment (Barlow 1981).
The concept under which the interactions genotype- environment affects the heterosis amplitude was approached by numerous researchers at different animal species (Sang, 1964; Knight, 1973; Orozco, 1976; Griffing and Zsiros, 1978; Ruban et al., 1988; Ehiobu, Goddard, 1989; Santiago et al., 1989).
3.8. VOLTIMISM TYPE AND HETEROSIS
The most studies realized until now (Hirobe, 1985; Udupa and Gowda, 1988; Nagaraju, 1990) shows that the hybrids obtained by triple and quadruplicate crossing are lowers that those obtained from simple crossings. The results obtained by Nagaraju (1990) for the hybrids obtained by triple crossing (polivoltines x polivoltines) x bivoltines and (polivoltines x bivoltines) x bivoltines join to these observations.
Such a difference between the hybrids obtained by simple crossings, triple and quadruplicate can be interpreted like a consequence of the fact that one of the parents implicated in triple crossing and the bought parents from quadruplicate crossing are they self F1 exemplars. More than that, the population obtained from
39

SUMMARY
triple or quadruplicate hybrids is a mixture of genotypes which can be obtained by simple crossings.
Hinajawa and Ohtsuka (1975) used for the first time an anticipation system for hybrids resulted by triple and quadruplicate crossing performance using the performances of simple’s crossing.
CHAPTER IV
SILKWORM’S RACES AND HYBRIDS
4.1. GENERAL DESCRIPTION OF BOMBYX MORI L. RACES AND HYBRIDS
By „silkworms race” notion is described a group of individuals reproductively isolated, which present some requesting for environmental conditions. Within the races can be differentiated lines, which represents genetically under populations, reproducible isolated, which evolves under the selection influence; these groups differ by morphological or productive characters. From origin point of view the silkworm races represents biological entities or selected by locale populations with a high degree of adaptability at different environmental conditions.
The 60’s mulberry silkworm’s races are classified function of some criteria, like: the race’s origin, the color and the cocoon’s shape, voltinism, etc. In this way, function of voltinism (number of generations per year) can be distinguished:
- mono-voltine races (with one generation per year); - bi-voltine races (with two generations per year); - poli-voltine races (with 5- 8 generations per year).
Function of origin the races are classified in the next groups: - Japanese races; - Chinese races; - European races; - Tropical races.
Functions of cocoons shape are distinguished: - races with oval cocoons; - races with cylindrical cocoons; - races with spherical cocoons; - races with fusiform cocoons; - races with belted cocoons.
Functions of cocoons color are distinguished: - races with white cocoons; - races with yellow cocoons; - races with golden cocoons; - races with green cocoons; - races with pink cocoons.
Functions of hybridization degree are distinguished: - simple hybrids - obtained by directly and reciprocal crossing of two races of
silkworms.
40

SUMMARY
- triple hybrids - obtained by directly and reciprocal crossing between a simple hybrid and a silkworm race.
- quadruplicate hybrids – obtained by directly and reciprocal crossing between simple hybrids of silkworms.
4.2. PRODUCTIVE CHARACTERISTICS OF BOMBYX MORI L. RACES
In this subchapter are presented the productive characteristics of the next races: Alb Cislău, Băneasa 75, Alb Chinez 29, Alb Orşova, Alb Băneasa, S – 76, Alb Băneasa 1, Hebar 2/1, Hebar 1/18, EM5, SK2/F, N5, Saniş 8, Belokokonnaia 1, J90. 4.3. PRODUCTIVE CHARACTERISTICS OF BOMBYX MORI L. HYBRIDS AND POLY-
HYBRIDS The data regarding the biologically and technological characters of hybrids and poly-hybrids are extremely limited in literature; in this chapter are described the following hybrids: simple hybrid Băneasa 28, simple hybrid Băneasa 29, simple hybrid Cislău 8, simple hybrid Super 1, tetra-hybrid Băneasa 28 x Băneasa 29, tetra-hybrid Băneasa 8 x Băneasa 9, tetra-hybrid Băneasa 75 x Băneasa 9, tetra-hybridul Băneasa 76 x Băneasa 4, tetra-hybrid Cislău 8 x Cislău 1.
CHAPTER V BREEDING TECHNOLOGIES OF MULBURRY SILWORMS IN
PEDOCLIMATERIC CONDITIONS OF TRANSYLVANIA
The breeding technology of mulberry silkworms contains mainly aspects of eggs incubation, hatching, microclimate conditions and nutritional aspects.
The incubation is a complex represented by embryo development from egg and it is influenced by the insurance of optimal conditions of temperature, humidity, light and air. It can be natural or artificial.
The space preparation is realized during spring times; a cleaning and a space and used inventory disinfection are necessary. After itself cleaning of space, the wood inventory is washed with caustic soda 2-5%, then is washed again with clean water and then is dried. The disinfection is realized with formalin 3%, after the sealing of the incubation space. Under the condition of 23 – 250C, the formalin solution is applied and the space is left close for 2 days. After this disinfection the glue and the cleaning is made, including the doors, the windows and the floor.
The incubation technology consists in eggs entering in incubation cassette in 2 mm thickness layer; on those are put a cotton net, a perforated sheet; those are fixed whit a frame.
The temperature has an important role, and it influence the embryo development (Ifrim, 1998; Reddy et al, 2002), generally being used the method of constant temperature and the gradual lifting of temperature.
41

SUMMARY
The incubation period takes normally 12-14 days and is strictly connected whit the microclimate conditions described before.
The hatch is the process by which the larvae leaves the egg by micropil and takes place in 2 – 3 days. Whit 2- 3 days before hatching the larvae consume the serous shell of egg, this decolorized himself and inside of it can be heard clicks. The hatching takes place between 5 and 11 a.m. In the firs day are coming out the “spies”, which are the first larvae hatched and which are not maintained for breeding. In the first day 43 to 44% of larvae are hatching, in the second between 40 and 42%, and the rest ones in the third day. The hatched larvae after the third day are removed for realize than breeding groups as uniform as possible.
An important role in silkworm breeding technologies is occupied by the space insurance. The places recovery can be realized by increasing the useful space and the vertically disposure of larvae.
The young larvae are more resistant at high temperature; a normal development of larvae takes place only if the next temperatures are respected: 26-270C for the first and second instar, 24-250C for the 3rd instar, and for the 4th and the 5th instar 23-240C. The overcoming of these values creates the physiological process intensification, having an unwanted effect on organism’s viability (Rajasekhar R., 1993; Tsurumaki J. et al., 1999).
The growing surface is other important factor of silkworm breeding and this assure a uniform feeding of larvae. For one gram of eggs is necessary a surface of 0.1 m2 in the first instar, 0.2 m2 in the second instar, 0.5 m2 in the third stage, 1.5 m2 in the 4th instar and 3.0 m2 in the 5th instar.
The leaves harvesting must be done during morning or evening on the cool weather and the warm period will be avoided because the leaves has a low nutritional value (Vlaic, B., 2007). The leaves administration on shoots is made 3-4 time per day, in this way the leaves withering slowly.
Mărghitaş L. Al., (1995) and Morar R., (1999) recommends for the larvae breeding from one gram of eggs 35-40 kg mulberry leaves administrated as follows: in the first instar 0.2 kg, in the second instar 0.6 kg, in the third instar 2 kg, in the fourth instar 5 kg, and in the fifth instar 26-31 kg.
The failure of these conditions can have serious consequences, the moult became impossible, and the larvae’s death takes place. The first feeding after moult must be done carefully because the new skin of larvae is very sensitive and can be easily hearted.
CHAPTER VI
NATURAL FEED AND DIFFERENT FEED ADDITIVES ADMINISTRATED TO SILKWORMS
Nutrition is an important factor of silkworm breeding. Researches regarding this argument knew a real progress after the appearance and the use of artificial diet; important benefits were done for the industry. The only stage in which the silkworms are fed is the larval one. In this stage the reserves of nutrients are accumulate and there are enough for the chrysalides and butterfly stage.
42

