
INVESTIGAREA MOLECULARĂ A INFLUENȚEI · procesele biologice ale organismului. În majoritatea...
42
UNIVERSITATEA „ALEXANDRU IOAN CUZA” din IAŞI ŞCOALA DOCTORALĂ A FACULTĂŢII DE FIZICĂ Laboratorul de Biofizică Moleculară şi Fizică Medicală INVESTIGAREA MOLECULARĂ A INFLUENȚEI STRUCTURII UNOR PEPTIDE ASUPRA INTERACȚIUNII CU SISTEMELE LIPO-PROTEICE Rezumatul tezei de doctorat Conducător ştiinţific, Prof. Dr. Tudor LUCHIAN Doctorand, Irina ŞCHIOPU - 2014, IAŞI -
Transcript of INVESTIGAREA MOLECULARĂ A INFLUENȚEI · procesele biologice ale organismului. În majoritatea...

UNIVERSITATEA „ALEXANDRU IOAN CUZA” din IAŞI
ŞCOALA DOCTORALĂ A FACULTĂŢII DE FIZICĂ
Laboratorul de Biofizică Moleculară şi Fizică Medicală
INVESTIGAREA MOLECULARĂ A INFLUENȚEI
STRUCTURII UNOR PEPTIDE ASUPRA
INTERACȚIUNII CU SISTEMELE LIPO-PROTEICE
Rezumatul tezei de doctorat
Conducător ştiinţific, Prof. Dr. Tudor LUCHIAN Doctorand,
Irina ŞCHIOPU
- 2014, IAŞI -

UNIVERSITATEA "ALEXANDRU IOAN CUZA" DIN IAŞI Şcoala Doctorală de Fizică
Vă facem cunoscut faptul că în data de 1 septembrie 2014, ora 11:00, în
sala de conferinţe „FERDINAND”, domnişoara Irina ŞCHIOPU, va susţine, în
şedinţă publică, teza de doctorat:
„Investigarea moleculară a influenţei structurii unor peptide asupra interacţiunii
cu sistemele lipo-proteice”
în vederea obţinerii titlului ştiinţific de doctor în domeniul fundamental Ştiinţe
Exacte, domeniul Fizică.
Comisia de examinare a tezei: Prof. Dr. Diana MARDARE Preşedinte
Directorul Şcolii Doctorale de Fizică Universitatea “Alexandru Ioan Cuza”, Iaşi Prof. Dr. Tudor LUCHIAN Coordonator ştiinţific
Facultatea de Fizică Universitatea “Alexandru Ioan Cuza”, Iaşi
Prof. Dr. Nicoleta DUMITRAŞCU Referent Facultatea de Fizică Universitatea “Alexandru Ioan Cuza”, Iaşi
C.S.I. Dr. Ioan TURCU Referent INCDTIM, Cluj – Napoca Conf. Dr. Dragoş GRIGORIU Referent
Universitatea de Medicină şi Farmacie „Gr. T. Popa”, Iaşi
Vă invităm pe această cale să participaţi la şedinţa publică de susţinere a tezei.

☼ Mulţumesc coordonatorului ştiinţific, profesor dr. Tudor Luchian şi colegelor mele dr. Aurelia Apetrei, dr. Loredana Mereuţă, dr. Alina Asandei pentru colaborare,
incredere şi sfaturi utile oferite pe parcursul perioadei de doctorat.
☼
Mulţumesc doamnelor profesoare dr. Dorina Creangă şi dr. Nicoleta Dumitraşcu şi domnişoarei lector dr. Loredana Mereuţă pentru sfaturile constructive
acordate în calitate de membri ai comisiei de îndrumare.
☼
Mulţumesc doamnei profesoare dr. Nicoleta Dumitraşcu, domnului cercetător ştiinţific dr. Ioan Turcu şi domnului conferenţiar dr. Dragoş Grigoriu
pentru sprijinul acordat în calitate de refereţi ai acestei teze.

CUPRINS
I. Motivaţia studiului: Arhitectura inteligentă a peptidelor – cheia obţinerii unor molecule cu proprietăţi terapeutice superioare ............................................ 5
II. Peptidele antimicrobiene şi interacţiunea lor cu membranele lipidice artificiale ...................................................................................................... 9
II.1. Caracteristici generale ale peptidelor antimicrobiene ................................................... 9
II.2. Mecanisme de destabilizare a membranelor lipidice artificiale. Tipuri de pori transmembranari ................................................................................................................. 15
II.3. Rolul aminoacidului Triptofan în interacţiunea peptidă – membrană lipidică ............ 19
III. Detalii structurale şi funcţionale ale membranelor lipidice artificiale. Proprietăţi electrice ale membranelor lipidice şi importanţa acestora în modularea activităţii peptidelor ............................................................................ 24
IV. Detalii structurale şi funcţionale ale peptidelor antimicrobiene utilizate în cadrul studiilor experimentale .............................................................................. 32
IV.1 Caracteristici specifice ale peptidei antimicrobiene melitina ..................................... 32
IV.2 Caracteristici specifice analogilor sintetici ai peptidei HPA3NT0:
Pep1, Pep2, Pep3 ....................................................................................................... 35
V. Interacţiuni manifestate la nivel de singură moleculă ale proteinelor, peptidelor şi moleculelor mici cu nanoporii proteici ........................................... 38
V.1. Descrierea detaliată a interacţiunilor manifestate la nivelul porului proteic ............... 38
V.2. Detalii structurale şi funcţionale ale porului proteic de α-hemolizină ........................ 40
VI. Detalii structurale şi funcţionale ale peptidelor utilizate în cadrul studiilor experimentale la nivel de singură moleculă ......................................................... 42
VI.1 Caracteristici specifice peptidei chimerice CA(1-8)MA(1-12)................................... 42
VI.2 Caracteristici specifice peptidei amiloidice uman-Aβ(1-16), de provenienţă umană şi murin-Aβ(1-16), de provenienţă murină ......................................................................... 46
VI.3 Efectul metalelor de tranziţie asupra conformaţiei peptidelor amiloidice .................. 52
VI.4 Natura chimică a interacţiunii aminoacidului histidină cu ionii de cupru ................... 56

VII. Tehnici şi metode aplicare în studiul influenţei structurii unor peptide asupra modului de interacţiune cu sistemele lipo-proteice ................................. 61
VII.1. Metoda „Montal-Muller” de realizare a membranelor lipidice artificiale ................ 61
VII.2. Înregistrări de electrofiziologie la nivel de singură moleculă. Utilizarea proteinei α-hemolizină ca nanosenzor ............................................................................................... 65
VII.3. Prepararea lipozomilor unilamelari mici prin metoda deshidratării ......................... 67
VII.4. Metode spectrofluorometrice de analiză şi control bazate pe emisia aminoacizilor aromatici ............................................................................................................................. 72
VII.5. Tehnica monitorizării în timp real a efluxului de calceină încapsulat în interiorul lipozomilor unilamelari mici sub acţiunea peptidelor......................................................... 76
VII.6. Tehnica stingerii emisiei de fluorescenţă a floroforilor din structura peptidelor ...... 79
VIII. REZULTATE EXPERIMENTALE: .......................................................... 84
VIII.1. Studiul interacţiunilor dintre peptida antimicrobiană melitin şi sisteme lipidice biomimetice .............................................................................. 84
VIII.1.1. Modularea activităţii peptidei antimicrobiene melitina de către agentul modificator al potenţialului de dipol phloretin .............................................................. 84
VIII.1.2. Influenţa tăriei ionice asupra adsorbţiei şi formării de pori transmembranari de către peptida antimicrobiană melitin ............................................................................. 91
VIII.2. Modularea activităţii unor peptide scurte de către modificarea poziţiei şi orientării spaţiale a aminoacidului triptofan .................................. 94
VIII.2.1. Studiul activităţii analogilor sintetici ai peptidei HPA3NT0: Pep1, Pep2, Pep3 asupra membranelor lipidice prin tehnici de electrofiziologie .............................. 94
VIII.2.2.Studiul adsorbţiei peptidelor Pep1, Pep2, Pep3 în sisteme de lipozomi prin tehnici de spectroscopie de fluorescenţă ..................................................................... 102
VIII.2.3.Studiul cineticii de inserţie şi formare a porilor transmembranari de către Pep1, Pep2, Pep3 prin monitorizarea spectrofluorimetrică a efluxului de calceină din lipozomi .............................................................................. 109

VIII.3. Monitorizarea schimbărilor conformaţionale a unor peptide în prezenţa unor metale relevante fiziologic ...................................................... 114
VIII.3.1. Studiul deplasării spectrului de emisie a peptidei CA(1-8)MA(1-12) în interacţiunea cu ionii de cupru. Identificarea constantei de disociere Kd cu ajutorul modelul matematic Langmuir ........................................................................ 114
VIII.3.2. Modelul matematic propus pentru caracterizarea cineticii reacţilor de asociere şi disociere dintre porul proteic de α – hemolizină şi peptida chimerică CA(1-8)MA(1-12), complexată şi necomplexată de ionii de cupru ............................ 120
VIII.4. Studiu comparativ asupra peptidelor amilodice de provenienţă umană uman-Aβ(1-16) şi murină, murin-Aβ(1-16) în interacţiunea cu ionii de cupru ............................................................................................... 136
VIII.4.1. Utilizarea porului proteic de α – hemolizină ca detector molecular a diferenţelor conformaţionale ale peptidelor amiloidice Ab(1-16)h şi Ab(1-16)r induse de ionii de cupru .............................................................................................. 136
VIII.5. Investigarea efectelor metalelor relevante fiziologic asupra conformaţiei spaţiale a peptidei amiloidice de provenienţă umană Aβ(1-16) ........................................................................................................................... 151
VIII.5.1. Studiul la nivel de singură moleculă a cineticii reacţiei α-HL – Ab(1-16)h în prezenţa ionilor de Cu2+, Zn2+, Fe3+ şi Al3+. Determinarea constantelor de disociere Kd ................................................................................................................ 151
Bibliografie ............................................................................................................ 152

I. Motivaţia studiului: Arhitectura inteligentă a peptidelor – cheia obţinerii unor molecule cu proprietăţi terapeutice superioare
Motivaţia studiului peptidelor antimicrobiene Peptidele antimicrobiene reprezintă o parte esenţială a sistemului imunitar care
acţionează împotriva infecţiilor microbiene. Acestea se regăsesc atât la plante, insecte cât şi la vertebratele superioare. Acestea peptide naturale sunt compuşi bazici, cu o structură primară de 12-50 de aminoacizi, în bazele de date existente până la ora actuală fiind inregistrate peste 800 de asemenea compuşi. Câte va exemple de astfel de peptide antimicrobiene împreuna cu sursa lor, ar fi: insect defensin A (fungi), cecropin A (insecte), melittin (venin de albine), magainin 2 (amfibieni), tachyplesin 1 (crabi), α-defensin HNP-1 (oameni).
Compoziţia lor în aminoacizi, amfipaticitatea, încărcarea electrică cationică şi structurile lor sescundare, permit acestor peptide să se adoarbă şi să se insere în membranele lipidice pentru a forma pori transmembranari [1,2]. Deşi au existat numeroase încercări de a elucida mecanismul lor de acţiune prin diferite modele de tipul porilor transmembranari clasici, a porilor toroidali sau de distrugere a membranelor [3], există încă dezbateri privind modalitatea sau modalităţile de acţiune asupra microorganismelor in vivo.
În acest studiu ne-am propus investigarea influenţei poziţiei şi orientării geometrice a aminoacizilor aromatici prezenţi în structura primară a unor peptide antimicrobiene derivate din peptida HPA3NT0 asupra mecanismului acestora de formare a porilor transmembranari în bistraturi lipidice reconstituite. Rezultatele experimentale şi interpretarea acestora vor oferi o mai bună înţelegere a mecanismelor de acţiune litică a acestor peptide in vivo, esenţială pentru asamblarea unor noi molecule derivate din peptide antimicrobiene naturale, dar cu proprietăţi superioare acestora.
Motivaţia studiului interacţiunilor dintre peptide şi metale de tranziţie În ultimii ani interesul faţă de etiologia bolilor neurodegenerative a crescut
considerabil, iar una din ipotezele propuse de studiile efectuate până în prezent în această direcţie indică tulburarea homeostaziei metalelor la nivelul creierului ca şi cauză principală pentru inducerea unei conformaţii anormale [4] a peptidelor şi proteinelor implicate în procesele biologice ale organismului. În majoritatea cazurilor, împachetarea greşită a peptidelor şi proteinelor induce la oligomerizarea, agregarea acestora şi în final la formarea de plăgi insolubile [4]. O gamă variată de boli neurodegerantive sunt afectate de procesele de agregare a peptidelor şi proteinelor, cu toate că încă nu se cunoaşte dacă formarea de fibrile este cauza apariţiei bolilor sau doar o consecinţă.
În acest studiu ne-am propus ca în acest studiu să dezvoltăm o serie de experimente în care să urmărim pe de-o parte schimbarea conformaţiei peptidelor amiloidice de proveniență umană și murină în urma procesului de complexare cu ionii de cupru, prin tehnici de electrofiziologie la nivel de singură moleculă, şi pe de altă parte să demonstrăm importanţa structurii primare a peptidelor în împachetarea greşită a acestora prin prisma diferențelor existente între cele două tipuri de peptide amiloidice. În acest scop ne-am folosit de porul
1

