I. MICROORGANISME PROCARIOTE … citoplasmatică Membrana citoplasmatică acoperă citoplasma...
92
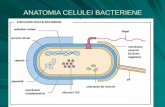
I. MICROORGANISME PROCARIOTE – ULTRASTRUCTURA CELULEI BACTERIENE Elementele structurale ale celulei bacteriene au fost clasificate în: elemente structurale constante (permanente) şi elemente structurale inconstante (nepermanente). Elemente structurale constante Peretele celular bacterian Celula bacteriană este delimitată de un perete celular bine definit structural, având o consistenţă rigidă. Peretele acoperă membrana citoplasmatică şi poate la rândul său să fie acoperit la unele specii de o capsulă sau de un strat mucos. Datorită indicelui de refracţie mic, peretele celular este invizibil, ori foarte greu vizibil, la celulele vii examinate la microscopul fotonic. El poate fi pus în evidenţă prin colorare selectivă sau prin examinare la microscopul electronic. La bacteriile tinere, care au citoplasmă abundentă, compactă şi omogenă peretele celular aderă intim la conţinutul celular subiacent. La celulele bătrâne însă, sau la cele în curs de liză, peretele celular devine evident datorită îndepărtării de citoplasmă. Examinat la microscopul electronic, peretele celular izolat are aspectul unui sac gol, care conservă forma bacteriei din care provine şi o constituţie asemănătoare ochiurilor unei plase. Grosimea peretelui celular variază între 15 – 35 nm, excepţional putând să ajungă la 80 nm. Este mai subţire la celulele tinere faţă de celulele bătrâne. Ultrastructura peretelui celular este diferită la cele două grupe mari de bacterii (clasificate în funcţie de afinităţile tinctoriale): bacterii Gram pozitive şi bacterii Gram negative. La bacteriile Gram pozitive peretele celular apare la microscopul electronic ca un strat unic, omogen. Componentul care se găseşte în cea mai mare parte în peretele celular al acestor bacterii (80 – 90 % din greutatea uscată a celulei) este peptidoglicanul sau mureina. În afara acestui component peretele celular mai conţine proteine, polizaharide şi acizi teichoici. Peptidoglicanul este un component parietal comun şi caracteristic tuturor bacteriilor, cu excepţia celor din genul Mycoplasma şi a bacteriilor halofile extreme. Din punct de vedere chimic peptidoglicanul este un heteropolimer compus dintr-o porţiune glicanică şi o porţiune peptidică. Porţiunea glicanică, foarte uniformă este formată din două N – acetil – hexozamine diferite: N – acetilglucozamina şi acidul N – acetilmuramic, legate între ele prin legături β – 1,4. Acizii teichoici (gr. teichos = zid, perete) sunt prezenţi numai în peretele celular al bacteriilor Gram pozitive, conferindu-le o serie de proprietăţi importante. La bacteriile Gram negative peretele celular este mai subţire, dar are o structură mai complexă. El este alcătuit din următoarele structuri: complexul peptidoglican – lipoproteine care reprezintă 2 – 10 % din greutatea peretelui celular şi are o compoziţie chimică foarte asemănătoare la toate bacteriile Gram negative; membrana externă a peretelui celular, cu o grosime de 6 – 20 nm, formată din fosfolipide, proteine şi lipopolizaharide; spaţiul periplasmic sau periplasmatic întâlnit numai la bacteriile Gram negative, cuprins între membrana citoplasmatică şi membrana externă a peretelui celular. În spaţiul periplasmic se depozitează un mare număr de enzime şi pigmenţi. Enzimele au rolul de a hidroliza substanţele nutritive pătrunse prin porii membranei externe, pregătindu-le pentru transportul activ prin membrana citoplasmatică. Semnificaţia biologică a peretelui celular. Peretele celular reprezintă sistemul de susţinere mecanică a întregii celule. Datorită rigidităţii sale determină forma caracteristică a celulelor bacteriene şi o apără de acţiunea factorilor nefavorabili din mediu. Asigură protecţia celulei faţă de şocul osmotic. Participă la procesul de creştere şi diviziune celulară, urmând membrana citoplasmatică în formarea septului transversal care separă celula mamă în cele două celule fiice. Datorită porilor, peretele celular permite pătrunderea în celulă a substanţelor nutritive şi eliminarea substanţelor rezultate din metabolism. Participă indirect la deplasarea bacteriilor ciliate, constituind suportul rigid pe care se sprijină cilii în contracţie. Conţine substanţele care dau specificitatea antigenică a celulelor şi receptori specifici pentru fixarea bacteriofagilor (virusuri care parazitează bacteriile).
Transcript of I. MICROORGANISME PROCARIOTE … citoplasmatică Membrana citoplasmatică acoperă citoplasma...

I. MICROORGANISME PROCARIOTE – ULTRASTRUCTURA CELULEI BACTERIENE
Elementele structurale ale celulei bacteriene au fost clasificate în: elemente structurale constante
(permanente) şi elemente structurale inconstante (nepermanente).
Elemente structurale constante
Peretele celular bacterian
Celula bacteriană este delimitată de un perete celular bine definit structural, având o consistenţă rigidă.
Peretele acoperă membrana citoplasmatică şi poate la rândul său să fie acoperit la unele specii de o capsulă
sau de un strat mucos. Datorită indicelui de refracţie mic, peretele celular este invizibil, ori foarte greu vizibil,
la celulele vii examinate la microscopul fotonic. El poate fi pus în evidenţă prin colorare selectivă sau prin
examinare la microscopul electronic. La bacteriile tinere, care au citoplasmă abundentă, compactă şi omogenă
peretele celular aderă intim la conţinutul celular subiacent. La celulele bătrâne însă, sau la cele în curs de liză,
peretele celular devine evident datorită îndepărtării de citoplasmă. Examinat la microscopul electronic,
peretele celular izolat are aspectul unui sac gol, care conservă forma bacteriei din care provine şi o constituţie
asemănătoare ochiurilor unei plase. Grosimea peretelui celular variază între 15 – 35 nm, excepţional putând
să ajungă la 80 nm. Este mai subţire la celulele tinere faţă de celulele bătrâne.
Ultrastructura peretelui celular este diferită la cele două grupe mari de bacterii (clasificate în funcţie
de afinităţile tinctoriale): bacterii Gram pozitive şi bacterii Gram negative.
La bacteriile Gram pozitive peretele celular apare la microscopul electronic ca un strat unic, omogen.
Componentul care se găseşte în cea mai mare parte în peretele celular al acestor bacterii (80 – 90 % din
greutatea uscată a celulei) este peptidoglicanul sau mureina. În afara acestui component peretele celular mai
conţine proteine, polizaharide şi acizi teichoici. Peptidoglicanul este un component parietal comun şi
caracteristic tuturor bacteriilor, cu excepţia celor din genul Mycoplasma şi a bacteriilor halofile extreme. Din
punct de vedere chimic peptidoglicanul este un heteropolimer compus dintr-o porţiune glicanică şi o porţiune
peptidică. Porţiunea glicanică, foarte uniformă este formată din două N – acetil – hexozamine diferite: N –
acetilglucozamina şi acidul N – acetilmuramic, legate între ele prin legături β – 1,4. Acizii teichoici (gr.
teichos = zid, perete) sunt prezenţi numai în peretele celular al bacteriilor Gram pozitive, conferindu-le o serie
de proprietăţi importante.
La bacteriile Gram negative peretele celular este mai subţire, dar are o structură mai complexă. El este
alcătuit din următoarele structuri: complexul peptidoglican – lipoproteine care reprezintă 2 – 10 % din
greutatea peretelui celular şi are o compoziţie chimică foarte asemănătoare la toate bacteriile Gram negative;
membrana externă a peretelui celular, cu o grosime de 6 – 20 nm, formată din fosfolipide, proteine şi
lipopolizaharide; spaţiul periplasmic sau periplasmatic întâlnit numai la bacteriile Gram negative, cuprins
între membrana citoplasmatică şi membrana externă a peretelui celular. În spaţiul periplasmic se depozitează
un mare număr de enzime şi pigmenţi. Enzimele au rolul de a hidroliza substanţele nutritive pătrunse prin
porii membranei externe, pregătindu-le pentru transportul activ prin membrana citoplasmatică. Semnificaţia
biologică a peretelui celular. Peretele celular reprezintă sistemul de susţinere mecanică a întregii celule.
Datorită rigidităţii sale determină forma caracteristică a celulelor bacteriene şi o apără de acţiunea
factorilor nefavorabili din mediu. Asigură protecţia celulei faţă de şocul osmotic. Participă la procesul de
creştere şi diviziune celulară, urmând membrana citoplasmatică în formarea septului transversal care separă
celula mamă în cele două celule fiice. Datorită porilor, peretele celular permite pătrunderea în celulă a
substanţelor nutritive şi eliminarea substanţelor rezultate din metabolism. Participă indirect la deplasarea
bacteriilor ciliate, constituind suportul rigid pe care se sprijină cilii în contracţie. Conţine substanţele care dau
specificitatea antigenică a celulelor şi receptori specifici pentru fixarea bacteriofagilor (virusuri care
parazitează bacteriile).

Membrana citoplasmatică
Membrana citoplasmatică acoperă citoplasma celulei bacteriene separând-o de suprafaţa internă a peretelui
celular de care este, de obicei, strâns lipită. Poate fi pusă în evidenţă şi la microscopul fotonic, după o
colorare selectivă cu albastru Victoria, sau fără colorare, la bacteriile vii examinate în câmp întunecat, caz
în care apare ca o linie netă, luminoasă, strălucitoare. La microscopul electronic, pe secţiuni ultrafine
membrana citoplasmatică prezintă o structură tristratificată, tipică tuturor membranelor biologice: două
foiţe opace la electroni ce limitează o foiţă internă electronotransparentă. Grosimea membranei
citoplasmatice este de 7,5 – 10 nm şi nu prezintă pori. Din punct de vedere chimic este formată din
proteine, lipide şi glucide. Membrana citoplasmatică este prevăzută cu numeroase invaginări de formă
veziculară, lamelară sau tubulară care pătrund în citoplasmă. Aceste invaginări sunt denumite mezozomi şi
împreună cu membrana citoplasmatică din care derivă formează sistemul membranar unic sau “unit
membrana”. Mezozomii au caracteristicile structurale ale membranei plasmatice din care derivă. Sunt mai
numeroşi şi mai bine dezvoltaţi la bacteriile Gram pozitive comparativ cu bacteriile Gram negative unde
sunt, în general, rudimentari, slab dezvoltaţi ca mărime şi mai puţini ca număr. Mezozomii sunt structuri
multifuncţionale. Au o deosebită plasticitate structurală ca răspuns la nevoile variate ale celulei. Formarea
lor reprezintă o modalitate prin care celula bacteriană îşi măreşte suprafaţa membranei citoplasmatice, în
funcţie de condiţiile de mediu. De aceea, se consideră că nu ar avea funcţii diferite faţă de cele ale
membranei plasmatice. Mezozomii sunt legaţi atât de membrana plasmatică, cât şi de materialul nuclear,
având rol important în controlul replicării cromozomului bacterian. Mezozomii sunt implicaţi şi în sinteza
membranei plasmatice, a peretelui celular şi a septului transversal care separă celulele după diviziune.
Semnificaţia biologică a membranei citoplasmatice
Membrana citoplasmatică reprezintă o componentă permanentă a celulei bacteriene, având rolul de a
delimita spaţiul intracelular. Pentru celula bacteriană membrana citoplasmatică reprezintă o barieră
osmotică semipermeabilă, asigurând o rapidă echilibrare a ionilor de sodiu între interiorul şi exteriorul
celulei, o concentrare intracelulară a ionilor de potasiu şi magneziu, precum şi a unor metaboliţi.
Membrana citoplasmatică este sediul a numeroase enzime respiratorii, reprezentând o importantă structură
cu rol energetic. Membrana citoplasmatică coordonează creşterea şi diviziunea celulară, iniţiind replicarea
cromozomului bacterian şi asigurând separarea cromozomilor fii, prin formarea septului transversal de
diviziune.
Materialul nuclear
Spre deosebire de celulele eucariote care au un nucleu cu structură bine definită, mărginită de o membrană,
conţinând un număr definit de cromozomi capabili de diviziune mitotică, materialul nuclear bacterian are o
formă primitivă de organizare, lipsit de membrană, situat în partea centrală a celulei şi care nu suferă
modificări de tip mitotic în cursul ciclului de diviziune. Datorită caracteristicilor sale particulare, această
structură a fost desemnată cu numeroşi termeni: nucleoid, nucleozom, material nuclear, nucleoplasmă,
cromozom bacterian etc. Din punct de vedere chimic materialul nuclear conţine 60% ADN, 30% ARN şi
10% proteine. Se pare că ARN-ul şi proteinele contribuie la conservarea unităţii structurale a
nucleoplasmei, în lipsa unei membrane proprii. Evidenţierea materialului nuclear bacterian la microscopul
fotonic nu se poate face prin colorare selectivă decât după îndepărtarea ARN-ului citopasmatic, realizată
prin hidroliza acidă sau enzimatică cu ribonuclează. Cu ajutorul acestor tehnici materialul nuclear apare
sub diferite forme: granule sferice sau ovalare, halteră, bastonaşe izolate sau dispuse în V etc.,
reprezentând 5 – 16% din volumul celulei. Pe micrografiile electronice se constată localizarea materialului
nuclear în partea centrală a celulei. El se prezintă ca o zonă clară, cu o densitate medie mai mică decât cea
a citoplasmei înconjurătoare. Pe secţiuni ultrafine s-a observat că regiunea nucleară este plină cu filamente
foarte fine, uneori aranjate în şiruri ondulate, paralele asemănătoare unei papiote de aţă, câteodată răsucită.
Aceste filamente sunt formate din ADN şi corespund cromozomului bacterian. În mod normal, bacteriile
aflate în faza de repaus, în culturi staţionare şi vechi au un singur cromozom, deci sunt uninucleate. În faza

de creştere activă, în culturi tinere, pe medii optime ele apar multinucleate, având 2 – 4 cromozomi,
identici genetic, deoarece provin prin replicarea cromozomului parental. De aceea, indiferent de aspectul
morfologic al materialului nuclear, din punct de vedere genetic bacteriile sunt haploide. Apariţia bacteriilor
multinucleate este rezultatul unei lipse de sincronizare între ritmul de creştere şi ritmul de diviziune
celulară. Bacteriile multinucleate apar şi sub influenţa unor condiţii de mediu. Astfel, sub acţiunea
penicilinei, care blochează sinteza mureinei din peretele celular, aceasta nu mai poate participa la formarea
septului transversal de diviziune şi apar astfel celule filamentoase sau globuloase cu mai mulţi nuclei.
Semnificaţia biologică a materialului nuclear
Cromozomul bacterian poartă în structura sa întreaga informaţie genetică necesară pentru biosinteză, creştere,
diviziune, reglarea activităţii celulare etc. Determină arhitectura celulei bacteriene, ereditatea şi capacitatea ei
de evoluţie. Datorită lipsei membranei proprii, un contact direct între ADN şi ribozomii din citoplasmă face
ca transferul informaţiei genetice să se facă foarte rapid.
Incluziunile citoplasmatice
Incluziunile sunt formaţiuni structurale ce cuprind o serie de substanţe inerte depuse în citoplasma celulei,
cu rol de substanţă de rezervă. După structura lor, evidenţiată la microscopul electronic, ele sunt de două
tipuri:
A - incluziuni înconjurate de membrană: incluziuni de poli β-hidroxibutirat, incluziuni cu sulf şi
carboxizomii;
B - incluziuni neînconjurate de membrane: incluziuni de polimeri anorganici, incluziuni polizaharidice,
rhapidozomii şi magnetozomii.
Incluziunile de poli β-hidroxibutirat sunt specifice numai procariotelor. Aceste incluziuni se observă uşor
datorită gradului mare de refringenţă sau pot fi puse în evidenţă după o colorare specifică pentru grăsimi
(negru de Sudan). În celulele vii apar ca granulaţii rotunde foarte refringente şi reprezintă un material de
rezervă tipic pentru procariote.
Incluziunile cu sulf apar sub forma unor corpusculi strălucitori, birefringenţi la două grupe ecologice de
bacterii: bacterii sulfuroase purpurii (genul Chromatium) şi la bacterii sulfuroase nepigmentate (genurile
Beggiatoa, Thiothrix). Granulele de sulf se găsesc în cantităţi mari la bacteriile care cresc în medii bogate
în hidrogen sulfurat şi reprezintă un depozit, deoarece dispar prin oxidarea sulfului, dacă aceste bacterii
sunt transferate într-un mediu sărac în hidrogen sulfurat.
Carboxizomii sunt incluziuni cu profil poliedric în număr variabil (1 – 200) prezenţi la bacteriile care
fixează CO2 pe calea pentozofosfaţilor. Sunt localizaţi în vecinătatea nucleoplasmei şi reprezintă locul de
depozitare a enzimelor cu activitate în fixarea CO2 .
Incluziunile de polimeri anorganici se mai numesc și incluziuni de volutină, denumire dată de Grimm
(1902) deoarece au fost găsite în număr mare la bacteria Spirillum volutans. Aceste incluziuni au fost
descoperite de către V. Babeş în 1897 la Corynebacterium diphtheriae şi Mycobacterium tuberculosis şi
apoi de Ernst în 1899 la alte bacterii, fapt pentru care mai sunt numiţi şi corpusculii Babeş – Ernst. Datorită
conţinutului mare în polifosfaţi anorganici produc efecte metacromatice (se colorează în roşu - violet cu
albastrul de metilen) motiv pentru care au fost denumite şi granulaţii metacromatice. Granulele de volutină
reprezintă o rezervă de fosfor şi energie intracelulară.
Incluziunile polizaharidice cuprind incluziunile de glicogen şi amidon. Acestea se întâlnesc ca
substanţe de rezervă la bacteriile aerobe sporulate. La microscopul fotonic pot fi observate după o
prealabilă tratare a celulelor cu iod în iodură de potasiu. În aceste condiţii incluziunile de glicogen se
colorează în brun roşcat, iar cele de amidon în albastru, în contrast cu citoplasma care apare colorată în
galben pal.

Rhapidozomii sunt particule ribonucleoproteice sub formă de bastonaşe scurte, rigide, intra- sau
extracelular, care pot fi eliminate din celule prin liza lor. Sunt întâlniţi la numeroase specii de mixobacterii,
la Proteus, Pseudomonas, Actinomyces. Conţin proteine şi ARN în proporţie de 2:1.
Magnetozomii sunt incluziuni care conţin fier şi au rol în orientarea şi migrarea unor bacterii sub influenţa
câmpurilor magnetice slabe. Se întâlnesc la unele bacterii magnetotactice din sedimentele marine sau de
apă dulce. Nu se găsesc dispersaţi în celulă ci formează una, două formaţiuni intracelulare alcătuite din 5 –
41 particule de formă cubică, cu colţurile rotunjite, situate în apropierea suprafeţei interne a membranei
plasmatice. Magnetozomii conţin fier sub formă de magnetită. Ei acţionează ca o busolă biomagnetică,
asigurând orientarea şi deplasarea bacteriilor în câmpurile magnetice.
Elemente structurale inconstante
Capsula şi stratul mucos
Unele bacterii au proprietatea de a elabora, în anumite condiţii de mediu, substanţe macromoleculare cu
caracter vâscos, gelatinos – mucos.
În funcţie de structura şi raporturile cu celula bacteriană substanţa mucoidă se poate prezenta sub
următoarele forme:
a) Microcapsula - la care substanţa mucoidă alcătuieşte un strat foarte fin în jurul celulei bacteriene,
având o grosime de 0,2 µm. Nu poate fi observată la microscopul fotonic, dar este detectabilă prin
metode imunologice.
b) Macrocapsula sau capsula adevărată - are o grosime ce depăşeşte 0,2 µm şi se prezintă ca o
formaţiune morfologic distinctă care înveleşte de jur – împrejur fiecare celulă sau perechi de
celule.
c) Stratul mucos - apare ca o masă amorfă, neorganizată în jurul celulei bacteriene.
d) Zooglea sau masa zoogleică - în care stratul mucos leagă între ele mai multe celule, formând
adevărate colonii mucilaginoase de bacterii.
Macrocapsula sau capsula adevărată apare la microscopul fotonic ca un halou incolor în jurul celulei
bacteriene. Dimensiunile ei variază foarte mult în raport cu corpul bacterian. Spre deosebire de stratul mucos
care formează în jurul bacteriilor o reţea laxă şi difuză de exopolizaharide, fără o structură definită, capsula
adevărată este strâns aderentă de peretele celular. Exemple de bacterii capsulate: Bacillus anthracis,
Streptococcus pneumoniae, Leuconostoc mesenteroides, Enterobacter aeruginosa, Klebsiella pneumoniae.
Capsula este greu de observat prin microscopie directă. Are o afinitate slabă pentru coloranţi şi de
aceea ea poate fi evidenţiată numai prin coloraţii speciale. După coloraţii uzuale ea apare ca un halou
incolor. Fiind puţin opacă pentru electroni capsula nu apare clar sau este invizibilă la microscopul
electronic.
Semnificaţia biologică a capsulei
Capsula este un produs inert care rezultă din activitatea metabolică a celulei bacteriene şi care poate
fi îndepărtat, celula bacteriană păstrându-şi viabilitatea. Capsula are un rol ecologic în supravieţuierea
bacteriilor în natură. Ea protejează celula bacteriană de efectul nociv al desicaţiei datorită proprietăţilor
sale higroscopice şi reprezintă un rezervor pentru stocarea substanţelor nutritive sau a celor rezultate din
metabolism. La bacteriile patogene, capsula are rol protector faţă de acţiunea fagocitelor. Bacteriile
patogene capsulate au o virulenţă mai mare pentru om şi animale. Astfel, pneumococul capsulat este
virulent, în timp ce tulpinile necapsulate sunt nevirulente şi fagocitabile.
Glicocalixul
Glicocalixul este alcătuit dintr-o masă de filamente polizaharidice ataşate de lipopolizaharidele de
pe suprafaţa bacteriilor, formând în ansamblu o structură ca o pâslă pe suprafaţa celulei, asigurând fixarea

acesteia de alte celule sau suporturi. Adeseori, diviziunea bacteriilor în interiorul glicocalixului poate
produce o microcolonie de celule acoperite de un glicocalix comun.
Semnificaţia biologică a glicocalixului
Glicocalixul reprezintă un rezervor de hrană pentru bacterii, prin legarea diferiţilor ioni şi molecule
care provin din mediul înconjurător sau sunt eliberate prin digestia enzimatică a celulelor moarte. Prezenţa
glicocalixului la bacteriile patogene reprezintă o protecţie pentru celulă, asigurând fixarea şi menţinerea ei
în mediile naturale sterile (ex. căile urinare), fără a fi îndepărtată, precum şi o rezistenţă mărită faţă de
bacteriofagi sau protozoare. În natură, în apele repezi curgătoare de munte, glicocalixul asigură fixarea
bacteriilor pe diferite substraturi inerte (pietre, resturi de lemn etc.) pe care formează depozite vâscoase.
Flagelii
Flagelii sunt apendici filamentoși dispuși la suprafaţa bacteriilor, reprezentând organite de locomoţie.
Posibilitatea de deplasare se întâlneşte numai la anumite specii bacteriene ca: bacili, vibrioni şi spirili.
Morfologie. Flagelii sunt structuri filamentoase subţiri, flexibile, de formă helicală. Lungimea lor este mult
mai mare decât a celulelor care îi poartă. De asemenea, ei sunt mai lungi la celulele bătrâne şi la cele care
trăiesc în medii lichide. Forma lor este cilindrică deşi, uneori, apar turtiţi ca o panglică. Numărul flagelilor
pe suprafaţa unei celule variază între 1 şi 100, fiind caracteristic pentru o anumită specie.
În funcţie de dispunerea flagelilor pe suprafaţa celulelor bacteriene acestea pot fi: atriche (bacterii imobile,
fără flageli - majoritatea bacteriilor); monotriche (cu un singur flagel polar, exemplu Vibrio cholerae);
lofotriche (cu un smoc de flageli situat la un pol al celulei, exemplu Spirillum marinus); amfitriche (cu
câte un flagel la ambii poli ai celulei, exemplu Bacillus megaterium); peritriche (cu flageli pe toată
suprafaţa celulei, exemplu Proteus vulgaris).
Ultrastructură. Un flagel este alcătuit din trei elemente principale: corpul bazal, articulaţia (cârligul) şi
filamentul helical extracelular.
La bacteriile Gram negative corpul bazal este alcătuit din patru discuri interconectate şi anume: discul
“M” dispus în membrana plasmatică şi montat fix pe axul de transmisie, dar care se roteşte liber în
membrana plasmatică; discul “S” localizat în regiunea supramembranară plasmatică (spaţiul periplasmic);
discul “P” inclavat în stratul peptidoglicanic; discul “L” legat de membrana externă lipopolizaharidică a
peretelui celular.
La bacteriile Gram pozitive structura corpului bazal este mai simplă, fiind format din două discuri
corespunzătoare discurilor “P” şi “M”.
Modul de deplasare. Filamentul flagelului pus în mişcare de “motorul rotativ” se învârte în jurul
propriului său ax, propulsând astfel celula. Când flagelii se rotesc în sens opus acelor de ceasornic aceştia
formează un fascicul în urma celulei care este propulsată în linie dreaptă. Când sensul rotirii se schimbă, în
sensul acelor de ceasornic, celula se “rostogoleşte” deoarece flagelii se răsfiră, fiecare trăgând celula în altă
direcţie. Deci, rotaţia în sens “antiorar” asigură deplasarea celulei în linie dreaptă, iar cea în sens “orar”
rostogolirea.
Pilii şi fimbriile
Sunt apendici filamentoşi, pericelulari prezenţi la bacteriile Gram negative (Escherichia coli, Proteus,
Klebsiella, Shigella, Salmonella) şi mai rar la bacteriile Gram pozitive. Pot fi evidenţiaţi numai prin
microscopie electronică. Sunt mai numeroşi decât flagelii, mai subţiri şi mai casanţi.
După structură, funcţie şi determinismul genetic aceşti apendici au fost grupaţi în două categorii: pili
de sex - apendici filamentoși care reprezintă receptori specifici pentru bacteriofagi şi calea de transfer a ADN-

ului bacterian şi plasmidial în procesul de conjugare; fimbriile - care nu sunt implicate în transferul ADN-
ului cromozomal, viral sau plasmidial.
Pilii de sex sunt în număr de 1 – 10 per celulă. Sunt codificaţi de informaţia genetică extracromozomală
(plasmida de sex). La microscopul electronic apar ca nişte structuri tubulare, cu un canal axial delimitat de
un perete alcătuit din molecule de pilină (o fosfoglicoproteină) asamblate după o simetrie helicală.
Semnificaţia biologică a pililor
Pilii sunt organite esenţiale pentru transferul ADN-ului cromozomal sau plasmidial în cursul
conjugării celulelor bacteriene mascule şi femele. Ei poartă receptori pentru fixarea bacteriofagilor de
celulele bacteriene şi servesc drept conduct prin care se realizează transferul genomului fagic în celula
bacteriană. Pierderea pililor prin mutaţie determină rezistenţa faţă de infecţia cu bacteriofagi.
Fimbriile sunt apendici filamentoși, rectilinii, dispuși pericelular, polar sau bipolar şi a căror sinteză este
controlată de gene cromozomale. Numărul lor variază între 1 – 1000 per celulă.
Semnificaţia biologică a fimbriilor
Creşterea suprafeţei de absorbţie a substanţelor nutritive. Creşterea activităţii respiratorii a bacteriilor.
Facilitarea aderării bacteriilor de diferite substraturi sau alte organisme. Servesc ca organite de transport a
unor metaboliţi. Contribuie la stabilizarea cuplurilor de conjugare. Influenţează virulenţa bacteriilor
patogene prin: fixarea bacteriilor patogene de celule şi ţesuturi, determinarea unei specificităţi de ţesut;
inducerea formării de anticorpi datorită antigenelor din fimbrii; conferirea unei rezistenţe deosebite a
bacteriilor la fagocitoză.
Sporul bacterian
La unele bacterii forma vegetativă se poate transforma într-un spor. Aceste bacterii se numesc sporogene.
La bacterii se întâlnesc mai multe tipuri de spori:
Endosporul (sporul propriu- zis) - este o formaţiune intracelulară foarte refringentă şi rezistentă la
factorii nefavorabili de mediu. Este caracteristic pentru genurile Clostridium şi Bacillus.
Exosporul – reprezintă o formaţiune rezistentă care se întâlneşte la bacteriile metanooxidante şi la unele
tulpini de Rhodomicrobium vannielii.
Artrosporul – constituie o formaţiune intracelulară mai puţin rezistentă decât endosporul, dar superioară
formei vegetative. Se formează prin fragmentarea celulei vegetative. A fost descris la Bacillus megaterium.
Chlamidosporul – este întâlnit la mixobacterii şi la Azotobacter chroococcum. Rezultă din îngroşarea
peretelui celulei vegetative şi acumularea unor substanţe de rezervă.
Gonidia – reprezintă o formaţiune intracelulară sferică rezultată din condensarea şi fragmentarea
conţinutului celular. Nu are calităţi specifice sporului adevărat dar are în schimb rol în înmulţire. A fost
descris la Leptothrix ochracea, Beggiatoa, Crenothrix.
Sporii de la actinomicete – sunt rezistenţi la uscăciune dar nu şi la temperatură. Au rol în reproducere.
Poartă diferite denumiri în funcţie de reprezentanţii unor familii şi anume: oidiospori, conidiospori,
sporangiospori.
Adevăratul spor bacterian sau endosporul este întâlnit la speciile genului Bacillus (aerob) şi Clostridium
(anaerob). La coci endosporul este întâlnit numai excepţional la Sporosarcina ureae. Datorită calităţilor
sale deosebite de rezistenţă endosporul conservă în stare dormantă, timp îndelungat, toate proprietăţile
genotipice ale celulei vegetative din care provine. Indiferent de morfologia celulei în care se constituie,
forma sporului (endosporului) este rotundă sau ovală, cu diametrul variind între 0,2 şi 2 µm. Poziţia
sporului în celulă este diferită şi reprezintă un criteriu taxonomic. Astfel, sporul ocupă o poziţie centrală la

Bacillus anthracis, subterminală la Bacillus cereus sau o poziţie terminală la Bacillus subtilis (la care
sporul poate avea și pozițiile centrală sau subterminală). Diametrul sporilor poate fi mai mic decât al
celulelor vegetative (ex. genul Bacillus) și se numesc spori nedeformanți. La unele specii, diametrul
sporului este mai mare decât cel al celulei în care se formează, deformând astfel celula. Sporii deformanţi
îi întâlnim la bacilii telurici, anaerobi, aparţinând genului Clostridium şi pot avea poziţie centrală dându-le
acestora aspect de fus (Clostridium pasteurianum), poziţie subterminală – aspect de rachetă de tenis
(Clostridium sporogenes) sau băţ de tobă dacă sporul este terminal (Clostridium tetani).
Ultrastructura
În general, ultrastructura sporului este, cu mici variaţii, aceeaşi la toate bacteriile. Microscopia electronică
a pus în evidenţă următoarele elemente structurale: tunicile (învelişurile sporale) – reprezentând trei
straturi suprapuse: unul extern (exina), unul intern (intina), între care se găseşte stratul mijlociu; toate sunt
electronoopace şi reprezintă 20 – 30 % din greutatea sporului; cortexul – o zonă electronotransparentă,
situată sub tunica internă, format dintr-un peptidoglican (altul decât mureina) şi acid dipicolinic;
membrana sporoplasmei – identică cu membrana citoplasmatică a celulei vegetative; sporoplasma
(inima sporului) – reprezintă 60 – 70% din greutatea sporului şi conţine elementele biologic active (ADN,
ARN, mai puţin ARN mesager, ribozomi). La unele specii (Bacillus cereus, Bacillus anthracis) la
suprafaţa tunicilor sporale se află un strat mucos, de formă neregulată numit exosporium. Sporul conţine,
în general, aceeaşi componenţi ca celula vegetativă dar în raporturi diferite. Astfel, comparativ cu celula,
sporul are un conţinut scăzut în apă, mai ales în apă liberă; este mai bogat în Ca2+
şi Mg2+
dar mai sărac în
K. Caracteristic sporului este prezenţa acidului dipicolinic, care sub forma dipicolinatului de calciu conferă
sporului termorezistenţă. Sporul este mai sărac în enzime decât celula vegetativă, iar enzimele existente
sunt inactive, motiv pentru care sporul este metabolic inactiv.
Semnificaţia biologică a sporului
Sporul reprezintă o etapă normală a ciclului de dezvoltare a bacteriilor sporogene, deoarece sporogeneza
necesită condiţii de mediu similare celor reclamate de celula vegetativă, uneori cu limite de variaţie mult
mai mici. El se deosebeşte de celula vegetativă prin mai multe caractere şi se comportă ca o formă de
rezistenţă ce păstrează intacte toate caracterele genotipice şi fenotipice ale speciei. Sporul bacterian nu
reprezintă o formă de multiplicare ca la fungi. Rezistenţa sporului bacterian la factorii nefavorabili de
mediu este deosebit de mare; pot rezista câteva ore la 1200C căldură umedă sau 200
0C căldură uscată.
II. STRUCTURA ȘI ULTRASTRUCTURA CELULELOR EUCARIOTE
1. Structura si ultrastructura organitelor specifice celulei vegetale (cloroplastul, peretele celular si
vacuola)
Cloroplastele sunt plastide care contin pigmentul verde numit clorofilă, cu ajutorul căruia realizează
fotosinteza, ceea ce conferă plantelor rol de producători primari. Clorofila este concentrată în mici granule
lenticulare numite grana, dispuse mai mult sau mai puţin regulat şi dând cloroplastului o structură granulară.
Ansamblul regiunilor clare, nepigmentate, ce separă grana se numeşte stromă (matrix).
Morfologie. La alge cloroplastele se numesc cromatofori; forma acestora variază şi este utilizată în
taxonomie pentru a diferenţia speciile, genurile şi grupele între ele: la Chlamydomonas şi Chlorella, alge
unicelulare, are aspect de cupă sau de clopot; la Ulothrix, algă pluricelulară, are aspect de inel ecuatorial, cu
marginile dantelate; la Cladophora, algă pluricelulară ramificată, este reticulat; la Spirogyra, algă
pluricelulară filamentoasă, are aspect de panglică spiralată. În ceea ce priveşte numărul de cromatofori /
celulă, acesta poate fi: între 1-6 la Spirogyra, 2 în formă de stea la Zygnema, mai multe zeci cu aspect
lenticular la Bryopsis.
Majoritatea cromatoforilor au una sau mai multe granule proteice numite pirenoizi în jurul cărora sunt
granule de amidon. Pirenoizii sunt corpusculi sferici, având 1-10 micrometri în diametru. La algele verzi şi la
unele alge roşii, pirenoizii sunt intraplastidiali, iar la algele brune, extraplastidiali, fixaţi în afara

cromatoforului cu ajutorul unui pedicel. Corpul central al pirenoidului este de natură lipoproteică şi se
numeşte pirenozom. La unele alge verzi, pirenoizii mai conţin ADN şi clorofilă .
La plantele superioare, cloroplastele sunt lenticulare, mari, vizibile la microscopul fotonic.
Dimensiunile lor variază cu specia, organul analizat, tipul de ţesut vegetal (la frunze, cloroplastele din celulele
ţesutului palisadic sunt de 2,5 ori mai mari decât cele din ţesutul lacunos).
Ultrastructură. Cloroplastul este limitat de o anvelopă (membrană dublă), ce închide o stromă
incoloră. Fiecare din cele două membrane simple are grosimea de 7,5 nm, între ele aflându-se un spaţiu clar
de 10 nm.
Membrana externă este netedă. Membrana internă emite invaginaţii, din care unele se prelungesc în
lamele stromatice, dispuse paralel cu axa mare a organitului. Între lamelele stromatice (stromatilacoizi) se
formează, pe alocuri, un anumit număr de saci mici, aplatizaţi, numiţi saculi granari (granatilacoizi) dispuşi în
teancuri (de la 2 până la 60 într-un teanc); aceştia provin din dilatări ale lamelelor stromatice, care se etalează
paralel cu aceste lamele şi de care se detasează. Ansamblul format de saculele granare şi porţiunile de lamele
stromatice care le mărginesc constituie aşa-numitul granum.
Tilacoizii cuprind: - unităţi pătratice de 15 nm, care apar pe cele două feţe ale membranei care conţin
aproximativ 300 de molecule de clorofile şi aproximativ 50 de molecule de pigmenţi carotenoizi, toate
înglobate prin polul lor hidrofob în pătura lipidică a membranei tilacoidale (fotosintetizantă); aceste unităţi
pătratice au fost asimilate cu unităţi funcţionale numite cuantozomi; - unităţi mai mici (8 nm), tot
transmembranare; - unităţi intermediare (9 nm) pedicelate, situate exclusiv pe faţa stromatică a membranei
tilacoidale, îndeosebi la extremităţile tilacoidelor (ele sunt factori de cuplaj clorofilieni, care servesc la sinteza
de ATP, ca şi în mitocondrie).
Stroma este incoloră, având o structură asemănătoare cu cea a hialoplasmei. Este de natură proteică,
conţinând picături lipidice şi câteva granule de amidon; acestea se formează la sfârşitul zilei, dar sunt
hidrolizate în cursul nopţii în glucoză, care migrează în plantă. În stromă s-au evidenţiat şi plastoribozomi,
ARN, ADN.
Peretele celular - Marea majoritate a celulelor vegetale sunt acoperite la exterior de un înveliş solid
numit perete celular, care separă celulele între ele, le protejează întreg conţinutul şi le conferă o anumită
formă stabilă. Aceste celule sunt cunoscute şi sub numele de dermatoplaste, prin contrast cu celulele animale,
numite nude sau gimnoplaste.
Peretele celular a fost prima componentă structurală observată la microscopul fotonic de către Robert
Hooke, în 1665.
Peretele celular este un produs al protoplastului şi rămâne permanent în contact cu citoplasma, prin
intermediul plasmalemei. Formarea sa începe la sfârşitul telofazei când se constituie un nou perete
intercelular în fragmoplast. În zona ecuatorială a fragmoplastului apar numeroase formaţiuni granulare mici,
ce conţin pectine şi care nu sunt altceva decât vezicule ale aparatului Golgi. Numărul şi mărimea acestora
cresc treptat, se apropie şi fuzionează într-o placă celulară (viitoarea lamelă mediană), care creşte centrifugal.
În cursul apropierii şi fuzionării veziculelor golgiene, membranele lor produc plasmalemele celor două celule
vecine. Între vezicule se află profile de RE, ce formează plasmodesmele, care trec prin punctuaţiile ce rămân
în lamela mediană. Ulterior se formează pereţii primari ai celulelor vecine. Noul perete se sudează cu pereţii
laterali ai celulei iniţiale şi separă astfel celula veche în două celule noi.
Peretele celulelor meristematice şi parenchimatice rămâne primar, celulozo-pectic tot timpul vieţii
acestora. În schimb, peretele celulelor ţesuturilor mecanice, conducătoare şi protectoare se îngroaşă prin
depuneri de material nou peste peretele primar, alcătuind peretele secundar. Spre deosebire de peretele
primar, care creşte mai mult în suprafaţă, peretele secundar creşte numai în grosime .
Compoziţia chimică a peretelui celular este foarte variată, în funcţie de grupa taxonomică.
Pereţii celulelor conţin 4 grupe de substanţe: glucide, lipide, proteine şi substanţe minerale. De regulă,
predomină glucidele, găsindu-se în special sub formă de polizaharide, cea mai frecventă fiind celuloza.
Alături de celuloză, în mod constant se găsesc hemiceluloze şi substanţe pectice.

Celuloza este un polimer liniar, neramificat, alcătuit din 5000-35000 molecule de D-glucoza.
Prin tehnica curentă de dublă colorare a preparatelor microscopice (verde iod şi carmin alaunat),
coloraţia roz a peretelui celular nu este specifică numai celulozei, ci şi hemicelulozei, calozei şi substanţelor
pectice.
Microscopia electronică a permis decelarea unităţilor de constituţie ale celulozei: formaţiunea
submicroscopică fundamentală este fibrila elementara. Fibrilele elementare se asociază în unităţi superioare
numite microfibrile. Prin asocirea a circa 250 microfibrile rezultă o fibrilă, iar prin unirea a 1 500 de fibrile
rezultă o fibră macroscopică.
Biodegradarea celulozei se realizează prin hidroliză catalizată de celulaz. Relativ puţine organisme
sintetizează celulază şi, deci, pot utiliza direct această biomasă disponibilă. Plantele verzi, care produc cea
mai mare cantitate de celuloză, nu au activitate celulolitică deosebită, fiind practic incapabile să catabolizeze
acest compus. Dintre animale, rumegătoarele, realizând o digestie simbiotică, reuşesc să catabolizeze
celuloza; activitatea de celuloliză este restrânsă practic la microorganisme: bacterii, protiste, ciuperci.
Pectinele constituie cea mai mare parte din lamela mediană, care menţine împreună celulele unui
ţesut. Locul de sinteză al pectinelor îl reprezintă aparatul Golgi. Se colorează specific în roz cu roşu ruteniu.
Plantele dispun de pectinaze care hidrolizează pectinele. Remanierea acestor polizaharide reprezintă un
proces natural frecvent, fie în cursul creşterii, fie la sfârşitul maturării celulei.
Hemicelulozele sunt macromolecule cu un grad mai scăzut de polimerizare, comparativ cu celuloza
Spre deosebire de celuloză, hemiceluloza poate fi utilizată de plantă ca substanţă de rezervă, deoarece conţine
o enzimă, hemicelulaza, care o poate hidroliza până la oze simple.
Ultrastructura peretelui celular. Se apreciază că primele straturi dinspre lamela mediană sunt mai
bogate în substanţe pectice şi hemicelulozice, cantitatea de celuloză crescând treptat spre interior. Cercetările
de microscopie electronică au evidenţiat că, in peretele primar, textura fibrilară are o orientare determinată;
fibrilele celulozice prezintă o dispoziţie paralelă pe faţa externă a peretelui şi aproape perpendiculară pe cea
internă, în raport cu axul longitudinal al celulei.
Peretele secundar conţine mai multă celuloză, iar în plus are şi substanţe de încrustaţie: cutina,
suberina, ceara, lignina, mucilagii, săruri, răşini. Este heterogen, fiecare din cele trei straturi ale sale (S1, S2,
S3) având structură proprie, situaţie vizibilă îndeosebi la vasele de lemn şi fibrele de sclerenchim.
Stratul S1 (extern) este gros de 0,12 – 0,35 micrometri (ocupă 5-11% din perete); este format din 4-6
lamele de microfibrile celulozice, având structură aparent reticulată.
Stratul S2 (mijlociu) ese cel mai gros (ocupă 74-84% din perete); are 30-150 de lamele foarte
apropiate, iar microfibrilele de celuloză au dispoziţie helicoidală, deci textura este paralelă, dar diferă direcţia
de înfăşurare în diferite lamele.
Stratul S3 (intern) este gros de 0,07-0,08 micrometri; are doar 2-3 lamele de microfibrile celulozice,
iar direcţia lor de înfăşurare în lamele alternează, deci textura este reticulată.
Peretele primar este plastic, extensibil şi, deci, permite alungirea organelor; peretele secundar este
elastic, dar nu plastic (el se formează atunci cand celulele, deci şi organele, au încetat să mai crească).
Modificările chimice secundare ale peretelui celular pot fi clasificate astfel:
1. apoziţii lipidice: cutinizare, cerificare, suberificare;
2. încrustaţii cu polifenoli: lignificare;
3. încrustaţii cu săruri minerale: mineralizare (silicificare, calcificare);
4. modificări cu rol degenarativ: gelificare şi lichefiere.
Cutinizarea şi cerificarea constau în impregnarea şi acoperirea pereţilor externi ai celulelor
epidermice cu substanţe de natură lipidică, numite cutina şi ceara. În general, celulele situate la suprafaţa
organelor aeriene au pereţii acoperiţi de o cuticulă, în care se disting ceruri (extractibile cu cloroform, benzen,
hexan) şi un polimer insolubil numit cutină.
Cuticula are un rol foarte important în echilibrul hidric al plantei, reducând evaporarea apei în
atmosferă; apariţia sa în cursul evoluţiei a fost esenţială pentru dezvoltarea plantelor în mediul aerian. În caz

de uscăciune, stomatele se închid şi atunci transpiraţia cuticulară devine determinantă pentru adaptarea la
viaţa în mediul uscat. Cuticula intervine, de asemenea, în reglarea umidităţii de la suprafaţa organelor,
eliminarea substanţelor volatile, apărarea împotriva factorilor externi (vânt, poluanţi, paraziţi); de cuticulă
depinde puterea de pătrundere a erbicidelor în plante.
Ceara şi cutina constituie complexul cuticular. Cuticula se colorează cu diferiţi reactivi (roşu Sudan
III, tetraoxid de osmiu). Grosimea sa variază în funcţie de condiţiile ecologice în care creşte planta: astfel,
plantele submerse au o cuticulă foarte subţire (uneori chiar poate lipsi), în timp ce xerofitele au cuticula foarte
groasă; la frunzele de măslin, laur, afin ea poate atinge 10-20 de micrometri.
Cerurile se află incluse fie în ochiurile polimerului de cutină (ceruri intracuticulare), fie se prezintă sub
forma unor proiecţii superficiale de forme variate – plăci, tubuli, bastonaşe, panglici – reprezentând
aproximativ 90% din ceara totală (ceruri epicuticulare).
În ceea ce priveşte biodegradarea cutinei: în natură, aceasta este reciclată de microflora solului
(bacterii, levuri); unele ciuperci saprofite pot fi cultivate pe substrat de cutină ca unică sursă carbonată.
Mamiferele posedă lipaze care degadează acest polimer.
Suberificarea constă în impregnarea pereţilor celulozici cu suberină (de natură lipidică). Procesul de
suberificare priveşte organe relativ în vârstă, adesea cu structură secundară; rareori, suberificarea reprezintă
un răspuns la răniri (când epiderma este distrusă), rezultând un suber de cicatrizare. În celulele de suber
adcrustarea de lipide are loc centripetal afectând toţi pereţii celulari, făcându-i impermeabili pentru lichide şi
gaze, împiedicând schimburile dintre celule şi mediul extern, conducând la moartea acestora. De aceea
funcţiile de izolare şi protecţie caracteristice suberului se realizează numai ce către pereţi atunci când celulele
mor.
Suberificarea poate fi totală la fag, salcie sau parţială la plop, arţar.
Lignificarea constă în impregnarea pereţilor celulozici cu lignină, compus de natură polifenolică, care
înconjoară microfibrilele de celuloză, umplând spaţiile dintre ele, mărind duritatea, rezistenţa mecanică şi
chimică a peretelui, micşorându-i flexibilitatea. Poate fi totală (în sclerenchimul târziu) sau parţială (în
xilemul timpuriu). Este un proces ireversibil, propriu cormofitelor, îndeosebi vaselor de lemn şi elementelor
de sclerenchim, rareori celulelor epidermice (ca la poacee).
Pereţii lignificaţi au o organizare ternară, cuprinzând un schelet microfibrilar de celuloză foate bine
organizat, o matrice amorfă predominant glucidică şi o reţea de polifenoli încrustaţi.
Între biopolimerii elaboraţi pe suprafaţa globului, ligninele ocupă locul al doilea după celuloză,
împreună reprezentând 60-80% din biomasa terestră. Celulele care işi lignifică pereţii celulari sunt decelabile
la microscopul fotonic prin utilizarea unor coloranţi specifici; caracteristice pentru lignificare sunt reacţiile cu
sulfat de anilină şi acid sulfuric, care conferă ligninei o culoare galben aurie (deci nuanţează culoarea naturală
a ligninei, care este cea galbenă), precum şi cea cu fluoroglucină şi acid clorhidric, care o colorează în roşu.
Lignina este foarte greu biodegradabilă; numai anumite bacterii şi câteva ciuperci (Polyporus) sunt
capabile să produca peroxidaze şi în prezenţa apei oxigenate să producă o lignoliză completă.
Mineralizarea este procesul de impregnare a pereţilor celulari cu diferite săruri minerale şi organice,
ceea ce le conferă rigiditate, soliditate şi rezistenţă. Substanţele încrustante sunt reprezentate mai cu seamă de
carbonat de calciu şi dioxid de siliciu.
Calcificarea - La plantele superioare, calcificarea pereţilor este localizată: astfel, în pereţii unor peri
protectori numiţi peri cistolitici (de la cânepă, hamei, mierea ursului) întâlnim concreţiuni amorfe care le
conferă o anumită rigiditate; în frunza de la Ficus există structuri numite cistoliţi, în care un repliu intern din
perete funcţionează ca loc de acumulare calcaroasă pe un schelet celulozic. Dizolvarea carbonatului de calciu
se poate realiza, pe o secţiune transversală prin frunză, cu ajutorul unei picaturi de acid.
Silicificarea poate fi întâlnită la algele unicelulare din grupa diatomeelor, peretele acestora fiind
constituit din două valve silicificate, inextensibile, foarte fin ornamentate. Tulpinile poaceelor şi ciperaceelor
au pereţii celulelor epidermice mineralizaţi,cu o cantitate apreciabilă de dioxid de siliciu. În cazul perilor de la
Urtica dioica, cea mai mare parte a peretelui este calcificată; doar vârful este silicificat, ceea ce determină

ruperea lui în punctele diferit mineralizate şi transformarea într-un minuscul vârf de ac de seringă, care se
adânceşte în piele, lăsând să difuzeze conţinutul celular urticant.
Gelificarea este procesul prin care compuşii pectici se îmbibă cu apă, se hidrolizează, transformându-
se într-o masa gelatinoasă. Lichefierea poate urma gelificării şi constă în dizolvarea şi dispariţia peretelui
celular. Ambele sunt procese de degradare controlată a peretelui celular (autoliză), frecvent întâlnite la
celulele ţesuturilor.
Vacuolele provin fie din dilatările locale ale membranelor RE, fie din diviziunea celor preexistente.
Sunt incluziuni “inerte” apoase, prezente în citoplasmă; ele nu efectuează transformări metabolice şi sunt
limitate de tonoplast. Pot fi colorate natural atunci când conţin antociani sau flavone, ori artificial cu coloranţi
selectivi de tipul roşului neutru şi albastrului de crezil.
În celulele tinere (meristematice) din apexurile radicular şi caulinar vacuolele sunt numeroase, dar
foarte mici; pe măsură ce procesul de diferenţiere avansează, vacuolele confluează, ajungând ca în celula
adultă să existe o singură vacuolă foarte mare, care ocupă circa 90% din volumul celular .
Conţinutul vacuolar este o soluţie de compoziţie complexă, a cărei natură chimică şi concentraţie sunt
foarte variabile în funcţie de specie, tip celular şi stare fiziologică. Compuşii încorporaţi în vacuole pot fi
grupaţi în două categorii:
1. compuşi intermediari ai metabolismului celular: acizii carboxilici răspunzători de pH-ul conţinutului
vacuolar; glucide (zaharoză la trestia de zahar şi sfeclă; inulină îndeosebi la asteracee); acizi aminici şi
proteine (care sunt transferaţi în vacuole dupa sinteză de către poliribozomii citoplasmici).
2. compuşi finali ai metabolismului celular: pigmenţii flavonoizi (exclusiv vegetali, semnale atractive
esenţiale pentru polenizatori); taninurile (inhibitori ai enzimelor; conferă astringenţă fructelor; oferă protecţie
organelor vegetale contra agenţilor patogeni); alcaloizii (cu proprietăţi farmacodinamice foarte variate).
Aparatul vacuolar al celulelor din embrion şi acela din celulele albumenului are un comportament
particular. În ovulele care se transformă în seminţe, celulele au vacuole mari, hidratate; în cursul maturării
seminţelor, vacuolele se deshidratează şi se fragmentează, în acelaşi timp acumulâdu-se o mare cantitate de
proteine. La sfârşitul evoluţiei lor, vacuolele vor conţine proteina precipitată (ne referim la granulele de
aleurona , care pot fi simple sau compuse, în ultimul caz proteina fiind organizată sub forma unui cristaloid
însoţit de o parte neproteică numită globoid).
Funcţiile vacuolei: - constituie pentru celulă principala sa rezervă de apă, compus indispensabil multiplelor
reacţii de biosinteză: - acumulează substanţe de rezervă sau deşeuri toxice; - contribuie la creşterea celulară; -
exercită presiune asupra peretelui celular extensibil şi elastic, care se întinde (starea de turgescenţă celulară,
care implică şi fenomene osmotice).
2. Structura si ultrastructura nucleului; diviziunea celulara (mitoza si meioza).
Nucleul - Este organitul descoperit pentru prima dată de R. Brown în 1831 (1835) în celulele petalelor de
orhidee. Ulterior a fost semnalat şi în celula animală. În 1838, M. Schleiden generalizează prezenţa nucleului
în toate celulele vegetale. Rolul nucleului în procesele de înmulţire, diferenţiere, cancerizare, sinteza
proteinelor citoplasmice i-a determinat pe citologi şi biochimişti să-i acorde o atenţie prioritară în cercetările
lor.
Morfologia nucleului interfazic. Nucleul eucariotelor este o achiziţie a evoluţiei; la procariote (virusuri,
bacterii, cianobacterii) întâlnim un nucleoid (substanţă nucleară, îndeosebi ARN şi ADN, fără existenţa unei
anvelope nucleare).
Număr: la eucariote, fiecare celulă are în general un singur nucleu; filamentele dicariotice ale unor
basidiomicete au câte doi nuclei; unele alge verzi (Caulerpa, Cladophora) au mai mulţi nuclei. Deosebim
astfel: plasmodii, în cazul cariocinezelor neurmate de citocineze şi sinciţii, ce rezultă în urma lizei pereţilor
despărţitori. Există şi celule anucleate: articulele tuburilor ciuruite, la care nucleul, devenit inutil în cursul
difereţierii celulare, a dispărut. În cazul elementelor moarte la maturitate (vasele de xilem, elementele de
sclerenchim, celulele de suber) nucleul a degenerat şi a dispărut o dată cu tot ce era viu în celulă.