SUMMARY
The main feed source for Bombyx mori specie is represented by mulberry leaves, these containing enough quantities of proteins, carbohydrate, fats minerals and water. The assimilation of those nutrients depends on microclimates conditions provided to larvae and on the leaves quality.
CHAPTER VII MATERIALS AND METHODS
7.1. EXPERIMENTAL MATERIALS
7.1.1. Biological material
The biological material used in two experimental years consisted in 4
silkworm races: Alb de Băneasa (AB), Băneasa 1 (B1), Alb de Cislău (AC), Alb chinezesc line 29 (AC29), from the genetic bank of Bombyx mori L. specie, provided by S.C. Sericarom Băneasa S.A (Bucarest). It were study the next hybrids, too:
♦ 4 simple hybrids: ABxAC, ACxAB, B1xAC29 and AC29xB1; ♦ 8 trihybrids: AC29x(AB x B1); (AB x B1)xAC29; (AB x B1)xAC;
ACx(AB x B1), (AC x AC29)xB1; B1x(AC x AC29); (AC29 x AC)xAB and ABx(AC29 x AC);
♦ 2 tetrahybrids: (AB x B1) x (AC x AC29); (AC x AC29) x (AB x B1); The breeding of races and hybrids previously mentioned were realized in
bought experimental years (2007 and 2008) in two rehearsals (C1 and C2), using 300 larvae/rehearsal, and all data presented in the following tables and graphics from the next chapters represents the average values of two rehearsals done in the same experimental year.
The number of studied samples was different function of analyzed characters. For the biological parameters of eggs (prolificacy, number of hatched eggs and hatching percent) the results is the expression of 5 different clocks of races and hybrids; the larvae weight was studied for 20 exemplars, the larvae length for 10, the weight of the serigene gland for 15 exemplars, the crude cocoons weight was determined for 100 larvae for every sex, the incartament weight and the silk percent for 15 cocoons, and the longitudinal and transversal axes and the thread length for 10 larvae.
7.1.2. Materials used in organization of breeding batches
The silkworm breeding was possible due to the existence of mulberry plantation appertained to Morus alba specie, from Ukraine 107 variety, in the University of Agricultural science and Veterinary Medicine. This variety, original from Ukraine, is cold resistant and is suitable for Transylvania climacterics conditions, being very resistant at bacteriosis.
43

SUMMARY
From physical and qualitative point of view the leaves were superior. The harvest was done on cold weather, during morning and evening, because in these moments of the day the leaf has a high nutritive value.
The leaves administration was done 4 times on day. The leaves were kept in a dark room at 100C, and before administration it
was putted in a room for feed preparation at 250C. For the younger larvae the leaves were cut before administration at 0.5cm thickness. For the larvae from the second instar the leaves thickness was 1-2 cm, and for the larvae from the third instar this was 2-3 cm. Before sleeping period the leaves were not cut for avoiding the premature fading of those and for maintain the larvae’s health.
For keeping the leaves freshness for a long period of time the larvae from the 4th and the 5th instar were fed with leaves left on the branches.
The space for experimental series breeding has a 40 square meter whit nordic position.
Before starting the experimental series growing, the growing room, de sericulture module and the entire inventory necessary were mechanical washed, and then disinfection with formaldehyde steams was done. For a good disinfection all windows and doors were closed, and for a high efficiency the indoor temperature were increased at 300C, and the air humidity at 85%. The steams were let to action for 48 hours, and then the space was ventilating for 3 days.
At the entrance a disinfector containing sodium hydroxide 2% was fixed. In the growing space overlapped shelves were mounted for four levels
(Figure 1); they have stable feet and mobile growing surfaces. The distance between growing beds is 50 cm, and that one between bed and floor is 60 cm, for temperature differences equilibration. In this manner uniform conditions are assured for all studied larvae. On the growing bad a white paper was sat and then the hatched larvae.
The bed changing was realized using perforated paper. The holes made in paper presents different diameters function of larvae’s instars: 6 mm for the first three instars, 10 mm for the fourth instar and 13-15 mm for the last instar of larval stage. In the sleep period the larvae are not fed, and they are started to eat when more than 90% of them returned for the sleep period. The larvae which from different reasons can not change theirs tegument soon will be death, because the old tegument is to small for permitting their growing.
44

SUMMARY
Figure 1. The familial sericulture module (MSF) (original)
An important factor in the growing period of larvas is their density. Between two larvae a space equal with their thickness was assured during growing period.
7.2. Experimental method
The experiments take place in the Department of Apicultural and Sericulture Products Technologies from University of Agricultural Science and Veterinary Medicine Cluj-Napoca and at S.C. Sericarom Băneasa S.A. (Bucarest), in 2007 and 2008.
The silkworm’s larvae from experimental variants were fed with leaves harvested from mulberry plantation held by University; its surface is equal at 2 ha and it’s found in Didactical Farm Cluj-Napoca.
The data’s regarding the followed index (detailed in Chapter 7.1.) were gradually recorded during the experimental period in 2007 and 2008, by measurements and weighing and the data’s statistical interpretation was made using recognized statistical methods (Tacu, A, 1968; Petre A. şi col., 1989).
45

SUMMARY
CHAPTER VIII
RESULTS AND DISCUSSIONS
8.1. THE INCUBATION AND HATCHING OF SILKWORMS
8.1.1. Hatching biological parameters of silkworm breeds and hybrids
For eggs incubation of studied breeds and hybrids the method of gradually increased temperature was used; this technique was described by Mărghitaş, L. Al., (1995): from 150C maintained for 3 days, at 18 - 200C, for 2 days, the temperature is increased then at 23-240C for 4 days, and at the end of this period the eggs were maintained at 25-260C for all the rest of the period until the hatching had place. The air humidity was between 75-85%, being higher in the last part of hatching period, for helping the hatching.
An important parameter is represented by hatching percent; the lowest percent was obtained for Alb chinezesc line 29 breed (90.76%), and this value is similar at that obtained by Benţea (2006) (90.71%). The maximum value for this parameter was registered for Alb de Cislău breed (95.42%), this value being very closely by those obtained by others researchers (Paşca, 2004; Benţea, 2006). The average of hatching percent of these breeds was 93.68%.
The average number/ hatching at the four simple hybrids varies between 558 at AC29xB1 and 694 at ABxAC, and this hierarchy is maintained for the number of hatching eggs.
Regarding the hatching percent the minimum value of this was recorded at ACxAB breed, 94.78%, and the highest percent of 96.90% was recorded for ABxAC and B1xAC29 hybrids. The average of hatching percent for the four simple hybrids was 96.36%. The presented data shows an increasing of 2.68% for the average of simple hybrids, this fact being explained by heterosis effect, manifested from the egg’s stage.
The average number of eggs in the case of trihybrids varies between 619 at the trihybrid B1x (ACx AC29) and 749 at (ABx B1)xAC29. The minimum number of hatching eggs was obtained at (ABx B1)x AC, 616 eggs, and the maximum number was registered at (ABx B1)x AC29, this registering 730 eggs from which resulted larvae. The hatching percent presented variations between 94.06 for the trihybrid (ACx AC29)x B1 and 97.67% for AC29x (ABx B1). The hatching average at trihybrids was 96.61%. In this case the difference of 2.93% was registered at the hatching percent comparative whit that obtained for parental breeds, and it can be considered significant.
A variation regarding the average number of eggs per hatching between 672 at tetrahybrid (ACx AC29)x (ABx B1) and 785 at hybrid (ABxB1)x (ACx AC29) can be observed. This difference is probably due to the different combinative capacity of those. The hatching eggs vary in the same way at two studied tetrahybrids, and the hatching percent was 98.22 at (AB x B1) x (AC x AC29) and 97.76 at (AC x AC29) x (AB x B1).
46
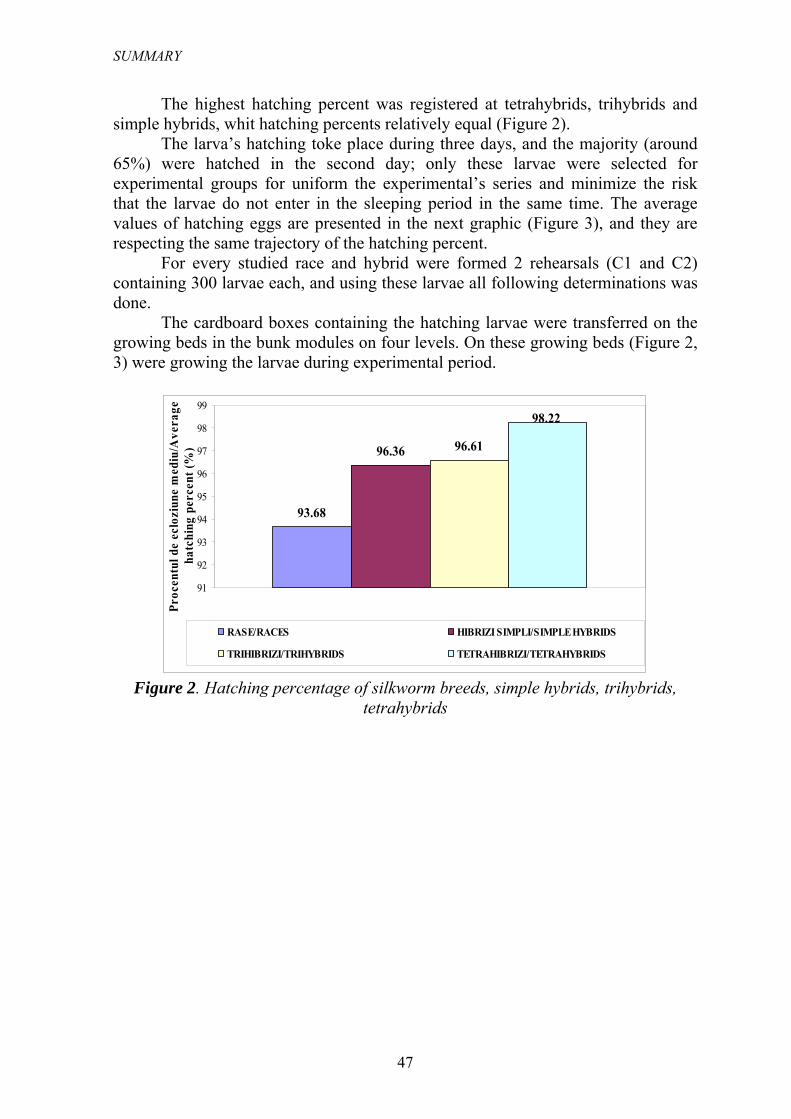
SUMMARY
The highest hatching percent was registered at tetrahybrids, trihybrids and simple hybrids, whit hatching percents relatively equal (Figure 2).
The larva’s hatching toke place during three days, and the majority (around 65%) were hatched in the second day; only these larvae were selected for experimental groups for uniform the experimental’s series and minimize the risk that the larvae do not enter in the sleeping period in the same time. The average values of hatching eggs are presented in the next graphic (Figure 3), and they are respecting the same trajectory of the hatching percent.
For every studied race and hybrid were formed 2 rehearsals (C1 and C2) containing 300 larvae each, and using these larvae all following determinations was done.
The cardboard boxes containing the hatching larvae were transferred on the growing beds in the bunk modules on four levels. On these growing beds (Figure 2, 3) were growing the larvae during experimental period.
93.68
96.36 96.61
98.22
91
92
93
94
95
96
97
98
99
Proc
entu
l de
eclo
ziun
e m
ediu
/Ave
rage
ha
tchi
ng p
erce
nt (%
)
RASE/RACES HIBRIZI SIMPLI/SIMPLE HYBRIDS
TRIHIBRIZI/TRIHYBRIDS TETRAHIBRIZI/TETRAHYBRIDS
Figure 2. Hatching percentage of silkworm breeds, simple hybrids, trihybrids,
tetrahybrids
47