proteic de α-HL pentru a testa modul de interacţiune a peptidelor amiloidice cu acest nanopor, în prezenţa şi în absenţa ionilor de cupru, dezvoltând și un model matematic complex ce ne permite analiza cantitativă a acesor interacțiuni și determinarea unor parametri esențiali (i.e. constanta de disociere) în descrierea efectelor ionilor de cupru asupra conformației peptidelor amiloidice. Relevanţa aplicării acestei tehnici de studiu al interacţiunilor la nivel uni-molecular constă în uşurinţa şi eleganţa implementării acesteia cu succes în analiza la rezoluţie subnanometrică a moleculelor de ADN [5] şi în diverse studii de detecţie moleculară. Importanţa implementării acestei tehnici, descrisă pe larg în această lucrare, o prezintă posibilitatea de a descrie mecanismele moleculare ce au loc între o singură peptidă în interacţiune cu ionii de metal.
II. Peptidele antimicrobiene şi interacţiunea lor cu membranele lipidice artificiale
Numeroase studii au descris influenţa unor parametri structurali importanţi ai peptidelor: sarcina electrică netă, helicitatea, hidrofobicitatea intrinsecă, momentul hidrofob, şi mărimea domeniilor polare respectiv hidrofobe asupra efectului de permeabilizare a membranelor, a efectului antimicrobial şi hemolitic. Experimentele arată că aceşti parametri pot constitui o bază puternică în optimizarea structurii peptidelor, corelată cu activitatea lor antimicrobială. Predominanţa unui mecanism de permeabilizare a membranei este determinată de efectul cumulat al caracteristicilor structurale ale peptidei şi de influenţa acestora asupra etapelor de adsorbţie şi permeabilizare a membranelor. Principalele caracteristici care determină activitatea şi specificitatea peptidelor antimicrobiene sunt: secvenţa primară, structura secundară, helicitatea, momentul hidrofob, hidrofobicitatea, unghiul de hidrofobicitate şi sarcina electrică a peptidei.
Până în prezent au fost descrise două tipuri distincte de canale ionice formate prin agregarea peptidelor citotoxice: porii clasici („barrel stave”) al căror interior este format numai din părţile polare ale peptidelor şi porii toroidali la a căror structură interioară contribuie şi capetele polare ale lipidelor ce intră în componenţa membranei (Fig. II.2.1).
Un alt mod de interacţiune cu bistratul lipidic ar fi reprezentat de mecanismul de carpetare, conform căruia peptidele se acumulează la interfaţa bistrat/mediu apos sub forma unui „covor”şi, când se depăşeşte o valoare prag a concentraţiei de monomeri, membrana este
Fig.II.2.1. Reprezentarea schematizată a modelelor de formare a porilor transmembranari. (a) Modelul porilor clasici; (b) Modelul porilor toroidali [10]
2

permeabilizată şi dezintegrată într-un mod asemănător acţiunii detergenţilor, fără formarea unor canale ionice distincte (Fig. II.2.2).
Fig. II.2.2. Reprezentare schematică a modului de disrupere a membranei celulare prin mecanismul de carpetare. Peptidele cu structură de α-helix se adsorb iniţial la interfaţa membrană/mediu apos (a) apoi se acumulează (b) orientate paralel la suprafaţa membranei. Acumularea continuă de peptide la nivelul bistratului conduce la „acoperirea (carpetarea)” acesteia cu peptide (c) şi în final, când se atinge o concentraţie critică, la dezintegrarea ei (d) [11].
Învestigaţii de înaltă rezoluţie a structurii proteinelor inserate în membranele lipidice şi studii recente ale rolului aminoacizilor individuali în medierea interacţiunilor proteine – lipide au arătat că reziduurile polare – aromatice, triptofan (Trp) şi tirozină (Tyr), prezintă o afinitate specifică pentru regiunea lipidică aflată la interfaţa membrană – mediu apos. Interfaţa membrană – mediu apos reprezintă o parte relativ mare din grosimea totală a bistratului lipidic şi prezintă un mediu chimic complex, care oferă multe posibilităţi pentru interacţiuni necovalente cu catenele laterale ale peptidelor şi proteinelor. Această interfaţă prezintă un gradient de polaritate, de la foarte apolar în apropierea regiunii cozilor hidrocarbonate ce formează miezul hidrofob al membranei, până la foarte polar, în apropierea fazei apoase. Grupările carboxil ale lipidelor, grupare fosfatcolină, cât şi moleculele de apă din jurul capetelor polare ale fosfolipidelor oferă posibilitatea unor interacţiuni de tip dipol – dipol, cation – π, formarea de legături de hidrogen cu lanţurile laterale ale aminoacizilor din structurile peptidelor şi proteinelor [12,13].
Fig.II.3.1 Interacţiunea cation-π dintre un cation oarecare şi un inel aromatic de benzen [adaptare după UC Davis ChemWiki].
Interacţiunea cation – π reprezintă o interacţiune puternică de natură necovalentă dintre electronii orbitalilor π (în care densitatea electronică este distribuită în mod uniform deasupra şi dedesubtul planului moleculei) şi un cation aflat în vecinătate. În cazul interacţiunilor de tip cation – π manifestate la interfaţa dintre bistratul lipidic şi mediul apos, legătura se formează între inelul indol al aminoacidului triptofan şi azotul încărcate electric pozitiv din gruparea colină a capetelor polare a fosfolipidelor ce compun bistratul lipidic. Legăturile de hidrogen se manifestă între hidrogenul din gruparea NH a inelului indol a triptofanului şi oxigenul din gruparea carboxil (C=O). Tăria acestor legături depinde de tipul de fosfolipide din componenţa membranei, calcule energetice din dinamica moleculară au
3

arătat că preponderenţa interacţiunilor cation – π creşte în cazul lipidelor în care cationul azot este mai expus (e.g.: POPE, 1-palmitoyl-2-oleoylphosphatidylethanolamine, unde azotul este legat doar de trei atomi de hidrogen, faţă de POPC, 1-palmitoyl-2-oleoylphosphatidyl-choline, unde azotul este legat de trei grupări CH) [14].
III. Detalii structurale şi funcţionale ale membranelor lipidice artificiale. Proprietăţi electrice ale membranelor lipidice şi importanţa acestora în modularea activităţii peptidelor.
Membranele biologice sunt structuri polimerice alcătuite în principal din lipide şi proteine, cu rol esenţial în menţinerea integrităţii celulare prin aceea că reprezintă o barieră de permeaţie foarte selectivă între mediile intra şi extracelular. Acestea servesc drept ‘suport’ pentru o clasă vastă de proteine membranare ce sunt practic solubilizate în bistratul lipidic şi care au rol esenţial în procesele de transducţie celulară, comunicare celulară, replicarea ADN-ului, în procesele de recunoaştere celulară, etc. Deşi caracteristica tuturor celulelor ce intră în compoziţia materiei vii este diversitatea funcţională şi structurală, toate biomembranele sunt structuri planare, cu grosimi cuprinse între 60Å şi 100Å, alcătuite în principal din lipide şi proteine. Mai mult, biomembranele sunt suprastructuri moleculare necovalente, organizate sub formă de bistrat lipidic al căror monostraturi prezintă asimetrie.
În general, se poate spune că profilul electric al biomembranelor se compune din: contribuţia potenţialului transmembranar, potenţialul de dipol şi diferenţa potenţialelor de suprafaţă dintre cele două feţe ale membranei. În timp ce potenţialul transmembranar rezultă în urma gradientului de sarcină de o parte şi de alta a membranei şi potenţialul de suprafaţă este dat de excesul de sarcină netă prezentă la suprafaţa membranei, potenţialul de dipol al membranei îşi are originea în dipolii moleculari localizaţi la nivelul moleculelor lipidelor.
Potenţialul de dipol ( dψ ) (Fig.III.4) reprezintă cea de-a treia componentă a
potenţialului transmembranar total. Studii structurale ale biomembranelor au relevat faptul că originea potenţialului de dipol este dată de doi factori principali şi anume: pe de o parte orientarea grupărilor dipolare localizate în moleculele lipidice (dipolul corespunzător grupării
carbonil din legătura esterică şi dipolul capătului terminal - +P - N ), pe de altă parte momentele dipolare corespunzătoare moleculelor de apă situate la interfaţa membrană/soluţie apoasă.
4

Fig.III.4. Reprezentare schematizată a unui bistrat lipidic (a) alcătuit din fosfolipide a căror structură prezintă o regiune polară (colină-fosfat-glicerol) şi două cozi hidrocarbonate hidrofobe (acizi graşi) (bReprezentare schematică a potenţialului de dipol
dψ , corelat cu existanţa momentelor dipolare diferite de zero existante în capetele hidrofile ale lipidelor ce intră în compoziţia membranelor biologice ( P Pδ δ− +− ), ( C Oδ δ+ −− ) şi a momentelor dipolare ale moleculelor de apă.
În literatura actuală, sunt cunoscute anumite molecule care pot modula amplitudinea potenţialului de dipol membranar atunci când ajung la suprafaţa membranară. Dintre acestea, phloretinul are abilitatea de a scădea amplitudinea potenţialului de dipol. Phloretinul este un acid slab, are valoarea pKa = 7.3 iar forma activă a acestuia este cea neutră, la un pH < 7.3.
Fig.III.6 Schemă reprezentativă pentru absorbţia moleculelor de phloretin la interfaţa membrană – mediu apos. Structura chimică a moleculei de phloretin, precum şi momentul de dipol corespunzător acestuia dat de gruparea carboxil (C=O) este reprezentată în stânga, iar bistratul lipidic este reprezentat în dreapta împreună cu potenţialul de dipol dat de capetele polare. Direcţia pe care moleculele de phloretin se adsorb este cu momentul de dipol orientat invers faţă de cel al capetelor polare, astfel ducând la o scădere a potenţialului de dipol al membranei.
IV. Detalii structurale şi funcţionale ale peptidelor antimicrobiene utilizate în cadrul studiilor experimentale
Melitină (Apis Mellifera): H2N-Gly-Ile-Gly-Ala-Val-Leu-Lys-Val-Leu-Thr-Thr-Gly-Leu-Pro-Ala-Leu-Ile-Ser-Trp-Ile-Lys-Arg-Lys-Arg-Gln-Gln-CONH2
5

analogi HPA3NT0 (Helicobacter pylori): Phe-Lys-Arg-Leu-Glu-Lys-Leu-Phe-Ser-Lys-Ile-Trp-Asn-Trp-Lys
Pep1: Leu-Lys-Arg-Leu-Gln-Lys-Leu-Leu-Ser-Lys-Ile-Trp-Asn-Lys-Trp - NH2 Pep2: Phe-Lys-Arg-Trp-Gln-Lys-Leu-Leu-Ser-Lys-Ile-Trp-Trp-Lys-Asn - NH2 Pep3: Leu-Lys-Arg-Leu-Gln-Lys-Leu-Leu-Ser-Lys-Ile-Trp-Trp-Lys-Asn - NH2
V. Interacţiuni manifestate la nivel de singură moleculă ale proteinelor, peptidelor şi moleculelor mici cu nanoporii proteici
Procesul de translocare prin proteine, de dimensiuni nanometrice, inserate în membranele celulelor eucariote este semnificativ pentru menţinerea funcţiilor acestora. În ultimii ani, interesul pentru astfel de procese a crescut, sesizându-se potenţialul unor proteine de a fi utilizate în domeniul biotehnologiilor, ca şi sisteme de detecţie bimoleculară. Astfel, această tehnică a fost folosită pentru caracterizarea unor suspensii de particule [15-17], secvenţierea ADN-ului [18,19] sau detecţia schimbărilor conformaţionale ale peptidelor [20, 21].
Pe scurt, atunci când o moleculă translocă printr-un por proteic de dimensiuni nanometrice, inserat într-o membrană lipidică ce separă două compartimente cu conţinut de soluţie electrolitică, aceasta va bloca accesul ionilor să mai parcurgă traseul din interiorul porului, ducând la o scădere a curentului ionic iniţial, corespunzător porului liber. Profilul şi succesiunea de fluctuaţii de curent ionic generate de trecerea diverselor tipuri de molecule printr-un singur por proteic pot oferi informaţii utile cu privire la caracteristicile structurale ale acestora.
Fig.V.1.1. Reprezentarea unui singur por proteic de α-HL inserat în membrana lipidică. Peptida amiloid-beta prezentată în prezenţa şi în absenţa β-ciclodextrinei (Aβ 42-CD) sau a moleculei de congo red. (a) Reprezentarea a blocajelor de curent ionic datorate inteacţiunii dintre por şi complexul Aβ 42-CD. (b) Reprezentarea a blocajelor de curent ionic datorate inteacţiunii dintre por şi complexul Aβ 42. (c) Reprezentarea a blocajelor de curent ionic datorate inteacţiunii dintre por şi complexul Aβ42-congo red. [22]
α-hemolizina (α-HL) este o proteină monomerică de 33.2 kDa solubilă în apă, secretată de bacteria Staphylococcus aureus, care prezintă proprietăţi de autoasamblare, formând canale ionice heptamerice în membrane lipidice sau artificiale (Fig.V.2.1).
6

Fig.V.2.1 (a) Vedere laterală a complexului heptameric de α-hemolizină care arată modul de inserţie în bistratul lipidic membranar. Complexul are o lungime şi un diametru comparabile de aproximativ 100 Å. (b) Vedere a α-hemolizinei din partea cis a membranei lipidice. (c) şi (d) Secţiunea longitudinală respectiv modelul ‘space-filling’ al proteinei care arată diametrul interior al canalului acesteia la diferite poziţii. Fiecare culoare reprezintă un monomer diferit de α-hemolizină [23].
Structura cristalină a acestei proteine este bine cunoscută [24]. Datorită posibilităţii de a introduce numeroase mutaţii locale în structura α-HL, proteina este un bun candidat pentru o gamă largă de aplicaţii. Având în vedere importanţa biologică deosebită a interacţiunii dintre peptide şi pori proteici, au fost realizate numeroase studii care investighează partiţionarea peptidelor în porul proteic de α-HL [24-33].
VI. Detalii structurale şi funcţionale ale peptidelor utilizate în cadrul studiilor experimentale la nivel de singură moleculă
VI.1 Caracteristici specifice peptidei chimerice CA(1-8)MA(1-12):
CA(1-8)MA(1-12) - formată prin alăturarea primilor opt aminoacizi a peptidei cecropin A cu primii doisprezece aminoacizi a peptidei magainin 2:
Lys-Trp-Lys-Leu-Phe-Lys-Lys-Ile-Gly-Ile-Gly-Lys-Phe-Leu-His-Ser-Ala-Lys-Lys-Phe-NH2
VI.2 Caracteristici specifice peptidei amiloidice uman-Aβ(1-16), de provenienţă umană şi murin-Aβ(1-16), de provenienţă murină: uman - Aβ(1-16): Asp - Ala - Glu - Phe - Arg - His - Asp - Ser - Gly - Tyr - Glu - Val - His - His - Gln – Lys
murin - Aβ(1-16): Asp - Ala - Glu - Phe - Gly - His - Asp - Ser - Gly - Phe - Glu - Val - Arg - His - Gln – Lys
7

