

39385592 Www Referat Ro Studii Privind Biosinteza Unui Complex Multienzimatic Utilizabil CA Aditiv...
58
‘UNIVERSITATEA DE ŞTIINŢE AGRONOMICE ŞI MEDICINĂ VETERINARĂ BUCUREŞTI FACULTATEA DE BIOTEHNOLOGII SPECIALIZAREA BIOTEHNOLOGII MEDICAL-VETERINARE PROIECT DE DIPLOMĂ Şef lucrări dr.chimist LUMINIŢA IONESCU CSI dr. chimist : GETA AVRAM Absolvent : Marinescu Georgiana www.referat.ro www.referat.ro
-
Upload
teo-brunetzik -
Category
Documents
-
view
108 -
download
3
Transcript of 39385592 Www Referat Ro Studii Privind Biosinteza Unui Complex Multienzimatic Utilizabil CA Aditiv...

‘UNIVERSITATEA DE ŞTIINŢE AGRONOMICE ŞI
MEDICINĂ VETERINARĂ BUCUREŞTI
FACULTATEA DE BIOTEHNOLOGII
SPECIALIZAREA BIOTEHNOLOGII MEDICAL-VETERINARE
PROIECT DE DIPLOMĂ
Şef lucrări dr.chimist LUMINIŢA IONESCU
CSI dr. chimist : GETA AVRAM
Absolvent : Marinescu Georgiana
www.referat.ro
www.referat.ro

BUCUREŞTI
2010
UNIVERSITATEA DE ŞTIINŢE AGRONOMICE ŞI MEDICINĂ VETERINARĂ – BUCUREŞTI
FACULTATEA DE BIOTEHNOLOGII
SPECIALIZAREA BIOTEHNOLOGII MEDICAL
VETERINARE
STUDII PRIVIND BIOSINTEZA UNUI COMPLEX
MULTIENZIMATIC UTILIZABIL CA ADITIV
FURAJER
Coordonator Ştiinţific : LUMINIŢA IONESCU
CSI dr. chimist : GETA AVRAM

Absolvent : Marinescu Georgiana
BUCUREŞTI
2010

Cuprins
REZUMAT………………………………………………………………………………
…………..3
INTRODUCERE ................................................................................................................ 7 PARTEA I ......................................................................................................................... 11 STUDII DOCUMENTARE ............................................................................................. 11 Capitolul 1. Enzime hidrolitice de uz furajer şi microorganisme producătoare ....... 11
α-amilaza ........................................................................................................................ 11 Celulazele ....................................................................................................................... 13
1.3. Hemicelulazele .................................................................................................... 14 1.4. Microorganisme producătoare de enzime hidrolitice de uz furajer .................... 16
Capitolul 2. Utilizarea deşeurilor şi/sau subproduselor agricole în medii de biosinteză a enzimelor hidrolitice de uz zootehnic ........................................................ 20 Capitolul 3. ........................................................................................................................ 24 CERCETĂRI EXPERIMENTALE ............................................................................... 27 Capitolul 4. Materiale şi metode ..................................................................................... 27
4.1 Tulpini de lucru ........................................................................................................ 27 4.2. Obţinerea culturilor de intreţinere ........................................................................... 27
................................................................................................. 27 4.3. Cultivarea în laborator şi testarea activităţii hidrolitice .................................... 28 4.4. Determinarea activităţilor enzimatice ................................................................. 30 4.5. Testarea produsului enzimatic în hrana animalelor de fermă ............................. 39
Capitolul 5. Rezultate şi discuţii ..................................................................................... 42 5.1. Compoziţia mediilor pentru cultura de întreţinere ................................................. 42 5.2 Analiza comparativă a biosintezei enzimelor hidrolitice cu tulpini Aspergillus niger şi Trichoderma viride pe diferite medii ......................................................................... 42 5.3. Temperatura optimă de cultivare ............................................................................ 45 5.3 Alegerea pH-ului optim de cultivare ........................................................................ 47 5.4. Testarea complexului enzimatic ca aditiv furajer ................................................. 51
CONCLUZII ....................................................... 54 BIBLIOGRAFIE ............................................................................................................. 56

REZUMAT
Numeroase nutreţuri ieftine sunt limitate ca utilizare în hrana animalelor datorită
disponibilităţii reduse a nutrientilor pe care îi contin. Astfel, anumite reţete pe bază de
cereale, care, de altfel, se folosesc şi în ţara noastră, conţin cantităţi importante de
poliglucide neamidonoase PNA (polizaharide fără amidon). Acestea sunt reprezentate în
principal de beta-glucani (în orz şi ovăz), arabinoxilani (în grâu şi tărâţe), polimeri pe
bază de rafinoză (în şrotul de soia) celuloză şi hemiceluloză (şrotul de floarea soarelui),
fapt care limiteaza sever folosirea lor în hrana animalelor. PNA din cereale şi tărâţele
acestora manifesta o activitate antinutritivă când sunt prezenţi în cantităţi mari în hrana
păsărilor şi porcilor. Ele nu pot fi degradate de enzimele din tubul digestiv al acestor
animale. Efectele antinutritive ale PNA se concretizeaza în diminuarea digestibilitatii
amidonului, proteinelor şi grăsimilor, determinând astfel un efect general de reducerea
ratei de creştere şi de valorificare a hranei. PNA măresc vâscozitatea conţinutului
intestinal şi împiedica difuzia enzimelor endogene şi mixarea aceatora în masa
continutului digestiv.
Având în vedere structura complexă a PNA din materiile prime furajere, este
evident că un singur tip de activitate enzimatică nu poate asigura disponibilizarea tuturor
substanţelor nutritive din componentele hranei, fiind necesară combinarea mai multor
produse diferite (de unde necesitatea obţinerii unor complexe multienzimatice).
În acest context, in proiectul de faţã s-a urmărit selecţia unor tulpini fungice cu
capacitate crescutã de producere a unui complex de enzime hidrolitice prin utilizarea unor
formule noi de substraturi nutritive, precum şi stabilirea condiţiilor optime de biosinteză a
acestor enzime cu microorganismele selecţionate.
In prima parte a lucrării sunt prezentate studii documentare referitoare la enzime
hidrolitice de uz furajer şi microorganisme producătoare ale acestora, utilizarea deşeurilor
şi/sau subproduselor agricole în medii de biosinteză a enzimelor hidrolitice de uz
zootehnic, caracteristici ale proceselor de fermentatie cu microorganisme producatoare de
enzime hidrolitice utilizabile ca aditivi furajeri.

În cadrul cercetărilor experimentale prezentate în proiect s-au testat două tulpini
de Aspergillus niger şi o tulpină Trichoderma viridae din colecţia Centrului de
Biotehnologii Microbiene şi respectiv a Facultăţii de Biologie, Universitatea Bucureşti, în
ceea ce priveşte capacitatea acestora de a produce un complex de enzime hidrolitice
(proteaze, amilaze, celulaze, xilanaze) utilizabil ca aditiv furajer.
Cele trei tulpini de fungi au fost cultivate în sistem submers, pe 4 medii de
cultură preluate din literatura de specialitate; activitatea polienzimatica a mediilor de
fermentaţie şi a produselor finite (obţinute prin filtrarea mediului de fermentaţie,
concentrare şi adsorbţie pe CaCO3 tehnic) s-a evaluat comparativ cu preparatul
KEMZYME®VP DRY (Belgia).
Rezultatele obţinute au evidenţiat capacitatea bioproductivă superioară a tulpinii
Aspergillus niger CMGB-401 în ceea ce priveşte complexul enzimatic studiat.
În scopul determinării condiţiilor optime de cultivare a tulpinii de Aspergillus
niger CMGB-401 pentru biosinteza enzimelor hidrolitice (amilaze, proteaze, celulaze,
xilanaze), au fost cercetaţi o serie de parametrii care au o importanţă semnificativă pentru
acest tip de bioprocese şi anume: compoziţia mediul de cultură, temperatura, pH-ul
mediului de cultură şi gradul de agitare şi aerare al culturii.
În ceea ce priveşte compoziţia mediului de cultură, dintre cele cinci medii testate
(patru medii lichide şi un mediu semisolid pe bază de tărâţe de grâu), cele mai bune
rezultate s-au obţinut cu mediul lichid M 4, având următoarea compoziţie (g%):
Făină de porumb-2%; Făină de soia-3%; (NH4)2HPO4-0,5%; MgSO4x7H2O-0,05%; CaCl2-0,1%
În cazul diferitelor variante de pH şi temperatură folosite, s-a observat că tulpina
de Aspergillus niger CMGB-401 conduce cele mai bune activităţi enzimatice, în urma
cultivării la pH = 5,5 şi la temperatura de 280C (activitatea amilolitică 141,6 UDNS/g,
activitatea proteolitică 1,95 UP/g, activitatea celulozolitică 2,27 UC1-Cx/g, activitatea
xilanazică 1366,33 U/ml).