SUMMARY
555592 593
715
0
100
200
300
400
500
600
700
800
num
ar o
ua e
cloz
iona
te/n
umbe
r of
hat
chin
geg
gs
RASE/RACESHIBRIZI SIMPLI/SIMPLE HYBRIDSTRIHIBRIZI/TRIHYBRIDSTETRAHIBRIZI/TETRAHYBRIDS
Figure 3. Average hatching number of silkworm breeds, simple hybrids, trihybrids, tetrahybrids
8.2. EVOLUTIONARY STAGES OF LARVAE
The larval stage in the case of parental race lasted 30 days, from 18.05.2007
until 16.06.2007 in the first experimental year and 31 days, from 15.05.2008 until 14.06.2008, in the second experimental year. The differences between these periods and those reported in the literature are insignificant, being of maximum 2 days (Paşca, 2004). The studied hybrids have the larval stage insignificant shorter (with 1-2 days) comparing with parental races, and the differences between them were only for some hour in the both experimental years.
8.2.1. Climatic conditions during experimental period
The climatic conditions in experimental years 2007 and 2008, meaning the average temperature and relative humidity’s of the air during the experiments were registered by automated Meteorological Station from University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca, and the differences between them and the normal values for the period of time are showed in the next graphic (Figure 4).
48

SUMMARY
15.519.31 17.94
68.35 69.3 67.5
0
10
20
30
40
50
60
70
80
Conditii normale 2007 2008
grad
e C
elsi
us s
i %/C
elsi
us D
egre
es a
nd
%
Temperatura/Temperature
Umiditateaaerului/airhumidity
Figure 4. Graphical evolution of temperature and air`s humidity in may-june 2007
and 2008
8.2.2. The microclimate conditions supplied at experimental groups
For the good development of the larvae’s growing the assurance of optimal conditions of temperature, humidity, ventilation, brightness, space and hygiene is very important.
The strictly control of these parameters from growing room ensure the success of larvae’s development; from this aspects depends the cocoon’s making and the obtaining of a high production of cocoons and a high qualitative silk; as we showed before all these parameters influence the duration of larval stage.
For temperature and humidity control the termo-hydrograph was used; it register the temperature and air’s humidity from growing room, and these characteristics are showed in Figure 5.
26.5 26.5 25.6 23.5 22.5
82.5 82.5 82.5
70 67.5
0
20
40
60
80
100
120
1 2 3 4 5
Varsta larvelor/Larvae's age
grad
e C
elsi
us/% Umiditate/Air
hymidity
Temperatura/Temperature
Figure 5. Microclimatic conditions ensured in growing room
49

SUMMARY
8.2.3. The breeding technologies
8.2.3.1. The breeding supports and their sanitation The growing beds of this module are formed by a wood rectangular border.
The larvae’s support is formed by a string net with holes, adapted to stretching and which can be fixed on the bed’s border. These supports are constructed in such a manner that they are easily to manipulate and they are cheap. The distance between two beds is 50 cm, and the distance between the first bad and the floor was about 60 cm. This fact is due to the necessity of temperature smoothing between the first and the last growing bed.
The bed’s sanitation was done by brushing, using caustic soda 5%; a rinsing with clean water was done, and then the beds were dried.
On the growing surface were put sheets of paper about 100/70 cm, and over them were settle the hatching larvas.
The space assured to the larvae’s hatched was 1m2/300 larvae, as the literature provides; around 1000 larvae can be obtained from 1 gram of eggs, and those need a space of 2,5-3m2 (Mărghitaş, 1995).
8.2.3.2. The bed changing
For bad changing the technique is the following: first of all the sheet of paper
is put on the larvae’s, during the morning, before the feed administration, and over this is sprinkle the fresh leaf, possibly cut it for being attractive for the larva’s. They are climbing by the holes on the paper surface.
8.2.4. The succession of the larvae instars and their sleep
The first larval instar last 3-4 days and was followed by a day of sleep; the
second instar last 2-3 days and it was followed by a day of sleep; the third larval instar last 3-4 days and has finished with a day and a half of sleeping period. The forth larval instar last 4-5 days, and at the end of it the larvae passed by two days of sleep. The last larval instar (the fifth) last 8-9 days, and at its end the cocoon’s making took place.
8.2.5. Natural larvae feed - harvesting, processing and administration
The feed used during experiments for all larval instar, for the studied races and hybrids was the mulberry leaves harvested from Ukraine 107 specie of mulberry trees from plantation appertained to USAMV Cluj-Napoca. It was administrated in the same number of halves at all variants. For the larvae from the first three instars the leaves were administrated mashed (Figure 6), and then wholly or not stemmed from the branches.
The leaves harvesting was realized during the morning and the evening because in this moments of the day the leaves have the highest contents of nutrients,
50

SUMMARY
and in this manner it is possible to assure at the larvae the necessary proteins and minerals. The leaves administrated were clean, without pollutants, and the transport in the growing room was done whit vehicles in the shortest time.
Figure 6. The administration of mashed leaves (original)
8.3. BIOLOGICAL INDEXES OF LARVAE
8.3.1. Larvae weight
For determine these morphological measurements 20 larvae were random, and weight’s determination was done using an electronic balance. In the fourth larval instar an only one determination of the body weight was done, in the fourth day, and in the fifth instar two measurements were done, in the second and in the fifth day.
In the next graphic is represented the evolution of body weight at studied races and hybrids (Figure 7).
51