VI.3 Efectul metalelor de tranziţie asupra conformaţiei peptidelor amiloidice
Din studiile efectute până la ora actuală reiese clar faptul că dereglarea homeostaziei metalelor de tranziţie prezente în organism joacă un rol important în etiologia bolii neurodegerative Alzheimer. Astfel că, elucidarea detaliilor de ordin structural ce stau la baza interacţiunilor dintre aceste metale şi peptidele amilodice ar facilita înţelegerea apariţiei neurotoxicităţii şi proiectarea unor substanţe chimice care să aibă o acţiune terapeutică. Studiile efectuate asupra modificărilor de conformaţie a acestor peptide induse de ionii de zinc indică implicarea celor trei reziduuri de histidină (His6, His13 şi His14), ca şi în cazul cuprului. A patra legătură coordonativă fiind încă necunoscută, unele studii propunând ca posibili candidaţi: oxigenul acidului glutamic (Glu11), capătul N – terminal atunci când acesta este acetilat sau acidul aspartic (Asp1), atunci când capătul N-terminal nu este acetilat [34]. Studii comparative, de rezonanţă magnetică nucleară, 1H RMN, ce vizează efectului ionilor de zinc asupra segmentului de peptidă Aβ(1-28) de provenienţă umană şi murină dizolvată în apă sau SDS (sodium dodecyl sulphate), la pH neutru [35] au arătat că acidul glutamic (Glu11) ar avea potenţialul cel mai mare pentru a forma a patra legătură coordonativă cu ionii de zinc, iar diferenţa de o histidină (H13R) dintre cele două peptide ar fi responsabilă pentru conformaţia mai stabilă adoptată de peptida de provenienţă umană, cea care conţine trei histidine, decât cealaltă, ce prezintă doar două histidine.
VI.4. Natura chimică a interacţiunii aminoacidului histidină cu ionii de cupru
Nivelul de cupru regăsit la nivel neuronal este crescut semnificativ la pacienţii care deja se află într-o stare avansată a bolii Alzheimer faţă de pacienţii sănătoşi. Cifrele indicând valori ale cuprului de la 4.4 ± 1.5 µg/g în cazul eşantionului control, până la 19.3 ± 6.3 µg/g în cazul eşantionului test, ajungându-se chiar până la valori de 30.1 ± 11 µg/g în analizei în mijlocul plăcii senile [36].
Histidina (His) este un unul din cei 20 de aminoacizii principali care intră în componenţa proteinelor şi a peptidelor. Acesta conţine un inel imidazol ca şi catenă laterală, aromatic la orice valoare a pH-ului. Conţine şase electroni π: patru din două legături duble şi doi de la azot, aceştia din urmă fiind cei cu perechi de electroni liberi ce pot contribui la formarea legăturilor de coordinare cu metalele. Electronii liberi ai azotului din gruparea amină contribuie la caracterul aromatic al inelului imidazol. Prin hibridizarea orbitalului 2sp şi completarea acestuia cu încă un electron de la atomul de hidrogen acesta nu mai are electroni liberi pentru a forma legăruri coordinative cu metalele. Pe de altă parte, azotul din grupare imină, care are două legături sigma şi una pi, se află deja în stare hibridizată 2sp.
8

Fig.VI.4.2 Configurațiile electronice și gruparea electronilor pe subnivelele energetice a atomului de azot simplu și în a celor din legăturile grupărilor amină (dreapta), respectiv imina (stânga) a aminoacidului histidină. Pentru simplitate reprezentării grupările amină (-NH2) și acidul carboxilic (COOH) care intră în lanțul polipeptidic al unei peptide au fost notate cu R, de interes fiind doar catena laterală a aminoacidului histidină (inelul imidazol). În cazul grupării amină hibridizarea nivelului energetic se face la interacțiunea acestuia cu atomul de hidrogen (acesta având doar trei legături de tip σ), contrat grupării imina în care nivelul energetic este deja hibridizat (acesta având două legături de tip σ și una de tip π).
Astfel că perechea de electroni liberi se află deja pe orbitalul 2sp hibridizat şi cu care poate contribui la legătura coordinativă cu ionii de cupru, mai exact cu orbitalul 4sp. Din acest motiv, ionii de cupru se leagă preferenţial de atomii de azot din anumiţi aminoacizi, cum ar fi histidinele din structura primară a peptidei β – amiloid.
VII. Tehnici şi metode aplicare în studiul influenţei structurii unor peptide asupra modului de interacţiune cu sistemele lipo-proteice
Metoda „Montal-Muller” de realizare a membranelor lipidice artificiale. Monitorizarea canalelor voltaj dependente formate de peptidele citotoxice în bistraturi lipidice reprezintă una din tehnicile utile de investigare a formării şi stabilităţii porilor transmembranari induşi de acestea la nivelul membranele celulare biologice. Amplitudinea curentului electric mediat de porii formaţi de peptidele citotoxice în membranele artificiale reprezintă o măsură a capacităţii de penetrare a biomembranelor de catre aceste molecule peptidice. Atunci când peptidele se adsorb la interfaţa bistrat/mediu apos, se inseră în miezul hidrofob al bistratului formând pori apoşi transmembranari ce pot fi monitorizaţi înregistrând modificările de curent electric ce apar în sistem în urma aplicării unei deferenţe de potenţial de o parte şi cealaltă a membranei lipidice. (Fig.VII.1.1)
9

Fig.VII.1.1 Reprezentare schematizată a componentelor dispozitivului experimental utilizat pentru realizarea biomembranelor artificiale. Bistratul lipidic se formează la nivelul aperturii de diametru de 100 µm din filmul de teflon cu grosime de 25 µm vedere detaliată a biomembranei formate la nivelul partiţiei de teflon ce separă cele două cuve de teflon, în care, pentru exemplificare, se consideră inserat un canal ionic de alameticină. Curentul electric a fost detectat şi amplificat cu un amplificator integrat Axopatch 200 B (Molecular Devices, USA) setat pe modul de voltaj fixat, prin intermediul a doi electrozi de Ag/AgCl. Achiziţia datelor s-a realizat cu o placă de achiziţie NI PCI 6221 pe 16 biţi (National Instruments USA), cu o frecvenţă de achiziţie de 30 kHz în mediul grafic LabVIEW 8.20.
Înregistrări de electrofiziologie la nivel de singură moleculă. Utilizarea proteinei α-hemolizină ca nanosenzor. Înregistrarea fluctuațiilor de curent electric mediat de canale ionice membranare prin tehnici de electrofiziologie moleculară este esențială pentru vizualizarea directă a tranziţiilor moleculare realizate de un singur canal ionic și pentru a studia fenomenele de transport ionic sau a unor molecule mici prin aceste canale ionice. Aceste tranziții au un caracter pur stochastic astfel încât, în descrierea comportamentului acestor molecule sunt importante estimarea numărului exact de stări moleculare (de tip „închis” și „deschis”) prin care poate trece un canal ionic, determinarea ratelor de tranziție între aceste stări precum și analiza proceselor de transport ionic mediate de acele canale.
Fig.VII.2.1 Fluctuaţii specifice ale curentului ionic printr-un singur por proteic de α-HL induse în urma adiţiei în partea trans a membranei lipidice a peptidelor, înregistrate la o valoare a diferenţei de potenţial aplicate de -50 mV, observate sub forma unor alterări de tip treaptă ale curentului ionic spre valori mai pozitive ale acestuia.
10

Peptidele interacţionează rapid cu zona de constricţie a porului proteic conducând la evenimente de blocaj complet sau parţial, cu o durată suficient de mare pentru ca monitorizarea lor să fie posibilă. Valorile amplitudinii blocajelor porului proteic de către peptidă sunt date de conformaţia în care se regăseşte peptida, aceasta poate bloca complet porul în zona de constricţie, ceea ce sugerează o aranjare spaţială mai voluminoasă a peptidei, sau poate bloca parţial porul, ceea ce indică o aranjare liniară a pepdei la trecea acesteia prin por.
Prepararea lipozomilor unilamelari mici prin metoda deshidratării. Lipozomii, cele mai întâlnite nanoparticule alcătuite din molecule lipidice, pot fi reprezentați printr-o sferă a cărei diametru variază între 30 nm şi 10 μm. Lipozomii se formează atunci când un film lipidic subțire este hidratat iar mulțimile de bistraturi lichid-cristaline devin fluide și cresc în mărime. Straturile de lipide hidratate se desprind în timpul procesului de agitație iar capetele acestora se aproprie pentru a evita contactul apei cu miezul hidrofob al bistratului, formând vezicule multilamelare. Pentru a reduce dimensiunile acestor vezicule este nevoie de aport energetic sub formă de energie sonică (sonicare) sau energie mecanică (extrudare), rezultând astfel vezicule unilamelare mici (SUVs).
Fig.VII.3.3 Reprezentare schematizată a etapelor de formare a vezicuelor unilamelare mici (SUV). În general, procedura de obținere a lipozomilor presupune: prepararea lipidelor pentru hidratate, hidratarea prin agitație și aducerea la o distribuție omogenă a dimensiunii veziculelor prin sonicare.
Metode spectrofluorometrice de analiză şi control bazate pe emisia aminoacizilor aromatici. Spectroscopia de fluorescenţă (fluorometria sau spectrofluorometria) este un tip de spectroscopie electromagnetică ce analizează fluorescenţa unei probe de studiu. Acest tip de analiză implică utilizarea unui fascicul de lumină, de obicei cu raze ultraviolete, care excită electronii moleculelor din anumiţi compuşi ceea ce duce la emisia unei lumini cu energie mai mică de obicei, dar nu neapărat, în lumina vizibilă.
11

Fig. VII.4.4 Dacă inițial triptofanul se află într-un solvent polar iar ulterior trece într-un mediu hidrofob (apolar), spectrul de emisie va fi deplasat spre regiunea albastră a lungimilor de undă.
Triptofanul prezintă un spectru de absorbție cu maximul la 280 nm, pe când maximul spectrului de emisie variază între 300 și 350 nm, în funcție de polaritatea solventului [37,38]. Triptofanul existent în structura primară a proteinelor poate fi deci utilizat pentru estimarea naturii mediului în care se află. În funcție de polaritatea, flexibilitatea și natura chimică a mediului în care se găsește triptofanul, acesta va interacționa diferit la redistribuirea densității electronice în inelul indol al aminoacidului.
Tehnica monitorizării în timp real a efluxului de calceină încapsulat în interiorul lipozomilor unilamelari mici sub acţiunea peptidelor. Fluoroforul calceină se prezintă sub formă de cristale de culoare portocalie, având lungimea de undă de excitaţie la 495 nm, respectiv maximul spectrului de emisie la 525 nm (Fig.VII.5.1). În literatura de specialitate este specificat faptul că fluoroforul calceină îşi auto-stinge emisia fluorescentă chiar şi la valori ale concentraţiei de sub 100 mM [39]. Pe parcursul experimentelor, calceina a fost solubilizată în soluţie într-o concentraţie de 50 mM, care asigură autostingerea fluorescenței. Sistemele lipidice utilizate în aceste experimente sunt lipozomi sferici formați prin același protocol ca cel descris în subcapitolul VII.3., hidratați cu ajutorul soluției de calceină. Pentru a elimina calceina din mediul extern al veziculelor unilamelare mici astfel formate, suspensia de lipozomi a fost supusă unui proces de separare cromatografică pe coloane cu gel [40]. Eliminarea calceinei libere din suspensia lipidică s-a făcut folosind Coloane PD-10 cu un conţinut de gel Sephadex G-25, ce asigură separarea compuşilor cu masă moleculară mare de cei cu masă moleculară mică. Peptidele citotoxice se inseră în bistratul lipidic şi formează pori transmembranari prin intermediul cărora calceina, aflată în interior la o concentraţie care asigură stingerea fluorescenţei, este eliberată în mediul extern şi emite fluorescent. Evoluția în timp real a intensității fluorescenței poate fi corelată cu cinetica de formare a porilor de către peptidele citotoxice studiate.
Tehnica stingerii emisiei de fluorescenţă a floroforilor din structura peptidelor. Stingerea fluorescenţei (quenching) se referă la orice proces care induce scăderea intensităţii de fluorescenţă a unor anumite substanţe denumite florofori în urma interacţiunii acestora cu alte molecule, denumite molecule quencher.
12
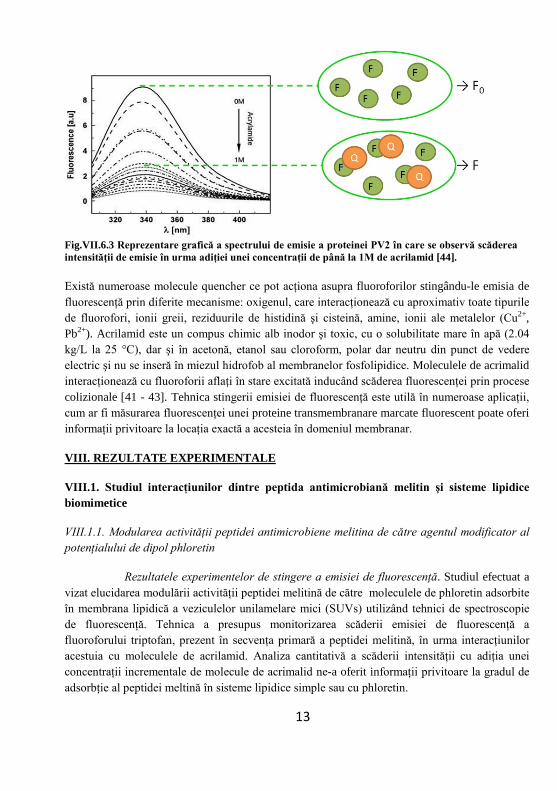
Fig.VII.6.3 Reprezentare grafică a spectrului de emisie a proteinei PV2 în care se observă scăderea intensităţii de emisie în urma adiţiei unei concentraţii de până la 1M de acrilamid [44].
Există numeroase molecule quencher ce pot acţiona asupra fluoroforilor stingându-le emisia de fluorescenţă prin diferite mecanisme: oxigenul, care interacţionează cu aproximativ toate tipurile de fluorofori, ionii greii, reziduurile de histidină şi cisteină, amine, ionii ale metalelor (Cu2+, Pb2+). Acrilamid este un compus chimic alb inodor şi toxic, cu o solubilitate mare în apă (2.04 kg/L la 25 °C), dar şi în acetonă, etanol sau cloroform, polar dar neutru din punct de vedere electric şi nu se inseră în miezul hidrofob al membranelor fosfolipidice. Moleculele de acrimalid interacţionează cu fluoroforii aflaţi în stare excitată inducând scăderea fluorescenţei prin procese colizionale [41 - 43]. Tehnica stingerii emisiei de fluorescenţă este utilă în numeroase aplicaţii, cum ar fi măsurarea fluorescenţei unei proteine transmembranare marcate fluorescent poate oferi informaţii privitoare la locaţia exactă a acesteia în domeniul membranar.
VIII. REZULTATE EXPERIMENTALE
VIII.1. Studiul interacţiunilor dintre peptida antimicrobiană melitin şi sisteme lipidice biomimetice
VIII.1.1. Modularea activităţii peptidei antimicrobiene melitina de către agentul modificator al potenţialului de dipol phloretin
Rezultatele experimentelor de stingere a emisiei de fluorescenţă. Studiul efectuat a vizat elucidarea modulării activităţii peptidei melitină de către moleculele de phloretin adsorbite în membrana lipidică a veziculelor unilamelare mici (SUVs) utilizând tehnici de spectroscopie de fluorescenţă. Tehnica a presupus monitorizarea scăderii emisiei de fluorescenţă a fluoroforului triptofan, prezent în secvenţa primară a peptidei melitină, în urma interacţiunilor acestuia cu moleculele de acrilamid. Analiza cantitativă a scăderii intensităţii cu adiţia unei concentraţii incrementale de molecule de acrimalid ne-a oferit informaţii privitoare la gradul de adsorbţie al peptidei meltină în sisteme lipidice simple sau cu phloretin.
13