Forma variază în funcţie de vârstă, forma şi funcţia celulei. În celulele tinere este sferic, iar în cele
diferenţiate este eliptic. Forme particulare: semilunară (celulele stomatice ale frunzei de Tradescantia);
fusiformă (parenchimul medular din tulpina de la Tilia); filamentoasă (celulele epidermice ale frunzei de
Aloe).
Poziţia în celulă poate fi: centrală (în celulele meristematice); parietală (în celulele diferenţiate);
parietală, dar numai lângă peretele extern (celulele epidermice de la frunza de Aloe); subterminală (în perii
absorbanţi radiculari).
Mărimea variază de la 0,5-3 micrometri la unele ciuperci şi alge, până la 1000 de micrometri în cazul
zigotului de Cycas revoluta; în medie, dimensiunile sunt cuprinse între 5 şi 15 micrometri.
Ultrastructură. Nucleul prezintă patru componente:
- anvelopa (membrana dublă);
- unul sau câţiva nucleoli sferici sau ovoizi;
- nucleoplasma, substanţa fundamentală optic omogenă ;
- cromatina, substanţă cu aspect variabil, dispersată în nucleoplasmă.
Anvelopa nucleară cuprinde două membrane simple (fiecare având grosimea de 7,5 nm), separate de
un spaţiu perinuclear clar, gros de 10-100 nm. Anvelopa prezintă pori nucleari, ce ocupă până la 25% din
suprafaţa ei, reprezentând deschideri prin care se pun în contact direct hialoplasma şi nucleoplasma.
Prin intermediul membranei externe, spaţiul perinuclear este în contact direct cu cavităţile RE (unii
autori consideră că membrana nucleară este o porţiune specializată a RE).
Membrana internă este dublată (în celula animală) pe faţa internă de o pătură groasă (60 nm) de
proteine, numită lamină fibroasă .
Mitoza (diviziunea ecvaţională)
Are loc la formarea celulelor somatice (diploide); plecând de la o celulă diploidă (2n) se ajunge la două
celule-fiice, tot diploide.
1. Cariocineza (mitoza în sens restrâns) cuprinde: interfaza, profaza, metafaza, anafaza, telofaza, toate
alcătuind ciclul celular, care durează 20-24 ore, uneori chiar câteva zile.
Interfaza este etapa în care se dublează numărul constituenţilor nucleari, mai ales ADN; cuprinde trei
perioade : G1 (perioada presintetică, cuprinde 25-50% din interfază), când fiecare filament cromatic are o
moleculă de ADN combinat cu histone; are loc sinteza de ARNm; cromozomii sunt monocromatidici; S
(perioada sintetică, cuprinde 35-40% din interfază), când continuă sinteza de ARNm; se dublează
cantitatea de ADN; cromozomii sunt bicromatidici; G2 (perioada postsintetică, cuprinde 10-40% din
interfază), cand continuă sinteza de ARNm; are loc sinteza proteinelor cu rol contractil, necesare formării
viitorului fus de diviziune.
Profaza: - se individualizează, prin spiralizare şi prin scurtare, cromozomii clivaţi în cele două cromatide
unite încă prin centromeri; - începe formarea fusului de diviziune (500-600 fibre proteice cu diametrul de
25 nm) pe seama citoplasmei perinucleare (întâi sub forma unor calote polare, ce se alungesc în formă de
conuri; în metafază ele ating polii celulei şi cu bazele se unesc la mijloc); - dispare nucleolul; - membrana
nucleară se fragmentează şi dispare; - citoplasma şi nucleoplasma se amestecă, rezultând mixoplasma, în
care sunt dispersaţi cromozomii.
Metafaza: - se încheie formarea fusului acromatic de diviziune; - cromozomii bicromatidici clivaţi se
dispun în planul ecuatorial al celulei, unde se inseră prin centromer de filamentele fusului de diviziune
(singurul nedivizat); - braţele cromozomilor sunt orientate spre polii celulei; - se formează astfel placa
celulară.
Anafaza: - are loc disocierea (separarea) cromatidelor, începând de la centromer (care s-a divizat), astfel
încât acum fiecare cromatidă este autonomă, individualizată, cu valoare de cromozom; - cromozomii fii
(cromatidele) migrează sincron spre polii fusului de diviziune, cu centromerul spre poli şi braţele spre
vechea placă ecuatorială; - cromozomii ating polii fusului, formând două grupe simetrice compacte, cu

braţele tasate unele lângă altele; - rol motor în migrarea cromozomilor îl are centromerul, dar ei se mişcă
datorită scurtării filamentelor fusoriale cromozomice.
Telofaza: - se formează cei doi nuclei fii, tot cu număr dublu de cromozomi, dar aceştia din urmă sunt
monocromatidici; - cromozomii se despiralizează, rezultând o nouă reţea cromatică; - ia naştere nucleolul,
lângă o zonă specială heterocromatică, numită nucleolară, a unor cromozomi cu satelit, organizatori de
nucleol; - se formeză membrana nucleară, din fragmente ale membranei iniţiale, care au fost antrenate
spre cei doi poli ai fusului de diviziune şi pe seama RE.
2. Citocineza începe la sfârşitul anafazei sau la începutul telofazei.
La cormofite, mici vezicule golgiene, bogate în glucide, protide şi lipide, se dispun la ecuatorul fusului
de diviziune, cresc, confluează (exceptând locurile ce corespund punctuaţiilor primare, pe unde trec
plasmodesmele) şi astfel se organizează fragmoplastul sau placa celulară. Fragmoplastul se întinde treptat
centrifugal spre peretele celulei iniţiale, intrată în diviziune, pentru a forma lamela mediană dintre cele
două celule fiice. În paralel, fusul de diviziune se dezorganizează, după ce şi filamentele sale participaseră
la formarea lamelei mediane. Fiecare din cele două citoplasme sintetizează câte un perete primar şi astfel
sunt separate cele două celule- fiice, în care s-au repartizat mai mult sau mai puţin egal diversele organite.
Meioza (diviziunea reducţională)
Fecundaţia şi meioza apar ca două mecanisme compensatoare: fecundaţia dublează numărul de
cromozomi, iar meioza îi reduce la jumătate. La covârşitoarea majoritate a plantelor, meioza precede
formarea sporilor.
Meioza cuprinde două diviziuni succesive: mitoza reducţională sau heterotipică (un nucleu 2n dă doi
nuclei n) şi mitoza ecvaţională sau homeotipică (cei doi nuclei n dau patru nuclei n). Aşadar, meioza are
loc la formarea celulelor de reproducere.
Mitoza reducţională
Profaza I durează câteva zile şi cuprinde mai multe subfaze:
- leptoten: cromozomii sunt filamente lungi, foarte fine, sinuoase; deşi s-au format cele două cromatide,
nu este vizibil clivajul;
- zigoten: cromozomii sunt tot filamente lungi, dar împerecheate; cromozomii omologi, paterni şi
materni, se apropie şi se unesc doi câte doi, fenomen numit sinapsă, ce conduce la formarea de bivalenţi;
- pachiten: cromozomii perechi sunt mai scurţi şi devin mai groşi prin spiralizare unul în jurul altuia;
clivajul apare vizibil la extremităţi;
- diploten: clivajul este vizibil, cromozomii din fiecare bivalent tind să se separe, dar cromatidele rămân
ataşate în mai multe porţiuni numite chiasme;
- diacineza: se accentuează spiralizarea, cromozomii devenind foarte scurţi şi groşi; cromozomii
omologi din fiecare bivalent se separă şi la nivelul chiasmelor are loc fenomenul de crossing-over (schimb
de gene); acum dispar nucleolul şi membrana nucleară, începe să se formeze fusul de diviziune.
Metafaza I: - se termină de format fusul de diviziune; - perechile de cromozomi (bivalenţii clivaţi) se
dispun în planul ecuatorial al celulei, cu centromerii de o parte şi de alta a planului ecuatorial.
Anafaza I: - disocierea anafazică separă cromozomi întregi (clivaţi în cele două cromatide) şi nu
cromatide ca în mitoza ecvaţională; - rezultă două loturi de cromozomi bicromatidici care migrează spre
poli.
Telofaza I: - cromozomii bicromatidici s-au regrupat la cei doi poli ai fusului de diviziune; - se formează
nucleolul şi anvelopa nucleară.
Uneori se formează peretele dintre cele două celule-fiice, deci are loc citocineza. Nu se mai observă
interfaza, deci nu are loc replicarea ADN.
Mitoza ecvaţională
Profaza II: - este scurtă;
Metafaza II: cromozomii se inseră cu centromerul la ecuatorul fusului de diviziune (cele două fuse se
dispun perpendicular pe fusul primei mitoze).

Anafaza II: disocierea anafazică separă cromatide, care migrează spre cei doi poli.
Telofaza II: - are loc regruparea cromozomilor monocromatidici la polii fusului de diviziune; - se
formează pereţi între celule, rezultând o tetradă de spori, mai rar gameţi; - împreună, cele două telofaze
reconstiuie patru nuclei haploizi; - se formează nucleolul şi anvelopa nuleară; - cantitatea totală de ADN /
nucleu este egală cu jumătate din cea a unui nucleu somatic la sfârşitul mitozei.
STRUCTURA ŞI ULTRASTRUCTURA CROMOSOMULUI EUCARIOT
Cromosomii fiecărei specii eucariote, de plante sau de animale, au particularităţi morfologice şi
numerice caracteristice speciei.
Reprezentarea schematicã a morfologiei şi organizăriii cromosomului la eucariote.
Pe lângă constricţia primară, determinantă a poziţiei centromerului, cromosomii au şi constricţii
secundare, cu rol în formarea nucleolului, fapt pentru care au primit şi denumirea de organizatori
nucleolari. Uneori, cromosomii au la unul din capete un segment numit satelit. Numărul nucleolilor dintr-un
nucleu este egal cu numărul cromosomilor cu satelit.
Centromerul (constricţia primară): - formaţiune de natură proteică, care permite ataşarea
cromosomilor la fibrele fusului de diviziune.
Regiunile telomerice: - sunt de natură proteică şi se formează prin activarea telomerazei în
primele stadii de dezvoltare;
- după formare, telomeraza se inactivează, iar regiunile telomerice se scurtează la fiecare ciclu
de diviziune.
Rolul regiunilor telomerice:
- împiedică alipirea cromosomilor în timpul diviziunii
- rol de protecţie enzimatică împotriva nucleazelor
- se presupune că intervine în vârsta celulei
Organizatorul nucleolar (constricţia secundară): - deţine informaţia pentru formarea nucleolilor. Structura internă a cromosomilor la eucariote.
Referitor la compoziţia chimică a cromosomilor eucariotelor, s-a stabilit că principala componentă o
constituie substanţa cromatică, denumită cromatină, alcătuită, la rândul ei, din acizi nucleici şi proteine. S-a
constatat că partea proteică este reprezentată de proteine histonice, notate cu H1, H2A, H2B, H3 şi H4. Aceste
cinci histone au secvenţă aminoacidică asemănătoare la majoritatea plantelor şi animalelor, ceea ce sugerează
telomer
satelitorganizator nucleolar
(constricţie secundară)
cromatide
centromer
(constricţie primară)
telomer
braţ scurt
braţ lung
Regiune
distală
Regiune
distală
Regiune
proximală
cromoneme

că gena care este responsabilă de biosinteza lor are o origine foarte îndepărtată şi nu a suferit prea multe
mutaţii, pe întregul parcurs al procesului filogenezei.
Patru dintre proteinele histonice (H2A, H2B, H3 şi H4) formează octameri, particule în jurul cărora se
înfăşoară ADN, care, împreună cu proteina H1 alcătuiesc nucleosomul. Nucleosomul este considerat,
structura de bază a cromosomului. Analizele cu raze X au dovedit că nucleosomii sunt legaţi între ei prin
ADN neasociat cu histone, fiind sub forma unei fibrile de aproximativ 10 nm grosime.
În celulele eucariotelor cu organizare superioară, nucleosomii conţin aproximativ 195 - 200 pb de ADN,
un octamer histonic alcătuit din câte două molecule de H2A, H2B, H3 şi H4 şi o moleculă de H1. De
asemenea, există şi un fragment de 27pb (perechi de baze), care nu este direct asociat cu octamerul histonic.
Acest fragment conectează, între ei, doi nucleosomi consecutivi. Lungimea ADN pliat în jurul octamerului
histonic este variabilă. În general, octamerul histonic are în jurul lui aproximativ 1,7 ture de ADN (adică 146
pb). Octamerul histonic împreună cu cele 146 perechi de nucleotide alcătuiesc miezul, sâmburele, particula
centrală a nucleosomului.
Astfel, în conformitate cu acest model, există un disc de 11nm diametru, care are 1,7 ture de acid
dezoxiribonucleic, înfăşurat în jurul lui. Cele 146 de pb ale acidului dezoxiribonucleic nu sunt repartizate
uniform în jurul octameruiui histonic. Tetramerii 2H3 şi 2H4 interacţionează cu macromolecula de ADN. S-a
demonstrat că partea centrală a octameruiui are histonele H3 şi H4, celelalte două histone fiind la exteriorul
lor (pe feţele discului).
Integritatea octameruiui este asigurată de interacţiunile puternice între moleculele de histone (H2A
interacţionează cu H2b şi cu H3, H2B cu H2A şi cu H4, H3 cu H4).
Nucleosomii împreună cu acidul dezoxiribonucleic linker alcătuiesc o fibră cromatică de 11 nm
grosime care se spiralizează, la rândul ei, dând un solenoid de 30 nm
grosime.
În solenoid nucleosomii sunt aranjaţi într-un helix de stânga, câte 6 pe tura
de helix, cu o grosime a spiralei solenoidului egalã cu diametrul
nucleosomului - adicã 11 nm.
Histona H1 este localizatã în sau lângã centrul solenoidului. ADN-
ul din componenţa solenoidului este, la rândul lui, constituit din douã pãrţi
- o parte ataşatã la miezul histonic şi o altã parte cu rol de linker, care
asambleazã nucleosomii în solenoid.
Cromosomii metafazici, care sunt eliberaţi de aproximativ 99% din
conţinutul lor histonic, îşi pãstreazã ADN-ul în formã aproximativ
constantã (asemenea cromosomilor iniţiali, intacţi), probabil tocmai din
cauzã cã ADN-ul este ataşat, în numeroase puncte, la scheletul proteic
nonhistonic intern. Acest schelet constã, în primul rând, din douã proteine,

desemnate prin Sc1 şi Sc2, cu greutãţi moleculare de 170 kD şi, respectiv, de 135 kD. Sc1 este
Topoizomeraza II, buclele radiale ale ADN-ului fiind ataşate la aceastã proteinã, în punctele numite SAR (în
englezã scaffolding attaching regions - regiuni de ataşare la schelet).
Eucromatina reprezintã materialul normal, izopicnotic, deţinãtorul informaţiei genetice, cu
comportament tipic în cazul diviziunii celulare (se spiralizeazã, se condenseazã, se decondenseazã şi se
coloreazã). La rândul ei, eucromatina este de douã tipuri: eucromatina activã şi eucromatina permisivã.
Eucromatina activã conţine genele ce vor fi transcrise în ARNm.
Eucromatina permisivã este reprezentatã de acea porţiune din eucromatinã care devine activã doar dupã
ce acceptã (permite) semnale declanşatoare (din categoria hormonilor, enzimelor etc.). Procesul autoreplicãrii
semiconservative a ADN-ului, în faza S din ciclul diviziunii celulare, începe la nivelul eucromatinei. În
consecinţã, replicarea eucromatinei este mult mai timpurie, în comparaţie cu cea a heterocromatinei.
Heterocromatina reprezintã materialul unor regiuni (uneori al unor întregi cromosomi) heteropicnotice,
caracterizate prin structurã densã şi compactã, inclusiv în telofazã, interfazã şi profaza timpurie. Din aceste
cauze, heterocromatina se coloreazã intens şi este vizibilã şi în interfaza ciclului celular. Heterocromatina
este componenta preferenţialã a cromosomilor sexuali şi a celor suplimentari (cromosomii B). Sunt
unele specii (broaştele ţestoase, unii viermi) la care întregul set cromosomial este heterocromatic.
TRANSCRIPŢIA ŞI TRANSLAŢIA INFORMAŢIEI GENETICE Transcripţia este procesul prin care regiuni din ADN (gene) sunt copiate în ARNm cu ajutorul enzimelor
ARN-polimeraze. Transcripţia mai este cunoscută şi sub numele de expresie a genelor. Toate cele trei tipuri
de ARN celular (ARNm, ARNr, ARNt) se sintetizează printr-un mecanism complex cunoscut sub numele de
transcrierea ARN, sau transcripţia ADN. Rolul principal în acest proces biosintetic îl joacă enzima ARN-
polimeraza-ADN-dependentă care necesită în calitate de substrat cei patru ribonucleosid-5'-trifosfaţi (ATP,
GTP, CTP, UTP), a ionilor de Mg2+
şi Mn2+
, precum şi prezenţa în calitate de template a ADN dublu-catenar.
Secvenţele de ARNm sunt în aşa fel prezentate, astfel încât sinteza să se desfăşoare de la stânga la
dreapta, în direcţia 5’ 3’
Copierea informaţiei de pe
catena antisens a ADN şi sinteza
ARNm
La eucariote există diferite
ARN-polimeraze pentru cele trei
tipuri de ARN transcrise, în timp
ce la procariote există o singură
ARN-polimerază care este
implicată în transcrierea celor
trei tipuri de ARN. Biosinteza
ARN sub acţiunea ARN-polimerazei-ADN-dependente se desfăşoară în patru etape: legarea matriţei, iniţierea
transcrierii, elongarea catenei ARN, terminarea transcrierii.
Prima etapă (legarea matriţei) constă în interacţiunea ARN-polimerazei cu ADN-ul matriceal, când se
formează un complex binar ce poate lega în continuare ribonucleosid-trifosfatul pentru a iniţia sinteza lanţului
polinucleotidic de ARN.
ARN-polimeraza se ataşează stabil la nivelul unei secvenţe specifice de ADN, secvenţă numită
promotor. Promotorii reprezintă fragmente cu până la 50 de dezoxiribonucleotide, localizate imediat înaintea
genei (la eucariote), sau înaintea grupului de gene (la procariote) a cărei informaţie genetică trebuie transcrisă
în ARN. Promotorul conţine mai multe secvenţe aflate la distanţe diferite de situsul start, care cuprind până la
6 nucleotide (GGCGGG – secvenţă numită şi GC box, TATAAT – TATA box).

După legarea la promotor, are loc desfacerea
(despiralizarea) dublului helix de ADN, formându-se aşa-
numita buclă de transcriere, sau complex deschis la
nivelul căruia punţile de hidrogen scindează.
A doua etapă: Din situsul start se iniţiază biosinteza
lanţului de ARN când ARN-polimeraza debutează prin
reacţia dintre ATP sau GTP cu o a doua moleculă de
ribonucleosid-trifosfat, rezultând un dinucleotid care mai
conţine un radical ortofosfat la capătul 3'. Succesiunea
bazelor azotate din promotor nu este transcrisă în catena
de ARN nou sintetizat.
Elongarea (a treia etapă) catenei de ARN se realizează
prin legarea succesivă a câte unui nou ribonucleotid la
gruparea OH liberă din poziţia 3' a dinucleotidului, respectiv a polinucleotidului precedent.
Formarea legăturilor fosfodiesterice între ribonucleotide se realizează numai în direcţia 5'3
', ceea ce
înseamnă că ARN-polimeraza începe să acţioneze la capătul 3' al catenei de ADN ce urmează a fi transcrisă.
Catena matricială (de ADN) formează cu catena de ARN aflată în
creştere un hibrid molecular ADN-ARN temporar, prin intermediul
punţilor de hidrogen ce se stabilesc între bazele azotate complementare.
Deci, catena de ARN ce se sintetizează este complementară din punctul
de vedere al structurii sale primare cu catena de ADN utilizată drept
matriţă.
Alungirea lanţului de ARN se continuă până când se ajunge la o altă
secvenţă specială de pe ADN, secvenţă formată din trei nucleotide,
numită semnal stop (a patra etapă – terminarea transcrierii). Semnalul
stop, numit şi semnal de terminare este reprezentat de un anumit bloc de
baze azotate perechi. În momentul în care ARN-polimeraza ajunge în
dreptul situsului stop, ea se desprinde de pe ADN, eliberându-se în
acelaşi timp şi molecula de ARN transcris.
Spre deosebire de replicarea ADN, care este semiconsevativă,
transcrierea acestuia este un proces complet conservativ.
Macromoleculele de ARN astfel sintetizate sunt apoi supuse
maturării post-trascripţionale.
Eucariotele au trei tipuri de ARN-polimeraze, fiecare
fiind responsabilă pentru transcrierea a diferite seturi de gene.
Aceste enzime au fost notate cu ARN-polimeraza I, II şi III.
ARN-polimerazele sunt molecule proteice mari, unele din
ele având molecula alcătuită din mai multe subunităţi. De
exemplu, ARN-polimeraza din Escherichia coli este o enzimă
care conţine în moleculă patru subunităţi, notate cu , , şi
, alături de ioni de zinc.

Deosebirea esenţială dintre cele trei ARN-polimeraze ale eucariotelor, constă în localizarea celulară şi în
funcţiile acestora:
- ARN-polimeraza I este localizată preponderent în nucleol şi catalizează biosinteza ARN ribosomal.
- ARN-polimeraza II este localizată în principal în nucleoplasmă şi este implicată în biosinteza ARN
mesager (ARN informaţional) ce va fi decodificat în procesul translaţiei informaţiei genetice.
- ARN-polimeraza III este localizată în nucleoplasmă şi participă la biosinteza ARN transportor, precum şi
la transcrierea moleculelor foarte mici de ARN 5S, molecule care intră în structura ribosomilor.
Prelucrarea ARN transcris primar
În procesul de transcripţie a eucariotelor, produşii sintetizaţi sunt reprezentaţi de ARNm precursor,
numit şi pre-ARNm, ARN premesager sau ARN heterogen, alături de precursorii ARNt şi ARNr.
Macromoleculele de ARN precursor, înainte de a fi exportate în citoplasmă, ca să devină funcţionale sunt
supuse la o serie de modificări: tăiere, ordonare şi unire, modificări cunoscute sub denumirea de matisare sau
splicing. Acest proces este necesar pentru ca intronii prezenţi în macromoleculele precursor să fie eliminaţi,
în aşa fel ca numai secvenţele exonice să fie unite, pentru a realiza macromoleculele de ARN matur.
Prima modificare constă în metilarea capătului 5' (capăt care este primul sintetizat în timpul transcripţiei)
reprezentată de o moleculă de 7-metil guanozin trifosfat. Acest proces se numeşte 5'-capping. Capătul 5
'-
metilat va avea ulterior, un rol foarte important în iniţierea sintezei proteice, precum şi de asigura protecţia
împotriva degradării macromoleculei de ARN care este transcrisă.
A doua modificare, are loc la nivelul capătului opus al lanţului ARN şi anume la capătul 3'. La această
extremitate a ARN, se ataşează o „coadă” poliadenilică. Acest lanţ poliadenilic se adaugă la ARN transcris,
imediat după ce ARN-polimeraza a depăşit regiunea terminator a genei transcrise. Lanţul poliadenilic este un
polimer format din 50 – 250 de nucleotide cu adenină (acid adenilic). Respectivul proces este realizat de către
o polimerază numită polimeraza poli-A.
A treia modificare constă în acoperirea întregului lanţ de ARNm cu un strat de proteine care îl protejează
de atacul endonucleazelor.
După realizarea acestor modificări, toţi intronii sunt îndepărtaţi, iar exonii se îmbină unii cu alţii.
Rezultatul este o macromoleculă de ARN mult mai scurtă, care conţine o secvenţă informaţională continuă.
Când şi această etapă (matisare, splicing) s-a încheiat, rezultă ARNm matur sau funcţional, care poate părăsi
nucleul pentru a iniţia translaţia.
Intronii care trebuiesc înlăturaţi din ARN transcris primar prin procesare, au mărimi variabile, cuprinse
între 80 şi 10 000 de nucleotide. Singurele secvenţe importante din structura intronilor sunt, cele implicate în
excizia lor prin ARN-procesare. Aceste secvenţe care acţionează ca markeri, numite secvenţe de consens sunt
situate strict la cele două capete ale fiecărui intron. Secvenţa specifică de la capătul 5' al intronului poartă
numele de situs donor, iar cea de la capătul 3' se numeşte situs acceptor.
În timpul procesării ARN transcris primar, are loc ruperea catenei de ARN la nivelul secvenţelor de
consens ale intronului, îndepărtarea întregului intron şi legarea la loc a catenei de ARN, respectiv a celor doi
exoni care erau despărţiţi iniţial de intronul excizat.
În nucleul celulelor există o serie de complexe macromoleculare formate din proteine şi ARN cu secvenţă
foarte scurtă, de aproximativ 250 de nucleotide. Aceste complexe, notate cu U1, U2, U3, ... U12, sunt denumite
ribonucleoproteine mici (snRNP- small nuclear ribonucleoprotein particles). S-a dovedit că snRNP au un
rol esenţial în procesarea ARN transcris primar.
În timpul procesării, moleculele de snRNP (U1 şi U2) se leagă la situsul donor şi respectiv la situsul
acceptor, apoi asociindu-se cu alte molecule de snRNP, formează complexe moleculare mari, numite
spliceosomi. Aceste particule de mărimea unui ribosom, produc excizia intronilor. Unul din rolurile ARN din
snRNP este de a recunoaşte complementaritatea dintre perechile de baze folosind secvenţele de nucleotide
care marchează începutul şi sfârşitul fiecărui intron (situsurile donor şi acceptor). După asamblarea
spliceosomului procesul de excizie a intronului decurge în două trepte:

- în prima etapă are loc ruperea lanţului ARN la nivelul situsului donor şi legarea capătului liber 5' la o
adenină din apropierea situsului acceptor, formându-se astfel un lasou (buclă, laţ);
- în etapa a doua are loc ruperea ARN la nivelul situsului acceptor şi legarea capetelor libere ale exonilor.
După terminarea procesării ARN, rezultă ARN matur care conţine informaţia genetică într-o secvenţă
neîntreruptă pentru sinteza unui lanţ polipeptidic.
Diferențe ale procesului de transcripție la procariote și eucariote
Procariote Eucariote
transcripţia are loc concomitent cu translaţia,
informaţia genetică fiind prezentă în
citoplasmă, adică acolo unde este şi utilizată
transcripţia este decalată în timp şi spaţiu
de translaţie, deoarece are loc în nucleu,
în timp ce proteinele sunt sintetizate în
citoplasmă
este realizată de o singură enzimă ARN-
polimerază
este realizată de trei ARN-polimeraze
diferite
macromoleculele de ARNm sintetizate nu
suferă modificări posttranscripţionale
sinteza unor precursori numiţi transcripţi
primari, care, posttranscripţional suferă
modificări importante
Asemănări ale procesului de transcripție la procariote și eucariote
Procariote şi Eucariote
copierea unei singure catene din segmentul de ADN care reprezintă gena
dezrăsucirea macromoleculei de ADN, pentru ca ARN-polimeraza să aibă acces la nucleotide, iar catena de
ADN să poată servi ca matriţă pentru sinteza ARN şi refacerea dublului helix pe măsură ce gena a fost
transcrisă
iniţierea şi stoparea transcripţiei la nivelul unor regiuni specifice recunoscute de ARN-polimerază
Translația
Proteinele sunt macromolecule liniare, alcătuite din aminoacizi. Secvenţa aminoacizilor din proteină –
numită structură primară – este determinată de structura unei gene specifice.
La baza procesului complex de biosinteză a proteinelor stă aşa-numitul principiu al sintezei
complementare pe (template) matriţă, principiu ce se mai întâlneşte atât la replicarea ADN cât şi la
biosinteza ARN. În procesul de biosinteză a proteinelor se foloseşte drept matriţă ARNm,
În sens strict, prin translaţia informaţiei genetice se înţelege traducerea secvenţei de nucleotide din
ARNm, într-o secvenţă de aminoacizi, acesta din urmă constituind structura primară a unei anumite proteine.
Procesul propriu-zis de biosinteză a proteinelor se realizează la nivelul ribosomilor în mai multe etape:
activarea aminoacizilor, iniţierea translaţiei, elongarea, terminarea translaţiei şi modificarea post-
translaţională a proteinelor.
Activarea aminoacizilor: Această etapă constă în faptul că, fiecare tip de aminoacid din citoplasmă
recunoaşte prin intermediul unei enzime specifice, un ARNt specific, alături de care formează un complex
activ de tipul aminoacil-ARNt. Această recunoaştere este înalt specifică în sensul că fiecărui aminoacid

proteinogen îi corespunde un anumit ARNt şi se realizează datorită înaltei specificităţi de substrat a enzimei
ce catalizează această reacţie, enzimă care se numeşte aminoacil-ARNt-sintetază.
Deci, codul genetic este translatat de două seturi de adaptori, care acţionează secvenţial şi cu mare
specificitate: aminoacil-ARNt-sintetaza care leagă un anumit aminoacid la ARNt corespunzător şi molecula
de ARNt care se ataşează prin anticodon, la codonul corespunzător din secvenţa ARNm.
2. Iniţierea translaţiei:
recunoaşterea semnalului de start din catena polinucleotidică a ARNm,
asocierea subunităţilor ribosomale, legarea complexului aminoacil - ARNt corespunzător la aceste subunităţi
formarea complexului ribosomal activ.
Complexele aminoacil-ARNt sintetizaţi în citoplasmă, sunt transportaţi spre ribosomi unde are loc
procesul propriu-zis de biosinteză al lanţurilor polipeptidice. În acest caz, ribosomii au rol de suport-
catalizator al biosintezei proteice.
Un ribosom conţine patru situsuri de legare pentru macromoleculele de ARN: unul pentru macromolecula
de ARNm şi trei numite A, P şi E pentru ARNt (A, de la aminoacil- ARNt; P, de la peptidil-ARNt şi E, de la
exit – ieşire).
ARNm se leagă de subunitatea mică (de 30 S la procariote, respectiv de 40 S la eucariote) printr-o
secvenţă pe care o conţine, secvenţă ce precede regiunea ce codifică proteina propriu-zisă, numită şi situs de
legare la ribosom (RBS – Ribosome Binding Sequence). La acest situs se formează un complex de iniţiere la
care se ataşează subunitatea mare. Subunitatea mare catalizează formarea legăturii peptidice între aminoacizi.
Ea conţine trei situsuri: A, P, E.
Situsul A, numit şi situs acceptor, sau aminoacilic, sau situs de intrare, ţine legată o macromoleculă de
ARNt care corespunde codonului ARNm ataşat de ribosom. Înainte ca aminoacil-ARNt să intre, acest situs
expune codonul care reprezintă următorul aminoacid ce trebuie să fie adăugat la lanţul polipeptidic.
Situsul P, numit şi situs donor, sau situs peptidilic, leagă primul aminoacil-ARNt în momentul iniţierii
sintezei proteinei, apoi va conţine lanţul polipeptidic în creştere.
Situsul E marchează locul de eliberare al ARNt din ribosom.
Situsul de legare al ARNm, situat pe subunitatea mică, asigură asocierea ARNm cu ribosomul în aşa fel
încât, doi codoni succesivi din ARNm să fie dispuşi exact în dreptul situsurilor A şi P.
Iniţierea biosintezei proteice debutează prin asocierea moleculei de ARNm la suprafaţa subunităţii mici a
ribosomului, într-un punct situat la aproximativ 10 nucleotide de capătul 5 al catenei, deoarece „citirea”
programului genetic stocat în ARNm se face în sensul 53.
Translaţia unui ARNm începe cu codonul AUG, iar pentru iniţierea translaţiei este necesar un ARNt
special, numit ARNt iniţiator. În cazul eucariotelor, ARNt iniţiator poartă întotdeauna o metionină, iar la
bacterii, ARNt iniţiator poartă formil-metionina. De aceea, lanţurile polipeptidice au întotdeauna metionina,
respectiv formil-metionina ca prim aminoacid al capătului aminoterminal.
Subunitatea ribosomală încărcată se leagă la capătul 5 al unui ARNm care este recunoscut datorită
acoperirii sale de către 7 metil-guanozin trifosfat. Subunitatea mică se mişcă în direcţia 53 de-a lungul
ARNm căutând prima secvenţă AUG (secvenţa START). Când a întâlnit AUG, factorii de iniţiere se
disociază de subunitatea ribosomală mică făcând loc subunităţii ribosomale mari, care se asamblează cu cea
mică formând ribosomul funcţional.
La bacterii, mecanismul pentru selectarea unui codon start este diferit. ARNm bacterian nu are un capăt
5 acoperit care să semnaleze ribosomului de unde să înceapă translaţia. El are însă secvenţe specifice pentru
legarea ribosomilor, secvenţe cu o lungime de şase nucleotide care sunt localizate cu câteva nucleotide
înaintea codonului AUG de la care începe translaţia.
Spre deosebire de ribosomul eucariot, ribosomul procariot se poate lega direct de un codon start din
interiorul ARNm, dacă acesta este precedat de câteva nucleotide faţă de situsul de legare al ribosomilor.
ARNm procariot este policistronic, adică deține informaţia necesară pentru sinteza mai multor proteine

diferite care pot rezulta din aceeaşi macromoleculă de ARNm. Spre deosebire de procariote, ARNm de la
eucariote poartă informaţia pentru sinteza unei singure proteine, deci este monocistronic.
3. Elongarea lanţului polipeptidic
Procesul elongării lanţului polipeptidic pe ribosom, poate fi considerat ca un ciclu în care se succed trei
etape (Figura 4):
1. În prima etapă, o moleculă de aminoacil-ARNt se leagă la situsul A liber de pe ribosom (adiacent
situsului P ocupat), prin formarea bazelor perechi dintre ARNt (anticodonul) şi cele din codonul de pe ARNm
expus în acest situs.
2. În etapa a doua, capătul carboxil (-COOH) al lanţului polipeptidic este decuplat de pe ARNt din situsul
P şi legat printr-o legătură peptidică la gruparea amino liberă a aminoacidului legat la ARNt din situsul A.
Reacţia catalizată de peptidil-transferază este însoţită de alunecarea subunităţii mari, faţă de subunitatea mică.
Prin această alunecare cei doi ARNt se deplasează în situsurile E şi P ale subunităţii mari.
3. În etapa a treia, subunitatea mică se deplasează exact cu trei nucleotide în lungul lanţului ARNm spre
capătul 3 şi se alătură la subunitatea mare, iar ARNt părăseşte situsul E. Deci, situsul A rămâne liber, legând
un nou aminoacil-ARNt corespunzător codonului ARNm expus în acest situs.
Acest ciclu în trei etape se repetă de fiecare dată când un aminoacid este adăugat în lanţul polipeptidic
care creşte de la capătul amino la capătul carboxil, până când întâlneşte un nou codon.
4. Terminarea sintezei lanţului polipeptidic
Sinteza unui lanţ polipeptidic este oprită printr-un semnal determinat de prezenţa codonilor STOP: UAA,
UAG, AGA. Aceştia nu pot fi recunoscuţi de nici un ARNt şi nu specifică un aminoacid. Rolul codonilor stop
este de a semnaliza ribosomului să oprească translaţia. Proteinele numite factori de eliberare se leagă de
codonul stop care ajunge în situsul A al ribosomului. La procariote sunt doi factori de eliberare, notaţi: EF1 şi
EF2. Factorul EF1 recunoaşte codonii UAA şi UAG, iar factorul EF2 recunoaşte codonii UAA şi UGA.
Celulele eucariote conţin doar un singur factor de terminare notat CRF care recunoaşte toţi cei trei codoni
stop.
Ribosomul eliberează ARNm şi se disociază în cele două subunităţi, care se pot asambla pe o altă
moleculă de ARNm pentru a începe un nou ciclu de sinteză proteică.
ARNm poate fi tradus simultan de mai mulţi ribosomi, atât la celulele eucariote, cât şi la cele procariote.
Odată ce un ribosom s-a deplasat de pe situsul de iniţiere, un alt ribosom se poate lega de ARNm şi poate
începe sinteza unui nou lanţ polipeptidic. În această manieră, un ARNm este tradus de o serie de ribosomi
aşezaţi la intervale de 100-200 de nucleotide. Grupul de ribosomi ataşaţi la o macromoleculă de ARNm este
numit poliribosom sau polisom. Fiecare ribosom din grup funcţionează independent pentru a sintetiza un lanţ
polipeptidic separat.
III. FOTOSINTEZA
Fotosineza este procesul fiziologic prin care plantele verzi îşi sintetizează cu ajutorul luminii şi cu
participarea pigmenţilor asimilatori substanţele organice proprii, folosind ca materii prime o sursă
anorganică de carbon (CO2 sau ionii bicarbonat), apă şi săruri minerale.
Procesul este însoţit, la plantele superioare de o degajare de O2, ca produs secundar; spre deosebire
de acstea, bacteriile fotoautotrofe realizează procesul de fotosinteză fără producere de O2.
Pentru a se realiza, procesul necesită trei condiţii principale: prezenţa în celule a pigmenţilor asimilatori;
existenţa în mediu a CO2; prezenţa luminii.
Fotosinteza are loc în celulele organelor verzi ale plantelor expuse la lumină, celule ce conţin un complex
de pigmenţi asimilatori, localizaţi în cloroplaste.
La algele unicelulare - fotosinteza este îndeplinită de însăşi celulele, ce reprezintă indivizii, iar la
algele cu tal masiv – (Laminaria) – numai primele 6-8 straturi de celule de la suprafaţa talului formează un
înveliş asimilator, cu feoplaste voluminoase, ovoidale.

La plantele superioare activitatea fotosintetică este îndeplinită predominant de către frunze, limbul
acestora prezentând adaptări ce favorizează desfăşurarea procesului: forma turtită a limbului oferă o
suprafaţă mare de contact cu mediul înconjurător; poziţia frunzelor favorizează prin aşezarea, în general
orizontală a limbului, absorbţia luminii necesară în fotosinteză; structura anatomică a frunzelor evidenţiază
la nivelul epidermei aparatul stomatic, prin care are loc difuziunea CO2 din aerul înconjurător în mezofil,
proces favorizat şi de prezenţa spaţiilor intercelulare între celulele ţesutului palisadic şi ţesutului lacunos.
Fotosinteza poate fi îndeplinită nu numai de limbul frunzelor, ci şi de celelalte părţi ale acestora: stipele,
peţiol, teacă, precum şi de alte organe - tulpini tinere nesuberificate, părţi ale învelişurilor florale - bractee şi
sepalele, precum şi fructe verzi în curs de creştere; este posibilă, de asemenea, o fotosinteză de intensitate redusă
şi la tulpinile mai vârstnice, sub stratul subţire de suber, precum şi în rădăcinile aeriene ale plantelor epifite, sub
stratul de velamen ce le acoperă, în cazul când acesta devine transparent, ca urmare a îmbibării cu apă.
Procesul fotosintezei are loc la toate plantele fotoautotrofe în organite speciale din celulele mezofilului
frunzei, numite cloroplaste. Numărul acestora la plantele superioare variază de la câteva zeci, până la câteva sute
în fiecare celulă, în dependenţă de condiţiile mediului de existenţă şi de tipul de ţesut; de exemplu, la una şi
aceeaşi plantă, în frunzele expuse la lumină puternică se formează mai multe cloroplaste (în ţesutul palisadic), iar
în cele expuse la lumina difuză mai puţine.
Suprafaţa totală a cloroplastelor în plantă este foarte mare: de exemplu, la o frunză de fag aceasta
depășește de 20 de ori suprafaţa frunzei, iar la un arbore de 100 de ani este estimată a fi de aproximativ 20.000
m2 (2 hectare), valori cu importanţă deosebit de mare legată de productivitatea procesului.
Cloroplastele se găsesc în celule într-o continuă mişcare, deplasându-se în hialoplasma celulară în funcţie
de intensitatea luminii printr-o cicloză continuă pasivă sau printr-o cicloză activă amoeboidală și orientându-se
spre sursa de lumină astfel încât să se ferească de supraîncălzire sau de fotooxidare.
Celulele asimilatoare conţin un ansamblu de pigmenţi care intervin în procesul de fotosinteză, cunoscuţi
sub denumirea de pigmenţi asimilatori, împărţiţi, pe baza particularităţilor lor structurale, în trei grupe: pigmenţi
clorofilieni, pigmenţi carotenoizi şi pigmenţi ficobilini. Dintre aceştia pigmenţii clorofilieni au o deosebită
importanţă în procesul fotosintezei, pigmenţii carotenoizi însoţesc pigmenţii clorofilieni în celulele asimilatoare
ale tuturor plantelor, iar cei ficobilini se întâlnesc împreună cu ceilalţi la algele albastre-verzi şi la algele roşii.
Din grupa pigmenţilor clorofilieni fac parte: clorofila a - în celulele asimilatoare ale tuturor platelor
fotosintetizante, clorofila b - la plantele superioare şi la algele verzi, clorofila c - la diatomee şi algele brune,
clorofila d - la unele alge roşii şi clorofila e - la algele galben-aurii. La bacteriile autotrofe fotosintetizante s-a
detectat bacterioviridina la bacteriile verzi, bacterioclorofila a la bacteriile purpurii şi verzi şi bacterioclorofila
b într-o tulpină a unei bacterii purpurii aparţinând genului Rhodopseudomonas.
Din punct de vedere chimic moleculele de clorofilă reprezintă gruparea prostetică a unor cromoproteine.
Caracteristica esenţială a lor este prezenţa în structură a unui nucleu tetrapirolic cu un atom de magneziu în
centru, legat de atomii de azot ai celor patru nuclei pirolici prin două valenţe chimice şi prin două legături fizice.
Pigmenţii asimilatori au rolul de a absorbi, conform cu particularităţile lor de absorbţie, radiaţiile de
lumină a căror energie este utilizată la sinteza substanţelor organice, în cursul procesului de fotosinteză.
Dintre toţi pigmenţii asimilatori ai plantelor fotoautotrofe un rol deosebit îl are clorofila a, care este
acceptorul final al energiei radiaţiilor absorbite de ceilalţi pigmenţi şi care este capabilă să absoarbă, ea însăşi,
anumite radiaţii ale spectrului solar. În timpul fotosintezei moleculele de clorofila a, ajunse în stare de excitaţie,
revin la starea fundamentală eliminând un electron cu un potenţial energetic ridicat şi acceptând un alt electron,
cu un potenţial energetic normal. În consecinţă, în ansamblul reacţiilor ce caracterizează mecanismul fotosintezei
au loc procese de oxidoreducere, în cadrul cărora moleculele de clorofila a transferă electroni unei substanţe ce
se reduce şi primesc alţi electroni de la substanţă ce se oxidează. Prin acest transfer are loc transformarea energiei
luminii absorbite în energie chimică, deci reacţia fotochimică a fotosintezei.
În frunzele bătrâne sau în condiţii nefavorabile de viaţă (secetă, exces de umiditate, insolaţie puternică,
exces de substanţe toxice, temperaturi scăzute) are loc degradarea peristromei cloroplastului, dezorganizarea
stromei, granelor şi, în final, descompunerea moleculelor de clorofilă, până la dispariţia culorii verzi. Se formează

în această situaţie epiporfirina - pigment roşu, feofitina - brun, pigmenţii carotenoizi - mai rezistenţi la
degradare şi pigmenţii antocianici; ca urmare, apar coloraţii specifice frunzelor în perioada de toamnă, cu nuanţe
de galben, portocaliu, brun roşietic şi albăstrui, funcţie de raportul dintre aceste componente.
MECANISMUL FOTOSINTEZEI I. Faza de lumină a fotosintezei
FAZA I - a, luminoasă, sau reacţia de lumină, faza HILL - constă dintr-un ansamblu de reacţii în care
esenţială este reacţia fotochimică, pe baza ei se realizându-se transformarea energiei luminoase în energie
chimică, înmagazinată în substanţele organice produse în timpul fotosintezei. Aceste procese se desfăşoară în
granele cloroplastelor şi în prezenţa pigmenţilor asimilatori, cuprinzând următoarele procese: captarea CO2 în
cloroplastele celulelor asimilatoare, stimularea clorofilei pentru fixarea energiei luminoase, fosforilare ADP în
ATP, fotoliza apei cu fixarea de H+ în NADP redus şi eliminarea O2. Toate aceste procese necesită în mod
obligatoriu prezenţa luminii şi sunt grupate în trei etape distincte: pătrunderea CO2 în cloroplaste; absorbţia
energiei luminoase; transformarea energiei luminoase în energie chimică.
● Pătrunderea CO2 în cloroplaste
În această etapă aerul din mediul înconjurător pătrunde prin ostiolele stomatelor, deschise prin reacţie
fotoactivă, trecând în camera substomatică, de unde difuzează în spaţiile intercelulare, și ajunge în contact direct
cu pereţii celulozici ai celulelor asimilatoare din ţesutul palisadic şi lacunos. Dacă pereţii celulelor asimilatoare
sunt umezi, permanent irigaţi cu apa absorbită din sol, CO2 din aerul circulant în spaţiile intercelulare, cu o mare
capacitate de hidrosolubilizare, se dizolvă în apă şi trece în acid carbonic, care - după disociere - dă formele
ionice HCO3- şi CO3
-, care difuzează în citoplasma celulară, ajungând până la cloroplaste.
Noaptea, când stomatele sunt închise prin reacţie fotoactivă, precum şi în perioadele de secetă, când
stomatele sunt închise prin reacţie hidroactivă, pereţii celulelor din mezofilul frunzei sunt uscaţi, fotosinteza este
blocată, iar creşterea plantelor stagnează, din lipsa asimilatelor nou sintetizate.
● Absorbţia energiei luminoase
Constă din captarea luminii de către pigmenţii asimilatori sub formă de fotoni. Intensitatea acestui proces
fotofizic este direct proporţională cu numărul de fotoni absorbiţi, ceea ce face ca necesarul de lumină pentru
fotosinteză să se exprime prin numărul de cuante luminoase/moleculă.
În grana cloroplastelor clorofila a există sub două forme: una cu maximum de absorbţie la 700 nm care,
alături de un alt pigment accesoriu formează sistemul fotochimic I şi o altă formă, cu maximum de absorbţie la
690 nm care, alături de clorofila b, pigmenţii carotenoizi şi de pigmenţii ficobilini la algele roşii - formează
sistemul fotochimic II. Fiecare din aceste sisteme fotochimice posedă câte un centru de reacţie ce funcţionează
ca o pompă de electroni, întrucât moleculele de clorofilă a constituite în centrul de reacţie emit electroni de
înaltă energie când ajung în starea de excitaţie şi revin la starea fundamentală, acceptând electroni cu potenţial
energetic normal.
Cea mai mare parte a moleculelor de clorofilă a din aceste fotosisteme, precum şi celelalte molecule de
pigmenţi asimilatori au un rol comparabil cu acela al unei antene, care constă în captarea radiaţiilor de lumină şi
în transferarea energiei acestora moleculelor din centrii de reacţie. În captarea energiei luminoase rolul
principal îl prezintă clorofila a690 din fotosistemul II de pigmenţi, care acţionează ca un sensibilizator optic şi
participă direct la captarea cuantelor de lumină, îndeosebi a fotonilor roşii şi albaştri. Energia luminoasă fixată de
clorofila a690 este transferată pe moleculele de clorofilă a700 într-un timp foarte scurt (10-8
- 10-9
secunde). Acest
transfer de energie este posibil numai atunci când distanţa între moleculele de clorofilă este sub 50-100 Å,
condiţie realizată în cuantozomi unităţi elementare funcţionale ale cloroplastelor, ce răspund de absorbţia şi de
transformarea energiei luminoase. Se consideră că în acest proces 200 molecule de clorofila a690 aranjate într-un
fascicul au funcţia asemănătoare unei lentile ce captează fotonii şi îi concentrează într-un punct focal, respectiv
pe molecula de clorofilă a700, cu rol de depozit primar al energiei luminoase.
Pigmenţii din fotosistemul II de pigmenţi participă indirect în această etapă fiziologică, prin captarea unei
cantităţi reduse de fotoni, pe care îi transferă pe clorofila a690 prin inducţie moleculară sau prin fenomene de
rezonanţă.
● Transformarea energiei luminoase în energie chimică

