
e n z i m e
237
Cursul 5 E N Z I M E • Caracteristici generale ale enzimelor • Însuşiri caracteristice enzimelor • Complexul ES • Centrul activ al enzimelor • Energia de activare a reacţiilor catalizate enzimatic
-
Upload
iuliana-july -
Category
Documents
-
view
40 -
download
2
description
enzime biochimie
Transcript of e n z i m e

Cursul 5
E N Z I M E
• Caracteristici generale ale enzimelor
• Însuşiri caracteristice enzimelor
• Complexul ES
• Centrul activ al enzimelor
• Energia de activare a reacţiilor catalizate enzimatic

Caracteristici generale ale enzimelor
• Ce sunt enzimele ?
catalizatori ai reacţiilor chimice din organismele vii

Însuşirile comune cu catalizatorii
• acţionează în concentraţii foarte mici;
• nu se consuma în cursul reacţiei;
• nu modifică constantele de echilibru ale reacţiilor pe care le catalizează;
• grăbesc numai atingerea stării de echilibru pentru reacţiile termodinamic posibile.

Însuşiri caracteristice enzimelor
A.Sunt macromolecule proteice
B. Au eficienţă catalitică mult mai mare
C.Au specificitate
D. Activitatea lor poate fi reglată

A.Cu excepţia ribozimilor (ARN)
enzimele sunt invariabil
proteine

• Structură:
a. enzime cu structura strict proteică:
ribonucleaza
chimotripsina
lizozimul din mucusul nazal şi lacrimi;

b. enzime cu structura heteroproteică: parte proteica numită „apoenzimă“
partea neproteică numită cofactor:
coenzimă = componentă neproteică organică, legată slab necovalent de apoenzimă;
grupare prostetică = componentă organică legată puternic de apoenzima (ex. hemul)
metal- legat permanent sau temporar

B.Enzimele asigură viteze mari reacţiilor
• reacţiile enzimatice sunt de 106 - 1012 ori mai rapide ca cele neenzimatice
a.hidratarea CO2 este catalizată de anhidraza carbonică , cea mai
activă enzimă:
105 molecule/ secundă
reacţia este de 107 ori mai rapidă decât reacţia necatalizată
CO2 + H2O H2CO3
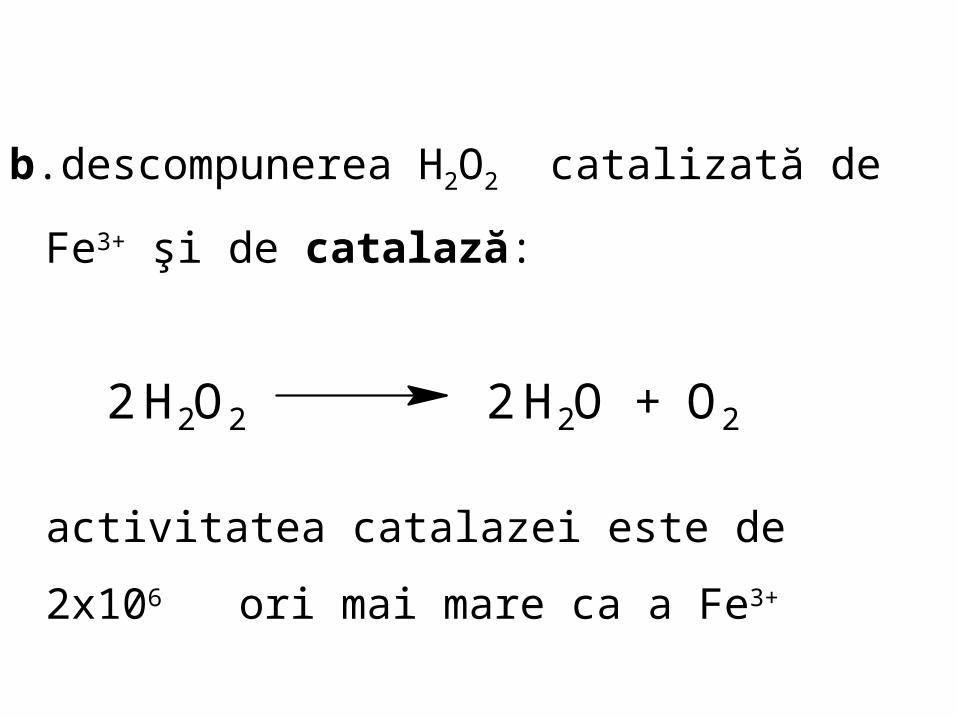
b.descompunerea H2O2 catalizată de Fe3+ şi de
catalază:
activitatea catalazei este de 2x106 ori mai
mare ca a Fe3+
2 H2O2 2 H2O + O2

C.Specificitate
1. de reacţie
2. de substrat: -absolută
-relativă: -de grup
-mai largă
3. stereospecificitate

1.Specificitate de reacţie
Substrat + Reactant Produşi
• clasificarea enzimelor:
Oxido-reductaze;
Transferaze;
Hidrolaze;
Liaze;
Ligaze.

2.Specificitate de substrat (selectivitate în alegerea substratului)
a.Specificitate absolută
E utilizează ca S un singur compus chimic:
-ureaza catalizează hidroliza ureei:
-acetilcolinesteraza catalizează hidroliza esterului colinei cu acidul acetic:
-anhidraza carbonică
H2N CO NH2 + 2 H2O CO2 + 2 NH3
H3C COO CH2 CH2
N+(CH3)3
+ H2O H3C COOH + H2C CH2
N+(CH3)3HO
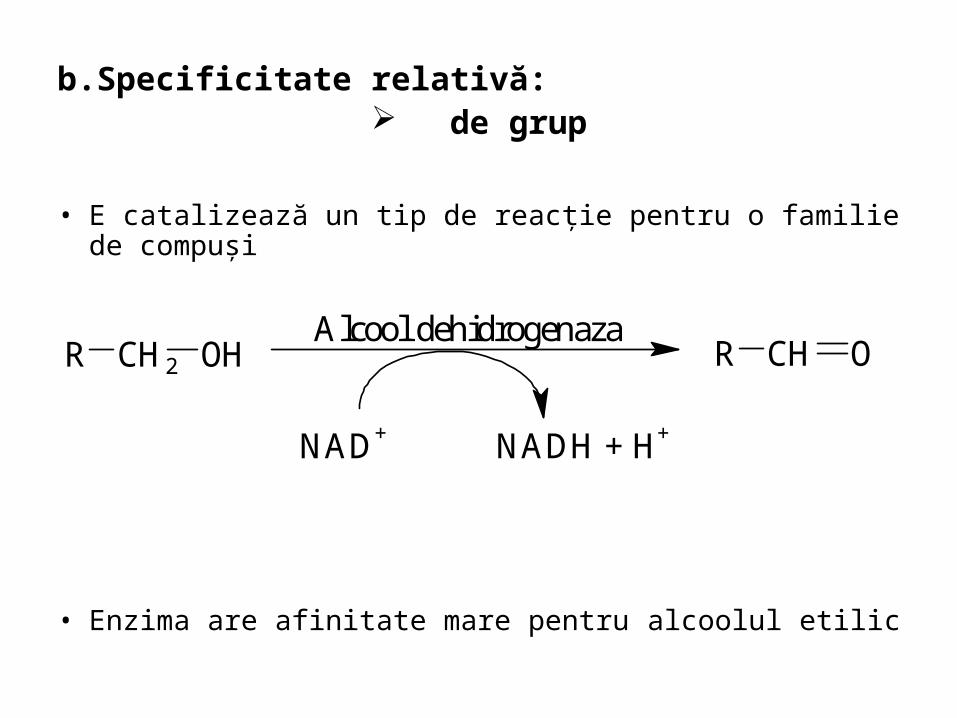
b.Specificitate relativă: de grup
• E catalizează un tip de reacţie pentru o familie de compuşi
• Enzima are afinitate mare pentru alcoolul etilic
R CH2 OHAlcool dehidrogenaza
NAD+ NADH + H+
R CH O
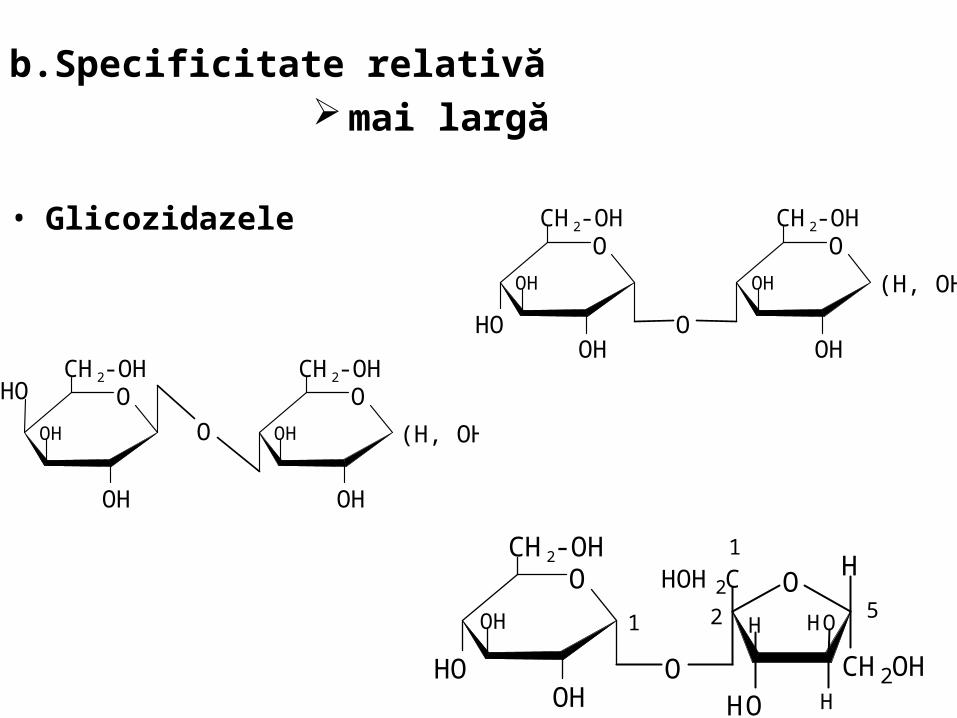
b.Specificitate relativă mai largă
• Glicozidazele
(H, OH)
O
HO
OH
OH
CH2-OHO
OH
OH
CH2-OH
O
O
HO
OH
OH
CH2-OH
O
O
OH
H
OH
CH2OHH
H
HOH2C
1
1
2 5
(H, OH)
OHO
OH
OH
CH2-OHO
OH
OH
CH2-OH
O

Proteazele
NH CH CO NH
R2
CH CO
R3
R3 = fenilalaninã, tirozinãmetioninã, leucinã
Pepsinã

NH CH CO NH
R2
CH CO
R3
R2 = lizinã, argininã
Tripsinã
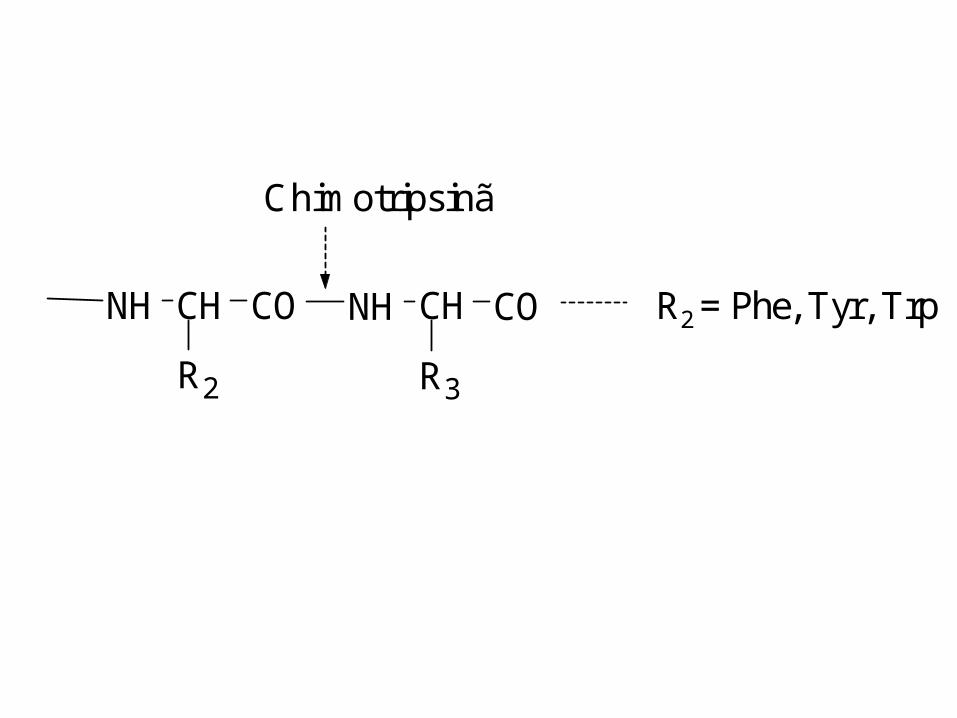
NH CH CO NH
R2
CH CO
R3
R2 = Phe, Tyr, Trp
Chimotripsinã
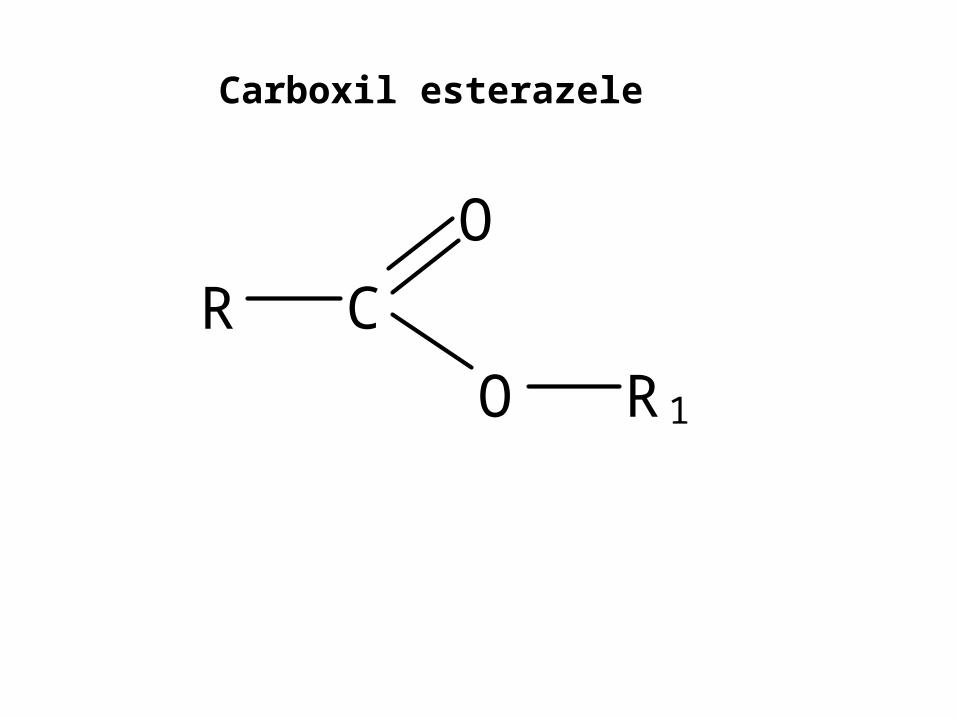
Carboxil esterazele
C
O
O R1
R