Condiţionarea produsului polienzimatic s-a realizat prin concentrarea mediului
integral din finalul fermentaţiei, tratarea cu CaCO3 a concentratului şi uscarea sub vid a
amestecului rezultat.
Testarea ca aditiv furajer a produsului polienzimatic condiţionat a evidenţiat
faptul că utilizarea acestuia nu a influenţat semnificativ greutatea corporală şi sporul
mediu zilnic a purceilor în perioada crizei de înţărcare, posibil din cauza faptului că
nutreţul combinat a fost optimizat în principii nutritivi specifici acestei categorii de
porcine;
Consumul specific la loturile de purcei la care s-a utilizat preparat enzimatic este
cu 1,52% - 4,04% mai mic faţă de martor, in funcţie de cantitatea de produs introdusă în
raţia zilnică.
Rezultatele obţinute sugerează eficienţa utilizării preparatelor enzimatice în
recepturi de nutreţ combinat deficitare în principii nutritive.
INTRODUCERE
Odatã cu integrarea în Uniunea Europeanã, România s-a aliniat la Standardele
Europene în ceea ce priveşte asigurarea calitãţii nutreţurilor cu un impact deosebit asupra
sãnãtãţii animalelor şi în consecinţã a oamenilor, s-a interzis utilizarea pulberilor de
origine animalã în furajarea animalelor şi prin urmare s-au căutat alte cãi pentru ca
necesarul de proteinã furajerã sã fie acoperit. Printre sursele alternative de proteine
furajere se numără:
− surse semiconvenţionale, cum sunt proteinele seminţelor de
oleaginoase (soia) şi din concentratele de peşte, utilizate curent
astãzi în diferite ţãri;
− surse neconvenţionale, cum sunt proteinele furnizate de
microorganisme: mucegaiuri, bacterii, drojdii, alge.
De peste 60 de ani domeniul aditivilor furajeri este explorat prin numeroase
cercetări, timp în care au fost elaborate şi testate o multitudine de produse experimentale

şi comerciale. De la simple substanţe de tipul acizilor organici pâna la sisteme de aditivi
cu peste 10 substanţe active încorporate, aceasta nouă clasă de compuşi a devenit una din
principalele căi de sporire şi eficientizare a performanţelor de valorificare a hranei de
către animalele de interes zootehnic.
Numeroase nutreturi ieftine sunt limitate ca utilizare în hrana animalelor datorita
disponibilitatii reduse a nutrientilor pe care îi contin. Astfel, anumite reţete pe bază de
cereale, care, de altfel, se folosesc şi în ţara noastră, conţin cantităţi importante de
poliglucide neamidonoase PNA (polizaharide fără amidon). Acestea sunt reprezentate în
principal de beta-glucani (în orz si ovaz), arabinoxilani (în grâu si tarâte), polimeri pe
bază de rafinozã (în şrotul de soia) celuloză si hemiceluloză (şrotul de floarea soarelui),
fapt care limiteaza sever folosirea lor în hrana animalelor. PNA din cereale si tarâtele
acestora manifesta o activitate antinutritiva când sunt prezenti în cantitati mari în hana
pasarilor si porcilor. Ele nu pot fi degradate de enzimele din tubul digestiv al acestor
animale. Efectele antinutritive ale PNA se concretizeaza în diminuarea digestibilitatii
amidonului, proteinelor si grasimilor, determinând astfel un efect general de reducerea
ratei de crestere si de valorificare a hanei. PNA maresc vâscozitatea continutului
intestinal si împiedica difuzia enzimelor endogene si mixarea aceatora în masa
continutului digestiv. Ca rezultat, digestibilitatea nutreţurilor se reduce şi apar o serie de
tulburări ca: diaree (simptom asociat frecvent cu prezenta grâului în cantităţi mari în
raţii), consum excesiv de apă, dejecţii lipicioase sau cu umiditate ridicată (care afectează
negativ microclimatul). În acelaşi timp pentozanii si betaglucanii se pot combina cu
enzime specifice ale tractusului digestiv, reducându-le activitatea. Astfel eficienţa
conversiei hranei este redusă şi costul/kg spor este ridicat, sporul mediu zilnic fiind redus.
O soluţie pentru diminuarea efectului negativ al PNA asupra producţiei animaliere
este reprezentată de adiţia de enzime în hrana animalelor.
Enzimele sunt catalizatori biologici de natura proteica care participa la numeroase
reactii chimice ce au loc in sistemele vii şi fara de care viata nu ar putea exista. Ele au rol
important in reglarea matabolismului substantelor nutritive, a proceselor de crestere a
tesuturilor la animale si pasari. Desi ele se formeaza si functioneaza ca si catalizatori doar
in celule vii, cele mai multe pot fi separate si-si pot continua activitatea in vitro.

În ultima perioada, eforturile cercetatorilor din intreaga lume s-au indreptat spre
studierea efectului enzimelor de natura exogena in cresterea disponibilitatii si
digestibilitatii substantelor nutritive si valorii energetice a nutreturilor, imbunatatirea
performantelor productive, reducerea costurilor hranei pe unitatea de produs. Cerintele
noilor enzime furajere sunt: activitati specifice mari in conditiile de lucru impuse,
rezistenta mare la inactivare prin tratament termic, pH scazut si la actiunea enzimelor
proteolitice, costuri scazute de productie, stabilitate marita la pastrare.
Prin încorporarea enzimelor exogene specifice în nutreţul combinat pentru porci
sau păsări, se poate realiza creşterea performanţelor productive şi, totodată, protejarea
mediului prin reducerea conţinutului excretei în azot şi fosfor. Aceste aspecte sunt
susţinute de cercetările de specialitate care demonstrează, de exemplu, corelaţia între
progresul genetic la porc şi schimbările în raţiile specifice categoriei şi vârstei, astfel că,
răspunsul animalelor la adiţia de enzime nu depinde numai de substrat dar şi de
capacitatea fiziologică a animalelor de a utiliza nutrienţii.
Având în vedere structura complexă a PNA din materiile prime furajere, este
evident că un singur tip de activitate enzimatică nu poate asigura disponibilizarea tuturor
substanţelor nutritive din componentele hranei, fiind necesară combinarea mai multor
produse diferite (de unde necesitatea obţinerii unor preparate enzimatice complexe). De
exemplu, la grâu şi secară, unde predomină arabinoxilanii este necesar ca în complexul
enzimatic utilizat, activitatea esenţială să fie orientată spre acest substrat, chiar dacă
pentru cele mai multe din dietele pe bază de proteine vegetale este necesară adăugarea de
alfagalactozide şi pectinaze.
Hemiceluloza este o componentă a fracţiunii polizaharidice care se găseşte la un
nivel ridicat în peretele celular al plantei. Acesta este formată dintr-un grup de
heteropolizaharide solubile în mediu alcalin, care sunt asociate cu celuloza si intra in
constitutia peretelui celular vegetal, incluzând β-D-glucani necelulozici, substanţe pectice
(poligalacturonani) şi mai multe heteropolizaharide cum sunt cele formate în mod
predominant din galactoză (arabinogalactani), manoză (galactogluco- şi glucomanani) şi
xiloză (arabinoglucurono- şi glucuronoxilani). De fapt, heteropolizaharidele cu un grad
de polimerizare redus (100 – 200 unităţi) comparativ cu al glucozei (10000 – 14 0000),
sunt în general denumite cu termenul de hemiceluloze. Principalii componenţi glucidici ai

heteropolizaharidelor hemicelulozice sunt: D-xiloza, D-manoza, D-glucoza, D-galactoza,
L-arabinoza, acidul D-glucuronic, acidul D-galacturonic şi mai puţin L-rhamnoza, L-
fucoza şi diferite glucide O-metilate. Dintre aceştia β-glucanii şi arabinoxilanii sunt
recunoscuţi ca factori antinutritivi în cereale.
Adiţia enzimelor exogene precum amilaza, celulaza, proteaza, xilanaza, în dietele
furajere, oferă nutriţioniştilor posibilităţi multiple pentru stimularea proceselor digestive
şi metabolice la animale, contribuind la ridicarea eficienţei biologice şi economice a
hranei.
Datele din literatura de specialitate atestă faptul că enzimele hidrolitice sus-
menţionate se obţin fie prin fermentaţie în sistem submers, fie pe substraturi solide, cu
microorganisme specifice genului Aspergillius.
În acest context, in proiectul de faţã s-a urmărit selecţia unor tulpini fungice cu
capacitate crescutã de producere a unui complex de enzime hidrolitice prin utilizarea unor
formule noi de substraturi nutritive, pe baza utilizãrii reziduurilor agricole ca materii
prime în procesele fermentative precum şi stabilirea condiţiilor optime de biosinteză a
acestor enzime cu microorganismele selecţionate.