Fig.VIII.1.1.2 Spectrele de emisie a 1µM peptidă melitin în sisteme lipidice simple. Spectrul de emisie iniţial a melitinei este reprezentat cu negru. Se observă scăderea intensităţii de fluorescenţă a fluoroforului triptofan din structura peptidei melitină în urma adiţiei moleculelor de acrilamid în concentraţii de 75 mM (verde) şi respectiv 150 mM (roşu).
Fig.VIII.1.1.3 Spectrele de emisie a 1µM peptidă melitin în sisteme lipidice cu 20 µM phloretin. Spectrul de emisie iniţial a melitinei este reprezentat cu negru. Se observă scăderea intensităţii de fluorescenţă a fluoroforului triptofan din structura peptidei melitină în urma adiţiei moleculelor de acrilamid în concentraţii de 75 mM (albastru) şi respectiv 150 mM (roz).
Se poate observa, că pentru o aceeaşi valoare a concentraţiei de acrilamid, de exemplu 75 mM, scăderea maximului spectrului de emisie a melitinei este dimnuată în cazul sistemelor lipidice care conţin molecule de phloretin. Acest fapt este datorat gradului de expunere diminuat a fluoroforului (aminoacidul triptofan) către moleculele de quencher (acrilamid), interacţiunea dintre aceştia fiind diminuată. Analiza cantitativă a scăderii emisiei de fluorescenţă şi fitarea cu ecuaţia Stern-Volmer a variaţiei liniare a acesteia de concentraţia de acrilamid, ne oferă posibilitate calculului constantei Stern-Volmer în cazul proceselor de quenching colizional, a cărei valoare ne indică gradul de adsorbţie a peptidei în sistemele lipidice.
Fig.VIII.1.1.4 Reprezentarea fitului liniar al datelor experimentale obţinute în urma monitorizării scăderii intensităţii fluorescenţei cu creşterea concentraţiei de acrilamid în sisteme lipidice simple (verde) şi sisteme lipidice cu phloretin (maro). Scăderea intensităţii este reprezentată sub forma raportului dintre maximul intensităţii fluorescenţei iniţiale, în absenţa acrilamidului şi maximul intensităţii după adiţia acestuia.
MLT in: KSV (M-1)
SUV 7.41 ± 0.07
SUV+Phl 3.73 ± 0.30
14

Analiza valorilor constantelor obţinute ne indică faptul că monomerii de peptidă melitin sunt adsorbiţi într-o mai mare măsură în sistemele lipidice cu phloretin decât în cele formate doar din lipide simple. Prezenţa moleculelor de phloretin favorizează adsorbţia peptidelor la nivelul membranei lipidice datorită modulării de către acesta a proprietăţilor electrice a membranei (potenţialul de dipol), jucând un rol important în cazul interacţiunilor între peptide antimicrobiene şi membrane specifice, cum ar fi cele bacteriene.
Rezultatele experimentelor de eliberare a calceinei din vezicule unilamelare mici. Scopul acestui studiu a fost acela de a investiga efectul moleculelor de phloretin asupra proceselor de inserție și de agregare a peptidelor în urma adsorbției lor la interfața membranei lipidice. Cinetica de formare a porilor de către peptide poate fi astfel monitorizată prin înregistrarea în timp real a modificării maximului de emisie fluorescentă a moleculelor de calceină eliberate în mediul apos. Astfel, am înregistrat evoluția în timp a intensității emisiei fluorescente a moleculelor de calceină încapsulate într-o concentrație inițială de 50 mM în vezicule unilamelare mici formate din fosfatidilcolină, la lungimea de undă corespunzătoare maximului de emisie (λmax = 520 nm), în urma eliberării lor prin porii transmembranri formați de peptida melitină adăugată în sistem într-o concentrație de 3 µM. Acțiunea peptidei a fost studiată prin comparație între sisteme lipidice formate din vezicule unilamelare mici în concentrație lipidică de 100 µM, în absența, respectiv în prezența a 20 µM molecule phloretin.
Fig.VIII.1.1.5 Evoluția temporală a intensității emisiei fluorescente a calceinei, ca urmare a eliberării acesteia din interiorul lipozomilor formați din fosfatidilcolină, în absența (curba verde), respectiv în prezența moleculelor de phloretin inserate în bistratul lipidic (curba maro), sub acțiunea peptidei melitină.
Analizând cele două curbe, se constată că prezența moleculelor de phloretin inserate în bistratul lipidic al lipozomilor a condus la creșterea concentrației moleculelor de calceină eliberate (intensitatea de emisie fluorescentă la saturație este mai mare în acest caz), ca o consecință directă a formării unui număr mai mare de pori transmembranari în bistraturile lipidice care conțin phloretin. Prezența moleculelor de phloretin conduce la creșterea activității peptidei melitină, prin reducerea energiei libere necesare pentru inserția monomerilor și agregarea lor sub formă de pori.
15
VIII.1.2. Influenţa tăriei ionice asupra adsorbţiei şi formării de pori transmembranari de către peptida antimicrobiană melitin
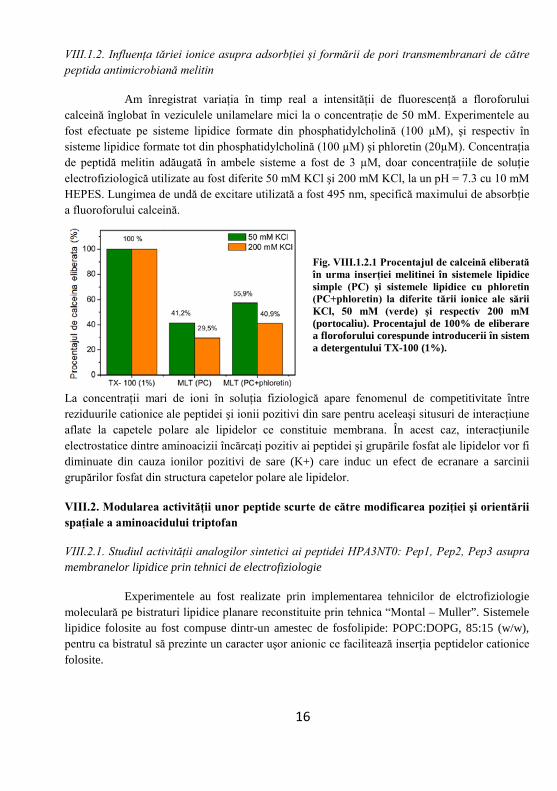
Am înregistrat variaţia în timp real a intensităţii de fluorescenţă a floroforului calceină înglobat în veziculele unilamelare mici la o concentraţie de 50 mM. Experimentele au fost efectuate pe sisteme lipidice formate din phosphatidylcholină (100 µM), şi respectiv în sisteme lipidice formate tot din phosphatidylcholină (100 µM) şi phloretin (20µM). Concentraţia de peptidă melitin adăugată în ambele sisteme a fost de 3 µM, doar concentraţiile de soluţie electrofiziologică utilizate au fost diferite 50 mM KCl şi 200 mM KCl, la un pH = 7.3 cu 10 mM HEPES. Lungimea de undă de excitare utilizată a fost 495 nm, specifică maximului de absorbţie a fluoroforului calceină.
Fig. VIII.1.2.1 Procentajul de calceină eliberată în urma inserţiei melitinei în sistemele lipidice simple (PC) şi sistemele lipidice cu phloretin (PC+phloretin) la diferite tării ionice ale sării KCl, 50 mM (verde) şi respectiv 200 mM (portocaliu). Procentajul de 100% de eliberare a floroforului corespunde introducerii în sistem a detergentului TX-100 (1%).
La concentraţii mari de ioni în soluţia fiziologică apare fenomenul de competitivitate între reziduurile cationice ale peptidei şi ionii pozitivi din sare pentru aceleaşi situsuri de interacţiune aflate la capetele polare ale lipidelor ce constituie membrana. În acest caz, interacţiunile electrostatice dintre aminoacizii încărcaţi pozitiv ai peptidei şi grupările fosfat ale lipidelor vor fi diminuate din cauza ionilor pozitivi de sare (K+) care induc un efect de ecranare a sarcinii grupărilor fosfat din structura capetelor polare ale lipidelor.
VIII.2. Modularea activităţii unor peptide scurte de către modificarea poziţiei şi orientării spaţiale a aminoacidului triptofan
VIII.2.1. Studiul activităţii analogilor sintetici ai peptidei HPA3NT0: Pep1, Pep2, Pep3 asupra membranelor lipidice prin tehnici de electrofiziologie
Experimentele au fost realizate prin implementarea tehnicilor de elctrofiziologie moleculară pe bistraturi lipidice planare reconstituite prin tehnica “Montal – Muller”. Sistemele lipidice folosite au fost compuse dintr-un amestec de fosfolipide: POPC:DOPG, 85:15 (w/w), pentru ca bistratul să prezinte un caracter uşor anionic ce facilitează inserţia peptidelor cationice folosite.
16
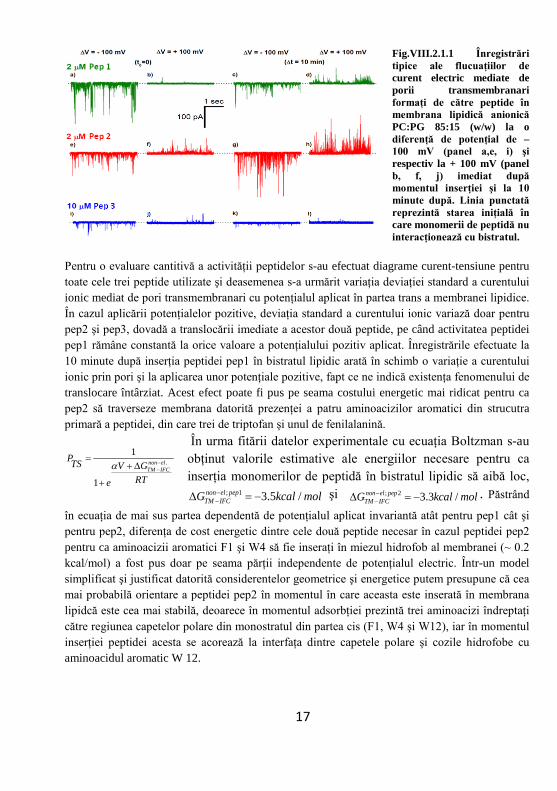
Fig.VIII.2.1.1 Înregistrări tipice ale flucuaţiilor de curent electric mediate de porii transmembranari formaţi de către peptide în membrana lipidică anionică PC:PG 85:15 (w/w) la o diferenţă de potenţial de – 100 mV (panel a,e, i) şi respectiv la + 100 mV (panel b, f, j) imediat după momentul inserţiei şi la 10 minute după. Linia punctată reprezintă starea iniţială în care monomerii de peptidă nu interacţionează cu bistratul.
Pentru o evaluare cantitivă a activităţii peptidelor s-au efectuat diagrame curent-tensiune pentru toate cele trei peptide utilizate şi deasemenea s-a urmărit variaţia deviaţiei standard a curentului ionic mediat de pori transmembranari cu potenţialul aplicat în partea trans a membranei lipidice. În cazul aplicării potenţialelor pozitive, deviaţia standard a curentului ionic variază doar pentru pep2 şi pep3, dovadă a translocării imediate a acestor două peptide, pe când activitatea peptidei pep1 rămâne constantă la orice valoare a potenţialului pozitiv aplicat. Înregistrările efectuate la 10 minute după inserţia peptidei pep1 în bistratul lipidic arată în schimb o variaţie a curentului ionic prin pori şi la aplicarea unor potenţiale pozitive, fapt ce ne indică existenţa fenomenului de translocare întârziat. Acest efect poate fi pus pe seama costului energetic mai ridicat pentru ca pep2 să traverseze membrana datorită prezenţei a patru aminoacizilor aromatici din strucutra primară a peptidei, din care trei de triptofan şi unul de fenilalanină.
În urma fitării datelor experimentale cu ecuaţia Boltzman s-au obţinut valorile estimative ale energiilor necesare pentru ca inserţia monomerilor de peptidă în bistratul lipidic să aibă loc,
; 1 3.5 /non el pepTM IFCG kcal mol−
−∆ = − şi ; 2 3.3 /non el pepTM IFCG kcal mol−
−∆ = − . Păstrând
în ecuaţia de mai sus partea dependentă de potenţialul aplicat invariantă atât pentru pep1 cât şi pentru pep2, diferenţa de cost energetic dintre cele două peptide necesar în cazul peptidei pep2 pentru ca aminoacizii aromatici F1 şi W4 să fie inseraţi în miezul hidrofob al membranei (~ 0.2 kcal/mol) a fost pus doar pe seama părţii independente de potenţialul electric. Într-un model simplificat şi justificat datorită considerentelor geometrice şi energetice putem presupune că cea mai probabilă orientare a peptidei pep2 în momentul în care aceasta este inserată în membrana lipidcă este cea mai stabilă, deoarece în momentul adsorbţiei prezintă trei aminoacizi îndreptaţi către regiunea capetelor polare din monostratul din partea cis (F1, W4 şi W12), iar în momentul inserţiei peptidei acesta se acorează la interfaţa dintre capetele polare şi cozile hidrofobe cu aminoacidul aromatic W 12.
.1
1
non elTM IFC
PTS V GRTe
α −−
=+ ∆
+
17