3.Specificitate stereochimică
• Enzimele disting
-un enantiomer levogir de cel dextrogir;
-un izomer cis de cel trans.
Lactat dehidrogenaza
NAD+ NADH + H+
COOH
C
CH3
HHO
Acid L-lactic
COOH
C
CH3
O
Acid piruvic
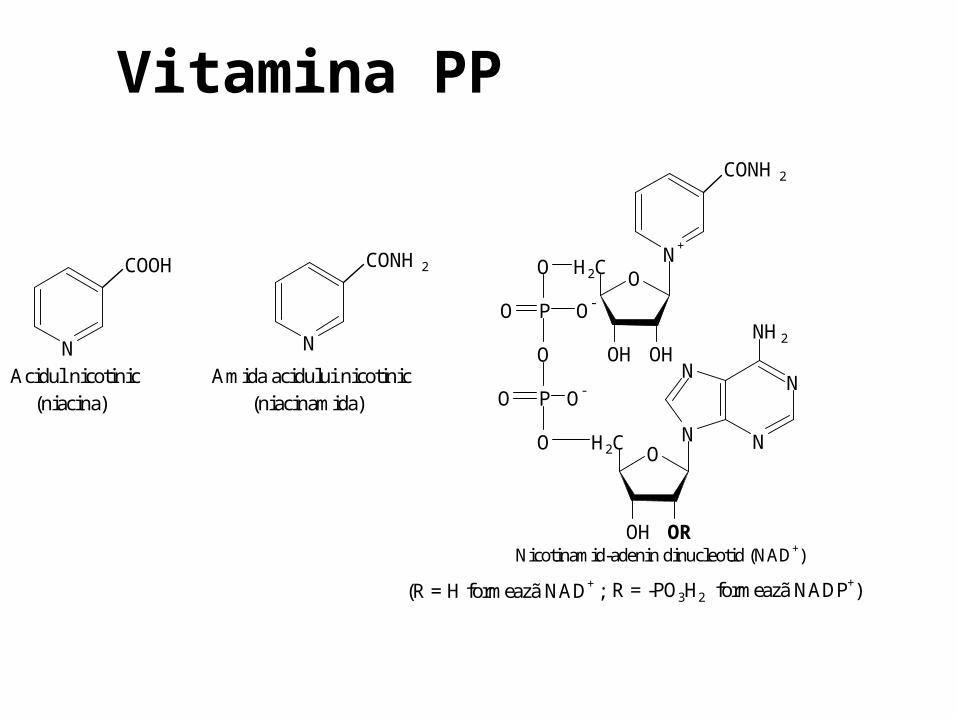
O N
N
N
N
NH2
OH2C
OROH
O P O-
O
O P O-
ON+
OH2C
OHOH
CONH2
N
COOH
Acidul nicotinic Amida acidului nicotinic (niacina) (niacinamida)
N
CONH2
Nicotinamid-adenin dinucleotid (NAD+)
(R = H formeazã NAD+ ; R = -PO3H2 formeazã NADP+)
Vitamina PP
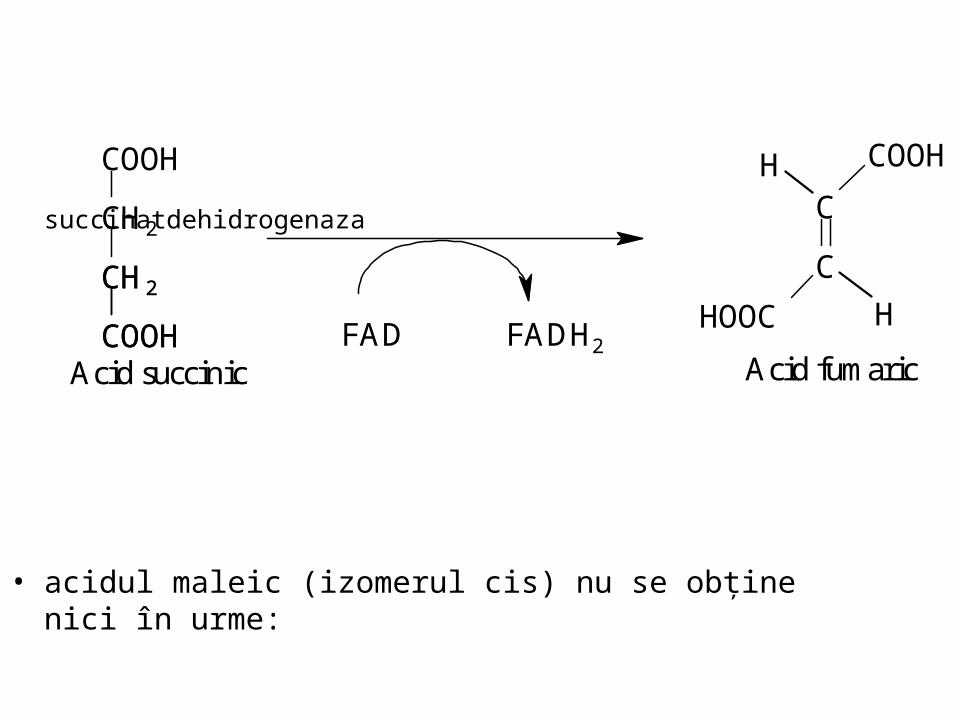
succinatdehidrogenaza
• acidul maleic (izomerul cis) nu se obţine nici în urme:
FAD FADH2
CH2
CH2
COOH
COOH
Acid succinic
COOH
C
C
HOOC
H
H
Acid fumaric
CH2
COOH

Vitamina B2
• Conţine heterociclul numit izoaloxazină
N N
NNH
CH2
O
OCH3
CH3
CH CH CH
OH OH OH
OCH2 P O
O-
O
P O
O-
O
N
N
N
N
NH2
OH2C
OHOH
lumicrom
lumiflavinã
riboflavinã
flavin adenin mononucleotid (FMN)
adenozinã
adenozin 5`-monofosfat(AMP)
flavin adenin dinucleotid (FAD)
(H)
(H)
(H)(H)
(OH)

• 6.Activitatea catalitică a enzimelor poate fi
modulată de anumiţi agenţi reglatori.


Enzimele se deosebesc de catalizatorii anorganici prin:A.Nu modifică echilibrele chimice la care conduc reacţiile reversibile
B.Prezintă specificitate de reacţie
C.Majoritatea sunt proteine
D.Intervin numai în reacţii chimice posibile din punct de vedere termodinamic
E.Acţiunea lor este susceptibilă de reglare

Complexul ES
• reprezentare reacţii enzimatice :
E + S ES E + Produsi

Centrul activ al enzimelor
• Enzimele sunt macromolecule
care prezintă
Centrul activ = entitate tridimensională
unde se leagă substratul

• Centrul activ al enzimelor =
– o regiune restrânsă din proteină
– cu structură
chimică şi
geometrică
bine
determinate

Cine conferă structura Centrului Activ ?
–natura resturilor aminoacidice şi
–organizarea secundară,
terţiară şi
cuaternară a
proteinei.

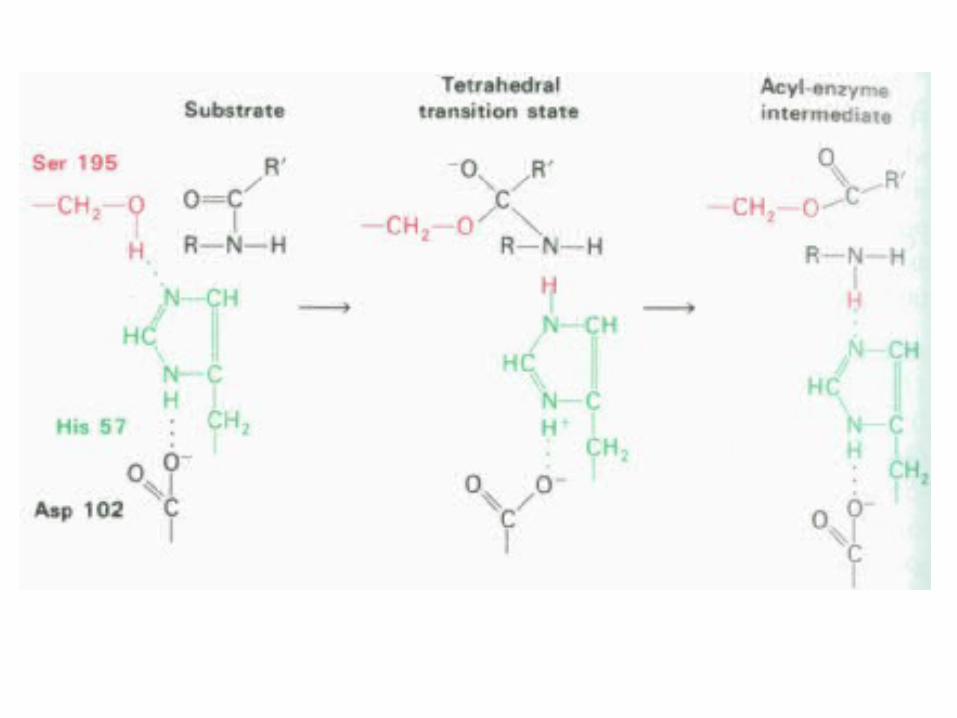
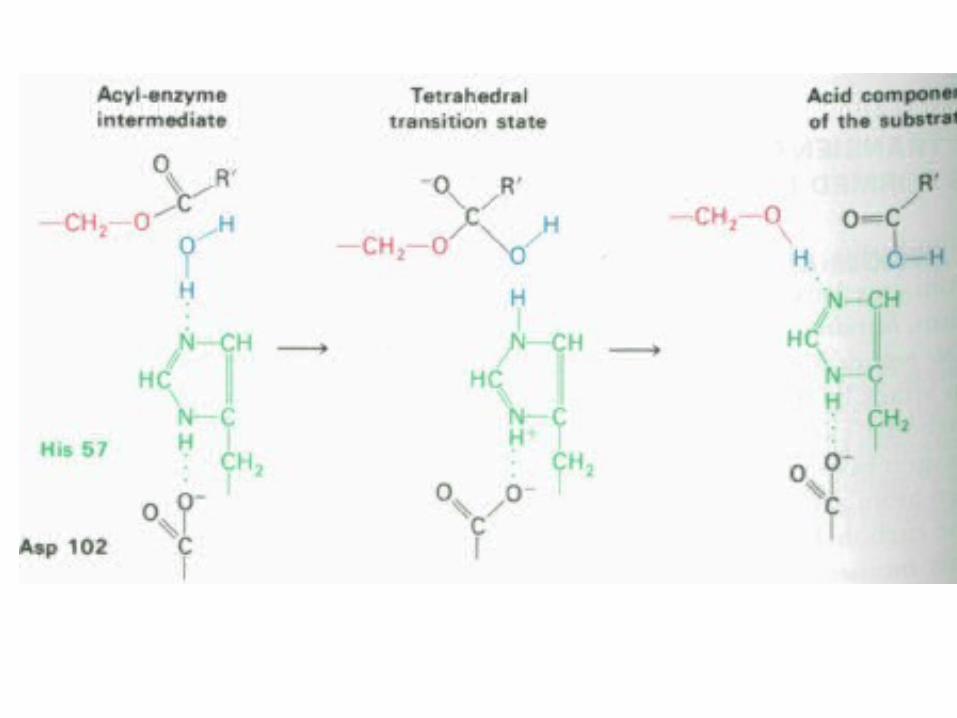
Centrului activ al chimotripsinei
• His - 57 din lanţul B;
• Ser - 195 din lanţul C; • Asp - 102 din lanţul B.
A B C
S S S S-chimotripsina contine 5 legãturi disulfurice
S S S S S S
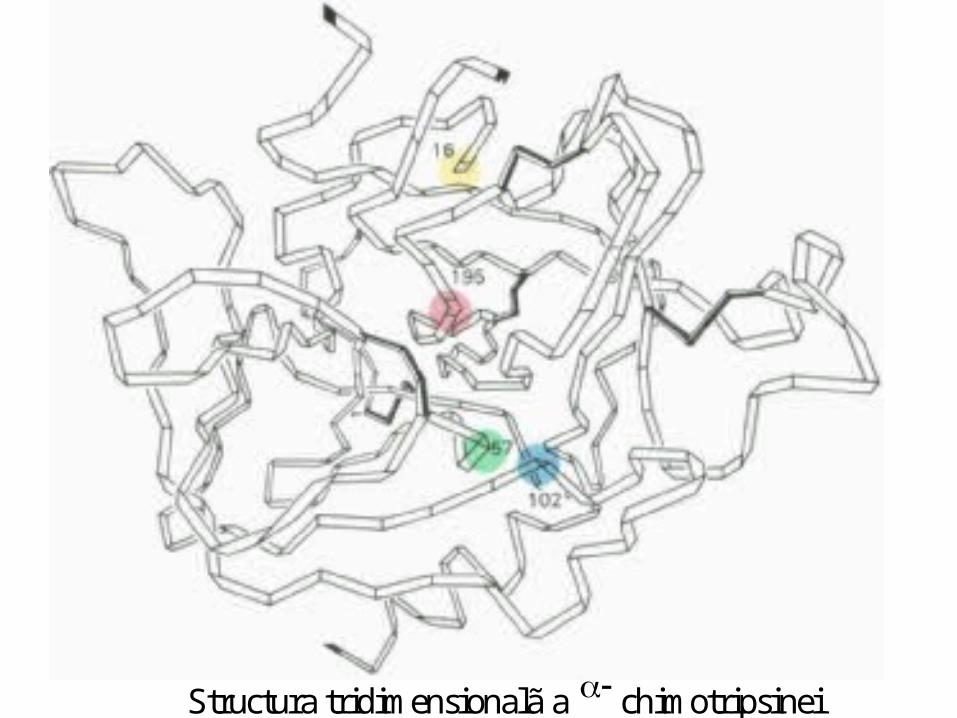
Structura tridimensionalã a chimotripsinei

Triada Asp-His-Ser

• Centrul activ
conţine resturi aa
ce crează o entitate tridimensională
complementară cu substratul


• Inhibitorii interferă cu proteazele utilizate de HIV pentru transformarea proteinelor precursor (gag, pol, env) în proteine active
• Inhibitorul proteazelor blochează centrul activ al enzimelor

• Fatty Acid Desaturases - Integral Membrane Metalloenzymes
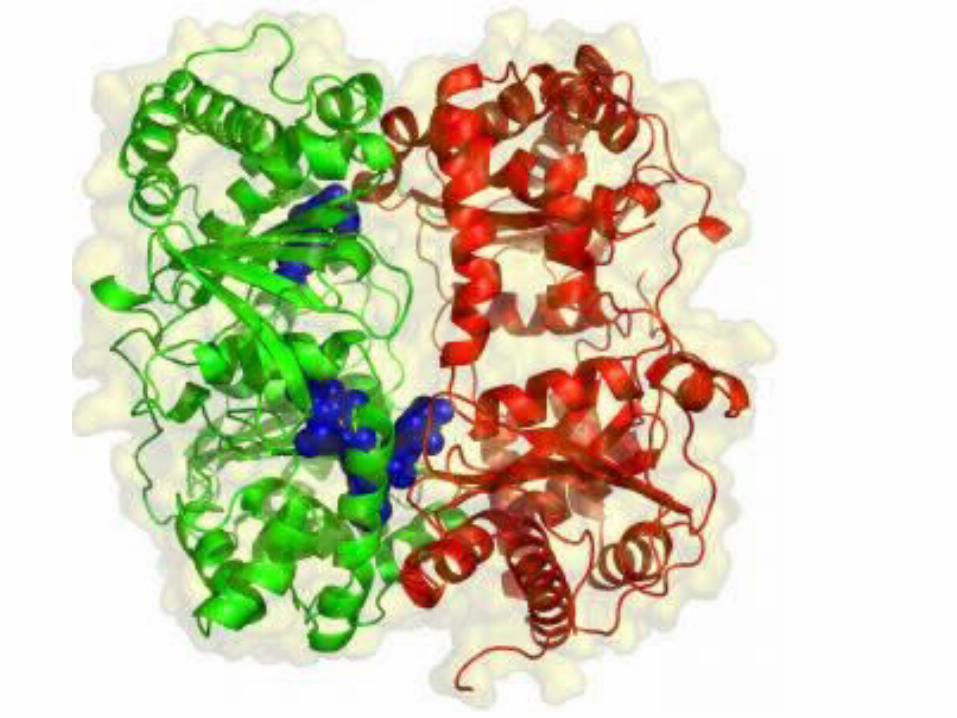

• ScienceDaily (Oct. 16, 2006) — Univ Chicago
• S-a identificat structura tridimensională a enzimei de degradare a insulinei, care degradează şi amiloidul implicat în boala Alzheimer.
• Enzima este o ţintă promiţătoare
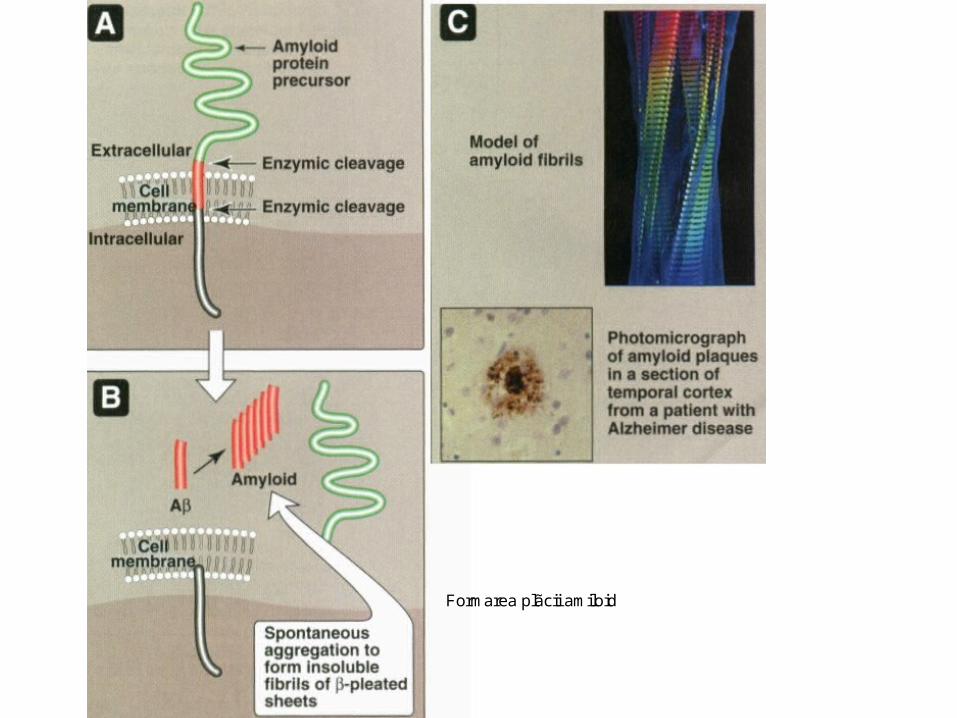
Formarea plãcii amiloid

Centrul activ al enzimelor
• centru de legare prezintă resturi aminoacidice care asigură legarea substratului
• centru catalitic prezintă resturi aa responsabile de catalizarea propriu-zisă
• Resturile de aa din situsurile catalitice ale enzimelor sunt: Cys, Ser, His, Tyr, acid Glu şi Asp, Lys cu grupările funcţionale:
OH, SH, NH3+, COO-
HN+
NH
imidazol



Complexul enzimă – substrat (ES)
• structuri labile, cu o viaţă foarte scurtă • între E şi S se stabilesc:
- forţe necovalente (punţi de hidrogen, interacţiuni hidrofobe, interacţiuni electrostatice) sau
-covalente reversibile
• Complementaritatea chimică şi geometrică determină legarea E-S

Modelul clasic (lacăt – cheie)
• al CA al enzimei în raport cu structura substratului, propus de Emil-Fischer
• Este un model rigid.
Modelul lacãt cheie
SubstratEnzimã Complexul ES

Modelul dinamic „centrul indus“sau „complementaritate indusă“,
• produs de Koshland, este acela de mâna în mănuşă
• Modelul indus presupune o flexibilitate a zonei în care se află centrul activ
Modelul centrului indusSubstrat Enzimã Complexul ES



Cum acţionează enzimele ?
• Mecanismul de acţiune privit din 2 perspective:
– modificarea energiei în timpul reacţiei;
– rolul centrului activ în realizarea catalizei din punct
de vedere chimic

• Ce este energia liberă de activare ?