Cuprinde trei subetape: fotofosforilarea aciclică, fotofosforilarea ciclică şi fotoliza apei.
Prin procesele de fosforilare plantele verzi înmagazinează o parte din energia liberă ce provine din
activitatea metabolică proprie sau de la soare şi conservă un anumit potenţial energetic sub formă de molecule
stabile, bogate în energie, moleculele de ATP, care este utilizat, după hidroliză, în satisfacerea nevoilor
termodinamice ale reacţiilor endergonice din metabolism. În acest proces ATP-ul este într-o continuă sinteză şi
hidroliză.
La plantele verzi fotoautotrofe există, pe lângă fosforilarea oxidativă, procese de sinteză a ATP-ului prin
fosforilare fotosintetică sau fotofosforilare.
În cazul sistemului fotochimic II, reducerea moleculelor de clorofilă a din centrul de reacţie se face prin
intermediul unei substanţe necunoscute care conţine Mn şi care ia electroni de la moleculele de apă ce participă
ca materie primă în fotosinteză, transformându-se în protoni de hidrogen şi în oxigen. În cazul sistemului
fotochimic I rolul de reducător al moleculelor de clorofilă a din centrul de reacţie este îndeplinit de citocromul f,
iar cel de oxidant de către ferredoxină - o proteină ce conţine fier.
Cele două sisteme fotochimice a căror centri de reacţie funcţionează ca o pompă de electroni sunt legate
în serie şi nu în paralel, electronii emişi de sistemul fotochimic II fiind preluaţi de plastochinonă şi transmişi,
prin intermediul unui lanţ de citocromi (citocromul b6 şi citocromul f) la sistemul fotochimic I, ale cărui
molecule de clorofilă a din centrul de reacţie sunt readuse astfel la starea stabilă. Electronii emişi de sistemul
fotochimic I sunt captaţi de ferredoxină şi sunt transferaţi prin intermediul unei flavoproteine, împreună cu
hidrogenul rezultat în urma sciziunii moleculelor de apă prin intervenţia sistemului fotochimic II, la
nicotinamid-adenin dinucleotid-fosfat, forma redusă (NADPH) + H+.
Deci, în cursul fazei de lumină, electronii furnizaţi de apă sunt excitaţi succesiv de radiaţiile de lumină la
nivelul sistemului fotochimic II şi apoi la nivelul sistemului fotochimic I. În cursul acestui transfer de electroni
se produce şi o fosforilare, care constă în înglobarea unei părţi a energiei luminoase în energie chimică, sub
formă de grupări fosforice macroergice în ATP, la nivelul transportului de electroni de la citocromul b6 la
citocromul f. Se crede că ferredoxina poate să reducă în aceeaşi măsură şi citocromul b6, fosforilarea fiind astfel
posibilă şi fără intervenţia apei şi degajarea de O2. În acest caz este excitat doar sistemul fotochimic I şi electronii
emişi de centrul de reacţie al acestuia îşi reiau locul după ce au trecut prin ferredoxină, citocromul b6 şi
citocromul f. Această fosforilare - în cadrul căreia moleculele de clorofilă a reprimesc chiar electronii pe care i-
au emis - se numeşte fotofosforilare ciclică, spre deosebire de fosforilarea aciclică, în cursul căreia moleculele
de clorofila a din centrul de reacţie al fotosistemului II revin la starea stabilă pe seama electronilor proveniţi de
la apă. Fotofosforilarea aciclică este cuplată şi cu reducerea nicotinamid-adenin dinucleotid-fosfatului, forma
oxidată (NADP+) la (NADPH) + H
+.
Grupările macroergice de ATP rezultate prin fosforilare nu pot difuza spre citoplasmă, întrucât membrana
dublă a cloroplastelor este impermeabilă. Transferul energiei din cloroplaste în citoplasmă şi în diferite organite
citoplasmatice se face prin intervenţia triozofosfaţilor fotosintetizanţi şi a acidului fosfogliceric, ce rezultă în
reacţiile de glicoliză.
Organizarea - din punct de vedere funcţional - a moleculelor de pigmenţi asimilatori din sistemele
fotochimice pe de o parte în centri de reacţie, iar pe de altă parte în antene de pigmenţi reprezintă o adaptare ce
permite realizarea unui randament fotosintetic mai bun; aceasta, deoarece pigmenţii antenelor ce pot absorbi
individual doar un număr mic de fotoni pe secundă colectează energia fotonilor absorbiţi şi o transmit centrilor de
reacţie, permiţându-le să pompeze un mare număr de electroni de înaltă energie pe secundă.
Dacă fiecare moleculă de clorofilă a ar funcţiona şi ca pompă de electroni, fiecare dintre aceste molecule
ar trebui să dispună de un set propriu de substanţe, ce participă la transportul de electroni caracteristic fiecărui
sistem fotochimic. Aceasta ar face ca din lumina ce ajunge la pigmenţii asimilatori să fie absorbită de către
aceştia doar o fracţiune mică, restul radiaţiilor devenind ineficiente, printr-o absorbţie inutilă, de către moleculele
de substanţe ce intervin în transportul de electroni. Într-o astfel de situaţie randamentul fotosintezei s-ar micşora
pe de o parte prin înrăutăţirea condiţiilor de absorbţie a luminii de către pigmenţii asimilatori, iar pe de alta prin

faptul că moleculele de substanţe ce intervin în transportul de electroni nu ar fi folosite eficient, întrucât fiecare
moleculă ar transporta un număr mic de electroni în unitatea de timp.
II. Faza de întuneric a fotosintezei
FAZA a II - a, de întuneric, faza BLACKMANN sau faza termo-chimică, enzimatică - constă într-un
ansamblu de reacţii independente de lumină, dar dependente de temperatură, în care intervin anumite enzime
localizate în stroma nepigmentată a cloroplastelor. Aceste reacţii permit fixarea CO2 pe o substanţă acceptoare,
regenerarea acceptorului şi sinteza substanţelor ce conţin carbonul preluat din CO2 absorbit. Astfel se sintetizează
substanţe organice primare (trioze fosforilate), care polimerizează în substanţe organice complexe (glucide,
lipide, proteine, acizi nucleici, pigmenţi).
Reacţiile din cursul fazei de întuneric sunt grupate în ciclu CALVIN–BENSON-BASSHAM, după
numele cercetătorilor care au contribuit la clarificarea a numeroase aspecte ale mecanismului fotosintezei şi care
au primit în anul 1961 Premiul Nobel pentru chimie. Durata ciclului este de aproximativ 5 secunde.
Prima substanţă organică sintetizată în frunze prin procesul de fotosinteză este ribulozo-1,5-difosfatul
(RuDP), cu 5 atomi de carbon în catenă. Această substanţă poate fixa CO2 în prezenţa apei, în urma fixării
rezultând două molecule de acid fosfogliceric (A.P.G.).
Reducerea acidului fosfogliceric la aldehidă fosfoglicerică este fenomenul chimic, cel puţin parţial cel
mai important al fotosintezei, deoarece această reacţie îmbogăţeşte catena de atomi de carbon în hidrogen şi
energie. Hidrogenul necesar acestei reacţii este furnizat de NADPH + H+, iar energia necesară reducerii este
dată de ATP, substanţe ce se formează în cursul reacţiilor fazei de lumină a fotosintezei.
Până nu de mult se considera că fixarea la lumină a CO2 atmosferic şi încorporarea carbonului în
substanţe organice s-ar face la toate speciile vegetale pe o cale unică, conform reacţiilor din ciclul CALVIN,
denumit mai recent ciclul C3, întrucât primul compus stabil format după cuplarea CO2 pe acceptorul organic este
un compus cu 3 atomi de carbon - acidul-3-fosfogliceric. S-a constatat însă că există unele specii la care fixarea
fotosintetică a CO2 atmosferic se realizează pe o altă cale - calea HATCH – SLACK - KORTSCHAK (H.S.K.)
sau ciclul C4 - deoarece primii compuşi stabili formaţi după cuplarea CO2 sunt doi produşi cu 4 atomi de carbon,
acidul malic şi acidul aspartic, care derivă dintr-un compus instabil cu 4 atomi de carbon - acidul oxalilacetic.
La plantele la care CO2 se acumulează în cantitate mare sub formă de acid malic această specializare
biochimică este reflectată în majoritatea cazurilor atât în structura frunzelor, cât şi în cea a cloroplastelor. Celulele
mezofilului de la aceste plante posedă cloroplaste cu tilacoizi şi grane normale, conţin granule de amidon puţine
şi sunt aşezate radiar în jurul tecii fasciculare. Celulele din teaca fasciculară, în care se înmagazinează mult
amidon, conţin cloroplaste cu granele slab dezvoltate.
Se cunosc în prezent peste 100 de genuri de mono- şi de dicotiledonate ce aparţin tipului C4 , (planta tip
trestia de zahăr). Acest tip de fixare şi reducere a CO2 este întâlnit la specii care cresc în deşerturile aride şi în
pădurile tropicale umede şi umbroase, deci în condiţii ce nu favorizează desfăşurarea fotosintezei la nivel normal.
Aceste plante sunt totuşi capabile de o asimilaţie a carbonului foarte intensă, deoarece acidul fosfoenol piruvic
(PEP) pe care se fixează CO2 în celulele mezofilului, în prima etapă a reacţiilor de întuneric, este mult mai
reactiv cu CO2 decât ribulozo -1,5 - difosfatul (Ru DP) format, ca prim produs de sinteză, în frunzele plantelor
aparţinând tipului C3. Aceasta face ca în spaţiile intercelulare ale frunzelor de tip C4 concentraţia CO2 să fie foarte
mică, fapt ce favorizează o difuziune intensă a CO2 din atmosferă, prin ostiolele stomatelor extrem de puţin
deschise, în spaţiile intercelulare. Ca rezultat al acestei adaptări în regiunile aride, unde lipsa de apă limitează
fotosinteza, plantele de tip C4 pot desfăşura o fotosinteză intensă chiar în cazul unor temperaturi ridicate, fără să
rişte deshidratări fatale. De asemenea, în zonele tropicale şi ecuatoriale, cu vegetaţie luxuriantă, această adaptare
le permite să utilizeze în procesul de fotosinteză mici cantităţi de CO2 existente în aerul din jurul plantelor, care
formează desişuri masive.
Multe plante suculente din familiile Crassulaceae, Cactaceae, Liliaceae etc. fixează în timpul nopţii
cantităţi mari de CO2 pe acidul fosfoenolpiruvic (PEP), cu formare de malat, ce se acumulează în sucul vacuolar
al celulelor asimilatoare. Vacuolele mari ale celulelor asimilatoare servesc, deci, în acest caz, nu numai pentru
acumularea şi reţinerea apei, ci şi ca rezervoare de CO2 , necesar în activitatea fotosintetică.

Separarea în timp a proceselor care conduc la acumularea CO2 în timpul nopţii şi la utilizarea lui în
fotosinteză în cursul perioadei de lumină a zilei prezintă un deosebit avantaj pentru plantele suculente care, trăind
în locuri secetoase, îşi asigură prin aceste mecanisme o bună aprovizionare cu CO2, fără a risca pierderi din
conţinutul de apă, întrucât acestea au stomatele deschise la minimum în timpul perioadei de lumină a zilei.
Plantele cu acest mecanism de fixare a CO2 se numesc plante de tip C.A.M. (crassulacean acid metabolism,
planta tip ananasul).
Din punct de vedere anatomic o frunză de Crassula de exemplu prezintă un mezofil omogen. Celulele de
la exterior (de sub epiderme) prezintă un număr mai mare de cloroplaste, comparativ cu cele din centrul frunzei,
structura acestor frunze fiind, în general, mult mai apropiată de aceea a plantelor de tip C3, structura lor ne
prezentând nici un dimorfism celular sau cloroplastic. În apropierea fasciculelor conducătoare din mezofilul
acestor frunze se găsesc celule cu granule de amidon, dar nu există o teacă asimilatoare perifasciculară.
Ca adaptări structurale pentru factorul apă frunzele de la speciile din familia Crassulaceae prezintă o
textură cărnoasă, ce favorizează reţinerea apei în ţesuturi şi reduce viteza schimburilor gazoase.
Separarea în timp între perioada nocturnă, favorabilă absorbţiei intense a CO2 şi aceea din timpul zilei,
când are loc formarea puterii reducătoare a CO2, contribuie la reglarea randamentului fotosintetic, ceea ce
permite acestor plante să supravieţuiască în condiţiile secetei din mediul lor de viaţă.
Diferenţa dintre tipurile fotosintetice HATCH – SLACK - KORTSCHAK şi C.A.M. constă în aceea că la
primele reacţiile biochimice sunt separate spaţial, având loc în celule aparţinând la ţesuturi diferite (mezofil şi
teaca fasciculară asimilatoare), pe când la ultimele aceste reacţii sunt separate în timp (ziua şi noaptea),
desfăşurându-se în aceleaşi celule.
În natură există puţine situaţii ecologice şi geografice strict specifice pentru o anumită cale fotosintetică.
Plantele de tip C4 şi CAM prezintă un înalt grad de halofilie şi rezistenţă la secetă. S-a emis chiar ipoteza
conform căreia ciclul C4 ar constitui o modificare a ciclului C3, ca un răspuns al plantelor la presiunea selectivă a
mediului xerofitic sau salin, iar carboxilarea acidului fosfoenolpiruvic (PEP), prin acumularea acidităţii pe care
o produce, ar avea la origine o adaptare faţă de neutralizare a cationilor minerali de către anionii organici.
Plantele de tip C4 necesită ioni de Na+
pentru desfăşurarea normală a metaboismului lor. În lipsa Na+ apar
necroze şi cloroze speifice, în timp ce acest cation nu este absolut necesar plantelor de tip C3. Balanţa între tipul
C3 şi tipul C4 poate fi influenţată, de asemenea, de vârsta frunzelor sau de nutriţia minerală, în special de ionii
de Na+, aportul acestora favorizând trecerea la tipul C4 sau la tipul CAM. Astfel, dacă se suprimă NaCl din
mediul de cultură, specia Mesembryanthemum crystallinum, de tip CAM, tinde să devină de tip C3.
Pentru plantele de tip CAM (Agave americana sau speciile de Opuntia) regimul hidric, asociat cu
temperatura şi fotoperioada sunt hotărâtoare în reglarea metabolismului. Astfel, pentru Opuntia, după căderea
ploii, când filocladiile au un potenţial hidric ridicat, fixarea nocturnă a CO2 este maximă, prin formarea acidului
malic. În timpul zilei stomatele lor sunt închise şi totuşi plantele asimilează CO2, luându-l din decarboxilarea
acizilor dicarboxilici acumulaţi în vacuole, utilizând calea C4. Această caracteristică reprezintă o adaptare
remarcabilă a respectivelor plante, ce le permite să-şi închidă stomatele în timpul zilei, fără a-şi opri procesul de
fotosinteză, pe când la plantele de tip C3 orice închidere a stomatelor în timpul zilei reduce serios fixarea CO2 şi,
deci, fotosinteza. Aceste strategii la care recurg plantele arată corelaţii ecofiziologice dintre cele mai uimitoare:
plantele de tip CAM sunt esenţial legate de mediile aride, în vreme ce plantele de tip C4 sunt mult mai frecvente
în ecosistemele semi-aride de latitudini joase, iar plantele de tip C3 predomină în zonele temperate şi nordice de
latitudini mai ridicate.
FACTORII DE MEDIU CARE INFLUENŢEAZĂ PROCESUL DE FOTOSINTEZĂ
a. Factorii externi
●Lumina. Frunzele plantelor nu utilizează toată energia luminii care cade pe suprafaţa lor: 20% din
aceasta este reflectată şi se risipeşte în spaţiu, iar 10% străbate frunzele, fără a fi absorbită. Deci, din lumina
care cade pe suprafaţa frunzelor acestea absorb 70%, din care utilizează în fotosinteză numai aproximativ 1%.
Din fracţiunea absorbită 20% ridică temperatura frunzelor cu circa 1-20
C peste valoarea temperaturii

mediului ambiant, iar restul de 49% este eliminată din frunze prin fenomenul de transpiraţie. Aceste valori
prezintă o serie de variaţii ce depind de structura plantelor (de exemplu cuticula netedă şi lucioasă la unele
specii sau mată la altele, care influenţează procentajul de lumină reflectată ori grosimea frunzelor şi
cantitatea de pigmenţi asimilatori, ce influenţează cantitatea de radiaţii care străbat frunzele).
În afară de faptul că intervine în declanşarea proceselor care duc la mărirea gradului de deschidere a
stomatelor (favorizând difuziunea O2 şi a CO2 din și, respectiv, în mezofil) lumina intervine în fotosinteză prin
intensitate, calitate şi prin acţiunea manifestată asupra procesului de respiraţie foliară la unele specii.
• În funcţie de intensitatea luminii incidente fotosinteza are loc în mod diferit: la plantele heliofile
acesta creşte în intensitate până la 50-60% din intensitatea maximă a luminii solare directe care poate atinge,
în zilele senine de vară, 100.000 lucşi. La plantele sciafile intensitatea fotosintezei creşte odată cu mărirea
intensităţi iluminării doar până la aproximativ 10% din intensitatea maximă a luminii solare directe, după
care, la iluminări mai mari, aceasta scade.
• Lumina influenţează fotosinteza şi prin compoziţie, (lungimea de undă a radiaţiilor incidente pe
frunze). Mersul fotosintezei în radiaţiile monocromatice depinde de garnitura de pigmenţi asimilatori de care
dispun diferite grupe de plante şi de însuşirile acestor pigmenţi de a absorbi diferite radiaţii ale spectrului
vizibil.
La plantele superioare şi la algele verzi fotosinteza decurge cu o intensitate mai mare în radiaţiile
albastre-violet şi în cele roşii, spectrul de acţiune urmând spectrul de absorbţie al luminii de către pigmenţii
asimilatori. La algele brune fotosinteza are loc aproape cu aceeaşi intensitate în radiaţiile verzi şi în cele
albastre datorită prezenţei fucoxantinei, care absoarbe radiaţiile verzi. La algele roşii fotosinteza decurge cu
intensitate maximă în lungimile de undă din zona mijlocie a spectrului vizibil, ca urmare a faptului că
ficoeritrina are absorbţie maximă în acest domeniu.
În cursul evoluţiei plantele s-au adaptat în sensul elaborării unor pigmenţi care să le permită
absorbţia în condiţii optime a acelor radiaţii luminoase care există în mediul lor natural de viaţă. Acest fapt
este deosebit de evident în repartiţia algelor pe adâncime în apă: în mediul acvatic diferitele radiaţii ale
spectrului solar pătrund cu atât mai la adânc cu cât lungimea lor de undă este mai mică: radiaţiile roşii
pătrund până la adâncimi mult mai mici (maxim 34 m), comparativ cu cele albastre (500m). Ca urmare,
algele verzi predomină în zona superficială a apelor, iar la adâncimi mai mari predomină algele brune şi
roşii, ce posedă pigmenţi accesorii, capabili să absoarbă radiaţiile de lumină ajunse în aceste zone.
●Temperatura. În mersul intensităţii fotosintezei în funcţie de temperatură se pot distinge: o
temperatură minimă - temperatura cea mai joasă la care procesul începe să se manifeste; o temperatură
optimă - temperatura la care procesul decurge cu intensitatea cea mai mare şi constantă în timp; o
temperatură maximă - temperatura cea mai ridicată la care procesul mai poate avea loc. Procesul de
fotosinteză începe, în general, la o temperatură uşor inferioară valorii de 00C, creşte în intensitate odată cu
ridicarea temperaturii, atingând la unele specii valoarea maximă la 30-400C după care, prin mărirea, în
continuare, a temperaturii, acesta descreşte rapid, încetinind în jurul temperaturii de 500C.
Ca limită inferioară a temperaturii la care fotosinteza mai are loc, poate fi discutat cazul plantelor cu
frunze persistente în timpul iernii, la care procesul are loc la temperaturi mult mai coborâte: - 60C (molid) ori
numai -20C (grâul de toamnă).
Temperatura optimă a fotosintezei, strâns legată cu condiţiile de iluminare şi cu concentraţia CO2 din
mediu variază, în general, la plantele mezoterme, caracteristice regiunilor noastre, între 20-300C.
Limita maximă a temperaturii la care procesul de fotosinteză se mai produce este situată, la plantele
din regiunile noastre, la valori sub 45-500C, iar la plantele din regiunile sudice la valori de 50-55
0C.
b. Factorii interni
● Gradul de hidratare al ţesuturilor asimilatoare. La frunzele plantelor superioare terestre fotosinteza
scade destul de repede în intensitate odată cu creşterea deficitului de apă al frunzelor; în acelaşi timp existenţa
unei cantităţi mai mari de apă în ţesuturile foliare duce la o scădere uşoară a procesului, probabil din cauza

reducerii spaţiilor intercelulare, ca urmare a saturării celulelor cu apă, fapt ce duce la o micşorare a difuziunii
gazelor implicate. La deficite de apă mai mari de 13% procesul se reduce progresiv şi încetează la un deficit
de apă cuprins între 40-50%.
● Cantitatea de asimilate. Pentru ca fotosinteza să decurgă cu o intensitate normală este necesar ca
produşii rezultaţi să fie transportaţi rapid din celulele asimilatoare în celelalte părţi ale corpului plantelor,
unde vor fi utilizaţi în diferite sinteze sau vor fi puşi în rezervă.
La plantele amilofile, înspre amiază, cloroplastele se încarcă cu cantităţi mari de amidon, fapt ce
deranjează desfăşurarea intensă a fotosintezei, procesul sporind din nou ca intensitate abia în timpul după
amiezii, după ce o parte din amidonul acumulat anterior, printr-o fotosinteză activă, va fi fost transportat din
frunze.
● Cantitatea de pigmenţi asimilatori. În mod obişnuit pigmenţii asimilatori se găsesc în exces în
celulele asimilatoare. Această cantitate devine un factor limitant doar în cazurile de etiolare a plantelor,
fenomen produs de lipsa luminii, care se soldează, la unele plante, cu reducerea substanţială a cantităţii de
pigmenţi în celulele asimilatoare.
IV. EVOLUȚIA CRANIULUI LA VERTEBRATE : CONDROCRANIUL, SPLANHNOCRANIUL,
DERMATOCRANIUL1
Craniul este o structură complexă, formată din trei părţi distincte din punct de vedere al originii
filogenetice. Cea mai veche parte este considerată a fi cea viscerală (splanhnocraniu), care iniţial susţinea
deschiderile faringiene la protocordate. A doua parte, numită condrocraniu, protejează şi susţine creierul, fiind
alcătuită din oase endocondrale sau cartilaje. Cea mai recentă parte ar fi cea a dermatocraniului, de origine
desmală, care contribuie la formarea cutiei craniene.
La majoritatea vertebratelor, condrocraniu este de la început (primar) o structură embrionară care
serveşte ca „schelă” pentru dezvoltarea creierului şi ca suport pentru capsulele senzoriale. Două tipuri de
celule embrionare participă la formarea condrocraniului: celulele crestelor neurale, ca ectomezenchim,
contribuie la capsula nazală, trabecule (posibil numai în partea anterioară) şi probabil parte din capsula otică;
mezenchimul, de origine mezodermică, contribuie pentru restul condrocraniului.
Diferenţierea condrocraniului debutează cu condensări mezenchimatoase în regiunea cefalică, din care
se formează cartilaje alungite în continuarea notocordului. Perechea anterioară de cartilaje poartă denumirea
de trabecule, cea posterioară este a cartilajelor paracordale, iar la unele vertebrate încă o pereche de cartilaje
polare dispuse între ele. In urma cartilajelor paracordale, apar cartilajele occipitale. Acestor cartilaje se alătură
capsulele cartilaginoase senzoriale nazale, optice, otice. Ulterior, cartilajele fuzionează. Regiunea rezultată
din unirea capsulele nazale cu partea anterioară a trabeculelor se numeşte platou etmoid. Cartilajele
paracordale se unesc între ele medial, rezultând placa bazală dintre capsulele otice. Cartilajele occipitale
înconjurând notocordul şi tubul neural, formează arcul occipital. Toate aceste cartilaje prelungite şi fuzionate
constituie condrocraniu. La ciclostomi, condrocraniul este rudimentar, cu pereţii reduşi, fără regiune
occipitală. La elasmobranhi, condrocraniu este cartilaginos, cartilajele crescând şi formând o capsulă
protectoare completă pentru creier. La majoritatea celorlalte vertebrate, condrocraniul se osifică parţial sau în
întregime.
Splanhnocraniul este o structură veche a cordatelor, fiind asociat iniţial cu suprafeţele de filtrare a
hranei. La vertebrate, primar, susţine branhiile şi oferă suprafaţă de inserţie pentru muşchii respiratori; la
gnatostomieni diferenţiază şi fălcile. Elementele splanhnocraniului contribuie la formarea fălcilor şi a
aparatului hiobranhial la gnatostomieni. Sursa embrionară a splanhnocraniului diferă la cordate. La
protocordate, celulele crestelor neurale sunt absente, iar arcurile faringiene, care constituie coşul branhial,
sunt formate din ţesut conjunctiv fibros, niciodată osos, provenit din mezoderm. La vertebrate,
splanhnocraniul provine din celulele crestelor neurale (material ectomezenchimal). Aceste celule migrează
1 Pentru ilustrarea structurilor descrise să se consulte bibliografia recomandată

din preajma tubului neural spre pereţii faringelui, între fantele faringiene, unde se vor diferenţia arcurile
faringiene (în general, în număr de 7 perechi la gnatostomieni). Fiecare arc visceral este alcătuit din segmente,
după cum este reprezentat sintetic în tabelul următor.
Tabel. Diferenţierea arcurilor viscerale la gnatostomieni.
Denumirea Stadiile dezvoltării
arcului mezenchimatos cartilaginos osos - endocondral
proximal distal proximal distal proximal distal
oral (I) nesegmentat
cartilajul palatopătrat cartilajul lui Meckel pătrat articular
hioidian (II) nesegmentat
hiomandibular hioid hiomandibular stilohial; epihial;
ceratohial; hipohial;
bazihial*
Branhiale
(III-VII)
nesegmentate
faringobranhial; epibranhial;
ceratobranhial; hipobranhial;
bazibranhial*
Faringobranhial; epibranhial;
ceratobranhial; hipobranhial;
bazibranhial*
* segment impar
Oasele dermice care contribuie la formarea craniului constituie dermatocraniu. Filogenetic, aceste
oase provin din plăcile osoase ale tegumentului (rezultate din osificarea directă a ţesutul mezenchimal şi
ectomezenchimal al dermei) prezente la primele vertebrate, plăci care s-au scufundat, asociindu-se cu
splanhnocraniu şi condrocraniu (dar şi cu centura pectorală constituind componentele dermice ale acesteia).
Dermatocraniu formează laturile şi plafonul (cupola) craniului, delimitând cutia craniană, care protejează
encefalul; de asemenea, formează majoritatea oaselor din bolta cavităţii bucale şi înconjoară mult din
splahnocraniu.
Oasele dermatocraniului pot fi convenţional grupate în mai multe serii: facială (premaxilare, maxilare,
nazale); orbitală (lacrimale, prefrontale, frontale postfrontale, postorbitare, jugale); temporală (pătratojugal,
intertemporal, supratemporal, tabular, scvamozal); a bolţii craniului (frontalul, parietal, postparietal); palatină
(vomer, palatin, ectopterigoid, pterigoid, parasfenoid); mandibulară (dentar, splenial, angular, supraangular,
coronoid, prearticular).
Morfologia craniului. Din punct de vedere morfologic, craniul prezintă ca părţi componente:
neurocraniul (cutia craniană), fălcile, aparatul hiobranhial/hoidian.
Ciclostomii au un neurocraniu cu podeaua cartilaginoasă şi bolta conjunctivă. La peştii condrihtieni,
cutia craniană este o capsulă cartilaginoasă; dermatocraniul este absent, scheletul fiind în întregime
cartilaginos. La majoritatea peştilor osoşi şi tetrapode, cutia craniană este osificată. Platforma (podeaua) are
elemente endocondrale ce susţin encefalul, iar în completarea cutiei craniene sunt oase desmale. Podeaua
endocondrală este asamblată dintr-o serie de oase sfenoide (bazisfenoid, presfenoid, pleurosfenoide,
orbitosfenoide). Oasele occipitale (bazioccipitalul, supraoccipitalul şi occipitalele laterale), care au origine
vertebrală, sunt situate la capătul podelei sfenoidale, închizând posterior cutia craniană; regiune occipitală
lipseşte la ciclostomi. Occipitalele delimitează gaura occipitală, prin care trece nevraxul. Capsulele otice
rămân în partea posterioară a platformei endocondrale şi închid părţile senzoriale otice. Splanhnocraniul
contribuie cu epipterigoidele (alisfenoidele în cazul mamiferelor) la platforma endocondrală; de asemenea,
furnizează columela (la tetrapodele nemamifere), respectiv cele trei oase ale urechii medii – ciocanul,
nicovala şi scăriţa (la mamifere).
Fălcile, caracteristice gnatostomienilor, au apărut prima dată la peştii acantodieni şi placodermi,
permiţând consumarea unor prăzi mai mari.
Falca superioară constă în cartilajul palatopătrat, complet funcţional la condrihtieni. La peştii osoşi şi
tetrapode, acesta de regulă participă limitat, prin doar două derivate: epipterigoidul, care fuzionează cu
neurocraniul, şi pătratul, care suspendă falca în cazul tetrapodelor – cu excepţia mamiferelor la care constituie
nicovala din urechea medie. O serie de oase dermice completează falca superioară: premaxilare, maxilare,
palatine, pterigoide.

Falca inferioară (mandibula) este reprezentată de cartilajul lui Meckel, în exclusivitate la condrihtieni.
La mulţi peşti şi tetrapode, cartilajul lui Meckel persistă, dar este îmbrăcat în oase desmale; capătul posterior
al cartilajului se osifică dând osul articular (la mamifere constituie ciocanul din urechea medie). La unele
tetrapode, şi capătul anterior al cartilajului se osifică, rezultând osul mental. La mamifere, mandibula constă
într-un singur os dermic, numit dentar.
Aparatul hio-branhial este un derivat ventral al splanhnocraniului, situat în urma fălcilor. La peşti
aparatul hio-branhial, constituit din partea ventrală a perechii arcului hioidian (II) şi din arcurile branhiale,
susţine podeaua cavităţii buco-faringiene şi sistemul branhial. La tetrapode, arcurile branhiale regresează
(resturi se regăsesc în scheletul laringo-traheal), persistă aparatul hioidian, ale cărui elemente susţin podeaua
buco-faringiană şi limba.
ANATOMIA COMPARATĂ A SISTEMULUI NERVOS LA VERTEBRATE2
La vertebrate sistemul nervos cuprinde, din punct de vedere topografic, sistemul nervos central (dispus
pe partea dorsală a corpului) şi sistemul nervos periferic. Sistemul nervos central, este reprezentat de măduva
spinării şi encefal, iar sistemul nervos periferic de nervi (spinali și cranieni) și ganglioni nervoși.
A. Măduva spinării. Măduva spinării are forma unui cordon turtit la ciclostomi sau cilindric la
gnatostomieni. Calibrul măduvei spinării este, de regulă, uniform la vertebratele primar acvatice. Specia
Trigla lucerna constituie o excepţie, prezentând la capătul anterior al măduvei cinci perechi de intumescenţe
corelate cu dezvoltarea puternică a aripioarelor pectorale. Tetrapodele au de-a lungul măduvei spinării două
intumescenţe determinate de dezvoltarea membrelor: una anterioară, numită şi brahială, alta posterioară –
lombo-sacrală sau pelviană.
Secţiunile transversale realizate prin măduva diferitelor specii de vertebrate ilustrează structura
măduvei: substanţă cenuşie la interior şi substanţă albă spre exterior. Aranjarea substanţei cenuşii variază în
seria vertebratelor. La ciclostomi apar două coarne laterale de substanţă cenuşie. Substanţa cenuşie a măduvei
la peşti prezintă un corn dorsal şi două ventrale. Toate tetrapodele au substanţa cenuşie, pe secţiune
transversală, de forma literei H, cu două coarne dorsale şi două ventrale.
B. Encefalul. Encefalul, deşi prezintă o evoluţie cu puternice transformări progresive în seria
vertebratelor, păstrează un plan structural destul de unitar, ilustrat în tabelul 2.
Tabelul 2. Planul structural al encefalului la vertebrate.
Segment encefalic Structuri nervoase
dorsal ventral
telencefal palium striatum şi septum
diencefal epitalamus talamus (lateral) hipotalamus
mezencefal tuberculi bigemeni
(cvadrigemeni la mamifere)
tegmentum (pedunculi cerebrali la mamifere)
metencefal cerebel tegmentum (punte la mamifere)
mielencefal (bulb) ventricul mielencefalic tegmentum (+piramide şi olive la mamifere)
B.1. Encefalul la peşti (Pisces)
Peşti elasmobranhi (Pisces, Chondrichthyes). Caracteristicile de grup, precum şi variabilitatea
morfologică a encefalului de elasmobranhi pot fi urmărite la patru specii: Raja clavata, Squalus acanthias,
Dasyatis pastinaca, Torpedo marmorata. Telencefalul este relativ mare (proporţional mai voluminos la
Dasyatis pastinaca), cu emisferele cerebrale incomplet delimitate între ele şi continuate anterior cu lobii
olfactivi pedunculaţi. Diencefalul apare relativ larg pe faţa dorsală. Ventral, hipotalamusul delimitat anterior
de chiasma optică prezintă lobul median cu hipofiza bine dezvoltată, doi lobii inferiori (formaţiuni
caracteristice peştilor) şi doi saci vasculari. Mezencefalul prezintă tuberculii bigemeni ovali bine delimitaţi
între ei, mediu dezvoltaţi – mai mari apar la Squalus acanthias. Ventral, tegmentumul mezencefalic se
2 Pentru ilustrarea structurilor descrise să se consulte bibliografia recomandată

continuă cu cel de la nivelul metencefalului şi mielencefalului. Metencefalul poartă dorsal cerebelul alcătuit
dintr-un corp (paleocerebelul) şi două auricule laterale (arhicerebelul). Corpul este mare, de formă ovală şi cu
un şanţ cruciforn la Squalus acanthias şi Raja clavata. La Dasyatis pastinaca corpul cerebelos este
hipertrofiat şi cu numeroase şanţuri, revărsat anterior până la telencefal şi posterior până peste bulb.
Mielencefalul, moderat dezvoltat, prezintă dorsal ventriculul mielencefalic, cu prelungiri anterioare, care
pătrund în auriculele cerebeloase. La Torpedo marmorata pot fi identificaţi la nivelul bulbului doi lobi
„electrici” corespunzători hipertrofierii componentei motorii, ce asigură inervaţia musculaturii branhiomerice
transformată, în mare parte la această specie, în organe electrice.
Acipenseride (Pisces, Chondrostei). Telencefalul prezintă emisferele cerebrale incomplet separate
între ele, cu un început de eversiune. Lobii olfactivi situaţi anterior sunt sesili. Diencefalul poate fi observat
pe partea dorsală, apărând ca o bandă relativ lată. Tuberculii bigemeni reprezintă o parte voluminoasă a
encefalului. Cerebelul este slab dezvoltat, de forma unei lame transversale cu două auricule laterale
(arhicerebelul). Bulbul este tipic.
Peşti teleosteeni (Pisces, Teleostei). Encefalul teleosteenilor prezintă particularităţi de grup, dar şi o
variabilitate morfologică determinată atât de factori sistematici cât şi de cei eto-ecologici. Cele cinci segmente
encefalice apar aliniate, fără a delimita curburi. Telencefalul, de tip eversat, este reprezentat de două emisfere
cerebrale continuate anterior cu lobii olfactivi. Majoritatea teleosteenilor (cum apar şi la Gobius
melanostomus) au lobii olfactivi sesili şi doar puţine grupe (de exemplu, ciprinidele, siluridele, gadiidele) îi
au pedunculaţi. Emisferele cerebrale apar proporţional mai mari la speciile cu simţul olfactiv mai dezvoltat,
precum Silurus glanis şi unele ciprinide (Cyprinus carpio, Carassius carassius). Diencefalul este aproape
complet acoperit dorsal. Ventral, la nivelul hipotalamusului pot fi indentificate următoarele structuri: chiasma
optică, lobul median cu hipofiza, lobii inferiori şi sacul vascular. Sacul vascular, o structură impară, de
origine ependimo-meningeală, poate avea dimensiuni mai reduse, cu un lumen simplu, aşa cum apare la
Cyprinus carpio, ori poate fi mare, cu un lumen foarte sinuos, ca la specii marine bentonice precum
Scophthalmus maeoticus. Mezencefalul este reprezentat dorsal de tuberculii bigemeni, ventral de tegmentum
(continuat de cel de la segmentele posterioare - metencefalic, mielencefalic). Pe o secţiune transversală
realizată la nivelul mezencefalului pot fi observate structurile din interiorul ventricului mezencefalic
(torusurile longitudinale – intruziuni ale scoarţei tectale, torusurile semicirculare şi valvula cerebelului –
componentă cerebeloasă), precum şi scoarţa tuberculilor bigemeni (tectală). Tuberculii bigemeni la toţi
teleosteenii reprezintă porţiunea cea mai voluminoasă a encefalului. La speciile răpitoare, atât de apă dulce
(Stizostedion lucioperca, Esox lucius, Aspius aspius) cât şi marine (Scomber scombrus, Sarda sarda),
tuberculii bigemeni sunt proporţional foarte mari, văzul fiind un simţ foarte bine dezvoltat. Metencefalul are
cerebelul bine dezvoltat, reprezentat de un corp (paleocerebelul), eminenţele granulare (arhicerebelul) şi
valvula cerebelului. Corpul apare constant bine dezvoltat, proporţional mai mare la speciile cu mobilitate mai
mare (de exemplu, Scomber scombrus, Aspius aspius). Valvula cerebelului reprezintă o prelungire anterioară
a cerebelului, tipică teleosteenilor, situată în ventriculul mezencefalic. Morfologia ei variază de la un grup la
altul: este mai voluminoasă şi trilobată la ciprinide sau mai mică şi tetralobată la percide. Mielencefalul apare
simplu la unele specii precum Gobius melanostomus, Stizostedion lucioperca, Esox lucius, Scomber
scombrus, Sarda sarda, Aspius aspius. La teleosteenii care prezintă simţul gustativ foarte bine dezvoltat se
produce o hipertrofiere a centrilor nervoşi corespunzători (nucleii gustativi ai nervilor VII, IX şi X) cu o
complicare a morfologiei bulbare prin formarea unor lobi: patru la Silurus glanis, trei la ciprinide.
B.2. Encefalul la amfibieni (Amphibia)
Urodele (Urodela). Encefalul de Triturus are telencefalul de tip inversat, ca la toate tetrapodele, cu
emisferele alungite reprezentând de acum segmentul cel mai voluminos al encefalului. Tuberculii bigemeni
sunt mai mici, ovali. Cerebelul, slab dezvoltat, este reprezentat de o bandă transversală iar bulbul apare
simplu.
Anure (Anura). O secţiune sagitală realizată prin encefalul de Rana ilustrează alinierea segmentelor
(lipsa curburilor), de asemenea, caracterul inversat al telencefalului. Emisferele cerebrale care reprezintă

aproape jumătate din lungimea encefalului se continuă anterior cu lobii olfactivi uniţi între ei. Diencefalul
apare dorsal ca o bandă destul de largă, iar ventral, în urma chiasmei optice, prezintă lobul median cu hipofiza
voluminoasă. Mezencefalul are tuberculii bigemeni ovali, orientaţi uşor oblic faţă de axul longitudinal al
encefalului; scoarţa tectală poate fi observată pe secţiunile encefalului. Torusurile semicirculare sunt bine
dezvoltate şi unite între ele median, încât ventriculul mezencefalic apare etajat după cum se observă pe
secţiunea transversală realizată la acest nivel. Metencefalul prezintă un cerebel redus la o simplă lamă dispusă
transversal a cărei scoarţă poate fi observată pe secţiunea sagitală a encefalului. Mielencefalul este simplu, cu
ventriculul de formă triunghiulară.
B.3. Encefalul la reptile (Reptilia). Segmentele encefalice la reptile configurează de acum, prin
aranjarea lor, cele trei curburi caracteristice amniotelor. Ele pot fi observate pe secţiunea sagitală a encefalului
de Lacerta: curbura vertexului, la nivelul mezencefalului, convexă în sus; curbura pontină, la nivelul
metencefalului, convexă în jos; curbura nucală, la nivelul mielencefalului, convexă în sus. Telencefalul
prezintă cele două emisfere cerebrale nu prea mari la Lacerta, uşor mai voluminoase la Natrix şi Emys. Pe
secţiunea sagitală a encefalului de Lacerta poate fi observată scoarţa cerebrală apărută pentru prima dată la
reptile. Lobii olfactivi cu care se continuă rostral emisferele cerebrale sunt alungiţi. Diencefalul este aproape
în întregime acoperit dorsal, iar ventral prezintă formaţiunile hipotalamice specifice tetrapodelor (întâlnite şi
la amfibieni). Mezencefalul prezintă tuberculii bigemeni încă relativ mari, mai cu seamă la şopârlă. La şarpe,
înapoia lor, torusurile semicirculare sunt parţial ieşite la suprafaţă. Metencefalul are dorsal cerebelul de forma
unei lame transversale mai groasă la şarpe şi mai ales la broasca ţestoasă. Mielencefalul este simplu.
B.4. Encefalul la păsări (Aves). Encefalul la păsări, aşa cum apare la Columba livia, este voluminos,
cu particularităţi evolutive şi adaptative. Telencefalul are emisferele cerebrale voluminoase, fiecare
prezentând pe faţa dorsală câte un şanţ superficial numit valeculă. Secţiunea sagitală realizată prin encefalul
de porumbel ilustrează o caracteristică importantă: dezvoltarea puternică a corpilor striaţi cu reducerea
ventriculelor laterale la simple fante. Scoarţa cerebrală este subţire, slab dezvoltată. Caracterul microsmatic al
telencefalului este sugerat de lobii olfactivi cu dimensiuni reduse. Diencefalul este acoperit dorsal în totalitate
pe de o parte de emisferele cerebrale, pe de altă parte de cerebel. Hipotalamusul are morfologia caracteristică
tetrapodelor. Mezencefalul prezintă tuberculii bigemeni relativ mari, împinşi în poziţie latero-ventrală. Pe o
secţiune realizată la nivel mezencefalic poate fi observată dorsal scoarţa tectală foarte dezvoltată, în interior
ventriculul mezencefalic cu torusuri semicirculare, iar ventral tegmentumul. Metencefalul are cerebelul
voluminos, întins anterior până la baza emisferelor cerebrale, iar posterior peste o mare parte din bulb.
Cerebelul are o structură plurilamelară, cu o alternanţa caracteristică de substanţă cenuşie şi substanţă albă
(„arborele vieţii”) pe secțiune. Mielencefalul este acoperit dorsal, în mare parte, de cerebel.
B.5. Encefalul la mamifere (Mammalia). Encefalul mamiferelor apare modificat datorită, în
principal, dezvoltării accentuate a paliumului şi a prezenţei neoformaţiunilor. Telencefalul prezintă emisferele
cerebrale foarte voluminoase, continuate anterior cu lobii olfactivi foarte bine dezvoltaţi la mamiferele
macrosmatice, precum Oryctolagus cuniculus şi Bos taurus. La cele două specii, suprafaţa emisferelor
cerebrale este reprezentată ventral de paleocortex şi dorso-lateral de neocortex – a cărui dezvoltare
pronunţată, caracteristică mamiferelor, este corelată cu diferenţierea neoformaţiunilor encefalice. Cele două
teritorii paleale care apar la exterior, paleocortexul şi neocortexul, sunt separate între ele de un şanţ rinal. Cel
de-al treilea teritoriu paleal, arheocortexul, este scufundat (înrulat) în interiorul ventriculului lateral; cea mai
importantă parte a sa numită cornul lui Amon poate fi evidenţiată printr-o secţiune combinată a telencefalului
de Bos taurus. Suprafaţa neocortexului poate fi netedă – deci mai redusă ca întindere (neocortex lisencefalic),
ca la Oryctolagus cuniculus, sau cutată (neocortex girencefalic) ca la Bos taurus. Secţiunea sagitală realizată
prin encefal evidenţiază comisura corpului calos (comisura care leagă neocortexul celor două emisfere) mult
mai bine dezvotată la Bos taurus (neocortexul fiind mai voluminos) decât la Oryctolagus cuniculus.
Diencefalul este acoperit dorsal de emisferele cerebrale, iar ventral are morfologia tipică tetrapodelor;
talamusul poate fi observat pe secţiunea sagitală realizată prin encefalul celor două specii de mamifere luate
în studiu. Mezencefalul este reprezentat dorsal de tuberculii cvadrigemeni (cea de-a doua pereche fiind

considerată ca reprezentând torusurile semicirculare ieşite înafară), iar ventral de pedunculii cerebrali.
Pedunculul cerebral conţine neoformaţiunea numită piciorul peduncului (adăugată ventral peste tegmentumul
mezencefalic), mult mai pronunţată la Bos taurus, datorită mai bunei dezvoltări a neocortexului. Cele două
componente ale pedunculului cerebral pot fi observate pe secţiunile sagitale ale celor două encefale.
Metencefalul prezintă dorsal cerebelul, iar ventral puntea. Cerebelul, bine dezvoltat, apare situat posterior
emisferelor cerebrale, întinzându-se înapoi şi peste bulb. Morfologic, cerebelul prezintă dorsal vermisul,
lateral emisferele cerebeloase – flancate la rândul lor de lobulii arhicerebelului. Structura cerebelului
polilamelară este evidenţiată de către numeroasele şanţuri existente la suprafaţă iar la interior – pe secţiunea
sagitală a organului – prin alternanţa caracteristică de substanţă albă şi substanţă cenuşie numită „arborele
vieţii”. Puntea, după cum apare pe fiecare din secţiunile sagitale, conţine partea veche (tegmentumul) şi
neoformaţiunea numită puntea propriu-zisă, mai consistentă la Bos taurus. Mielencefalul poartă dorsal doar
porţiunea posterioară a ventriculului IV, cea anterioară aparţinând de metencefal (situată sub cerebel). Pe faţa
ventrală apar neoformaţiunile: median – piramidele bulbare mai bine dezvoltate la Bos taurus, iar lateral –
olivele bulbare evidente la Bos taurus.
C. Nervii spinali. Nervii spinali sunt conectaţi la măduva spinării, având o dispoziţie metamerică.
Numărul lor variază odată cu cel al somitelor troncale (de exemplu, 10 perechi la Rana, 31 perechi la om,
peste 100 de perechi la unii şerpi). Nervul spinal prezintă în alcătuirea sa două rădăcini, trunchi şi ramuri.
Rădăcinile sunt: dorsală şi ventrală. Cea dorsală, are pe traiectul său un ganglion (ganglion dorsal) în care se
găsesc corpii protoneuronilor senzitivi somatici şi viscerali. La Amphioxus, protoneuronul este situat în
măduva spinării, neexistând ganglionul dorsal. La ciclostomi, rădăcinile se formează altern (cea dorsală dintr-
un metamer „n”, iar cea ventrală dintre două metamere), nu se unesc între ele constituind fiecare câte un nerv
separat: cel dorsal cu fibre senzitive (somatice şi viscerale) şi unele visceromotorii, iar cel ventral cu restul
fibrelor visceromotorii, plus cele somatomotorii. La gnatostomieni, ambele rădăcini se formează în acelaşi
metamer, apoi se unesc într-un trunchi mixt. Numai la mamifere, cele două rădăcini sunt complet separate
funcţional: dorsala senzitivă, ventrala motorie. Ramurile principale sunt: dorsal şi ventral. Ramul dorsal
aparţine musculaturii epaxonice şi pielii spatelui, iar cel ventral este pentru musculatura hipaxonică şi pielea
din regiunea ventro-laterală a corpului. La vertebratele care au un lanţ ganglionar laterovertebral, din ramul
ventral se desprinde un ram comunicant (visceral) cu fibre visceromotorii preganglionare. La mamifere, din
acest ram comunicant se diferenţiază unul alb (preganglionar) şi altul cenuşiu (posganglionar) pentru periferie
(glande tegumentare, muşchiul erector al firului de păr). Ramul dorsal este tot mai subţire în seria
vertebratelor, cu cât musculatura epaxonică îşi pierde din importanţă. O dată cu ştergerea metameriei
musculare se pierde şi dispoziţia metamerică a nervilor rahidieni, ei formând plexuri. Tetrapodele au două
plexuri: cervico-brahial, lombo-sacral; nervii toracici păstrează metameria. La vertebratele superioare se
formează plexurile: cervical, brahial, lombar, sacral. Trebuie de menţionat că la formarea plexurilor participă
numai ramurile ventrale, cele dorsale păstrându-şi metameria.
D. Nervii cranieni. Nervii cranieni au rădăcinile cuprinse în encefal. Denumirile lor utilizate frecvent
sunt cele din anatomie umană. Amniotele au 12 perechi, anamiotele au 10 perechi plus alţii nenumerotaţi (de
exemplu nervul lateral). Nervii cranieni pot fi grupaţi în: somatomotori, somatosenzitivi şi branhiomerici.
D.1. Nervii somatomotori. Nervii somatomotori (perechile III, IV, VI, XII) inervează musculatura
somatică a capului: muşchii extrinseci ai globului ocular, musculatura hipobranhială. Nervul oculomotor
comun (III) inervează trei din muşchii drepţi ai gobului ocular (superior, inferior, intern) şi muşchiul micul
oblic. De asemenea, acest nerv mai include fibre parasimpatice care inervează musculatura intrinsecă a
globului ocular (muşchii ciliari, ai irisului). Nervul patetic (IV) inervează muşchiul marele oblic al globului
ocular. Nervul oculomotor extern (abducens) inervează muşchiul drept extern şi muşchiul retractor bulbi
când există. Nervul hipoglos (XII) este prezent numai la amniote, inervând la acestea musculatura limbii.
D.2. Nervii somatosenzitivi. Nervii somatosenzitivi (0, I, II, VIII, nervul lateral) aparţin organelor de
simţ. Nervul terminal (0) apare la toate vertebratele (cu excepţia ciclostomilor şi păsărilor), cel puţin în stadiu
embrionar. Culege aferenţe din mucoasa olfactivă la peşti sau din organul vomero-nazal Jacobson la

tetrapode. Nervul olfactiv (I) are caracter primitiv întrucât neuronii de origine sunt localizaţi periferic, chiar în
epiteliul mucoasei olfactive. Ca şi precedentul, este legat de telencefal. Nervul optic (II), legat de diencefal,
este de fapt un tractus intracerebral (cupa optică fiind de origine diencefalică). Adevăratul nerv optic ar fi
reprezentat de axonii celulelor bipolare din retină. Nervul acustico-vestibular sau octavus (VIII). La
vertebratele inferioare conţine predominant fibre vestibulare şi mai puţine acustice. Acestea din urmă sporesc
de la amfibieni, atingând ponderea cea mai mare la mamifere, la care axistă două porţiuni ale nervului:
vestibulară (cu origine în ganglionul Scarpa) şi acustică (cu origine în ganglionul Corti). Nervul lateral este
reprezentat de nervii liniei laterale, pentru neuromaste, existând la agnate, peşti, formele acvatice de amfibieni
(urodele exclusiv acvatice, larvele anurelor). Iniţial, nervii liniei laterale au ganglioni proprii, ulterior aceştia
se unesc cu cei ai nervilor branhiomerici. Astfel, nervul lateral anterior însoţeşte trigemenul şi facialul,
culegând informaţii de la neuromastele de pe cap; nervul lateral posterior însoţeşte glosofaringianul şi vagul,
culegând informaţii de la neuromastele din teritoriul branhial; alte fibre independente culegând din regiunea
trunchiului.
D.3. Nervii branhiomerici. Nervii branhiomerici sunt asociaţi arcurilor viscerale. Nervul trigemen
(V) este o formaţiune complexă, inervând arcul oral. La vertebratele inferioare conţine doi nervi: nervul
oftalmic profund, senzitiv, care ar aparţine unui arc preoral, şi nervul trigemen propriu-zis, care are un ram
maxilar (senzitiv) şi un ram mandibular (mixt). La mamifere, există un singur nerv, cu trei ramuri: oftalmic,
maxilar şi mandibular. Nervul trigemen este lipsit de componenta viscerosenzitivă şi de cea visceromotorie
generală. Are componenta somatosenzitivă, care asigură sensibilitatea generală a feţei şi motorie pentru
musculatura arcului oral. Nervul facial (VII) inervează arcul hiodian. A pierdut aproape complet componenta
somatosenzitivă, dar o are pe cea viscerosenzitivă generală şi specială. Ramul palatin al facialului dă
sensibilitatea plafonului regiunii faringiene, la care se adaugă fibrele gustative. Aceste din urmă elemente se
reduc la vertebratele superioare, în schimb apar componentele parasimpatice pentru glandele mucoase şi
glanda lacrimală. Ramul mandibular intern asigură inervarea gustativă a părţii anterioare a limbii şi secreţia
glandelor salivare submaxilară şi sublinguală. Ramul mandibular extern este motor, inervând musculatura
pieloasă a feţei. Nervul glosofaringian (IX) este nervul primului arc branhial. În el predomină componenta
viscerosenzitivă specială (sensibilitatea gustativă de la baza limbii). Componenta motorie este slab dezvoltată
la vertebratele superioare, inervând o serie de muşchi ai laringelui. Fibrele parasimpatice sunt pentru glanda
salivară parotidă. Nervul vag (X) este pentru următoarele arcuri branhiale. La peşti, prezintă patru ramuri
branhiale şi un ram intestinal pentru trunchi. Componenta viscerosenzitivă specială (gustativă) este bine
dezvoltată la peşti şi redusă la tetrapode. Componenta somatosenzitivă este mai importantă la mamifere.
Ramul intestinal devine la amniote componenta cea mai importantă, fiind pentru motricitatea viscerală
generală (fibre parasimpatice pentru organele cavităţii generale). Nervul spinal sau accesoriu (XI) este
caracteristic amniotelor, existând complet numai la mamifere. Este un nerv motor care inervează muşchiul
trapez, iar la mamifere şi derivatele acestuia: sternomastoidianul şi cleidomastoidianul
(sternocleidomastoidianul la om). Nervul spinal are două componente: una desprinsă din vag (cu origine în
partea caudală a nucleului ambiguu) şi una de origine spinală. Nervul hipoglos (XII) este, de asemenea,
caracteristic amniotelor. Cu origine în bulb, fibrele sale somato-motorii inervează musculatura intrinsecă şi
extrinsecă a limbii.
V. ORGANIZAREA SISTEMICĂ ȘI NIVELURI DE ORGANIZARE A LUMII VII
1. Teoria sistemelor – noțiuni generale.
Din perspectiva teoriei sistemice studiază sistemele supraindividuale de organizare ale materiei vii
(populații, biocenoze, biosfera) integrate în mediul lor abiotic, inclusiv sistemul socio-economic uman și
relațiile acestuia cu componentele ecosferei. Astfel teoria generală a sistemelor este bază conceptuală și
metodologică pentru abordarea ecologiei. Conform acestei teorii, întreaga materie este organizată în corpuri
de natură diferită (atomi, molecule, planete, galaxii, ființe vii) care reprezintă tot atâtea sisteme.
Sistemul este un ansamblu de elemente, identice sau diferite, unite prin conexiuni într-un întreg. Deci,
sistemul apare ca un ansamblu organizat, astfel încât conexiunile și funcțiile elementelor componente