PARTEA I
STUDII DOCUMENTARE
Capitolul 1. Enzime hidrolitice de uz furajer şi
microorganisme producătoare
α-amilaza
α-amilazele (α-1,4-D-glucan-4-glucohidrolaza EC 3.2.1.1.) se întâlnesc în toate
organismele vii (microorganisme, plante, animale) realizând hidroliza substraturilor
amidonoase până la oligozaharide. Ele sunt sunt proteine mici, cu mase moleculare
cuprinse între 50.000 şi 60.000. Situsul catalitic al α-amilazelor este format din doi
aminoacizi (Asp şi Glu) care se găsesc în zona de legare a substratului [2]. Mecanismul
hidrolizei, asemănător celui întâlnit în cazul lizozimului, începe printr-o slăbire a legăturii
C1-O4 în vederea hidrolizei, urmată de formarea complexului enzimă-substrat. Această
legătură continuă să fie slăbită prin acţiunea unuia din cei doi aminoacizi, care se
comportă ca donor de proton pentru O4. Ruperea legăturii glicozidice duce la formarea
unui ion intermediar oxicarbonic (stare de tranziţie) care este stabilizat de încărcătura
negativă a celorlalţi aminoacizi. În final, acest ion reacţionează cu o moleculă de apă şi
cele două oligozaharide formate părăsesc situsul catalitic (fig. 1).
pH-ul optim al α-amilazelor depinde de originea acestora, situându-se între 4,8 şi
6,9. Valori extreme de pH se întâlnesc la α-amilaza acidă din Bacillus subtilis (pH 3,5)
sau α-amilaza bazică din Bacillus licheniformis (pH 9,0). Prezenţa ionilor de calciu
favorizează uneori creşterea stabilităţii enzimei în condiţii extreme de pH.
Temperatura optimă de activitate variază şi ea în funcţie de originea enzimei.
Temperatura medie este de 40-500C, dar poate ajunge până la 70-800C în cazul α-
amilazelor bacteriene din Bacillus stearothermophilus, Bacillus subtilis, Bacillus
licheniformis (tabelul 1).

Figura 1. Mecanismul propus pentru reacţia de hidroliză catalizată de α-amilază
Tabelul 1. Proprietăţile α-amilazelor (E.C. 3.2.1.1)
Originea enzimei Masa moleculara pH optim Temperatura optima (0C)
AnimalăPancreas de porc 50.000 6,9 37
VegetalaMalţ de orz 59.500 4,7 – 5,4 50 - 55Microbiană
Bacillus amyloliquefqciens 49.000 5,9 65B. licheniformis 22.500 5,0 – 9,0 76
B. stearothermophilus 49.000 5,4 – 6,1 70B. subtilis 47.000 5,3 – 6,4 50A. oryzae 52.600 5,5 – 6,9 40

Celulazele
Celulazele, enzimele responsabile de hidroliza celulozei, sunt compuse dintr-un
amestec complex de proteine enzimatice, cu diferite specificităţi de hidroliză a legăturilor
glicozidice. În funcţie de activitatea lor enzimatică, ele pot fi împărţite în trei clase
majore: endoglucanaze sau endo-1,4-β-glucanaze (E.C.3.2.1.4), celobiohidrolaze
(E.C.3.2.1.91) şi β-glucozidaze (E.C.3.2.1.21). Se consideră că endoglucanazele, deseori
denumite carboximetilcelulozo (CM)-celulaze iniţiază atacul aleatoriu la multiple situsuri
din interiorul regiunilor amorfe ale fibrei celulozice, deschizând astfel calea
celobiohidrolazelor. Celobiohidrolazele, deseori denumite exoglucanaze, formează
componenta majoră a sistemului celulazic fungic, reprezentând aproximativ 40-70% din
proteina celulazică totală şi se caracterizează prin faptul că hidrolizează celuloza
cristalină. Celobiohidrolazele indepărtează mono- şi dimeri de la capătul lanţului
celulozic. β-glucozidaza hidrolizează dimerii şi în anumite cazuri, celo-oligozaharide la
glucoză. În general, endoglucanazele şi celobiohidrolazele acţionează în mod sinergic,
dar detaliile mecanismelor lor de acţiune încă nu sunt cunoscute. Se pare că
microorganismele au mai multe variante distincte de endo- şi exoglucanaze. Ca şi
hemicelulazele, celulazele sun proteine formate din mai multe domenii. Definiţia curentă
şi clasificarea “adevăratelor” celulaze este încă incertă [3].

Figura 2. Degradarea enzimatica a celulozei
1.3. Hemicelulazele
Hemicelulazele, ca multe alte enzime care hidrolizează polizaharide, respectiv
hemiceluloza, din peretele celular al plantelor, sunt proteine alcătuite din mai multe
domenii.
Hemiceluloza este un polimer complex, în componenţa căruia predomină
molecule de glucide cu 5 atomi de carbon, în special xiloză şi arabinoză şi care mai
conţine şi hexoze (în special manoză) şi câţiva acizi glucidici (figura 2).
Figura 3. Structura hemicelulozei

Hemicelulazele conţin în general module catalitice şi necatalitice structural
distincte. Cele mai importante module necatalitice constau în domenii de legare a
carbohidraţilor (CBD) care facilitează fixarea acestora, liganzi interdomenii şi module
dockerin care mediază legarea domeniului catalitic al enzimei, prin interacţii de coeziune-
dockerin, fie de suprafaţa celulei microbiene, fie de complexe enzimatice precum
celulozomul Shallom şi Shoham. În funcţie de secvenţa de aminoacizi sau de acizi
nucleici specifică modulelor lor catalitice, hemicelulazele sunt fie glicozid-hidrolaze
(GHs), fie carbohidrat esteraze (CEs)[4]. Acestea din urmă hidrolizează legăturile
esterice ale acetatului sau ale esterilor acidului ferulic şi, în funcţie de omologia secvenţei
lor primare, ele au fost grupate în diferite familii.
În funcţie de specificitatea de subtract, hemicelulazele se împart în:
- xilanaze: hemicelulaze majore care ataca legăturile β-1,4-glicozidice din
scheletul xilanului → xilooligomeri;
- β-xilozidaze: hidrolizeaza xilooligomerii la xiloza;
- xilozidaz-arabinozidaze: GHs bifunctionale, care actioneaza asupra legaturilor
dintre xiloza si arabinoza (Lee şi colab., 2003) si au o secventa primara asemanatoare cu
unele dintre β-xilozidaze;
- β-mananaze: hidrolizează hemicelulazele pe bază de manan → β-1,4-mano-
oligomeri mici;
- β-manozidaza: hidrolizeaza β-1,4-mano-oligomeri la manoză
-α-L-arabinofuranozidaze si α-L-arabinanaze: hidrolizează arabinofuranozil-
hemicelulozele; unele au o specificitate largă de substrat, acţionând asupra uneia din
legăturile O-5, O-2 si/sau O-3 ale arabinofuranozidelor ca si supra legăturilor O-2 şi O-3
din xilanii dublu substituiţi;
- alte xilanaze si α-D-glucuronidaze: hidrolizează legătura α-1,2-glicozidică din
catena laterală a xilanilor, formată cu acid 4-O-metil-D-glucuronic
- acetil esteraze: hidrolizează substituenţii acetil de pe molecula xilozei
- feruloil esteraze: hidrolizează legătura esterică dintre substituenţii arabinozei si
acidul ferulic. Feruloil esterazele contribuie la ruperea legaturilor dintre hemiceluloza si

lignină, polizaharidulul devenind accesibil biodegradării ulterioare de către glicozid-
hidrolazele specific[5].
Figura 4. Enzime implicate in degradarea hemicelulozei
O clasificare comprehensivă a hemicelulazelor şi a altor hidrolaze este disponibilă
pe site-ul http:/afmb. cnrs-mrs.fr/CAZY.
1.4. Microorganisme producătoare de enzime hidrolitice de uz furajer
O preocupare permanentã a cercetãtorilor din domeniul biotehnologiilor o
constituie ridicarea potenţialului productiv al tulpinilor implicate în procesele de
biosinteza. Este cunoscut faptul cã rentabilitatea bioproceselor depine în mare masurã de
potenţialul productiv al suşelor producãtoare. Mãrirea gradului de eficacitate a
tehnologiilor de biosintezã se realizeazã astãzi pe scarã largã, prin utilizarea tehnicilor de
recombinare geneticã.