SUMMARY
0.37
3
0.43
9
0.37
5
0.36
1
0.37
3
0.38
5
0.44
2
0.37
1
0.36
9
0.37
2
0.37
3
0.36
3
0.36
2
0.42
2
0.37
2
0.37
2
0.34
6
0.35
4
1.26
1.19 1.
42
1.26 1.35
1.30 1.47
1.36 1.40
1.05 1.
37
1.15 1.26 1.39
1.30
1.32
1.25
1.19
5.87 6.
74 6.99
5.80
7.37
7.00 7.11
6.55 6.79
6.46
6.00
5.67 5.84
5.88
5.73
5.76
6.79 6.99
0
1
2
3
4
5
6
7
8
AB
AC B1
AC
29
AB
X A
C
AC
X A
B
B1 X
AC
29
AC
29 X
B1
AC
29x
(ABx
B1)
(ABx
B1)
x A
C29
(ABx
B1)
xAC
AC
x (A
BxB1
)
(AC
xAC
29)x
B1
B1x(
AC
xAC
29)
(AC
29xA
C)x
AB
ABx
(AC
29xA
C)
(ABx
B1)x
(AC
xAC
29)
(AC
xAC
29)x
(ABx
B1)
Rasele si hibrizii/Races and hybrids
Gre
utat
ea/W
eigh
t (g)
Greutatea medie a larvelor in ziua 4 a varstei a IV-a
Greutatea medie a larvelor in ziua 2 a varstei a V-aGreutatea medie a larvelor in ziua 5 a varstei a V-a
Figure 7. The comparative evolution weight of larvae breeds, simple hybrids, tri-hybrids, and tetrahybrids from experimental’s groups
8.3.2. Larvae length
The measurements regarding the larvae’s length (table 1) were done in the
same days of body’s weight, using measuring tool, and a number of 20 samples were measured for every experimental group.
Comparing the results obtained after weight and length determination it can be observed that, generally, (excepting the simple hybrids) the body’s weight is directly correlated with theirs length, and this fact demonstrates that these two characteristics are directed correlated.
8.3.3. The weight of the serigene glands
The next graphic (Figure 8) presents that the obtained values for the weight
of serigene gland and the variability coefficient of those. After a hybrids hierarchy function of followed characters until this point of
researchers we can conclude that at the simple hybrids it can be remarked B1 x AC29 for the larvae’s length and the weight of serigene gland, and for the larvae’s weight the hybrid AB x AC. Trihybrids AC29 x (AB xB1), (AC x AC29) x B1 and AB x (AC29 x AC) demonstrates a special larvae’s weight and serigene gland (this aspects demonstrates once again a special race’s combinative capacity); between the studied tetra-hybrids it remarks the (AC x AC29) x (AB x B1) for the larvae’s weight and length and it’s reciprocal hybrid (AB x B1) x (AC x AC29) can be remarked for the weight of serigene glands.
52
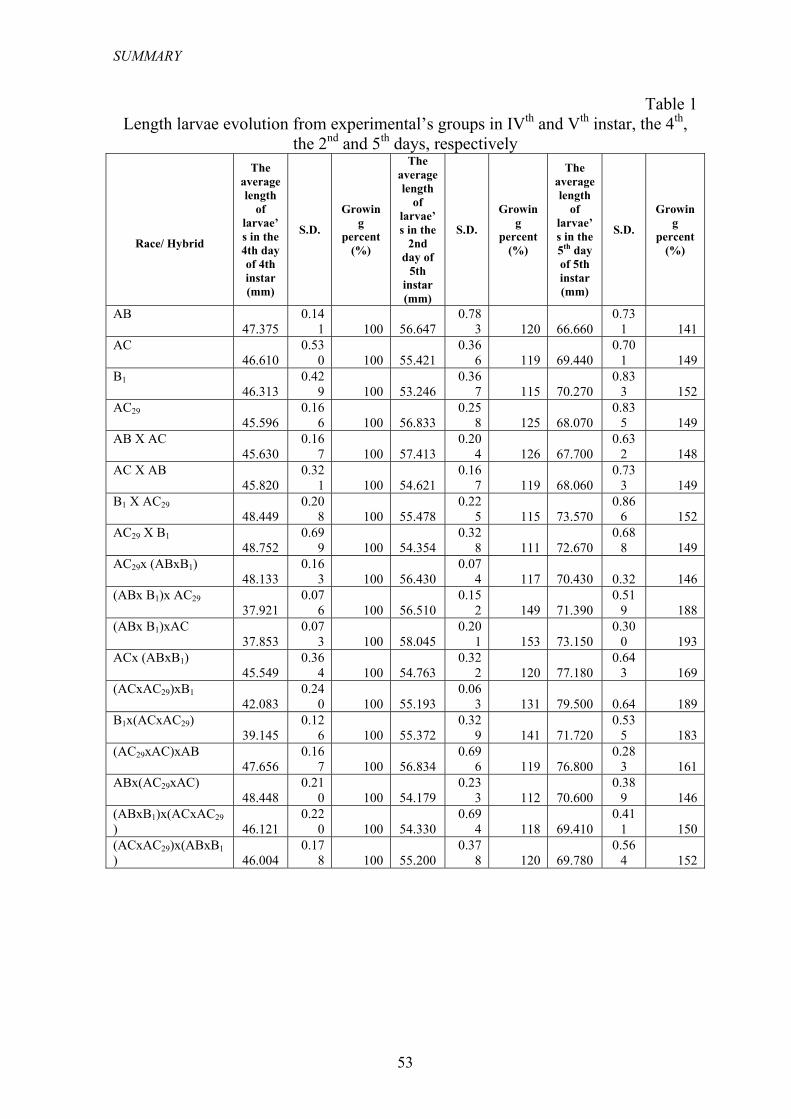
SUMMARY
Table 1 Length larvae evolution from experimental’s groups in IVth and Vth instar, the 4th,
the 2nd and 5th days, respectively
Race/ Hybrid
The average length
of larvae’s in the 4th day of 4th instar (mm)
S.D.
Growing
percent (%)
The average length
of larvae’s in the
2nd day of
5th instar (mm)
S.D.
Growing
percent (%)
The average length
of larvae’s in the 5th day of 5th instar (mm)
S.D.
Growing
percent (%)
AB 47.375
0.141 100 56.647
0.783 120 66.660
0.731 141
AC 46.610
0.530 100 55.421
0.366 119 69.440
0.701 149
B1 46.313
0.429 100 53.246
0.367 115 70.270
0.833 152
AC29 45.596
0.166 100 56.833
0.258 125 68.070
0.835 149
AB X AC 45.630
0.167 100 57.413
0.204 126 67.700
0.632 148
AC X AB 45.820
0.321 100 54.621
0.167 119 68.060
0.733 149
B1 X AC29 48.449
0.208 100 55.478
0.225 115 73.570
0.866 152
AC29 X B1 48.752
0.699 100 54.354
0.328 111 72.670
0.688 149
AC29x (ABxB1) 48.133
0.163 100 56.430
0.074 117 70.430 0.32 146
(ABx B1)x AC29 37.921
0.076 100 56.510
0.152 149 71.390
0.519 188
(ABx B1)xAC 37.853
0.073 100 58.045
0.201 153 73.150
0.300 193
ACx (ABxB1) 45.549
0.364 100 54.763
0.322 120 77.180
0.643 169
(ACxAC29)xB1 42.083
0.240 100 55.193
0.063 131 79.500 0.64 189
B1x(ACxAC29) 39.145
0.126 100 55.372
0.329 141 71.720
0.535 183
(AC29xAC)xAB 47.656
0.167 100 56.834
0.696 119 76.800
0.283 161
ABx(AC29xAC) 48.448
0.210 100 54.179
0.233 112 70.600
0.389 146
(ABxB1)x(ACxAC29) 46.121
0.220 100 54.330
0.694 118 69.410
0.411 150
(ACxAC29)x(ABxB1) 46.004
0.178 100 55.200
0.378 120 69.780
0.564 152
53