(subsisteme) concură la menținerea și îndeplinirea funcțiilor întregului.
Din perspectiva relațiilor cu mediul (cu alte sisteme), sistemele pot fi de trei categorii:
- izolate: nu au schimburi de materie și energie cu mediul; nu există în natură; unele sisteme artificiale
(recipiente de tip termos) funcționează ca sisteme izolate, temporar.
- închise: au doar schimburi energetice cu mediu; nu există sisteme naturale absolut închise; un
recipient închis ermetic este un astfel de sistem; planeta Terra poate fi considerată cu o anumită larghețe un
sistem închis, deoarece schimburile de materie cu Universul sunt infime dacă le raportăm la volumul planetei
(sistemului).
- deschise: prezintă schimburi atât energetice cât și materiale cu mediu. Cum materie și energia sunt
suporturi (purtătoare) ale informației, aceste sisteme prezintă implicit și schimburi informaționale cu mediul;
toate sistemele naturale sunt sisteme deschise, iar sistemele vii (biologice) sunt sisteme deschise în mod
obligatoriu, prezentând o serie de însușiri dintre care unele sunt deosebit de importante din punct de vedere
ecologic, și care le deosebesc de sistemele nevii.
2. Însușirile generale ale sistemelor biologice. [Botnariuc & Vădineanu 1982]
Caracterul istoric (diacronismul). Însușirile structurale și funcționale ale sistemelor vii sunt
rezultatul istoriei, a evoluției lor în timp. Explicarea structurii și funcționării unui organism sau a unei
populații nu se poate face fără cunoașterea evoluției lor istorice. De ex. pentru a explica arealul actual al unei
specii trebuie cunoscută istoria speciei: centrul de origine, căile de dispersie, „pulsațiile” arealului cauzate de
modificări la scară spațio-temporală geologică (glaciar interglaciar) etc.
Dependența de condiția inițială și comportamentul neliniar. Condiția inițială a unui sistem poate
determina traiectoria comportamentului acestuia în timp. Acest comportament este însă neliniar: o schimbare
de o anumită magnitudine poate induce o modificare de magnitudine neproporțională cu cea a cauzei. După
unii autori, datorită comportamentului neliniar prognozarea traiectoriei este practic imposibilă. Ex:
modificarea concentrației N și P dintr-un lac poate declanșa procesul de „înflorire” algală cu consecințe
complexe la nivelul structurii și funcției ecosistemului.
Caracterul informațional. Sistemele biologice sunt sisteme informaționale, adică au capacitatea de a
recepționa, prelucra, stoca și transmite informație în mediul lor. Cu cât organizarea unui sistem este mai
avansată, cu atât cantitatea de informație deținută este mai mare. Existența oricărui sistem biologic depinde de
relațiile sale cu mediul, astfel tendința evolutivă constă în realizarea unei cantități optime de informație pentru
asigurarea persistenței sistemului. Perceperea și transmiterea fidelă a informației se realizează datorită
fenomenului de redundanță (suplimentarea informației unui mesaj fără alterarea sensului, pentru a-l proteja de
perturbația „zgomotului de fond”) atât la nivelul mesajului cât și al receptorului (doi ochi, două urechi).
Fidelitatea informației nu este absolută, de exemplu erorile de copiere a informației genetice (mutațiile)
reprezintă o sursă de evoluției prin selecția lor naturală de către mediu.
Integralitatea. Integralitatea. Orice sistem este alcătuit din mai multe părți care, datorită specializării
și interdependenței, se comportă ca un întreg. Cu cât specializarea (exacerbarea unor caracteristici în
detrimentul altora) părților este mai mare, cu atât interdependența acestora va fi mai puternică, iar
integralitatea sistemului va fi mai accentuată. Integralitatea sistemelor supraindividuale este inferioară celei
caracteristice organismelor individuale. Însușirile întregului sunt superioare însușirilor părților sau sumei
acestei însușiri, deoarece, datorită interacțiunii dintre părți, rezultă trăsături noi, emergente, proprii doar
întregului, care nu se regăsesc la nivelul părților acestuia. Nivel Individual Populațional Biocenotic
Trăsături
Emergente
metabolism,
existența în timp
limitată genetic,
ontogenia,
capacitatea de reacție la
modificările mediului,
sursă a variabilității, material
pentru selecție
existență nelimitată în timp,
rata reproducerii, dimensiunea,
densitatea,
dinamica,
structura pe vârste, sexe
reciclarea substanțelor, deci
regenerarea resurselor primare,
realizarea unei producții biologice

O altă consecință a integralității este că un subsistem integrat în sistem are alt comportament decât
înafara sistemului unde nu mai interacționează cu celelalte subsisteme. Din această cauză, rezultatele unor
experimente de laborator trebui completate cu observații sau experimente de teren și / sau naturale.
Autoreglarea. Sistemele biologice funcționează pe principii cibernetice, mecanismele implicând mai
multe tipuri de conexiuni:
Conexiuni externe: pun sistemul în relație cu mediul.
Conexiuni interne: directe (determină apariția unui răspuns la un anumit stimul) și inverse (feed-back)
(ajustează răspunsul inițial la caracteristicile stimulului) pozitive (amplifică răspunsul inițial) sau negative
(reduc răspunsul inițial).
Feed-before: mecanisme de prevenire și anticipație
Rezultatul acestor mecanisme constă în apariția unor oscilații în jurul unor valori programate ale
parametrilor sistemului. La organismele individuale există structuri specializate ce asigură funcționarea
mecanismelor cibernetice. La sistemele supraindividuale aceste structuri lipsesc sau sunt mai puțin evidente
(pot să apară indivizi specializați în cadrul grupurilor la organismele cu organizare socială complexă – unele
insecte, mamifere). Autoreglarea permite existența unui comportament activ al organismelor față de variația
variabilelor de mediu. Organismul alege hrana, caută sau construiește refugii, evită sau luptă cu inamicii,
alege locurile cu factori abiotici favorabili. Deci, sistemele biologice nu sunt entități pasive, ci se comportă
activ selectând sau chiar creând condiții/resurse de mediu.
Caracterul autopoetic. Este capacitatea sistemelor biologice de a se autoreface. Este rezultatul
capacității de autoorganizare a sistemelor biologice datorate conexiunilor funcționale dintre componentele
sistemului: componentele în interacțiune sunt capabile să reproducă sau să producă aceeași rețea de structuri
și procese care le-a dat naștere lor.
Caracterul antientropic. Este evidențiat de termodinamica sistemelor disipative ce mențin stabile
fluxurile materiale și energetice departe de echilibrul termodinamic, cu tendința de maximalizare a intrărilor
de energie din mediu, energie disipată pentru menținerea, dezvoltarea și creșterea complexității structurilor
sistemului. Comportamentul antientropic al sistemelor biologice contrazice în aparență legile termodinamicii.
Sistemele biologice se opun tendinței către echilibrul termodinamic prin creșterea continuă a entropiei, ele se
structurează acumulând negentropie: dezvoltarea ontogenetică, evoluția speciilor.
Caracterul fractal. Sistemele biologice depind de fluxul continuu de energie. Interfața sistem-mediu
are un rol important pentru menținerea acestui flux. Dar, suprafața crește la pătrat, iar volumul la cub, ceea ce
înseamnă că o creșterea biomasei organismelor implică amplificarea fluxului de energie la nivelul interfeței
rezultând un deficit de suprafață.
Deficitul de suprafață se poate rezolva pe două căi: (1) Fragmentarea suprafeței. Raportul
suprafață/volum rămâne constant dacă odată cu creșterea volumului, prin apariția de pliuri, excrescențe, prin
fragmentare se realizează mărirea și amplificarea schimburilor de suprafață. (2) Fluidizarea mediului. La
animalele reofile curenții de apă asigură un aport permanent de resurse, compensând imposibilitatea creșterii
suprafeței corpului care ar duce la alterarea hidrodinamicii acestuia.
Caracterul dinamic. Sistemele biologice pot exista timp nedefinit într-o fază de relativă stabilitate
sau de echilibru dinamic (Flussgleichgewicht = echilibru curgător cf. Ludwig von Bertalanffy). Rezultă
datorită mecanismelor cibernetice care realizează autocontrolul parametrilor sistemici ce suferă oscilații în
jurul unor valori medii. Acest caracter răspunde la oscilațiile periodice sau neperiodice ale factorilor de
mediu. Rezultă capacitatea sistemelor biologice de a-și menține homeostazia, de a rezita activ la variațiile,
adesea imprevizibile ale mediului. Mecanismele cibernetice asigură menținerea orientării, a tendinței inițiale a
proceselor biologice, chiar și sub influența perturbațiilor, fenomen numit homeorheză (stabilitate
homeodinamică). Datorită sensibilității lor, sistemelor dinamice au comportament neliniar ce reprezintă o
consecință a două mecanisme inițiale: bifurcații și haos.
Bifurcații. Comportamentul neliniar face ca traiectoriile sistemelor să fie imprevizibile deoarece în
dependență de condițiile inițiale și ca urmare a depășirii unor limite de către variațiile unor parametri sistemici

inițiali, oscilațiile sistemului se modifică, devin eratice, în aparență dezordonate. Consecința acestor fluctuații
constă în multiplicarea traiectoriilor posibile ale evoluției ulterioare a sistemului – fenomen numit bifurcație.
Haos. Reprezintă amplificarea oscilațiilor care face ca sistemul să intre într-un regim dinamic, aparent
dezordonat. La sistemele dinamice apare un haos determinist în sensul că are niște reguli proprii care nu sunt
cunoscute în totalitate. Starea de haos reprezintă o tranziție către o nouă stare de echilibru dinamic a
sistemului. Se caracterizează printr-un anumit grad de dezordine cu creștere de entropiei și imprevizibilitatea
traiectoriei ulterioare. Ulterior, are loc trecerea la un nou regim dinamic, la structurarea de noi cicluri
catalitice care duc și la scăderea entropiei.
Ierarhia de programe. Comportamentul în sens larg al unui sistem viu se poate schimba în anumite
limite permițând sistemului realizarea unor stări diferite. Programul este una din stările posibile pe care
sistemul dat este capabil să le realizeze în limitele permise de organizarea sa. Programul reprezintă
schimbarea posibilă, viitoare, a sistemului determinată de structura lui (Amosov 1965). Sistemele au mai
multe traiectorii posibile, motiv pentru care se realizează programul pentru care există condiții potrivite. Deci
un sistem este guvernat de mai multe programe care, datorită ierarhizării, se clasifică în trei categorii: (i)
proprii („pentru sine”): asigură autoconservarea sistemului; (ii) inferioare: stările subsistemelor din sistemul
dat; (iii) superioare: stările unui sistem dat menite să asigure existența suprasistemului din care face parte.
Programele inferioare sunt subordonate programelor proprii care se subordonează programelor
superioare. Însă, programele inferioare constituie limitări ale celor superioare, adică ultimele nu se pot derula
dacă primele nu se realizează corespunzător.
Teoria „gena egoistă” (Selfish Gene - Richard Dawkins, 1976) conform căreia genele tind să se
reproducă / răspândească uneori în defavoarea unor purtători explică dominarea programelor specie asupra
programelor proprii indivizilor din specia respectivă, precum și fenomene nefavorabile indivizilor ca selecția
de grup înrudit si altruismul.
3. Ierarhia sistemelor biologice, categorii ierarhice în lumea vie.
Orice sistem viu este alcătuit din elemente componente (subsisteme) și este la rândul său un subsistem
al unui sistem mai mare, integrator (suprasistem). Astfel rezultă un ansamblu de sisteme cu grade de
complexitate diferite constituite într-o structură ierarhizată: subsistem < sistem < suprasistem. O consecință a
acestei ierarhii este aceea că sistemele au proprietăți sumative (rezultate prin însumarea proprietăților
subsitemelor componente) și proprietăți emergente (proprietăți noi ale unui sistem ce nu se regăsesc la
nivelului subsistemelor componente).
Sistemele vii sunt extrem de complexe, dar pot fi grupate în patru mari categorii ierarhice: somatică,
organizatorică, ecologică și taxonomică.
Ierarhia somatică. Include totalitatea sistemelor ierarhizate, vii și nevii, ce intră ca elemente
componente în alcătuirea unui sistem biologic de nivel individual (organism). Din această cauză, ele se mai
numesc și infraorganismice sau subindividuale. Această ierarhie cuprinde: particule elementare, atomi,
molecule, macromolecule,organite, celule, țesuturi, organe și sisteme de organe.
Ierarhia organizatorică. Include sistemele biologice, adică nivelurile de organizare ale materiei vii.
Un nivel de organizare cuprinde sisteme biologice diferite din punct de vedere structural și funcțional,
capabile de o existență de sine stătătoare și cu însușiri universale comune. Această ierarhie cuprinde:
individul (organismul), populația (specia), biocenoza (comunitatea) și biosfera.
Individul reprezintă forma elementară și universală de existență a vieții. Indivizii au o existență în
timp limitată genetic și realizează un schimb permanent de materie, energie și informație cu mediul
înconjurător (proces numit metabolism). Toate sistemele supraindividuale sunt forme de grupare și organizare
a indivizilor. Fiecare organism prezintă funcții de relație, nutriție și reproducere. Indivizii pot fi reprezentați și
de organisme unicelulare, ca urmare, celula poate fi nivel al ierarhiei somatice dar și al ierarhiei
organizatorice.
Populația este alcătuită dintr-un grup de indivizi aparținând aceleiași specii, care își desfășoară
activitatea în cadrul unei biocenoze. Indivizii unei populații sunt interconectați prin relații intraspecifice și

mai ales prin relații de reproducere. Populația este capabilă de existență de sine stătătoare, din punct de vedere
reproductiv, timp nedefinit.
Specia este o comunitate reproductivă alcătuită din una sau mai multe populații. Specia este în același
timp un sistem biologic (un nivel în ierarhia organizatorică) și un nivel în ierarhia taxonomică. Astfel, specia
ca sistem biologic, devine veriga de legătură dintre ierarhia organizatorică și ierarhia taxonomică.
Biocenoza include totalitatea indivizilor grupați în populații distincte ce ocupă un anumit biotop
(spațiu sau volum fizic). Principala caracteristică a biocenozei o reprezintă relațiile dintre speciile componente
(relații interspecifice), dintre care cele mai importante sunt cele trofice.
Biosfera este învelișul viu al planetei, ce include totalitatea indivizilor de pe Pământ, grupați în
populații și biocenoze.
Ierarhia sistemelor ecologice. Include totalitatea sistemelor ce intră în componența ecosferei.
Unitatea de bază a ecosferei este ecosistemul. Acesta este alcătuit dintr-o biocenoză împreună cu biotopul pe
care aceasta îl ocupă și include inter-relațiile dintre aceste două componente. Ecosfera integrează biosfera cu
suportul ei neviu (toposfera: totalitatea biotopurilor de pe Terra), alcătuit din hidrosferă și părți ale litosferei
și atmosferei. Ecosistemul este constituit din biocenoza integrată cu biotopul ei. Reprezintă unitatea
structurală elementară a ecosferei, capabilă să asigure transferul de materie, energie și informație. Biotopul
este constituit din totalitatea factorilor abiotici (nevii) de la nivel local.
Ierarhia taxonomică. Este alcătuită din categorii taxonomice (numite taxoni) de diferite ranguri,
mergând de la specie la regn.
4. Populația: efectiv, densitate, natalitate, mortalitate, structura de vârstă, structura de sex,
structura spațială.
Populația reprezintă forma elementară, necesară și suficientă, de existență de sine stătătoare a speciei
din punct de vedere genetic și ecologic. Ca subsistem al biocenozei, populația este influențată atât de factorii
abiotici cât și de alte populații din cadrul biocenozei, fie că este vorba de cele cu care stabilește relații trofice
(populațiile ce reprezintă hrană și populațiile de prădători), relații de competiție pentru resursa trofică și/sau
habitat, prezența unor paraziții sau agenți patogeni.
Statica populației. Reprezintă descrierea formală, cantitativă a unei populații indiferent de specia
căreia ii aparține. Din această cauză parametrii statici permit compararea populațiilor unor specii aflate pe
trepte evolutive diferite (populația de insecte cu populație de pești). Parametrii statici sunt: efectivul,
densitatea, natalitatea și mortalitatea.
Efectivul (dimensiunea populației). Numărul de indivizi sau biomasa (alge, bacterii, organisme
unicelulare), și reprezintă cantitatea totală de materie vie sub formă de indivizi existentă la un moment dat în
populație.
Dimensiunea unei populații la un moment dat este rezultatul interacțiunii dintre procesele care aduc
indivizi în populație (natalitatea și imigrarea) și procesele care scot indivizi din populație (mortalitatea și
emigrarea).
Efectivul este important deoarece asigură perpetuarea populației și implicit a speciei din care face
parte, și menține continuitatea lanțurilor trofice la nivelul biocenozei. O populației se menține viabilă dacă
efectivul oscilează între o valoare maximă și minimă. Depășirea maximului populației viabile poate duce la
intensificarea competiției intrapopulaționale și chiar la epuizarea unor resurse ceea ce face ca populația să
intre în declin numeric. Scăderea sub minimul populației viabile poate produce un declin, uneori ireversibil
prin reducerea diversității și implicit vigorii genetice a indivizilor din consangvinizării generalizate.
Densitatea. Efectivul raportat la unitatea de spațiu, suprafața sau volum. Unitatea de suprafață diferă
în funcție de talia speciilor (mamifere – hectare, nevertebrate – m2, la pești – ha luciu de apă). Determinarea
densității are importanță ecologică deosebită deoarece exprimă gradul de încărcare al biotopului – este o
măsură a capacității biotopului de a suporta o anumită cantitate de biomasă (resursele pe care le consumă
indivizii).
Densitatea optimă corespunde șansei maxime de supraviețuire a populației în respectivul biotop.

Conform principiului lui Allee supraviețuirea în populații e defavorizată atât de suprapopulație cât și de
subaglomerare. Supraaglomerarea are ca efect epuizarea resurselor de mediu, intensificarea concurenței
intraspecifice, ceea ce duce la destabilizarea populației. Subaglomerarea este defavorabilă deoarece poate
micșora șansa de reproducere a indivizilor și duce la o utilizare ineficientă a resurselor.
Natalitatea. Reprezintă numărul de indivizi nou apăruți pe unitatea de timp prin diviziune, eclozare,
germinație sau naștere. Natalitatea poate avea valori pozitive sau nule.
Natalitatea poate fi: (i) absolută – reprezintă numărul de indivizi ce pot fi produși de o populație în
condiții optime de existență; (ii) ecologică – reprezintă numărul efectiv de indivizi produși de o populație în
condiții concrete de existență.
Legat de natalitate cu influența asupra ei sunt fecunditatea și fertilitatea speciei. Fecunditatea
reprezintă performanța potențială sau capacitatea fizică a unei populații în ceea ce privește reproducerea.
Fertilitatea e gradul în care această capacitate fizică se concretizează în indivizi viabili .
Rata natalității reprezintă raportul dintre natalitate și numărul total de indivizi dintr-o populație sau
cifra etalon. Este un indicator al vitezei de creștere a populației. Rata natalității e condiționată ecologic și
genetic deoarece fiecare specie prezintă un anumit potențial biotic, are o anumită capacitate de a se înmulți și
de a supraviețui. Factorii ecologici pot limita potențialul biotic al speciei. Deci natalitatea este rezultatul
interacțiunii dintre potențialul biotic al unei specii și rezistența mediului. Pentru a evita acest fapt, unele specii
au strategii de supraviețuire care constau în apariția urmașilor într-un interval de timp îndelungat în mod
eșalonat deși ponta sau forma de rezistență a fost depusă la un moment dat. Prin această strategie (strategie de
evitare a riscurilor) se asigura supraviețuirea unui număr de urmași din totalitatea celor eclozați și anume a
acelora care au apărut atunci când condițiile de existență au fost favorabile vieții.
Mortalitatea. Reprezintă eliminarea indivizilor din populație fie prin moarte fiziologică, fie datorită
bolilor dăunătorilor, prădătorilor sau altor factori. În funcție de mortalitate se stabilește longevitatea unei
specii. Acesta poate fi fiziologici când exprima durata ciclului de dezvoltare a individului unei specii, sau
ecologică, aceasta reprezintă longevitatea medie realizată în condiții concrete de existență.
Rata mortalității e raportul dintre indivizii dispăruți într-un interval de timp și efectivul total al
populației sau o cifra etalon. Este un indicator al vitezei de descreștere a dimensiunii populației.
Prin analiza tabelei de mortalitate adică a mortalității în funcție de vârsta indivizilor dintr-o generație
se poate întocmi curba de supraviețuire a speciei care poate fi de 3 tipuri :
1. curbă puternic convexă. La populațiile la care mortalitatea se produce masiv la ultimele vârste
indicate de longevitate. La populațiile umane și la unele diptere (Drossophilla)
2. forma liniară. Pierderea unui număr constant de indivizi la fiecare categorie de vârstă. Foarte rar
întâlnită în natură. Apare la animale sesile (hidra, corali).
3. curbă puternic concavă. Mortalitatea foarte accentuată în primele vârste. In general la speciile cu
prolificitate mare și care nu își îngrijesc descendenții.
La acestea se mai pot adăuga încă două variante:
- forma în trepte – caracteristică organismelor cu stadii de dezvoltare cu mortalități diferite (insecte cu
dezvoltare cu metamorfoză).
- curba ușor concavă – este caracteristică unor pești și amfibienilor
Cunoașterea tipului de curbă de supraviețuire permite determinarea stadiilor cele mai vulnerabile din
viața unei specii. Intervenind în aceste stadii se poate modifica natalitatea sau mortalitatea și drept consecință
evoluția ulterioara a speciei.
Relația dintre mortalitate și vârstă depinde de strategia de supraviețuire a speciilor. Aceste strategii
alcătuiesc un continuum în lumea vie. Strategiile extreme sunt cunoscute sub denumirea de strategie r și
strategie K. Strategiile de tip r, caracterizează populațiile ce investesc mai mult în reproducere și mai puțin în
competitivitate (de exemplu, dintre mamifere populațiile de rozătoare). Produc un număr mare de
descendenți, dintre care foarte mulți nu ajung la maturitate. Grija părinților pentru descendenți este
cvasiinexistentă. Au o curbe de supraviețuire concave Populațiile cu strategii de tip K se caracterizează prin

indivizi ce investesc mai puțin în reproducere și mai mult în sporirea ratei de supraviețuire, prin mărirea
competitivității (de exemplu, populațiile de carnivore mari). Produc un număr mic de urmași, care sunt
îngrijiți de părinți și dintre care foarte mulți ajung la maturitate. Au curbă de supraviețuire convexă.
Structura populației poate fi privită din perspectiva:
Genetică: Se referă la distribuția alelelor și implicit haplotipurilor în genofondul populațional
(totalitatea genomurilor indivizilor componenți). Expresia structurii genetice poate fi evidentă în cazul
populațiilor polimorfice, în care diferite morfe (forme) au o valoare adaptativă diferită. Morfele contribuie la
perpetuarea populației atunci când au loc modificări ale condițiilor de existență (ex. molia mesteacănului,
forma melanică a permis perpetuarea populațiilor în contextul melanismului industrial). Poate avea implicații
ecologice în sensul în care reducerea diversității genetice din cauze ecologice (ex. scăderea dimensiunii
populației) contribuie la extincție
Ecologică: poate fi de: vârstă, sex și spațială
Structura de vârstă. Reprezintă proportia în care sunt reprezentate diferite grupe sau clase de vârste
față de efectivul total al populației. Criteriul cel mai important din punct de vedere populațional, de
delimitarea claselor de vârstă este participarea la reproducere a indivizilor componenți. Din această
perspectivă se pot delimita 3 clase de vârstă: (i) Prereproductivă (juvenilă). Durează de la formarea zigotului
din care se formează individul și până la prima reproducere (fără să o includă). (ii) Reproductivă(de
maturitate). De la prima reproducere până ultima. (iii) Post reproductivă (senescentă). După ultima
reproducere până la moarte.
La unele speciile de animale cu strategie r, viața adulților se încheie cu reproducerea (efemeroptere,
polichete, somoni) și astfel vârsta postreproductivă lipsește. Avantajul adaptativ constă în evitarea competiției
pentru resurse între indivizii de generații diferite și a canibalismului. La alte specii din aceeași categorie dar
cu generații suprapuse, relațiile antagonice dintre generații se rezolvă prin ocuparea unor nișe spațiale sau
temporale diferite (Naja pallida adulți nocturni, juvenili diurni; Ophiophagus hannah femela părăsește cuibul
înainte de eclozare și vânează în alte zone).
La speciile cu strategie k indivizii care nu participă la reproducere contribuie la educarea juvenililor
prin transmiterea experienței acumulate pe parcursul vieții (femela cea mai bătrână sau matriarhul conduce
turma de elefanți).
Într-o populația de plante se disting 6 clase de vârste caracterizate după faza de vegetație și stadiul
funcției reproductive : 1. plante în repaos (forme de semințe, tulpini subterane, forme de rezistență); 2. plante
care germinează; 3. plante imature, juvenile – incomplet dezvoltate vegetativ și din punct de vedere al
organelor sexuale; 4. plante virginale – complet dezvoltate vegetativ dar cu organele sexuale incomplet
dezvoltate; 5. plante reproducătoare – complet dezvoltate vegetativ și sexual; 6. plante senescente – mai sunt
capabile să vegeteze sau nu mai sunt capabile de reproducere.
Structura pe vârste se reprezintă grafic printr-o diagramă numită piramida vârstelor. Din analiza
structurii pe vârste la diferite specii s-au putut evidenția 3 tipuri de piramide ale vârstelor: 1. piramida cu bază
largă. Cuprinde cel mai mare număr de indivizi în stadiu juvenil și caracterizează populațiile aflate în creștere
rapida. 2. piramida normală. Are forma de clopot și exprimă un raport echilibrat între diferitele grupe de
vârstă. Caracterizează populațiile aflate în creștere staționară (natalitatea = mortalitatea). 3. piramida cu baza
îngustă. Reflectă un număr mic de indivizi tineri și o dominanță a adulților. Caracterizează populațiile aflate
în declin numeric.
Aceste predicții trebuie făcute doar considerând și alte aspecte demografice (ex. dimensiunea
populației, graficele de mortalitate / supraviețuire, strategii de reproducere / supraviețuire).
Structura pe sexe. Reprezintă proporția sexelor în populație = raportul dintre sexe sau sex ratio =
masuculi:femele. În funcție de vârsta unei generații la care este determinat, sex ratio este de tei feluri:
- Sex ratio primar: la momentul concepție; este de 1:1
- Sex ratio secundar: la naștere / eclozare; este de 1:1, dar există numeroase exemple de dominare a
unuia dintre sexe. De exemplu, la aligatori, din ouăle incubate la temperaturi mai scăzute (27.7-30oC)

eclozează femele; iar din cele incubate la temperaturi ridicate (33.2-33.8oC) eclozează masculi (determinism
sexual dependent de temperatură).
- Sex ratio terțiar: la atingerea maturității sexuale; este cel mai adesea balansat în favoarea unuia dintre
sexe. De exemplu, sex ratio secundar unei generații de reni a fost de 1:1 ; sex ratio terțiar al aceleași generații
a fost de 3:5.
- Sex ratio cuaternar: în clasa postreproductivă
În general sex ratio terțiar indică o predominare a femelelor la mamifere și o predominanță a
masculilor la păsări. Aceste situații sunt explicate de trei ipoteze:
Ipoteza cromozomială: sexul homogametic (femele XX la mamifere, masculi ZZ la păsări) are o
vigoare genetică mai ridicată decât cel heterogametic (masculi XY la mamifere, femele ZW la păsări),
datorită prezenței genelor majore (gene fără de care indivizii nu sunt viabili) în doză dublă. La sexul
heterogametic genele majore sunt în doză simplă (pe cromozomul X sau respectiv Z), cromozomii sexuali Y
și W fiind purtători de genele ce controlează expresia unor caractere sexuale secundare și a căror absență nu
determină moartea indivizilor.
Ipoteza mortalității: femelele sunt mai vulnerabile în timpul clocitului, masculii poligami folosesc
energie pentru atragerea și paza femelelor de alți masculi ceea ce îi face să fie mai vulnerabili față de
prădători.
Ipoteza competiției intraspecifice: unul dintre sexe domină în competiția pentru resurse. La unele
populații de păsări s-a observat o predominare a masculilor odată cu creșterea densității populaționale.
Masculii mamiferelor (elan) care nu-și apără resursele au de suferit deoarece talia mai mare a acestora
presupune cheltuieli energetice mai ridicate.
Multe situații sunt explicate parțial de mai multe ipoteze.
Au fost observate și excepții de la aceste generalizări. La multe păsări, masculii sunt mai vulnerabili
din cauza unei vizibilități mai ridicare (penaj cu culori contrastante, cântec complex reperabil de la distanțe
mari).
Se pare că în realitate, mai important decât sex ratio fizic este sex ratio operațional care raportul dintre
masculii competitori, gata de reproducere și femelele competitoare, gata de reproducere. Sex ratio operațional
diferă de sex ratio fizic prin faptul că nu ia în considerare indivizii sexual inactivi sau care nu intră în
competiție pentru parteneri de reproducere. Sex ratio operațional prezintă importanță pentru:
- Selecția sexuală: sexul predominant este supus unei competiții mai puternice pentru parteneri de
reproducere și implicit unei selecții sexuale mai puternice. Selecția sexuală poate fi intrasexuală (competiție
între indivizi de același sex) și intersexuală (realizată de către indivizii de sex opus prin selectarea unui
anumit partener de reproducere). Selecția sexuală favorizează purtătorii unor caractere ce contribuie la
succesul în competiția pentru partenerii de reproducere. Aceste caractere pot însă crește vizibilitatea și
vulnerabilitatea purtătorilor, motiv pentru care purtătorii unor caractere exagerat de dezvoltate deși avantajați
de selecția sexuală sunt eliminați prin selecție naturală.
- Intensitatea competiției: la indivizii din sexul mai abundent competiția este mai mare, în timp ce
indivizii celuilalt sex devin mai selectivi în alegerea partenerilor.
- Paza partenerilor: pe măsură ce sex ratio operațional se modifică în sensul unuia dintre sexe, cresc
șansele ca partenerii sin sexul mai puțin abundent să fie păziți de competitori.
Devierile de sex ratio de 1:1 pot fi compensate prin adoptarea unor tipuri diferite de sisteme de
împerechere care prezintă avantaje evolutive corespunzătoare:
- Monogamia: poate fi sezonieră (Anas acuta) sau permanentă (coiot, gâște canadiene). Partenerii nu
mai cheltuie energie pentru formarea perechilor și sunt mai puțin vulnerabili față de prădători.
Partenerii pot avea relații sexuale și înafara cuplului (lebădă).
- Poligamia:
- Poliginia: gorile, babuini, elefant de mare, fazan, cerb, elan, om; femelele îngrijesc
descendenții; costuri: diversitate genetică redusă, atacuri din partea masculilor competitori;

beneficii: abilitate de supraviețuire și succes reproductiv ridicate ale masculului posesor de
harem.
- Poliandria: la caradriiforme poate ajunge la 6:1, masculii preiau rolul de îngrijire a
descendenței iar femelele intră în competiție pentru parteneri de reproducere. Costuri:
diversitate genetică redusă; beneficii: abilitate de supraviețuire și succes reproductiv ridicate
femelei.
- Poliginandrie: 2 sau mai mulți masculi au relații sexuale exclusive cu 2 sau mai multe femel;
beneficii - diversitate genetică mare, competiție intersexuală redusă, îngrijirea descendenților
mai eficientă; la Prunella modularis.
- Promiscuitatea: împerecherea nediscriminantă, fără relații stabile. Multe specii social monogame
practică relații extraconjugale
Există specii la care sex ratio are o valoare informativă redusă deoarece au sisteme de reproducere
partenogenetică sau hermafrodită. La afide se succed generații de femele partenogenetice, masculii apărând la
finalul perioadei de activitate. Hermafroditismul secvențial: protoandrie (masculi devin femele) ex. peștele
claun, protoginie (femele masculi masculi) ex. unii pești, Rana temporaria. Hermafroditismul simultan: un
adult cu organle sexuale mascule și femele în același timp; melci, râme.
Reprezentarea grafică a structurii pe sexe se face în cadrul piramidei combinate de sex și vârstă. În
general prezența ambelor sexe în toate clasele de vârstă indică o populație viabilă, iar predominanța sexului
femel poate indica o creștere numerică a efectivului.
Structura spațială sau dispersia. Reprezintă modul de distribuție e în strânsă corelație cu limitele de
toleranță pe care le au indivizii față de factorii abiotici, cu interacțiunile acestora cu alte grupe de organisme și
cu comportamentul lor. Distribuția spațială influențează densitatea populației, cunoașterea sa fiind importantă
pentru descrierea mărimii și dinamicii populației. Există trei tipuri de distribuție spațială:
Distribuția uniformă. Indivizii au tendința de a se respinge. Presupune existența unor distanțe
aproximativ egale între indivizi. Se poate întâlni în acele medii care satisfac cerințele speciilor în orice punct
în aceeași măsură. Se întâlnește rar în natură, poate apărea ca rezultat al competiției pentru hrană și spațiu. De
exemplu, arborii dintr-o pădure matură se distribuie la distanțe aproximativ egale datorită comportamentului
uniform față de factorul lumină care duce la o concurența între indivizi, având ca rezultat menținerea unei
distanțe egale între aceștia. În cazul populațiilor animale o astfel de distribuție se întâlnește la speciile care au
comportament de teritorialitate. Teritoriul unui individ sau grup având o suprafață egală cu teritoriul altui
individ din aceeași specie.
Distribuția întâmplătoare. Indivizii nici nu se atrag, nici nu se resping. Aranjarea indivizilor unei
populații în habitat este neuniformă. Apare la speciile care trăiesc în medii omogene dar care nu au tendințe
de teritorialitate și nici gregare. De exemplu : păianjen, insecte galicole, protozoare.
Distribuția aglomerată (grupată, contagioasa). Indivizii au tendința de a se atrage reciproc. Este cea
mai răspândita în natura. indivizii sunt repartizați în habitat în grupuri de mărimi diferite, grupuri dispuse
uniform sau la întâmplare. De exemplu: afidele (păduchi de plante) distribuția grupată e determinată de
comportamentul gregar al indivizilor care caută să trăiască în apropierea semenilor lor și e determinată și de
variabilitatea condițiilor de biotop, indivizii fiind atrași de acele locuri unde condițiile de viață sunt optime.
Cauzele ce pot influența tipul de dispersie sunt: (i) Repartiția spațială a resurselor. O repartiție
uniformă poate determina orice tip de dispersie, în timp ce una concentrată, insulară, determină o dispersie
grupată. (ii) Modul de înmulțire. Înmulțirea vegetativă favorizează o dispersie grupată. Instinctul social. (iii)
Animalele cu instincte sociale adoptă o dispersie grupată (permanentă sau temporară).
5. Relațiile interspecifice.
Sunt relațiile ce se stabilesc între indivizi din populații diferite ce aparțin la specii diferite. În funcție
de modul în care relația avantajează (+), dezavantajează (-) sau lasă indiferenți (0) partenerii, acestea pot fi:

Mutualismul este o relație obligatorie, ambele populații
profitând de pe urma conviețuirii. Acest tip de relație este foarte
răspândit în natură, majoritatea speciilor conviețuind cu altele. De
exemplu, majoritatea animalelor conțin în sistemul digestiv
microorganisme care le permit digerarea sau asimilarea hranei.
Lichenii sunt un exemplu de mutualism împins la extrem, când
asocierea dintre ciuperci, alge și bacterii a dat naștere unor organisme
(respectiv specii) noi.
Comensalismul este o relație obligatorie pentru comensal, în timp ce organismul gazdă nu este afectat
de conviețuire. Cel mai adesea, organismul gazdă este folosit drept adăpost sau suport de către alte specii.
Dispersia semințelor realizată de unele specii de animale (zoochoria) se încadrează cel mai adesea în acest tip
de relație.
Prădătorismul este o relație obligatorie, benefică pentru prădător (+) și dăunătoare pentru pradă (-).
Indivizii din populația prădătorului consumă membrii populației pradă ca hrană. Acest tip de relație se
deosebește de parazitism, deoarece parazitul de obicei nu își omoară gazda. Are un rol important în reglarea
numărului de indivizi din ambele populații și stă la baza rețelei trofice din cadrul biocenozei. De exemplu,
dispariția unui prădător poate determina o explozie numerică a populației pradă ceea ce poate avea un impact
negativ la nivelul biocenozei. De exemplu, dispariția populațiilor de lupi poate duce la înmulțirea necontrolată
a cerbilor și căprioarelor, care pot determina distrugerea completă a plantelor prin suprapășunat, ceea ce
antrenează și moartea lor. Deci, la nivel populațional, efectul prădătorului asupra prăzii este benefic deoarece
prin prădătorism sunt eliminați indivizii tarați din populația pradă, și dimensiunea acesteia este controlată
evitându-se suprapopulația și epuizarea consecutivă a unor resurse.
Parazitismul este o relație obligatorie pentru organismul parazit care beneficiază de relația lui cu
organismul gazdă (efect pozitiv), în timp ce organismul gazdă este influențat negativ. Relația gazdă-parazit
este universală în lumea vie, practic neexistând specie care să nu fie ori parazitată (gazdă) sau să fie parazit.
Conceptul de populație și biocenoză nu au relevanță întotdeauna în acest caz. Pentru unii paraziți interni
organismul gazdă reprezentă habitatul iar aceștia reprezintă echivalentul unei populații. Întrucât majoritatea
speciilor au mai multe specii de paraziți vorbim de comunități de paraziți într-un organism gazdă, paraziții
interacționând la rândul lor unii cu alții.
Competiția interspecifică. Cauza acestei competiții este accesul la resursele limitate de mediu. În
funcție de particularitățile relației există mai multe tipuri de competiție:
- Consumptivă (de exploatare) – se realizează pentru resursele de hrană; indivizii concurează prin
utilizarea mai eficientă a resursei.
- Preemptivă (pt. spațiu) – la plante și animale sesile.
- Prin creștere – la plante, prin umbrire.
- Teritorială – pentru. ocuparea unui teritoriu.
- De întâlnire – prin luptă.
Există doua rezultate posibile ale interacțiunilor competitive:
- Excluderea (eliminarea) competitivă – destul de rară.
- Coexistența – este rezultatul cel mai frecvent; se poate realiza prin: - partajarea resurselor; accesarea
aceleași resurse dar din compartimente diferite; separarea temporală a utilizării resurselor; eficiența utilizării
resursei în funcție de condițiile de mediu.
Amensalismul reprezintă o relație care nu este obligatorie dintre două specii, nefavorabilă pentru una
dintre ele. De exemplu unele plante elimină substanțe toxice ce inhibă creșterea altor plante (alelopatie), acest
lucru conferindu-le un avantaj competițional. Acesta este cazul nucului ale cărui frunze căzute prin
descompunere generează compuși toxici pentru alte specii de plante.
Neutralism indică lipsa de interacțiuni directe între indivizi, populații sau specii. De exemplu,
indivizii unei populații de salamandre sau broaște dintr-o pădure de fag nu interacționează direct cu
Efectele Relația
+:+ Mutualism
+:0 Comensalism
+:- Prădătorism, Parazitism
-:- Competiție interspecifică
-:0 Amensalism
0:0 Neutralism

populațiile de păsări granivore (care se hrănesc cu semințe).
VI. PRINCIPII MODERNE DE CLASIFICARE A LUMII VII
ORGANIZAREA STRUCTURALĂ ŞI EVOLUŢIA TALULUI ORGANISMELOR DIN
REGNUL PROTISTA
Regnul Protista grupează organisme eucariote foarte heterogene unicelulare, coloniale sau
multicelulare, de tip haploid sau diploid, uneori cu celule plurinucleate, rareori şi cu o falsă diferenţiere
celulară (pseudoplectenchime). Se acceptă ideea că protistele actuale reprezintă mai multe linii evolutive
distincte şi de fapt celelalte organisme eucariote (fungi, muşchi, plante vasculare şi animale) îşi au originea în
protiste primitive.
Aceste organisme prezintă următoarele caracteristici:
organisme unicelulare heterotrofe cu organizare celulară de tip animal: Protozoare;
organisme heterotrofe cu stadii ameboidale şi plasmodiale în ciclul de viaţă cu o serie de caractere
apropiate de fungi: Myxomycota, Chytridiomycota, Oomycota;
organisme eucariote fotoautrofe, taloide unicelulare, coloniale şi pluricelulare simple: Algae;
nutriţia se realizează prin absorbţie, ingestie sau prin fotosinteză;
se înmulţesc prin meioză zigotică, sporică sau gametică;
peretele celular poate lipsi la unele specii, iar atunci când este prezent prezintă o compoziţie
chimică foarte variată;
flagelii sunt absenţi sau prezenţi (dar, în acest caz au o structură complexă, tipică).
Algele sunt considerate talofite care prezintă o mare diversitate morfologică şi structurală. După unii
autori sunt considerate organisme vegetale micro şi macrofite foarte heterogene care populează cu predilecţie
mediul acvatic (ape dulci, sărate şi salmastre).
Înmulţirea algelor poate fi vegetativă, asexuată şi sexuată.
Înmulţirea vegetativă se poate produce prin diviziunea celulară la formele unicelulare şi prin
fragmentarea talului la algele pluricelulare fără modificări evidente ale protoplasmei.
Înmulţirea asexuată se realizează prin spori asexuaţi specializaţi. Algele prezintă două tipuri de spori
asexuaţi: zoospori şi aplanospori.
Zoosporii (planosporii) sunt spori asexuaţi mobili care se deplasează cu ajutorul flagelilor. Celula în
care se formează zoosporii se numeşte zoosporange (zoosporocist).
Aplanosporii sunt spori asexuaţi specializaţi care nu prezintă flageli şi de aceea nu sunt mobili.
Înmulţirea sexuată este foarte diferită şi se poate realiza prin gametogamie şi somatogamie
(conjugare).
Gametogamia se realizează prin fecundaţia gameţilor cu polaritate sexuală diferită. Formarea
gameţilor întotdeauna este precedată de meioză şi din această cauză sunt celule haploide (n). Din punct de
vedere morfologic şi biochimic gameţii prezintă o mare variabilitate. Ţinând cont de aceste considerente se
deosebesc: izogameţi şi heterogameţi.
Izogameţii pot fi flagelaţi şi sunt identici din punct de vedere morfologic, dar diferiţi din punct de
vedere biochimic.
Heterogameţii sunt diferiţi din punct de vedere morfologic şi biochimic. În foarte multe cazuri,
gametul femeiesc este mai mare, mobil şi se numeşte macrogamet. Există situaţii când gametul femeiesc este
mare, imobil şi se numeşte oosferă. Gametul bărbătesc este mult mai mic decât cel femeiesc şi se numeşte

microgamet. Gameţii bărbăteşti pot fi mobili şi imobili. Gameţii bărbăteşti mobili se numesc anterozoizi sau
spermatozoizi, iar cei lipsiţi de flageli se numesc spermatii.
În funcţie de morfologia gameţilor se deosebesc următoarele tipuri de gametogamie: izogamia,
anizogamia şi oogamia.
La algele roşii se întâlneşte un caz particular de gametogamie: fecundaţia (carpogamia) are loc între
doi gameţi (oosferă, spermatie) care sunt imobili. Oosfera se formează în carpogon, iar spermatia în
spermatangiu.
Somatogamia constă în fuziunea întregului conţinut celular care provine de la două celule vegetative
cu polaritate sexuală diferită. Astfel, cele două celule fuzionează prin intermediul unui canal de conjugare
(copulaţie). După fecundaţie, rezultă zigotul (2n). Acest tip de înmulţire la alge poartă numele de conjugare
(Spirogyra).
Ciclul de dezvoltare şi alternanţa de generaţii la alge prezintă două faze diferite din punct de vedere
citologic şi fiziologic: haploidă şi diploidă.
Faza haploidă (n) este reprezentată de gametofit (generaţia sexuată, producătoare de gameţi), este
precedată de diviziunea reducţională (meioza), iar faza diploidă (2n) este reprezentată de sporofit (generaţia
asexuată, producătoare de spori) şi este precedată de fecundaţie.
Datorită faptului că, în ciclul de dezvoltare al organismelor care prezintă sexualitate, cele două faze
succed cu regularitate şi se generează reciproc, acest fenomen este cunoscut sub numele de alternanţă de
generaţii sau faze ale ciclului de dezvoltare.
Ţinând cont de acest aspect algele sunt grupate astfel: haplobionte, diplobionte şi haplodiplobionte.
Unele alge haplobionte includ în ciclul de dezvoltare ontogenetică două generaţii izomorfe
gametofitice şi sporofitice. Există alge haplodiplobionte includ în ciclul de dezvoltare două generaţii
heteromorfe gametofitice şi sporofitice, în care una poate fi predominantă. Algele haplodiplobionte pot
prezenta gametofit homotalic şi heterotalic. Speciile cu gametofit heterotalic prezintă gametofit bărbătesc şi
femeiesc pe care se formează gameţi caracteristici.
La speciile de alge evoluate din clasa Florideophyceae (Rodophyta) ciclul de dezvoltare este de tip
haplodiplobiont care include trei generaţii (trigenetic). Aceste specii prezintă o fază haploidă (gametofitul) şi
o fază diploidă cu două generaţii sporofitice independente: carposporofitul şi tetrasporofitul. Carposporofitul
formează spori asexuaţi care se numesc carpospori, iar tetrasporofitul produce spori asexuaţi care se numesc
tetraspori.
Morfologia şi evoluţia talului la alge. Tipul de tal şi modul de organizare prezintă caracteristici
specifice grupului considerat, de la forme solitare, până la forme coloniale şi pluricelulare. Algele care
prezintă structură simplă sunt unicelulare mobile (monade) sau imobile. Algele considerate evoluate au o
structură parenchimatică. Organizarea structurală a acestor organisme este corelată cu treapta de evoluţie
corespunzătoare.
Diversitatea morfologică a algelor evidenţiază faptul că formele mai puţin evoluate sunt unicelulare,
flagelate, fără perete celular. Din acest grup au evoluat pe de o parte algele rizopodiale, iar pe de altă parte
formele monadale. Algele rizopodiale includ reprezentanţi care prezintă stadii ameboidale şi plasmodiale.
Talul ameboidal este unicelular, uninucleat, lipsit de perete celular, prezentând prelungiri
protoplasmatice numite pseudopode.
Talul plasmodial este unicelular, plurinucleat, fără perete celular, consecință a anulării citochinezelor
şi fără posibilitatea de a forma pseudopode datorită creşterii consistenţei protoplasmei.