În plus, în ultima perioadã, ca tulpini producãtoare de enzime, sunt utilizate tot
mai des mutante, microorganisme modificate genetic, capabile sã sintetizeze
biocatalizatorul cu o ratã crescutã, într-un timp mai scurt şi pe medii de culturã cât mai
ieftine. Tehnicile utilizate în scopul schimbãrii structurii genetice a microorganismelor
constau în mutaţii şi selecţii tehnice recombinare in vitro şi tehnici de inginerie geneticã.
Datele recente din literaturã indicã obţinerea de enzime hidrolitice în special cu
ajutorul fungilor de Aspergillus niger, studiindu-se posibilitatea îmbunãtãţirii procesului
de fermentaţie sau selecţia de noi tulpini producãtoare.
Fungii apartinând genului Aspergillus sunt alãturi de cei din genul Penicillium,
cele mai importante microorganisme folosite în biotehnologiile de obţinere a enzimelor.
Fungii filamentoşi invadeaza substraturile solide, secretã enzime degradative conducând
în cele din urma la descompunerea şi transformarea substratului pe care se dezvoltã.
Din punct de vedere taxonomic, fungii, microorganisme eucariote, pot fi
încadrate dupã clasificarea datã de A.L. Demain şi N.A. Solomon (1985) în Biology of
Industrial Microorganisms [6]:
− Diviziunea Anastigomycotina
− Subdiviziunea Deuteromycotina
− Clasa Deuteromycetes
− Subclasa Hypomycotidae
− Ordinul Moniliales
− Familia Monialiaceae
− Genul Aspergillus
Majoritatea speciilor din genul Aspergillus prezintã miceliu septat, au un ciclu de
viatã sexuat şi sunt denumiţi în mod obişnuit fungi imperfecţi. Mai pot fi încadrati şi la
fungii perfecţi datorita prezenţei ciclului de viaţã parasexual. Acest ciclu ambiguu
asemãnãtor cu cel sexual, în special în etapa de recombinare a materialului genetic, cu
specificaţia cã recombinarea nu are loc în meioza, ci în cursul deviziunii mitotice.
Utilizarea mucegaiurilor pentru producerea de enzime hidrolitice prezintã o serie
de avantaje:
− avand o structurã filamentoasã pot fi separate mai usor din mediul de culturã faţã
de drojdii şi bacterii;

− au un conţinut mult mai mic de acizi nucleici decât bacteriile şi drojdiile;
− pot asimila din substrat oligozaharidele, care nu sunt accesibile drojdiilor.
Multe dintre microorganismele folosite produc aceste enzime extracelular, ceea ce
usureazã foarte mult condiţiile de prelucrare industrială pentru obţinerea de enzime.
Mucegaiurile pot fi cultivate pe substraturi lichide sau solide bogate în glucide,
cele mai importante genuri fiind: Aspergillus (Aspergillus niger, Aspergillus oryzae),
Trichoderma (Trichoderma viridae, Trichoderma reesei, Trichoderma koningii),
Fusarium, Penicillium si Rhizopus.
Carbohidrolazele şi enzimele celulozolitice derivate din Aspergillus niger pot fi
utilizate în siguranţa ca aditiv în hrana, în conformitate cu urmãtoarele condiţii:
(a) tulpina Aspergillus niger nu este toxicã sau patogenicã pentru oameni sau animale;
(b) aditivul nu este folosit în exces, fiind necesare cantitãţi minime pentru a produce
efectul dorit.
A fost stabilitã şi standardizatã o metodã de biosintezã a enzimelor celulozolitice
cu o tulpinã de Aspergillus niger IBT-90 [7].
În afarã de enzimele celulozolitice, fungul mai produce cantitãţi considerabile de
hemicelulaze, ca şi de enzime pectinolitice, 1,3-glucanaze, β-amilaze, proteinaze şi
ligninperoxidaze. Studiile asupra activitãţii enzimelor celulozolitice au evidenţiat 2-endo-
β-1,4-glucanaze si anume celobiohidrolaze si β-glucanaze. Aceste doua carbohidrolaze au
fost izolate şi au fost determinate proprietãţile lor catalitice şi moleculare, incluzand
temperatura, pH-ul optim şi greutatea molecularã. Enzimele celulozolitice derivate din
Aspergillus niger IBT-90 par sã atace celuloza într-un mod sinergic.
Endo-β-1,4-xilanaza hidrolizeazã arabinoxilanul, un constituent major al
peretelui celular al cerealelor şi de curand aceastã enzimã este utilizatã în mare mãsura în
procesele biotehnologice.
În industria alimentarã xilanazele au fost propuse pentru modificarea celulozei şi
hemicelulozei prin hidroliza parţialã. Hidroliza totalã a celulozei în glucozã, se poate
finaliza cu cu fermentaţia glucozei la etanol, izopropanol sau butanol.
Arabinofuranozidazele sunt enzime care scindeazã substituienţii arabinozici din
hemiceluloze (arabinoza la arabinoxilani, arabinani şi arabinogalactani). A fost

secvenţializatã parţial şi purificatã arabinofuranozidaza din Aspergillus niger crescut pe
mediul de cultura cu pulpa de sfecla de zahar/tãraţe de grâu.
Procentul de umiditate iniţial, timpul de cultivare, mãrimea inoculului şi
concentraţia mediului de bazã au fost optimizate în vederea producţiei de xilanaze de
cãtre Aspergillus niger prin fermentaţie în stare solidã. S-a demonstrat cã timpul de
cultivare şi concentraţia mediului de bazã reprezintã factorii cei mai importanţi care
afecteazã activitatea xilanazicã. Mãrimea inoculului de 52x105 spori/g, umiditatea de
65%, timpul de cultivare de 5 zile şi de 10 ori mai concentrat mediul bazal care conţine
50% extract de grâu, au reprezentat parametrii optimi pentru producţia de xilanazã prin
fermentaţie în stare solidã. Activitatea şi producţia de xilanazã dupã 5 zile de fermentaţie
utilizând ca sursã de carbon paie de orez a fost de 5,071 IU/g respectiv 14,790 IU I-1h-1.
Activitatea xilanazicã estimatã prin modelul polinomial a fost de 5,484 IU/g de paie de
orez [8].
Tulpina de Aspergillus niger 44 , cunoscutã ca producãtoare de xilanazã, a fost
studiatã de cãtre P.V. Gawande si M.Y. Kamat [9] pentru obţinerea de xilanazã (E.C.
3.2.1.8) prin fermentaţie în stare solidã, pe diferite substraturi lignocelulozice. Ca substrat
lignocelulozic utilizat pentru producţia de xilanazã cel mai bun a fost tãrâţele de grâu.
Mediul optim pentru Aspergillus niger a fost: tãrâţe de grâu în proporţie 1:5 cu solutţe
mineralã Mandels şi Strenberg (continând 0,1% extract de drojdie) la 35ºC şi
concentraţia inoculului fiind de 2x107-2x108 spori/5g substrat. Dupã 4 zile de incubare,
activitatea xilanazicã a fost de 74,5 IU/ml. Cultura filtratã proaspãtã a fost utilizatã pentru
hidroliza diferitelor deşeuri lignocelulozice. În timpul zaharificãrii, preparatele xilanazice
au îndepãrtat fracţia hemicelulozicã din toate deşeurile lignocelulozice testate,
accentuând potenţialul comercial al producţiei enzimelor xilanazice. Aspergillus sp. a
produs xilanaza cu un nivel scazut al activitãţii celulozolitice, facilitând un sistem de
purificare al enzimelor relativ simplu[9].