• Toate reacţiile chimice sunt însoţite de variaţii
ale energiei
• In organismele vii temperatura şi presiunea ==
constante
• Energia liberă de reacţie G caracterizează reacţiile
din lumea vie

• Reacţiile posibile termodinamic decurg cu
scăderea energiei libere a reactanţilor
Gproduşi –
Greactanţi <0
– reacţie „exergonică“, G<0
– reacţie „endergonică“, G>0

Cum se modifică energia în timpul reacţiei ?
• Virtual, toate reacţiile chimice au o barieră energetică ce separă
rectanţii de produşi
• Această barieră numită energie liberă de activare este
diferenţa dintre energia reactanţilor şi energia intermediarului
care se produce în timpul formării produsului.
BTA
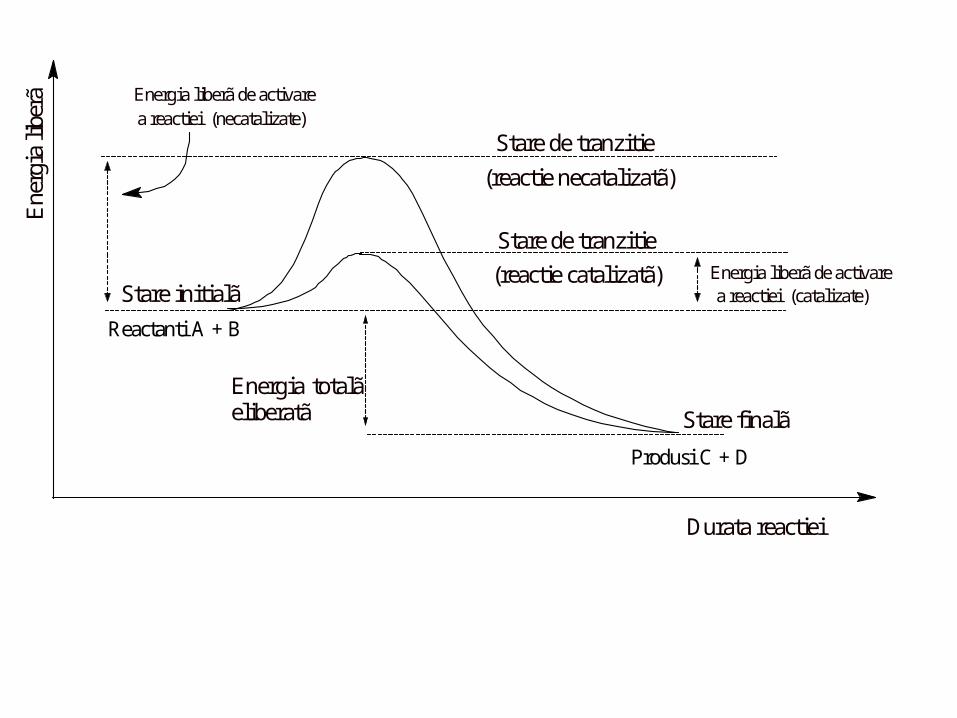
Durata reactiei
Stare de tranzitie
Ene
rgia
libe
rã
(reactie necatalizatã)
Stare de tranzitie
(reactie catalizatã)Stare initialã
Energia liberã de activarea reactiei (necatalizate)
Energia liberã de activarea reactiei (catalizate)
Stare finalã
Energia totalã eliberatã
Produsi C + D
Reactanti A + B

• Enzima nu modifică energia liberă a reacţiei
catalizate (G0), a reactanţilor sau produşilor, şi deci
nu modifică echilibrul recţiei (constanta de echilibru)
• Enzima furnizează o cale de reacţie alternativă, prin
micşorarea energiei libere de activare (ca urmare
a micşorării entropiei substratului)

Formarea stãrii de tranzitie

Mecanisme chimice folosite de centrul
activ pentru a transforma
Substrat Produşi

• Centrul activ al E stabilizează starea T* a
substratului

Cataliza covalentă
• Enzime care formează intermediari covalenţi enzimă-substrat: • Clasa serinei• Tripsina, Chimotripsina, Elastaza, Acetilcolinesteraza
• Clasa cisteinei• Gliceraldehid 3P-dehidrogenaza, Papaina
• Clasa histidinei• Glucozo 6-fosfataza, Succinil~CoA sintetaza • Clasa lizinei• D-aminoacid oxidaza, Transaldolaza

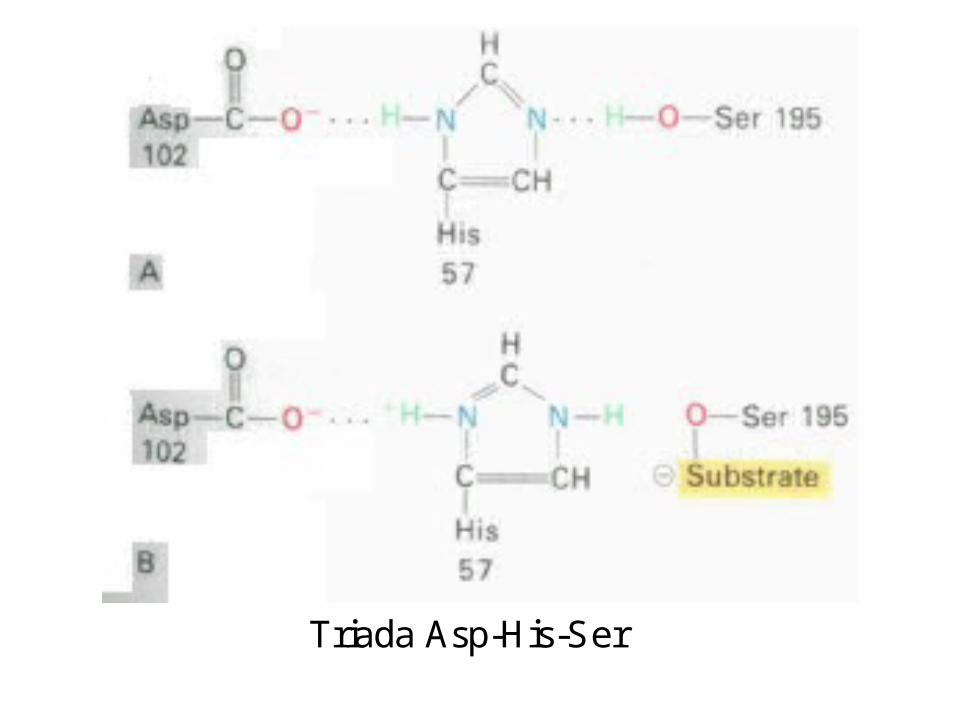
Triada Asp-His-Ser



Cataliza acido-bazică
• Enzimele conţin grupări funcţionale care pot acţiona ca donori sau ca acceptori de protoni: amino, carboxil, sulfhidril, imidazol, hidroxilul fenolic.
• hidroliza esterilor carboxilici şi fosforici, reacţii de izomerizare

Glucozo-6-fosfat izomeraza

Există un factor de tensiune în cataliza
enzimatică ?

• legarea S la E poate deforma atât enzima cât şi substratul
• centrul activ al unor enzime este relativ nepolar


Care dintre următoarele afirmaţii referitoare la
enzime sunt adevărate:
A.Scad G0 (energia liberă) a reacţiei catalizate
B.Scad energia de activare a procesului catalizat
C.Deplasează echilibrul reacţiei catalizate în sensul formării
produşilor
D.Catalizează ruperea şi/sau formarea unor legături
covalente
E.Cresc viteza de apariţie a produşilor de reacţie

• Enzimele:
A.Sunt polizaharide naturale
B.Au structură asemănătoare vitaminelor
C.Pot cuprinde în structura lor toţi aminoacizii naturali
D.Pot fi reprezentate de anumite molecule de acizi nucleici
E.Pot fi localizate în membranele celulare deoarece au
structură lipidică


Cursul 6 18.11.2009
Cinetica enzimatică
• Factori care influenţează activitatea enzimatică
• Teoria cinetică Michaelis-Menten• Ecuaţia Lineweaver Burk• Inhibiţia activităţii enzimelor• Enzime alosterice

Cinetica enzimatică
• Cinetica studiază reacţiile chimice:
–desfăşurarea în timp a reacţiei;
–factori care influenţează viteza reacţiilor;
–etapele intermediare prin care trec
reactanţii până devin
produşi.

• Cum caracterizăm cinetic o reacţie
chimică ?

• Prin viteza de reacţie

• Viteza unei reacţii = variaţia în timp a
concentraţiei reactanţilor sau a produşilor de reacţie.
• A B
• ecuaţii cinetice
dt
]A[dv
dt
]B[dv

Ce postulează Legea cinetică ?
☻ Viteza de reacţie este proporţională cu
produsul concentraţiilor reactanţilor

• Pentru o reacţie:
A Produşi V = K [A]
Pentru o reacţie:
A + B Produşi V = K [A][B]
K = constanta de viteză sau viteza specifică(V de reacţie la concentraţii 1M ale reactanţilor)

Activitatea enzimatică
• U.I. (unitatea internaţională) = activitatea enzimei care transformă 1 mol substrat/minut în cond standard de pH, temperatură, cofactori
• Activitatea moleculară = nr. molecule de S transformate în produs de către o moleculă de E /secundă.
• Activitatea specifică = nr. de unităţi de activitate enzimatică / mg de proteină totală.
• Constanta catalitică (turn-over number) = nr. de transformări S P catalizate de fiecare CA al E în unitatea de timp.

Factorii care influenţează activitatea enzimatică
• Temperatura;
• pH;
• [E];
• [S];
• prezenţa inhibitorilor

Temperatura
• T optimă = temp. la care activitatea enzimelor este maximă
50
100
20 40 60temperatura
activitatea enzimaticã
%
Toptimã

pH• pH-uri extreme denaturează enzimele• Variaţii mici de pH în zona de stabilitate a enzimei
modifică v.
Vite
za d
e r
ea
ctie
(v)
Pepsina Fosfataza alcalinã
amilaza salivara

[E]
• [S] = constantă, v este proporţională cu cantitatea de enzimă
activitatea enzimaticã
%
concentratia enzimei

Ecuaţia Michaelis Menten
• V = f([s]) la [E], temp., pH = constante
• Michaelis şi Menten au introdus
parametrii cinetici KM şi Vmax
activitatea enzimaticã
concentratia substratului
vmax
vmax
2
KM
saturatie
E + SK1
K2
ES E + ProdusK3

Unde E, ES şi S sunt în echilibru
[Etotal]=[ES]
Vmax=K3[Etotal]
• La echilibru• v1 = v2 + v3
• K1[E][S] = K2[ES] + K3[ES]• K1[E][S] = [ES](K2 + K3)
E + SK1
K2
ES E + ProdusK3
Etapa rapidã Etapa lentã

KM
• KM este o măsură a afinităţii dintre E şi S
• afinităţile enzimelor pentru substraturile lor
sunt cu atât mai mari
cu cât valorile KM sunt mai mici
M1
32 KK
KK
]ES[
]S][E[

Ecuaţia Michaelis - Menten
• Ecuaţia de viteză a reacţiilor enzimatice cu un singur
substrat sau ecuaţia lui Michaelis-Menten
• reprezentare grafică v = f([S]) este un arc de hiperbolă.
][
].[max
SK
SVv
M

activitatea enzimaticã
concentratia substratului
vmax
vmax
2
KM
saturatie

• KM este numeric egală cu [S]
la care viteza reacţiei este semimaximă
]S[]S[K
]S[V
2
V
M
maxmax
MK

Ecuaţia Lineweaver – Burk
• Ecuaţia Michaelis-Menten inversată este ecuaţia unei drepte.
maxmax V
1
][
11
SV
K
VM
][
].[max
SK
SVv
M

- 1/KM 0 1/[S]
1/V
1/Vmax
Panta = KM/Vmax

• Inhibiţia activităţii enzimelor

Inhibiţia activităţii enzimelor
• scăderea activităţii catalitice a enzimei
în prezenţa unui compus chimic denumit
inhibitor
Inhibitorii pot fi:
– endogeni (metaboliţi)
– exogeni (substanţe toxice,medicamente)

Inhibiţia poate fi:
- ireversibilă
- reversibilă

Inhibiţia ireversibilă
• legături covalente între E şi I
• Când [I] = [E]
activitatea catalitică a enzimei
este redusă la zero.
E + I EI
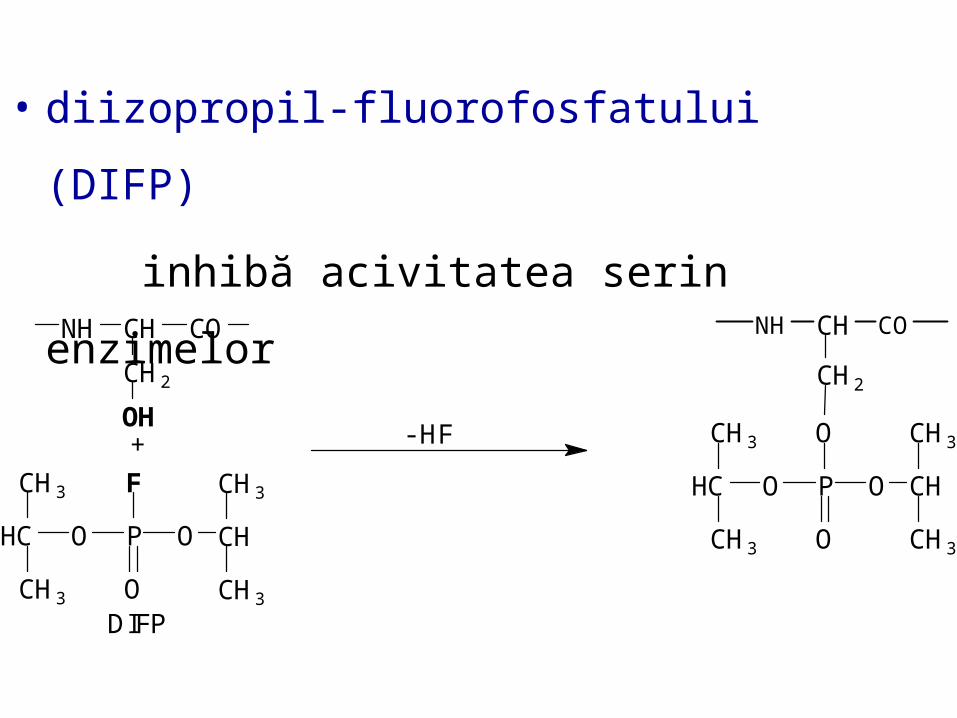
• diizopropil-fluorofosfatului (DIFP)
inhibă acivitatea serin enzimelor
NH CH CO
CH2
OH+
P
F
O
OOHC
CH3
CH3
CH
CH3
CH3
DIFP
- HF
NH CH CO
CH2
O
P
O
OOHC
CH3
CH3
CH
CH3
CH3