Din talul unicelular imobil şi/sau monadal a evoluat talul cenobial, iar din acesta talul colonial.
Cenobiul este alcătuit din celule formate din diviziunea aceleiaşi celule mamă şi rămân asociate
datorită mucilagiului, care rezultă din gelificarea peretelui celulei mamă. Celulele din cenobiu pot trăi
independente, dacă se desprind de acesta.
Coloniile realizează trecerea de la protofitele unicelulare la talofitele pluricelulare (Pediastrum,
Volvox). La aceste telofite se observă un început de diferenţiere a formei indivizilor, în legătură cu poziţia lor
în colonie. Colonia prezintă numeroase celule interdependente, între care se stabilesc numeroase legături
funcţionale. Celulele din colonie sunt legate unele de celelalte prin plasmodesme şi nu pot trăi independente.
Numărul celulelor din colonie diferă, în funcţie de specie.
La speciile genului Volvox, colonia este formată din aproximativ 15.000–20.000 de celule biflagelate,
diferenţiate din punct de vedere morfologic şi funcţional. Celulele formează peretele unei sfere goale, incluse
într-o masă gelatinoasă comună şi legate între ele prin intermediul plasmodesmelor. Celulele de la polul
anterior sunt mai mari, cu stigmă la baza flagelilor, iar cele de la polul posterior sunt mai mici, cu stigma mai
depărtată de baza flagelilor. Cenobiile şi coloniile pot fi mobile sau imobile, în funcţie de prezenţa sau de
absenţa flagelilor.
Din talul monadal au evoluat formele capsale şi cocale. Talul capsal (palmeloid) se caracterizează
prin lipsa peretelui, iar celulele se găsesc incluse într-o masă gelatinoasă. Forma acestui tal depinde de
proprietăţile masei gelatinoase.
Talul cocal (cocoid) a evoluat din talul capsal. Caracteristica cea mai importantă a acestui tip de tal o
constituie apariţia peretelui propriu-zis, cu o consistenţă evidentă, la multe specii peretele celular este
impregnat cu diferite substanţe.
Din talul cocal au evoluat talul sifonal şi talul filamentos (trihal).
Talul sifonal este unicelular, lipsit de pereţi transversali, plurinucleat, are forme diferite (veziculos sau
tubular) şi este denumit cenocit.
Talul filamentos este alcătuit din celule dispuse într-un singur rând sau pe mai multe rânduri, simplu
sau ramificat. Se deosebesc următoarele categorii de tal filamentos:
tal filamentos simplu, unistratificat (Spirogyra);
tal filamentos ramificat, unistratificat (Cladophora);
tal bistratificat, bidimensional, tubulos (Ulva, Enteromorpha);
tal diferenţiat morfologic în rizoid, cauloid şi filoid, prezentând chiar anumite zone anatomice
(protectoare, corticale, medulare), alcătuite din ţesuturi adevărate (prosoplectenchime şi paraplectenchime:
Fucus, Laminaria, Cystoseira).
Studierea algelor din diferite încrengături a evidenţiat faptul că celula de tip monadă cu flageli poate să
piardă uşor flagelii şi să se transforme temporar în tal de tip rizopodial, iar în alte situaţii în tal de tip capsal,
respectiv cocal. De regulă este posibilă revenirea la stadiul iniţial din aceste stadii temporare după o perioadă
scurtă de timp. Atunci când perioada de timp creşte considerabil şi devine caracteristică ciclului de viaţă, iar
organismul revine în stadiul monadal doar în perioada reproducerii, atunci se consideră corespunzător tipul de
tal rezultat în urma transformărilor menţionate.
Structura filamentoasă poate evolua din tipul monadal. Acest aspect poate fi evidenţiat la germinarea
zoosporilor în filamente, iar din filamente poate apare frecvent structura lamelară. Din această cauză, stadiul
monadal poate fi considerat iniţial pentru toate celelalte tipuri de organizare a talului. Deoarece în cadrul
încrengăturilor, etapele iniţiale sunt reprezentate de structuri monadale similare, dar care diferă în funcţie de
categoria de pigmenţi care dau culoarea caracteristică sau după produsele rezultate în urma metabolismului şi

structura flagelilor, evoluţia acestora se explică prin dezvoltarea paralelă a altor tipuri de organizare din cea
de monadă.
Complexitatea organizării structurale şi trecerea la organismele pluricelulare a avut loc odată cu
pierderea mobilităţii în stadiul vegetativ. Adesea, structura monadală iniţială s-a păstrat doar în celulele cu rol
în înmulţire. În decursul procesului de evoluţie stadiul monadal s-a redus semnificativ, păstrând esenţiale doar
acele trăsături ale formelor primare. Astfel, se poate explica cum celulele monadale cu rol în înmulţire
caracteristice pentru diferite încrengături reproduc atât de strict trăsăturile organismelor iniţiale. Dezvoltarea
paralelă a diferitelor grupe de alge poate fi explicată şi ca urmare a uniformităţii mediului acvatic în
comparaţie cu mediul terestru.
ROLUL BIOLOGIC ŞI TAXONOMIC AL PIGMENŢILOR ASIMILATORI SPECIFICI
PROTISTELOR
Clasificarea organismelor în regnul Protista constituie în prezent un subiect controversat. În acest regn
sunt incluse algele care prezintă structură celulară eucariotă. Caracterele genetice, citologice şi biochimice
relevă existenţa a trei grupe de alge (rodofite, cromofite şi clorofite) şi mai multe încrengături care corespund
unor linii evolutive fundamentale. Acestea se deosebesc prin categoriile de pigmenţi asimilatori şi prin
produsul de rezervă. (Tabelul 1).
Tabelul 1
Tipuri de pigmenţi asimilatori şi produsul de depozitare caracteristic algelor eucariote (din M. Pârvu, 2003)
Încrengătura Tipuri de pigmenţi Produsul de
depozitare clorofile ficobiline caroteni xantofile
Cryptophyta a şi c ficocianină α şi β diatoxantină amidon
ficoeritrină aloxantină
Dinophyta a şi c – β diadinoxantină amidon
diatoxantină
peridinină
Chrysophyta a şi c – β diadinoxantină crizolaminarină
diatoxantină
fucoxantină
Xanthophyta a şi c – β diadinoxantină crizolaminarină
diatoxantină
heteroxantină
vaucheriaxantină
Bacillariophyta a şi c – α şi β diadinoxantină crizolaminarină
diatoxantină
fucoxantină
Phaeophyta a şi c – β diadinoxantină laminarină
diatoxantină
fucoxantină manitol, picături
de ulei zeaxantină

Încrengătura Tipuri de pigmenţi Produsul de
depozitare clorofile ficobiline caroteni xantofile
Rodophyta a şi d ficocianină α şi β luteină amidon de
floridee ficoeritrină
aloficocianină zeaxantină
floridorubină
Euglenophyta a şi b – β diadinoxantină paramilon
diatoxantină
Chlorophyta a şi b – β luteină amidon (în
cloroplast) zeaxantină
În grupul rodofitelor sunt cuprinse algele care conţin clorofile a şi d care sunt incluse în încrengătura
Rodophyta. Grupul cromofitelor include algele eucariote care conţin clorofile a şi c. Speciile din acest grup
fac parte din următoarele încrengături: Cryptophyta, Dinophyta, Chrysophyta, Bacillariophyta, Xanthophyta
şi Phaeophyta.
Algele care fac parte din grupul clorofitelor conţin clorofile a şi b şi sunt incluse în încrengătura
Euglenophyta şi Chlorophyta.
ORGANIZAREA STRUCTURALĂ ŞI EVOLUŢIA MICELIULUI ORGANISMELOR DIN
REGNUL FUNGI
Structura celulară a fungilor filamentoşi este specifică organismelor eucariote. Forma, dimensiunile şi
structurile extra– şi intracelulare (perete celular, citosol, nucleu, mitocondrii, vacuole) se observă la
microscopul fotonic. Ultrastructurile (plasmalema, reticulul endoplasmic, aparatul Golgi, ribosomii) se
observă la microscopul electronic (x 20.000).
Din punct de vedere structural, celula fungilor prezintă următoarele componente: protoplasmatice
(citoplasma, nucleul) şi neprotoplasmatice (peretele celular, vacuomul, substanţele de rezervă).
Peretele celular şi plasmalema sunt structuri care delimitează celula şi intervin în procesele biologice
fundamentale care se desfăşoară la acest nivel.
Fungii prezintă peretele celular cu formă, grosime şi structură diferită de la un grup la altul.
Componentele care intră în alcătuirea peretelui celular sunt distribuite într-o matrice amorfă. Compoziţia
chimică a peretelui celular: 80–90% polizaharide, dintre care polizaharide complexe, în combinaţie cu
albumine, lipide, acizi graşi superiori, polifosfaţi, pigmenţi ş.a. Componentele scheletice ale peretelui celular
sunt microfibrile, respectiv chitină şi celuloză (micoceluloză, hemiceluloze, caloză). Chitina este un polimer
din reziduuri de -1-4-acetilglucozamină, care reprezintă constituientul glucidic cel mai important al peretelui
celular la fungi, cu excepţia speciilor reunite în clasa Oomycetes, la care lipseşte. Polizaharidele sunt asociate
în principal cu proteine, enzime şi lipide. Compoziţia chimică a polizaharidelor este diferită în funcţie de
grupul de fungi, ceea ce evidenţiază rolul acestor componente în taxonomie.
La speciile de fungi din încrengătura Zygomycota (regnul Fungi), peretele celular este alcătuit din
chitină şi chitosan (polimer al D-glucozaminei), iar cele din clasa Chytridiomycetes (regnul Fungi) prezintă
peretele celular alcătuit din chitină şi glucan.
În cazul speciilor incluse în încrengăturile Ascomycota, Basidiomycota, inclusiv fungii conidiali,
peretele celular este alcătuit din chitină şi glucan. La speciile din familia Saccharomycetaceae (Ascomycota)
cantitatea de chitină din peretele celular este mai mare în comparaţie cu alte specii de fungi ce conţin polimeri
ai manozei (Gorlenko, 1990).
Citoplasma este constituită din citosol şi organite citoplasmatice. Prezintă o reţea de proteine fibrilare

care formează citoscheletul, care asigură forma şi rigiditatea celulei. Hialoplasma sau matricea citoplasmatică
reprezintă substanţa fundamentală a citoplasmei, componentă structural funcţională prezentă în toate celulele
vii, localizată în spaţiul celular în afara compartimentelor delimitate de membranele intracelulare. Este
alcătuită dintr-o fracţiune solubilă (citosol), citoschelet şi o reţea de filamente (reţea microtrabeculară), care
interconectează elemente de citoschelet şi organitele celulare în citoplasmă.
Granulele de glicogen se prezintă sub forma unor corpusculi sferici reprezentând principala substanţă
de rezervă a celulei fungilor. Evidenţierea intracelulară se face prin suspendarea celulelor fungilor în soluţie
de Lugol, iar în prezenţa iodului, glicogenul se colorează brun–roşcat.
În celula fungilor se găseşte un număr variabil de nuclee, în funcţie de tipul miceliului şi stadiul
dezvoltării ontogenetice. Forma acestora poate fi diferită (sferică, ovală), iar dimensiunile variază între 1 µm
şi 20 µm. Nucleul prezintă membrană nucleară, nucleoplasmă şi unul sau mai mulţi nucleoli, acid
dezoxiribonucleic, precum şi 2 până la 16 cromosomi.
Numărul nucleelor din celule variază cu grupul de fungi şi cu stadiul de dezvoltare ontogenetică:
plasmodiul şi sifonoplastul conţin un număr mare de nuclee; miceliul primar haploid conţine un singur
nucleu, iar miceliul secundar de la fungii din încrengătura Ascomycota şi Basidiomycota este dicariotic, de
aceea în fiecare celulă se găsesc câte două nuclee care nu au fuzionat după contopirea citoplasmelor.
Corpul vegetativ al fungilor se numeşte miceliu (gr. mykes = ciupercă; lium = excrescenţă) şi este
caracteristic în funcţie de grupa taxonomică la care aparţin. Acesta este alcătuit din filamente septate şi
neseptate, denumite hife.
Hifele sunt filamente microscopice, ramificate, alcătuite dintr-un perete celular şi o cavitate (lumen)
care conţine sau este delimitată de citoplasmă. În cursul dezvoltării ontogenetice, miceliul trece prin două
stadii succesive: vegetativ (miceliul, corpul sau talul fungilor) şi fertil (sporomii, corpurile sporifere sau
carpoforii), cu structură şi formă diferită în funcţie de specie. Hifele îşi au originea într-un tip de
structură reproductivă, cel mai frecvent într-un spor care, în condiţii favorabile, formează un tub de
germinaţie. Acesta se alungeşte, se ramifică şi se transformă într-o hifă. Grosimea hifelor este variabilă, de la
0,5 la 100 µm, cel mai frecvent între 2 şi 10 µm.
Miceliul este de tipuri diferite, în funcţie de poziţia taxonomică a acestora în sistemul de clasificare al
fungilor:
1) gimnoplastul (gr. gymno = nud, neacoperit; plastum = modelat) – reprezentat printr-o celulă
amoeboidă, cu citoplasmă şi nucleu, lipsită de perete individualizat la speciile din clasa
Chytridiomycetes;
2) plasmodiul (gr. plasma, plasmodium = citoplasmă, protoplasmă; plastum = modelare) – reprezintă o
asociere de gimnoplaşti, cu citoplasmă şi numeroase nuclee, fără perete celular; există următoarele
categorii de plasmodii: extracelular, liber (saprofit) la speciile din clasa Myxomycetes şi endocelular
(parazit) la speciile din clasa Plasmodiophoromycetes;
3) sifonoplastul (gr. siphon = tubular, tubulos; plastum = modelare) – reprezentat prin filamente
neseptate, unicelulare, cu structură cenocitică (cu numeroase nuclee), la speciile reunite în regnul
Chromista şi încrengătura Zygomycota din regnul Fungi:
4) dermatoplastul (gr. derma = piele; plastum = modelare) – reprezentat printr-o celulă cu perete
individualizat; formează celule fiice prin înmugurire sau se divid şi pot rămâne asociate formând
pseudomicelii la speciile din familia Saccharomycetaceae;
5) cladomul (gr. cladus = ramură, aici cu sensul de ramificat) – reprezentat printr-un ax care este
prevăzut cu pleuridii, la speciile incluse în ordinul Laboulbeniales (încrengătura Ascomycota);
6) miceliu ramificat, septat, alcătuit din filamente sau hife (gr. hypha = pânză, aici filament), cu creştere
apicală, adesea cu ramificare laterală, uneori dichotomică, la speciile reunite în încrengăturile
Ascomycota şi Basidiomycota;
a) hifele se pot anastomoza (gr. anastomosis = joncţiune, împletire, reunire), formând o pâslă
deasă, hialină, albicioasă sau colorată diferit;

b) hifele se pot dezvolta fie în substrat având rol de absorbţie a substanţelor nutritive, fie la
suprafaţă formând organe de sporulare;
c) miceliul septat poate fi:
ramificat la specii de ascomicete şi de basidiomicete;
masiv în sporomi (basidiom), la specii reunite în clasele Hymenomycetes şi
Gasteromycetes;
d) structura septelor poate fi:
simplă – cu un por central la specii de ascomicete şi ocazional la specii de basidiomicete;
doliporă – cu un por tubular lărgit, diafragmatic, care asigură continuitatea citoplasmei în
miceliu, la specii de basidiomicete;
e) creşterea hifelor are loc prin diviziunea celulelor situate apical (la unele specii pot atinge câţiva
metri).
ROLUL BIOLOGIC ŞI TAXONOMIC AL SPORILOR SPECIFICI FUNGILOR
În cazul fungilor, termenul de spori vizează categorii de propagare, reproductive sau conservative de
mici dimensiuni, care sunt formate dintr-o hifă a miceliului vegetativ sau dintr-o celulă specială sporogenă,
din care se pot dezvolta independent hife mai mult sau mai puţin ramificate.
Sporii pot fi unicelulari sau sunt formaţi dintr-un număr mic de celule asociate care îşi păstrează
individualitatea. Prezenţa în ciclul de viaţă al aceleiaşi specii a două sau a mai multe forme morfologice
independente, fiecare prezentând modalităţi şi structuri reproductive proprii, este o caracteristică a unor fungi,
cunoscută sub numele de pleomorfism sau polimorfism.
Structuri de reproducere asexuată. Reproducerea asexuată este caracteristică anamorfelor. Acest tip
de înmulţire poate avea loc vegetativ şi prin spori – structuri reproductive diferenţiate (asexuată propriu-zisă).
Structurile de reproducere vegetativă sunt reprezentate prin porţiuni nespecializate ale aparatului
vegetativ care, diseminate pe diferite căi, pot genera un nou miceliu. Astfel, există foarte multe specii care se
înmulţesc uşor: Rhizoctonia solani (specie răspândită în sol); Agaricus bisporus, specie comestibilă introdusă
în cultură, care se înmulţeste prin fragmente de miceliu.
Artrosporii (gr. arthro, arthrus = articulat; spora = sămânţă; aici spor) se formează prin fragmentarea
miceliului în dreptul septului, atât terminal, cât şi intercalar. După separare aceşti spori au tendinţa de a se
aşeza în zig–zag (Geotrichum). Artrosporii terminali apar în urma septării în succesiune bazipetală, a părţii
distale a hifei.
Clamidosporii (gr. chlamydo, chlamydeus, chlamys = învelit, acoperit; spora = sămânţă; aici spor) se
formează în culturi mai vechi decât restul talului (Mucor) sau în interiorul conidiilor pluricelulare (Fusarium).
Clamidosporii pot avea rolul de structuri de propagare sau de rezistenţă. În mod obişnuit se formează
intercalar, mai rar terminal, prin modificarea celulelor miceliului vegetativ.
Reproducerea asexuată propriu-zisă se realizează prin mitospori, cu rol de înmulţire şi de propagare,
care se diferenţiază fie direct pe miceliul haploid sau dicariotic, fie pe ramificaţiile acestuia. Mitosporii
rezultă în urma diviziunii tipice a nucleelor, iar formarea acestora nu este precedată de un proces sexual.
Acest stadiu în care se formează mitosporii (sporii asexuaţi) este cunoscut sub numele de anamorf sau
„imperfect”. Structuri reproductive asexuate fac parte din ciclul de viaţă al fungilor şi se identifică cu
anamorfele acestora.
În unele cazuri mitosporii pot avea şi rol de rezistenţă (spori durabili), sau funcţie sexuală, fiind
organe de fecundare (spermatii).
După posibilităţile de mişcare, sporii sunt de două feluri: spori imobili sau aplanospori şi spori mobili
sau planospori, care prezintă în general 1–2 flageli.
După locul de formare sporii pot fi de două feluri: endogeni şi exogeni. Sporii endogeni (endospori)
se formează într-un număr foarte mare în celule speciale sau structuri sporogene, iar sporii exogeni (exospori)
se formează la exterior, pe filamente specializate care se diferenţiază pe miceliu.

Structurile specializate purtătoare de celule sporogene şi de spori sunt cunoscute şi sub numele de
sporofori, au o creştere limitată sau mult mai încetinită decât hifele vegetative.
Limitarea creşterii sporoforilor este însoţită de formarea unor structuri caracteristice, distincte de
hifele miceliului vegetativ, cu o mare valoare în caracterizarea şi determinarea unor taxoni. Sporoforii simpli
sunt ramificaţii ale hifelor somatice care, ajunse la o lungime strict determinată pentru o specie, formează
celule sporogene şi spori caracteristici. În mod obişnuit, sporoforii sunt erecţi şi, adesea, distinct ramificaţi.
Sporoforii simpli, care poartă sporangi sunt caracteristici speciilor din regnul Chromista, respectiv
încrengătura Zygomycota şi poartă numele de sporangiofori.
Sporii endogeni sau endosporii se formează în sporangi sau sporocişti, diferentiaţi tenninal sau
intercalar pe miceliul vegetativ. Sporangii pot fi sesili sau pot fi dispuşi pe un sporangiofor sau sporocistofor
distinct. Endosporii au rol în propagarea speciilor din încrengăturile Myxomycota, Chytridiomycota şi
Zygomycota.
În funcţie de posibilităţile de mişcare, endosporii pot fi imobili (aplanospori) sau mobili (planospori).
Zoosporii sunt mobili şi se formează în zoosporangi (zoosporocişti) cu forme diferite: sferici
(Pythium, Albugo), cilindrici (Saprolegnia, Aphanomyces, Leptomytus), fusiformi (Achlya) etc.
Sporoforii care formează conidii, sunt specifici ascomicetelor şi se numesc conidiofori. De obicei,
sporoforii îşi încetează creşterea odată cu formarea celulelor sporogene şi a sporilor. Există însă şi sporofori
cu creştere şi formare continuă de spori.
Sporoforii compuşi sunt structuri stromatice sau pseudostromatice, formate din asocieri de hife cu
creştere strict coordonată pe care se formează agregate de celule sporogene şi de spori.
Sporoforii compuşi, care formează spori asexuaţi (conidii), sunt cunoscuţi sub numele de conidiom.
Conidiomii reprezintă structuri de reproducere asexuată pentru majoritatea anamorfelor.
Conidiile propriu-zise sunt exospori haploizi (haploconidii), care se diferenţiază direct pe miceliu sau
pe structuri speciale, denumite conidiofori.
La unele specii conidiile sunt unicelulare (amerospore), la altele sunt bicelulare (didimospore) sau
multicelulare. Prezenţa septelor le împarte într-un număr mai mare sau mai mic de celule.
Conidiile bi sau multicelulare pot fi alungite, doar cu septe transversale (fragmospore), pot fi
muriforme, cu septe transversale şi longitudinale (didimospore), stelate, cu 3 sau mai multe braţe
(staurospore), alungite şi răsucite în spirală (helicospore) etc.
Fiecare din aceşti termeni au denumiri speciale pentru a indica culoarea sporilor: hialo= (hyalo=)
pentru spori hialini (fără culoare) şi feo= (phaeo=) pentru categorii de spori cu peretele celular de culoare
galbenă sau brună.
Conidiile unicelulare pot avea forme variate: sferică, ovată, obovată, piriformă, obpiriformă,
elipsoidală, cilindrică, oblongă, alantoidă, filamentoasă, falcată, fusiformă etc. Suprafaţa externă a peretelui
conidiilor poate fi netedă sau ornamentată: echinulată, verucoasă, reticulată, striată, alveolată, punctată etc. La
unele specii conidiile sunt prevăzute cu unul sau mai mulţi apendici. Conidiile pot germina imediat după ce
au fost eliberate de pe conidiofor, formând un filament de germinaţie care, alungindu-se, va forma un nou
miceliu. În cazul conidiilor pluricelulare, fiecare celulă germinează independent. Atunci când conidioforii nu
se deosebesc sau diferă foarte puţin de hifele vegetative pe care se formează, se numesc micronematici.
Atunci când sunt complet diferiţi ca aspect de hifele somatice, se numesc conidiofori macronematici. Rareori
conidioforii sunt simpli. Cel mai adesea, conidioforii se ramifică monopodial, simpodial, dicotomic sau în
verticil. De asemenea, conidioforii pot fi septaţi sau neseptaţi, hialini sau divers coloraţi.
Adesea, conidioforii sunt asociaţi în sporofori compuşi. Aceste agregate se pot forma la suprafaţa
substratului (coremii şi sporodochii) sau sunt organizate în diferite tipuri de strome, localizate superficial
(acervuli) sau profund (picnidii) în diferite ţesuturi ale organismelor gazdă.
Coremiile (gr. coremium = mătură), cunoscute şi sub numele de sinemata, sunt sporofori compuşi care
prezintă conidioforii grupaţi în fascicule compacte, cu extremităţile superioare libere. Partea bazală a

conidioforilor este sterilă şi are rol de susţinere a conidiilor care se formează terminal. Coremiile sunt
caracteristice genurilor Graphium, Isariopsis, Podosporium etc.
Sporodochiile (gr. sporodochium = recipient) au aspectul unor mici ridicături (mameloane) formate
dintr-o bază stromatică pe care se diferenţiază conidioforii şi conidiile (Monilia fructigena, M. laxa; specii din
genul Tubercularia, Ramularia, Fusarium etc.).
Acervulii (lat. acervulus = grămadă) se formează subepidermal sau subcuticular şi au aspectul unor
strome sau pseudostrome, pe care se formează conidiofori de obicei scurţi şi conidii cu forme, dimensiuni şi
culori diferite. Sunt caracteristice genurilor Colletotrichum, Gloeosporium, Melanconium, Coryneum etc.
Picnidiile (gr. pycnos = dens, compact) sunt structuri compacte, globuloase sau oval-alungite,
localizate în mod obişnuit în substrat. Forma caracteristică este menţinută de o structură externă
pseudoparenchimatică care se numeşte peridie, cu rol de apărare, iar pe partea internă se diferenţiază
conidiofori simpli sau ramificaţi, dispuşi într-un strat palisadic sau himenial. Conidiile se numesc fie
picnospori, atunci când celulele au formă sferică sau ovală, fie stilospori, atunci când celulele sunt
filamentoase unicelulare sau multicelulare, hialine sau pigmentate.
Conidiile pot fi eliberate din picnidie fie prin ruperea / putrezirea peridiei, fie prin ostiol. Conidiile se
găsesc într-o masă gelatinoasă răsucite sau încolăcite, cu aspectul unor cârcei, denumite cirri.
În anumite cazuri aceste conidii pot participa la fecundaţie şi sunt cunoscute sub numele de spermatii
(Uredinales, Basidiomycota).
Picnidiile sunt frecvent întâlnite în ciclul de viaţă al speciilor din genurile: Phyllosticta, Phoma,
Cytospora, Ascochyta, Septoria etc.
Structuri de reproducere sexuată. Reproducerea sexuată se realizează prin intermediul meiosporilor.
Din această categorie fac parte diferite tipuri de spori: sporii (mixosporii) de la mixomicete, oosporii de la
oomicete, zigosporii de la zigomicete, ascosporii de la ascomicete şi basidiosporii de la basidiomicete.
Oosporii şi zigosporii rezultă din fuziunea unor nuclee compatibile cu polaritate sexuală diferită. La
unele specii aceste categorii de spori nu se formează în corpuri sporifere diferenţiate.
Sporii mixomicetelor se formează în interiorul sau la exteriorul unor formaţiuni sporifere (sporofori),
denumite sporangi (mixosporangi), care au forme, structuri şi culori diferite, caracteristice speciei. Cele mai
multe mixomicete prezintă sporangi simpli, multispori, cu sporangi protejaţi de peridie. Există specii la care
plasmodiul în întregime se transformă într-un sporofor compus, cu o structură masivă, denumit etaliu, în care
se formează numeroşi sporangi ce conţin spori.
Zigosporii sunt spori de dimensiuni mari (100–200 µm) cu suprafaţa rugoasă, rezistenţi la condiţiile
nefavorabile de mediu. Prin copulare la zona de contact se formează progametangele, iar prin liza peretelui
celular rezultă zigosporul diploid (Mucor, Rhizopus).
Oosporii se formează întotdeauna endogen prin oogamie (o formă evoluată de heterogametangiogamie).
Gametangiul femeiesc se numeşte oogon şi conţine una sau mai multe oosfere, iar gametangiul bărbătesc se
numeşte anteridie şi conţine anterozoizi. După ce oosferele sunt fecundate de anterozoizi, rezultă o celulă
diploidă, cu un înveliş gros cu rol de apărare care se transformă în oospori.
Ascosporii sunt endogeni şi se formează în mod obişnuit câte 8 (rar 2 sau 4) într-o celulă sporogenă
care se numeşte ască.
Basidiosporii sunt exogeni şi se formează în mod obişnuit 4 (uneori 2 sau mai mulţi) pe basidii.
Ascele şi basidiile sunt meiosporocişti, deoarece formarea acestora întotdeauna este precedată de
procesul sexual prin care are loc trecerea de la haplofază la diplofază. În aceste celule are loc cariogamia
(fuziunea nucleelor), diviziunea reducţională (meioza) şi sporogeneza (diferenţierea sporilor sexuaţi
caracteristici). Cu excepţia drojdiilor, la care ascele se formează holocarpic, toate celelalte ascomicete şi
bazidiomicete sunt eucarpice.

În mod obişnuit ascele şi basidiile sunt asociate, de cele mai multe ori dispuse în palisadă, formând un
strat fertil, denumit himeniu, care se formează în sau pe sporomi. După caz, aceste structuri reproductive se
numesc ascomi sau ascocarpi pentru ascomicete şi basidiomi sau basidiocarpi pentru basidiomicete, iar
organismele respective carpomicete. Există unele excepţii, la care ascele (Taphrinales) şi basidiile
(Exobasidiales) se găsesc liber pe substrat.
Ascomii şi basidiomii sunt formaţiuni stromatice sau semistromatice care conţin straturi de celule
sporogene, asociate uneori cu celule sterile. Aceste straturi sporifere se formează la suprafaţa sau în diferite
tipuri de cavităţi ale sporomului. Sporomii reprezintă agregări de hife, care pot fi monocariotice la ascomicete
sau dicariotice la bazidiomicete, uneori şi heterocariotice. Stadiul diploid este limitat, fiind reprezentat de
nucleele rezultate în urma cariogamiei, care are loc în celulele zeugite ale sporomilor. Aceste celule terminale
sunt considerate celulele mamă ale ascelor la ascomicete şi probasidiile la bazidiomicete. De obicei, acestea
se formează în număr mare, aproape concomitent şi la acelaşi nivel al ascomilor şi bazidiomilor, alcătuind
straturi sporogene sau himeniale.
În funcţie de forma, culoarea şi structura ascomilor poartă denumiri specifice: cleistotecie, peritecie şi
apotecie.
Cleistoteciul (gr. cleisto = închis; rămas nedeschis; thecium = cutie) reprezintă un ascom complet
închis, de unde ascele cu ascospori sunt eliberate în mod pasiv, prin putrezirea peridiei. Morfologia acestor
organe este caracteristică speciilor din ordinul Erysiphales.
Periteciul (gr. peri = care înconjoară; thecium = cutie) reprezintă un ascom care prezintă o deschidere
(ostiol) pe unde sunt eliminate în mod activ ascele cu ascosporii atunci când au ajuns la maturitate.
Morfologia acestor organe este caracteristică speciilor din ordinul Pleosporales.
Apoteciul (gr. apo = fără; thecium = cutie) reprezintă un ascom cu forme caracteristice (disc, cupă,
farfurioară) şi mărimi diferite, de unde ascele şi ascosporii sunt eliminaţi în mod activ. Morfologia acestor
organe este caracteristică speciilor din ordinul Pezizales
REGNUL PLANTAE
Plantele sunt organisme eucariote imobile, primar fotosintetizatoare, pluricelulare, în al căror ciclu de
viaţă se succed două generaţii distincte: haploidă (gametofitul) şi diploidă (sporofitul). Din acest grup de
organisme fac parte muşchii şi plantele vasculare (cormofitele).
O serie de dovezi (morfologice, structurale, biochimice) arată că plantele au evoluat dintr-un grup de
alge verzi străvechi. Ca şi algele verzi, plantele conţin clorofile a şi b, şi carotenoizi ca pigmenţi accesorii, iar
amidonul (ca principal produs de fotosinteză) este depozitat în interiorul cloroplastelor (la alte organisme
eucariote fotosintetizatoare produşii de rezervă sunt depozitaţi în afara cloroplastelor). De asemenea,
principalul component al pereţilor celulari la plante şi la unele alge verzi este celuloza. Datorită
caracteristicilor comune pe care le prezintă, este foarte posibil ca plantele să descindă dintr-un strămoş algal
comun sau din diferite grupe de alge verzi înrudite, care au ocupat cu succes uscatul.
La plantele inferioare, sporofitul embrionic diploid este dependent de gametofitul haploid pentru o
perioadă mai lungă sau mai scurtă, în timp ce la grupele evoluate gametofitul devine dependent de sporofitul
matur pentru nutriţie. Una dintre generaţii (sau ambele) pot prezenta creştere nedefinită.
Plantele reacţionează lent la stimuli, prin creşterea ţesuturilor sau a unor celule individuale (tropisme,
mişcări nastice).
Organismele care au realizat tranziţia de la apă la uscat au dezvoltat structuri care le protejează de
uscăciune. Aceste organisme şi-au format un înveliş steril (perete) protector în jurul celulelor gametice din
gametangii (anteridii şi arhegoane) şi a celulelor producătoare de spori (arhesporiului) din sporange. De
asemenea, embrionii plantelor sunt protejaţi în interiorul arhegonului. Părţile supraterane ale plantelor sunt
acoperite cu cuticulă, care le protejează faţă de uscăciune. Prezenţa cuticulei este strâns legată de existenţa
stomatelor, care funcţionează, în principal, ca reglator al schimburilor de gaze.
SUBREGNUL CORMOBIONTA (TRACHEOBIONTA)

- Sunt organisme tipic terestre şi numai puţine sunt adaptate secundar la mediul acvatic;
- Sunt plante, în majoritate, autotrofe, pluricelulare, având aparatul vegetativ constituit din organe diferenţiate
nu numai morfologic ci şi anatomic, în rădăcină, tulpină şi frunze, alcătuind cormul, de unde şi numele de
cormofite;
- Cormul are o structură complexă, distingâdu-se prin prezenţa ţesuturilor conducătoare (liberiene şi
lemnoase) situate în cilindrul central numit stel, de aceea se mai numesc plante vasculare;
- Sunt plante diplobionte, cu alternanţă de faze obligatorie: faza sexuată – gametofaza, de scurtă durată şi faza
asexuată – sporofaza, care domină, deci planta reprezintă sporofitul;
- Organul de reproducere sexuată femel caracteristic numit arhegon este pluricelular, ca şi la briofite, de aceea
aceste plante se mai numesc arhegoniate;
- În urma fecundării, din zigot se formează un corp pluricelular, cu proprietăţi meristematice, numit embrion,
din care se vor diferenţia toate organele plantelor, de aceea aceste plante se mai numesc embriofite.
Originea algală a cormofitelor
Plantele vasculare, cormofitele, au fost precedate de talofite şi au derivat din alge. Strămoşii primelor
cormofite sunt algele verzi izomorfe, de tip Coleochaetae (Ulothrichales), cu o alternanţă de generaţii
independente, care trăiau în zonele litorale (cu flux şi reflux) ale mărilor devoniene. Aceste alge au un tal
heterotrich, ramificat, cu creştere, în general, intercalară, ale cărui ramuri sunt atenuate în prelungiri subţiri,
ca nişte peri. Ele se înmulţesc asexuat prin zoospori biflagelaţi, iar reproducerea sexuată este o oogamie
particulară. Oogonul, de o formă specială, prevăzut cu o prelungire subţire, după fecundare închide zigotul şi
se înconjoară cu filamente pluricelulare provenite din celulele vegetative învecinate. Aceste filamente
formează în jurul zigotului un înveliş protector, sugerând originea peretelui pluricelular al arhegonului.
Datorită climei calde şi umede, aceste alge s-au adaptat treptat la lipsa apei în perioada refluxului şi, cu
timpul, s-au extins în mlaştinile litorale şi apoi prin estuarele fluviilor au înaintat în interiorul continentelor, în
apele continentale. Analiza structurii celor două generaţii demonstrează că numai sporofitul a avut calităţile
necesare pentru edificarea unui organism nou, vascularizat. La algele marine mai evoluate, la care sporofitul
prefigurează elemente ale organizării plantelor vasculare, se constată o mare diversitate, o structură şi o
morfologie paralelă cu cea a plantelor vasculare. Astfel, la multe alge verzi (Ulva lactuca), brune (Laminaria
hyperborea) şi roşii (Delesseria sanguinea) sporofitul este diferenţiat în rizoid, cauloid şi filoid, asemănător
organelor vegetative ale cormofitelor. Delesseria sanguinea (Rhodophyta) are un tal în care se distinge o
parte bazală, cauloidul, peren, care poartă filoide anuale nervate, dar fără a fi dispuse cu o regularitate
filotaxică. Anthophycus longifolius (Phaeophyta) are o analogie de organizare mai pronunţată, prezentând un
cauloid purtând filoide foarte regulate, ramuri secundare cu creştere limitată, cu conceptacule, iar la bază 1 –
2 filoide. Talul la numeroase alge prezintă toate tipurile de ramificaţii, iar în vârful acestora se află fie o
singură celulă iniţială, fie un grup de celule meristematice apicale, la fel ca la cormofite. Unele alge au un
aparat conducător omolog liberului. Acesta este schiţat la Halimeda sp. şi Stilophora sp. (Phaeophyta), la care
celulele centrale sunt orientate în aşa fel încât să asigure funcţia conducătoare, iar la Alaria esculenta
(Phaeophyta) celulele centrale au pereţi ciuruiţi. La unele specii ale genului Laminaria, cauloidul prezintă la
exterior o cuticulă groasă, constituind un meristoderm, sub care se află un strat cortical extern, format din
celule mici, susceptibile de a se diviza activ, producând ţesuturi secundare, iar în interior un corp central cu o
măduvă formată din celule alungite cu pereţi transversali ciuruiţi, asemănătoare vaselor liberiene de la
fanerogame, îndeplinind rolul de conducere al substanţelor nutritive. Tot în zona medulară se află celule cu
pereţi puternic îngroşaţi, având rol mecanic, de susţinere a talului. Localizarea acestor celule în zona centrală
evocă dispoziţia protostelică. De notat că singurele elemente conducătoare existente la alge aparţin liberului,
vasele ciuruite, care apar primele în ontogenia fanerogamelor.
Existenţa unor plante vasculare în Silurian şi Devonian, al căror corp vegetativ era un adevărat tal
vascularizat (aşa cum le atestă resturile găsite în Ordovician) pare a reprezenta proba că unele alge foarte
diferenţiate morfologic şi anatomic, sunt la originea plantelor vasculare. O astfel de algă străveche ajunsă în

condiţii de mediu terestru mult mai heterogen, s-a putut adapta la acest mediu prin consolidarea şi
perfecţionarea structurii sporofitului şi prin apariţia stelului primitiv, alcătuit din vase liberiene înconjurând
celule cu pereţi lignificaţi, care prefigurează vasele lemnoase. Acest sporofit algal, a dat naştere, de-a lungul a
milioane de ani, unui corp vegetativ nou, vascularizat, din care s-a diferenţiat rădăcina, tulpina, frunzele şi
sporangii, alcătuind cormul primelor plante vasculare cunoscute în Silurian ca fosile. Prin urmare, dintre
algele verzi pluricelulare, au putut cuceri uscatul numai acele populaţii la care, după fecundarea efectuată
obligatoriu prin intermediul apei, zigotul dădea naştere la un embrion rezistent la insolaţie şi uscăciune. Din
acesta se putea dezvolta sporofitul cu posibilităţi de adaptare, care să asigure protecţia contra insolaţiei,
absorbţia continuă a apei, o cantitate mare de spori, iar gametofitul se dezvolta numai în sezonul sau locurile
umede. Adaptarea la viaţa de uscat a cormofitelor s-a petrecut prin transformarea succesivă de la simplitatea
morfologică şi primitivitatea anatomică a algelor ancestrale, spre complexitatea şi diferenţierea pe diverse
planuri a fanerogamelor.
Caracterele cormofitelor care dovedesc originea lor în algele verzi
- Au aceeaşi pigmenţi clorofilieni – clorofila a şi b, carotenii - şi şi xantofile – zeaxanthina;
- Sintetizează şi depozitează amidon intraplastidial;
- Au aceeaşi ultrastructură a nucleului, cloroplastelor şi condriozomilor;
- Au acelaşi tip de polaritate, simetrie şi ramificaţie;
- Creşterea în lungime se realizează prin diviziunea unei celule sau a unui grup de celule apicale;
- Vasele liberiene apar înaintea vaselor lemnoase, ca şi celulele floemice de la alge;
- Au aceleaşi organe morfofuncţionale, analoage şi diferite ca structură: rizoid – rădăcină, cauloid – tulpină,
filoid – frunză;
- Au acelaşi tip de alternanţă de faze, gametofaza şi sporofaza, cu predominarea sporofitului, ca şi la algele
verzi macrofite;
- Gameţii masculi la ferigi şi gimnospermele primitive (ca şi la briofite) sunt flagelaţi, ca la algele verzi.
Caracterele cormofitelor realizate prin adaptarea la viaţa terestră
- Realizarea unor organe noi, specializate la mediul terestru: rădăcina, tulpina, frunza;
- Structura acestor organe se caracterizează prin ţesuturi definitive: epiderma cu stomate, rizodermă, ţesut
conducător organizat în stel, ţesuturi mecanice (sclerenchim şi colenchim), parenchim de depozitare, ţesuturi
de excreţie;
- Gametangii realizează pereţi pluricelulari de protecţie;
- Sporangii migrează din vârful axului pe frunze;
- Apariţia staminelor (cu antere) şi ovarelor (cu ovule), ovulele transformate în sămânţă care devine organul
specific de înmulţire;
- Apariţia fecundării sifonogame etc.
Clasificarea cormobiontelor
- Polypodiophyta (Pteridophyta), plante lemnoase şi erbacee care se înmulţesc exclusiv prin spori, de aceea se
mai numesc Criptogame vasculare;
- Pinophyta (Gymnospermae, Gymnospermatophyta), plante lemnoase care se înmulţesc prin seminţe, lipsite
de fructe; flori fără înveliş floral;
- Magnoliophyta (Angiospermae, Angyospermatophyta), plante lemnoase şi erbacee care se înmulţesc prin
seminţe închise în fructe; florile prezintă, în general, înveliş floral. Cele 2 încrengături (Pinophyta şi
Magnoliophyta) sunt reunite, conform anumitor sisteme de clasificare, sub titulatura de Spermatophyta –
plante cu flori şi seminţe; înmulţirea se realizează prin seminţe şi vegetativ.
Încrengătura POLYPODIOPHYTA (PTERIDOPHYTA, Criptogame vasculare)
Caractere generale: - Majoritatea polipodiofitelor sunt plante erbacee perene, rar anuale (Selaginella sp.),
dar şi arbustive sau chiar arborescente, vasculare, cu ramificare dicotomică, rar monopodială;

- Gametofitul, liber sau inclus, foliaceu taloidic, mai rar masiv (Lycopodium sp.), sau vascularizat şi caulinar
(Psilotales), întotdeauna independent de sporofit;
- Embrionul are prima rădăcină laterală, în general endogenă;
- Rădăcinile toate sunt adventive, iar rădăcina principală lipseşte;
- Sporofitul este foarte divers, sporangii epifili, hipofili sau terminali, rar epicaulinari, dispersaţi sau mai mult
sau mai puţin grupaţi în sori sau strobili;
- Homosporia răspândită, anterozoizi ciliaţi;
- Apogamia frecventă;
- Aparatul vascular lemnos format din traheide, adesea scalariforme, rar însoţite de trahee (Selaginella sp.,
Athyrium filix-femina )
- Formaţiunile secundare sunt rare la ferigile actuale (tulpinile fertile de Equisetum sp., Botrychium sp.,
Isoëtes sp.);
- Au două generaţii heteromorfe independente, care se succed obligatoriu, cu predominarea sporofitului.
Sporofitul
Este reprezentat prin zigot, embrion, plantă şi sporangi. El se identifică cu însăşi planta vasculară care
are dimensiuni variabile, de la câţiva centimetri până la zeci de metri. Este inaugurat de zigotul diploid, situat
pe protal, care prin germinare generează embrionul, acesta fiind legat de protal printr-un masiv celular numit
talpa sau suspensorul, care-i înlesneşte nutriţia. La una din extremităţile embrionului se disting mugurele
tulpiniţei şi primordiile foliare (frunza cotiledonară), iar la cealaltă radicula. Prin creştere, embrionul
generează rădăcină, tulpină şi frunze.
Rădăcina are origine endogenă, cu rare excepţii în endoderm sau în periciclu (Lycopodiatae). Prima
rădăcină apare lateral, cea principală lipseşte, iar primele mor imediat, fiind înlocuite cu rădăcini adventive.
Rădăcina la polipodiofitele primitive lipseşte (Rhyniales, Psilotales), absorbţia având loc prin rizoizi care se
dezvoltă pe rizomoizi. Ramificarea este dicotomică sau pseudodicotomică.
Tulpina are creştere apicală şi poate fi erectă (ferigi arborescente, Lycopodiaceae), târâtoare, epigee
sau hipogee (Lycopodium, Pilularia ), unele sunt liane prin ramuri (Selaginella) sau prin frunze (Lygodium),
mai rar flotante (Salvinia). La ferigile erbacee tulpina este subterană, un rizom, iar la cele lemnoase, un stip
înalt de câţiva metri. Ramificarea este dicotomică, rar monopodială, iar la ferigile superioare este simpodial-
dicotomică.
Din punct de vedere anatomic, tulpina prezintă o epidermă (cu stomate), scoarţă (reprezentată printr-
un parenchim asimilator şi unul de depozitare şi uneori un endoderm) şi cilindru central sau stel (care uneori
are şi periciclu). În cilindrul central se află ţesutul conducător constituit din ţesut lemnos (xilem) reprezentat
prin traheide inelate şi spiralate în protoxilem şi frecvent scalariforme în metaxilem, însoţite uneori şi de
trahee (Selaginella, Athyrium filix-femina ), precum şi ţesutul liberian (floem), reprezentat numai prin tuburi
ciuruite. Meristemele secundare la ferigile actuale apar destul de rar şi atunci când ele există generează lemn
secundar. Cel mai primitiv tip de stel este protostelul (Rhyniales), dar apar şi actinostelul (Psilotales),
plectostelul (Lycopodiales), polistelul (Selaginellales) şi alte tipuri de stel.
Frunzele, numite fronde, sunt de două tipuri fundamentale: microfile cu originea în enaţiuni
epidermice şi macrofile (megafile), cu origine caulinară, provenite prin cladodificare. În general, frunzele
alcătuiesc aparatul fotosintetizator şi de aceea se numesc trofofile, cu limbul simplu, întreg sau divizat (palmat
sau penat), sau compus. Unele poartă sporangi, pierd funcţia asimilatoare şi se numesc sporofile, iar altele
sunt asimilatoare şi purtătoare de sporangi şi se numesc trofosporofile.
Sporangii au peretele pluricelular. La polipodiofitele arhaice sporangii se află în vârful tulpinilor
(Rhyniales), alteori se grupează formând sinangii (Psilotales). La ferigile evoluate sporofilele se grupează în
vârful tulpinilor, formând spice sporifere sau strobili (Lycopodiatae, Equisetatae), sau se aglomerează pe
dosul frunzelor, formând sori, uneori protejaţi de o membrană numită induzie sau induziu. După provenienţă

şi poziţie, sporangii sunt de două tipuri: eusporangi, la ferigile primitive, care se numesc şi eusporangiate, şi
leptosporangi, la ferigile evoluate, numite leptosporangiate.
Gametofitul
Este reprezentat prin spori, protal, gametangi şi gameţi şi se identifică cu protalul. El rezultă din
germinarea sporilor, rezultaţi prin diviziunea reducţională a celulelor sporogene din arhesporiul sporangelui.
Sporii au următoarele învelişuri: endosporul celulozic şi subţire şi exosporul, gros şi cutinizat; la unele
ferigi (Equisetum) mai există un al treilea înveliş numit epispor. Sporii pot fi identici – izospori, sau
diferenţiaţi pe sexe, homospori şi heterospori, microspori şi macrospori. Prin germinare, sporii dau naştere
unui masiv de celule numit protal.
Protalul produs din germinarea izosporilor şi homosporilor poartă numele de izoprotal pe care se
dezvoltă atât anteridiile cât şi arhegoanele, ferigile numindu-se homoprotaliene, iar protalul rezultat din
germinarea heterosporilor se numeşte heteroprotal, care poate fi microprotal provenit din germinarea
microsporilor şi pe care se află anteridiile, precum şi macroprotal, provenit din germinarea macrosporilor şi
pe care se dezvoltă arhegoanele. Aceste ferigi se numesc heteroprotaliene. Protalul este în general mic,
măsurând doar câţiva milimetri sau câţiva centimetri (de obicei 1-2 cm, mai rar 5 cm), cu structură
pluricelulară, cu parenchim omogen, nediferenţiat în ţesuturi. El se prezintă sub aspect lamelar (foliaceu),
cordiform sau lobat, uneori tuberculiform şi mai rar filamentos. De regulă, el este taloidic, fixându-se de
substrat cu ajutorul rizoizilor şi numai excepţional are forma unui corm vascularizat (cormoid - Psilotales),
dezvoltându-se independent de sporofit. Majoritatea polipodiofitelor au protal epigeu, verde, nutrindu-se
autotrof, însă în unele cazuri este lipsit de pigmenţi asimilatori, devenind hipogeu, nutrindu-se saprofit sau
trecând în simbioză cu ciupercile, deci micotrof. În general, protalul are o viaţă scurtă (anual), dar la unele
specii (Botrychium, Gymnograma sp.) acesta poate supravieţui mai mulţi ani (peren). La multe ferigi
(Selaginelales, Salviniaceae), protalul se dezvoltă în interiorul sporului, numit endoprotal, aceste ferigi
numindu-se endoprotaliene.
Pe protal se formează gametangiile - anteridiile şi arhegoanele. Anteridia este formată dintr-un perete
steril, pluricelular, cu celule dispuse într-un strat, care închide înăuntru anterozoizii spiralaţi pluriflagelaţi
(Polypodiatae) sau biflagelaţi (Lycopodiaceae, Selaginellaceae). Anteridia are formă sferică, la ferigile
primitive fiind împlântată în protal, în care caz peretele nu este întotdeauna distinct şi numărul anterozoizilor
este mare, iar la tipurile evoluate, anteridia este situată pe o proeminenţă, peretele fiind format numai din trei
celule, două celule inelare şi una operculară, iar numărul anterozoizilor este redus. Arhegonul are forma unei
butelii, având partea bazală mai umflată împlântată în protal şi conţine o oosferă mare, imobilă şi un gât
(trichogin) ce proliferează în exterior. Gâtul este format din patru rânduri de celule (acestea diminuâmd cu
evoluţia), iar numărul celulelor canalului gâtului este de 16 (Lycopodium) şi se reduce la una la ferigile
leptosporangiate.
Fecundarea se realizează prin intermediul apei, deci este zoidiogamă.
Ciclul de dezvoltare
În ciclul de dezvoltare al pteridofitelor se succed obligatoriu două generaţii heteromorfe, care îşi duc
viaţa independent, sporofitul şi gametofitul. Generaţia sporofitică este dominantă, fiind reprezentată prin
planta propriu-zisă, care începe cu zigotul diploid. Acesta, prin diviziuni mitotice repetate dă naştere
embrionului, iar acesta la planta pe care se dezvoltă sporangii. Generaţia gametofitică începe cu formarea
sporilor, prin diviziunea reducţională a celulelor sporogene din sporange. Generarea sporului dă naştere la
protal pe care se dezvoltă anteridiile şi arhegoanele ce vor produce anterozoizii şi oosferele. În urma
fecundării rezultă zigotul, diploid. Ca durată şi ontogenie predomină sporofaza, deci polipodiofitele sunt
plante diplonte.
Polipodiofitele se înmulţesc şi vegetativ prin rizomi, stoloni, tuberculi, tuberuli, frunze sau protal.
Uneori butăşesc sau marcotează prin formarea de rădăcini adventive la nodurile ramurilor sau chiar în vârful
frunzelor.
Clasa PSILOPHYTATAE (PSILOPSIDA)

Sunt cele mai vechi şi mai primitive ferigi, cele mai multe cunoscute numai în stare fosilă, care s-au
dezvoltat din Silurian până la începutul Carboniferului şi dintre care au supravieţuit până în zilele noastre
numai două genuri. Psilofitatele actuale sunt plante mici erbacee sau arbustive, cu ramificaţie dicotomică,
lipsite de rădăcini şi de frunze veritabile. Tulpina are stelul predominant de tip protostel, dar şi actinostel şi
sifonostel. Sunt plante izosporee, cu spori formaţi în sporangi uniloculari situaţi în vârful ramurilor şi
tulpinilor sau sporangi triloculari (sinangii), proveniţi dintr-un grup de celule subepidermice (eusporangi).
Clasa LYCOPODIATAE (LYCOPSIDA)
Sunt polipodiofite cu sporofitul un corm, alcătuit din rădăcină, tulpină şi frunze. Sunt erbacee şi
lemnoase (arbuşti sau arbori), cu tulpinile ramificate dicotomic, purtând microfile uninerve, spiralate sau
opuse. Sporangii sunt situaţi, de regulă, pe faţa adaxială a sporofilelor, rareori direct pe tulpină. Uneori
sporofilele sunt dispersate în vârful tulpinilor, iar alteori se grupează în vârful acestora formând spice
sporifere (strobili). Sporangii au originea într-un grup de celule subepidermice (tipul eusporangiat), cele mai
primitive sunt izosporee, iar cele mai evoluate – heterosporee. Protalul este epigeu sau hipogeu, adesea
micotrof, fiind de tip izoprotal sau heteroprotal (micro şi macroprotal), uneori endoprotal.
Clasa EQUISETATAE (SPHENOPSIDA, ARTICULATAE)
Equisetatele sunt actuale şi fosile: cele actuale sunt plantele erboase mici, în comparaţie cu cele
dispărute, care erau lemnoase, sub formă de arbori, arbuşti şi liane, arborii atingând până la 1 m în diametru.
Tulpinile şi ramurile sunt ramificate monopodial, (caracter care apare pentru prima dată), sunt articulate,
formate din internoduri lungi, canaliculate, separate de noduri scurte, de aici şi numele de Articulatae.
Anatomia tulpinii variază de la protostel şi sifonostel la artrostel, la care apare cambiul cilindric, care
generează ţesuturi secundare. Frunzele în general sunt microfile, de obicei îngustate spre bază, în formă de ic
(pană) dispuse în verticil la noduri, sau se unesc într-un manşon în jurul tulpinilor şi ramurilor, având la vârf
numeroşi dinţişori, formând vagina, neasimilatoare, caz în care funcţia de asimilaţie este preluată de tulpină.
La articulatele primitive sporangii sunt dispuşi în vârful sporofilelor ramificate dicotomic, iar la cele actuale
sporofilele sunt grupate în vârful tulpinii sau ramurilor în spice sporifere sau strobili. Ele sunt homosporee,
rar izosporee sau heterosporee.
Clasa POLYPODIATAE (FILICATAE, PTEROPSIDA, FILICOPSIDA)
Rădăcinile primare dispar de timpuriu, fiind înlocuite cu rădăcini secundare adventive, diarhe, mai
rar monarhe sau poliarhe. Tulpina majorităţii ferigilor actuale este subterană, perenă, un rizom de culoare
brună, prevăzut cu solzi şi resturile tecilor frunzelor, de pe care pleacă rădăcini adventive. Anatomic, această
tulpină este alcătuită dintr-o scoarţă parenchimatică, în care se află fie un strat sclerenchimatic, fie mai multe
cordoane sclerenchimatice şi din cilindru central de tip sifonostel, solenostel, polistel etc., cu vase lemnoase
de tip traheide scalariforme şi mai rar în amestec cu trahee (Athyrium filix – femina). Tulpina ferigilor fosile şi
a celor arborescente actuale este un stip. Ramificarea tulpinii este complexă, derivând însă din dicotomia
primitivă; ramificarea monopodială este foarte rară. Formaţiunile secundare sunt foarte rare (Zygopteridales,
Ophyoglossales etc.). Frunzele sunt macrofile, întregi, lobate, sectate sau compuse, cu nervatură dicotomică,
cu creştere terminală şi prefoliaţie circinată, exceptând frunzele ferigilor străvechi care erau caulinare,
formate din ramificaţii subţiri, numite pinule sau aveau forma unor foliole mici. Frunzele ferigilor se mai
numesc fronde şi sunt de trei categorii: trofofile, verzi, asimilatoare, sporofile, galben- brunii, purtătoare de
sporangi şi trofosporofile, verzi, asimilatoare, care poartă şi sporangi. Sporangii ferigilor primitive erau axiali,
situaţie care se mai păstrează şi la ferigile actuale din familiile Osmundaceae, Hymenophyllaceae,
Ophyoglossaceae etc. La ferigile evoluate, sporangii au o poziţie marginală şi hipofilă, de obicei grupaţi în
sori. După originea sporangilor, ferigile sunt eusporangiate şi leptosporangiate. Majoritatea ferigilor sunt
izosporee, cu sporangi pedicelaţi şi cu pereţi unistratificaţi (la leptosporangiate) şi pluristratificaţi (la
eusporangiate). Peretele sporangelui leptosporangiatelor prezintă un inel mecanic incomplet, alcătuit din
celule cu pereţi laterali şi bazali inegal îngroşaţi, care contribuie la deschiderea sporangilor. Prin uscare, inelul
mecanic se contractă prin pereţii neîngroşaţi, creând o tensiune în întreg inelul, iar porţiune din sporange
rămasă fără inel mecanic crapă şi sporii sunt puşi în libertate. Protalul este anual sau peren, fixat de substrat

cu rizoizi, de dimensiuni de până la 1 cm, având forma unei lame cordate pluristratificate, tuberculiform,
filamentos etc. În general protalul este epigeu, autotrof, dar sunt şi exemple la care protalul este hipogeu,
micotrof (Ophyoglossales) sau nutrindu-se saprofit. Pe partea inferioară a protalului se află anteridiile care
produc anterozoizi spiralaţi şi pluriflagelaţi şi arhegoane cu oosfere. Înmulţirea vegetativă se realizează prin
butaşi de rădăcini (Ophyoglossum), stoloni supraterani (Nephrolepis sp.), frunze (Asplenium), bulbile epifile
(Cystopteris, Asplenium), prin fragmente de protal etc.
Subclasa PROTOPTERIDIDAE
Sunt cele mai primitive ferigi fosile cunoscute din Devonian. Ele purtau pe tulpini pinule mici, verzi,
filiforme, dispuse terminal sau serial, numite filofore. Sporangii situaţi la extremitatea ramurilor, erau de tip
eusporangiat, cu numeroşi izospori. Din aceste protopterididae arhaice s-au desprins specii care pe lângă
filofore aveau şi foliole mici, iar sporangii erau situaţi pe sporofile.
Subclasa ARCHAEOPTERIDIDAE
Plante fosile lemnoase (arbori şi arbuşti) cu trunchiuri groase şi drepte cu vârful ramificat. Tulpina
creştea în grosime, formându-se lemn cu traheide areolate, asemănătoare cu cele ale gimnospermelor.
Frunzele erau variate: filofore, foliolate sau macrofile penat sectate.
Subclasa COENOPTERIDIDAE
Ferigi paleofitice, lemnoase, izosporee şi heterosporee, prevăzute cu filofore sau cu frunze mici. În
vârful ramificaţiilor se aflau sporangii mari, apicali, dehiscenţi, liberi sau uniţi.
Subclasa OPHYOGLOSSIDAE
Ferigi fosile şi actuale, cele actuale erbacee, având în sol un rizom de pe care se ridică o axă purtând o
singură frunză, sau un mic număr de frunze, fiecare alcătuită din două segmente, unul verde, steril, autotrof, şi
altul galben-bruniu, fertil, purtând numeroşi sporangi Sporangii sunt grupaţi în spice, sau un sinangiu foarte
alugit, masiv, cu perete gros pluristratificat, de tip eusporangiat, producând numeroşi izospori.
Subclasa MARATTIIDAE
Sunt ferigi tropicale, eusporangiate, cu tulpini drepte, înalte de până la 6 m, sau cu un rizom repent
purtând frunze mari, bipenate sau palmate, uneori întregi. Cele fosile erau arborescente, cu tulpina un stip
scurt. Sporangii sunt grupaţi în sinangii caracteristice sau în sori, pe faţa inferioară a pinulelor.
Subclasa OSMUNDIDAE
Sunt ferigi fosile şi actuale, ele făcând trecerea între ferigile eusporangiate şi leptosporangiate:
sporangele începe să se dezvolte dintr-o celulă epidermică ca la leptosporangiate şi ajunge ca unele celule
vecine să participe la formarea pedicelului pluricelular. La aceasta se mai adaugă o serie de caractere
considerate primitive: sporangii mari, negrupaţi în sori şi neprotejaţi, lipsiţi de inel mecanic; sporii sunt
numeroşi, fără perispor etc.
Subclasa POLYPODIIDAE
Majoritatea polipodiidelor sunt ferigi erbacee, numai cele tropicale sunt arborescente, atingând câţiva
metri înălţime. Ele aparţin ferigilor leptosporangiate, caracterizate prin: sporangi cu pereţi subţiri,
unistratificaţi, grupaţi în sori, în multe cazuri acoperiţi de induzie, în care se formează un număr mic de spori
(de obicei 32 – 64), adesea prevăzuţi şi cu perispor; anteridiile mai mici, neafundate adânc în protal şi produc
mai puţini anterozoizi; protalul este anual, de scurtă durată; organele vegetative cresc în lungime prin mitoza
unei singure celule apicale.
Sporofitul. Tulpinile polipodialelor sunt erbacee, perene, iar a celor tropicale, arborescente. Ferigile
erbacee au în sol un rizom brun, acoperit cu solzi şi cu resturi din tecile frunzelor moarte, purtând rădăcini
adventive. Cele arborescente au tulpina un stip, este erectă, neramificată, purtând în vârf un buchet de frunze.
Cilindrul central este variabil, de la protostel la eustel.