VIII.2.2. Studiul adsorbţiei peptidelor Pep1, Pep2, Pep3 în sisteme de lipozomi prin tehnici de spectroscopie de fluorescenţă
Aceste experimente sunt bazate pe proprietatea aminoacidului triptofan, prezent în secvența primară a celor trei peptide utilizate, de a-şi deplasare maximului de emisie fluorescentă la trecerea din mediul polar al soluției electrofiziologice în mediul mai hidrofob de la interfața membranei lipozomilor. Ținând cont de proporționalitatea care există între deplasarea maximului de emisie și concentraţia de peptidă adsorbită, dependența mărimii de concentrația lipidică oferă informații despre cum variază de fapt concentrația de peptidă adsorbită în funcție de concentrația lipidică.
Fig.VIII.2.2.3 Variaţia deplasării maximului de emisie a triptofanului în funcţie de concentraţia lipidică înjumătăţită pentru peptidele utilizate şi fiturile corespunzătoare acestora, reprezentate cu linie punctată pentru pep1 (verde), pep2 (roşu) şi respectiv pep3 (albastru), ca urmare a adsorbţiei peptidei în concentraţie de 0.5 mM la nivelul veziculelor unilamelare mici.
max a
a
K [L]1 K [L]
⋅∆λ = ∆λ ⋅
+
Din datele experimentale şi folosindu-ne de modelul matematic explica în detaliu anterior ce ne oferă valorile numerice ale constantelor aparente de asociere a peptidelor cu membrana lipidică, am calculat energia liberă de partiţionare a peptidelor din mediul apos a soluţiei electrofiziologice în mediul lipidic al monostratului extern al lipozomilor: pep1 (-9.29 kcal/mol), pep2 (-10.02 kcal/mol) şi pep3 (-9.01 kcal/mol).
VIII.2.3. Studiul cineticii de inserţie şi formare a porilor transmembranari de către Pep1, Pep2, Pep3 prin monitorizarea spectrofluorimetrică a efluxului de calceină din lipozomi
În cele ce urmează, ne-am propus efectuarea unui studiu de spectrometrie de fluorescență prin care să poată fi monitorizată etapa de formare a porilor transmembranari de către peptidele antimicrobiene în bistratul lipidic al veziculelor unilamelare mici.
18
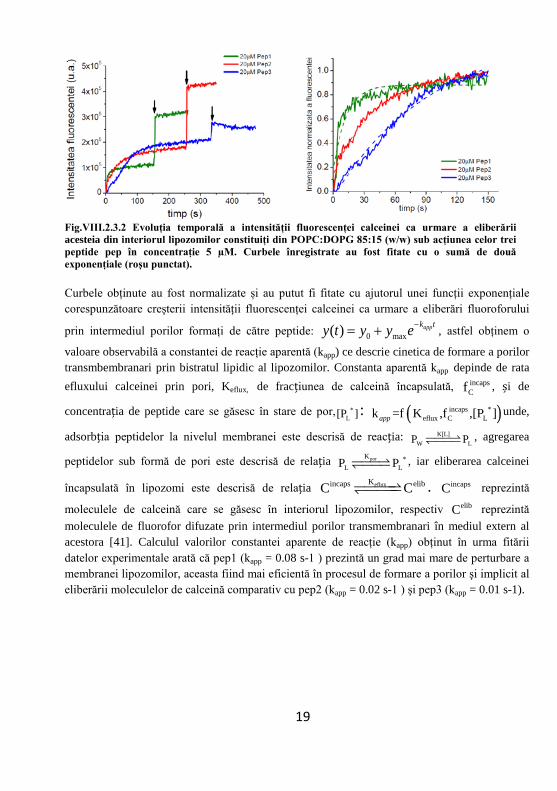
Fig.VIII.2.3.2 Evoluția temporală a intensității fluorescenței calceinei ca urmare a eliberării acesteia din interiorul lipozomilor constituiți din POPC:DOPG 85:15 (w/w) sub acțiunea celor trei peptide pep în concentrație 5 µM. Curbele înregistrate au fost fitate cu o sumă de două exponențiale (roșu punctat).
Curbele obținute au fost normalizate și au putut fi fitate cu ajutorul unei funcţii exponenţiale corespunzătoare creşterii intensităţii fluorescenţei calceinei ca urmare a eliberări fluoroforului
prin intermediul porilor formaţi de către peptide: 0 max( ) appk ty t y y e−= + , astfel obţinem o
valoare observabilă a constantei de reacţie aparentă (kapp) ce descrie cinetica de formare a porilor transmbembranari prin bistratul lipidic al lipozomilor. Constanta aparentă kapp depinde de rata efluxului calceinei prin pori, Keflux, de fracțiunea de calceină încapsulată, incaps
Cf , și de
concentrația de peptide care se găsesc în stare de por, *L[P ] : ( )incaps *
eflux C Lk =f K ,f ,[P ]appunde,
adsorbția peptidelor la nivelul membranei este descrisă de reacția: K[L]W LP P
, agregarea
peptidelor sub formă de pori este descrisă de relația porK *L LP P
, iar eliberarea calceinei
încapsulată în lipozomi este descrisă de relația efluxKincaps elibC C
. incapsC reprezintă
moleculele de calceină care se găsesc în interiorul lipozomilor, respectiv elibC reprezintă moleculele de fluorofor difuzate prin intermediul porilor transmembranari în mediul extern al acestora [41]. Calculul valorilor constantei aparente de reacţie (kapp) obţinut în urma fitării datelor experimentale arată că pep1 (kapp = 0.08 s-1 ) prezintă un grad mai mare de perturbare a membranei lipozomilor, aceasta fiind mai eficientă în procesul de formare a porilor şi implicit al eliberării moleculelor de calceină comparativ cu pep2 (kapp = 0.02 s-1 ) şi pep3 (kapp = 0.01 s-1).
19
VIII.3. Monitorizarea schimbărilor conformaţionale a unor peptide în prezenţa unor metale relevante fiziologic
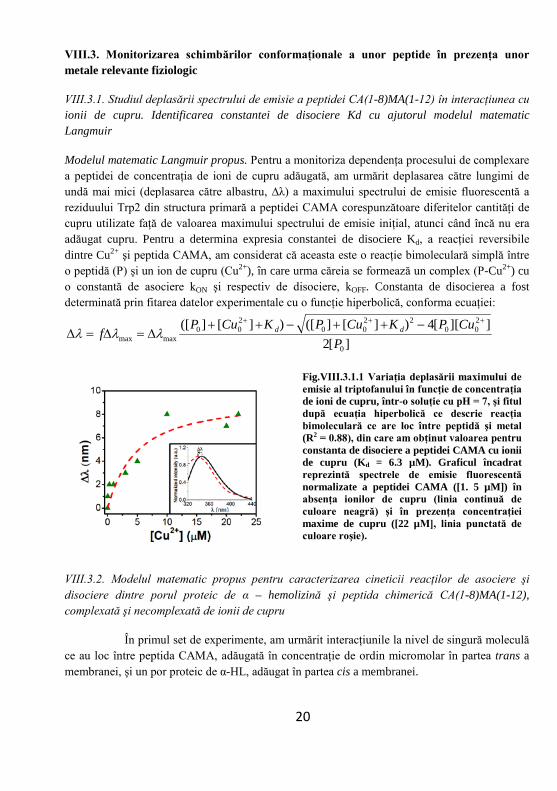
VIII.3.1. Studiul deplasării spectrului de emisie a peptidei CA(1-8)MA(1-12) în interacţiunea cu ionii de cupru. Identificarea constantei de disociere Kd cu ajutorul modelul matematic Langmuir
Modelul matematic Langmuir propus. Pentru a monitoriza dependenţa procesului de complexare a peptidei de concentraţia de ioni de cupru adăugată, am urmărit deplasarea către lungimi de undă mai mici (deplasarea către albastru, ∆λ) a maximului spectrului de emisie fluorescentă a reziduului Trp2 din structura primară a peptidei CAMA corespunzătoare diferitelor cantităţi de cupru utilizate faţă de valoarea maximului spectrului de emisie iniţial, atunci când încă nu era adăugat cupru. Pentru a determina expresia constantei de disociere Kd, a reacţiei reversibile dintre Cu2+ şi peptida CAMA, am considerat că aceasta este o reacţie bimoleculară simplă între o peptidă (P) şi un ion de cupru (Cu2+), în care urma căreia se formează un complex (P-Cu2+) cu o constantă de asociere kON şi respectiv de disociere, kOFF. Constanta de disocierea a fost determinată prin fitarea datelor experimentale cu o funcţie hiperbolică, conforma ecuaţiei:
][2]][[4)][]([)][]([
0
200
2200
200
maxmax PCuPKCuPKCuP
f dd+++ −++−++
∆=∆=∆ λλλ
Fig.VIII.3.1.1 Variaţia deplasării maximului de emisie al triptofanului în funcţie de concentraţia de ioni de cupru, într-o soluţie cu pH = 7, şi fitul după ecuaţia hiperbolică ce descrie reacţia bimoleculară ce are loc între peptidă şi metal (R2 = 0.88), din care am obţinut valoarea pentru constanta de disociere a peptidei CAMA cu ionii de cupru (Kd = 6.3 µM). Graficul încadrat reprezintă spectrele de emisie fluorescentă normalizate a peptidei CAMA ([1. 5 µM]) în absenţa ionilor de cupru (linia continuă de culoare neagră) şi în prezenţa concentraţiei maxime de cupru ([22 µM], linia punctată de culoare roşie).
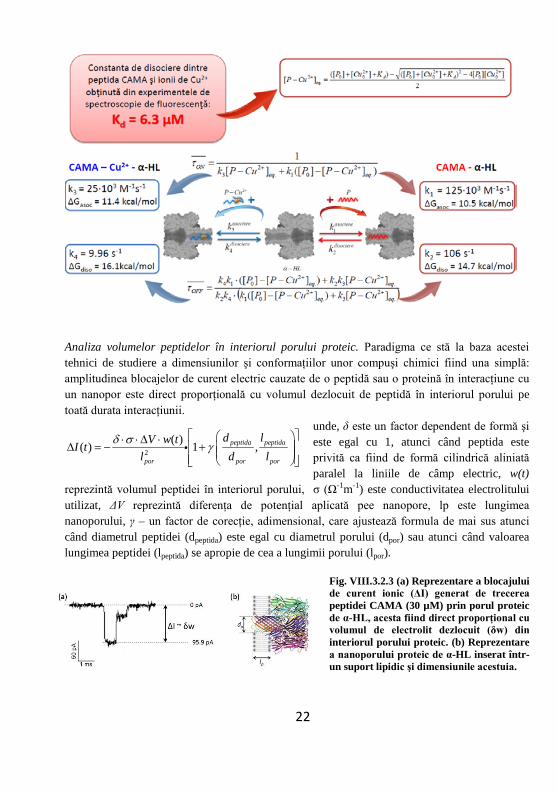
VIII.3.2. Modelul matematic propus pentru caracterizarea cineticii reacţilor de asociere şi disociere dintre porul proteic de α – hemolizină şi peptida chimerică CA(1-8)MA(1-12), complexată şi necomplexată de ionii de cupru
În primul set de experimente, am urmărit interacţiunile la nivel de singură moleculă ce au loc între peptida CAMA, adăugată în concentraţie de ordin micromolar în partea trans a membranei, şi un por proteic de α-HL, adăugat în partea cis a membranei.
20