Capitolul 2. Utilizarea deşeurilor şi/sau subproduselor agricole în medii de biosinteză a enzimelor hidrolitice de uz
zootehnic
Biomasa lignocelulozicã (în special deşeurile agricole) este cunoscutã ca fiind o
excelentã sursã de carbon pentru productia enzimelor microbiene. Biomasa
lignocelulozicã constituitã din reziduuri agricole (paie, coceni de porumb), unele
subproduse ale industriei alimentare (pulpe epuizate de fructe, tãiţei de sfecla de zahãr
epuizaţi), furajele şi lemnele neutilizabile reprezintã o rezervã şi insuficient valorificatã
de glucide fermentescibile. În ultimul deceniu s-au făcut eforturi considerabile în privinţa
valorificării superioare a biomasei lignocelulozice(BLC). Celuloza, împreună cu
hemiceluloza şi lignina intra în structura pereţilor celulelor vegetale. Pentru separarea
celulozei din complexul lignocelulozic se aplică tratamente mecanice şi cu vapori de apă.
Substratul pe care se cultivă fungii se obţine în urma hidrolizei în mediu acid, bazic sau
enzimatic a deşeurilor celulozice [10].
Reziduurile agro-industriale sunt în general considerate ca cele mai bune
substraturi pentru obţinerea de enzime prin fermentaţie în stare solidă. Marele avantaj al
acestor surse naturale de carbon este că sunt disponibile în cantităţi industriale şi că au un
preţ scăzut.
Câteva substraturi cum ar fi tărâţe de grâu, tărâţe de orez, ştiulete de porumb, paie
de grâu, paie de orez, coajă de orez, păstaie de soia, reziduuri de banană, resturile de
plante medicinale, cocean de porumb,pulpă de sfecla de zahăr, etc. sunt utilizate ca surse
de carbon în producţia de enzime. Dintre acestea, tărâţele de grâu sunt cel mai frecvent
utilizate în diferite procese.
S-a constatat că fungul Aspergillus niger 38 poate fi cultivat prin fermentaţie în
strat solid pentru obţinerea celulazelor folosind ca sursa de carbon deşeuri agricole uşor
de procurat cum ar fi tărâţele de grâu şi paiele de grâu în diferite proporţii. Mediul de
cultură (g/l) a avut urmatoarea compoziţie: 10 (NH4)2SO4; 3 KH2PO4; 0,5 MgSO4x 7H2O;
si 0,5 CaCl2xH2O.

Tărâţele de grâu şi paiele de grâu s-au folosit în proporţii diferite (9:1 la 1:9). Cu
o umiditate de 74%, un domeniu al pH-ului de fermentaţie cuprins între 4.5-5.5 si un
raport paie de grâu tărâţe de grâu de de 9:1, s-a obţinut o activiate endonucleazică de
14.80 UI/ml (unităţi internaţionale/ml). Raportul o parte paie de grâu şi nouă părţi tărâţe
de grâu a prezentat cea mai bună activitate.
Alti autori [11] au folosit fungul Trichoderma reesei ZU-0 2 pentru obţinerea de
celulază prin fermentatie în stare solidă, utilizând ca substrat reziduuri agricole: ştiulete
de porumb, deşeuri lignocelulozice din industria xilozei. Mediul de cultură pentru
cresterea microorganismului Trichoderma reesei ZU-02 a avut urmatoarea compoziţie
(%): ştiulete de porumb 66; tărâţe de grâu 30; (NH4)2SO4 2; uree 0.5; KH2PO4 0.5;
MgSO4x7H2O 0.5; CaCl2xH2O 0.45; CoCl2 0.05. Substratul solid a fost reutilizat de cel
puţin trei ori în sistem”batch” şi s-a constatat că activitatea celulozolitică cea mai mare a
fost obţinută (158 IFPU/g koji) în a doua fermentaţie batch. Enzimele koji produse prin
acest proces au putut fi utilizate direct în hidroliza ştiuleţilor de porumb. Când doza de
celulază este peste 20 IFPU/g substrat, producţia de zahăr este peste 84%.
Cinci tulpini de fungi filamentoşi (tulpinile Aspergillus niger NRRL 3122 şi
T00050007-2; Aspergillus oryzae CCT3940; Aspergillus awamori NRRL3112 şi
Trichoderma sp.) au fost comparate pentru a vedea capacitatea lor de a produce
endopoligalactouranaze în fermentaţie în stare solidă. Tărâţele de grâu au fost utilizate ca
suport şi ca unică sursă de carbon peste care s-au adăugat 45 ml soluţie de săruri minerale
conţinând (%): KH2PO4 0.2; MgSO4 0.1; (NH4)2SO4 0.4; FeSO4 6,3x10-4; MnSO4 1,0x10-
4 si ZnSO4 6,2x10-4 pentru fiecare 100g tărâţe, pentru a produce mediul bazic de tărâţe de
grâu. Activitatea pectolitică a fost atinsă după 72 de ore de creştere cele mai bune tulpini
de fungi fiind Aspergillus niger T00050007-2 şi Aspergillus oryzae CCT3940. Trei tipuri
de pectine comerciale purificate şi patru de pectine neprelucrate au fost folosite pentru a
evalua efectul pectinei asupra producţiei de endopoligalacturonase de Aspergillus niger
T00050007-2. Maximul activităţii pectolitice a fost atins folosind 6 şi 10 % pectină
purificată ca inductor. În funcţie de originea pectinei comerciale folosită ca inductor,
nivelele maxime endopoligalacturonase au variat de la 223-876 unităţi/gram pe mediu
uscat indicand că trebuie să se aibă grijă când se alege acest component al mediului[12].

Când pectinele crude au fost folosite ca inductori la aceeasi concentraţie ca
pectina purificată, maximul activităţii pectolitice a fost 250-300 unităţi/gram. Totuşi, prin
creşterea procentului de până la 50%, maximul endopoligalacturonazei a fost de 919
unităţi/gram, deschizând astfel posibilitatea scăderii costului mediului pentru producerea
de endopoligalacturonaze.
Resturile de grâu, grăunţele uscate şi silozul de porumb sunt bioproduse ale
prelucrării porumbului şi grâului având un conţinut scăzut de proteină şi un conţinut
ridicat de fibre crude. Aceşti doi factori limitează utilizarea lor în hrana păsărilor şi
porcilor, sector care constituie un mare consumator de furaje comerciale[13]. Este absolut
necesar să se sporească valoarea nutritivă a acestor bioproduse prin desfacerea
polizaharidelor neamidonoase. Aceast proces este realizabil prin utilizarea enzimelor
microbiene degradative din fungi. Fungii au capacitatea de a produce numeroase enzime.
Aspergillus niger, Aspergillus flavus si Penicillium sp. sunt menţionate ca fiind cele mai
bune surse de celulaze, amilaze, hemicelulaze, catalaze, pectinaze şi xilanaze. Aceste
enzime ajută la degradarea polizaharidelor neamidonoase din substrat până la glucide
solubile. Aspergillus niger a prezentat cel mai mare procent de reducere al celulozei
datorită creşterii sale mai rapide şi prin urmare abilităţii de a produce mai multe enzime
celulozolitice într-o perioadă mai scurtă. Au fost determinate modificări în proteină,
glucide şi celuloză a trei bioproduse agro-industriale prin fermentare în stare solidă cu
Aspergillus niger, Aspergillus flavus şi Penicillium sp . Celuloza a fost redusă
semnificativ în toate deşeurile agro de către toţi fungii după 14 zile. Reducerea
procentuală cea mai mare a fost realizată de Aspergillus niger în toate deşeurile agro:
36,51% pentru deşeuri de grâu; 35,87% pentru grăunţe uscate şi 35,80% pentru siloz de
porumb. După 14 zile nivelul zahărului a început să scadă şi nu a mai existat o degradare
semnificativă a celulozei. Protenia totală din deşeurile agro-industriale a crescut
semnificativ. După 14 zile creşterea procentuală cea mai mare în proteină (41%) a fost
obţinută din deşeuri de grâu inoculat cu Aspergillus niger. Rezultatele acestui studiu au
indicat posibilitatea sporirii valorii nutriţionale a acestor bioproduse printr-o tehnică
simplă, necostisitoare şi uşor de adaptat şi utilizarea acestora în hrana păsărilor şi porcilor
[14].