Frunzele sunt de tip macrofil, mari, lung peţiolate, de regulă sectate, rareori întregi. La majoritatea
polipodialelor, frunzele sunt trofosporofile şi numai la cele mai evoluate (Blechnum, Matteucia etc.) frunzele
sunt diferenţiate în trofofile şi sporofile.
Sporangii sunt pedicelaţi, cu cutia (capsula) prevăzută cu un inel mecanic incomplet şi sunt grupaţi în
sori. După locul de apariţie, sporangii pot fi marginali, cu o dispoziţie marginală, pe marginea frunzelor, sau
superficiali, pe faţa inferioară a frunzelor, uneori acoperiţi cu o induzie. Toate polipodiatele sunt izosporee.
Gametofitul. Germinarea izosporului dă naştere unui izoprotal lamelar, cordiform şi epigeu, verde,
autotrof, fixat prin rizoizi. Gametangii se află pe partea inferioară a protalului. Anteridiile sunt mici, alcătuite
numai din 3 celule parietale şi una centrală producând anterozoizi spiralaţi şi poliflagelaţi, iar arhegoanele
sunt împlântate numai cu partea bazală în protal.
Subclasa HYDROPTERIDIDAE
Ferigi acvatice şi palustre, foarte diferite de celelalte prin aparatul vegetativ: axa principală este
rizomatoasă purtând pe laturi frunze, rădăcini sau alte organe. Caracteristica esenţială este heterosporia, caz
unic la ferigile leptosporangiate. Sporangii sunt fără inel mecanic sau cu un rudiment, fiind grupaţi în sori
unisexuaţi sau bisexuaţi (microsporangi şi macrosporangi sau separaţi), incluşi într-o induzie, rezultând un
organ dur numit sporocarp. Microsporangele produce 64 microspori, iar macrosporangele voluminos produce
un singur macrospor. O altă caracteristică o reprezintă endoprotalia (protalele mici sunt incluse în spori)
amintind de Selaginellales şi Isoëtales. Microsporii şi macrosporii eliberaţi din sporangi şi sporocarpi
germinează dând protale (micro- şi macroprotale), care plutesc pe apă, are loc fecundarea cu formarea
zigotului, apoi a embrionului, din care rezultă o nouă plantă.
Încrengătura PINOPHYTA (Gymnospermatophyta, Gimnosperme)
Prin caracterele lor, pinofitele fac trecerea dintre polipodiofite şi magnoliofite:
- Fecundarea la polipodiofite se realizează prin anterozoizi ciliaţi, adică este zoidiogamă, caracter care există
şi la unele pinofite, în timp ce alte pinofite sunt sifonogame, ca şi magnoliofitele. De asemenea, fecundarea
are loc printr-un singur gamet mascul la polipodiofite, unul sau doi la pinofite şi doi la magnoliofite.
- Arhegonul de la polipodiofite este vizibil la pinofitele ancestrale, iar granulele de polen îşi continuă tranziţia
între tipul de dezvoltare al microsporului polipodiofitelor şi cel al polenului magnoliofitelor.
- Elementele conducătoare lemnoase de tip traheide caracteristice polipodiofitelor, există şi la pinofite şi chiar
la magnoliofitele primitive, după cum şi traheele caracteristice magnoliofitelor există şi la unele pinofite sau
chiar şi polipodiofite.
Caractere generale
Sunt plante exclusiv lemnoase (arbori, arbuşti, subarbuşti sau liane) constituite dintr-un corm complet, cu
rădăcină, tulpină şi frunze. Ele au lemnul format din traheide areolate, iar liberul este lipsit de celule anexe şi
prin activitatea meristemelor (cambiu şi felogen) se formează ţesuturi secundare, ce determină creşterea în
grosime a tulpinii şi rădăcinii;
Sunt diplobionte, eusporangiate şi heterosporee. Sporangii, împlântaţi în mezofil, se formează pe sporofile
specializate: microsporangii pe stamine şi macrosporangii pe carpele. Deoarece macrosporangele este învelit
în integument, apare un organ nou, numit ovul;
Sunt antofite, deoarece staminele sau carpelele se formează pe lăstari fertili, care constituie flori, unisexuate,
ahlamide şi anemogame, plantele fiind monoice sau dioice. Numai unele pinofite fosile (Bennettitales) au
avut flori bisexuate;
Ovulul după fecundare, la pinofitele evoluate se transformă în sămânţă, din care cauză ele se numesc şi
spermatofite. Sămânţa nu este închisă în carpele (care nu formează ovar), ea rămâne golaşă, de aceea se
numesc şi gimnosperme;

Primele diviziuni ale zigotului sunt nucleare (şi nu celulare ca la angiosperme), formându-se un proembrion
nediferenţiat, cu cca 1 000 nuclei fără pereţi celulari.
Sporofitul
Embrionul rezultă din diferenţierea histologică a proembrionului, format în urma diviziunii mitotice a
zigotului. Adesea, dintr-un singur zigot rezultă mai mulţi embrioni, fenomen numit poliembrionie
monozigotică. Prin diviziunea celulelor superficiale ale embrionului se formează un suspensor, care împlântă
embrionul în endospermul primar, apoi embrionul se diferenţiază în trei organe primordiale: radicula, tigela
şi gemula. Radicula va emite o rădăcină endogenă şi monopodială, tigela devine colet, iar gemula generează
tulpina şi cotiledoanele (2 – 18). În această fază, în care embrionul devine plantulă, ovulul se transformă în
sămânţă.
Rădăcina se ramifică în plan vertical: la pinofitele fosile (Cordaites sp.) ramificarea este dicotomică,
iar la cele actuale ramificarea este de regulă monopodială, dar şi simpodială (la unele specii din ordinul
Pinales). La unele specii (Pinus sp.) rădăcinile formează micorize, iar la Taxodium distichum, rădăcinile emit
pneumatofori. Rădăcina are structură endarhă şi prezintă şi creştere secundară, dar şi exarhă la pinofitele
primitive, fosile.
Tulpina este întotdeauna lemnoasă, cu ramificare dicotomică la fosile (Pteridospermales), iar la cele
actuale ramificarea este monopodială (Pinatae) sau simpodială (Ginkgoatae). De asemenea, tulpina este un
stip (Cycadatae), lianoidă (Gnetum sp.), un butuc (Welwitschia sp.) sau un cotor cu ramuri caduce (Ephedra
distachya). La genul Pinus sp, pe ramurile mari (macroblaste) se formează ramuri scurte (microblaste), care
cad împreună cu frunzele, iar la Ephedra distachya tulpina este articulată, virgată. Tulpina prezintă ritidoma
bine dezvoltată şi la unele specii (Pinus sp.) se desprinde în plăci. Scoarţa este relativ redusă. Stelul este de tip
protostelic, actinostelic, polistelic şi eustelic, endarh, mezarh (Pteridospermales) şi mai rar exarh. Lemnul
este constituit exclusiv din traheide scalariforme şi spiralate (Cycadatae) sau areolate (Pinatae) şi mai rar
traheidele sunt însoţite şi de trahee inelate (Gnetatae). Liberul este lipsit de celule anexe. Xilemul secundar
constituie adesea un duramen pronunţat, măduva este dezvoltată numai la Cycadatae, iar razele medulare sunt
omogene. La Pinales structura tulpinii se caracterizează prin prezenţa canalelor rezinifere schizogene, iar la
Cycadales există canale secretoare de gome, canalele secretoare fiind prezente şi în celelalte organe
vegetative.
Frunzele sunt diverse şi pot fi împărţite în două tipuri: microfiline şi megafiline (macrofiline). Cele de
tip microfilin pot fi scvamiforme neasimilatoare (Ephedra distachya) sau asimilatoare (Cupressaceae),
aciculare dispuse singular (Abies sp., Picea sp.), câte două sau mai multe (Pinus sp.) sau fasciculat (Larix
sp.), sau pot fi lăţite, asemănătoare cu ale magnoliofitelor (Gnetum sp.) sau bilobate (Ginkgo biloba).
Frunzele de tip macrofilin sunt în general de talie mare în raport cu tulpina, penate, asemănătoare cu ale
ferigilor pteridoide (Pteridospermales, Cycadatae, Bennettitatae) sau în formă de panglică (Welwitschia sp.).
Deşi se consideră că pinofitele au frunze persistente (sempervirente), totuşi ele cad pe rând după 2 până la 12
ani, cu excepţia frunzelor de la Welwitschia sp. care persistă toată viaţa (aproximativ 2 000 de ani) şi de la
Larix sp., Taxodium distichum sau Ginkgo biloba, care cad anual. Nervatura este diversă: uninervă
(Pinaceae), dicotomică (Ginkgo biloba), penată (Cycas sp., Gnetum sp.) sau paralelă (Welwitschia sp.).
Structura frunzei este adesea xeromorfă, cu epidermă cutinizată şi cerificată şi cu stomatele haplocheilice (la
pinofitele fosile) şi sindetocheilice (la cele actuale), cu mezofilul bogat în cloroplaste şi cu canale secretoare.
Floarea are o conformaţie strobilară, de con şi este lipsită de periant (ahlamidă) sau cu periantul redus
(Gnetatae), sporofilele purtând numele de stamine şi carpele, care se dispun spirociclic pe un lăstar scurt,
terminal sau axilar. Majoritatea pinofitelor au flori unisexuate şi în acest caz ele sunt dioice (Cycadatae,
Ginkgoatae, Gnetatae, Taxus baccata) sau monoice (Pinatae), iar flori bisexuate au avut numai unele
Bennettitatae.
Staminele (microsporofilele) au forme variate: penate (Pteridospermales şi Benettitales), scut peltat
(Taxus sp.), solzi purtători de două antere (Pinaceae), solzi cu numeroase antere concrescute (Cycadatae), sau
cu un filament simplu (Ginkgo biloba), sau ramificat (Ephedra sp.). Florile unisexuate mascule se pot grupa

în spice (Ginkgo biloba, Cordaites sp., Gnetum sp.) sau conuri compuse (Pinaceae). Anterele
(microsporangii) deşi nu au inel mecanic ca la polipodiofite, totuşi crapă longitudinal sau transversal şi au de
regulă doi saci polenici, uneori trei (Ginkgo biloba) sau mai mulţi (Taxus baccata). În sacii polenici, prin
diviziunea reducţională a arhesporiului se formează grăunciorele de polen (microsporii haploizi).
Carpelele (macrosporofilele) au forme diverse: cupuliforme (Ginkgo biloba, Taxus baccata),
solziforme (Pinaceae) sau foliacee şi penată (Cycas sp.). La unele pinofite fosile (Caytonia sp.) carpela se
răsucea circinat şi învelea ovulele într-o cavitate asemănătoare cu ovarul. La Pinaceae solzul carpelar fertil
este însoţit de un solz steril care reprezintă o bractee (frunză sterilă) ligulară. Ovulul (macrosporangele),
organul nou şi caracteristic apărut la pinofite, este constituit dintr-un integument pluristratificat, mai rar două
integumente (Gnetatae), care închide nucela (ţesutul sporogen – arhesporiul femel). Integumentul, provenit
din una sau mai multe macrosporofile concrescute şi transformate într-un înveliş protector, lasă o deschidere
apicală numită micropil, prin care va pătrunde tubul polenic. La Gnetatae, integumentele formează o
prelungire numită tubilus. La Cycadatae şi Ginkgoatae, la care anterozoizii sunt flagelaţi, sub micropil
rămâne o cameră polenică prin care intră anterozoizii spre oosferă. După poziţie, ovulele sunt atât ortotrope
(Bennettitatae, Cycadatae, Ginkgoatae), cât şi anatrope (Pinatae); de asemenea, ovulele pot fi pedicelate
(Bennettitatae şi Ginkgoatae) sau sesile (Pinatae şi Gnetatae).
Sămânţa este uneori cărnoasă (Cycas sp., Ginkgo biloba, Ephedra distachya) sau înconjurată numai
cu o cupă cărnoasă numită aril (Taxus baccata), dar cel mai frecvent sămânţa este uscată, cu testa sclerificată
provenită din integumentul ovulului. La unele (Pinaceae) la testă se adaugă o aripă provenită din ţesuturile
solzului carpelar, dând seminţei aspectul de samară.
La Cycadatae şi Gynkgoatae ovulul după fecundare nu suferă modificări morfologice şi poate fi
fecundat chiar şi după căderea lui de pe ramură, de aceea aceste plante sunt considerate prefanerogame, care
nu au sămânţă propriu-zisă cu embrion bine diferenţiat.
Gametofitul
Începe cu formarea microsporilor (grăuncioarele de polen) în microsporangi (sacii polenici din anteră)
şi a macrosporilor în macrosporange (nucela), fiind redus şi endoprotalian. Atât microsporii cât şi macrosporii
se formează în tetradă în urma diviziunii reducţionale a celulelor arhesporiului.
Grăunciorii de polen (microsporii) se formează câte patru din arhesporiul din sacii polenici din anteră
prin mai multe diviziuni. Grăunciorul de polen are un înveliş (epispor) compus din două straturi, intina
(intern) şi exina (extern), care la Pinales sunt îndepărtate pe laturi formând doi saci aeriferi, care ajută la
plutirea polenului, întrucât polenizarea este anemogamă. La Cycadatae şi Ginkgoatae episporul prezintă un
şanţ care permite creşterea conţinutului. De asemenea, la toate pinofitele exina prezintă cel puţin un por prin
care evaginează intina formând tubul polenic. Învelişul închide o celulă vegetativă, două celule protaliene sau
mai multe (protalul) şi o celulă generativă (anteridia). Prin urmare, microprotalul este redus la celulele
vegetative şi protaliene, iar anteridia (gametangiul mascul) la o singură celulă generativă. Gameţii masculi, la
Cycadatae şi Ginkgoatae sunt poliflagelaţi, iar fecundarea este zoidiogamă, pe când la Pinatae şi Gnetatae
gameţii masculi sunt spermatii, iar fecundarea este sifonogamă.
Macrosporii se formează câte patru după reducerea cromatică a unei celule sporogene a nucelei, dintre
care trei se reduc (resorb) şi numai unul, după numeroase diviziuni mitotice devine endospermul primar, care
reprezintă macroprotalul. Arhegoanele (gametangiile femele) se formează din celule ale endospermului
primar, fiind alcătuite din oosferă şi 2 – 3 celule gât şi se numesc corpuscule.
Gameţii femeli (oosferele) sunt imobili, formaţi în corpuscule, iar după fecundare, zigotul format
ramâne închis în endospermul primar al ovulului. Fecundaţia este simplă (este fecundată numai oosfera) şi
este foarte îndelungată, adică de la polenizare, formarea tubului polenic şi a gameţilor masculi şi până la
pătrunderea acestora la oosferă durează adesea un an.
Ciclul de dezvoltare
Gametofaza, mică şi de scurtă durată, începe cu diviziunea reducţională a celulelor arhesporiului din
sporangi, rezultând microsporul (grăunciorul de polen), cu microprotalul şi anteridia închise în microspor, şi

macrosporul care generează macroprotalul şi archegoanele. Prin urmare pinofitele sunt heterosporee şi
endoprotaliene. Sporofaza, predominantă, începe cu zigotul ce va da naştere la proembrion şi embrion în
sămânţă, care prin germinare va genera plantula şi apoi planta floriferă cu macro- şi microsporofile purtătoare
de macro- şi microsporangi. Deoarece sporofaza este predominantă, rezultă că plantele sunt diplonte.
Înmulţirea vegetativă este rară, ea realizându-se prin drajoni (Sequoia sp., Cephalotaxus sp.) sau
marcotaj (Ephedra sp., Taxus sp.).
Clasa CYCADATAE (Cycadopsida)
Cuprinde 100 specii actuale şi 100 specii fosile. Sunt plante lemnoase, arbori, arbuşti sau liane dioice,
monopodiale, cu frunze penate, pteridoide. Florile femele au ovule ortotrope, cu cameră polenică, care nu se
transformă în seminţe. Florile mascule au grăunciorii de polen fără saci aeriferi, iar gameţii masculi sunt
poliflagelaţi.
Clasa BENNETTITATAE (Bennettatopsida)
Cuprinde pinofite exclusiv fosile, cunoscute de la începutul Mezozoicului. Ele erau plante lemnoase,
arbuşti, cu trunchiul ca un butuc sau columnar, cu creştere monopodială, uneori ramificat, frecvent de 50 cm
înălţime. Din punct de vedere anatomic, tulpina prezenta un stel de tip sifonostel sau sifono-eustel, cu un
parenchim medular bine dezvoltat, străbătut de canale secretoare. Lemnul secundar avea traheide scalariforme
şi areolate. Frunzele erau penat sectate, lungi de până la 3 m, aşezate în vârful tulpinii, sau mici şi întregi,
lanceolate, cu nervatura penată, dispuse altern pe tulpină. Florile erau bisexuate sau unisexuate, în acest caz
plantele fiind dioice.
Clasa GINKGOATAE
Ginkgoatele sunt cunoscute din Permianul inferior, atingând cea mai mare dezvoltare în Mezozoic,
astăzi supravieţuind o singură specie – Ginkgo biloba, din familia Ginkgoaceae, ordinul Ginkgoales,
răspândită în Estul Chinei şi frecvent cultivată în Europa ca arbore ornamental. Sunt arbori dioici, cu creştere
simpodială. Ovulele după fecundare nu se modifică în seminţe veritabile şi prezintă cameră polenică, indicând
o fecundare zoidiogamă.
Clasa PINATAE
Cuprinde arbori sau arbuşti cu ramificare monopodială, mai rar simpodială (Pinus sp., Taxodium sp.,
Taxus baccata), cu ramuri lungi (macroblaste), scurte (microblaste) (Pinus sp., Cedrus sp.) sau numai lungi
(Abies sp., Picea sp.). Frunzele sunt simple, aciculare sau scvamiforme, alterne, izolate sau grupate câte două
sau mai multe până la fasciculate, decusate sau verticilate. Florile sunt unisexuate, iar plantele sunt monoice,
rar dioice (Taxus baccata, Juniperus sp.). Florile mascule, sub formă de stamine solziforme, mai rar peltate
(Taxus baccata), sesile sau peţiolate, fiecare purtând 2 – 20 antere dehiscente, sunt dispuse pe un ax formând
conuri. Grăunciorul de polen adesea are saci aeriferi, iar gameţii sunt neflagelaţi – spermatii. Florile femele
sunt dispuse în conuri formate dintr-un ax pe care se inseră carpele solziforme. Fiecare solz este dedublat,
constituit dintr-un solz extern steril, foarte dezvoltat la formele arhaice, şi un solz intern fertil, care poartă pe
faţa inferioară ovulele. Uneori toţi solzii carpelari sunt fertili. Când solzul extern este considerat bractee, iar
cel intern carpelă, conul este o inflorescenţă. Ovulul este protejat de un integument, cu sau fără cameră
polenică, iar în endospermul primar se formează 2 – 20 arhegoane. Ovulul fecundat devine sămânţă, cu
tegumentul sclerificat, la care aderă fie o aripă samaroidă, fie un aril cărnos (Taxus baccata), sau solzii
carpelari devin cărnoşi (Juniperus sp., Thuja sp.).
Toate organele vegetative şi reproducătoare conţin canale rezinifere, cu excepţia speciei Taxus
baccata
Clasa GNETATAE (Chlamidospermopsida)
Gnetatele sunt plante lemnoase dioice arbustoide, rar liane, nerăşinoase, fără reprezentanţi fosili, cu
lemnul secundar alcătuit din traheide cu punctuaţii areolate, dar şi din trahee, lipsite de canale secretoare, iar
liberul este uneori însoţit de celule anexe. Frunzele sunt variate ca formă şi dimensiuni, scvamiforme, late, în
formă de panglică, cu dispoziţie opusă. Florile, unisexuate, au perigon redus. Ovulul are două integumente

care se prelungesc deasupra micropilului într-un organ filiform numit tubilus. Embrionul are două
cotiledoane. La unele gnetate are loc dubla fecundare şi există endosperm secundar.
Încrengătura MAGNOLIOPHYTA (Angiospermatophyta, Angiospermae)
Sunt plante cu flori (antofite) grupând specii lemnoase (arbori şi arbuşti) şi erbacee (perene şi anuale).
Ele sunt plante tipic terestre, formând păduri şi pajişti şi numai secundar sunt adaptate la mediul acvatic
limnicol şi mai rar marin;
Majoritatea magnoliofitelor sunt verzi, autotrofe şi numai puţine sunt heterotrofe, nutrindu-se saprofit sau
parazit şi semiparazit;
Stelul este de tip eustel şi atactostel cu ţesutul lemnos, cu puţine excepţii, alcătuit exclusiv din trahee, care
nu conţine canale rezinifere, iar ţesutul liberian este compus şi din celule anexe;
Ramificarea tulpinii este preponderent simpodială, mai rar monopodială, dar niciodată dicotomică;
Frunzele sunt macrofile, mai rar aciculare sau reduse;
Florile sunt mai evoluate decât la gimnosperme, caracterizându-se prin prezenţa periantului (caliciu şi
corolă), apariţia ovarului (care închide ovulele lipsite de cameră polenică) ce se transformă în fruct, iar
stamina este redusă la filament şi anteră;
Gameţii masculi nu sunt niciodată ciliaţi (spermatii) şi realizează dubla fecundaţie (a oosferei şi a nucleului
secundar);
După fecundare se formează endospermul secundar din zigotul accesoriu al sacului embrionar;
Embrionul are numai 1 - 2 cotiledoane.
Sporofitul
Rădăcina primară provine din radicula embrionului, ea persistând la majoritatea magnoliatelor
(dicotiledonatelor), dar este efemeră la liliate (monocotiledonate), unde este înlocuită de rădăcini adventive cu
origine, de regulă, endogenă, în periciclu. Ea poate fi rămuroasă, pivotantă şi fasciculată. La unele plante
rădăcinile lipsesc, plantele fiind arize. Majoritatea rădăcinilor sunt prevăzute cu peri absorbanţi, dar pot lipsi
la unele magnoliate, lipsa acestora fiind suplinită de miceliul ciupercilor micorizante (rădăcini micorizante),
dar şi la unele liliate, lipsa lor fiind suplinită prin rădăcinile fasciculate care formează un sistem radicular
intensiv. Multe rădăcini sunt apte să drajoneze producând lăstari aerieni sau stoloni subterani.
Tulpina este generată de meristemele apicale ale gemulei embrionului şi poate avea dimensiuni de la 1
mm (Wolffia sp.) până la aproximativ 300 de metri la liane (Calamus sp.). Ea este de consistenţă lemnoasă
sau erbacee şi creşte în grosime, fie prin activitatea meristemelor secundare (cambiu şi felogen), la
magnoliate, fie prin sporirea numărului şi volumului celulelor corticale, la liliate. De asemenea, ca poziţie în
spaţiul aerian, tulpina poate fi erectă (la majoritatea magnoliofitelor), volubilă sau repentă.
După modul cum plantele trec sezonul nefavorabil, durata de viaţă a lor este foarte variabilă, ele fiind
anuale, bianuale şi perene (vivace).
Frunza provine din primordiile foliare ale mugurilor şi este întotdeauna în poziţie laterală şi niciodată
terminală, dar poate să ia o poziţie pseudoterminală. Frunzele pot fi simple, cu marginea întreagă, dinţată sau
serată, lobată, sectată (palmat penată) şi compuse, penate (impari- sau paripenate) şi palmate. Unele specii se
caracterizează prin heterofilie. Nervatura poate fi paralelă (în general la liliate) sau reticulat penată (la
majoritatea magnoliatelor).
Frunzele sunt inserate pe tulpini cu o mare regularitate, acestea având o dispoziţie alternă, opusă,
decusată, verticilată. Unele frunze au anexe foliare: stipele (Pisum L.), ligule şi auricule (Poaceae). Unele
plante parazite (Cuscuta L.) sunt complet lipsite de frunze, acestea numindu-se afile.
Floarea este organul caracteristic de reproducere al magnoliofitelor, fiind alcătuită din peduncul sau
receptacul pe care se inseră învelişurile protectoare, periantul şi elementele reproducătoare, staminele şi
pistilele.

Periantul este compus fie din caliciu (totalitatea sepalelor) şi corolă (totalitatea petalelor), deci este
dublu şi se numeşte diplohlamideu (heterohlamideu), sau poate fi simplu, cu un singur înveliş, şi se numeşte
monohlamideu (haplohlamideu) sau perigon, care atunci când este de culoare verde se numeşte sepaloid, iar
când este de culoare petalelor se numeşte petaloid, în ambele cazuri elementele periantului numindu-se
tepale. Unele flori sunt lipsite de periant, când se numesc flori nude sau ahlamidee. Caliciul este în general
caduc, dar poate persista şi participă la formarea fructului, când se numeşte acrescent (Physalis alkekengi L.).
Uneori caliciul este însoţit de organe foliacee, stipele sau bractee, alcătuind caliculul (Malvaceae, Rosaceae).
Atunci când caliciul are sepalele libere el se numeşte dialisepal, iar când sepalele sunt unite se numeşte
gamosepal. De asemenea, caliciul poate fi actinomorf sau zigomorf.
Corola este constituită din totalitatea petalelor. Ca şi caliciul, corola poate fi dialipetală sau
gamopetală. Ea poate lipsi, fenomen numit apetalie adevărată sau pseudopetalie. Apetalia adevărată poate fi
apetalie originară, atunci floarea este nudă şi nu a avut niciodată corolă, sau apetalie prin avortare, atunci când
floarea a pierdut corola prin avortare. Pseudopetalia se referă la existenţa corolei sub o altă formă, de exemplu
sub formă de nectarine (Aconitum L., Nigella L. etc.). De asemenea, ca şi caliciul, corola poate fi actinomorfă
sau zigomorfă.
Androceul este alcătuit din totalitatea staminelor, o stamină (microsporofilă) în formă clasică fiind
alcătuită din filament şi anteră divizată în două de un conectiv, antera având 4 saci polenici (microsporangi).
Androceul are o alcătuire diversă: cu stamine libere (dialistemon) sau concrescute prin filamentele staminelor
(gamostemon) sau prin antere (sinanter - Asteraceae). Androceul gamostemon poate fi monadelf, format
dintr-un singur mănunchi (Malvaceae), diadelf, format din două mănunchiuri (Fabaceae), triadelf sau
poliadelf, format din mai multe mănunchiuri de stamine (Hypericaceae, Tiliaceae). Uneori androceul este
redus la o singură stamină, alteori unele stamine sunt sterile, numite staminodii sau transformate în nectarine.
Gineceul reprezintă totalitatea pistilelor (macrosporofile), pistil rezultat din unirea uneia sau mai
multor carpele prin marginile lor, alcătuind ovarul (care închide ovulele), stilul şi stigmatul. Stilul este, în
general, terminal, dar poate fi lateral (Rosaceae) sau bazal - ginobazic (Boraginaceae, Lamiaceae). Stilul
poate avea lungimi diferite la florile heterostile: flori brevistile cu staminele depăşind stilul şi flori longistile,
cu staminele mai scurte decât stilul (Primula L.). Stigmatul poate fi întreg sau lobat. După poziţie, gineceul
poate fi super, sau hipogin, semi-infer sau perigin şi infer sau epigin. De asemenea gineceul este monomer şi
polimer, care este apocarpic (apocarp) sau dialicarpic, la care carpelele sunt independente, libere, fiecare
formând câte un pistil sau gamocarpic (sincarp), când gineceul este pluricarpelar, dar carpelele se unesc prin
marginile lor formând un ovar plurilocular. Când carpelele sunt unite numai prin baza lor, gineceul se
numeşte pseudo-apocarpic.
Elementele florale pot fi dispuse spirociclic, hemiciclic sau ciclic.
Florile pot fi izolate, singulare sau grupate în inflorescenţe simple sau compuse. Inflorescenţele simple
sunt formate dintr-un ax principal pe care se află bractee în axila cărora se găsesc ramuri secundare terminate
cu o floare. Se disting două tipuri de inflorescenţe simple: indefinite, la care florile au o dezvoltare centripetă
(florile mature se află la bază şi cele tinere la vârf) - racem, spic, corimb, umbelă, capitul, calatidiu şi definite,
la care florile au o dezvoltare centrifugă (au deschiderea florilor invers ca la cele indefinite), axul principal
terminându-se cu o floare ce se deschide prima, urmate de florile laterale, aceste inflorescenţe numindu-se
cime.
Inflorescenţele simple pot să se grupeze formând inflorescenţe compuse: racem compus, spic compus,
umbelă compusă etc.
Fructele rezultă după fecundare şi sunt de două tipuri: uscate şi cărnoase. Fructele uscate sunt:
indehiscente - achena, diachena, tetrachena, poliachena, samara, disamara, nuca sau nucula, cariopsa şi
dehiscente - capsula, silicva, silicula, folicula, păstaia, pixida. Fructele cărnoase: drupa, baca. De asemenea,
fructele pot fi adevărate, care provin din peretele gineceului şi false, la care pe lângă gineceu mai participă şi
alte elemente florale (receptacul, sepale etc.), precum şi fructe simple şi multiple (poliachene, polinucule,
polidrupe etc.).

Sămânţa provine din ovulul fecundat şi este protejată de un tegument seminal, la care se pot ataşa
diverse formaţiuni suplimentare: peri, aripi, caruncul etc.
Embrionul este format din tigelă, gemulă, radiculă, 1 - 2 cotiledoane şi hipocotilul (partea de tigelă de
sub cotiledoane).
Gametofitul
Grăunciorul de polen (macrosporul sau androsporul), care ia naştere în urma diviziunii reducţionale a
ţesutului sporogen din anteră, este învelit în exină (ce constituie un criteriu de identificare a speciilor) şi
intină, închizând o celulă vegetativă (microprotalul) şi o alta mică, lenticulară, celula generativă (anteridiola -
gametange). Celula generativă va da naştere la 2 gameţi masculi neciliaţi, spermatii (microgameţi). Acesta
reprezintă gametul mascul, foarte mult redus, închis în microsporangi (saci polenici) de pe sporofit.
Ovulul majorităţii magnoliofitelor este protejat de două integumente, numai la gamopetale şi apiacee
ovulul este prevăzut cu un singur integument, şi numai rar el lipseşte (Santalaceae). Sub integument se află
nucela (macrosporange), în care dintr-o celulă de sub micropil, prin meioză iau naştere macrosporii, din care
unul va da naştere sacului embrionar (macroprotalul), cu 8 nuclee din care se vor forma 7 celule: 3 celule
spre micropil, din care una centrală, oosfera (macrogametul) şi 2 laterale sinergide (gametangele); 3 celule la
polul opus, antipode (macroprotalul); o celulă centrală, nucleul secundar al sacului embrionar, diploid
(macrogamet). Sacul embrionar cu conţinutul său reprezintă gametofitul femel, redus şi închis în
macrosporange (nucela) de pe sporofit.
Fecundarea este dublă: o spermatie va fecunda oosfera, dând zigotul şi apoi embrionul, iar o alta va
fecunda nucleul secundar al sacului embrionar, dând zigotul accesoriu, triploid, din care prin mitoze se va
forma endospermul secundar, bogat în substanţe albuminoide, mult mai nutritiv.
Clasa MAGNOLIATAE (Dicotyledonatae)
Grupează aproximativ 171 000 (200 000) specii de plante lemnoase şi erbacee, reprezentând
componentele principale ale ecosistemelor din zonele tropicală, subtropicală şi temperată.
- Rădăcina principală endogenă este pivotantă sau ramificată, persistentă, exceptând unii reprezentanţi
(Ranunculaceae, Berberidaceae, Nymphaeaceae etc.) la care dispare de timpuriu, fiind înlocuită de rădăcini
adventive.
- Tulpina este erbacee sau lemnoasă, eustelică, cu fascicule libero-lemnoase de tip colateral, dispuse de regulă
pe un singur cerc concentric. Creşterea în grosime este determinată de către cele două zone generatoare
(cambiul, în cilindrul central şi felogenul, în scoarţă). Ramificarea este simpodială.
- Frunza este foarte diversificată ca formă: simplă cu marginea întreagă, dinţată, serată, penat sau palmat-
lobată sau sectată, sau penat- şi palmat-compusă, de regulă peţiolată şi uneori stipelată, cu nervaţiune penată
sau palmat-reticulată.
- Florile sunt în majoritate diplohlamidee, frecvent pe tipul 5, sau 4, mai rar pe tipul 2, spirociclice sau
hemiciclice la tipurile primitive şi ciclice la tipurile evoluate. Prezintă un aparat nectarifer floral, frecvent în
formă de disc.
- Embrionul, în general cu 2 cotiledoane, mai mult sau mai puţin egale, dar sunt şi cazuri de heterocotilie sau
de pseudocotilie, cu un singur cotiledon rezultat în urma reducerii unuia sau prin sincotilie. Mai rar
cotiledoanele lipsesc la unele specii parazite (Cuscuta L.).
Subclasa MAGNOLIIDAE (Polycarpice)
Sunt cele mai primitive magnoliofite, apărute în Jurasic. Caractere de primitivitate: vasele lemnoase de
tip primitiv, reprezentate prin trahee şi traheide, uneori lemnul secundar este homoxil, format numai de
traheide cu punctuaţii areolate; numărul mare şi inconstant de elemente florale (polimeria) dispuse de regulă
spirociclic (spirociclia) sau hemiciclic, mai rar ciclic, pe un receptacul conic, alungit, convex etc.; periantul
este simplu sau dublu, dialisepal şi dialipetal; floarea este de regulă actinomorfă, mai rar zigomorfă;
polenizarea este de regulă entomogamă, mai rar anemogamă.
Subclasa HAMAMELIDAE (Amentiferae)

Magnoliate în majoritate lemnoase, la unii reprezentanţi cu lemnul alcătuit şi din traheide cu punctuaţii
areolate, cu flori ciclice, haplohlamidee, rar diplohlamidee, sau cu periant rudimentar. Majoritatea au flori
unisexuate, plantele fiind monoice sau dioice, cu florile mascule grupate în amenţi, iar cele femele adesea
formând inflorescenţe variate, uneori în formă de con. Androceul este variabil, de la polimer până la
monomer. Gineceul apocarp sau sincarp, la care participă puţine carpele sau una singură, super sau infer, iar
ovlul are unul sau două integumente.
Subclasa ROSIDAE
Magnoliatele cele mai tipice, variate ca structură, dimensiuni, ecologie, corologie etc. Majoritatea au
flori bisexuate, diplohlamidee, dialisepale şi dialipetale, cu androceu de regulă polimer, până la oligomer sau
chiar monomer. Gineceul, de asemenea, polimer, apocarp sau sincarp, până la monomer, cu poziţie variabilă
în floare, superioară, semiinferioară şi inferioară. Dispoziţia elementelor florale, adesea ciclică.
Subclasa DILLENIIDAE
Dileniidele grupează plante lemnoase şi erbacee, răspândite mai ales în regiunile tropicale şi
subtropicale. Frunzele sunt predominant simple, iar florile pentamere, ciclice, diplohlamidee şi dialipetale, dar
şi gamopetale. Androceul poate fi diplostemon, rar haplostemon sau polimer prin multiplicare. Gineceul
sincarp, cu ovule numeroase, dispuse, în general, parietal.
Subclasa CARYOPHYLLIDAE (CENTROSPERMAE) Grupează plante în majoritate erbacee, rar lemnoase, caracterizate prin flori actinomorfe ciclice (cele
primitive hemiciclice), bisexuate sau unisexuate, în general pentamere. Androceul diplostemon, izomer sau
polimer, iar gineceul cu poziţie variabilă este sincarp, compus din numeroase până la o singură carpelă şi
placentaţie centrală.
Subclasa ASTERIDAE
Asteridele constituie una dintre cele mai evoluate linii filogenetice ale clasei Magnoliatae, incluzând
ordine de plante lemnoase şi erbacee răspândite pe tot globul. Printre caracteristicile principale ale plantelor din
această subclasă figurează: simpetalia şi tetraciclia florilor, androceul în majoritatea cazurilor concrescut cu tubul
corolei, lipsa poliandriei, iar gineceul este redus de obicei la 2 carpele.
Clasa LILIATAE (Monocotyledonatae)
Liliatele constituie un grup vast, cu cca. 2800 genuri şi 66000 specii de plante erbacee, mai puţin
lemnoase, răspândite pe tot globul. Cele lemnoase participă la constituirea pădurilor tropicale umede, iar cele
erbacee contribuie la edificarea savanelor şi mai ales a stepelor.
- Rădăcina principală rezultată din radicula embrionului piere de timpuriu, fiind suplinită de rădăcini
adventive fasciculate;
- Tulpina, de obicei, este neramificată sau ramificată mai ales monopodial, adesea este subterană sub formă de
rizomi, bulbi, tuberculi;
- Stelul este adesea de tip atactostel, cu fascicule libero-lemnoase închise, iar creşterea în grosime se
realizează rar pe seama meristemelor secundare;
- Frunze frecvent simple şi nestipelate, cu nervatura paralelă sau arcuată şi cu teaca bine dezvoltată;
- Florile sunt haplohlamidee, trimere, mai rar tetramere sau dimere, niciodată pentamere;
- La formarea tetradei de polen, citocineza are loc la prima diviziune, cu excepţia speciilor din familia
Orchidaceae;
- Embrion cu un singur cotiledon şi numai excepţional cu două sau un rudiment din al doilea.
Subclasa ALISMIDAE (Alismatidae) Plante acvatice, multe submerse, cu frunze alterne, heteromorfe. Florile actinomorfe hemiciclice şi
ciclice, foarte variate, unele simple, reduse şi nude, altele cu periant simplu sau dublu, cu androceu şi gineceu
polimer. Gineceul, întotdeauna apocarp, poate deveni pseudocarp prin sudarea bazei carpelelor. Polenizarea
hidrogamă, entomogamă şi anemogamă. Fructul capsulă, foliculă sau poliachenă.
Subclasa LILIIDAE

Liliidele se caracterizează prin plante cu flori actinomorfe şi zigomorfe, trimere, de regulă,
pentaciclice. Adesea au învelişul floral un perigon petaloid şi mai rar diferenţiat în caliciu şi corolă, sau lipsă.
Androceul din 6 stamine dispuse pe două cicluri, dar poate să fie redus până la o stamină, iar gineceul este
sincarp.
Subclasa ARECIDAE (SPADICIFLORAE)
Cuprinde ordine de plante erbacee şi lemnoase, caracterizate printr-o inflorescenţă numită spadice, cu
flori mici, înconjurate de o frunză modificată numită spat.
VII. REPRODUCEREA SEXUATĂ LA SPERMATOFITE: TENDINȚE EVOLUTIVE
Meioza şi fecundaţia constituie cei doi poli ai ciclului vital al organismelor; la plante, sporul şi respectiv,
zigotul sunt fiecare la originea generaţiilor distincte care alternează: haplofaza (n) şi diplofaza (2n),
realizându-se un ciclu haplo-diplofazic. La plantele vasculare modalităţile de reproducere sexuată nu pot fi
înţelese decât în strânsă corelaţie cu progresele continue legate de adaptarea la viaţa în mediul terestru.
La plantele vasculare se produce o regresie a haplofazei (sau a gametofitului), care se află progresiv
subordonată diplofazei (sau sporofitului). La cele mai primitive – pteridofitele, care sunt deja adaptate vieţii
terestre prin aparatul lor vegetativ, fecundaţia rămâne încă tributară apei, datorită producerii de anterozoizi
mobili. De aceea, ca şi algele, ciupercile şi briofitele la cere nu există organe reproducătoare distincte,
pteridofitele sunt incluse în grupa criptogamelor.
Să ne reamintim! La pteridofite, reproducerea sexuată este o heterogamie oogamă, dar cele două faze
care alternează sunt independente una de alta (deoarece protalul sau gametofitul este verde, asimilator, cu
rizoizi). Gametofitul (n) aminteşte de talul algelor şi se numeşte protal, redus la o lamă verde, purtătore de
organe sexuale (gametangi): anteridii (mascule) şi arhegoane (femele). Sporofitul (2n) este însăşi planta, care
domină gametofitul. După fecundaţie, zigotul va forma un proembrion, apoi un embrion, punctul de plecare
pentru formarea unei plante noi.
La ferigile izosporee (Dryopteris filix-mas, Polypodium), există fronde (frunze) ce produc un singur tip de
sporangi şi de spori; protalul (gametofitul) este hermafrodit, deoarece poartă în acelaşi timp şi anteridii şi
arhegoane.
La ferigile heterosporee (Selaginella), există un dimorfism sexual evident, deoarece microsporii (masculi)
şi macrosporii (femeli) sunt produşi în sporangi distincţi şi vor da microprotale cu anteridii şi macroprotale cu
arhegoane.
În ceea ce priveşte specializarea, la Dryopteris şi Polypodium frondele asimilatoare sunt şi reproducătoare
în acelaşi timp, în timp ce la Selaginella există două tipuri de fronde: unele numite trofofile, asimilatoare şi
altele numite sporofile (microsporofile şi macrosporofile). La speciile de Equisetum, sporangii apar grupaţi în
strobile, la extremitatea tulpinilor verzi sau pe axe neclorofiliene separate; sporii sunt asemănători din punct
de vedere morfologic (izospori), dar diferiţi din punct de vedere fiziologic (heterospori).
La fanerogame sau spermafite, microsporofilele se numesc stamine, iar macrosporofilele
(megasporofilele) se numesc carpele. Ele sunt încorporate într-un ansamblu ce include adesea şi pise sterile,
numit floare.
Staminele produc microsporii (granulele de polen tinere, uninucleate) a căror conţinut corespunde unui
gametofit mascul foarte redus.
Macrosporii (megasporii) nu mai sunt eliminaţi la exterior; ei rămân într-un macrosporangiofor numit
ovul, solidar cu carpela. Protalul femel sau endospermul se va dezvolta în interiorul ovulului, într-un ţesut
parenchimatic numit nucelă; în endosperm se vor forma gameţii femeli (oosferele).
Fecundaţia la gimnospermele primitive (Cycas, Ginkgo) se realizează încă prin participarea gameţilor
masculi mobili. La gimnospermele evoluate (Pinus, Abies), granulele de polen mature vor produce la
germinare un tub polinic, care va purta spre oosferă gameţii masculi, imobili, fenomen numit sifonogamie.

La gimnosperme, ovulul “dezvelit” pe carpelă, va veni în contact direct cu polenul. La angiosperme,
intervine o protecţie suplimentară: carpelele vor închide complet ovulul, apărând astfel ovarul. Tubul polinic
va trebui să traverseze parte din această carpelă până va întâlni oosfera (J. -C. Roland, F. Roland, 1987).
Aşadar, avem de-a face, în cursul evoluţiei, cu un paralelism în dezvoltarea individului superior diferenţiat
(sporofitul) şi reducerea progresivă a gametofitului. Un organism diploid (2n) este mai rezistent decât unul
haploid (n), căci dubla sa garnitură cromozomică rezistă mai bine erorilor şi accidentelor biochimice şi
genetice care pot surveni în celulele lor.
La gimnospermele primitive, unisexuat dioice, florile prezintă o organizare relativ simplă. Astfel, la
Cycas florile mascule (cu aspect de con) sunt formate din axe pe care se află dispuşi în spirală zeci ori sute
de solzi staminali lăţiţi, ce poartă la faţa inferioară numeroşi saci polinici grupaţi câte 3 sau 6; în sacii polinici
se vor forma prin meioză microsporii (granulele de polen tinere, uninucleate) din care vor rezulta mai apoi
granulele de polen mature, care în momentul eliberării din sacii polinici conţin 3 celule: bazală, vegetativă (cu
rol în formarea tubului polinic ce se va fixa în nucela ovulului, având rol de “haustor”) şi generativă (care va
forma doi gameţi masculi mobili).
Florile femele (cu aspect de con când sunt închise şi cu aspect de rozetă cand se deschid) sunt formate
din carpele de culoare gălbuie, extrem de pubescente, cu aspect foliar, ce prezintă o parte superioară
franjurată şi o parte inferioară purtătoare de ovule dispuse pe două şiruri. Ovulele pot atinge dimensiuni
impresionante (după unii autori, ajung de mărimea unui ou de găină) fiind protejate la exterior de un
integument (ce lasă în partea superioară un mic orificiu numit micropil) care prezintă trei pături distincte:
sarcotesta, cărnoasă, sclerotesta, dură şi endotesta, subţire, parenchimatică. La interior se află nucela (care
spre micropil se dezorganizează, formând o cameră polinică) în care, în urma meiozei, se vor forma
macrospori, unul din ei fiind la originea endospermului primar; în acesta din urmă se vor forma două
arhegoane, fiecare conţinând câte un gamet femel numit oosferă.
La Ginkgo biloba (arborele pagodelor) florile mascule sunt formate din numeroase “stamine”,
alcătuite dintr-un filament foarte redus şi prezintă terminal câte doi saci polinici; aici, granulele de polen
tinere, uninucleate ce iau naştere în urma meiozei, devin granule de polen mature cu 4 celule: bazală şi picior
(celule protaliene), vegetativă (având acelaşi rol ca şi la Cycas) şi generativă (care va forma, de asemenea, doi
gameţi masculi mobili).
Florile femele sunt reprezentate de pedunculi ramificaţi dichotomic, în vârful cărora se formează
iniţial două ovule, din care unul va avorta, iar celălalt va ajunge la maturitate. Ovulul este protejat la exterior
de un integument cu epidermă asimilatoare, care prezintă, ca şi la Cycas , sarcotestă, sclerotestă şi endotestă.
În nucela de la interior se va diferenţia endospermul primar clorofilian, cu substanţe de rezervă (aleuronă şi
lipide), în care se vor forma, spre micropil două arhegoane cu câte o oosferă.
În urma polenizării (realizată cu ajutorul vântului), atât la Cycas, cât şi la Ginkgo, granulele de polen
mature ajung pe o “picătură micropilară” emisă de nucelă şi prin resorbţia acesteia, în final, în camera
polinică, unde va începe germinarea granulelor de polen: celula vegetativă va forma un tub polinic simplu
(Cycas) sau ramificat (Ginkgo), ce se va fixa în nucelă, iar celula generativă va forma o celulă soclu şi o
celulă spermatogenă; din cea din urmă rezultând doi gameţi masculi mobili, care vor înota într-un “lichid de
fecundaţie”pentru a ajunge la oosfere.
Unii autori (H. Camefort, H. Boué, 1980) consideră că zoidogamia (picătura de lichid necesară
deplasării gameţilor masculi) şi acumularea substanţelor de rezervă în endosperm înainte de fecundaţie,
reflectă primitivitatea acestor grupe de plante în ceea ce priveşte reproducerea în general, incluzându-le din
acest punct de vedere într-o grupă specială, a “prefanerogamelor”. Mai mult chiar, datorită substanţelor de
rezervă de care dispun, imediat după polenizare, ovulele se pot desprinde de “planta mamă”, rupând orice
relaţie fiziologică cu aceasta, chiar dacă fecundaţia propriu-zisă nu a avut încă loc.
La gimnospermele cu conuri, unisexuat monoice (de exemplu, Pinus sylvestris), florile sunt lipsite
de învelişuri florale şi grupate în conuri mascule (ce apar la baza lăstarilor de un an) şi conuri femele (ce apar
la extremitatea lăstarilor anuali).

Conurile mascule, cu valoare de flori, prezintă o axă pe care sunt dispuse în spirală stamine cu pedicel
foarte scurt şi o anteră cu doi saci polinici, în care, prin meioză, se formează microspori sau granule de polen
cu doi saci aeriferi latero-bazali; fiecare granulă de polen matură care este eliberată în momentul dehiscenţei
sacilor polinici va avea 4 celule: bazală, picior (sau celule protaliene), vegetativă (cu rol în formarea tubului
polinic) şi generativă (cu rol în formare celor doi gameţi masculi imobili, numiţi spermatii).
Conurile femele, cu valoare de inflorescenţe, prezintă o axă pe care sunt dispuse în spirală carpele cu
câte două ovule la faţa superioară (o floare este formată dintr-o bractee, un solz carpelar şi două ovule); în
fiecare ovul, prin meioză, iau naştere macrospori, din diviziunile repetate ale unuia din ei rezultând
endospermul primar, în care se formează două arhegoane, fiecare cu câte un gamet femel, numit oosferă.
În urma polenizării (relizate de vânt) şi a fecundaţiei celor două tipuri de gameţi (conduşi de către
tubul polinic – sifonogamie simplă) rezultă zigotul (2n), din diviziunile căruia se formează embrionul; acesta,
împreună cu endospermul primar şi integumentul ovulului, formează sămânţa, organ neînchis în fruct (acesta
neformându-se la gimnosperme, deoarece carpelele nu se închid).
Microsporogeneza (formarea microsporilor sau a granulelor de polen tinere)
În complexul de celule sporogene primare din microsporange au loc mai întâi numeroase mitoze,
rezultând astfel celulele mamă ale granulelor de polen.
Fiecare din aceste celule (2n) suferă meioza, rezultând câte 4 microspori (n). Acesta este procesul de
microsporogeneză.
În paralel, celulele mamă ale granulelor de polen se disociază, se rotunjesc, se îmbibă cu apă şi cresc;
în interiorul lor microsporii îşi vor forma exina şi sacii aeriferi (la Pinus).
În ceea ce priveşte diviziunile celulelor mamă ale granulelor de polen distingem:
- tipul succesiv (succedan), când se formează pereţi după fiecare diviziune (2 pereţi perpendiculari);
tetrasporii au dispoziţie cruciată (Cycas);
- tipul simultan, întâlnit la majoritatea gimnospermelor, când microsporii au dispoziţie tetraedrică,
pereţii acestora edificându-se numai după ce au rezultat cei 4 nuclei (deci după cele două
diviziuni) (Ginkgo, Pinus, Larix).
După formarea tetradei de microspori, peretele celulei mamă se umflă, se gelifică şi apoi se resoarbe,
încât microsporii devin liberi în cavitatea microsporangelui, plutind într-un lichid bogat în substanţe nutritive,
rezultate din citoliza celulelor mamă ale microsporilor. Microsporii cresc în volum, luând forma şi structura
caracteristică pentru fiecare specie. Granulele de polen (iniţial uninucleate) sunt adesea supuse o perioadă
relativ îndelungată la condiţii extreme în timpul transportului şi polenizării. Protecţia lor este esenţială şi
asigurată de sporoderma formată din exină (care are 3 straturi: ectoexina, mezoexina şi endoexina) şi intină
(înconjoară protoplastul, este mult mai subţire, permeabilă, are multă pectină la periferie, ceea ce-i permite
detaşarea de exină; la germinarea polenului, intina va da tubul polinic).
Tipurile de polen sunt variabile, dar caracteristic este faptul că la conifere, în general, exina se
îndepărtează de o parte şi de alta de intină, rezultând astfel doi saci aeriferi, cu rol în răspândirea polenului.
Formarea gametofitului mascul (trecerea de la granula de polen tânără la granula de polen matură)
Una din trăsăturile caracteristice ale granulei de polen de la gimnosperme este structura sa bipolară: la
un pol se află celulele protaliene (celula bazală şi celula picior), la celălalt pol se află celula vegetativă ce va
forma tubul polinic.
La Pinus, dezvoltarea gametofitului mascul comportă 5 diviziuni şi are loc în două etape:
I. durează de la microspor până la formarea granulei de polen care se găseşte încă în
microsporange (sac polinic);
II. cuprinde dezvoltarea ulterioară a granulei de polen, după părăsirea microsporangelui (chiar
după polenizare, în ovul).