SUMMARY
0.66 0.85
0.67 0.79
0.62
0.65 0.74 0.87
0.69 0.79 1.00
2.24
4.73
3.18
2.71
3.68
5.79
2.33 2.
71
2.01
0.80 1.05
0.72
0.60 0.840.97
0.88
2.20
1.86
1.29
2.37
1.85
1.80
0.72
0.760.91
0
2
4
6
8
AB
AC B1
AC
29
ABx
AC
AC
x A
B
B1x
AC
29
AC
29x
B1
AC
29 x
(AB
xB1)
(ABx
B1)
x A
C29
(ABx
B1)
x A
C
AC
x (A
Bx B
1)
(AC
x A
C29
)x B
1
B1x
(AC
x A
C29
)
(AC
29x
AC
)x A
B
ABx
(AC
29x
AC
)
(ABx
B1)
x (A
Cx
AC
29)
(AC
x A
C29
)x (A
Bx B
1)
Rasele si hibrizii viermilor de matase/Silkworm races and hybrids
Gre
utat
e / W
eigh
t (%
)
Masa glandei sericigene/Sericine gland's weight Coeficienul de variabilitate/Variability coeficient
Figure 8. The weight of serigene gland and the variability coefficient of experimental groups
8.3.4. Cocoon yield percent
The larvae once arrived at the maturity, at the end of 5th instar, and after a larval period of 28-32 days (period which can be different function of growing conditions, races, and hybrids) are preparing themselves for cocoons making. In our studies this period was 30 days in the first experimental year and 31 days in the second one. The larvae’s behavior in this period change; they moves on the edge of growing surface, trying to arrive in darker places, preferring the corners; they stopped to feed themselves, and by regurgitation they empty their digestive tube, became transparent, their body became shorter, and remove a thin thread of silk.
In the experiment were used like cocoons materials the wood slats, plastic hedgehog, cardboard cells, oak and pine branches and the obtained results are showed in table 2.
Table 2 The cocoon making percent obtained at parental races on different cocoon making
devices From which obtained on:
Race
Total number of cocoons / Cocoons making
percent (%)
Oak leaves
Pine branches
Plastic hedgehog
Wood slats
Cardboard cells
AB 1280/100 791/61.79 125/9.76 119/9.29 104/8.12 141/11.01 AC 1305/100 785/60.15 145/11.11 130/9.96 124/9.50 121/9.27 B1 1216/100 809/66.5 134/11.01 109/8.96 103/8.47 61/5.01 AC29 1206/100 760/63 120/9.95 117/9.70 109/9.03 100/8.29
Average cocoon making
100 62.86 10.45 9.47 8.78 8.39
54
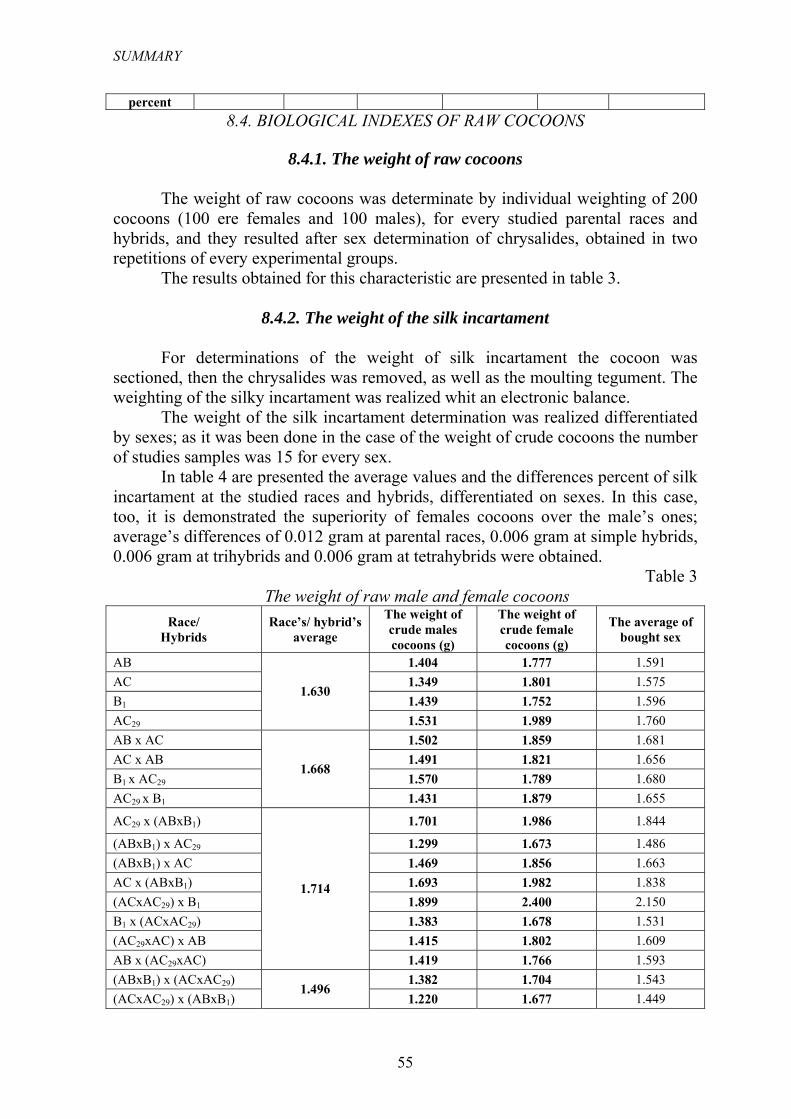
SUMMARY
percent 8.4. BIOLOGICAL INDEXES OF RAW COCOONS
8.4.1. The weight of raw cocoons
The weight of raw cocoons was determinate by individual weighting of 200
cocoons (100 ere females and 100 males), for every studied parental races and hybrids, and they resulted after sex determination of chrysalides, obtained in two repetitions of every experimental groups.
The results obtained for this characteristic are presented in table 3.
8.4.2. The weight of the silk incartament
For determinations of the weight of silk incartament the cocoon was sectioned, then the chrysalides was removed, as well as the moulting tegument. The weighting of the silky incartament was realized whit an electronic balance. The weight of the silk incartament determination was realized differentiated by sexes; as it was been done in the case of the weight of crude cocoons the number of studies samples was 15 for every sex.
In table 4 are presented the average values and the differences percent of silk incartament at the studied races and hybrids, differentiated on sexes. In this case, too, it is demonstrated the superiority of females cocoons over the male’s ones; average’s differences of 0.012 gram at parental races, 0.006 gram at simple hybrids, 0.006 gram at trihybrids and 0.006 gram at tetrahybrids were obtained.
Table 3 The weight of raw male and female cocoons
Race/ Hybrids
Race’s/ hybrid’s average
The weight of crude males cocoons (g)
The weight of crude female cocoons (g)
The average of bought sex
AB 1.404 1.777 1.591 AC 1.349 1.801 1.575 B1 1.439 1.752 1.596 AC29
1.630
1.531 1.989 1.760 AB x AC 1.502 1.859 1.681 AC x AB 1.491 1.821 1.656 B1 x AC29 1.570 1.789 1.680 AC29 x B1
1.668
1.431 1.879 1.655
AC29 x (ABxB1) 1.701 1.986 1.844
(ABxB1) x AC29 1.299 1.673 1.486 (ABxB1) x AC 1.469 1.856 1.663 AC x (ABxB1) 1.693 1.982 1.838 (ACxAC29) x B1 1.899 2.400 2.150 B1 x (ACxAC29) 1.383 1.678 1.531 (AC29xAC) x AB 1.415 1.802 1.609 AB x (AC29xAC)
1.714
1.419 1.766 1.593 (ABxB1) x (ACxAC29) 1.382 1.704 1.543 (ACxAC29) x (ABxB1)
1.496 1.220 1.677 1.449
55
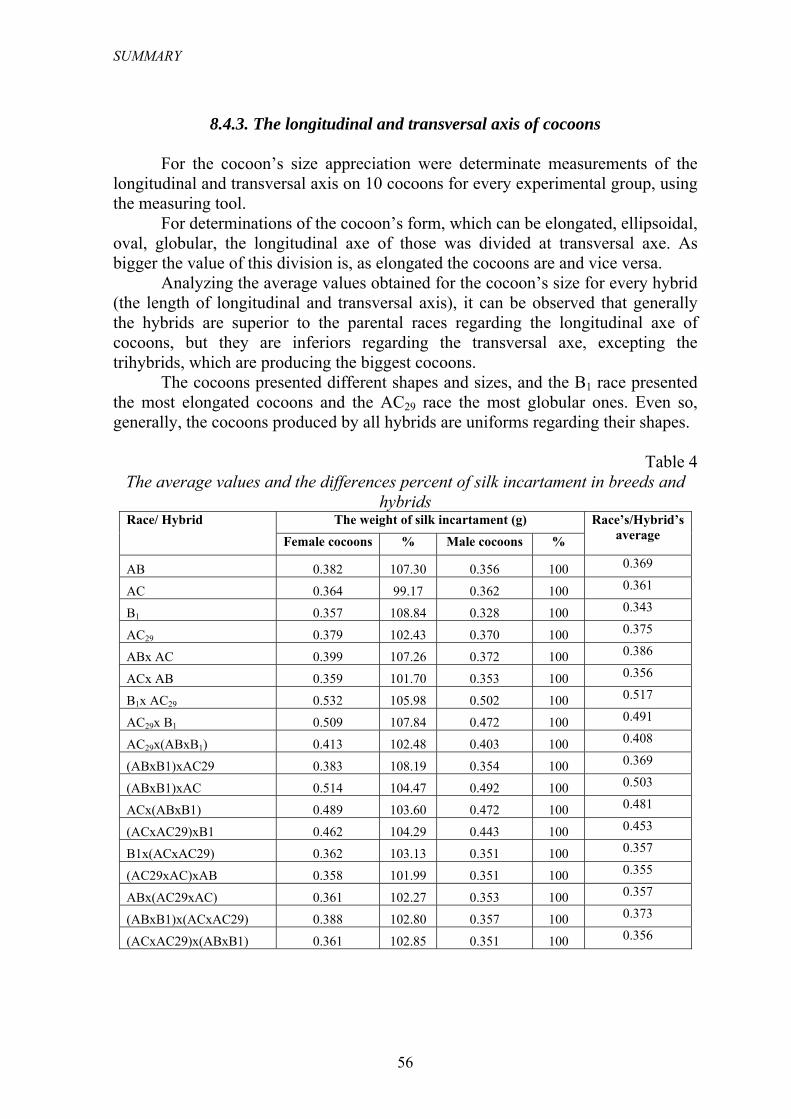
SUMMARY
8.4.3. The longitudinal and transversal axis of cocoons
For the cocoon’s size appreciation were determinate measurements of the
longitudinal and transversal axis on 10 cocoons for every experimental group, using the measuring tool.
For determinations of the cocoon’s form, which can be elongated, ellipsoidal, oval, globular, the longitudinal axe of those was divided at transversal axe. As bigger the value of this division is, as elongated the cocoons are and vice versa.
Analyzing the average values obtained for the cocoon’s size for every hybrid (the length of longitudinal and transversal axis), it can be observed that generally the hybrids are superior to the parental races regarding the longitudinal axe of cocoons, but they are inferiors regarding the transversal axe, excepting the trihybrids, which are producing the biggest cocoons.
The cocoons presented different shapes and sizes, and the B1 race presented the most elongated cocoons and the AC29 race the most globular ones. Even so, generally, the cocoons produced by all hybrids are uniforms regarding their shapes.
Table 4
The average values and the differences percent of silk incartament in breeds and hybrids
The weight of silk incartament (g) Race/ Hybrid
Female cocoons % Male cocoons %
Race’s/Hybrid’s average
AB 0.382 107.30 0.356 100 0.369
AC 0.364 99.17 0.362 100 0.361
B1 0.357 108.84 0.328 100 0.343
AC29 0.379 102.43 0.370 100 0.375
ABx AC 0.399 107.26 0.372 100 0.386
ACx AB 0.359 101.70 0.353 100 0.356
B1x AC29 0.532 105.98 0.502 100 0.517
AC29x B1 0.509 107.84 0.472 100 0.491
AC29x(ABxB1) 0.413 102.48 0.403 100 0.408
(ABxB1)xAC29 0.383 108.19 0.354 100 0.369
(ABxB1)xAC 0.514 104.47 0.492 100 0.503
ACx(ABxB1) 0.489 103.60 0.472 100 0.481
(ACxAC29)xB1 0.462 104.29 0.443 100 0.453
B1x(ACxAC29) 0.362 103.13 0.351 100 0.357
(AC29xAC)xAB 0.358 101.99 0.351 100 0.355
ABx(AC29xAC) 0.361 102.27 0.353 100 0.357
(ABxB1)x(ACxAC29) 0.388 102.80 0.357 100 0.373
(ACxAC29)x(ABxB1) 0.361 102.85 0.351 100 0.356
56
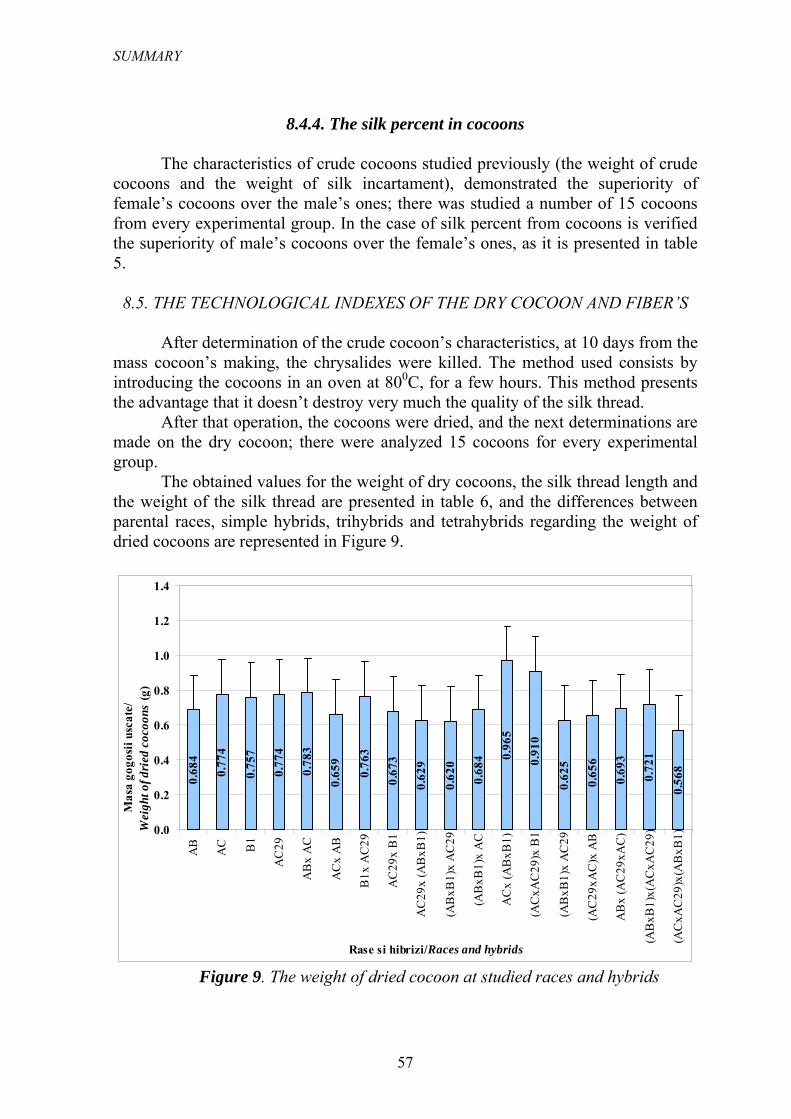
SUMMARY
8.4.4. The silk percent in cocoons
The characteristics of crude cocoons studied previously (the weight of crude
cocoons and the weight of silk incartament), demonstrated the superiority of female’s cocoons over the male’s ones; there was studied a number of 15 cocoons from every experimental group. In the case of silk percent from cocoons is verified the superiority of male’s cocoons over the female’s ones, as it is presented in table 5.
8.5. THE TECHNOLOGICAL INDEXES OF THE DRY COCOON AND FIBER’S
After determination of the crude cocoon’s characteristics, at 10 days from the mass cocoon’s making, the chrysalides were killed. The method used consists by introducing the cocoons in an oven at 800C, for a few hours. This method presents the advantage that it doesn’t destroy very much the quality of the silk thread.
After that operation, the cocoons were dried, and the next determinations are made on the dry cocoon; there were analyzed 15 cocoons for every experimental group. The obtained values for the weight of dry cocoons, the silk thread length and the weight of the silk thread are presented in table 6, and the differences between parental races, simple hybrids, trihybrids and tetrahybrids regarding the weight of dried cocoons are represented in Figure 9.
0.68
4
0.77
4
0.75
7
0.77
4
0.78
3
0.65
9
0.76
3
0.67
3
0.62
9
0.62
0
0.68
4 0.96
5
0.91
0
0.62
5
0.65
6
0.69
3
0.72
1
0.56
8
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
AB
AC B1
AC
29
AB
x A
C
AC
x A
B
B1x
AC
29
AC
29x
B1
AC
29x
(AB
xB1)
(AB
xB1)
x A
C29
(AB
xB1)
x A
C
AC
x (A
BxB
1)
(AC
xAC
29)x
B1
(AB
xB1)
x A
C29
(AC
29xA
C)x
AB
AB
x (A
C29
xAC
)
(AB
xB1)
x(A
CxA
C29
)
(AC
xAC
29)x
(AB
xB1)
Rase si hibrizi/Races and hybrids
Mas
a go
gosi
i usc
ate/
W
eigh
t of d
ried
coc
oons
(g)
Figure 9. The weight of dried cocoon at studied races and hybrids
57