☺acetilcolinesteraza este foarte sensibilă
la acţiunea
DIFP
DIFP este utilizat ca paralizant


• Hemoproteinele (citocromi, hemoglobina)
sunt inactivate de către:
CO -CN
care se leagă de ionul de Fe2+ din
hem

Inhibiţia reversibilă
• Ce efecte are Ic
asupra celor doi parametrii cinetici
vmax şi KM ?
E + I EI

• Inhibiţia reversibilă poate fi:
– Competitivă
– Necompetitivă
– Uncompetitivă

Inhibiţia reversibilă competitivă
• Ic este analog structural al S
• se leagă în centrul activ al enzimei
prin aceleaşi grupări ca S;

• Într-un sistem cu: E, S şi Ic au loc reacţiile:
E + S ES E + Produsi+I
EI

1/V
1/Vmax
activitatea enzimaticã
concentratia substratului
vmax
vmax
2
KM
saturatie
- 1/KM`
KM`
+ Ic
+ Ic
- 1/KM1/[S]0
• reacţia enzimatică atinge Vmax
dar la concentraţii mari de S.
• aparent creste KM,
deci afinitatea enzimei pentru S scade şi
nu se modifică Vmax;

Ic (compuşi naturali sau de sinteză)
• Acizii dicarboxilici: oxalic, malonic, malic, oxaloacetic, pirofosfat sunt inhibitori competitivi ai succinat dehidrogenazei (SD).
FAD FADH2
CH2
CH2
COOH
COOH
Acid succinic
COOH
C
C
HOOC
H
H
Acid fumaric
CH2
COOH
Succinat dehidrogenazã


Sulfanilamida
• Folat sintetaza, la bacterii este păcălită şi sintetizează un intermediar cu sulfanilamidă în locul acidului para-amino-benzoic care nu se poate transforma în acid folic.
• Cum acidul folic este indispensabil bacteriilor, acestea mor.
NH2
SO2NH2
NH2
COOHSulfanilamida Acid p-aminobenzoic

N
NN
N
H2N
OHCH2 NH CO NH
COOH
CH2
CH2
CH
COOH9 10
5
pterina acid p-aminobenzoic
acid glutamicacid pteroic

Analogi structurali ai bazelor purinice şi pirimidinice
• se utilizează în chimioterapia cancerului.
• inhibă sinteza de acizi nucleici, împiedicând diviziunea celulară care este mult mai rapidă în tumori
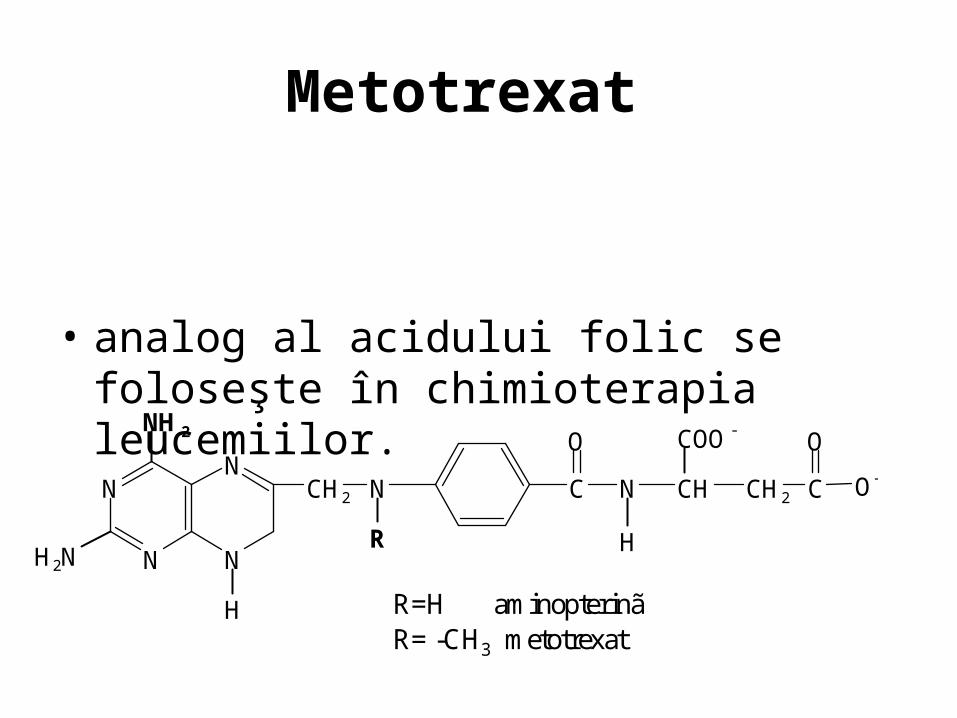
Metotrexat
• analog al acidului folic se foloseşte în chimioterapia leucemiilor.
NCH2
N
N
N
N C
H2N
NH2
H
R
N CH CH2 C O-
O
H
COO- O
R=H aminopterinã R= -CH3 metotrexat

Inhibiţia reversibilă necompetitivă
• Substratul şi inhibitorul necompetitiv (In)
– nu sunt analogi structurali
– nu au dimensiuni comparabile
– In nu se leagă în centrul activ

E + S ES E + Produsi+I
EI + S ESI
+I

• Se modifică Vmax
• KM nu se modifică în prezenţa inhibitorului
1/v
1/vmax
activitatea enzimaticã
concentratia substratului
vmax
KM
+ Ic
+ Ic
- 1/KM 1/[S]0
vmax / 2
v`max / 2
v`max 1/v`max

• grupări - SH libere (nu din centrul activ), pot lega slab:– Ag+
– Hg2+
– Pb care micşorează sau chiar anulează activitatea enzimelor
aşa se explică efectul otrăvitor al unor metale grele
• Pb inhibă ferochelataza
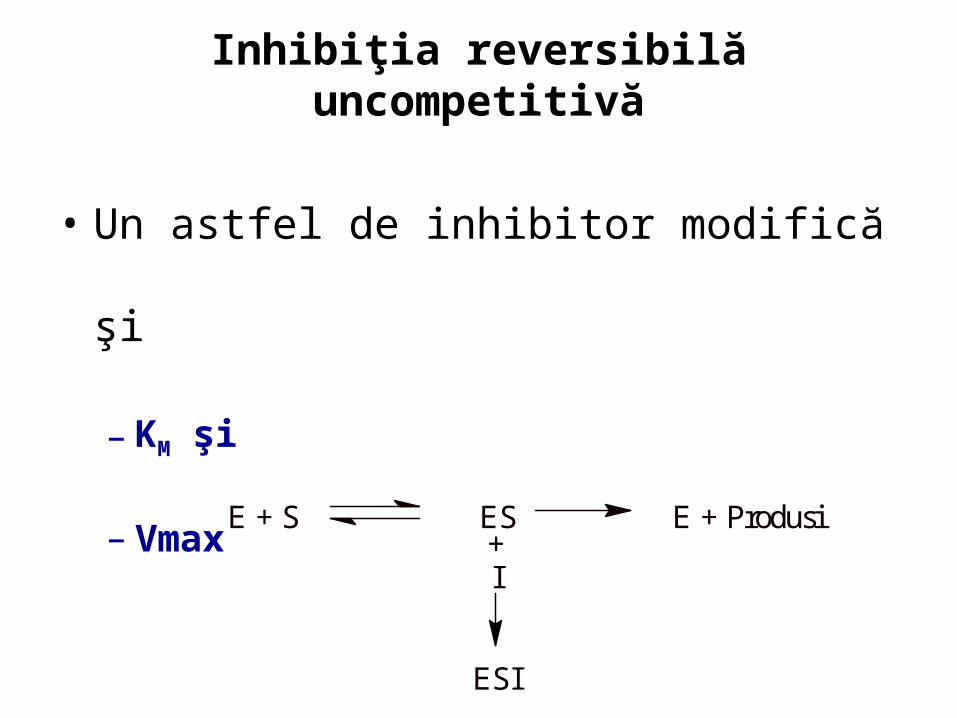
Inhibiţia reversibilă uncompetitivă
• Un astfel de inhibitor modifică şi
– KM şi
– Vmax
E + S ES E + Produsi+I
ESI

Enzime alosterice
• Sunt proteine oligomere (dimeri, tetrameri,
etc.)
• Au mai multe centre de legare pentru S
(câte un centru pentru fiecare subunitate).

• Curbele v = f([S]) sunt sigmoide
• Vmax se atinge atunci când toate centrele
active sunt ocupate cu S

• Parametrii cinetici pentru enzimele oligomere
sunt:
• -Vmax şi
-S0,5(S50) concentraţia substratului pentru
care reacţia are o viteză semimaximă

activitatea enzimaticã
concentratia substratului
vmax
KM
vmax / 2
• Aspectul sigmoid al curbei v = f([S]) reflectă o
modificare a afinităţii enzimei pentru substrat

• Enzimele care dau curbe v=f([s])
sigmoide sunt denumite
alosterice
• Liganzii fixaţi pot fi:– identici (de ex. Substraturi în ambele centre)
sau – diferiţi

• centru izosteric leagă substratul
• centru alosteric (allos=altul) (poate fi situat pe acelaşi lanţ polipeptidic dar în regiune
diferită)
leagă un efector alosteric

• Activatori alosterici
• Inhibitori alosterici
activitatea enzimaticã
vmax
KM
50
%100
1 23
[S]

• După natura centrelor de legare şi a liganzilor pentru aceste centre, efectele alosterice sunt:
• –Efecte homotrope (presupun cooperarea între două centre izosterice
• –Efecte heterotrope (cooperare între un centru izosteric şi un centru alosteric)

• Alosteria
capacitatea unor liganzi de a modifica funcţiile
unor proteine
prin
inducerea unor tranziţii alosterice (modificări
conformaţionale).

Caracteristici esenţiale pentru Enzimele alosterice:
-catalizează etape cheie din căile metabolice;
ireversibile, cu Go < 0
-au structură cuaternară

• Două Modele explică
mecanismul de acţiune
al Enzimelor Alosterice
1.modelul concertat (simetric)
2.modelul secvenţial

• Elemente comune ale modelelor
Fiecare monomer al oligomerului are centrul său activ
Monomerii cooperează în fixarea substratului :
–homotrop
–heterotrop
• Reglarea alosterică se face cu consum minim de energie

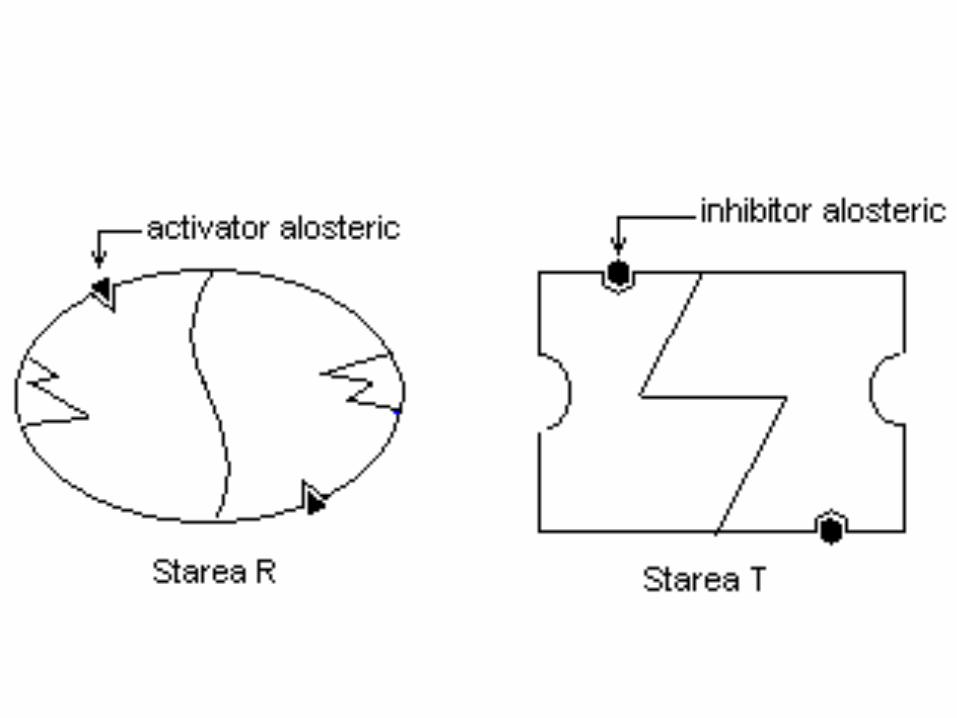
Modelul simetric
Subunităţile dimerului, pot avea două conformaţii;
-R (relaxată) cu afinitate mare pentru substrat
-T (tensionată) cu afinitate mică pentru substrat.
• Modelul impune simetria dimerului, care există numai în forma R sau T.


• La enzimele heterotrope:
• Activatorii stabilizează starea R
• Efectorii negativi stabilizează starea T
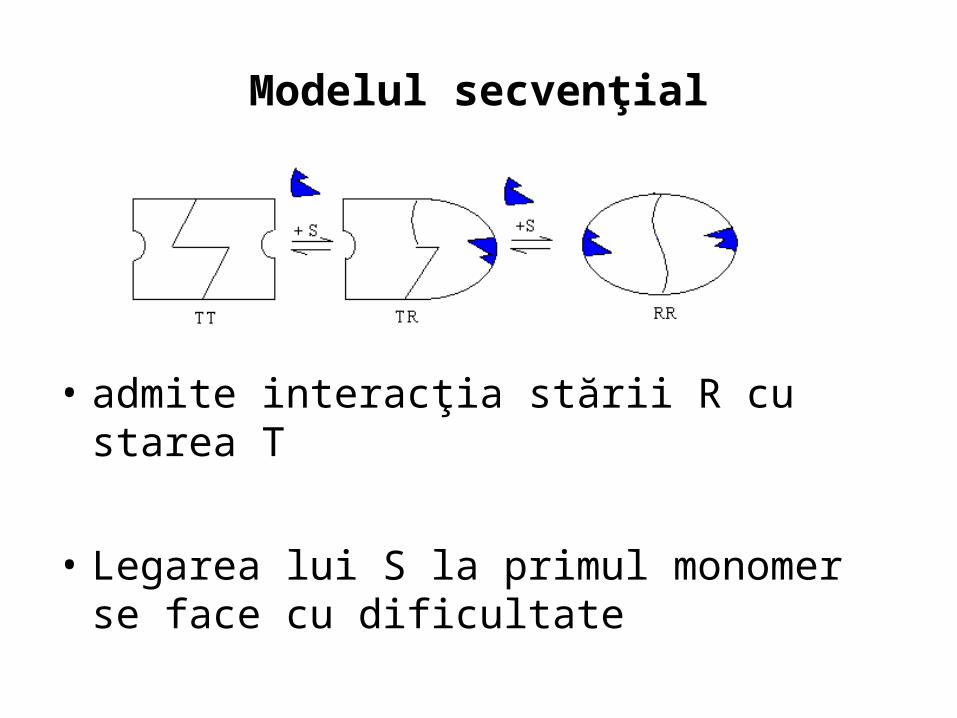
Modelul secvenţial
• admite interacţia stării R cu starea T
• Legarea lui S la primul monomer se face cu dificultate

• Ce tip de cooperare am întâlnit la
realizarea rolurilor Hemoglobinei ????

Hemoglobina fixează
• O2 prin cooperativitate (homotropă)
• coordonată de factorii: H+; CO2; 2,3-DPG (heterotropă)


Curs 7
• Reglarea cantitativă a enzimelor
• Reglarea eficienţei catalitice
• Complexe multienzimatice
• Antienzime
• Izoenzime
• Denumirea enzimelor
• Clasificarea enzimelor

Reglarea activităţii enzimelor
• Viteza reacţiilor enzimatice cheie din căile metabolice depinde de:
• –cantitatea de enzimă;
• –eficienţa enzimelor

Reglarea cantităţii de enzimă
• prin dinamica proceselor de biosinteză şi de degradare
•
Aminoacizi ProteineKs
Kd

Enzimele constitutive: Ks = Kd
• Catalizează procese fundamentale pentru existenţa celulei.
• Se găsesc în celule în cantităţi relativ constante.
• Ele nu suferă fluctuaţii mari în raport cu factorii de mediu.

• Enzimele inductibile: Ks > Kd
• Sunt implicate în căi catabolice.
• se sintetizată numai în prezenţa substratului
• Substratul acţionează ca un inductor, accelerând sinteza enzimelor.

• Enzimele represibile: Ks < Kd
• Sunt implicate în procese anabolice
• Sinteza lor este încetinită de produsul final al reacţiei catalizate;
• Represia se face de obicei, prin intermediul
complexelor corepresori - aporepresori.

• Controlul degradării proteinelor (enzimelor)
-se face independent de controlul sintezei;
-se face prin intermediul ubiquitinei;
-este proces dependent de ATP.

Reglarea eficienţei catalitice a enzimelor
Factorii ?????