Fig. VIII.3.2.1 Înregistrări ale curenţilor ionici reprezentative ilustrând efectul creşterii concentraţiei de ioni de cupru asupra interacţiunilor peptidei CAMA (30 μM) cu un singur por proteic de α-HL, în cazul unei soluţii electrofiziologice cu pH neutru de 2M KCl adăugată în ambele părţi ale membranei. În panelurile a, b şi c sunt reprezentate fluctuaţiile de curent ionic date de interacţiunea peptidei cu porul (sub forma scăderilor valorilor curentului faţă de linia de bază, corespunzătoare curentului ionic înregistrat pentru porul liber) la aplicarea unei diferenţe de potenţial de + 70 mV, în absenţa (panel a), respectiv în prezenţa ionilor de cupru într-o concentraţie de 10 μM (panel b) şi 100 μM (panel c).
În absenţa interacţiunii cu peptida CAMA, am propus ca starea porul proteic de α-HL să fie descrisă sub denumirea de „open state”. În această stare, nanoporul poate interacţiona fie cu peptida necomplexată cu ioni de cupru (notată cu P) ducând la generarea unui prim tip de blocaj, denumit „closed state”, fie poate interacţiona cu peptida complexată cu ioni de cupru notată cu P-Cu2+) generând un al doilea tip de blocaj posibil.
Fig. VIII.3.2.2 Analiza cantitativă a timpilor medii ce reflectă asocierea (τON; panel a) şi disocierea (τOFF; panel b) a unei singure peptide CAMA în interacţiunea sa cu un por proteic de α-HL la pH = 7, măsuraţi pentru diferite concentraţii de ioni de cupru (0, 3, 5, 10, 20, 50 and 100 μM) adăugate în partea trans a membranei.
Schematic, reacţiile ce pot avea loc, raţionamentul modelului matemati bazat pe calculul probabilităţilor de interacţiune dintre peptidă şi por precum şi rezultatele obţinute în urma aplicării acestuia sunt reprezentare în figura de mai jos:
21
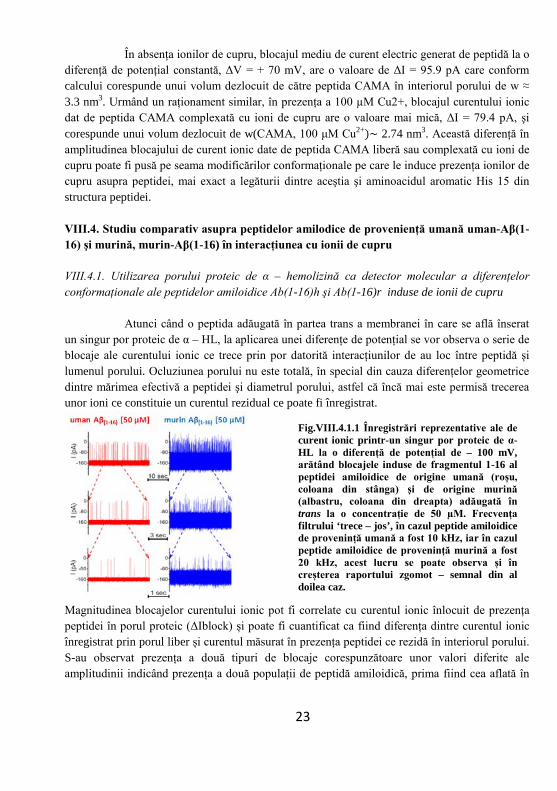
Analiza volumelor peptidelor în interiorul porului proteic. Paradigma ce stă la baza acestei tehnici de studiere a dimensiunilor şi conformaţiilor unor compuşi chimici fiind una simplă: amplitudinea blocajelor de curent electric cauzate de o peptidă sau o proteină în interacţiune cu un nanopor este direct proporţională cu volumul dezlocuit de peptidă în interiorul porului pe toată durata interacţiunii.
unde, δ este un factor dependent de formă şi este egal cu 1, atunci când peptida este privită ca fiind de formă cilindrică aliniată paralel la liniile de câmp electric, w(t)
reprezintă volumul peptidei în interiorul porului, σ (Ω-1m-1) este conductivitatea electrolitului utilizat, ΔV reprezintă diferenţa de potenţial aplicată pee nanopore, lp este lungimea nanoporului, γ – un factor de corecţie, adimensional, care ajustează formula de mai sus atunci când diametrul peptidei (dpeptida) este egal cu diametrul porului (dpor) sau atunci când valoarea lungimea peptidei (lpeptida) se apropie de cea a lungimii porului (lpor).
Fig. VIII.3.2.3 (a) Reprezentare a blocajului de curent ionic (ΔI) generat de trecerea peptidei CAMA (30 µM) prin porul proteic de α-HL, acesta fiind direct proporţional cu volumul de electrolit dezlocuit (δw) din interiorul porului proteic. (b) Reprezentare a nanoporului proteic de α-HL inserat într-un suport lipidic şi dimensiunile acestuia.
2
( )( ) 1 ,peptida peptida
por por por
d lV w tI tl d l
δ σ γ ⋅ ⋅∆ ⋅
∆ = − +
22
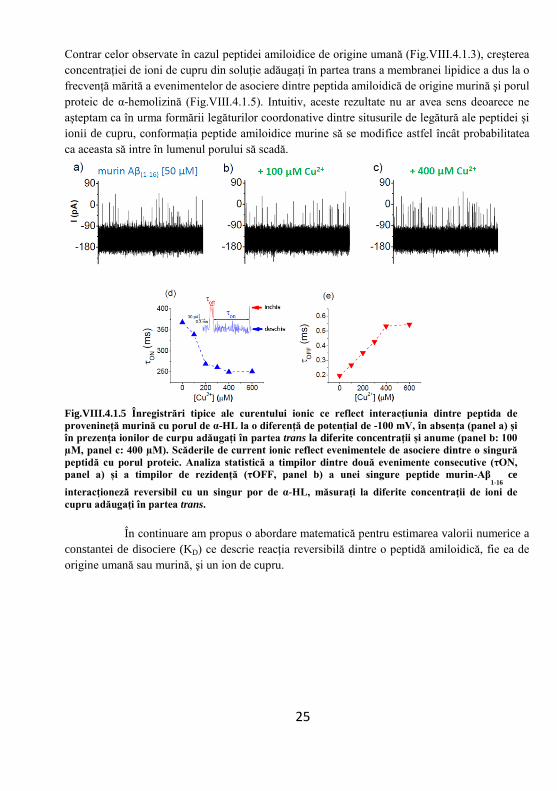
În absenţa ionilor de cupru, blocajul mediu de curent electric generat de peptidă la o diferenţă de potenţial constantă, ΔV = + 70 mV, are o valoare de ΔI = 95.9 pA care conform calcului corespunde unui volum dezlocuit de către peptida CAMA în interiorul porului de w ≈ 3.3 nm3. Urmând un raţionament similar, în prezenţa a 100 µM Cu2+, blocajul curentului ionic dat de peptida CAMA complexată cu ioni de cupru are o valoare mai mică, ΔI = 79.4 pA, şi corespunde unui volum dezlocuit de w(CAMA, 100 μM Cu2+)∼ 2.74 nm3. Această diferenţă în amplitudinea blocajului de curent ionic date de peptida CAMA liberă sau complexată cu ioni de cupru poate fi pusă pe seama modificărilor conformaţionale pe care le induce prezenţa ionilor de cupru asupra peptidei, mai exact a legăturii dintre aceştia şi aminoacidul aromatic His 15 din structura peptidei. VIII.4. Studiu comparativ asupra peptidelor amilodice de provenienţă umană uman-Aβ(1-16) şi murină, murin-Aβ(1-16) în interacţiunea cu ionii de cupru VIII.4.1. Utilizarea porului proteic de α – hemolizină ca detector molecular a diferenţelor conformaţionale ale peptidelor amiloidice Ab(1-16)h şi Ab(1-16)r induse de ionii de cupru
Atunci când o peptida adăugată în partea trans a membranei în care se află înserat un singur por proteic de α – HL, la aplicarea unei diferenţe de potenţial se vor observa o serie de blocaje ale curentului ionic ce trece prin por datorită interacţiunilor de au loc între peptidă şi lumenul porului. Ocluziunea porului nu este totală, în special din cauza diferenţelor geometrice dintre mărimea efectivă a peptidei şi diametrul porului, astfel că încă mai este permisă trecerea unor ioni ce constituie un curentul rezidual ce poate fi înregistrat.
Fig.VIII.4.1.1 Înregistrări reprezentative ale de curent ionic printr-un singur por proteic de α-HL la o diferență de potențial de – 100 mV, arătând blocajele induse de fragmentul 1-16 al peptidei amiloidice de origine umană (roşu, coloana din stânga) și de origine murină (albastru, coloana din dreapta) adăugată în trans la o concentrație de 50 µM. Frecvența filtrului ‘trece – jos’, în cazul peptide amiloidice de provenință umană a fost 10 kHz, iar în cazul peptide amiloidice de provenință murină a fost 20 kHz, acest lucru se poate observa și în creșterea raportului zgomot – semnal din al doilea caz.
Magnitudinea blocajelor curentului ionic pot fi correlate cu curentul ionic înlocuit de prezența peptidei în porul proteic (ΔIblock) și poate fi cuantificat ca fiind diferența dintre curentul ionic înregistrat prin porul liber și curentul măsurat în prezența peptidei ce rezidă în interiorul porului. S-au observat prezenţa a două tipuri de blocaje corespunzătoare unor valori diferite ale amplitudinii indicând prezenţa a două populaţii de peptidă amiloidică, prima fiind cea aflată în
23