Capitolul 3.Caracteristici ale proceselor de fermentatie cu microorganisme
producatoare de enzime hidrolitice utilizabile ca aditivi furajeri
Începand din anul 1970, au fost iniţiate studii privind obţinerea de proteaze acide
cu mucegaiuri din genul Aspergillus (A. niger, A. oryzae) prin cultivare în sistem submers
sau SSF. Preparatele enzimatice obţinute sunt complexe, conţin pe lângă proteaze şi
amilaze (α-amilaza şi glucoamilaza), celulaze, xilanaze, pectinaze. În cazul culturilor
submerse, comparativ cu procedeul de cultivare în sistem SSF, randamentele de obţinere
a preparatului enzimatic sunt mai scăzute [10].
Alături de tulpina microbiană şi compozţtia mediului de cultură, printre factorii
importanţi care influenţează procesul de biosinteză a enzimelor hidrolitice se numără
aeraţia, temperatura şi pH-ul mediului de biosinteză, precum şi gradul de agitare [15].
P.V. Gawande şi M.Y. Kamat au studiat influenţa temperaturii de cultivare asupra
obţinerii de xilanază (E.C. 3.2.1.8) prin fermentaţie în stare solidă pe diferite substraturi
lignocelulozice, cu 2 tulpini de Aspergillus sp. (A.terreus si A. niger 44), cunoscute ca
producătoare de xilanază. Tărâţele de grâu s-au remarcat ca cel mai bun substrat
lignocelulozic utilizat pentru producţia de xilanază. Mediul optim pentru Aspergillus
niger a fost: tărâţe de grâu în proporţie 1:5 cu soluţie minerală Mandels şi Strenberg
(conţinând 0,1% extract de drojdie), concentraţia inocului fiind de 2x107-2x108 spori /5 g
substrat. După terminarea procesului fermentative, s-a constatat că maximul producţiei de
xilanază, pentru ambele tulpini de Aspergillus, a fost atins la temperatura de 35°C.
Anterior zaharificării, preparatele xilanazice au îndepărtat fracţia hemicelulozică din toate
deşeurile lignocelulozice testate, dovedintu-şi astfel potenţialul commercial. Aspergillus
sp. a produs xilanază cu un nivel scăzut al activităţii celulozolitice, facilitând un sistem de
purificare al enzimelor relativ simplu.
Luiza Jecu [16] a constatat că fungul Aspergillus niger 38 poate fi cultivat prin
fermentaţie în strat solid, pentru obţinerea celulazelor folosind ca sursă de carbon deşeuri
agricole uşor de procurat cum ar fi tărâţele de grâu şi paiele de grâu în diferite proporţii.

Mediul de cultură (g/l) a avut urmatoarea compoziţie: 10 (NH4)2SO4;; 3 KH2PO4;; 0,5
MgSO4x7H2O; si 0.5 CaCl2xH2O. Tărâţele de grâu şi paiele de grâu s-au folosit în
proporţii diferite (de la 9:1 la 1:9). Cu o umiditate de 74% a substratului conţinând paie
de grâu/tărâţe de grâu în proporţie de 9/1, s-a obţinut o activitate endonucleazică de 14.80
UI/ml (unităţi internaţionale/ml), optimă, la un pH de 4.5-5.5. Raportul o parte paie de
grâu şi nouă pări tărâţe de grau a prezentat cea mai bună activitate.
Temperatura mediului în care are loc procesul de biosinteză a enzimelor
hidrolitice este un factor extrem de important pentru activitatea microorganismelor.
Variaţiile de temperatură au efect asupra randamentului de transformare a substratului în
produsul finit, asupra cerinţelor nutritive ale microorganismului şi compoziţiei biomasei
obţinute şi, nu în ultimul rând, asupra vitezei de creştere microbiană. Temperatura este un
factor care acţionează în mod direct asupra microorganismului viu. Temperaturile ridicate
pot dăuna microorganismelor prin denaturarea enzimelor, a proteinelor transportatoare,
sau a altor tipuri de proteine[17].
Valoarea pH-ului este, alături de temperatură, un parametru important în
procesele de biosinteză. Fiecare specie are definit un anumit interval de pH în care poate
creşte, precum şi un pH optim de dezvoltare. Influenţa valorii pH-ului asupra dezvoltarii
culturilor microbiene poate fi urmarită în două direcţii principale si anume : asupra
vitezei de creştere a microorganismelor şi asupra randamentului de conversie a
substratului la produsul polienzimatic. În general, microorganismele au un domeniu
optim de pH pentru dezvoltare, în care viteza specifică de creştere atinge valoarea
maximă. În cazul fungilor, pH-ul optim de dezvoltare este cuprins între 4,5-6,5. Fungii
prezintă avantajul de a creşte convenabil în medii de cultură acide, în care cele mai multe
specii de bacterii nu se dezvoltă. Formarea produsului dorit în urma procesului poate să
fie legată de desfaşurarea bioprocesului într-un domeniu foarte strict de pH. În timpul
dezvoltării unei culturi microbiene apar deviaţii ale pH-ului de la valoarea considerată
optimă, care pot avea urmări nedorite asupra procesului de biosinteză. Aceste modificări
se pot datora fie consumării unui nutrient, fie producerii unui acid organic de către
microorganism. Pentru corectarea pH-ului la valoarea prescrisă se folosesc diverse
substanţe chimice (acizi sau baze)[18].


PARTEA A II A
CERCETĂRI EXPERIMENTALE
Capitolul 4. Materiale şi metode
4.1 Tulpini de lucru
În vederea obţinerii unui complex enzimatic hidrolitic (amilaze, proteaze,
celulaze, xilanaze) s-au utilizat tulpini de fungi din Colectia de Microorganisme a
Facultatii de Biotehnologii – USAMV Bucureşti si din Colecţia de Microorganisme a
Facultăţii de Biologie a Universităţii Bucureşti, prezentate in continuare:
Denumire tulpină SursaAspergillus niger CBM-1 Facultatea de Biotehnologii – USAMV
BucureştiAspergillus niger CMGB-401 Facultatea de Biologie - Universitatea
BucureştiTrichoderma viride CMGB-405 Facultatea de Biologie - Universitatea
Bucureşti.
4.2. Obţinerea culturilor de intreţinere
4.2.1. Medii de întreţinere
Înainte de utilizare, tulpinile de Aspergillus niger CBM-1, Trichoderma viride
CBGM-405, Aspergillus niger CMGB-405 au fost activate prin treceri succesive pe mediu
înclinat Czapek-Dox simplu şi pe mediul Czapek-Dox - extract de malţ.
1. Mediul Czapek-Dox (g/l) :
- zaharoză- 30,0g ;
- NaNO3- 3,0g ;
- MgSO4- 0,5g ;

- KCl- 0,5g
- FeSO4- 0,01g ;
- K2HPO4-1,0g ;
- Agar- 2,0g ;
- Apă distilată-1000ml.
2. Mediul Czapek-Dox- extract de malţ (g/l) :
- Extract de malţ 20,0 g ;
- Componentele mediului Czapek-Dox.
După preparare, mediul s-a repartizat în eprubete şi s-a sterilizat la 110ºC timp
de 30 de minute, pH-ul mediului fiind de 6,4 corectat cu NaOH 40%.
4.2.2. Conservul vegetativ
După verificarea sterilităţii mediului prin menţinerea sa la termostat la 37ºC, timp
de 48 ore, acesta s-a însămânţat cu o suspensie de spori în apă distilată.
Pentru obţinerea inoculului pe mediu solid s-a folosit următoarea tehnică de
lucru : peste cultura de întreţinere bine sporulată s-a pipetat 5ml apă distilată sterilă, se
raclează foarte bine pentru desprinderea sporilor, obţinându-se o suspensie de spori[19].
Eprubetele cu mediu însamanţat s-au incubat la 28-30ºC timp de 7 zile. La
sfarşitul perioadei de incubare, eprubetele s-au examinat privind gradul de sporulare al
culturii, precum şi puritatea acesteia. Tuburile corespunzătoare au constituit cultura de
întreţinere. După această perioadă sporii se pot utiliza imediat sau se pot conserva la
frigider la +4ºC.
4.3. Cultivarea în laborator şi testarea activităţii hidrolitice
La nivel de laborator, lucrările s-au axat pe studiul diferitelor medii de cultură
în vederea stabilirii unui mediu optim pentru dezvoltarea microorganismului selecţionat
şi pentru biosinteza enzimelor hidrolitice precum şi pe stabilirea parametrilor de
bioproces optimi.
Au fost testate mai multe medii de cultura având următoarele compoziţii:

Mediu 1 (g/100ml) Mediu 2 (g/100ml)
Făină de soia -2% Amidon solubil-2%
Amidon solubil-1% Făină de soia -0,5%
Tărâţe de greu-0,5% CaCl2-0,1%
KH2PO4 -2,5% KH2PO4-0,1%
CaCl2-0,1% MgSO4x7H2O-0,01%
MgSO4x7H2O-0,01%
Mediu 3 (g/100ml) Mediu 4 (g/100ml)
Amidon solubil -1% Făină de porumb-2%
Peptonă-0,5% Făină de soia -3%
Extract drojdie -0,5% (NH4)2HPO4-0,5%
KH2PO4 -0,1% MgSO4x7H2O- 0,05%
MgSO4x7H2O -0,02% CaCl2-0,1%
CaCl2-0,1%
După preparare, mediile de cultură sterilizate şi răcite la 35ºC, s-au însă mânţat cu
o cultură de inocul sub formă de suspensie de spori obţinută din cultura de întreţinere.
Raportul de însamanţare este de 0,5 ml suspensie pentru 50 ml mediu. Cultivarea s-a
efectuat 72 ore la baloane agitate(220 rpm) cu 50 ml mediu de cultura/balon. Incubarea
s-a făcut la termostat la 28-30ºC.
Pentru urmărirea bioprocesului, la intervale de 24 ore s-au determinat următorii
parametri:

- pH-ul mediului de cultura;
- activitatea enzimatică a mediului ;
- dezvoltarea macroscopică si microscopică a tulpinii de la care s-a pornit.
4.4. Determinarea activităţilor enzimatice
4.4.1. Determinarea activităţii α-amilazice (E.C.3.2.1.1)
Pentru dozarea activiăţii α -amilazei produsă de fungi s-a aplicat metoda descrisă
de Hostettler şi colaboratorii [20] metodă care se bazează pe hidroliza enzimatică a
amidonului la pH=6,9 si la temperatura de 30ºC. Hidrolizatul format în special din
maltoză reactionează, datorită grupărilor reducătoare semiacetalice, cu acidul 3,5-
dinitrosalicilic, cu formare de acid nitroaminosalicilic de culoare roşie-portocalie, cu
maximum de adsorbţie la 546nm.
Concentraţia de acid diaminosalicilic format, măsurată colorimetric, este
proporţională cu activitatea enzimatică.
Conform acestei metode o unitate amilazică corespunde unui mmol de maltoză
eliberat de 1g preparat enzimatic într-un minut la 30ºC.
Reactivi necesari :
1. Soluţie tampon fosfat 0,2M,Ph=6,9 ;
7,16g Na2HPO4 x 12 H2O + 2,72g KH2PO4 + 0,58g NaCl se dizolvă şi se aduc la 1 litru
soluţie cu apă distilată. Valoarea de pH se verifică cu pH-metrul.
2. Soluţie amidon 1% in solutie tampon(1) ;
3. Reactiv acid dinitrosalicilic (DNS).
10g acid dinitrosalicilic se dizolvă la cald în aproximativ 300ml apă distilată. Se adaugă
400ml soluţie NaOH 1N şi 300g tartrat de sodiu şi potasiu. Se aduce la semn într-un
balon cotat de 1 litru.
4. Soluţie etalon de maltoză ;

0,2g maltoză monohidrat (0,19g maltoză anhidră) se dizolvă în apă îtr-un balon cotat de
100ml.
5. Soluţie probă de determinat.
Proba se dizolvă cu CaCl2, astfel încât să conţină 0,5-1,5 unităţi DNS.
Mod de lucru
Construirea curbei de etalonare
Din soluţia etalon de maltoză(4) se pipetează în 6 eprubete după cum urmează :
Tabelul 2. Prepararea diluţiilor de maltoză
Număr eprubetă Vol. soluţie maltoză (ml)
Vol. apă distilată (ml)
Conc. maltoză (µmoli)
1 - 2,1 -2 0,4 1,7 2,243 0,8 1,3 4,484 1,2 0,9 6,725 1,6 0,5 8,966 2,0 0,1 11,2
În fiecare eprubetă se adaugă 2ml DNS, apoi se tine în baie de apă la fierbere 5
minute.
Se răcesc şi se diluează pana la 12 ml cu apă distilată.
Se citeşte densitatea optică la 546nm dupa 20 de minute.
Datele oţinute se trec într-un grafic pentru construirea curbei de etalonare. Pe
abcisă se trec valorile reprezentând densitatea optică citită la spectrofotometru
corespunzătoare concentraţiei de maltoză înscrisă pe ordonată.
Dozarea activităţii enzimatice
Paralel cu proba de analizat se face o probă martor pentru a determina
concentraţia de maltoză existentă în probă, înainte de începerea reacţiei enzimatice.
Modul de lucru este prezentat schematic în tabelul 2.
Tabelul 3. Reprezentarea schematică a modului de lucru pentru dozarea activităţii

α-amilazice
Proba de analizat0,5ml soluţie tampon(1)
1ml soluţie amidon 1%(2)0,5ml proba
-agitare-
Proba martor0,5ml soluţie tampon(1)
1ml soluţie amidon 1% (2)2ml DNS(3)
–agitare0,5ml proba
Incubare 10 minute la 30ºC2ml DNS -
Incubare 5 minute pe baie de apă.Răcire.
Diluare până la 12ml cu apă distilatăCitirea valorilor absorbanţei la spectrofotometru la 546 nm
Calculul rezultatelor
Activitatea enzimatică se calculează şi se exprimă după formula :
Unităţi DNS/ml = mxD/vx10
unde :
- m = µmoli de maltoză, determinaţi din curba de etalonare ca fiind diferenţa dintre
µmoli de maltoză din probă şi µmoli de maltoză din martor ;
- D = factorul de diluţie al probei luate în lucru ;
- V = volumul (ml) de soluţie probă luată în lucru ;
- 10 = timpul de incubare (minute).
4.4.2. Determinarea activităţii celulazice
Principiul metodei
Determinarea activităţii celulozolitice se efectuează dupa metoda descrisă de
Petterson şi Porath [21].
Metoda se bazează pe dozarea glucidelor reducătoare eliberate de enzimă în urma
acţiunii asupra substratului respectiv carboximetil celuloza (CMC).
Reactivi şi soluţii:
1. Soluţie tampon Na2HPO4- acid citric 0,1N, pH=6,4 ;

2. Soluţie CMC 1%. 1g carboximetil celuloza se dizolvă în 100ml tampon(1) ;
3. Preparat enzimatic cu activitate celulazică ;
4. Reactiv DNS (20g acid 3,5 dinitrosalicilic, 4g fenol proaspat distilat, 1g sulfit de
sodiu, 400g tartrat de sodiu şi potasiu). Se dizolvă în 1 litru NaOH 2%, soluţia se
aduce la 2 litri cu apă distilată.
5. Solţtie glucoză 0,1% preaparată proaspăt în apă distilată.
Amestecul de reacţie, ce constă din 2ml soluţie de substrat (2), 0,2 ml preparat
enzimatic (3), este incubat 10 minute la temperatura de 50ºC, după care reacţia
enzimatică este stopată cu 3 ml rectiv DNS. Probele sunt incubate apoi 15 minute pe o
baie de apă la fierbiere, răcite şi colorimetrate la 640 nm faţă de apa distilată.
Fiecare probă este însoţită de un martor, în care dozăm glucidele reducătoare
preexistente în amestecul de reacţie prin introducerea reactivului DNS înaintea
preparatului enzimatic cu activitatea celulozolitică. Martorii sunt şi ei colorimetraţi la 640
nm faţă de apa distilată.
Calculul rezultatelor
Valorile densităţii optice măsurate la spectrofotometru pentru probe şi martori se
transformă cu ajutorul curbei etalon în mg glucoză.
(mg glucoză probă- mg glucoză martor) x5 x1/10 = U/ml/min/50ºC
Unitatea de activitate Cx este acea cantitate de enzimă care în condiţiile metodei
eliberează dintr-o solţtie de CM-celuloză o cantitate de glucide reducător, care dă cu
reactivul DNS acea densitate optică ca 1mg glucoză.
Construcţia curbei de etalonare
Din soluţia de glucoză 1,0% (5) se fac o serie de diluţii în apa distilată, la care se
adaugă reactiv DNS (câte 3 ml DNS/eprubeta cu diluţie). Probele sunt incubate 15 minute
pe o baie de apă la fierbere, răcite şi colorimetrate faţă de apa distilată.
Se citesc extincţiile probelor la 640 nm şi se trasează un grafic, notând pe abcisă
extincţia citită la spectrofotometru, iar pe ordonată mg glucoză.
Tabelul 4. Prepararea diluţiilor de glucoză
Vol. soluţie 5 (ml) Vol. apa distilată (ml) Cant. Glucoză (mg)