• Activitatea unor Enzime alosterice depinde de concentraţia substratului:
• Reglarea printr-un intermediar (feed forward stimulare):
A B C P E1 E2 E3
A B C D Produsi E1 E2 E3

Glucozã
Glucozo-6-fosfat
Fructozo-6-fosfat
Fructozo-1,6-bisfosfat
Gliceraldehid-3-fosfat Dihidroxiaceton-fosfat
Etapa hexozelor
ATP
ADP1
2
3
4
ATP
ADP
1,3-Bisfosfoglicerat
NAD+
NADH + H+
H3PO4
3-Fosfoglicerat
5
6ADP
ATPFosforilare la nivel de substrat
2-Fosfoglicerat
7
8H2O
FosfoenolpiruvatADP
ATP9 Fosforilare la nivel de substrat
Piruvat
Etapa triozelor

activitatea enzimaticã
concentratia substratului
vmax
KM
vmax / 2

• Hexokinaza: -prezentă în toate ţesuturile extrahepatice,-are afinitate mare pentru glucoza (KM = 0,15 mM) -ţesuturile preiau glucoza din sânge la orice valoare a glicemiei
• Glucokinaza: -prezentă numai în ficat -are afinitate mică pentru glucoza (KM = 10 mM) - este activă după ingestia de glucide
Glucozã + ATP Glucozo-6-P + ADP

Glucozo-6-fosfat
CH=O
C
C
C
C
CH2 OPO3H2
H OH
OH H
H OH
H OH
CH=O
C
C
C
C
CH2 OH
H OH
OH H
H OH
H OH
Glucozã
ATP ADP
Hexokinaza Mg2+ Glucozo-6-fosfat
izomeraza
CH2
C
C
C
C
CH2 OPO3H2
O
OH H
H OH
H OH
OH
Fructozo-6-fosfat

• Reglarea activităţii enzimelor alosterice prin produsul final (inhibiţie de tip feed back)
A B C D Produsi E1 E2 E3
A B C E1 E2
D P1
E P2
E3
E4

Reglarea covalentă
• Apare la organismele superioare
şi presupune activare prin
ruperea unor legături covalente

☺ enzime interconvertibile
Conversia are loc prin:
–fosforilare-defosforilare;
–nucleotidilare

☺activare prin proteoliză
proces unidirecţional

Enzime interconvertibile prin fosforilare <==> defosforilare
• Fosforilările:-sunt catalizate de kinaze (reglate hormonal, interconvertibile fosfo-defosfo);-sunt ireversibile;-necesită ATP
• Defosforilările: -sunt ireversibile;-se fac hidrolitic în prezenţa unor fosfataze; -fosfatazele sunt supuse unui control hormonal.

Enzimã (Ser-OH) Enzimã (Ser-OP)forma defosfo forma fosfo
ATP ADP
Pi H2O
proteinkinaze
fosfataze

• enzime active în forma fosforilată
glicogen fosforilaza implicată în catabolismul
glicogenului
• Enzime active în forma defosfo
glicogen sintaza implicată în proces anabolic
• activarea este implicată într-o cascadă de evenimente
declanşate de legarea unui hormon la o membrană celulară

Reglarea covalentă prin proteoliză
• Proenzimele (zimogenii)
– sunt produse la locul de sinteză în formă inactivă
– activarea se face la locul de acţiune.

• Activarea se face
– sub influenţa factorilor de mediu şi
– Autocatalitic• printr-un proces ireversibil de rupere a unor legături
peptidice specifice.

De ce este necesară existenţa zimogenilor ?
se obţin foarte rapid
• prin transformări minore
– cantităţi mari de enzime active.

• Enzimele proteolitice digestive
–secretate de stomac: pepsinogenul
–secretate de pancreas: tripsinogenul, chimotripsinogenul, proelastaza, procarboxipeptidaza
• Enzimele coagulării sângelui (secretate de ficat în formă de proenzime)
• Enzimele sistemului complement

• a.Enzimele proteolitice digestive hidrolizează proteine:
• Transformarea
HCl
• pepsinogen pepsina
(- segment terminal (44 aminoacizi) cu caracter bazic)

• Transformarea Tripsinogen tripsină enteropeptidază
(denumire veche enterokinază)
(fragment N-terminal de 6 aminoacizi din tripsinogen)
Tripsina activează tripsinogenul, chimotripsinogenul, proelastaza, şi procarboxipeptidaza.
Zimogenii pancreatici se activează când conţinutul stomacal a ajuns în duoden.

• Transformarea
chimotripsinogen chimotripsină
este activată de tripsină
se face prin îndepărtarea a două dipeptide
• Chimotripsina cuprinde trei lanţuri peptidice şi cinci punţi de sulf.
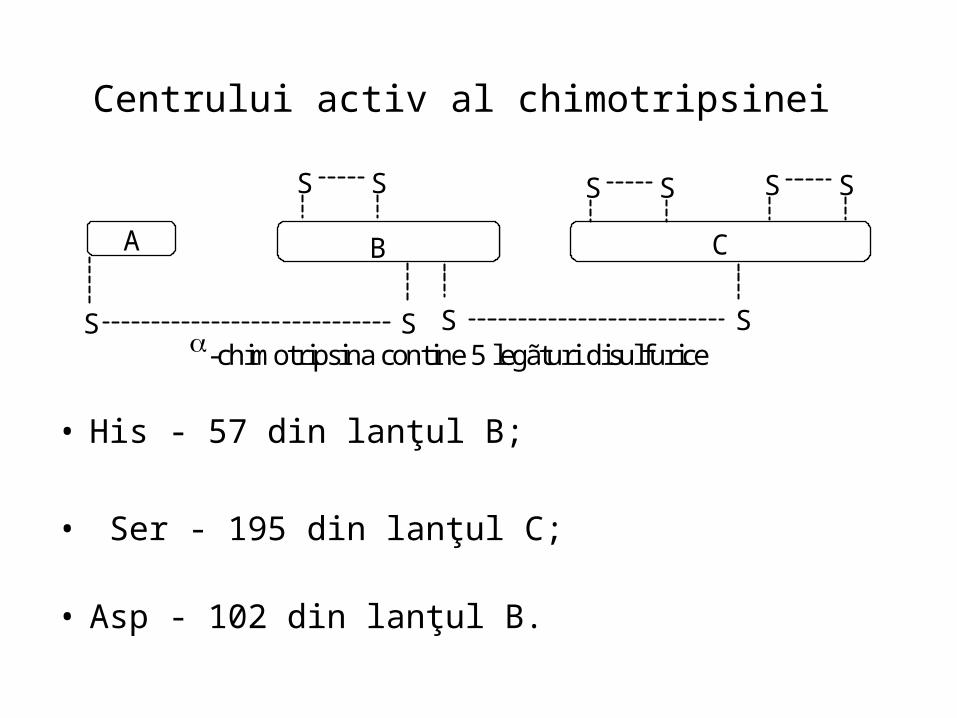
Centrului activ al chimotripsinei
• His - 57 din lanţul B;
• Ser - 195 din lanţul C; • Asp - 102 din lanţul B.
A B C
S S S S-chimotripsina contine 5 legãturi disulfurice
S S S S S S

Structura tridimensionalã a chimotripsinei

Enteropeptidazã
Tripsinogen
Tripsinã
Chimotripsinogen
Chimotripsinã
Procarboxipeptidaza A
Carboxipeptidaza A
Carboxipeptidaza B
Procarboxipeptidaza B
In intestin

• Enzimele coagulării şi fibrinolizei.
• Există două căi iniţiate de activatori diferiţi care converg într-o cale finală comună
–calea intrinsecă, cu participare de factori sanguini;
–calea extrinsecă, la care participă şi factori de origine exclusiv tisulară.

Iniţierea procesului de coagulare este urmată de activarea în cascadă a factorilor de coagulare
Care sunt avantajele unui răspuns reglator în cascadă ?
-o cantitate foarte mică de iniţiator declanşază un răspuns amplu.
-fiecare etapă a cascadei amplifică răspunsul

• Fibrinoliza. Componenţii necesari dezagregării chiagului se găsesc în sânge în formă inactivă şi se activează proteolitic
Plasminogen Plasminã
Cheag de fibrinã Peptide solubile
Factor tisular

Implică o modificare covalentă a enzimei:
A.modularea alosterică
B.inhibiţia competitivă
C.asocierea unei enzime cu un cofactor
D.inhibiţia necompetitivă
E.reacţiile de fosforilare în prezenţa kinazelor
specifice

Care dintre următoarele enzime proteolitice este activată prin hidroliza acidă a proenzimei inactive?
• a. tripsina
• b. chimotripsina
• c. elastaza
• d. pepsina
• e. carboxipeptidaza

Care dintre următoarele afirmaţii descriu corect enzimele alosterice?
a. efectorii pot stimula sau inhiba legarea substratului
b. activitatea lor nu este inhibată prin feed back
c. curbele v=f([S]) sunt sigmoide
d. difuzează rapid prin membranele celulare
e. sunt, de obicei, plasate la începuturile căilor metabolice

Antienzimele

• Antienzime (antiproteaze) = proteine reglatoare
Serpinele sunt inhibitori de serin proteaze
Se leagă necovalent în centrul activ al serin proteazelor:

• antitrombina III

1 antiproteaza
(denumită şi 1 antitripsina)

• inhibitorul pancreatic al tripsinei
• inhibitorul proteinei C (proteina C este o serin protează plasmatică dependentă de vitamina K, activată de trombină, care transformă factorii activi VIIIa şi Va ai coagulării în forme inactive).

Antitrombina III:
-prezentă în plasmă;
-inactivează trombina şi alţi factori ai coagulării: IXa, Xa, XIa
Cine măreşte viteza de formare a complexelor ireversibile
între antitrombina III şi serin proteazele coagulării ?
Heparina

1-antiproteaza:
-proteină plasmatică;
-este 1-globulină
inhibă elastaza, tripsina şi alte proteaze

1-antiproteaza sintetizată
• de ficat
• de monocitele şi macrofagele alveolare
– are rol în protejarea ţesuturilor de acţiunea elastazei
☺ Elastaza = enzimă proteolitică distructivă, secretată de
neutrofilele activate poate distruge elastina din pereţii
alveolelor pulmonare

• O deficienţă la nivelul 1-antiproteazei poate
duce la instalarea emfizemului pulmonar

• Inhibitorul pancreatic al tripsinei:
-este o proteină cu masa mică, produsă de pancreas.
-se leagă de tripsină, prevenind distrugerea pancreasului.

Izoenzimele
• Izoenzimele:
- proteine oligomere
-catalizează aceeaşi reacţie în toate ţesuturile;
-diferă prin:
☺structură
☺distribuţie tisulară.

• Diferenţele structurale pot influenta proprietăţile fizico-chimice:
• –pH izoelectric
• –mobilitate electroforetică
• –masa moleculară

• diferenţiere prin însuşirile catalitice:
– afinităţi diferite faţă de acelaşi substrat;
– sensibilităţi diferite la acţiunea unor efectrori
alosterici;
_ specificităţi distincte pentru coenzime
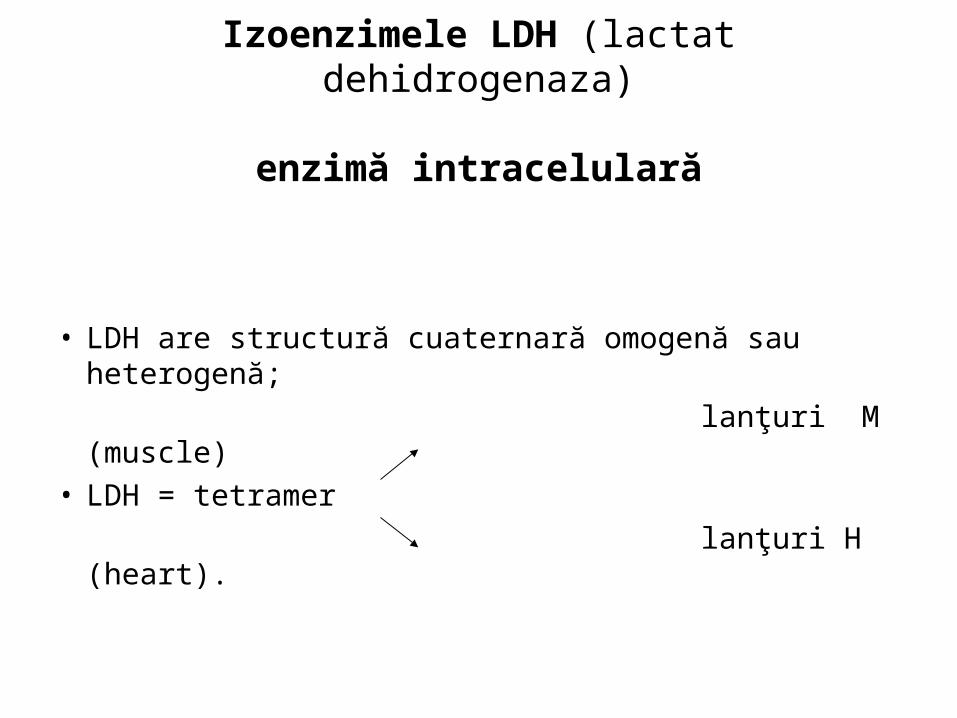
Izoenzimele LDH (lactat dehidrogenaza)
enzimă intracelulară
• LDH are structură cuaternară omogenă sau heterogenă;
lanţuri M (muscle)• LDH = tetramer
lanţuri H (heart).

• LDH are cinci izoenzime:
H4, H3M, H2M2, HM3, M4
sau
după mobilitatea electroforetică
LDH1 LDH5
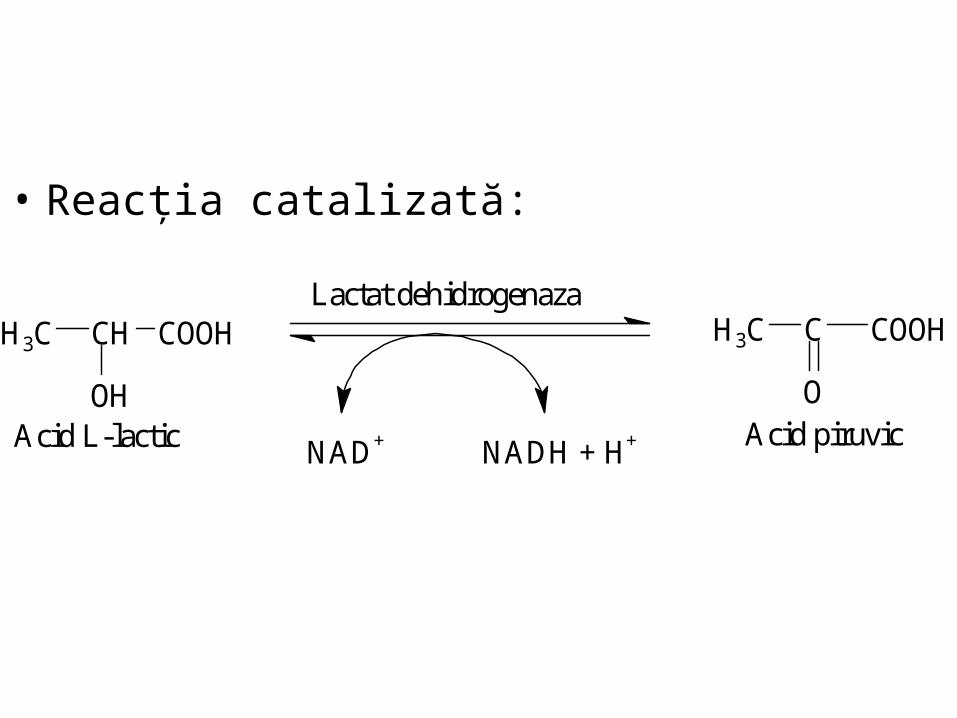
• Reacţia catalizată:
Lactat dehidrogenaza
NAD+ NADH + H+Acid L-lactic
COOHCH3C
OAcid piruvic
COOHCHH3C
OH

Izoenzimele LDH au afinităţi diferite pentru piruvat (sau lactat):
• H4 şi H3M inimă
afinitate mică pentru piruvat
(îl transformă greu în lactat)
• M4 şi M3H
muşchi scheletici şi ficat
afinitate mare pentru piruvat

Determinarea activităţii enzimelor serice
poate fi folosită în clinică pentru stabilirea
originii ţesutului lezat

Izoenzimele creatin kinazei (creatin fosfokinaza)
• Creatin kinaza:-enzimă intracelulară;-dimer format din două subunităţi:
☼ B (de la brain)☼ M (de la muscle)
• Are trei izoenzime:–MM (prezentă în muşchi şi miocard)–BB (prezentă în creier şi muşchi)–MB (prezentă în urme numai în miocard).