stare liberă (necomplexată, indicată de simbolul #) şi a doua corespunzătoare blocajului dat de peptida care şi-a schimbat conformaţia în urma interacţiunii cu ionii de cupru (complexată, indicată prin simbolul $).
Fig.VIII.4.1.2 Înregistrări tipice ale curentului ionic ce reflect interacțiunia dintre peptida de provenineță umană cu porul de α-HL la o diferență de potențial de -100 mV, în absența (panel a) și în prezența ionilor de curpu adăugați în partea trans la diferite concentrații, și anume 0, 25, 100 și 200 µM cupru (panel b, c și d). Scăderile de current ionic reflect evenimentele de asociere dintre o singură peptidă cu porul proteic. Histogramele de amplitudine ne indică prezeneța a două populații de peptide, complexate (indicate prin $) și necomplexate (indicate prin #) cu ioni de cupru, deasemenea și prezența unor interacțiuni ce blochează parțial porul fiind atribuite unor interacțiuni ce au loc între peptidele complexate și gura porului proteic (indicate prin &).
Analiza cantitativă a cineticii de interacţiune a peptidei amiloidice de origine umană cu porul proteic demonstrează că timpul mediu dintre două blocaje consecutive (τON), cauzate fie de peptida liberă fie de cea complexată cu ioni de cupru (Fig.VIII.4.1.3, panel (e)), precum şi durata medie a blocajelor τOFF, (Fig.VIII.4.1.3, panel (f)), cresc o dată cu adiţia unei concentraţii de cupru din ce în ce mai mare în partea trans a membranei.
Fig.VIII.4.1.3 Analiza statistică a timpilor dintre două evenimente consecutive a timpilor de rezidență a unei singure peptide uman-Aβ
1-16 ce
interacționeză reversibil cu un singur por de α-HL, măsurați la diferite concentrații de ioni de cupru adăugați în partea trans (i.e., 0, 25, 50, 75, 100, 150 și 200 μM).
24
Contrar celor observate în cazul peptidei amiloidice de origine umană (Fig.VIII.4.1.3), creşterea concentraţiei de ioni de cupru din soluţie adăugaţi în partea trans a membranei lipidice a dus la o frecvenţă mărită a evenimentelor de asociere dintre peptida amiloidică de origine murină şi porul proteic de α-hemolizină (Fig.VIII.4.1.5). Intuitiv, aceste rezultate nu ar avea sens deoarece ne aşteptam ca în urma formării legăturilor coordonative dintre situsurile de legătură ale peptidei şi ionii de cupru, conformaţia peptide amiloidice murine să se modifice astfel încât probabilitatea ca aceasta să intre în lumenul porului să scadă.
Fig.VIII.4.1.5 Înregistrări tipice ale curentului ionic ce reflect interacțiunia dintre peptida de provenineță murină cu porul de α-HL la o diferență de potențial de -100 mV, în absența (panel a) și în prezența ionilor de curpu adăugați în partea trans la diferite concentrații și anume (panel b: 100 µM, panel c: 400 µM). Scăderile de current ionic reflect evenimentele de asociere dintre o singură peptidă cu porul proteic. Analiza statistică a timpilor dintre două evenimente consecutive (τON, panel a) și a timpilor de rezidență (τOFF, panel b) a unei singure peptide murin-Aβ
1-16 ce
interacționeză reversibil cu un singur por de α-HL, măsurați la diferite concentrații de ioni de cupru adăugați în partea trans.
În continuare am propus o abordare matematică pentru estimarea valorii numerice a constantei de disociere (KD) ce descrie reacţia reversibilă dintre o peptidă amiloidică, fie ea de origine umană sau murină, şi un ion de cupru.
25
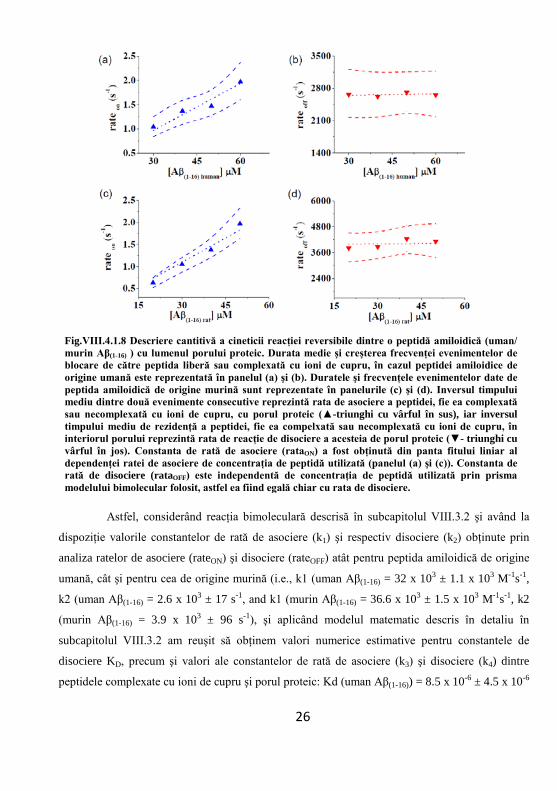
Fig.VIII.4.1.8 Descriere cantitivă a cineticii reacţiei reversibile dintre o peptidă amiloidică (uman/ murin Aβ(1-16) ) cu lumenul porului proteic. Durata medie şi creşterea frecvenţei evenimentelor de blocare de către peptida liberă sau complexată cu ioni de cupru, în cazul peptidei amiloidice de origine umană este reprezentată în panelul (a) şi (b). Duratele şi frecvenţele evenimentelor date de peptida amiloidică de origine murină sunt reprezentate în panelurile (c) şi (d). Inversul timpului mediu dintre două evenimente consecutive reprezintă rata de asociere a peptidei, fie ea complexată sau necomplexată cu ioni de cupru, cu porul proteic (▲-triunghi cu vârful în sus), iar inversul timpului mediu de rezidenţă a peptidei, fie ea compelxată sau necomplexată cu ioni de cupru, în interiorul porului reprezintă rata de reacţie de disociere a acesteia de porul proteic (▼- triunghi cu vârful în jos). Constanta de rată de asociere (rataON) a fost obţinută din panta fitului liniar al dependenţei ratei de asociere de concentraţia de peptidă utilizată (panelul (a) şi (c)). Constanta de rată de disociere (rataOFF) este independentă de concentraţia de peptidă utilizată prin prisma modelului bimolecular folosit, astfel ea fiind egală chiar cu rata de disociere.
Astfel, considerând reacţia bimoleculară descrisă în subcapitolul VIII.3.2 şi având la
dispoziţie valorile constantelor de rată de asociere (k1) şi respectiv disociere (k2) obţinute prin
analiza ratelor de asociere (rateON) şi disociere (rateOFF) atât pentru peptida amiloidică de origine
umană, cât şi pentru cea de origine murină (i.e., k1 (uman Aβ(1-16) = 32 x 103 ± 1.1 x 103 M-1s-1,
k2 (uman Aβ(1-16) = 2.6 x 103 ± 17 s-1, and k1 (murin Aβ(1-16) = 36.6 x 103 ± 1.5 x 103 M-1s-1, k2
(murin Aβ(1-16) = 3.9 x 103 ± 96 s-1), şi aplicând modelul matematic descris în detaliu în
subcapitolul VIII.3.2 am reuşit să obţinem valori numerice estimative pentru constantele de
disociere KD, precum şi valori ale constantelor de rată de asociere (k3) şi disociere (k4) dintre
peptidele complexate cu ioni de cupru şi porul proteic: Kd (uman Aβ(1-16)) = 8.5 x 10-6 ± 4.5 x 10-6
26
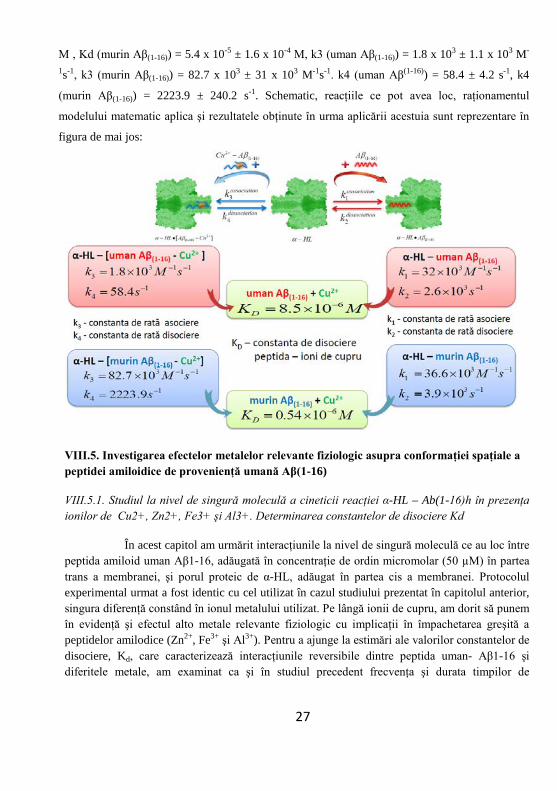
M , Kd (murin Aβ(1-16)) = 5.4 x 10-5 ± 1.6 x 10-4 M, k3 (uman Aβ(1-16)) = 1.8 x 103 ± 1.1 x 103 M-
1s-1, k3 (murin Aβ(1-16)) = 82.7 x 103 ± 31 x 103 M-1s-1. k4 (uman Aβ(1-16)) = 58.4 ± 4.2 s-1, k4
(murin Aβ(1-16)) = 2223.9 ± 240.2 s-1. Schematic, reacţiile ce pot avea loc, raţionamentul
modelului matematic aplica şi rezultatele obţinute în urma aplicării acestuia sunt reprezentare în
figura de mai jos:
VIII.5. Investigarea efectelor metalelor relevante fiziologic asupra conformaţiei spaţiale a peptidei amiloidice de provenienţă umană Aβ(1-16)
VIII.5.1. Studiul la nivel de singură moleculă a cineticii reacţiei α-HL – Ab(1-16)h în prezenţa ionilor de Cu2+, Zn2+, Fe3+ şi Al3+. Determinarea constantelor de disociere Kd
În acest capitol am urmărit interacţiunile la nivel de singură moleculă ce au loc între peptida amiloid uman Aβ1-16, adăugată în concentraţie de ordin micromolar (50 µM) în partea trans a membranei, şi porul proteic de α-HL, adăugat în partea cis a membranei. Protocolul experimental urmat a fost identic cu cel utilizat în cazul studiului prezentat în capitolul anterior, singura diferenţă constând în ionul metalului utilizat. Pe lângă ionii de cupru, am dorit să punem în evidenţă şi efectul alto metale relevante fiziologic cu implicaţii în împachetarea greşită a peptidelor amilodice (Zn2+, Fe3+ şi Al3+). Pentru a ajunge la estimări ale valorilor constantelor de disociere, Kd, care caracterizează interacţiunile reversibile dintre peptida uman- Aβ1-16 şi diferitele metale, am examinat ca şi în studiul precedent frecvenţa şi durata timpilor de
27

asociere/disociere dintre peptida liberă şi porul proteic de α-HL, la diferite concentraţii ale peptidei.
Fig.VIII.5.1.1 Înregistrări tipice ale curentului ionic la nivel de singură moleculă reprezentative pentru reacţiile reversibile dintre peptida uman Aβ1-16 şi porul proteic α-HL la aplicarea unei diferenţe de potenţial constante de ΔV = - 100 mV, în absenţa (panel a) şi prezenţa ionilor de Cu2+, Zn2+ , Fe3+ şi Al3+ adăugaţi în partea trans la o concentraţie de 200 µM.
Înregistrările arată că spre deosebire de Cu2+ şi Zn2+, prezenţa ionii de Al3+ şi Fe3+ presupune o schimbare mai puţin semnificativă în frecvența blocajelor cauzate de peptidă în interacțiunea cu porul α-HL. Aceast efect este corelat cu conformaţiile diferite pe care le adoptă peptida uman-Aβ1-16 în urma interacţiunii cu diferitele metale şi cu afinitatea diferită pe care o acestea faţă de peptidă.
Utilizând constantele de rată de reacţie obţinute din această analiză, şi fitând dependenţele timpilor de asociere/disociere de variaţia concentraţiei de metal după modelul matematic explicat în subcapitolul VIII.3.2, am obţinut valorile constantelor de disociere pentru ionii de Cu2+ şi respectiv Zn2+ cu peptidele uman- Aβ1-16: Kd(Cu2+) = 4.5x10-7 M ; Kd (Zn2+) = 9.2x 10-5 M. Se poate observa că o afinitate mai mare pentru peptidele uman- Aβ1-16 o prezintă ionii de cupru.
Concluzii:
Prezenţa moleculelor de phloretin determină eliberarea calceinei în urma formării porilor
transmembranari de metitină într-un procentaj mai mare decât în cazul absenţei acestora
în sistemele lipozomale;
28

Probabilitatea de inserţie a peptidei melitin în membrană şi formarea de pori
transmembranari este favorizată la tării ionice ale sării mai mici. Acest aspect a fost
observat atât pentru sistemele simple de lipozomi cât şi în cele cu phloretin.
Activitatea peptidei pep1 este mai mare decât în cazul peptidei pep2, fapt pus pe seama
costului energetic mai mare necesar inserţiei în miezul hidrofob al membranei lipidice a
peptidei ce conţine două puncte de ‚ancoarea’ (pep2) faţă de peptida ce prezintă doar un
punct de ‚ancoare’ la interfaţa bistratului cu mediul apos (pep1).
Diferenţa de ~ 0.2 kcal/mol a fost atribuită energii necesare inserării aminoacizilor
aromatici F1 şi W4 către partea trans a membranei, prezenţi în secvenţa primară a
peptidei pep2, poziţionaţi la capătul N-terminal, opus poziţiei aminoacidului W12, ce
rămâne ancorat în cazul ambelor peptide în partea cis a membranei.
Afinitiatea pentru membrana lipozomilor a peptidei cu cei mai mulţi aminoacizi
aromatici distribuiţi de ambele părţi ale α-helixului (pep2) este cea mai mare.
Deasemenea, pep1 având triptofanii W12 şi W15 orientaţi de aceeaşi partea a axului α-
helixului va avea o afinitate mai mare pentru membrane decât pep3, în care triptofanii
W12 şi W13 sunt orientaţi de o parte şi de alta a axului.
aminoacidul histindă este situsul de legătură principal şi pentru care ionii de cupru
prezintă cea mai mare afinitate, prezenţa acestuia în secvenţa primară a peptidei CAMA
fiind cauza principală a schimbării conformaţiei suferite de către aceasta.
datorită schimbărilor conformaţionale suferite de peptida CAMA în urma complexării cu
ionii de cupru, această întâmpină o barieră energetică mai mare în asocierea cu porul,
costul energetic asocierii fiind mai mare, dar o dată ce peptida complexată intră în por
interacţiunile ce au loc sunt favorabile energetic.
În prezenţa cuprului, ambele peptide prezintă o afinitate crescută pentru porul proteic de
α – hemolizină. Valorile obţinute pentru constantele de rată de asociere şi disociere
dintre por şi peptidele complexate, comparativ cu cele obţinute în cazul reacţiei por -
peptidă liberă, pot fi puse pe seama scăderii hidrofobicităţii locale în urma schimbării
conformaţiei peptidelor în urma formării legăturilor coordonative cu ionii de cupru.
Totuşi, având în vedere faptul că modificările conformaţionale induse de ionii de cupru
asupra peptidei de provenienţă umană produc un aranjament spaţial al acesteia astfel
încât să fie mai restricţionată trecerii prin por, componenta energetică ce priveşte
29

hidrofobicitatea locală crescută să fie determinantă pentru creşterea afinităţii acesteia
pentru por. Cu toate că există mulţi factori ce joacă un rol important în dezvoltarea bolii
Alzheimer, această scădere a hidrofobicităţii locale în cazul peptidei complexate cu ioni
de cupru, ar putea să explice, parţial, probabilitatea mai mică de agregare în prezenţa
cuprului a peptidelor amiloidice la şoareci.
Analiza statistică a blocajelor induse de o singură peptidă Aβ(1-16) asupra fluxul de
curent mediat de porul de α-HL indică următoare ordine în probabilitatea de a
interacționa cu peptida: Cu2+ > Zn2+ > Fe3+ > Al3+. Aceasta implică şi probabilitatea
de a induce schimbări conformaționale în structura peptidei amiloid.
Importanţa determinării constantei de asociere dintre peptida amiloid şi ionii metalelor
relevante prin metoda dezvoltată în această teză: această tehnică de determinare a constantei
de disociere, faţă de cele utilizate până acum (calorimetrie cu titrare izotermică, dicroism
circular, rezonanţă paramagnetică electronică sau spectrocopie RAMAN), care raportează
valori ale acesteia cuprinse între pM şi µM [6, 7, 8], unii raportând chiar valori de oridinul
attomolar [9], prezintă avantajul utilizării de concentraţii mici de peptidă pentru analizele
efectuate şi cel mai important asigură utilizatorul că rezultele obţinute sunt date de interacţiuni
dintre o singură peptidă, aflată în forma sa monomerică, şi ionii metalelor.
30