La Pinus, microsporul este uninucleat, cu doi saci aeriferi. După o primă diviziune rezultă o celulă
mică, bazală, care va degenera şi o celulă mare, apicală. A doua diviziune priveşte celula apicală din care vor
rezulta o celulă picior (lângă celula bazală) şi o celulă obişnuită. A treia diviziune priveşte celula obişnuită din
care se vor forma o celulă generativă (gametogenă) şi o celulă vegetativă.
Aşadar, granula de polen matură care pleacă din microsporange are 4 celule, dar primele două formate
(celula bazală şi celula picior) sunt împinse spre perete şi vor degenera rapid.
Creşterea tubului polinic începe din momentul în care granula de polen matură ajunge pe nucela
ovulului. În timp ce tubul polinic se alungeşte în nucelă (în jur de două luni după începutul germinării), are
loc a patra diviziune care vizează celula generativă ce va da o celulă soclu (sterilă) şi o celulă spermatogenă
(fertilă).
Abia în anul următor, continuă germinarea granulei de polen, când are loc cea de-a cincea diviziune,
iar celula spermatogenă va da doi gameţi masculi, imobili (spermatii).
Angiosperme. Formarea şi structura gametofitului femel (sacului embrionar)
Tipuri de gametofiţi:
- monosporic: unul dintre cei 4 macrospori participă la formarea scaului embrionar; este tipul normal
(Polygonum), întâlnit la 80 % dinte angiosperme;
- bisporic: cei doi nuclei din prima diviziune a meiozei iau parte la formarea sacului embrionar;
- tetrasporic: toţi cei 4 nuclei ai macrosporului cenocitic contribuie la formarea sacului embrionar.
Gametofit monosporic (tipul Polygonum). Sacul embrionar se formează din macrosporul chalazal
(bazal) al tetradei de macrospori. El creşte (pe seama afluxului de substanţe din placentă, asimilării ţesutului
nucelar şi chiar tegumentar), se alungeşte o dată cu dezvoltarea ovulului, provocând comprimarea celorlalţi
trei macrospori, a celulelor nucelare vecine şi chiar a calotei nucelare. Citoplasma se vacuolizează. Nucleul
suferă trei diviziuni mitotice, citocineza este simultană (după prima diviziune, cei 2 nuclei migrează la cei doi
poli ai sacului embrionr, vacuolele confluează într-una mare, centrală, care îi separă). După ce au rezultat cele
două grupe de câte 4 nuclei, se conturează cele 7 celule:
- aparatul oosferei (la polul micropilar) cuprinde oosfera (gametul femel, cu nucleu bazal) şi două
sinergide mici, cu nuclei apicali;
- aparatul antipodial (la polul chalazal) cuprinde 3 antipode;
- celula centrală (secundară) a sacului embrionar, cu 2 nuclei haploizi nefuzionaţi sau cu un nucleu 2n.
FECUNDAŢIA
Reprezintă fenomenul prin care doi gameţi de sex diferit se unesc şi rezultă zigotul (oul).
Când cei doi gameţi provin de la acelaşi individ procesul se numeşte autofecundaţie (autogamie), iar
când provin de la doi indivizi diferiţi, fecundaţie încrucişată sau alogamie. Autogamia este legată de
autofecundaţie (în special la plantele cu flori cleistogame), iar fecundaţia încrucişată, de alopolenizare.
FECUNDAŢIA SIMPLĂ
Se întâlneşte la grupele de plante mai puţin evoluate (briofite, pteridofite, gimnosperme). Are loc în
mediul acvatic la plantele mai puţin evoluate şi în mediul aerian la gimnosperme.
La gimnosperme, din celula spermatogenă a granulei de polen se diferenţiază doi gameţi masculi
neflagelaţi (imobili), care sunt transportaţi de tubul polinic. Acesta, după ce a traversat nucela, duce gameţii
masculi până la gâtul arhegonului. Ajuns în arhegon, vârful tubului polinic se gelifică, gameţii sunt eliberaţi şi
unul din ei (de regulă cel mai mare dacă dimensiunile sunt inegale) se contopeşte cu nucleul oosferei, ducând
la formarea zigotului. Avem de - a face, aşadar, cu o sifonogamie simplă. Cel de al doilea gamet mascul,
nucleul vegetativ şi celula soclu degenerează rapid, neluând parte la fecundaţie. Există însă şi excepţii, când
cel de al doilea gamet poate fecunda oosfera din al doilea arhegon sau (ca la Abies balsamea) se poate uni cu
nucleul celulei ventrale, fiind astfel la originea unui ţesut nucelar foarte puţin dezvoltat, inclus în sporofitul
tânăr. Acest proces ar preconiza, după unii autori, dubla fecundaţie de la angiosperme.
FECUNDAŢIA DUBLĂ

A fost descoperită de Navaşin (1898) şi apoi de Guinard (1899) la două specii de liliacee (Fritillaria
tenella şi Lilium martagon). Se cunoaşte în prezent faptul că dubla fecundaţie (în cursul căreia cei doi gameţi
masculi care sunt transportaţi prin tubul polinic fuzionează cu două celule din sacul embrionar – oosfera şi
celula centrală) caracterizează toate angiospermele. Oosfera fecundată formează zigotul principal, care se va
dezvolta în embrion. Celula centrală diploidă devine triploidă în urma fecundaţiei şi formează zigotul
accesoriu, din care se va dezvolta endospermul secundar (albumenul) cu rol în nutriţia embrionului.
Mecanismele dublei fecundaţii
După ce tubul polinic pătrunde în ovul, se îndreaptă spre sacul embrionar, unde va elibera gameţii
masculi. De regulă, intrarea în sacul embrionar se realizează printr-o sinergidă degenerescentă. De obicei,
înainte de ajungerea tubului polinic la sacul embrionar, o singură sinrergidă degenerează, aceasta
reprezentând locul de pătrundere a tubului polinic.
Sinergidele joacă un rol activ în intrarea tubului polinic în sacul embrionar; aceasta se datorează
faptului că una din sinergide degenerează înainte ca tubul polinic să ajungă în sacul embrionar. Această
degenerare are loc doar dacă floarea a fost polenizată. Rezultă, deci, că sinergida nu este o structură pasivă, ce
este distrusă de tubul polinic la pătrunderea în ea. Se presupune că sinergida degenerată, prin conţinutul ei
vacuolar, ar avea rol de ghidare a tubului polinic, de stabilire a direcţiei de creştere al acestuia.
După ce tubul polinic intră în nucelă, este ghidat spre sacul embrionar şi, chiar dacă ajunge la
sinergida intactă, îşi continuă drumul spre sinergida degenerată.
O dată pătruns în sinergidă, diametrul tubului polinic se reduce la jumătate şi creşterea încetează;
vârful tubului polinic se lizează datorită unor variaţii bruşte ale presiunii osmotice şi îşi eliberează conţinutul
(fig. 65 B). Nucleul vegetativ al granulei de polen, nucleul şi membrana sinergidei degenerează: cei doi
gameţi masculi lipsiţi de perete se îndreaptă spre cele două celule ce urmează a fi fecundate (oosfera şi
nucleul secundar), care sunt, la rândul lor, parţial lipsite de perete. Migrarea celor doi gameţi masculi este
strict controlată. Ajungerea tubului polinic la sacul embrionar provoacă o reorganizare importantă a
citoscheletului din sinergida receptoare, din oosferă şi celula centrală. La nivelul situsurilor posibile de
fuziune apar coroane de filamente actinice, despre care se crede că ar juca un rol în migrarea gameţilor
masculi spre locul de fuziune.
Fuziunea gameţilor este încă puţin cunoscută.Plasmalemele gameţilor masculi şi femeli fuzionează,
nucleul gamtului mascul pătrunde în citoplasma gametului femel, unde fuzionează cu nucleul acestuia. Celula
centrală a sacului embrionar poate avea doi nuclei şi, în acest caz, gametul mascul fuzionează cu unul dintre
aceştia, urmând ca apoi nucleul diploid astfel rezultat să fuzioneze cu celălalt nucleu al celulei centrale. În alte
cazuri (Arabidopsis), cei doi nuclei ai celulei centrale fuzionează înainte de fecundaţie, astfel încât gametul
mascul fuzionează direct cu un nucleu diploid.
La unele specii (Gossypium, Lilium), în apropierea gameţilor femeli fecundaţi s-au observat corpi
citoplasmatici enucleaţi, ceea ce indică faptul că citoplasma gameţilor masculi nu participă la fecundaţie. La
cele mai multe angiosperme, ereditatea extranucleară (reprezentată de moleculele de ADN plastidial şi
mitocondrial) este în totalitate de origine maternă. Totuşi, se cunosc unele specii (Plumbago zeylanica,
Nicotiana tabacum) la care ereditatea extranucleară are origine bipaternală.
În cursul cariogamiei, nucleul spermatic se alătură nucleului oosferei şi se aplatizează, apoi
membranele celor doi nuclei se resorb, iar nucleul mascul migrează în cel femel. În urma acestei fuziuni
nucleul rezultat se îmbogăţeşte în ADN, iar nucleolul mascul – care se menţine în timpul cariogamiei sau care
se reconstituie în momentul contopirii celor doi nuclei – se uneşte cu nucleolul femel. Aceasta are loc fie
imediat, fie în momentul începerii mitozelor care marchează începutul embriogenezei. Fecundaţia celulei
centrale are loc după acelaşi mod ca şi în cazul oosferei; deosebirile provin din faptul că există variaţii ale
momentului contopirii celor doi nuclei polari (înainte sau după fecundaţie), aşa cum am menţionat anterior.
EMBRIOGENEZA
După fecundaţie, în cursul ontogenezei, zigotul suferă diviziuni repetate, modificându-se ca structură
şi funcţii.

Citodiferenţierea se realizează prin activarea sau inactivarea materialului genetic, astfel încât în
celulele specializate funcţionează numai una sau câteva gene, restul fiind inactivate. Citodiferenţierea
reprezintă, în esenţă, o transcripţie diferenţiată a informaţiei genetice, care are drept consecinţă o sinteză
proteică diferenţiată.
Fenomenul prin care un grup de celule controlează diferenţierea altui grup de celule reprezintă
inducţia embrionară. În embrionul pe cale de dezvoltare se realizează astfel un control reciproc între celule,
grupe de celule, ţesuturi şi organe, ceea ce determină citodiferenţierea.
Aşadar, citodiferenţierea este, în esenţă, un fenomen de reglaj genetic la nivel celular, prin care are loc
stimularea activităţii unor gene şi represia altora.
Se ştie că transcripţia şi translaţia informaţiei genetice poate fi stimulată în diferite ţesuturi cu ajutorul
fitohormonilor, fără a se cunoaşte însă prea bine mecanismul lor de acţiune.
Prezenţa embrionului este caracteristică cormofitelor. Totuşi, unii autori consideră embrion şi primele
stadii de diviziune ale zigotului la briofite, până la diferenţierea sporogonului.
EMBRIOGENEZA LA ANGIOSPERME După fecundaţie, zigotul primar rămâne în stare latentă sau de maturare un timp mai îndelungat decât
zigotul accesoriu. Aceasta se explică prin activitatea fiziologică mai intensă a nucleului secundar, apoi a
zigotului accesoriu cu garnitură triplă de cromozomi. Totodată, această succesiune este normală, deoarece din
activitatea zigotului accesoriu va lua naştere albumenul, care va servi drept sursă de hrană pentru embrionul
principal, până când acesta va fi capabil de nutriţie autotrofă.
Perioada de repaus a zigotului principal este variabilă. De exemplu, la arborele de cacao (Theobroma
cacao) zigotul accesoriu se divide la 4-5 zile după fecundaţie, iar cel principal la 14-15 zile după aceasta. La
brânduşă (Colchicum autumnale), însă, fecundaţia are loc toamna, zigotul accesoriu se divide imediat după
aceasta, iar zigotul principal rămâne în stare de latenţă 4-5 luni, până în primăvara următoare.
Zigotul, care are o zestre ereditară dublă (maternă şi paternă), va genera un nou organism, mai adaptat
mediului de viaţă decât dacă acesta ar fi rezultat dintr-o oosferă nefecundată sau dintr-o celulă somatică
oarecare (prin înmulţire vegetativă).
Forma şi mărimea zigotului sunt variabile. În general, zigotul angiospermelor este piriform sau
cilindric, scurt sau alungit. Zigotul este în mod obişnuit drept, cu simetrie axial-longitudinală, rareori curbat.
În perioada imediat următoare fecundaţiei, în zigotul primar au loc modificări la nivel ultrastructural:
vacuolele mari prezente la partea superioară a celulei dispar treptat, locul lor fiind luat de citoplasmă.
Vacuolele vor reapărea în momentul premergător primei diviziuni a zigotului. Nucleul este situat de regulă
excentric, iar în jurul său se găsesc mitocondrii, plastide şi picături lipidice.
Zigotul prezintă o polaritate externă (morfologică), dublată de una internă (fiziologică) care, de regulă,
coincid. Polaritatea morfologică se manifestă prin aceea că la partea superioară se găseşte nucleul, iar la cea
inferioară vacuola. Polaritatea fiziologică rezidă din intensitatea crescută a metabolismului celular la polul
superior al celulei. De asemenea, partea superioară a embrionului prezintă un geotropism negativ, iar cea
inferioară unul pozitiv.
În primele stadii de dezvoltare a embrionului nu există diferenţe fundamentale între dicotiledonate şi
monocotiledonate. Însă, pe măsură ce embrionul se dezvoltă, diferenţele dintre între cele două grupe apar tot
mai pregnant.
De regulă, prima diviziune a zigotului este transversală. În urma acestei diviziuni rezultă două celule:
o celulă proximală (situată în dreptul micropilului) şi una distală. Între aceste două celule apare imediat un
perete despărţitor, spre deosebire de cazul de la gimnosperme, unde primele diviziuni ale zigotului nu erau
urmate de citocineză.
A doua diviziune poate varia ca orientare în cele două celule: de regulă celula proximală se divide tot
transversal, în timp ce celula distală se divide fie transversal, fie longitudinal sau chiar oblic. Ulterior,
diviziunile celulare – atât anticline cât şi pericline – au loc cu o frecvenţă mai mare în regiunea distală,
situată spre chalază,; în această parte se va forma embrionul propriu-zis (corpul embrionar). Atâta timp cât

simetria este radiară, vorbim de proembrion, iar după apariţia cotiledoanelor şi trecerea la simetria bilaterală,
de embrion. În regiunea proximală, dinspre micropil, diviziunile sunt mai reduse numeric; în această parte se
va forma suspensorul, alcătuit, de regulă, din unul sau mai multe şiruri de celule suprapuse.
Diferenţierea ulterioară a embrionului urmează un tipar (model) specific pentru fiecare grup de plante
în parte. Apariţia polilor radicular şi caulinar reprezintă primul pas în stabilirea polarităţii, care se va accentua
pe măsura derulării etapelor următoare ale embriogenezei.
Tipuri de dezvoltare şi structura embrionilor la dicotiledonate
În urma diviziunilor succesive ale zigotului se formează, aşa cum am menţionat şi mai sus, un
proembrion globulos cu simetrie radiară. În acest stadiu, diferenţele dintre dicotiledonate şi monocotiledonate
sunt nesemnificative. Numărul de primordii cotiledonare (unul la monocotiledonate şi două la dicotiledonate)
este considerat a fi prima deosebire dintre cele două grupe de angiosperme.
Ulterior embrionul dicotiledonatelor capătă o formă bilobată datorită dezvoltării celor două
cotiledoane, în timp ce embrionul monocotiledonatelor are o formă mai mult sau mai puţin cilindrică, indusă
de expansiunea unicului cotiledon.
Pe măsura dezvoltării proembrionului, la baza sa, lângă suspensor, apare hipofiza (ce grupează
celulele iniţiale ale scufiei, rizodermei şi scoarţei rădăcinii), iar la vârful lui se diferenţiază epifiza (din care
va rezulta epiderma şi scoarţa tulpinii).
La dicotiledonate, primordiile cotiledonare apar ca două proeminenţe meristematice situate la partea
apicală a embrionului şi rezultate în urma diviziunilor pericline ale straturilor superficiale din această regiune.
Schimbarea formei duce, implicit, la schimbarea simetriei embrionului: din radiară, în bilaterală.
Formarea şi creşterea cotiledoanelor este asemănătoare cu cele ale unei frunze normale. În acest caz
putem vorbi de o creştere apicală şi de una marginală a cotiledoanelor.
Regiunea apicală a embrionului (la dicotiledonate), delimitată de cele două cotiledoane, reprezintă
apexul caulinar al epicotilului. La monocotiledonate, apexul este evident într-o depresiune situată la baza
cotiledonului unic şi apare complet înconjurat de acesta la embrionul matur.
Sub cotiledoane se diferenţiază hipocotilul şi radicula, iar la germinarea seminţei apare vizibilă
tulpiniţa primordială (gemula sau epicotilul).
Tipurile de dezvoltare şi structură a embrionilor la angiosperme variază prin modul de apariţie a
pereţilor despărţitori în proembrion şi chiar în embrion, prin structura suspensorului, poziţia embrionului în
sămânţă etc. De aceea structura definitivă a unui embrion constituie, deseori, un caracter în diagnoza
diferitelor grupe de plante.
Tipul Crucifer
Fiind primul descris, se numeşte şi tipul clasic. Părţile principale ale embrionului derivă din celula
apicală. Suspensorul şi hipofiza derivă din celula bazală.
Dezvoltarea embrionului începe cu diviziunea transversală a zigotului în două celule:
- una bazală (inferioară), mai mare, spre micropil;
- una apicală (superioară), mai mică, orientată spre centrul nucelei.
a) Prima celulă care se divide este cea bazală ; prin numeroase segmentări transversale rezultă un
suspensor lung, filamentos, format dintr-un şir de celule, dintre care cea bazală, mare, veziculoasă, are
rol de haustor, iar cea terminală, dinspre proembrion, funcţionează ca hipofiză. Această celulă se va
divide transversal, rezultând două celule fiice, fiecare dintre ele suferind apoi două diviziuni prin pereţi
perpendiculari unul faţă de altul. Rezultă astfel un ansamblu de opt celule, dintre care patru inferioare
reprezintă iniţialele scoarţei rădăcinii, iar cele patru superioare generează scufia şi rizoderma rădăcinii.
b) Celula apicală se divide mai târziu longitudinal şi apoi transversal, rezultând proembrionul 4-celular
(stadiul de quadrant. Apoi, fiecare din celulele cvadrantului se divide transversal, rezultând
proembrionul 8-celular (stadiul de octant): cele patru celule inferioare vor forma tulpiniţa (epicotilul)

şi cotiledoanele, iar cele patru celule superioare vor forma hipocotilul şi iniţialele cilindrului central al
rădăcinii.
În continuare, toate celulele octantului se divid periclin, formându-se: un strat de celule externe ce vor
da tunica şi apoi epiderma şi un masiv de celule interne ce vor da corpusul, iar apoi scoarţa şi cilindrul
central. Diviziunile celulelor embrionului continuă şi, mai târziu, se schiţează cotiledoanele (pe flancurile unei
depresiuni embrionare apicale). La sfârşitul embrigenezei tulpiniţa se schiţează, apoi se conturează clar.
În acest stadiu, embrionul apare mai mult sau mai puţin cordat în secţiune longitudinală. Curând,
cotiledoanele şi hipocotilul se alungesc (prin diviziuni transversale). Ulterior embrionul creşte mult mai rapid
decât suspensorul, se îndoaie, luând forma ovulului (nucelei) (campilotrop la crucifere) .
În final embrionul matur ocupă cea mai mare parte din sămânţă, iar albumenul se resoarbe aproape
complet.
VIII. ZOOLOGIA NEVERTEBRATELOR
Tema I. Organizarea corpului la protozoare, clasificare, exemple, mod de viaţă
Protozoarele (gr. protos – primul, zoon – animal, Goldfuss, 1817) sunt eucariote unicelulare,
majoritatea de dimensiuni microscopice, cu nutriţie heterotrofă. Conform cu clasificarea lumii vii în mai
multe regnuri, ele aparţin Regnului Protista. Celula uni protozoar are valoare de individ biologic deoarece ea
singură îndeplineşte toate funcţiile fundamentale ale vieţii: funcţiile de relaţie, nutriţie şi de reproducere.
Mediul de viaţă al protozoarelor este variat. Majoritatea sunt organisme libere, acvatice (marine sau de
apă dulce) sau în solul umed; altele sunt simbionte, comensale sau parazite la suprafaţa sau în interiorul
corpului altor organisme; unele sunt chiar prădătoare.
Organizarea corpului. Celula unui protozoar este alcătuită din 3 părţi componente principale:
membrana, citoplasma şi nucleul.
Membrana celulară. Este formată din macromoleculele stratului superficial al citoplasmei, dispuse
aproape perpendicular pe suprafaţa celulei. O proprietate esenţială a membranei celulare este elasticitatea,
care conferă celulei mişcări metabolice.
Membrana celulară poate prezenta o serie de învelişuri de protecţie. Acestea pot fi de natură celulozică (la
unele fitoflagelate), chitinoasă ori silicioasă (testacee), sau proteică (gregarine).
Citoplasma. Citoplasma sau plasma fundamentală a celulei este compusă dintr-un amestec de proteine,
lipide şi glucide. La unele protozoare, citoplasma se diferenţiază într-un strat superficial – ectoplasma, hialin,
şi un strat intern, dens – endoplasma.
În masa citoplasmei se găsesc organite citoplasmatice. Acestea pot fi generale (prezente în toate celulele) şi
specifice (numai la anumite protozoare).
Organite citoplasmatice generale: mitocondriile (respiraţia aerobă); dictiozomii (= aparatul Golgi) –
sistem de canalicule cu rol secretor şi excretor; reticulul endoplasmatic – sistem de canalicule cu rol
circulator. Acesta este de două feluri: neted, cu rol în metabolismul glicogenului, şi rugos (prezentând
ribozomi pe suprafaţă), cu rol în sinteza proteinelor; ribozomii – sediul sintezei proteice; centrozomul – cu rol
în diviziunea celulară. Este un corpuscul sferic situat în apropierea nucleului, ce formează fusul de diviziune.
Organite citoplasmatice specifice: vacuolele digestive – vezicule intracitoplasmatice cu rol în digestie;
vacuolele pulsatile (sau contractile) – vezicule cu rol principal osmoregulator şi secundar excretor şi
respirator; organite fotosensibile: stigma, la flagelate; organite cu rol secretor: corpusculii parabazali (=
aparatul parabazal) – corpusculi derivaţi din aparatul Golgi, situaţi lângă nucleu; organite cu rol contractil:
1. mioneme – fibrile citoplasmatice lungi, care prin contracţie scurtează corpul şi pedunculul protozoarelor
fixate (la ciliate); 2. miofrisce - fibrile citoplasmatice scurte fixate la baza baghetelor scheletice de la acantari.
Prin contracţie, ele tracţionează stratul extern al citoplasmei, mărind volumul corpului şi facilitând plutirea
sau ridicarea la suprafaţă; kinetodesme – fibrile ectoplasmatice ce unesc corpusculii bazali, cu rol în mişcarea

coordonată a cililor (ciliate); organite cu rol în apărare: trihocistele (la ciliate) – vezicule ectoplasmatice
alungite care, prin „explodare” proiectează un filament prevăzut cu un spicul ascuţit; pot fi toxice sau
netoxice; capsula polară cu filament extensibil de fixare (Microsporidia); organite cu rol de schelet intern: 1.
axostilul (la unele flagelate) – baghetă de natură proteică dispusă în lungul celulei, cu rol în susţinerea
corpului celulei. 2. baghete de acantină (la acantari); 3. baghete de silice (la radiolari), 4. axoneme (=
filamente axiale proteice) – la heliozoare, folosesc ca loc de prindere a pseudopodelor (numite aici axopode);
5. centroplastul – o granulă centrală derivată din centrozom pe care se sprijină axonemele (la unele
heliozoare).
Pe lângă organitele celulare, în citoplasmă se mai pot găsi constituenţi paraplasmatici – produşi de
metabolism sau înglobaţi din afară: substanţe de rezervă: lipide, glucide, proteine (înglobate în vacuole);
învelişuri de protecţie: membrana chistului, substanţe minerale (carbonat de calciu, silice, etc.).
Nucleul. Este o formaţiune constantă (nu lipseşte niciodată), individualizată în citoplasmă morfologic
şi prin compoziţia chimică (conţine acizi nucleici şi proteine). Protozoarele pot avea unul (la euglenoidine,
amibe), doi (la Giardia) sau mai mulţi nuclei, fie de acelaşi fel (la opaline, unele heliozoare, acantari), fie
diferenţiaţi în micronuclei (cu rol generativ) şi macronuclei (cu rol vegetativ) – la ciliate.
Forma corpului la protozoare poate fi constantă, la cele cu membrana citoplasmatică întărită cu
învelişuri rezistente, sau variabilă, la cele cu membrana extrem de subţire; acestea din urmă îşi pot modifica
forma corpului prin emiterea de prelungiri protoplasmatice numite pseudopode (la Sarcodina) sau prin
acţiunea fibrilelor contractile (mioneme şi miofrisce).
Scheletul. Multe protozoare prezintă formaţiuni, în general dure, situate la interior sau la exterior, ce
constituie scheletul. Scheletul intern este reprezentat de organitele citoplasmatice cu rol de schelet, prezentate
mai sus. Scheletul extern este compus din constituenţi paraplasmatici de diferite naturi: învelişuri gelatinoase
sau pseudochitinoase, care se pot întări prin alipirea unor corpuri străine (granule de nisip), la Testacea;
cochilii calcaroase – la foraminifere.
Mişcarea. Protozoarele se deplasează activ cu ajutorul organitelor de mişcare. Acestea pot fi
temporare – pseudopodele, sau permanente – flagelii şi cilii.
Pseudopodele sunt prelungiri citoplasmatice produse de Sarcodine. Au forme diferite: lobate
(lobopode), filiforme (filopode), reticulate (reticulipode).
Flagelii şi cilii sunt prelungiri citoplasmatice permanente ce conţin la interior fibrile proteice flexibile.
La baza flagelilor şi cililor există o diferenţiere numită corpuscul bazal (= blefaroplast = kinetozom), care
constituie centrul cinetic al flagelului sau cilului respectiv. Corpusculii bazali sunt situaţi sub membrana
celulară. Distrugerea corpusculului bazal duce la imposibilitatea de mişcare a flagelului sau cilului respectiv.
Numărul flagelilor este variabil, la diferitele grupe de protozoare; în majoritatea cazurilor, flagelii sunt mai
lungi decât corpul.
Cilii sunt asemănători ca structură internă flagelilor, dar sunt mai scurţi şi întotdeauna în număr mare.
Ultrastructura flagelilor şi cililor este, în esenţă, asemănătoare la toate organismele eucariote. Un flagel (sau
cil) este alcătuit din: 2 microtubuli centrali (= fibrile axiale), înconjuraţi de 9 perechi de microtubuli externi (=
fibrile secundare). Un microtubul din fiecare dublet poartă două braţe îndreptate spre dubletul adiacent. Între
aceste două categorii de fibrile se poate intercala un cerc de 9 fibrile intermediare (la Polymastigina şi
spermatozoidul mamiferelor). Întregul mănunchi de microtubuli este închis într-o teacă ce se continuă cu
membrana celulară. Pe teaca învelitoare, anumiţi flageli (cei foarte activi) prezintă perişori laterali rigizi
extrem de fini, numiţi mastigoneme (nu se văd la microscopul optic). Flagelul (cilul) are la bază un corpuscul
bazal (blefaroplast = kinetozom) situat sub membrana celulară, în ectoplasmă. Acesta are o ultrastructură
aasemănătoare flagelului, cu excepţia că cele 9 grupe de fibrile externe sunt formate din triplete de
microtubuli, doi din cei trei microtubuli fiind continuaţi cu dubletele flagelului. În partea bazală a
corpusculului bazal, microtubulul intern din fiecare triplet este conectat printr-un fascicul radiar la un inel
central.

Protozoarele libere pot realiza deplasări orientate numite taxii, iar protozoarele fixate se pot orienta
într-o parte sau alta prin mişcări numite tropisme. Taxiile sau tropismele pot fi pozitive sau negative dacă
produc apropierea, respectiv îndepărtarea, sub acţiunea unui anumit factor de mediu (foto-, termo-, chimio-,
galvano- etc.).
Sensibilitatea. Se realizează cu ajutorul unor cili tactili, stigme fotoreceptoare sau prin existenţa unor
concreţiuni cu rol în percepţia gravitaţiei (la ciliate).
Nutriţia. Protozoarele sunt protiste cu nutriţie heterotrofă. Aceasta poate fi 1. fagotofă (= holozoică),
când particulele alimentare înglobate în citoplasmă sunt cuprinse într-o vacuolă digestivă, delimitată de o
membrană vacuolară derivată din plasmalemă, 2. osmotrofă, când hrana trece, la nivel macromolecular, prin
membrana citoplasmatică prin difuziune. Se întâlneşte la endoparazite (sporozoare) şi la unele flagelate
saprozoice.
O parte din nutrimente sunt oxidate, cu obţinerea de energie necesară proceselor metabolice, o altă
parte se depune sub formă de substanţe de rezervă (glicogen, paramilon, grăsimi). Acestea sunt oxidate
treptat, în funcţie de necesităţi. Părţile nedigerate se elimină prin apropierea vacuolei digestive de suprafaţa
corpului. La ciliate, eliminarea reziduurilor se face printr-un anumit loc numit citoproct.
Excreţia. În urma proceselor metabolice, în celulă apar substanţe toxice (acid uric, uree, amoniac),
care sunt eliminate prin difuziune pe toată suprafaţa celulei şi prin vacuolele pulsatile. La ciliate, o parte din
sărurile acumulate sunt eliminate prin trihociste.
Respiraţia. Poate fi aerobă sau anaerobă. Respiraţia aerobă se întâlneşte la formele libere, la care
schimbul de gaze se face prin difuziune, pe toată suprafaţa corpului. La protozoarele simbionte (heliozoare,
radiolari) cu algele fotosintetizante (zooclorele, zooxantele), oxigenul este produs în celulă prin fotosinteza
algelor. La formele parazite din sânge, oxigenul este luat din sângele gazdei. Respiraţia anaerobă se întâlneşte
la paraziţii intestinali. Energia este produsă prin descompunerea paraglicogenului în acizi graşi şi CO2.
Reproducerea. Poate fi asexuată sau sexuată. Reproducerea asexuată. Se întâlneşte la majoritatea
protozoarelor şi este singurul mod de reproducere la unele specii. Constă în diviziunea celulei (numită şi
fisiune), care poate fi de două feluri: binară sau multiplă. 1. diviziunea binară: celula creşte până îşi dublează
volumul, apoi se divide, mai întâi nucleul, apoi citoplasma. Rezultă 2 celule fiice de dimensiuni egale.
Diviziunea binară poate fi longitudinală (la flagelate) sau transversală (la ciliate). Atunci când una din cele
două celule fiice este mult mai mică decât cealaltă, procesul se numeşte înmugurire (la suctorii). 2. diviziunea
multiplă: în cazul celulelor vegetative este numită şi schizogonie şi constă în diviziunea repetată a nucleului
însoţită de plasmotomie simultană. Rezultă mai multe celule (= schizonţi sau merozoizi). Diviziunea multiplă
apare şi în procesul de sporogonie, când se divide de mai multe ori nucleul zigotului, rezultat dintr-un proces
de copulaţie a gameţilor; rezulte mai multe celule haploide numite spori. Diviziunea multiplă se întâlneşte la
foraminifere, radiolari şi sporozoare.
Reproducerea sexuată. Este numită şi gamogonie şi este reprezentată de: copulaţie, conjugare şi
autogamie. Copulaţia constă în fuziunea (singamia) a două celule numite gameţi. După forma, mărimea şi
sexul gameţilor, există trei tipuri de copulaţie: izogamie, heterogamie şi anizogamie. Izogamia. Constă în
copularea a doi gameţi identici ca formă, mărime şi potenţial sexual numiţi izogameţi. Heterogamia. Constă
în copularea a doi gameţi care diferă ca mărime, numiţi heterogameţi. Anizogamia. Constă în copularea a doi
gameţi care diferă ca mărime, formă şi sex: un microgamet flagelat, mascul, şi un macrogamet, neflagelat,
femel.
În general, meioza apare în procesul de formare a gemeţilor, care sunt haploizi (meioză prezigotică),
caz în care indivizii adulți sunt diploizi. Dar, la multe flagelate şi sporozoare, meioza este post-zigotică (ca la
majoritatea algelor), caz în care individul vegetativ este haploid. Conjugarea. Este un proces sexual
incomplet, cunoscut la ciliate, în care nu se formează gameţi. Doi indivizi aderă unul la celălalt şi fac schimb
de nuclei haploizi printr-o punte citoplasmatică temporară. Fiecare nucleu migrator (+) fuzionează cu un
nucleu staţionar (-) din conjugantul opus, formând un nucleu diploid numit sincarion. Apoi, cei doi
conjuganţi se separă şi continuă să se dividă binar.

Mai puţin comun este procesul numit autogamie, în care doi nuclei cu valoare de gamet fuzionează
formând un nucleu diploid în interiorul aceluiaşi individ. La multe protozoare, reproducerea se face în cadrul
unor cicluri reproductive, în care alternează reproducerea asexuată şi cea sexuată (foraminifere, sporozoare).
Închistarea. Este o caracteristică a ciclului de viaţă la numeroase protozoare ce includ majoritatea
speciilor de apă dulce. Are loc în condiţii nefavorabile (uscăciune, variaţii de temperatură, poluarea apei). În
timpul închistării, celula îşi micşorează volumul, ia o formă sferică şi secretă un înveliş îngroşat, devenind
inactivă. În aceste condiţii, protozoarele pot rezista perioade lungi de timp şi pot fi deplasate la distanţe mari.
Atunci când condiţiile favorabile revin, chistul se desface şi protozoarul îşi reia viaţa normală. Cel mai simplu
ciclu de viaţă cuprinde numai 2 faze: o fază activă şi o fază închistată. Ciclurile de viaţă mai complexe se
caracterizează prin zigoţi închistaţi sau prin formarea de chisturi reproductive în care au loc diviziunea,
gametogeneza sau alte procese reproductive.
Clasificarea protozoarelor. Clasificările moderne ale protozoarelor, considerate ca parte din regnul
protistelor sunt foarte variate, în funcție de autori. Prezentăm aici o clasificare tradițională, didactică, ce
reflectă mai clar etapele evoluției în organizarea corpului la protozoare. Se cunosc circa 64000 de specii de
protozoare, clasificate în 5 clase:
1. Clasa Flagellata (= Mastigophora) – libere sau parazite, posedă, cel puţin o parte din viaţa lor,
flageli. Exemple: 1. Ordinul Euglenoidina: Euglena gracilis; 2. Ordinul Protomonadina: Bodo saltans,
Codosiga botrytis, Trypanosoma gambiense; 3. Ordinul Polymastigina: Trichomonas vaginalis, Giardia (=
Lamblia) intestinalis; 4. Ordinul Hypermastigina: Trichonympha chattoni, Lophomonas blattarum; 5. Ordinul
Opalinida: Opalina ranarum; 6. Ordinul Rhizomastigina: Mastigamoeba aspera.
2. Clasa Sarcodina – libere sau parazite, prezintă pseudopode. Exemple: 2.1. Subclasa Rhizopoda: Ordinul
Amoebina: Amoeba proteus, Entamoeba histolytica; Ordinul Testacea: Difflugia oblonga, Euglypha
alveolata; Ordinul Foraminifera: Cornuspira sp., Lagena sp., Biloculina sp., Triloculina sp., Nodosaria sp.,
Globigerina sp., Rotalia sp., Elphidium sp., Nummulithes sp. 2.2. Subclasa Actinopoda: 2.2.1. Ordinul
Heliozoa: Actinophrys sol, Actinosphaerium eichorni, Acanthocystis aculeata; 2.2.2. Ordinul Acantharia:
Acanthometra elastica; 2.2.3. Ordinul Radiolaria: Thalassicolla sp., Collozoum sp., Eucyrtidium sp.,
Coelacantha ornatha.
3. Clasa Sporozoa – toate endoparazite, lipsite de organite locomotoare, dar prezintă gameţi flagelaţi.
Exemple: 3.1. Ordinul Gregarinida: Gregarina sp., Monocystis sp., Stylorhynchus sp., 3.2. Ordinul Coccidia:
Eimeria schubergi; 3.3. Ordinul Haemosporidia: Plasmodium falciparum.
4. Clasa Microsporidia – specii endoparazite, la care sporii, foarte mici, prezintă o singură capsulă polară
cu filament lung. Exemple: Nosema bombycis, Nosema apis.
5. Clasa Ciliophora – prezintă cili vibratili şi nuclei diferenţiaţi în micro- şi macronuclei. Exemple: 5.1.
Subclasa Ciliata. 5.1.1. Ordinul Holotricha: Paramaecium sp., Coleps sp., Ichthyophthyris sp.; 5.1.2. Ordinul
Spirotricha: Stentor sp., Balantidium sp.; 5.1.3. Ordinul Peritricha.: Vorticella sp. 5.2. Subclasa Suctoria.
Sphaerophrya sp., Tokophrya sp., Ephelota sp.
Tema II. Organizarea corpului la metazoarele diploblastice, clasificare, exemple, relații filogenetice,
mod de viaţă
Animalele (metazoarele) sunt cuprinse în două categorii organizatorice fundamentale: diploblastice şi
triploblastice. Animalele diploblastice se caracterizează prin faptul că în timpul dezvoltării embrionare se
formează numai două foiţe embrionare (straturi de celule): ectodermul şi endodermul. Din acest grup fac parte
spongierii, celenteratele. La aceste animale simetria fie lipsește (majoritatea spongierilor), fie este radiară
(cnidarii) sau biradiară (acnidarii).
1. Încrengătura Porifera (=Spongiaria)
Cuprinde spongierii, cele mai simple metazoare - animale solitare sau coloniale, marea majoritate;
organisme marine, puţine de apă dulce. Trăiesc fixate de substrat, în stadiul adult. Nu prezintă ţesuturi
specializate, nici organe; nu au musculatură, nici sistem nervos sau organe specializate de simţ. Nu au cavitate

digestivă (digestia intracelulară). Spongierii prezintă un schelet intern (produs în mezoglee), de natură
minerală sau organică. Scheletul mineral poate fi calcaros sau silicios, reprezentat de spiculi (sclerite) de
forme diferite (monaxoni, triaxoni, tetraxoni). Scheletul organic este reprezentat de fibrele de spongină, o
substanţă proteică.
Mod de viaţă. Spongierii trăiesc fixaţi de substrat, majoritatea fiind marini; puțini de apă dulce
(Spongilla, Ephidatia). Nutriţia este microfagă. Deseori participă la simbioze.
Clasificare. În funcţie de natura scheletului, spongierii se clasifică în 3 clase: Calcarea (ex.
Leucosolenia, Sycon), Hexactinellida (ex. Hyalonema, Euplectella) şi Demospongi (ex. Suberites, Spongilla,
Euspongia, Dendrilla).
Subdiviziunea Coelenterata. Celenteratele cuprind metazoare diploblastice cu simetrie radiară şi
prevăzute cu o cavitate digestivă (= gastrală), care este chiar cavitatea corpului (gr. coelos – cavitate; enteron
- intestin). Cavitatea digestivă comunică cu exteriorul printr-un singur orificiu – orificiul buco-anal. Apare
digestia extracelulară (cavitară), alături încă de digestia intracelulară. La celenterate apar diferenţieri tisulare:
musculatură, sistem nervos, organe de simţ.
Clasificare. Celenteratele cuprind două încrengături: Cnidaria şi Acnidaria.
Structura peretelui corpului La hidră, dintre cnidari (Clasa Hydrozoa), o specie foarte cunoscută,
peretele corpului este alcătuit din două foiţe epiteliale: ectodermul şi endodermul, între care se află un strat
subţire de mezoglee.
Ectodermul conţine mai multe feluri de celule: celule epitelio-musculare (sau mio-epiteliale) în forma
de T culcat, cu baza celulei prevăzută cu fibrile musculare netede, dispuse longitudinal în raport cu axa
corpului; aceste fibrile formează pătura musculară longitudinală, externă; ectodermul mai cuprinde: celule
senzitive, celule nervoase (care formează o reţea nervoasă – primul sistem nervos din seria animală), celule
urzicătoare (cnidoblaste sau nematoblaste, cu rol în atac şi în apărare), celule interstiţiale, mici din care se
formează cnidoblastele.
Endodermul este format din celule digestive mari, prevăzute cu 1-2 flageli, ce se mişcă în interiorul
cavităţii gastrale. Ele sunt capabile să emită şi pseudopode, cu ajutorul cărora fagocitează particulele
alimentare. O mare parte din celulele endodermice au baza diferenţiată în fibrile musculare orientate circular
faţă de axa longitudinală a corpului, formând în ansamblu pătura musculară circulară, internă. Printre
celulele digestive se găsesc şi celule mici, glandulare, ce conţin enzime digestive, şi celule mucoase.
1. Încrengătura Cnidaria cuprinde celenterate cu simetrie radiară, majoritatea marine, prevăzute cu
celule urzicătoare de origine ectodermică, numite cnidocite, cnidoblaste sau nematoblaste (gr. knide – urzică;
blastos – mugure; nema - filament). Cnidarii sunt unici printre metazoare prin prezența unui ciclu de
dezvoltare dimorfic, care presupune două morfologii ale stadiului adult: polipul și meduza.
Polipul este, în general, forma fixată de substrat şi se reproduce, de regulă, asexuat. Polipul poate fi
solitar sau colonial. Atunci când forma de meduză lipseşte, polipul poate deveni sexuat (la hidră sau la corali).
Coloniile de polipi pot avea indivizii asemănători între ei sau diferenţiaţi morfo-funcţional, fenomen numit
polimorfism colonial. Meduza este forma liberă şi sexuată. În ontogenie se realizează o alternanţă de polipi şi
meduze numită metageneză. Larva caracteristică cnidariilor este planula.
Clasificare. Se cunosc circa 11000 de specii de cnidari, clasificaţi în trei clase: Hydrozoa, Scyphozoa
şi Anthozoa.
1.1. Clasa Hydrozoa. Cuprinde circa 3200 de specii de cnidari cu cavitatea gastrală simplă,
necompartimentată. Se prezintă sub formă de polip sau de meduză. Majoritatea speciilor sunt marine, puţine
de apă dulce. Exemple:1.1.1. Subclasa Hydroida: Hydra sp., Podocoryne (=Hydractinia) sp., Campanularia
sp., Lizzia sp., 1. 1. 2. Subclasa Siphonophora: Halistemma sp., Stephalia sp., Physalia sp., Velella sp.
1. 2. Clasa Scyphozoa. Cuprinde cnidari reprezentaţi, mai ales sub formă de meduză. Polipul apare
tranzitoriu în timpul ontogenezei, mai rar ca stadiu adult. Cavitatea gastrală este compartimentată prin 4 septe
endodermice numite sarcosepte, în 4 buzunare gastrale. Orificiul buco-anal are formă pătrată. Exemple:
Lucernaria sp., Charybdea sp., Periphylla sp., Aurelia sp., Rhizostoma sp.

1. 3. Clasa Anthozoa. Cuprinde cnidari reprezentaţi numai sub forma de polip (gr. anthos – floare;
zoon - animal), cunoscuţi şi sub numele de corali. Pot fi solitari sau coloniali. Cavitatea gastrală este împărţită
prin septe în numeroase loji, fapt ce conferă o mare suprafaţă digestivă. Prezintă un schelet calcaros, în
general bine dezvoltat. Coralii sunt exclusiv marini. Se cunosc circa 6225 de specii. Exemple: 3.1. Subclasa
Octocorallia: Tubipora sp., Alcyonium sp., Corallium sp. Gorgonia sp., Pennatula sp.. 3.2. Subclasa
Hexacorallia: Actinia sp., Anemonia sp., Madrepora sp., Fungia sp., Cerianthus sp.
2. Încrengătura Acnidaria. Cuprinde celenterate cu simetrie bilradiară, lipsite de cnidoblaste dar
purtătoare de coloblaste – formaţiuni lipicioase cu rol în captarea hranei. Nu prezintă alternanţă polip-meduză
(metageneză). Exemple: 2.1. Clasa Ctenophora: Hormiphora plumosa, Beroe ovata.
Tema III. Organizarea corpului la metazoarele triploblastice acelomate, psudocelomate și celomate,
clasificare, exemple, relații filogenetice, mod de viaţă
Metazoarele triploblastice se caracterizează prin faptul că în timpul dezvoltării embrionare se
formează trei foiţe embrionare: ectodermul, endodermul şi între ele mezodermul. La triploblastice apare
simetria bilaterală, fapt ce va permite dezvoltarea sistemului nervos şi a principalelor organe de simţ la
extremitatea anterioară, proces numit cefalizare.
În funcție de originea embrionară a orificiilor bucal și anal, metazoarele triploblastice au evoluat în
două ramuri: ramura protostomienilor şi ramura deuterostomienilor.
Protostomienii se caracterizează prin: blastoporul dă naştere gurii (gr. protos – primul; stoma - gură);
segmentaţia zigotului de tip spiral (planul de diviziune este oblic faţă de axa lungă a oului (axa polară, între
polul animal şi polul vegetal al oului), astfel că celulele se aşează altern unele faţă de altele; privite de la polul
animal, celulele apar dispuse în spirală); segmentaţia oului este determinată (dacă după câteva diviziuni ale
oului celulele sunt separate, fiecare celulă prin diviziune va forma numai o parte din embrion); mezodermul
derivă din celule de pe marginea blastoporului (teloblaste); grupează acelomate (inclusiv pseudocelomate) şi
celomate. La protostomienii celomaţi, celomul se formează prin schizocelie (cavitatea celomică rezultă prin
scindarea la interior a masivelor mezodermice perechi de pe fiecare latură a corpului).
Deuterostomienii se caracterizează prin: blastoporul dă naştere orificiului anal iar gura este o
neoformaţie apărută la polul opus al corpului; segmentaţia zigotului este radiară (adică planul de diviziune
este paralel sau în unghi drept faţă de axa polară); segmentaţia oului este nedeterminată, adică celulele
separate după mai multe diviziuni pot încă să se dezvoltă într-un embrion complet; toate sunt celomate;
mezodermul derivă din evaginări ale endoblastului intestinal (prin enterocelie);
Din ramura protostomienilor fac parte încrengăturile (cele mai importante): Plathelminthes, Nemertini,
Rotifera, Gastrotricha, Kinorhyncha, Nematoda, Nematomorpha, Acanthocephala, Priapulida, Sipunculida,
Mollusca, Echiurida, Annelida, Pogonophora, Tardigrada, Onychophora, Arthropoda şi grupele de lofoforate:
Phoronida, Bryozoa, Brachiopoda.
Lofoforatele prezintă caractere comune cu protostomienii și deuterostmienii. Lofoforatele se apropie
de anelide, dintre protostomieni, prin structura nefridiilor, modul de formare a gonadelor, existenţa larvei
trochofore dar se apropie şi de deuterostomieni prin formarea pe cale enterocelică a mezodermului şi prin
segmentarea trimeră a celomului.
Din ramura deuterostomienilor fac parte încrengăturile: Echinodermata, Hemichordata şi Chordata.
Separaţia între protostomieni şi deuterostomieni se crede că a avut loc acum cel puţin 520 de milioane de ani,
judecând după prezenţa ambelor grupe în fauna Cambrianului inferior.
Cavitatea corpului. În sens larg, prin cavitatea corpului se poate înţelege orice spaţiu intern, în general
plin cu un fluid, care se întinde între peretele corpului organele interne. Fluidul din cavitatea corpului poate
îndeplini o varietate de funcţii: ca schelet hidrostatic sau ca mediu circulator. Spaţiul poate servi temporar

pentru acumularea fluidelor în exces şi a excretelor, sau ca loc pentru maturarea gameţilor. Cavitatea corpului
oferă un spaţiu pentru dezvoltarea dimensională a organelor interne.
La triploblastice, apariţia mezodermului permite o organizare diferită a cavităţii corpului, în funcţie de
care triploblasticele se împart în două categorii mari: acelomate şi celomate.
Acelomate. La acelomate cavitatea corpului este cea primară (blastoceliană) şi plină cu o reţea de
celule de origine mezodermică, numită parenchim. De reținut două încrengături de acelomate: Plathelminthes
și Nemertini.
1. Încrengătura Plathelminthes (= Platyhelminthes). Acelomate cu simetrie bilaterală şi corpul puternic
turtit dorso-ventral, anterior diferenţiat într-o regiune cefalică iar posterior în una caudală. Tub digestiv
incomplet, cu un orificiu buco-anal; apare pentru prima dată în seria metazoarelor un aparat excretor –
protonefridia. Mod de viaţă. Forme libere şi parazite, majoritatea speciilor fiind parazite. Clasificare. Se
cunosc circa 5500 de specii, ce aparţin la 3 clase: Turbellaria (Gyrodactylus Dendrocoelum), Trematoda
(Fasciola, Dicrocoelim) şi Cestoidea (Taenia, Diphyllobothrium).
2. Încrengătura Nemertini. Viermi liberi (650 de specii), marini, mai puţini dulcicoli sau tereştri, cu
tegument ciliat. Prezintă o trompă protractilă și retractilă, caracteristică. Apare pentru prima dată în seria
metazoarelor un aparat circulator, format dintr-un sistem închis de vase. Exemple: 1. Clasa Anopla (trompa
fără stilet veninos) - Lineus longissimus (Marea Nordului); 2. Clasa Enopla (trompa cu stilet veninos) -
Amphiporus pulcher (Marea Mediterană), Prostoma graecense (dulcicol, Europa), Geonemertes agricola
(tericol, Australia).
Pseudocelomate. O categorie aparte de acelomate sunt pseudocelomatele. La acestea cavitatea
corpului este reprezentată de un spaţiu liber plin cu lichid în care se găsesc organele corpului, imitând o
cavitate celomică (pseudocel). Dar această cavitate nu este formată din saci mezodermici cu pereţi şi cavitate
proprii ci rezultă prin regresia parenchimului. Din această categorie fac parte încrengăturile (mai importante):
Rotifera, Gastrotricha, Kinorhyncha, Nematoda, Nematomorpha, Acanthocephala, care formează grupul
numit Nemathelminthes. Tubul digestiv este complet, deschis prin două orificii: bucal și anal. Organele
exretoare sunr reprezentate de protonefridii (rotifere) sau glande sau organe tubulare (nematode).
1. Încrengătura Rotatoria (Rotifera). Pseudocelomate (1800 specii) de dimensiuni mici sau
microscopice (0,04-3 mm lungime), care se deplasează cu ajutorul cililor; majoritatea trăiesc în ape dulci.
Exemple: Epiphanes, Brachionus.
2. Încrengătura Nematoda. Viermi cilindrici (circa 10000 de specii), lungi, filiformi, nesegmentaţi, de
dimensiuni variate; Forme libere (marine, ape dulci, terestre) şi parazite (la plante și animale). Exemple:
Enoplus sp. (marin); Dorylaimus thermae (în ape termale), D maximus (tericol), Anguillula tritici (parazit la
grâu), Ascaris sp., Ancylostoma sp. Filaria sp. (parazite la animale și om).
Celomate. La celomate, mezodermul se organizează în cavităţi ce pereţi proprii numite saci celomici.
Pe parcursul dezvoltării embrionare, cavitatea sacilor celomici, dispuşi în perechi, va substitui cavitatea
primară, blastoceliană, prin mărirea sacilor celomici şi ataşarea pereţilor acestora la peretele extern al corpului
şi la intestin. Noua cavitate de origine mezodermică, numită celomică, se numeşte secundară pentru că a
înlocuit cavitatea primară şi generală pentru că reprezintă toată cavitatea corpului. Celomatele au corpul în
mod primar segmentat. La unele grupe, precum moluştele, metameria s-a şters din primele etape ale evoluţiei
prin dezvoltarea secundară a parenchimului şi musculaturii. Există astăzi şi părerea că celomatele nu au o
origine unică, celomul apărând de mai multe ori independent, iar platelminţii sunt secundar acelomaţi.
Celomatele au un aparat circulator închis (anelide, sipunculide, foronide) sau deschis, lacunar
(moluște, artropode). Aparat respirator branhial (moluște, multe anelide, artropode primitive - limulul),
traheal (onicofore, multe artropode), pulmonar (unele artropode - araneele). Organul excretor este
metanefridia.
Dintre celomate fac parte încrengăturile (mai importante): Mollusca, Sipunculida, Echiurida,
Annelida, Pogonophora, Tardigrada, Onychophora, Arthropoda, Phoronida, Bryozoa, Brachiopoda (dintre
protostomieni), Echinodermata, Hemichoprdata şi Chordata (dintre deuterostomieni).