SUMMARY
Table 5
The crude silk content of the male and female cocoons (%) Silk percent (%)
Race/ Hybrid Female’s cocoons
(%) S.D. %
Male’s cocoons
(%) S.D. %
Race’s/ hybrid’s avearge
(%)
S.D.
AB 21.654 0.299 100 24.677 0.341 113.96 23.166 0.320
AC 19.941 0.276 100 25.401 0.351 127.38 22.671 0.314
B1 20.241 0.280 100 23.171 0.320 114.48 21.706 0.300
AC29 19.305 0.267 100 24.123 0.333 124.96 21.714 0.300
Race’s average 22.289 0.311 100 26.257 0.362 117.80 24.273 0.337 ABx AC 20.873 0.288 100 24.353 0.336 116.67 22.613 0.312
ACx AB 19.710 0.272 100 23.664 0.327 120.06 21.687 0.300
B1x AC29 20.488 0.283 100 24.506 0.339 119.61 22.497 0.311
AC29x B1 20.578 0.284 100 23.9 0.330 116.14 22.239 0.307 Simple hybrid’s average 20.412 0.282 100 24.106 0.333 118.09 22.259 0.307
AC29x(ABxB1) 20.755 0.287 100 24.075 0.333 116.00 22.415 0.310
(ABxB1)xAC29 21.847 0.302 100 27.611 0.381 126.38 24.729 0.342
(ABxB1)xAC 22.524 0.311 100 26.259 0.363 116.58 24.392 0.337
ACx(ABxB1) 20.583 0.284 100 25.487 0.352 123.83 23.035 0.318
(ACxAC29)xB1 19.287 0.266 100 24.034 0.332 124.61 21.661 0.299
B1x(ACxAC29) 21.235 0.293 100 25.424 0.351 119.73 23.330 0.322
(AC29xAC)xAB 20.337 0.281 100 24.715 0.341 121.53 22.526 0.311
ABx(AC29xAC) 20.284 0.280 100 24.325 0.336 119.92 22.305 0.308
Trihybrids average 20.857 0.288 100 25.241 0.349 121.02 23.049 0.318 (ABxB1)x (ACxAC29) 19.736 0.273 100 26.138 0.361 132.44 22.937 0.317
(ACxAC29)x (ABxB1) 21.644 0.299 100 28.97 0.400 133.85 25.307 0.350 Tetrahybrids average 20.690 0.282 100 27.554 0.381 133.18 24.122 0.333
58