• reacţia catalizată este reversibilă:
Creatină + ATP Fosfo-creatină + ADP
• Creşterea activităţii creatin kinazei-MB în ser, indică lezarea miocardului
(infarct de miocard)
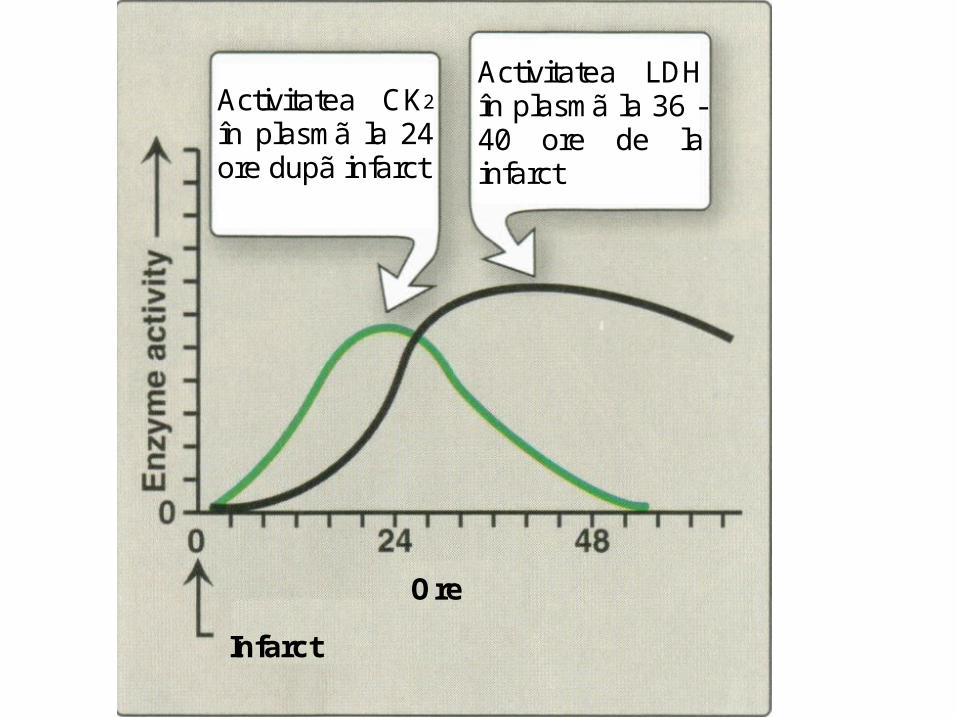
Infarct
Ore
Activitatea CK2 în plasmã la 24 ore dupã infarct
Activitatea LDH în plasmã la 36 - 40 ore de la infarct


Complexe multienzimatice
• Asamblări de proteine într-un complex, cu scopul de a creşte eficienţa globală a procesului.
• Rolul lor:• –cataliza coordonată a unei succesiuni de
reacţii;• –minimalizarea unor reacţii secundare.
• Exempl: acid gras sintază, complexul piruvat dehidrogenazei, etc.

Clasificarea şi denumirea enzimelor
• Denumirea enzimelor• se pot utilza trei modalităţi:
-adăugarea sufixului –ază la numele substratului (zaharază, urează, glucozidază)
-denumire după acţiunea specifică a enzimei (guanilat ciclază, lactat dehidrogenază, glutation reductază)
-denumiri particulare fără legătură cu reacţia catalizată: pepsină, chimotripsină, tripsină.

• Numele enzimei cuprinde:-denumirea substratului, -tipul reacţiei catalizate, -coenzima
• Fiecare enzimă este codificată de patru cifre, care definesc: -clasa; -subclasa; -sub-subclasa;-numărul de ordine al enzimei din şir.

Clasificarea enzimelor
• Sunt 6 clase de enzime după tipul de reacţie catalizat:
• 1.Oxidoreductaze• 2.Transferaze• 3.Hidrolaze• 4.Liaze• 5.Izomeraze• 6.Ligaze

Oxidoreductazele catalizează reacţii redox(transfer de electroni sau de hidrogen)
Subclase: • Dehidrogenazele catalizează
transferul de hidrogen
de la substrat (SH2)
pe o coenzimă redox
(NAD+; NADP+; FMN; FAD):
SH2 + Co Sox + CoH2
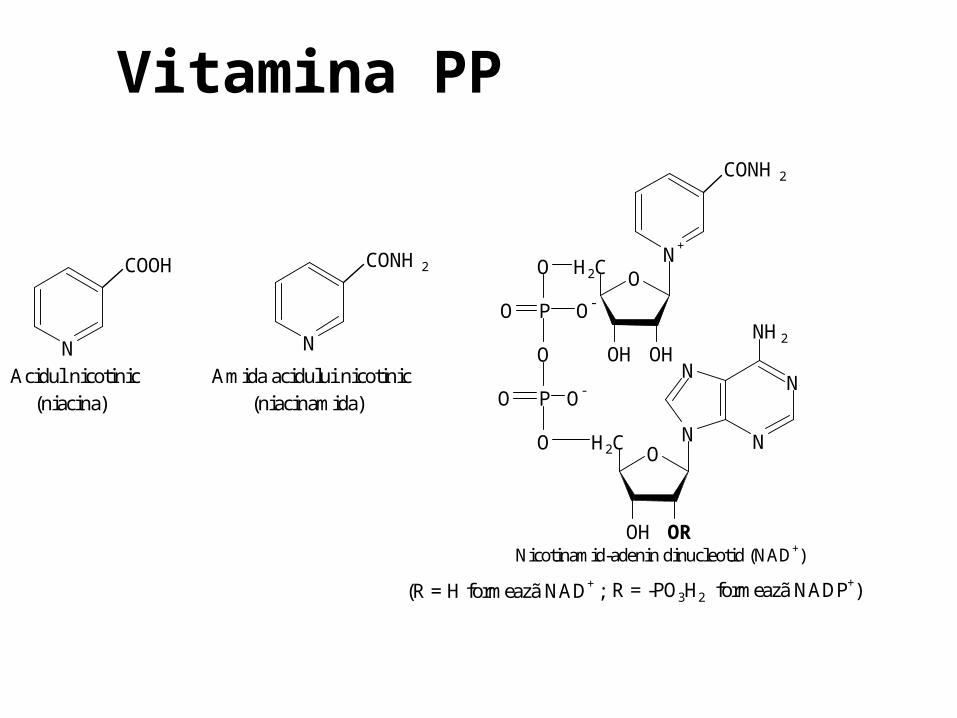
O N
N
N
N
NH2
OH2C
OROH
O P O-
O
O P O-
ON+
OH2C
OHOH
CONH2
N
COOH
Acidul nicotinic Amida acidului nicotinic (niacina) (niacinamida)
N
CONH2
Nicotinamid-adenin dinucleotid (NAD+)
(R = H formeazã NAD+ ; R = -PO3H2 formeazã NADP+)
Vitamina PP
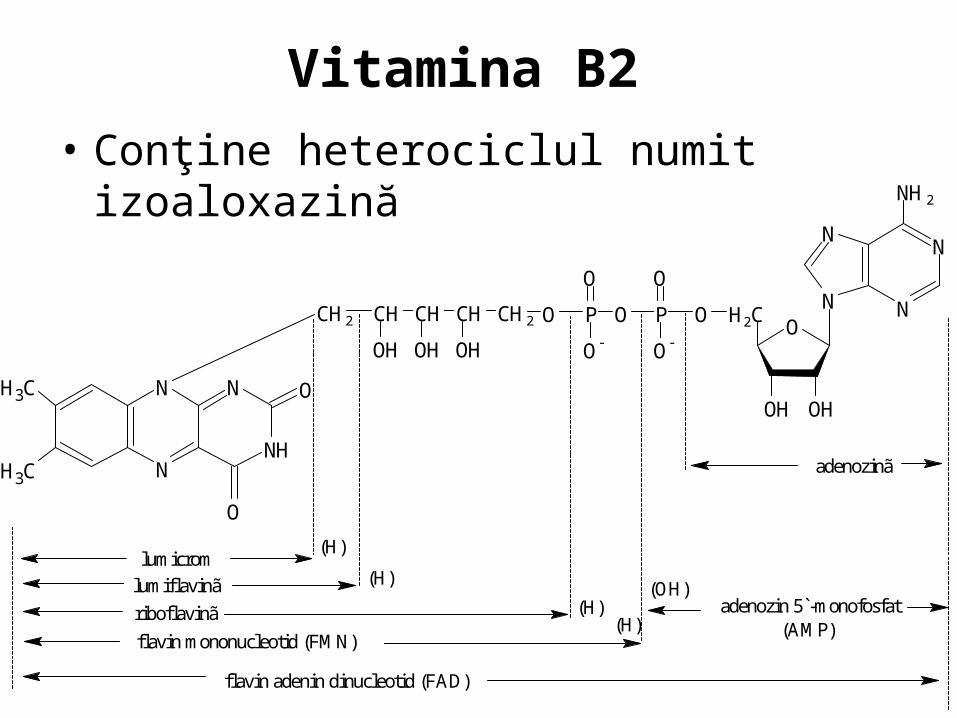
Vitamina B2
• Conţine heterociclul numit izoaloxazină
N N
NNH
CH2
O
OCH3
CH3
CH CH CH
OH OH OH
OCH2 P O
O-
O
P O
O-
O
N
N
N
N
NH2
OH2C
OHOH
lumicrom
lumiflavinã
riboflavinã
flavin mononucleotid (FMN)
adenozinã
adenozin 5`-monofosfat(AMP)
flavin adenin dinucleotid (FAD)
(H)
(H)
(H)(H)
(OH)
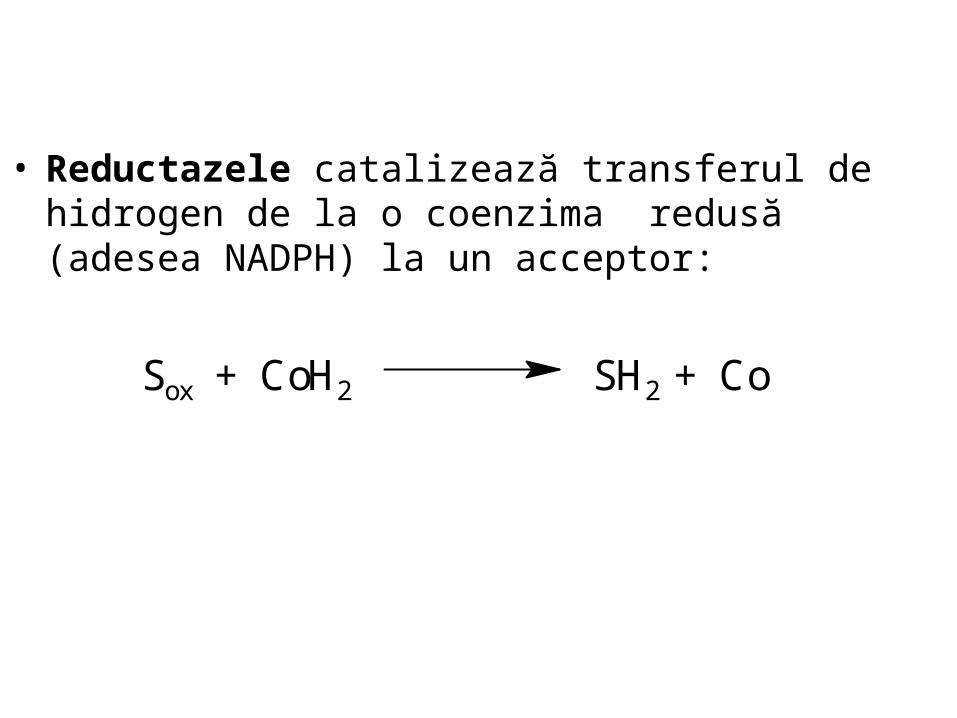
• Reductazele catalizează transferul de hidrogen de la o coenzima redusă (adesea NADPH) la un acceptor:
Sox + CoH2 SH2 + Co

• Oxidazele catalizează
transferul de echivalenţi reducători de la un substrat la dioxigen,
reducându-l fie la apă, fie la peroxid de hidrogen:
SH2 + 1/2 O2 Sox + H2O
SH2 + O2 Sox + H2O2
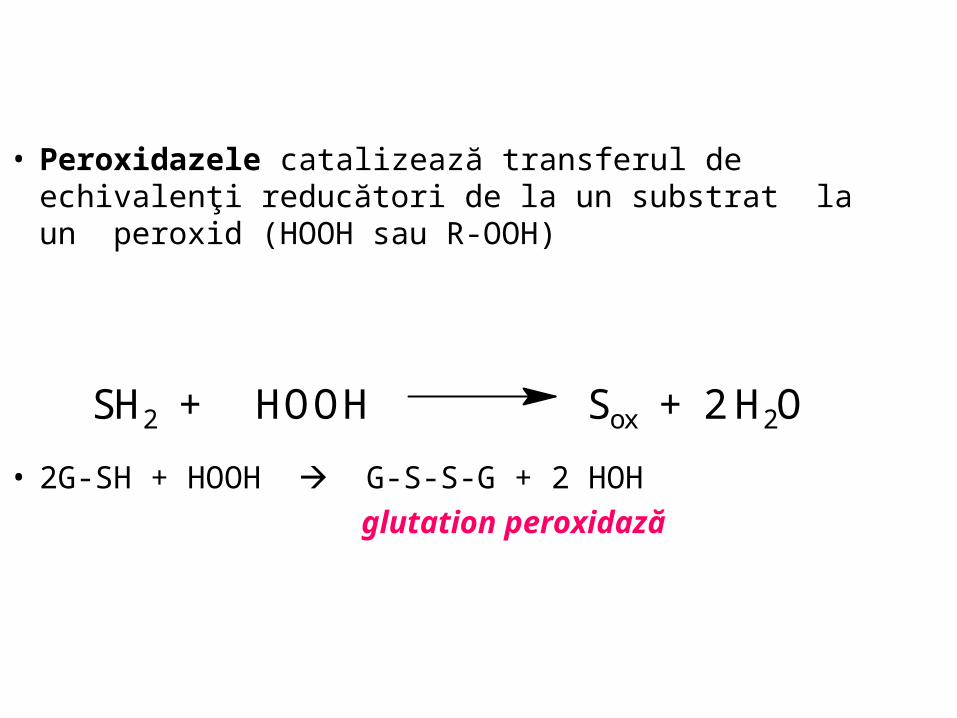
• Peroxidazele catalizează transferul de echivalenţi reducători de la un substrat la un peroxid (HOOH sau R-OOH)
• 2G-SH + HOOH G-S-S-G + 2 HOH
glutation peroxidază
SH2 + HOOH Sox + 2 H2O
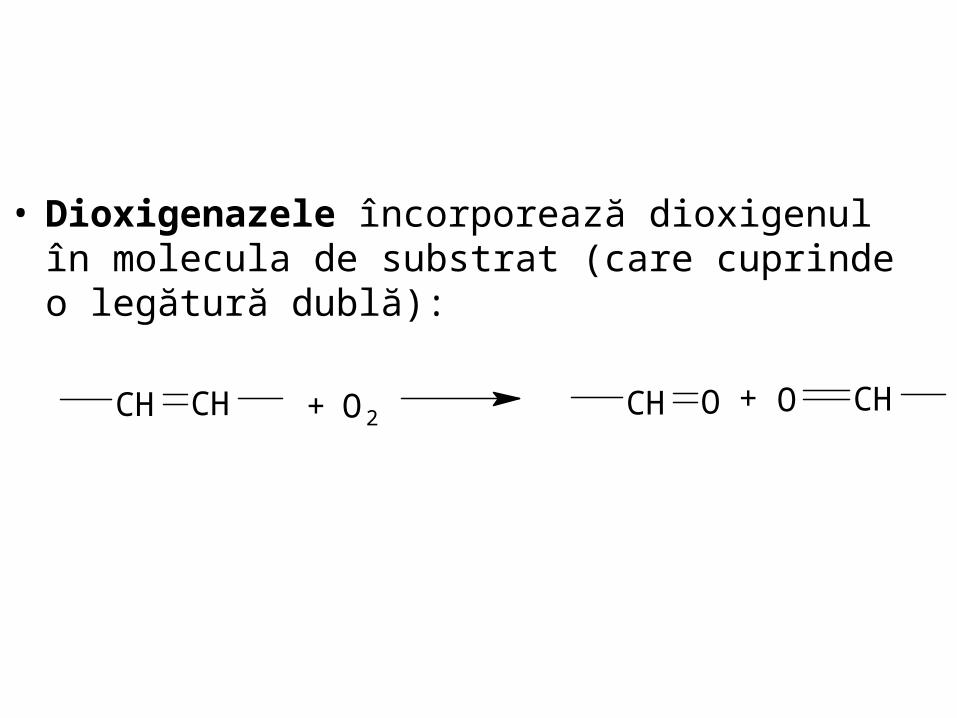
• Dioxigenazele încorporează dioxigenul în molecula de substrat (care cuprinde o legătură dublă):
CHCH + O2OCH CHO+