1. Încrengătura Mollusca. Moluştele sunt celomate protostomiene, marine, dulcicole sau terestre, cu
corpul moale, protejat, în general, total sau parţial de o cochilie calcaroasă; simetrie bilaterală sau secundar
asimetrice (gastropodele). Corpul este format din 3 regiuni: cap, picior şi masă viscerală, este acoperită de o
foiţă epidermică numită manta (=pallium). Celomul redus la cavitatea pericardică și perigonadică. Aparat
circulator deschis. Primul stadiu larvar: trocofora. Clasificare: Moluştele actuale (peste 100000 de specii) sunt
clasificate în 7 clase: Polyplacophora (ex. Chiton), Monoplacophora (ex. Neopilina), Aplacophora (ex.
Proneomenia), Gastropoda (ex. Conus, Murex, Pleurobranchus, Dendronotus, Clione, Helix, Planorbis),
Scaphopoda (ex. Dentalium), Bivalvia (ex. Nucula, Mytilus, Anodonta, Mya, Cardium) şi Cephalopoda
(Nautilus, Sepia, Loligo).
2. Încrengătura Annelida. Celomate protostomiene, cu simetrie bilaterală şi corp metamerizat
(segmentat), cu aspect vermiform; sunt numite şi „viermi inelaţi” (lat. annulus - inel). Corpul diferențiat în
cap, trunchi și pigidiu. Aparat circulator închis. Branhii sau respirație tegumentară. Metanefridii perechi.
Larva trocoforă, caracteristică multor forme marine (polichete). Clasificare: 3 clase: Polychaeta (majoritatea
marine; ex. Nereis sp., Eunice sp.), Oligochaeta (marine, dulcicole, terestre. Ex. Tubifex, Lumbricus) şi
Hirudinea (ectoparazite sau prădătoare, marine, dulcicole, terestre. Ex. Acanthobdella, Glossiphonia,
Piscicola, Hirudo).
3. Încrengătura Arthropoda. Celomate protostomiene, cu simetrie bilaterală şi corp metamerizat,
acoperit de o cuticulă chitinoasă şi prevăzut cu picioare articulate (gr. arthros – articulaţie, podos - picior).
Cavitatea corpului apare la embrion sub formă de saci celomici metamerici, care la adult se reduc, ducând la
formarea unui mixocel (= hemocel) – cavitatea primară plină cu sânge. Celomul este redus la cavităţile
gonadelor şi ale nefridiilor. Mod de viaţă. Artropodele trăiesc în toate mediile (acvatice şi terestre), libere sau
parazite. Clasificare. Sunt descrise circa 1,2 milioane de specii de artropode, clasificate în 3 subîncrengături:
Trilobitomorpha (exclusiv fosile paleozoice – Clasa Trilobita), Chelicerata (clasele Merostomata, Arachnida,
Pantopoda) şi Mandibulata (clasele: Crustacea, Myriapoda, Insecta).
4. Lophophorata. Celomate protostomiene, cu morfologie heterogenă şi mod de viaţă asemănător,
majoritatea fiind marine, sedentare, cu nutriţie microfagă şi muco-ciliară. Caracterele generale comune:
prezenţa unei coroane de tentacule ciliate, numite lofofor (gr. lophis = potcoavă; phorein = a purta), situată
în jurul gurii, pe un pliu al peretelui corpului, în formă de potcoavă sau circular. Capul este redus, modul de
viaţă sedentar; tubul digestiv are formă de U, cu anusul situat dorsal, în afara lofoforului. Corpul secretă un
înveliş cuticular, uneori îngroşat, în formă de tub (foronide), căsuță (briozoare) sau de cochilie (brahiopode).
Lofoforatele prezintă o serie de caractere comune cu deuterostomienii: segmentaţia oului este radiară la
briozoare şi brahiopode; celomul este de origine enterocelică la unele brahiopode; celomul este trimer, ca la
echinoderme. Pe baza acestor caractere, lofoforatele sunt plasate sistematic între protostomieni şi
deuterostomieni. Clasificare. Grupul Lophophorata cuprinde 3 încrengături: Phoronida (marine, solitare),
Bryozoa (majoritatea marine, puține de apă dulce, coloniale) şi Brachiopoda (marine, solitare, cochilie
bivalvă).
5. Încrengătura Echinodermata. Echinodermele sunt celomate deuterostomiene, exclusiv marine, stenohaline,
cu simetrie pentaradiară, secundar bilaterală, prevăzute cu un schelet dermic, calcaros, reprezentat de plăci şi
spini. Morfologie externă este variată, fiind numite crini de mare, stele de mare, cozi de şarpe de mare, arici
de mare, castraveţi de mare. Sunt lipsite de regiune cefalică. Cavitatea corpului este reprezentată de un celom
trimer (3 perechi de saci celomici). Caracteristic este sistemul ambulacrar – sistem acvifer de canale și
apendici (podii) cu rol locomotor și respirator. Aparatul circulator este reprezentat prin sisteme originale,
sistemul perihemal şi sistemul hemal, cu dispoziţie radiară ca şi sistemul ambulacrar. Nu există inimă. Primul
stadiul larvar este dipleurula, cu simetrie bilaterală şi celom trimer. Ea se transformă în alte tipuri de larve,
caracteristice fiecărei clase, proces în care se realizează simetria pentaradiară a adultului, după fixarea pe
substrat.
Clasificare. Se cunosc aproape 7000 de specii de echinoderme actuale, grupate în 2 subîncrengături: 1.
Pelmatozoa (cu Clasa Crinoidea – ex. Antedon, Pentacrinus) şi 2. Eleutherozoa (cu 4 clase mai importante:

Asteroidea (ex. Astropecten, Asterias), Ophiuroidea (ex. Ophioderma, Gorgonocephalus), Echinoidea (ex.
Echinus, Paracentrotus, Clypeaster), Holothuroidea (ex. Cucumaria, Holothuria)). În 1986 a fost descrisă o
nouă clasă de echinoderme actuale, margaretele de mare, Clasa Concentricycloidea, cu numai două specii din
genul Xyloplax.
Originea şi filogenia metazoarelor. Se consideră, în general, că metazoarele au o origine
monofiletică, adică provin dintr-un singur strămoş protist. Această ipoteză se bazează în special pe
asemănarea coanocitelor, celulele digestive ale spongierilor, cu coanoflagelatele (Choanoflagellida). Totuşi,
celule cu guleraş citoplasmatic, deşi nu identice cu cele ale spongierilor, au fost găsite la unele larve de
echinoderme, în oviductul unor castraveţi de mare şi la unii corali. Originea comună a spongierilor cu
celelalte metazoare este susţinută şi de datele de genetică moleculară (secvenţierea ARNr).
IX. ZOOLOGIA VERTEBRATELOR
Trecerea şi adaptări pentru viaţa în mediul terestru
Mediul terestru a constrâns vertebratele la modificări morfo-structurale, fiziologice şi
comportamentale menite să le asigure supravieţuirea şi succesul în cucerirea uscatului, oferindu-le şi
argumente suficiente pentru părăsirea mediului acvatic.
Deosebiri între mediul acvatic şi mediul terestru
Oxigenul are o concentraţie în aer de peste douăzeci de ori mai mare decât în apă; astfel, într-un litru
de aer se găsesc 210 ml O2, în timp ce într-un litru de apă volumul de oxigen variază între 3 – 9 ml, fiind
influenţat de temperatura apei şi de prezenţa celorlalte elemente dizolvate în apă. Totodată, rata difuziei
oxigenului este mult mai scăzută în apă decât în aer, ceea ce măreşte efortul animalelor acvatice pentru
procurarea oxigenului necesar.
Densitatea apei este de circa o mie de ori mai mare decât cea a aerului, iar vâscozitatea este de
aproximativ cincizeci de ori mai mare în apă. De aceea, locomoţia în apă este mai dificilă, dar densitatea
ridicată a apei asigură susţinerea corpului. Pentru deplasarea pe uscat a fost necesară remodelarea scheletului
în vederea susţinerii corpului, dezvoltarea unor membre puternice, precum şi transformarea musculaturii
scheletice.
Temperatura apei rămâne aproximativ constantă pentru lungi intervale de timp; animalele acvatice
sunt poichiloterme şi prezintă fluctuaţii mici ale temperaturii corpului. În mediul terestru, temperatura aerului
prezintă variaţii largi între zi şi noapte, de la o zi la alta şi de la un anotimp la altul astfel încât animalele
terestre au fost obligate să dezvolte structuri, mecanisme fiziologice şi tipare etologice care să le protejeze.
Homeotermia reprezintă mecanismul perfect de termoreglare, dar apare târziu, păsările şi mamiferele fiind
singurele organisme homeoterme.
Un alt parametru caracteristic mediului terestru este uscăciunea, precipitaţiile atmosferice având
regim neregulat. Supravieţuirea în aceste condiţii a fost posibilă numai prin controlul şi limitarea pierderilor
de apă.
Diversitatea ecosistemelor şi habitatelor, implicit a nişelor ecologice, sunt mai mari în mediul terestru
decât în cel acvatic, oferind condiţii pentru colonizarea lor de către o mare varietate de animale. În acelaşi
timp, mediul terestru oferă o protecţie mai bună pontei şi puilor, posibilităţi numeroase pentru refugiu şi
resurse trofice foarte diferite.
Adaptări pentru viaţa în mediul terestru În vederea trecerii la viaţa terestră, cele mai importante modificări adaptative au privit structurile care
puteau asigura respiraţia oxigenului din aer şi locomoţia în afara apei.
Plămânii au apărut ca diverticule bogat vascularizate ale segmentului iniţial al intestinului primitiv cu
care comunică, printr-un duct aerian scurt şi deschis printr-un orificiu (glotă), la nivelul faringelui, la specii de
peşti care aveau branhii funcţionale (dipnoi, crosopterigieni). Se admite că dipnoii actuali nu diferă prea mult

de speciile fosile care au trăit în Devonian; dipnoii au plămâni saciformi cu pereţi netezi, uşor plisaţi în partea
lor anterioară şi căptuşiţi cu un epiteliu ciliat, susţinut de un ţesut conjunctiv bogat vascularizat. Cele două
specii de crosopterigieni actuale – celacanţii (Latimeria chalumnae şi Latimeria menadoensis) – posedă un
singur plămân nefuncţional, fiind transformat în organ adipos; la unii crosopterigieni fosili (ripidiştii ce
populau ape dulci continentale), plămânii erau funcţionali, fiind probabil foarte asemănători cu cei de la
dipnoi.
Cavitatea nazală se separă într-un segment anterior respirator, care prezintă o pereche de nări interne
(coane) asigurând trecerea aerului spre faringe şi un segment posterior olfactiv, căptuşit cu epiteliu olfactiv.
Apariţia plămânilor care puteau realiza schimburi gazoase a impus modificarea aparatului circulator: o
pereche de arcuri aortice a format arterele pulmonare care transportă sângele venos în plămâni, iar sângele
oxigenat este adus de venele pulmonare în jumătatea stângă a inimii. A apărut astfel circuitul inimă – plămân
– inimă, similar circulaţiei mici de la tetrapode.
Consolidarea scheletului axial şi dezvoltarea membrelor, precum şi pierderea metameriei musculaturii
s-au realizat în vederea susţinerii greutăţii corporale şi pentru asigurarea locomoţiei pe uscat. La peştii dipnoi
actuali, se observă un proces de regresie evidentă a înotătoarelor perechi – dintre speciile actuale, Lepidosiren
paradoxa are înotătoarele perechi reduse la un filament mobil şi solzii acoperiţi de epidermă, ceea ce a făcut
posibilă includerea sa în grupul urodelelor multă vreme după descoperirea speciei în secolul al XIX-lea. La
dipnoii şi crosopterigienii fosili, dar şi la cele două specii de celacanţi actuali, înotătoarele perechi sunt
puternice şi musculoase, în special cele pectorale, fiind folosite în mod normal pentru înot, dar la speciile ce
populau apele dulci, puteau deveni şi vâsle care să le croiască drum pe uscat atunci când bălţile secau. În plus,
centura scapulară şi piesele de schelet ale înotătoarele pectorale prefigurează membrele anterioare ale
tetrapodelor primitive.
Un alt pas important în evoluţia tetrapodelor terestre pare să fi fost pierderea legăturii centurii
scapulare cu neurocraniul (care împiedică rotirea capului în cazul peştilor) şi apariţia gâtului care a permis
apariţia mobilităţii capului. Se presupune că, într-un stadiu iniţial, centura scapulară se termina în grosimea
musculaturii, ulterior, stabilindu-se legături noi ale centurilor cu scheletul axial, la nivelul coloanei vertebrale.
Importante modificări se înregistrează la nivelul organelor de simţ. Neuromastele liniei laterale
regresează şi apar receptori cutanaţi sub forma unor terminaţii nervoase libere sau încapsulate. Apare urechea
medie delimitată între timpan şi urechea internă, având legături cu faringele prin trompa lui Eustachio,
permiţând echilibrarea presiunii pe cele două feţe ale timpanului; în urechea medie se găseşte columela,
asigurând transmiterea vibraţiilor timpanului spre structurile urechii interne. Ochii prezintă pleoape cu rol
protector şi glande ale căror secreţii îi protejează împotriva uscăciunii; muşchii ciliari asigură acomodarea
vederii la distanţă.
Glandele tegumentare intervin în reglarea pierderilor de apă la nivelul tegumentului, iar rinichii se
perfecţionează pentru a reduce pierderile de apă prin urină.
Adaptări pentru cucerirea uscatului
Reptilele au cucerit rapid uscatul ca urmare a unor adaptări la condiţiile terestre de viaţă:
1. tegumentul prezintă un strat cornos superficial, funcţionând ca barieră împotriva uscăciunii; totodată,
glandele tegumentare sunt cvasiabsente astfel încât la acest nivel nu se pierde apă;
2. membrele sunt bine dezvoltate, permiţând susţinerea corpului şi deplasarea activă în mediul terestru;
3. encefalul prezintă emisfere bine dezvoltate și apare scoarţa cerebrală; conexiunile superioare dintre centrii
nervoşi permit realizarea unui comportament complex;
4. plămânii sunt parenchimatoşi, pereţii interni formând numeroase septuri, iar; respiraţia pulmonară asigură
în întregime necesarul de oxigen al organismului; ventilaţia pulmonară se realizează prin mişcări respiratorii
active - apare musculatura intercostală;
5. aparatul circulator este mai eficient decât cel al amfibienilor, iar septul interventricular incomplet şi
contracţia succesivă a pereţilor ventriculari reduc la minimum amestecarea sângelui venos cu cel arterial;

6. rinichiul de tip metanefros prezintă nefroni cu glomerul redus, astfel încât urina se prezintă ca o pastă
semisolidă, conţinând acid uric şi foarte puţină apă;
7. fecundaţia este internă, masculii având organe copulatoare. Embrionul prezintă anexe embrionare (amnios
şi alantoidă), fiind înconjurat de albumen şi coajă calcaroasă sau membranoasă, ceea ce permite dezvoltarea
embrionară în mediul terestru. Probabil, apariţia cojii oului a fost unul dintre factorii decisivi în ascensiunea
explozivă a reptilelor.
Încrengătura Vertebrata
Vertebratele sunt animale la care, de regulă, notocordul apare doar în timpul dezvoltării embrionare, la
adulţi, scheletul axial fiind reprezentat de coloana vertebrală şi craniu. Encefalul este adăpostit de craniu, iar
măduva spinării este protejată în canalul neural, format în partea dorsală a coloanei vertebrale.
1. Subîncrengătura Agnatha Agnatele sunt vertebrate acvatice primitive, reprezentate prin specii fosile şi prin ciclostomii actuali, care au
gura lipsită de fălci. Nu au înotătoare perechi, iar tegumentul este nud, lipsit de solzi. Notocordul este
persistent, iar la nivelul craniului, lipseşte regiunea occipitală (vertebrate hemicraniate). Prezintă un sac nazal,
deschis la exterior printr-o singură nară (monorine). Urechea internă prezintă unul sau două canale
semicirculare. Respiraţia este branhială, iar circulaţia simplă şi completă. Prezintă rinichi de tip pronefros sau
mezonofros. Au o singură gonadă, iar fecundaţia este externă.
Clasa Cyclostomata
Ciclostomii actuali sunt vertebrate acvatice şi poichiloterme. Adulţii au gura circulară, căptuşită cu odontoizi,
fiind prădători sau paraziţi, în timp ce larvele au gura sub formă de potcoavă şi sunt planctonofage şi
detritofage. Se cunosc, aproximativ, 120 de specii actuale, populând apele marine, salmastre şi dulci.
Caractere generale
1. Corpul este cilindric, serpentiform; nu au înotătoare perechi. Tegumentul este nud.
2. Notocordul este persistent, vertebrele fiind acentrice, iar craniul este lipsit de regiunea occipitală.
Musculatura flancurilor are structură metameră.
3. Sistemul nervos cuprinde encefalul redus, cu zece perechi de nervi cranieni, măduva spinării, nervii
rahidieni având rădăcinile separate.
4. Prezintă dinţi cornoşi (odontoizi); intestinul prezintă valvulă spirală şi se deschide în faţa înotătoarei anale
prin anus; glandele anexe sunt reprezentate de glande salivare, ficat şi câmpuri pancreatice.
5. Branhiile se găsesc în interiorul unor saci branhiali (marsipobranhii).
6. Aparatul circulator este închis; inima este bicamerală; prezintă sistem port-hepatic.
7. Excreţia se realizează prin rinichi de tip pronefros sau mezonefros.
8. Sexele sunt separate, fiecare individ prezentând o singură gonadă; fecundaţia este externă.
În apele curgătoare din ţara noastră trăiesc trei specii de cicari sau chişcari (Eudontomyzon sp.). Fac
parte din subclasa Petromyzontida care cuprinde specii parazite, ce se hrănesc cu lichidele interne ale
animalelor-gazdă. Sunt ciclostomi care prezintă faringe branhial, au ochii funcţionali, dar nu au chiasmă
optică. Pe laturile corpului, prezintă şapte perechi de fante branhiale externe. Nu au tentacule cefalice.
Dezvoltarea embrionară se desfăşoară cu metamorfoză, pe parcursul mai multor ani.
Chișcarii sunt ciclostomi de talie mică (15 – 25 cm), fiind prezenţi în sectorul montan al rîurilor. Coloritul
este cafeniu-cenuşiu, cu luciu metalic, abdomenul fiind alb-gălbui; înotătoarea codală este transparentă sau
negricioasă.
Eudontomyzon danfordi (chişcarul carpatic) este răspândit în tot bazinul dunărean, la noi fiind prezent, mai
ales, în rîurile din vestul, nordul şi estul ţării; stadiul larvar are o durată de 3,5 – 4 ani, în timp ce adulţii
trăiesc 17 – 21 luni şi parazitează, în special, specii de salmonide.
Eudontomyzon vladykovi (chişcarul dunărean) trăieşte în bazinele Jiu şi Argeş, iar Eudontomyzon mariae
(chişcarul ucrainean) este prezent doar, local, în bazinul Oltului, Jiului, Argeş, Siret şi Dunărea inferioară;

aceste două specii nu sunt considerate parazite, deoarece adulţii nu se hrănesc, iar larvele consumă alge
microscopice şi detritus organic.
II. Subîncrengătura Gnathostomata Gnatostomatele sunt vertebrate acvatice (peşti) şi terestre (tetrapode - amfibieni, reptile, păsări şi mamifere)
care prezintă gură cu fălci mobile. Peştii prezintă înotătoare perechi şi neperechi, iar tetrapodele au două
perechi de membre. De regulă, tegumentul prezintă produse cornoase – solzi, pene, peri - care formează un
înveliş protector al corpului. Gnatostomatele au doi saci nazali şi două nări, iar urechea internă prezintă trei
canale semicirculare. De regulă, notocordul este prezent doar în stadiul embrionar, iar craniul este complet.
Respiraţia este branhială sau pulmonară, iar circulaţia poate fi simplă sau dublă, completă sau incompletă.
Rinichii sunt de tip mezonefros sau metanefros, iar gonadele sunt perechi, sexele fiind separate. La grupele
inferioare (peşti şi amfibieni), de regulă, fecundaţia este externă şi are loc în apă; tetrapodele terestre au
fecundaţia internă, iar embrionul prezintă anexe embrionare (amnios şi alantoidă).
II. 1. SUPRACLASA Pisces
Peştii reprezintă cel mai numeros grup de vertebrate actuale - circa 32700 de specii. Sunt vertebrate primar
acvatice, populând toate tipurile de ape continentale şi marine. De regulă, corpul este hidrodinamic, acoperit
cu solzi; prezintă înotătoare perechi şi neperechi. Scheletul este cartilaginos sau osos, craniul prinzându-se
direct la coloana vertebrală. Respiraţia este branhială, dar la câteva grupe, apar şi plămânii. Inima este
bicamerală, iar circulaţia simplă şi completă, fiind prezente două sisteme port: port-hepatic şi port-renal.
Rinichii sunt de tip mezonefros. La majoritatea speciilor, fecundaţia este externă.
II. 1. 1. Clasa Chondrichthyes
Peştii cartilaginoşi, cunoscuţi sub numele de rechini, sunt peşti care au scheletul cartilaginos. Se cunosc
aproximativ 980 de specii actuale.
Caractere generale
1. Corpul este fusiform, hidrodinamic sau evident comprimat dorso-ventral; de regulă, prezintă două
înotătoare dorsale, iar codala este heterocercă. Înapoia ochilor, prezintă o pereche de spiracule.
2. Tegumentul est nud sau acoperit cu solzi placoizi.
3. Scheletul este cartilaginos. Articulaţia craniană poate fi autostilică, hiostilică sau amfistilică. Notocordul
este persistent, dar pierde metameria, vertebrele prezintă corp vertebral şi arcuri vertebrale. De regulă,
înotătoarele perechi nu au legături cu scheletul axial. Musculatura flancurilor are structură metameră.
4. Encefalul este redus, dar mai complex decât la peştii osoşi, cu zece perechi de nervi cranieni, iar nervii
rahidieni sunt micşti. Organele de simţ sunt foarte bine dezvoltate, acuitatea olfactivă şi electrorecepţia fiind
esenţiale pentru supravieţuire.
5. Intestinul prezintă valvulă spirală şi se deschide în cloacă. Glandele anexe tubului digestiv sunt
reprezentate de ficat şi pancreas.
6. Respiraţia este branhială; prezintă patru – şapte perechi de branhii.
7. Aparatul circulator este închis; inima este bicamerală, ventriculul continuându-se cu un con arterial;
prezintă sistem port-hepatic şi sistem port-renal.
8. Excreţia se realizează prin rinichi de tip mezonefros, ureterele deschizându-se în cloacă.
9. Sexele sunt separate; fecundaţia este internă, masculii având o pereche de mixopterigii, la nivelul
înotătoarelor ventrale. În funcţie de locul desfăşurării dezvoltării embrionare, sunt specii ovipare, ovo-
vivipare sau vivipare.
Speciile prezente în apele Mării Negre aparțin subclasei Elasmobranchii (Selachii) cuprinde rechinii de
suprafaţă (selacieni) şi rechinii bentonici (batoidei). Prezintă cinci perechi de branhii, iar cavitatea branhială
se deschide la exterior prin tot atâtea perechi fante branhiale. Tegumentul este acoperit cu solzi placoizi.
Prezintă rostru, iar nările şi gura sunt situate ventral. Articulaţia craniană este de tip amfistilic sau hiostilic. În
cavitatea bucală, dinţii sunt dispuşi în şiruri, înlocuindu-se pe măsura uzurii.
Sphyrna zygaena (rechin-ciocan) trăieşte în Marea Mediteraneană, de unde poate pătrunde accidental
în sectorul sudic al Mării Negre. Forma capului este determinată de dezvoltarea exagerată a cartilajelor

preorbitale şi postorbitale, rezultând cele două expansiuni la capătul cărora se găsesc ochii. Coloritul spatelui
şi flancurilor este gri-verzui, abdomenul fiind alburiu. Se hrăneşte cu peşti, dar poate consuma şi diferite
nevertebrate; execută migraţii estivale în apele nordice. Este vivipar.
Scyliorhinus canicula (rechin pătat) este prezent în Marea Mediteraneană, de unde, accidental, poate
pătrunde şi în Marea Neagră. Nu depăşeşte 80 cm lungime, iar coloritul este cenuşiu deschis, cu pete mici,
brun-întunecate. Vînează, mai ales, nevertebrate şi peşti de dimensiuni mici. Este o specie ovo-vivipară.
Squalus acanthias (rechin ţepos) este o specie prezentă şi în Marea Neagră. Dorsalele sunt precedate
de câte un spin, iar anala lipsește; coloritul flancurilor este cenuşiu, abdomenul fiind alburiu. Poate atinge 1,5
m lungime. Se hrăneşte cu peşti şi crustacee, fiind o specie ovo-vivipară.
Raja clavata (vulpe de mare, vatos), specie costieră, trăieşte şi în Marea Neagră. Corpul este romboidal
şi poate depăşi 1 m în diametru, prezentând butoni ţepoşi (inclusiv, pe pedunculul codal), formaţi prin
fuzionarea unor solzi. Dorsalele şi codala se reduc la câteva radii. Manifestă homocromie schimbătoare. Se
hrăneşte cu peşti, crustacee şi moluşte. Specie ovipară, depune ouă învelite într-o ootecă prevăzută cu
prelungiri filiforme, lungi, cu ajutorul cărora se fixează de diferite suporturi.
Dasyatis pastinaca (pisică de mare) se întâlneşte şi în Marea Neagră. Corpul este ovoid, nud, atingând
1 m lungime; dorsalele şi codala lipsesc, dar prezintă un spin caudal, rezultat prin modificarea unei radii a
înotătoarei dorsale, care poate depăşi 30 cm lungime şi la baza căruia se deschide o glandă cu secreţii
veninoase. Se hrăneşte cu peşti, crustacee şi moluşte. Este o specie ovo-vivipară.
II. 1. 2. Clasa Osteichthyes
Cuprinde peste 31500 de pești osoşi, având scheletul parţial sau complet osificat și populează apele
marine, salmastre şi dulcicole.
Caractere generale
1. De regulă, corpul este fusiform, hidrodinamic, dar se întâlnesc şi alte tipuri morfologice (serpentiform,
discoidal, sferic, cu corpul culcat pe o latură etc.); de regulă, codala este homocercă.
2. Tegumentul este acoperit cu solzi elasmoizi, mai rar ganoizi sau cosmoizi.
3. Scheletul este parţial sau complet osificat. Articulaţia craniană este hiostilică. De regulă, notocordul nu
este persistent, vertebrele prezintă corp vertebral şi arcuri vertebrale. Înotătoarele pectorale au legături cu
scheletul axial, fiind articulate la regiunea temporală a craniului. Musculatura flancurilor are structură
metameră.
4. Encefalul este redus, înconjurat de substanţă grasă, cu zece perechi de nervi cranieni, iar nervii rahidieni
sunt micşti. Organele de simţ sunt bine dezvoltate.
5. Prezintă o vezică gazoasă cu rol hidrostatic; în funcţie de legăturile acesteia cu esofagul, peştii osoşi pot fi
fizostomi sau fizoclişti.
6. Intestinul nu prezintă valvulă spirală, dar poate prezenta apendici pilorici şi se deschide pe marginea
anterioară a papilei ano-genito-urinare, în faţa analei. Glandele anexe tubului digestiv sunt reprezentate de
ficat şi câmpurile pancreatice.
7. Respiraţia este branhială; prezintă patru perechi de branhii, cavitatea branhială fiind acoperită de un
opercul osos, format din trei – patru oase. La numeroase grupe, apar organele respiratorii accesorii.
8. Aparatul circulator este închis; inima este bicamerală, ventriculul continuându-se cu un bulb arterial;
prezintă sistem port-hepatic şi sistem port-renal.
9. Excreţia se realizează prin rinichi de tip mezonefros, ureterele deschizându-se pe marginea posterioară a
papilei ano-genito-urinare.
10. Gonoductele se prind direct de gonade şi se deschid în segmentul median al papilei ano-genito-urinare.
Sexele sunt separate; fecundaţia este externă, majoritatea speciilor fiind ovipare.
Huso huso (morun), cel mai mare dintre sturioni (atinge 5 – 6 m lungime), este prezent și în Marea
Neagră, fiind o specie vulnerabilă. Prezintă cinci şiruri longitudinale de scuturi dermice. Are rostrul scurt, uşor
arcuit în sus, iar mustăţile aplatizate sunt prinse în apropierea gurii; deschiderea gurii este foarte largă,

semilunară. Este o specie ihtiofagă, dar consumă şi crustacee. Primăvara, migrează în Delta Dunării şi pe
cursul inferior al Dunării pentru reproducere.
Alosa immaculata (scrumbie de Dunăre) trăieşte în Marea Neagră; migrează în Dunăre pentru
reproducere (migraţie amfibiotică anadromă). Prezintă câte doi solzi alari pe fiecare faţă a codalei, iar carena
ventrală din fața înotătoaei anale este foarte evidentă. Se hrăneşte cu peşti mărunţi şi crustacee. Carnea are un
conţinut foarte ridicat de lipide (18 - 20%).
Salmo trutta fario (păstrăv indigen), la noi, este prezent în zona montană, preferând apele reci şi repezi.
Are două dorsale, a doua fiind adipoasă. Coloritul este verde-măsliniu deschis, cu pete negricioase spre spate,
inclusiv pe dorsala cu radii, iar pe flancuri, prezintă pete ocelare (roşu tivit cu alb). Se hrăneşte cu
nevertebrate, dar adulţii pot consuma peşti şi amfibieni; execută o migraţie anadromă, spre izvoare, în vederea
reproducerii.
Cyprinus carpio (crap) este larg răspândit în apele lin curgătoare şi stagnante din zona de câmpie şi de
deal. Corpul este relativ înalt şi comprimat lateral, acoperit cu solzi cicloizi. Dorsala are baza lungă; dorsala şi
anala au prima radie tare şi zimţată. Gura are buze groase şi prezintă două perechi de mustăţi. Coloritul
spatelui este brun-verzui, abdomenul fiind alb-gălbui.
Perca fluviatilis (biban) populează toate apele dulci din ţara noastră, exceptând sectorul montan al
apelor curgătoare. Are două dorsale, prima având radiile tari, iar solzii sunt ctenoizi. Coloritul este verzui, cu 5
– 9 benzi transversale întunecate, iar înotătoarele ventrale şi cea anală, precum şi vârfurile codalei sunt
roşietice. La marginea posterioară a primei dorsale, există o pată neagră.
II. 2. SUPRACLASA Tetrapoda
Tetrapodele sunt vertebrate terestre, prezente în cele mai diverse ecosisteme şi habitate. Forma corpului este
variată; exceptând amfibienii care au corpul nud, celelalte tetrapode au corpul acoperit cu produse cornoase
ale tegumentului - solzi, pene, peri etc. Au două perechi de membre, conformate pentru diverse tipuri de
locomoţie. Scheletul este osos, iar articulaţia dintre craniu şi coloana vertebrală este, de regulă, mobilă.
Respiraţia este pulmonară, exceptând amfibienii care pot respira şi prin intermediul tegumentului sau
branhiilor. Inima este tri- sau tetracamerală, iar circulaţia dublă, incompletă sau completă; sunt prezente două
sisteme port: port-hepatic şi port-renal, ultimul fiind în regresie (dispare la mamifere). Rinichii sunt de tip
mezonefros sau metanefros. La majoritatea speciilor, fecundaţia este internă (exceptând amfibienii anuri).
Amfibienii sunt tetrapode non-amniote, dezvoltarea embrionară realizându-se prin metamorfoză, în apă;
celelalte grupe sunt vertebrate amniote - embrionul prezintă anexe embrionare: amniosul şi alantoida.
II. 2. 1. Clasa Amphibia Amfibienii sunt vertebrate care trăiesc pendulând între mediul terestru şi cel acvatic, amfibienii actuali (circa
7000 de specii) fiind încă incomplet adaptaţi pentru viaţa pe uscat.
Caractere generale
1. Există trei tipuri morfologice: trituriform, raniform şi serpentiform.
2. Prezintă două perechi de membre; degetele sunt adeseori unite prin membrane interdigitale şi nu au
gheare sau unghii.
3. Tegumentul este subţire şi umed; nu prezintă solzi cornoşi; au numeroase glande mucoase, iar la unele
specii, există şi glande veninoase; uneori, prezintă homocromie schimbătoare.
4. Scheletul este complet osificat; craniul prezintă doi condili occipitali; notocordul dispare la adulţi, iar
numărul vertebrelor este variabil; coastele lipsesc la unele specii. Nu prezintă exoschelet.
5. La nivelul encefalului, emisferele cerebrale se individualizează. Au zece perechi de nervi cranieni.
6. Gura este largă; prezintă dinţi pe ambele fălci sau numai pe cea superioară; cavitatea nazală comunică prin
coane cu cea bucală. Prezintă cloacă.
7. Respiraţia poate fi branhială, pulmonară, tegumentară sau faringiană; la unele specii, branhiile persistă
întreaga viaţă.
8. Inima este tricamerală (două atrii şi un ventricul), circulaţia fiind dublă şi incompletă; prezintă două
sisteme port: port-hepatic şi port-renal; vascularizaţia tegumentului este foarte bogată.

9. Sunt tetrapode poichiloterme cu rata metabolică redusă.
10. Prezintă doi rinichi de tip mezonefros; urina este diluată şi conţine uree.
11. Sexele sunt separate; fecundaţia este externă la anure, internă la urodele şi gimnofioni. Majoritatea sunt
specii ovipare, ovo-viviparitatea fiind foarte rară; la unele grupe, se manifestă fenomenul de neotenie. Ouăle
sunt mezolecite, cu membrană gelatinoasă. Dezvoltarea embrionară se face prin metamorfoză.
Bombina bombina (buhai de baltă/izvoraş cu burtă roşie), specie diurnă şi crepusculară, prezentă în
regiunea de câmpie şi de deal. Coloritul este brun-cenuşiu pe spate, iar abdomenul este negru-albăstrui cu pete
roşii neregulate. Tegumentul are aspect rugos (cu negi) şi conţine glande cu secreţii iritante. Masculii au doi
saci vocali. În caz de pericol, în apă se ascunde în mâl, iar pe uscat, simulează moartea răsucindu-se cu
abdomenul în sus şi rămânând nemişcat.
Pelobates fuscus (broască brună de pământ) este prezentă în regiunile cu soluri afânate sau nisipoase la
altitudini mai mici de 600 m. Pe latura externă a tarsului, prezintă o lamă cornoasă cu aspect semilunar, iar
tegumentul este neted şi pupila verticală. Coloritul este brun-deschis cu pete întunecate pe spate, iar
abdomenul este alb-cenuşiu, pătat uneori cu brun-cenuşiu. Masculii nu au saci vocali şi calozităţi nupţiale, dar
prezintă o glandă humerală.
Bufo viridis (broască râioasă verde) este o specie ubicvistă prezentă chiar și în interiorul localităților.
Tegumentul dorsal prezintă negi, iar înapoia ochilor se găsesc glandele parotoide cu aspect oval-semilunar
(secretă bufonina). Pupila este orizontală. Nu are dinţi. Dorsal, pe fond cenuşiu-deschis prezintă pete verzi-
măslinii, iar abdomenul este alburiu cu pete întunecate. Masculii au un sac vocal submandibular, iar
calozităţile nupţiale sunt negre.
Salamandra salamandra (sălămâzdră) este o specie de urodele prezentă în munții Carpaţi, zona
Subcarpaţilor şi în Podişul Transilvaniei. Înapoia ochilor prezintă glande parotoide cu aspect reniform.
Coloritul este negru strălucitor cu pete neregulate galbene. Este nocturnă şi ovo–vivipară (femela naşte larve
cu branhii şi membre, având dimensiuni mici şi care rămân în apă aproximativ 12 săptămâni).
Triturus cristatus (sălămâzdră/triton cu creastă) se întâlneşte din zona de și deal; pentru reproducere,
intră în bălţi cu vegetaţie submersă. Flancurile şi spatele au culoarea brun-roşcată, iar abdomenul este galben-
portocaliu cu pete neregulate cenuşii-întunecate. În perioada de reproducere, masculul prezintă o creastă
dorsală înaltă şi adânc crestată, iar regiunea ventrală a cozii se colorează în alb-albăstrui.
II. 2. 2. Clasa Reptilia
Reptilele sunt primele tetrapode care cuceresc definitiv uscatul; majoritatea speciilor actuale (circa 10030 de
specii) se deplasează prin târâre, membrele asigurând doar susţinerea corpului.
Caractere generale
1. Forma corpului este variată, întâlnindu-se trei tipuri morfologice principale: cheloniform (reptile cu corpul
turtit dorso-ventral şi exoschelet bine dezvoltat), lacertiliform (reptile cu corpul fusiform şi două perechi de
membre egale) şi serpentiform (reptile cu corp cilindric şi apode).
2. Au două perechi de membre pentadactile adaptate pentru mers, căţărat sau înot; la şerpi şi unele şopârle,
membrele lipsesc.
3. Tegumentul uscat, cu puţine glande; stratul cornos superficial este înlocuit prin năpârlire periodică.
4. Craniul prezintă un condil occipital; apare cuşca toracică formată din stern, coaste şi câteva vertebre
toracice; la şerpi sternul lipseşte. Broaştele ţestoase au exoschelet foarte bine dezvoltat.
5. La nivelul encefalului, apare scoarţa cerebrală. Au doisprezece perechi de nervi cranieni.
6. De regulă, gura este largă şi prezintă dinţi pe ambele fălci; broaştele ţestoase prezintă cioc cornos; la
crocodili, apare bolta palatină secundară, separând complet calea digestiva de cea respiratorie. Stomacul este
de tip triturator. Prezintă cloacă.
7. Respiraţia este strict pulmonară; arcurile branhiale apar în timpul vieţii embrionare.
8. Inima este tricamerală, cu excepţia crocodililor care au inima tetracamerală; prezintă o pereche de cârje
aortice. Circulaţia este dublă şi incompletă.

9. Sunt vertebrate poichiloterme; la unele specii există adaptări comportamentale pentru asigurarea
termoreglării.
10. Rinichii sunt de tip metanefros; urina conţine acid uric.
11. Sexele sunt separate; fecundaţia este internă. Sunt vertebrate ovipare, viviparitatea fiind extrem de rară.
Ouăle au coaja membranoasă. Apar anexele embrionare: amniosul şi alantoida; embrionul prezintă, de
asemenea, corion şi sac vitelin.
Emys orbicularis (broască ţestoasă de lac) trăieşte în ape lin curgătoare şi stătătoare. Are carapacea
aplatizată, iar membrele lungi, cu autopodul lăţit şi degetele unite prin membrane interdigitale. Coloritul este
negricios-verzui cu pete mici galbene. Este o specie crepuscular-nocturnă şi iernează îngropată în mâl.
Lacerta viridis (guşter) este o specie larg răspândită în poieni şi liziere de pădure, putând ajunge până
la 1000 m altitudine. Nu depăşeşte 40 cm lungime şi este cel mai mare reprezentant al familiei în România.
Prezintă dimorfism sexual: masculul este verde şi are guşa azurie în sezonul de reproducere, iar femela este
verde-măslinie cu punte negre şi are treimea posterioară cafenie.
Anguis colchica (şarpe de sticlă, năpârcă) trăieşte în locuri deschise; lipseşte din Dobrogea. Specie
apodă, are corpul acoperit cu solzi netezi şi lucioşi, coloritul fiind arămiu; își poate rupe coada.
Eryx jaculus (şarpe de nisip, boa de nisip) este singurul reprezentant al familiei Boidae în România
(deosebit de rar, apare în zone cu soluri nisipoase sau afânate din Dobrogea și lunca Dunării). Nu depăşeşte 80
cm lungime şi are coloritul gălbui-cenuşiu cu pete neregulate brune. Este scurt și gros; aspectul general și
coloritul capului, respectiv, al cozii este similar. Gura este plasată ventral, iar partea anterioară a corpului, dură
și ascuțită, servește pentru săparea galeriilor subterane în care se adăpostește, fiind o specie nocturnă. Este
ovipar.
Natrix natrix (şarpe de casă) se întâlneşte de la ţărmul mării până la altitudinea de 1100 m, în
apropierea apelor, fiind un foarte bun înotător. Are corpul zvelt acoperit cu solzi carenaţi; coloritul flancurilor
şi spatelui este cenuşiu, cu două pete temporale alb-gălbui; abdomenul prezintă un desen negru cu alb, negrul
predominând.
II. 2. 3. Clasa Aves
În prezent, păsările constituie cel mai divers grup de vertebrate terestre - circa 10050 de specii actuale;
numărul variază de la un autor la altul, în funcţie de criteriile taxonomice folosite.
Caractere generale
1. Corpul este fusiform, aerodinamic, alcătuit din cap, gât, trunchi şi două perechi de membre. Cele
anterioare sunt transformate în aripi, iar membrele posterioare au patru degetele libere sau unite prin
membrane interdigitale, terminate cu gheare.
2. Tegumentul este uscat, lipsit de glande; prezintă o serie de produse cornoase – pene, cioc, solzi şi gheare.
3. Scheletul este alcătuit din oase subţiri şi pneumatice; craniul prezintă un condil occipital; la nivelul
coloanei vertebrale, sunt prezente osul sinsacrum şi pigostilul; coastele sunt alcătuite dintr-un segment sternal
şi unul vertebral, prezentând apofiză uncinee; sternul poate fi carenat sau acarenat. La nivelul autopodului, se
formează carpometacarpul, respectiv, tarsometatarsul. Musculatura care asigură zborul este foarte
voluminoasă.
4. La nivelul encefalului, emisferele cerebrale sunt voluminoase şi netede, iar cerebelul este bine dezvoltat şi
cutat. Au doisprezece perechi de nervi cranieni.
5. Nu au dinţi, oasele fălcilor fiind acoperite de cioc. Stomacul prezintă un segment triturator şi unul
glandular. Prezintă cloacă tricompartimentată.
6. Sirinxul este organul fonator, plămânii sunt parenchimatoşi, de tip tubular şi prezintă saci aerieni. În timpul
zborului, respiraţia este dublă.
7. Inima este tetracamerală (două atrii şi două ventricule), circulaţia fiind dublă şi completă; prezintă cârjă
aortică dreaptă, iar sistemul port-renal intră în regresie; vascularizaţia pulmonară este deosebit de bogată.
8. Sunt tetrapode homeoterme cu rata metabolică foarte ridicată.
9. Prezintă doi rinichi de tip metanefros şi nu au vezică urinară; urina este concentrată şi semisolidă.

10. Sexele sunt separate, iar femela prezintă doar jumătatea stângă a aparatului genital; fecundaţia este
internă. Sunt tetrapode ovipare, embrionul şi anexele sale fiind acoperite cu mai multe membrane şi o coajă
calcaroasă. Asigură clocitul ouălor şi au un comportament parental complex.
Cygnus olor (lebădă de vară) este o specie migratoare parţial în România, mai ales în regiunile sudice;
majoritatea exemplarelor care iernează la noi, provin din populaţii nordice. Preferă bălţile şi lacurile cu mult
stufăriş. Are penajul alb strălucitor, iar ciocul adultului este roşu-portocaliu, cu o protuberanţă bazală neagră.
În timpul zborului, gâtul şi picioarele palmate sunt perfect întinse, auzindu-se şuieratul aripilor. Păsările tinere
au penajul maroniu în primul an de viaţă.
Pelecanus onocrotalus (pelican comun) este oaspete de vară, cuibărind în colonii, în stufărişuri şi
mlaştini sau pe plauri. Penajul este alb, însă are remige negre şi subalare albe, pe faţa inferioară, aripa fiind
albă cu o bordură neagră. Sacul gular galben, iar picioarele de tip stegan sunt roz-maronii. Apare în Cartea
Roşie a Vertebratelor din România, având statut de specie vulnerabilă.
Ciconia ciconia (barză albă, cocostârc) preferă să-şi instaleze cuibul pe suporturi înalte, în perimetrul
localităţilor. Penajul este alb cu negru: gâtul, jumătatea anterioară a aripilor, spatele şi abdomenul sunt albe;
jumătatea posterioară a aripilor are culoare neagră. Ciocul este lung, de culoare roşu-portocaliu; la fel şi
picioarele. Apare în Cartea Roşie a Vertebratelor din România, cu statut de specie vulnerabilă.
Haliaeetus albicilla (codalb), numit impropriu „vultur”, este cea mai mare acvilă care cuibăreşte în
prezent în România, în special, în Delta Dunării. Apare în Cartea Roşie a Vertebratelor din România, cu
statut de specie critic periclitată. Coloritul adulţilor este predominant brun, cu capul cafeniu deschis şi coada
rotunjită, albă. Cuibăreşte solitar, amenajând cuiburi voluminoase în vârful unor arbori bătrâni.
Dendrocopos major (ciocănitoare pestriţă mare) specie sedentară comună în păduri şi, relativ,
frecventă în parcuri, grădini. Coloritul este o combinaţie de negru şi alb, obrajii fiind albi, traversaţi de o linie
neagră. La masculi, pe ceafă, este prezentă o pată roşie. Piciorul este de tip scansor, iar ciocul lung, conic şi
puternic, folosindu-l ca daltă pentru a sparge scoarţa arborilor, în căutarea insectelor şi larvelor acestora, pe
care le „culege” cu ajutorul limbii lungi şi lipicioase. Coada este folosită pentru sprijinire, rectricele fiind
rigide. Cuibul este săpat în trunchiul arborilor, cu ajutorul ciocului.
II. 2. 4. Clasa Mammalia
Mamiferele constituie grupul celor mai evoluate vertebrate, fiind prezente în toate ecosistemele terestre, din
zonele calde până în regiunile polare, dar şi în ecosisteme acvatice. În prezent, pe Terra, trăiesc aproximativ
5488 de specii de mamifere.
Caractere generale
1. Corpul are forme variate, fiind alcătuit din cap, gât, trunchi, coadă şi două perechi de membre, conformate
pentru diferite moduri de locomoţie.
2. Tegumentul prezintă mai multe tipuri de glande (sebacee, sudoripare, odorante, mamare) şi numeroase
produse cornoase (peri, solzi, gheare, unghii sau copite, coarne etc.).
3. Scheletul este alcătuit din oase mari, pline cu măduvă; craniul de tip platibazic şi prezintă doi condili
occipitali; în urechea medie, există un lanţ de trei oscioare (ciocanul, nicovala şi scăriţa). La nivelul coloanei
vertebrale, este prezent osul sacrum; coastele sunt arcuri osoase, care, în funcţie de articularea la stern, pot fi
adevărate, false sau flotante. Muşchii caracteristici mamiferelor sunt diafragmul, muşchii pieloşi şi muşchii
feţei.
4. La nivelul encefalului, emisferele cerebrale şi cerebelul sunt voluminoase, acoperind toate celelalte părţi
componente, iar scoarţa cerebrală atinge dezvoltarea maximă. Au doisprezece perechi de nervi cranieni.
5. Tubul digestiv se întinde între orificiul bucal şi cel anal, deschis direct, la exterior; nu au cloacă. Au dinţi
tecodonţi, dentiţia fiind, de regulă, heterodontă, monofiodontă sau difiodontă. Stomacul poate fi simplu sau
pluricompartimentat. Glandele anexe tubului digestiv sunt glandele salivare, ficatul şi pancreasul.
6. Laringele este organ fonator, iar plămânii sunt parenchimatoşi şi lobaţi, căile intrapulmonare formând
arborele bronşic.

7. Inima este tetracamerală (două atrii şi două ventricule), circulaţia fiind dublă şi completă; prezintă cârjă
aortică stângă, o singură venă cavă anterioară (superioară), precum şi un singur sistem-port, cel hepatic.
8. Sunt tetrapode homeoterme.
9. Prezintă doi rinichi de tip metanefros şi au vezică urinară, care se deschide, prin uretră, în sinusul uro-
genital.
10. Sexele sunt separate, iar fecundaţia este internă. Cu puţine excepţii, mamiferele nasc pui, dezvoltarea
embrionară desfăşurându-se la nivelul uterului, pe seama unei placente. Puii sunt alăptaţi, fiind dependenţi de
femele pentru un interval de timp mai lung sau mai scurt. Numeroase specii de mamifere sunt animale
sociale, cu manifestări comportamentale complexe.
Rhinolophus ferrum-equinum (liliac mare cu potcoavă) este un liliac care prezintă excrescenţe
foliacee în jurul nărilor, dispuse sub formă de potcoavă, dar nu are tragus. Blana este moale şi pufoasă, brun-
cenuşie; este unul dintre cei mai mari lilieci din Europa (anvergura aripilor este de 40 cm); specie sedentară,
se deplasează pe distanţe foarte scurte (cel mult 30 – 40 km) între locurile de iernat şi teritoriile de hrănire
folosite în timpul verii.
Erinaceus europaeus (arici) este o specie palearctică, lipsind din munţii Himalaya şi din nordul
Africii. Blana moale de pe partea ventrală a corpului şi bot este cenuşiu-negricioasă, iar ţepii sunt cenuşii, cu
baza şi vârful alb-gălbui. Este omnivor, sapă vizuini şi poate hiberna în unele regiuni ale arealului său
(inclusiv, în România).
Capreolus capreolus (căprior) se întâlneşte în păduri rare şi locuri semideschise. Blana de vară este
roşcat-cafenie, iarna devenind brun-cafenie. Masculii au coarne cu două – trei ramuri scurte.
Delphinus delphis (delfin comun) este o specie prezentă în largul mării; prezintă mai multe subspecii,
inclusiv, una endemică în Marea Neagră – Delphinus delphis ponticus. Ciocul nu este prea lung, iar melonul
este rotunjit. Coloritul este cenuşiu-întunecat, cu flancurile în nuanţe deschise – crem-gălbui în regiunea
toracică şi cenuşiu deschis pe pedunculul codal.
Lynx lynx (rîs) populează pădurile montane, fiind un căţărător deosebit de agil; la noi, este specie
protejată. Blana este cenuşiu-roşcată, cu pete mărunte negre, iarna devenind suriu-argintie; partea ventrală a
corpului este albă; în vârful pavilioanelor urechilor prezintă smocuri de peri lungi, iar pe marginile obrajilor,
peri lungi şi deşi formează „favoriţi”; coada este scurtă.