SUMMARY
Table 6
The technological indices of the dry cocoon and fiber
Race/Hybrids
The weight of
dried cocoons
(g)
S.D. The length of
silk thread (m)
S.D. The weight
of silk thread (g)
S.D.
AB 0.684 0.011 970 26.29 0.243 0.010AC 0.774 0.018 1241 38.16 0.258 0.013B1 0.757 0.017 937 24.11 0.192 0.011AC29 0.774 0.021 963 20.42 0.246 0.013Race’s average 0.747 0.017 1028 27.25 0.235 0.012AB x AC 0.783 0.030 1142 48.87 0.291 0.012AC x AB 0.659 0.015 1071 21.51 0.199 0.017B1 x AC29 0.763 0.021 1226 19.82 0.272 0.012AC29 x B1 0.673 0.025 1142 29.20 0.211 0.012Simple hybrid’s average 0.720 0.023 1145 29.85 0.243 0.013
AC29x (ABxB1) 0.629 0.012 970 24.12 0.271 0.011(ABxB1)x AC29 0.620 0.026 1092 20.38 0.215 0.015(ABxB1)x AC 0.684 0.022 1058 28.20 0.223 0.013ACx (ABxB1) 0.965 0.041 1139 26.76 0.282 0.021(ACxAC29)x B1 0.910 0.033 1039 28.11 0.313 0.015(ABxB1)x AC29 0.625 0.018 942 11.12 0.221 0.012(AC29xAC)x AB 0.656 0.017 1160 29.23 0.183 0.017ABx (AC29xAC) 0.693 0.020 1081 31.28 0.274 0.012Trihybrid’s average 0.723 0.024 1060 24.90 0.248 0.014(ABxB1)x(ACxAC29) 0.721 0.015 1218 27.51 0.210 0.012(ACxAC29)x(ABxB1) 0.568 0.013 889 15.21 0.195 0.012Tetrahybrid’s average 0.645 0.014 1054 21.36 0.203 0.012
CHAPTER IX COMPARATIVE STUDY OF MAIN BIOLOGICAL AND
TECHNOLOGICAL INDEXES IN TWO EXPERIMENTAL YEARS
The experiments which have the specific aim to make possible the hierarchies of silkworm races and hybrids function of the main biological index of larvae were developed in two consecutive years 2007 and 2008, on the same race’s and hybrids, for making possible the compare of theirs characteristics.
There were realized comparative analyses of the results obtained in two experimental’s years for incubation and hatching percent (chapter 9.1), the period of larval stage (chapter 9.2), the biological indexes of crude cocoons (chapter 9.3), as well as the technological index of dried cocoons and silk thread (chapter 9.4).
59

SUMMARY
CHAPTER X THE HIERARCHY OF STUDIED RACES AND HYBRIDS IN
CORRELATION WITH STUDIED PARAMETERS There were analyzed all 14 characteristics, respectively: the average of
hatching percent (%), the weight of larvae’s in the 5th day of the 5th instar (mg), the average of growing percent (%), the larvae’s length (mm), the average of growing percent (%), the weight of serigene gland (g), the crude cocoons weight (g), the weight of silk incartament (mg), the ratio between longitudinal and transversal axis, the silk percent, the weight of the dried cocoons (g), the length of silk thread (m) and the weight of the silk thread (g).
Analyzing the parental races it can be observed that the most valuable race is AC, which registered maximum values for 6 characteristics and it is followed by the parental races B1 and AC29, which registered maximum values for 4 studied characteristics.
Between the simple hybrids it can be remarked the AB x AC one, at who 8 characteristics registered maximum values; this is followed by the hybrid B1 X AC29, whit 6 maximum value.
The trihybrids are less uniform, regarding the hatching percent and the biological index of larvae’s, being more obvious the (AC x AC29) x B1 and (ABx B1) x AC, which registered 3 maximum values at followed characteristics.
The most valuable tetrahybrids after hierarchy was (ABxB1)x(ACxAC29).
CONCLUSIONS AND RECOMMENDATIONS
1. In this research work are presented data regarding the silkworm incubation and egg’s hatching as well as data regarding the main larva’s biological indexes. This study permitted a hierarchy of parental races and hybrids, taking in consideration the followed biological characteristics, in correspondence whit the proposed aim of the research.
2. The microclimate conditions from growing room, the quality of administrated leaves, the sanitation of growing room and applied breeding technology, represents important factors for the good development of experiments; all this parameters were assured at optimal levels for Bombyx mori silkworms.
3. The highest hatching percent (%) was observed in the case of tetrahybrids (98.22%), and a percent of only 93.68% was registered for parental races. Default, this tendency is maintained for the number of hatching eggs.
4. The uniformity of biological material is proved by the period of larval stage; it was 30 days at parental races in 2007 and 31 days in 2008, and the differences obtained in our researches were insignificants comparing whit those obtained by other authors. Other important aspect for demonstrate the uniformity of biological material is the simple hybrid’s insignificantly shorter larval stage (with 1-2 days shorter) comparing whit parental races, and the differences between those were only for a few hours.
60