CH3
CH3H3C CH3 CH3H3C
H3C CH3CH3CH3-caroten
caroten dioxigenazaO2 în prezenta sãrurilor biliare
OH
CH3
CH3H3C
CH2
CH3 CH3
O
CH3
CH3H3C
CH
CH3 CH3
CH3
CH3H3C
COOH
CH3 CH3
Retinal (retinalaldehidã)
Reducere Oxidare
Retinol (Vitamina A1) Acid retinoic

• Monooxigenazele încorporează numai un atom de oxigen în substrat, celalalt atom al dioxigenului este redus la apă.
Multe monooxigenaze sunt hidroxilaze care implică participarea sistemului redox:
NADP+ - (NADPH+ H+)
R-H + O2 + (NADPH + H+) R-OH + H2O + NADP+

Transferazele
catalizează reacţii de transfer a unei grupări de la un donor către un acceptor
• Subclase: Aminotransferaze (transaminaze) catalizează transferul unei grupări amino între un amino şi un cetoacid. Coenzima participantă este piridoxal fosfatul.
AX + BY AY + BX
COOHCHR1
NH2
COOHCR2
O
COOHCR1
O
COOHCHR2
NH2
+ +
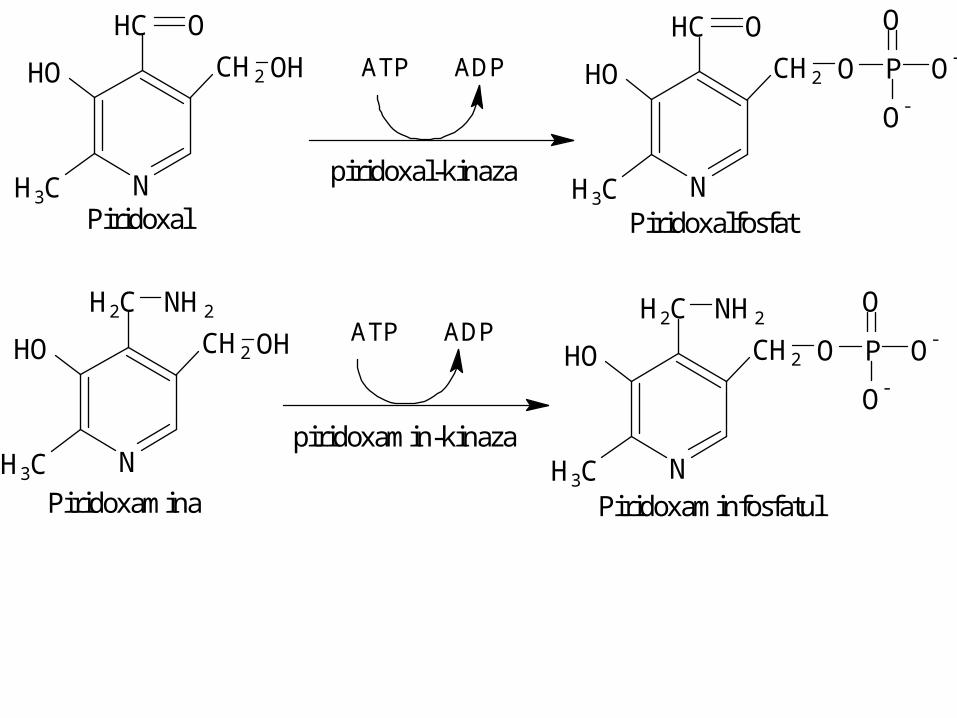
N
CH2HO
H3C
HC
OH
O
N
CH2HO
H3C
H2C
OH
NH2
Piridoxal N
CH2HO
H3C
HC
O
O
O-
O-
O
PATP ADP
piridoxal-kinaza
Piridoxalfosfat
Piridoxamina
ATP ADP
N
CH2HO
H3C
H2C NH2
O O-
O-
O
P
Piridoxaminfosfatul
piridoxamin-kinaza

+
COOH
CH2
CH2
HC
COOH
NH2
Acid glutamic
COOH
CH2
CH2
C
COOH
O
Acid-cetoglutaric
H3C CH
NH2
COOH + H3C C
O
COOH
Alaninã Acid piruvic
GPT

COOH
CH2
CH2
CH
COOH
NH2
Acid glutamic
COOH
CH2
CH2
C
COOH
O
Acid-cetoglutaric
GOT
COOH
CH2
CH
COOH
NH2
COOH
CH2
C
COOH
O+ +
Acid aspartic Acid oxaloacetic
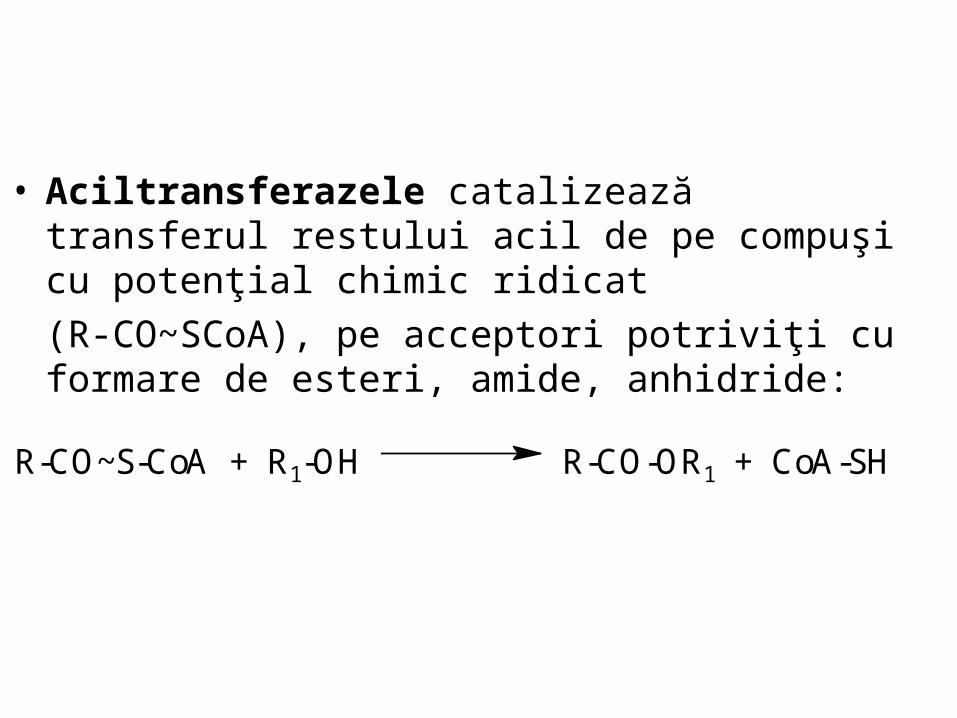
• Aciltransferazele catalizează transferul restului acil de pe compuşi cu potenţial chimic ridicat
(R-CO~SCoA), pe acceptori potriviţi cu formare de esteri, amide, anhidride:
R-CO~S-CoA + R1-OH R-CO-OR1 + CoA-SH

Acid pantotenic
OCH2CCHCNH
CH3
CH3OH
O
CH2CH2CO
O NO
H2C
OOH
O P O-
O
O P O-
N
NN
NH2
NH
SH
CH2
CH2
O-
O-
O
P
Tio
etan
olam
ina
AD
P
Coenzima A - SH

• Fosforil transferazele (kinaze)
catalizează transferul resturilor fosforil
de pe ATP
pe substraturi:
Glucozã + ATP Glucozo-6-P + ADP
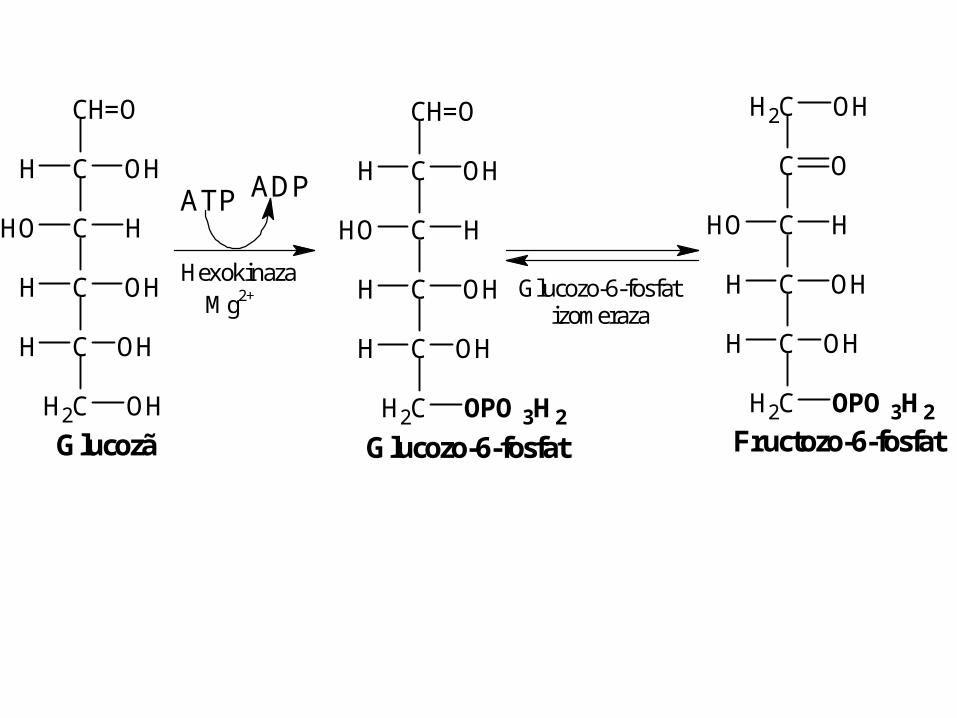
Glucozo-6-fosfat
CH=O
C
C
C
C
CH2 OPO3H2
H OH
OH H
H OH
H OH
CH=O
C
C
C
C
CH2 OH
H OH
OH H
H OH
H OH
Glucozã
ATP ADP
Hexokinaza Mg2+ Glucozo-6-fosfat
izomeraza
CH2
C
C
C
C
CH2 OPO3H2
O
OH H
H OH
H OH
OH
Fructozo-6-fosfat

• Metiltransferazele catalizează transferul
grupărilor cu un atom de carbon
• coenzima care poartă aceste grupări este
tetrahidrofolatul (FH4).
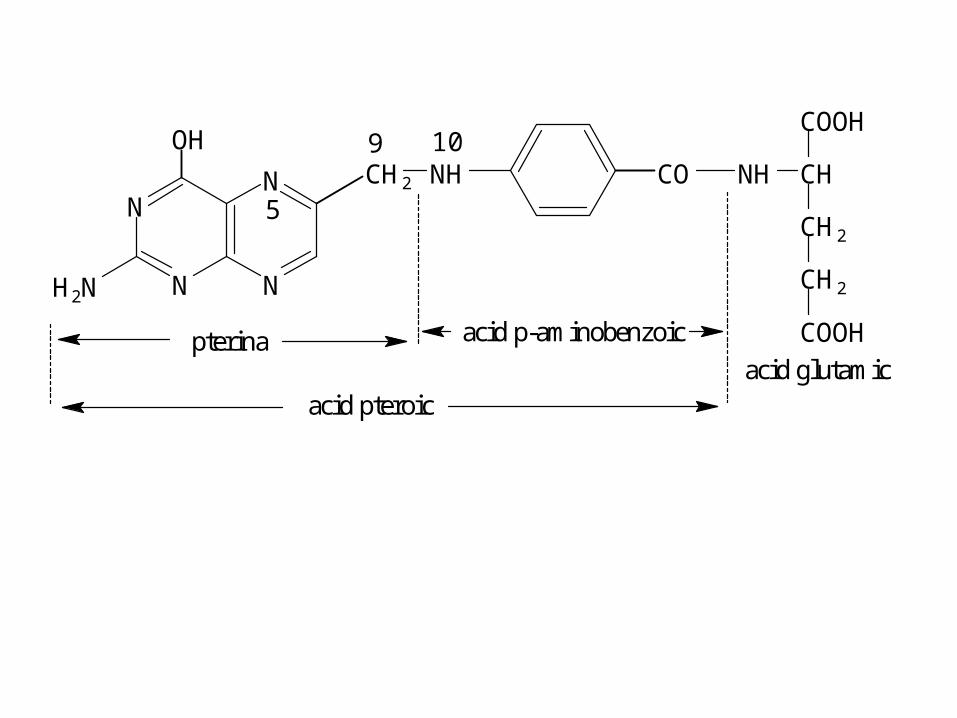
N
NN
N
H2N
OHCH2 NH CO NH
COOH
CH2
CH2
CH
COOH9 10
5
pterina acid p-aminobenzoic
acid glutamicacid pteroic

• Transportă grupări:
• metil (-CH3)
• hidroximetil (-CH2OH)
• formil (-CHO)
• formimino (-CH=NH)
• metilen (-CH2-),
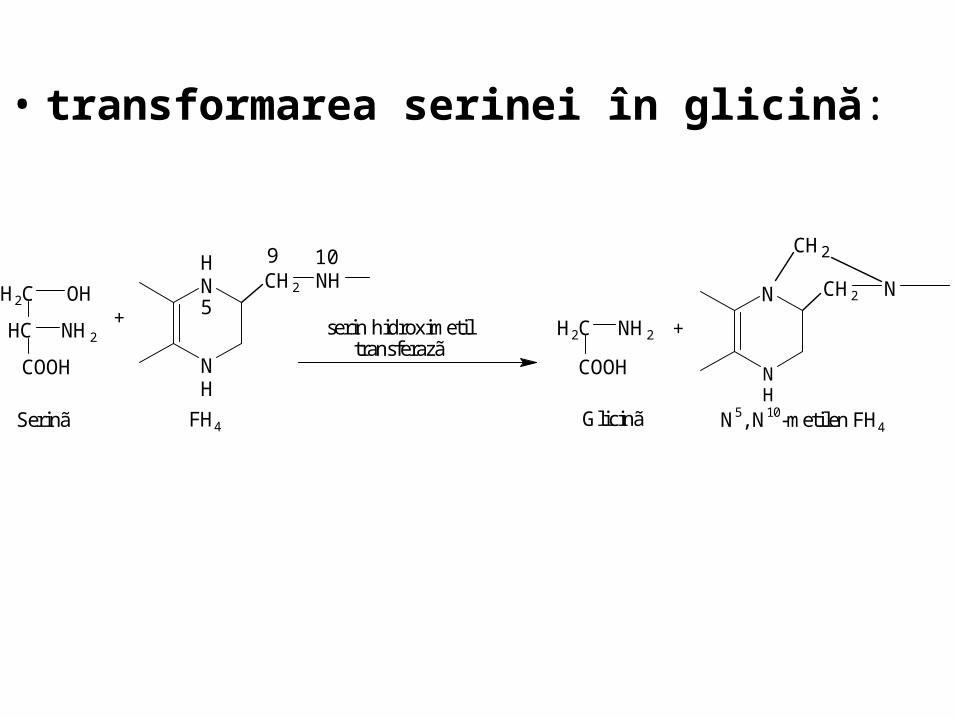
• transformarea serinei în glicină:
5
COOH
HC
H2C
NH2
OH
Serinã
9 10
NH
NH
CH2 NH
+
FH4
serin hidroximetiltransferazã
COOH
H2C NH2
Glicinã
+
N
NH
CH2 N
CH2
N5, N10-metilen FH4