Bibliografie
[1] J.I. Kourie et al., Properties of cytotoxic peptide-formed ion channels, 2000, Am. J. Physiol. Cell Physiol. 278, 1063-1087
[2] M.S.P. Sansom et al., Ion channels formed by amphipathic helical peptides, 1991, Eur. Biophys. J. 20, 229-240
[3] H.Sato et al., Peptide-membrane interactions and mechanisms of membrane destruction by amphipathic α-helical antimicrobial peptides, 2006, BBA 9, 1245-1256
[4] D.W. Hoskin et al., Studies on anticancer activities of antimicrobial peptides, 2008, BBA – Biomembranes 1778, 357-375
[5] Clarke, J., Wu, H., Jayasinghe, L., Patel, A., Reid, S. and Bayley, H. Continuous base identification for single-molecule nanopore DNA sequencing. Nature Nanotechnology 4, 265-270 (2009).
[6] Garzon-Rodriguez, et al., Binding of Zn(II), Cu(II), and Fe(II) ions to Alzheimer's A beta peptide studied by fluorescence, Bioorg Med Chem Lett. 1999, 9(15), 2243-2248
[7] Ha C, Ryu J, Park CB., Metal ions differentially influence the aggregation and deposition of Alzheimer's beta-amyloid on a solid template, Biochemistry 2007, 46(20), 6118-6125
[8] Tõugu V, et al, Binding of zinc(II) and copper(II) to the full-length Alzheimer's amyloid-beta peptide, J Neurochem. 2008, 104(5), 1249-12459
[9] Atwood CS, et al., Dramatic aggregation of Alzheimer abeta by Cu(II) is induced by conditions representing physiological acidosis, J Biol Chem. 1998, 273(21), 12817-12826.
[10] Vicente M. Aguilella et al., Lipid charge regulation of non-specific biological ion channels, Phys. Chem. Chem. Phys., 2014,16, 3881-3893
[11] H.Sato et al., Peptide-membrane interactions and mechanisms of membrane destruction by amphipathic α-helical antimicrobial peptides, 2006, BBA 9, 1245-1256
[12] J. Antoinette Killian and Gunnar von Heijne, How proteins adapt to a membrane–water interface, TIBS 25 – SEPTEMBER 2000
[13] D.A. Dougherty, Cation-pi interactions in chemistry and biology: a new view of benzene, peh, tyr and trp, Science, New Series 1996, 271 (5246), 163-168
[14] Stephen H White and William C Wimley, Peptides in lipid bilayers: structural and thermodynamic basis for patitioning and folding, Current Opinion in structural biology 1994 4:79-86
[15] Gregg, E. C., Steidley, K. D., Kinetics of Cell Volume Changes of Murine Lymphoma Cells Subjected to Different Agents In Vitro, Biophys. J. 1965, 5, 393–405.
[16] DeBlois, R. W.; Bean, Counting and sizing of submicron particles by resistive pulse technique, C. P. ReV. Sci. Instrum. 1970, 41, 909–916.
[17] Bezrukov, S. M. J., Ion channels as molecular coulter counters to probe metabolite transport, Membr. Biol. 2000, 174, 1–13.
[18] Deamer, D.; Branton, D., Characterization of nucleic acids by nanopore analysis , Acc. Chem. Res. 2002, 35, 817–825.
31

[19] J. Clarke et al., Continuous base identification for single-molecule nanopore DNA sequencing, Nat. Nanotechnol. 2009, 4, 265–270.
[20] Q. Zhaoet al., Real-time monitoring of peptide cleavage using a nanopore probe , J. Am. Chem. Soc. 2009, 131, 6324–6325.
[21] H. Wanget al., Peering into Biological Nanopore: A Practical Technology to Single-Molecule Analysis, Chem. Asian J. 2010, 5, 1952–1961.
[22] David S. Talaga and JialiLi, Single-Molecule Protein Unfolding in Nanopores, J. Am. Chem. Soc. 2009, 131, 9287–9297
[23] L.Z. Song et al., Structure of staphylococcal alpha-hemolysin, a heptameric transmembrane pore., 1996, Science 274, 1859–1866
[24] S.-H. Shin et al, Kinetics of a reversible covalent-bond forming reaction observed at the single molecule level.,. 2002, Angew. Chem. Int. Edit. 41, 3707-3709
[25] O. Braha et al., H. Simultaneous stochastic sensing of divalent metal ions,2000, Nature Biotechnology 18, 1005-1007
[26] X. Guan et al., Stochastic sensing of TNT with a genetically engineered pore, 2005, ChemBioChem6, 1875-1881
[27] J.J. Kasianowicz et al., Characterization of individual polynucleotide molecules using a membrane channel., 1996, Proc. Natl. Acad. Sci. USA 93, 13770–13773
[28] A. Meller et al., Rapid nanopore discrimination between single polynucleotide molecules., 2000, Proc. Natl. Acad. Sci. USA 97, 1079-1084
[29] C. James et al., Continuous base identification for single-molecule nanopore DNA sequencing., 2009, Nature Nanotechnology 4, 265-270
[30] A.J. Wolfe et al., Catalyzing the translocation of polypeptides through attractive interactions., 2007, J. Am. Chem. Soc. 129, 14034-14041
[31] L. Movileanu et al., Interaction of peptides with a protein pore, Biophys. J. 2005, 89, 1030–1045
[32] Q. Zhao et al.,Study of Peptide Transport through Engineered Protein Channels, 2009, J. Phys. Chem. B113, 3572–3578
[33] Ayumi Hirano-Iwata et al., The design of molecular sensing interfaces with lipid bilazer assemblies, 2008, Trends in analytical chemistry 27(6), 512-520
[34] Danielsson, J., Pierattelli, R., Banci, L.& Graslund, A. (2007). High-resolution NMR studies of the zinc-binding site of the Alzheimer’s amyloid β-peptide, FEBS Journal, 274, pp. 46-59.
[35] Gaggelli,E., Janicka-Klos, A., Jankowska, E., Kozlowski, H., Migliorini, C., Molteni, E., Valensin, D., Valensin, G., & Wieczerzak, E. (2008). NMR Studies of the Zn2+ Interactions with Rat and Human β-Amyloid (1-28) Peptides in Water-Micelle Environment, Journal of Physical Chemistry B, 112, 100-109.
[36] M.A. Lovell, et al., Copper, iron and zinc in Alzheimer's disease senile plaques, J Neurol Sci. 1998, 158, 47–52
[37] E. Burstein et al., Decomposition of protein tryptophan fluorescence spectra into log-normal components. I. Decomposition algorithms, 2001, Biophys, J, 81, 1699 - 1709
[38] J.R. Lakowicz, Principle of fluorescence spectroscopy, 2006 3rd edition, Springer Scicence + Bussines Media, LLC, New York
[39] T. Ruysschaert et al., Liposome retention in size exclusion chromatography, 2005, BMC Biotechnology 1186/1472
[40] L. Silvestro et al., The Concentration-Dependent Membrane Activity of Cecropin A, 1997, Biochemistry 36, 11452-11460
32

[41] Eftink MR, Ghiron C. 1981. Fluorescence quenching studies with proteins. Anal Biochem 114:199–227.
[42] Eftink MR. 1991. Fluorescence quenching reactions: probing biological macromolecular structures. In Biophysical and biochemicalaspects of fluorescence spectroscopy, pp. 1–41. Ed TG Dewey. Plenum Press, New York.
[43] Eftink MR. 1991. Fluorescence quenching: theory and applications. In Topics in fluorescence spectroscopy, Vol. 2: Principles, pp. 53–126. Ed JR Lakowicz. Plenum Press, New York.
[44] M.V. Frassa et al., A. Structure and stability of the neurotoxin PV2 from the eggs of the apple snail Pomacea canaliculata, Biochim. Biophys. Acta 2010, 1804 (7), 1492–1499
33

ACTIVITATE ŞTIINŢIFICĂ:
1. Articole publicate in extenso în reviste științifice indexate ISI cu factor de impact
1. Alina Asandei, Sorana Iftemi, Loredana Mereuta, Irina Schiopu and Tudor Luchian, Probing of Various Physiologically Relevant Metals–Amyloid-β Peptide Interactions with a Lipid Membrane-Immobilized Protein Nanopore, 2014, The journal of membrane biology, 247 (6), pp. 523 - 530 Factor de impact: 2.478 Prim autor: nu, coautor
2. Alina Asandei*, Irina Schiopu*, Sorana Iftemi, Loredana Mereuta and Tudor Luchian, Investigation of Cu2+ Binding to Human and Rat Amyloid Fragments Aβ (1−16) with a Protein Nanopore, 2013, Langmuir, 29 (50), pp. 15634 – 15642 Factor de impact: 4.186 Prim autor: da, cu contribuţii egale cu CS III Dr. Alina Asandei
3. Loredana Mereuta*, Irina Schiopu*, Alina Asandei, Yoonkyung Park, Kyung-Soo Hahm, Tudor Luchian, Protein nanopore-based, single-molecule exploration of copper binding to an antimicrobial-derived, histidine-containing chimera peptide, 2012, Langmuir, 28 (49), pp. 17079 – 17091 Factor de impact: 4.186 Prim autor: da, cu contribuţii egale cu Lect. Univ. Dr. Loredana Mereuta
4. Irina Schiopu*, Loredana Mereuță*, Aurelia Apetrei, Yoonkyung Park, Kyung-Soo Hahm, Tudor Luchian, The role of tryptophan spatial arrangement for antimicrobial-derived, membrane-active peptides adsorption and activity, 2012, Molecular BioSystems, 8 (11), pp. 2860 - 2863 Factor de impact: 3.534 Prim autor: da, cu contribuţii egale cu Lect. Univ. Dr. Loredana Mereuta
2. Lucrări prezentate la conferințe internaționale
1. Irina Schiopu, Alina Asandei, Sorana Iftemi, Loredana Mereuta, Liliana Chiribasa,
Tudor Luchian, Single – molecule probing of Cu2+ induced folding on human versus rat amyloid Aβ (1-16) fragments, IC-ANMBES 2014, Brasov, Romania, 13 – 15 Iunie 2014 (prezentare poster)

2. Irina Şchiopu, Loredana Mereuţă, Alina Asandei, Tudor Luchian, Copper(II) binding to a histidine - containing chimera peptide: a single protein nanopore study, FEBS Workshop: Biological surfaces and interfaces, Sant Feliu de Guixols, Catalonia, Spania, 30 iunie – 5 iulie 2013 (prezentare poster).
3. Irina Șchiopu, Loredana Mereuță, Aurelia Apetrei, Tudor Luchian, Electrophysiology and fluorescence studies of the key role played by the position of aromatic amino acids in antimicrobial peptide activity and translocation, 9th ISCBPU, International Student Conference of Balkan Physical Union, Constanța, România, 5-7 iulie 2012 (prezentare orală).
4. Aurelia Apetrei, Irina Șchiopu, Loredana Mereuță, Tudor Luchian, Electrophysiology study of amyloid beta channel formation and activity in reconstructed planar lipid membranes, 8th BPU, The 8th General Conference of Balkan Physical Union, Constanța, România, 5-7 iulie 2012 (prezentare poster).
5. Irina Șchiopu, Loredana Mereuță, Aurelia Apetrei, Tudor Luchian, Tryptophan anchor position determines antimicrobial peptide activity and translocation, Third International Symposium on Antimicrobial Peptides – “Today knowledge and future applications”, Lille, Franța, 13-15 iunie 2012 (prezentare poster).
3. Lucrări prezentate la conferințe naționale
1. Liliana Chiribasa, Irina Şchiopu, Tudor Luchian, The modulatory effects exerted by electrical properties of lipid membranes and ionic strength upon peptides - biomimetic systems interactions, FTEM National Conference, Iaşi, 26 octombrie 2013 (prezentare orala)
2. Irina Şchiopu, Loredana Mereuţă, Alina Asandei, Tudor Luchian, Analysis of copper ion induced peptide folding through a nanopore sensing technique, 12th National Conference on Biophysics „CNB 2013” – Biophysics of Health, cu participare internaţională, Iaşi, 13-16 iunie 2013 (prezentare orală)
3. Liliana Chiribaşa, Irina Şchiopu, Tudor Luchian, Mellitin affinity enhancement
through a dipole potential modifying agent, 12th National Conference on Biophysics „CNB 2013” – Biophysics of Health, cu participare internaţională, Iaşi, 13-16 iunie 2013 (prezentare poster - Premiul pentru cel mai bun poster)
4. Liliana Chiribaşa, Irina Şchiopu, Tudor Luchian, Importanţa aminoacidului
Triptofan în studiul adsorbţiei peptidei antimicrobiene melitină, National Conference on Fundamental and Applied Research in Physics, Iaşi, 26 octombrie 2012 (prezentare poster).

5. Liliana Chiribaşa, Irina Şchiopu, Tudor Luchian, Tryptophan fluorescence study of
the adsorption of antimicrobial peptide Melitin, FTEM National Conference, Iaşi, 12 – 14 mai 2012 (prezentare orală – Premiul I)
6. Irina Schiopu, Aurelia Apetrei, Tudor Luchian, The modulatory role of cholesterol
on the activity of the antimicrobial peptide Cecropin B: a fluorescence approach, National Conference of Biophysics, Sibiu, 10-12 noiembrie 2011 (prezentare poster – Premiul pentru cel mai bun poster)
4. Granturi de cercetare
1. PN-II-ID-PCCE-2011-2-0027/01.06.2012 - BIOSENS: „Detecţia şi separarea ionică prin intermediul peptidelor ciclice, al ciclodextrinelor şi al porilor proteici”
2. PN-II-PT-PCCA-2011-3.1-0595 - BIOPEP: „Generarea şi investigarea unor noi
peptide antimicrobiene, cu dimensiune redusă. Corelarea structurii peptidelor cu funcţia lor”















![Conținutul spațiului informațional televizat din Republica ... · COMPORTAMENTE ELECTORALE. CU O EVALUARE A INFLUENȚEI RUSE ASUPRA OPȚIUNILOR GEO-POLITICE.] Studiul examinează](https://static.fdocumente.com/doc/165x107/5b2dc0627f8b9a94168b4cc7/coninutul-spaiului-informaional-televizat-din-republica-comportamente.jpg)