SUMMARY
5. The larva’s weight (g) presented values very similar at those obtained by other researchers, emphasizing the B1 race, the simple hybrid AB x AC, the trihybrid (AB x B1) x AC and tetrahybrid (AB x B1) x (AC x AC29).
6. Regarding the larvae’s length (mm) emphasizing the race AC, the hybrid B1 x AC29, the trihybrid (AC x AC29) x B1 and the tetrahybrid (AC x AC29) x (AB x B1).
7. For the weight of serigene glands (g) the race AC, the simple hybrid B1 x AC29, the trihybrid (AB x B1) x AC and the tetrahybrid (AB x B1) x (AC x AC29) were superior.
8. The study of the weight of crude cocoons (g) demonstrated the superiority of parental race AC29, of simple hybrid AB x AC, of trihybrid (AC x AC29) x B1, and of the tetrahybrid (AB x B1) x (AC x AC29).
9. The race AC29, the simple hybrid B1 x AC29, the trihybrid (AB x B1) x AC, and its reciprocal hybrid AC x (AB x B1), as well as the tetrahybrid (AB x B1) x (AC x AC29) presented the highest values regarding the weight of silk incartament.
10. The analyze of average values obtained for the cocoon’s size for every hybrid (the length of longitudinal and transversal axis), shows that generally the hybrids are superior to the parental races regarding the longitudinal axe of cocoons but they are inferiors regarding the transversal axe, excepting the trihybrids, which produce the biggest cocoons.
11. The highest silk percent was registered at parental races (26.257%), followed by tetrahybrids (27.554%) and trihybrids, which presents very similar values (25.241%), and on the last position are the simple hybrids whit an average percent of 24.106%.
12. Between the biological indexes of crude cocoons (the weight of crude cocoons, the weight of silk incartament, the ratio between longitudinal and transversal axes, the silk percent) there was not found positive correlation.
13. The studied technological indexes of dried cocoons and of silk thread were the weight of dried cocoons, the length of silk thread, the weight of silk thread and the weight of cocoons waste.
14. The weight of dried cocoons is superior at parental races (0.747 gram), and is followed by the simple hybrids (0.720 gram), which are very similar with the trihybrids (0.723 gram).The tetrahybrids are situated on the last place whit 0.645 gram.
15. The studied parental races present an average length of silk threat of 1028 m, being inferior to the simple hybrids, which presents a value equal at 1145 m; the trihybrids recorded 1060 m and the tetrahybrids 1054 m; these values are similar to those of other Romanian and abroad researchers.
16. Regarding the weight of silk thread outlined the race AC (0.258 g), the simple hybrid (0.291 g), the trihybrid (AC x AC29) x B1, and the tetrahybrid (AB x B1) x (AC x AC29). Generally, the highest average of silk thread’s weight is registered at trihybrids (0.248 grams), followed by simples hybrids (0.243 grams), parental races (0.235 grams) and tetrahybrids (0.203 grams).
17. The weight of cocoon’s waste demonstrated the superiority of tetrahybrids, which present for this character the lowest average (0.031 grams), followed by the trihybrids, monohybrids and parental races. For this character the coefficient
61

SUMMARY
of variability for this character is high, and this fact demonstrates the low level of homogenously of the experimental’s groups.
18. After the final hierarchy of studied parental races, hybrids, trihybrids and tetrahybrids it can be concluded that taking in consideration the followed parameters the tetrahybrid (ABxB1)x(ACxAC29) is superior at all the others hybrids; it registered for 9 of 14 studied characters the maximum value. It is nearly followed by the simple hybrid AB X AC, which registered maximum values for 8 characteristics. For 6 studied characteristics the parental race AC and the hybrid B1 X AC29 presented higher values; for these considerations we can recommend these hybrids and parental races for the growing in Transylvania area.
62

SUMMARY
REFERENCES 1. BARLOW R., 1981, Experimental evidence for interaction between heterosis and environment in
animals, Animal Breeding Abst. 49, p.715- 739. 2. BENŢEA, M., 2006, Teză de Doctorat. USAMV Cluj-Napoca. 3. BURA M., 1992, Curs de tehnologia producţiei sericicole. Universitatea de Ştiinţe Agricole a
Banatului Timişoara. 4. DAS S.K., PATTNAIK S., GHOSH B., SINGH T., NAIR B.P., SEN S.K. AND SUBBA RAO G.,
1994, Heterosis analysis in some three way crosses of Bombyx mori L., Sericologia, 34 (1) p. 51-61. 5. EHIOBU N. G., GODDARD M. E., 1989, Heterosis in cross between lines of Drosophila
melanogaster selected for adaption to different environments. Theo. Appl. Genet., 77, p.253- 259. 6. GRIFFING B., ZSIROS E., 1971, Heterosis associated with genotype – environment interactions,
Genetics, 68, p. 443- 445. 7. HINAJAWA C., OHTSUKA M., 1975, Heterosis value in three- way and tetra- way crosses of
Bombyx mori L. hybrids, Theo. Appl. Genet., 43, p. 275- 278. 8. HIROBE T., 1985, On the recent advancement of silkworm breeding in Japan. Tamogawa Univ.
Japan, p. 157- 161. 9. IFRIM S., 1985, Ce ştim despre mǎtase, Editura Tehnicǎ Bucureşti,. 10. KNIGHT R., 1973, The relation between hybrid vigour and genotype and environment interaction.
Theo. Appl. Genet., 43, p.311- 318. 11. LERNER I. M., 1954, Genetic homeostasis, John Wiley and Sons Inc., New York. 12. MĂRGHITAŞ LIVIU AL., 1995, Creşterea viermilor de mătase, Ed. Ceres, Bucureşti. 13. MǍRGHITAŞ LIVIU AL., DEZMIREAN D., PAŞCA I., 2003, Sericicultura, Ed. Mediamira, Cluj-
Napoca. 14. MORAR R., DANA PUSTA, 1999, Zootehnie specială, III, Ed. Relief, p. 88. 15. MOROHOSHI S., 1949, Developmental mechanism in Bombyx mori. Meibundo, Tokyo. 16. NAGATOMO T., 1942 On the inheritance of the voltinism in the silkworm, J. Sericult. Sci. Japan,
Vol. 13, p. 114- 115. 17. NAKADA T., 1970, Researches on the sex- linked inheritance of the cocoon weight of reciprocal
crossings, J. Fac. Agric., Hokkaido University, 56, p. 348- 358. 18. OROZCO, 1976, Heterosis and genotype environment interaction. Theoretical and experimental
aspects. Bulletin technique, Departament de Genetique Animale, INRA, 24, p. 43- 52. 19. PAŞCA I., 2004, Teză de Doctorat. USAMV Cluj-Napoca. 20. PETRE, A., A. VLAIC, MĂRIOARA POP, 1989, Lucrări practice de Genetică animală, Tipo
Agronomia, Cluj-Napoca. 21. RAJASEKHAR, R., 1993, Studies on the temperature induced modulations on the growth and
reproductive physiology of silkworm, Bombyx mori L, PhD Thesis, S. V. University Tirupathi, India.
22. REDDY P. L., S.S. NAIK, P. M. MOHAN, 1998, Hatching pattern of silkworm, Bombyx mori L., as influenced by light intensity, Indian Journal of Sericiculture, 37 (2), p. 116- 122.
23. RUBAN P. S., CUNNINGHAM E. P., SHARP P. M., 1988, Heterosis and nutrition interaction in Drosophila. Theo. Appl. Genet., 76, p.136- 142.
24. SANG J. H., 1964, Nutritional requirements of inbred lines and crosses of Drosophila melanogaster. Chart. Res., 5, p. 50- 67.
25. SANTIAGO E., DIMINGUEZ A., ALBORNOZ J. PINEIRO R., IZOUIERDO J. I., 1989, Environmental sensivity and heterosis for egg laying in Drosophila melanogaster. Theor. Appl. Genet., 78, p. 243- 248.
26. SAVEL I., 1998, Mǎtasea naturalǎ, Ed. Ceres, Bucureşti. 27. SINGH R., SUDHAKOVA R. P., DATTA R.K., 1998, Studies on hybrid vigor in different crosses
of the silkworm Bombyx mori L., Sericologia, 38 (1), p. 155- 158. 28. SOHN K.W., 1983, Studies on variability in single, three- way and double crosses of silkworm,
Bombyx mori. Dissertation submitted to CSRTI, Mysore, India. 29. TACU, A., 1968, Metode statistice în zootehnie şi medicină veterinară, Ed. Agrosilvică, Bucureşti. 30. TAZIMA T., 1988, A view on the improvement of Mysore breeds. Proc. Intern. Cong. Trop.
Sericult. Bangalore, IV, 1- 5. 31. TSURUMAKI J., J. ISHIGURO, A. YAMANAKA, K. ENDO, 1999, Effects of photoperiod and
temperature on seasonal morph development and diapause egg oviposition in bivoltine race (Daizo) of the silkmoth, Bombyx mori L., Journal of Insect Physiology, 45, p. 101 – 106.
32. VLAIC, B., 2007, Teză de doctorat, USAMV Cluj-Napoca.
63

SUMMARY
64
33. WATANABE H., 1961, Studies on difference in the variability of larval body and cocoon weights between single cross and three- way cross or double cross hybrids in the silkworm, Bombyx mori., J. Sericult. Sci. Japan, 30, p. 463 – 467.
34. YOKOYAMA T., 1973, The history of sericulture science in relation to industry. In History of Entomology. R. F. Smith, T. E. Mittler and C. N. Smith (Eds.), Ann. Rev. Palo Alto, California, p. 267- 284.
