• formarea grupărilor metil:
N5,N10-metilen-FH4 + (NADH + H+) N5-metil-FH4 + NAD+
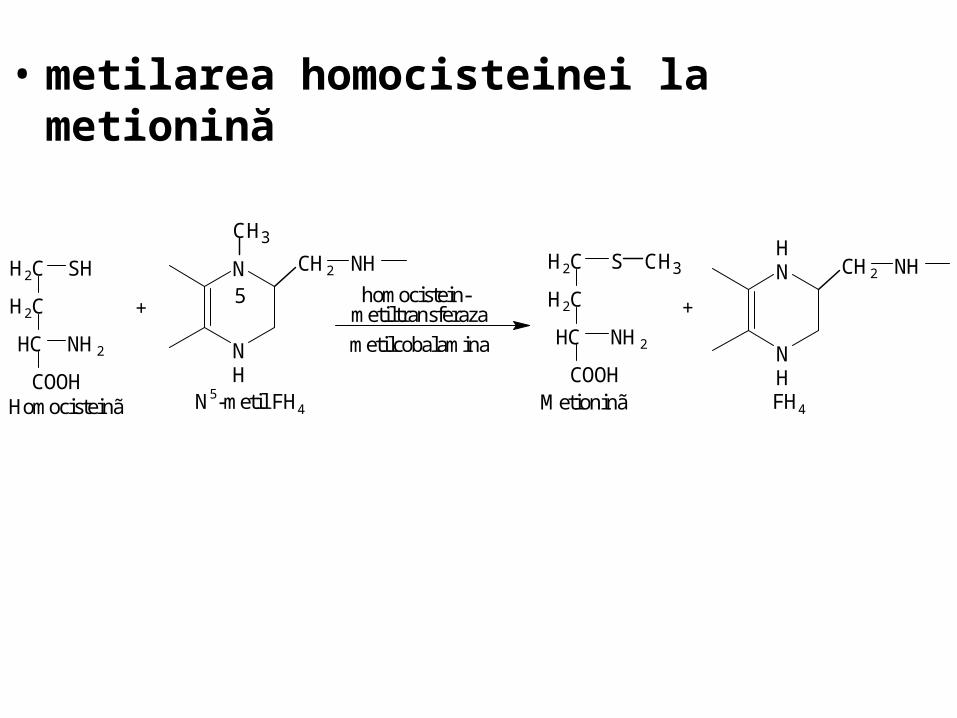
• metilarea homocisteinei la metionină
5
COOH
HC
H2C
NH2
H2C SH N
NH
CH2 NH
CH3
+
N5-metil FH4Homocisteinã
homocistein-metiltransferaza
metilcobalamina
COOH
HC
H2C
NH2
H2C S CH3
+
NH
NH
CH2 NH
FH4Metioninã
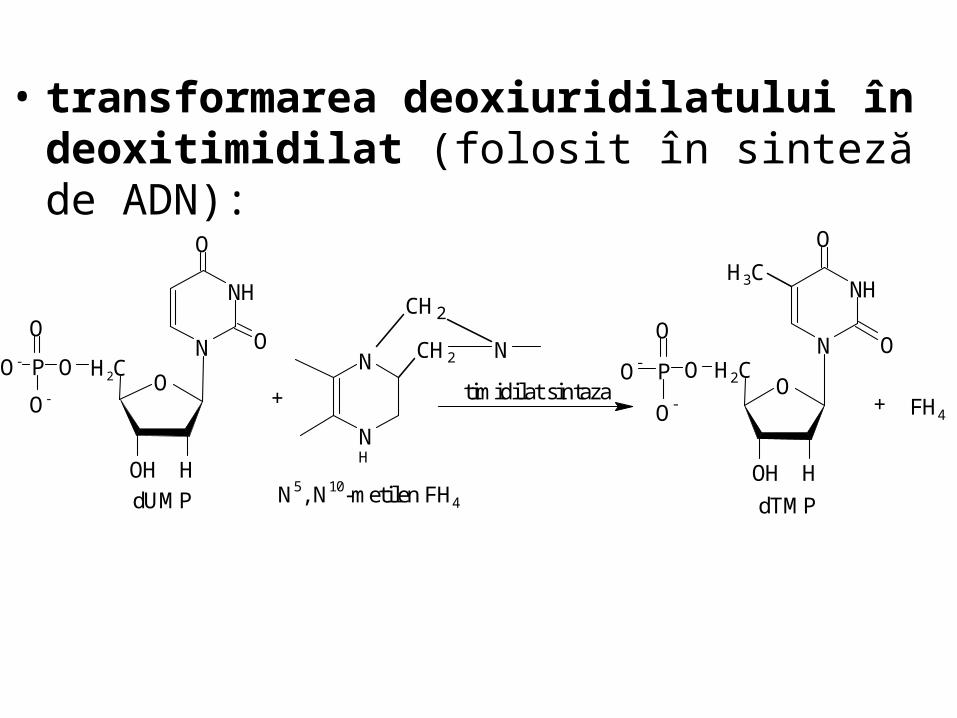
• transformarea deoxiuridilatului în deoxitimidilat (folosit în sinteză de ADN):
ON
OH2C
HOH
NH
O-
O
PO-
O
O
FH4+
N
NH
CH2 N
CH2
N5, N10-metilen FH4 dUMP
timidilat sintazaO
N
OH2C
HOH
NH
O-
O
PO-
O
O
H3C
dTMP
+

Hidrolazele
Subclase
• Esterazele care pot fi:
–carboxilesteraze (cu substrat R-COOR’)
–tiolesteraze (cu substrat R-CO -SR’)
–fosfomonoesteraze (substrat R-O-PO3H2)
–fosfodiesteraze sau fosfataze (substrat R-O-PO2H-O-R1)
• Glicozidaze• Peptidaze
A-B + H2O A-H + B-OH
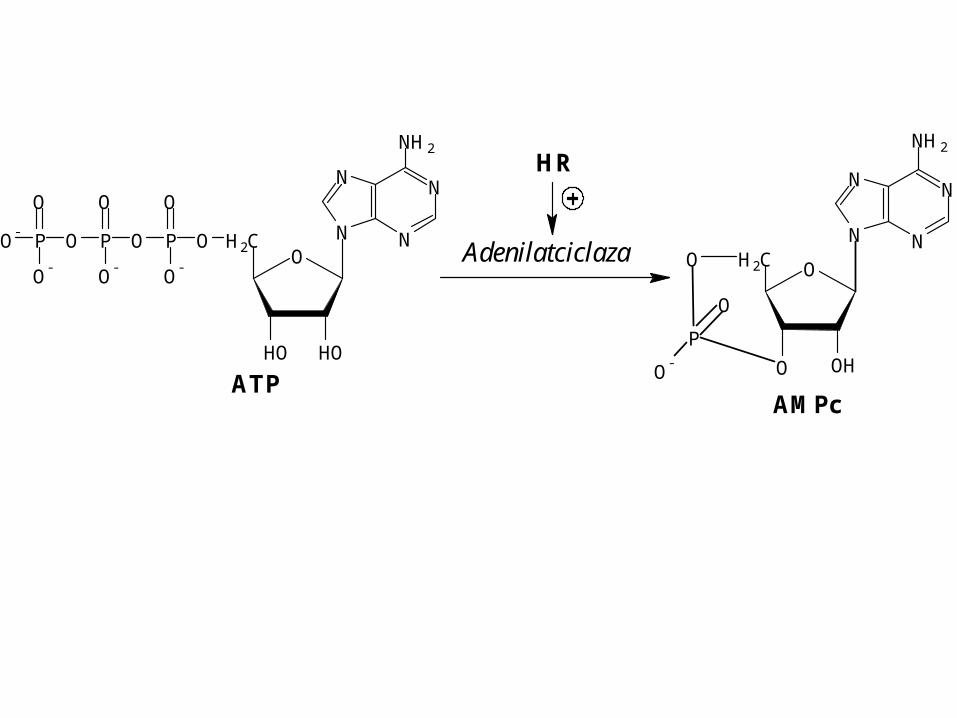
NO
H2CO
HOHO
P
O
O-
P
O
O
O-
O-
O-
O
O
P N
NN
NH2
ATP
OH2CO
OHO
N N
NN
NH2
P
O-
O
Adenilatciclaza
HR
AMPc

OH2CO
OHO
N N
NN
NH2
P
O-
O
AMPc
Fosfodiesteraza
Metilxantine
Insulinã
Ca2+-Calmodulinã
5`-AMP
NO
H2CO
HOHO
P
O
O-
O- N
NN
NH2

Liazele
• catalizează reacţii de adiţie reversibile, la legături multiple.
• Anhidraza carbonică catalizează hidratarea dioxidului de carbon:
CO2 + H2O H2CO3

• Fumaraza catalizează hidratarea acidului fumaric la acid malic:
+ H2O HC
CH2
COOH
COOH
OH
Acid malic
COOH
C
C
HOOC
H
H
Acid fumaric
Fumaraza

Izomerazele
• catalizează reacţii de izomerizare:
Racemazele transformă un enantiomer dextrogir în enantiomerul levogir
(+)-Alanina (-)-Alanina
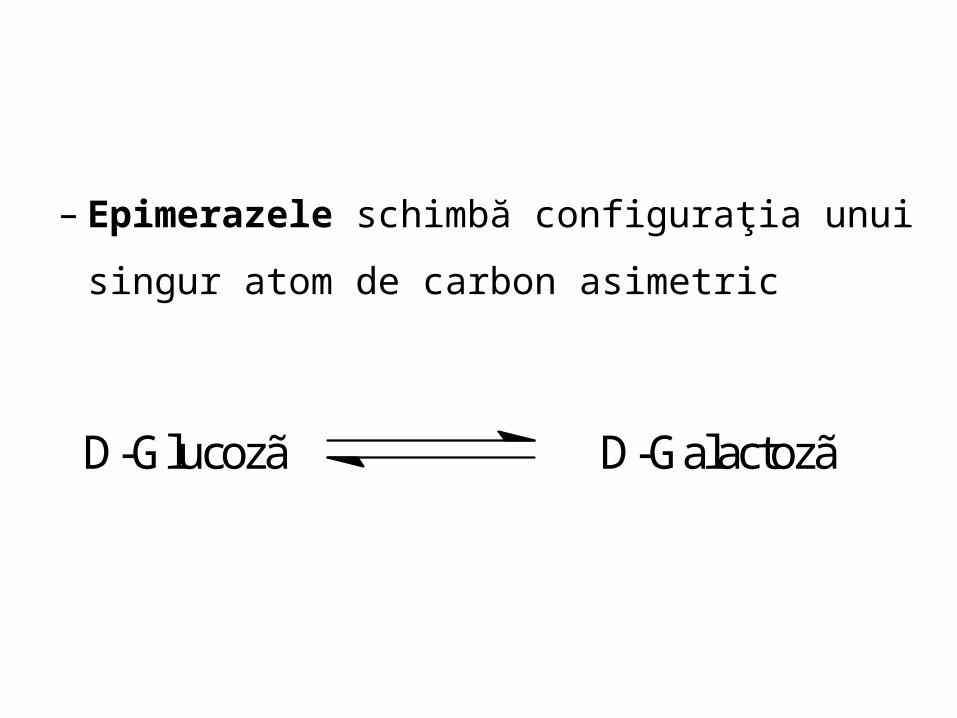
– Epimerazele schimbă configuraţia unui singur
atom de carbon asimetric
D-Glucozã D-Galactozã

D-Galactozã
CH
C
C
O
OHH
C
C
H
H
OH
OH
CH2 OH
H
OH
CH
C
C
O
OHH
C
C
H
OH
OH
H
CH2 OH
H
OH
CH
C
C
O
HOH
C
C
H
OH
OH
H
CH2 OH
H
OH
D-Glucozã D-Manozã
4020
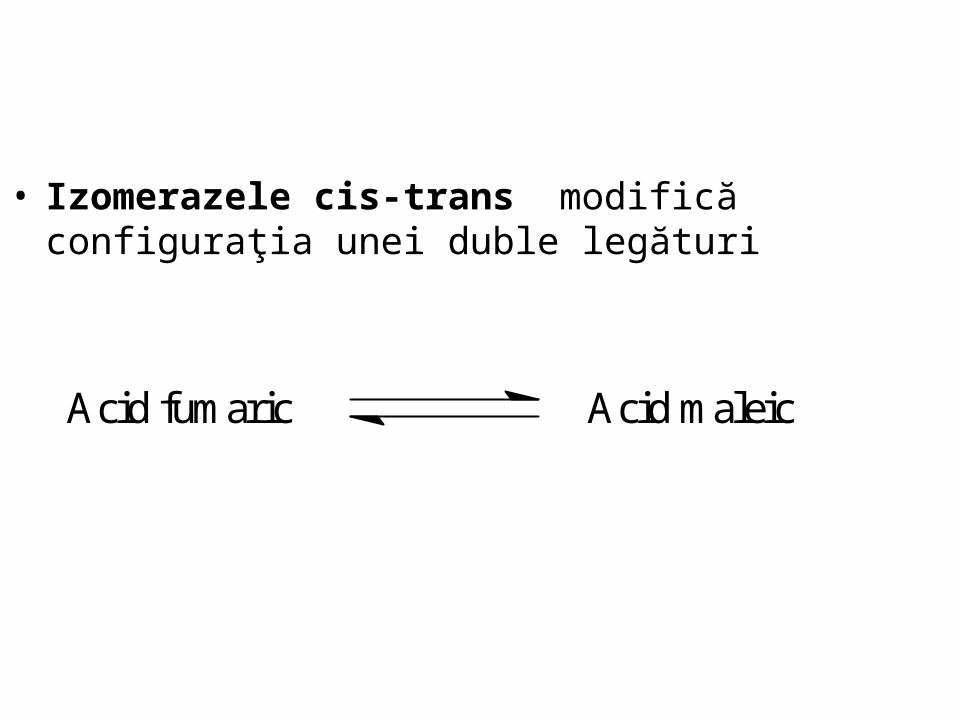
• Izomerazele cis-trans modifică configuraţia unei duble legături
Acid fumaric Acid maleic
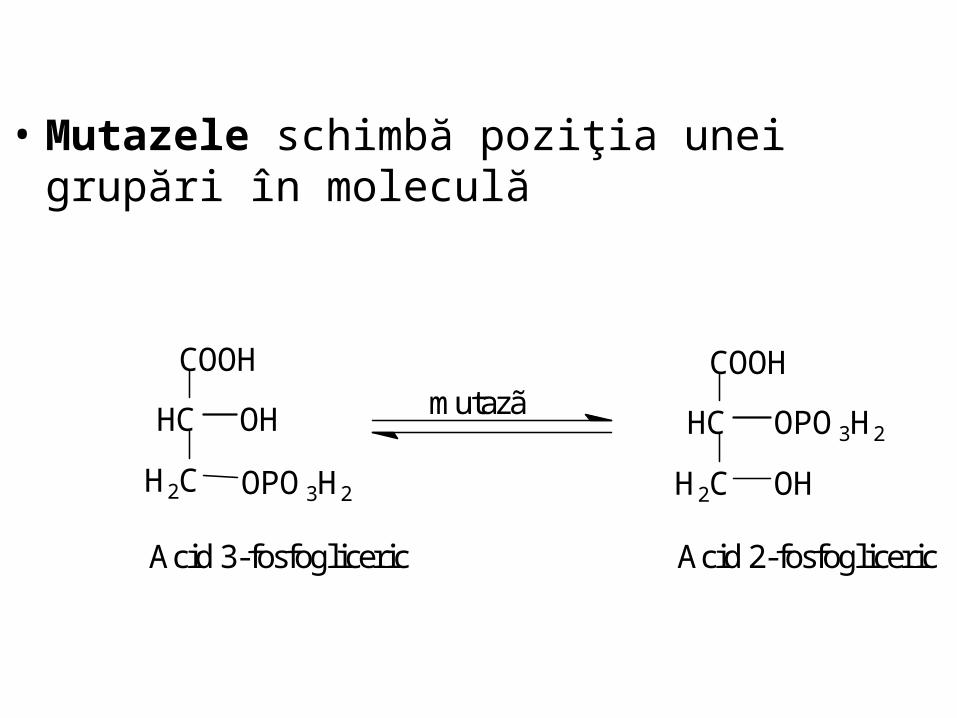
• Mutazele schimbă poziţia unei grupări în moleculă
HC
H2C OPO3H2
COOH
OH HC
H2C OH
COOH
OPO3H2
Acid 3-fosfogliceric Acid 2-fosfogliceric
mutazã

Ligazele
• catalizează reacţii de condensare cuplate cu scindare de ATP
• Sinteza glutaminei catalizată de glutaminsintetază:
A-H + B-OH A-B
ATPADP + Pi
Acid glutamic + NH3 GlutaminãATP
ADP

NH3
COOH
CH2
CH2
HC
COOH
NH2
Acid glutamic
ATPADP+Pi
C
CH2
CH2
HC
COOH
NH2
O
NH2
Glutaminã
Glutamin sintetazã+

• Sintazele catalizează legarea a două molecule fără
implicarea ATP
☺aminolevulinat sintaza
☺glicogen sintaza
☺NOS - nitric oxid sintaza

• Vitaminele care pot fi transformate în coenzime pentru a
participa la reacţii de oxido-reducere sunt:
A.Nicotinamida şi riboflavina
B.Vitamina C şi biotina
C.Biotina şi nicotinamida
D.Tiamina şi riboflavina
E.Tiamina şi Biotina



![16 Programe TV i Z S e d ` n u m a i c u I n f o r m a ] …...16 Informa\ia zilei Programe TV vineri 8 martie 2019S e d ` n u m a i c u I n f o r m a ] i a z i l e i ! S`n`tate &](https://static.fdocumente.com/doc/165x107/5e9682041b881c5c485610ed/16-programe-tv-i-z-s-e-d-n-u-m-a-i-c-u-i-n-f-o-r-m-a-16-informaia-zilei.jpg)
![16 Programe TV i Z S e d ` n u m a i c u I n f o r m a ] i a z i l e i ! & … · 2019-03-14 · 16 Informa\ia zilei Programe TV vineri 1 martie 2019 S e d ` n u m a i c u I n f o](https://static.fdocumente.com/doc/165x107/5f890f162979536e4238cc63/16-programe-tv-i-z-s-e-d-n-u-m-a-i-c-u-i-n-f-o-r-m-a-i-a-z-i-l-e-i-2019-03-14.jpg)