0,1 2,1 0,10,2 2,0 0,20,3 1,9 0,30,4 1,8 0,40,5 1,7 0,50,6 1,6 0,60,7 1,5 0,70,8 1,4 0,80,9 1,3 0,91,0 1,2 1,0
4.4.3. Determinarea activităţii proteazice
Activitatea proteolitică se determină după metoda Anson modificată [22] pe
substrat de cazeină.
Principiul metodei
Enzimele proteolitice catalizează hidroliza caseinei permiţând formarea unor
compuşi solubili în acid tricloracetic. Conţinutul în triozina şi triptofan , din aceşti
compuşi, este determinată spectrofotometric cu reactivul Folin-Ciocâlteu.
Reactivi şi soluţii:
1. Tampon fosfat 0,2 M , pH=7,0 (39,0 ml NaH2PO4 0,2M + 61,0 ml soluţie
Na2HPO4 0,2M) ;
2. Soluţie caseină 1%. 1g de caseină se dizolvă pe un agitator magnetic în cca 30
ml NaOH 1N. Se neutralizează cu H3PO4 10%, adăugat în picături sub continuă
agitare. Se aduce la pH-ul dorit şi se completează cu tampon fosfat (pH= 7în
cazul proteazelor neutre).
3. Soluţie de L- tirozină 1mM ( se dizolvă 181,19 mg de tirozină în 1000ml HCl
soluţie 0,2N) ;
4. Soluţie HCl 0,01N-0,06N-0,2N ;
5. Soluţie NaOH 0,5N ;
6. Soluţie acid tricloracetic 5% ;
7. Reactiv Folin-Ciocâlteu (se dizolvă 10 g Na2 WO4 x 2H2O + 2,5g Na2MoO4 x 2
H2O + 15 g Li2SO4 + 10 ml HCl + 5ml H3PO4 conc. şi se aduce la 100ml cu apa
distilată ). Înainte de utilizare se diluează o parte din această soluţie cu două părţi
apă distilată .
8. Soluţie enzimatică.
Mod de lucru
Amestecul de reacţie format din 0,5ml soluţie enzimatică (mediu de cultură
separat de biomasă) şi 1 ml solţtie caseină 1% în tampon fosfat 0,2 M (pH=7) se
incubează la 37º C timp de 10 minute. Reacţia enzimatică se stopează cu 2 ml acid
tricloracetic 5%. Amestecul de reacţie se menţine 30 minute la temperatura camerei şi
apoi se filtrează. La 0,5ml filtrat se adaugă 0,5 ml HCl 0,2 N, 2ml NaOH 0,5 N şi
reactiv Folin – Ciocâlteu diluat 1 :2. Se lasă 30 minute la temperatura camerei şi se
măsoară extincţia la 578nm faţă de martor. La proba martor se lucrează asemănător , cu
excepţia că în amestecul de reacţie se introduce acid tricloracetic imediat peste soluţia de
caseină, se lasă în repaus 30 minute la temperatura camerei şi apoi se introduce soluţia
enzimatică. În continuare se lucrează identic ca la proba de determinat.
Construirea curbei etalon
Se pipetează în 11 eprubete volumele de tirozină, HCl 0,2N, NaOH 0,5N,
menţionate în tabelul 4, după care, în condiţii de agitare puternică, se introduce reactivul
Folin-Ciocâlteu.
Tabelul 5. Obţinerea diluţiilor de L-tirozina
Eprubeta nr.
L-Tyr (ml) L-Tyr (µmoli)
HCl 0,2N(ml)
NaOH 0,5N(ml)
F-C(ml)
Extinctia
1 2 3 4 5 6 71 0 0 2,00 4 1,2 0,0433 0,04 0,04 1,96 4 1,2 0,0824 0,08 0,08 1,92 4 1,2 0,1255 0,12 0,12 1,88 4 1,2 0,556 0,16 0,16 1,84 4 1,2 0,227 0,20 0,20 1,80 4 1,2 0,298 0,24 0,24 1,76 4 1,2 0,329 0,26 0,26 1,72 4 1,2 0,3410 0,28 0,28 1,68 4 1,2 0,37511 0,32 0,32 1,64 4 1,2 0,45011 0,40 0,40 1,60 4 1,2 0,475

Se lasă la temperatura camerei 30 minute,apoi se citeşte extincţia la
spectrafotomertu Spekol la 578 nm , faţă de apa distilată. Se construieşte un grafic cu
extincţia citită la 578 nm pe ordonată şi µmoli L-tyr pe abcisă.
Calculul rezultatelor
Din curba de etalonare se determină µmoli tirozină corespunzători extincţiei
măsurate. Valorile citite pe grafic vor fi introduse în următoarea formulă:
(micromoli Tyr proba- micromoli Tyr martor) x 3,5/10 x 0,5 x 0,5 = micromoli Tyr/ml/min
unde:
3,5- volum total amestec ;
1/10- factorul de transformare pentru un minut de reacţie enzimatică ;
0,5- volumul de lichid cu activitate proteolitică luată în lucru ;
0,5- volumul de lichid filtrat luat în lucru ;
O unitate de activitate proteazică este definită ca fiind cantitatea de enzimă care în
condiţiile de reacţie indicate eliberează 1µmol tirozină pe minut.
4.4.4. Determinarea activităţii xilanazice
Activitatea xilanazică s-a determinat printr-o metodă combinată, având la bază
metoda ‘clasică’ a lui Miller ce foloseşte reactiv DNS [23].
Principiul metodei
Xilanaza catalizează hidroliza iar zaharurile reducătoare formate (xiloza) sunt
determinate spectrofotometric cu reactiv DNS la 540 nm.
Reactivi şi soluţii:
1. Tampon acetat de sodiu 0,05M, pH 5,3
2. Soluţie xilan (Fluka) 0,6% în tampon acetat
3. Soluţie de D-xiloza 0,0025 M în apă distilată
4. Reactiv DNS (se dizolvă 10 g acid 3,5 dinitrosalicilic în 300 ml apă distilată, se
adaugă 400ml NaOH 1N şi 300g tartrat de sodiu şi potasiu şi se aduce la 1 litru).
5. Soluţie enzimatică în tampon acetat.
Mod de lucru

În eprubetele prevăzute cu dop se introduce amestecul de reacţie format din 0,5
ml soluţie enzimatică şi 0,5ml soluţie substrat (xilan 0,6% în tampon acetat (pH=5,3), ce
se incubează pe baie de apă la 40ºC, timp de 30 de minute. Reacţia enzimatică se
stopează cu 1 ml reactiv DNS.
Amestecul de reacţie se fierbe 5 minute pe baie de apă, se aduce la 5 ml cu apă
distilată şi se măsoară extincţia la 540 nm faţă de apa distilată. La fiecare probă se face şi
un martor. La proba martor se lucrează asemănător, cu excepţia că în amestecul de reacţie
se introduce reactivul DNS imediat peste soluţia de substrat şi apoi se introduce soluţia
enzimatică. În continuare se lucrează identic ca la proba de determinat.
Construirea curbei etalon
Se pipetează în 11 eprubete volumele de soluţie de xiloză menţionate în tabelul 5,
după care se introduc 0,5 ml substrat (soluţie xilan) şi 1 ml reactiv DNS.

Tabelul 6. Obţinerea diluţiilor de D-xiloza
Eprubeta nr.
D-Xiloza (ml)
D-Xiloza (nmoli)
Apa (ml) Xilan (ml)
DNS (ml)
Extincţia
1 0 0 0,5 0,5 1,0 0,0392 0,05 250 0,45 0,5 1,0 0,04383 0,10 500 0,40 0,5 1,0 0,06314 0,15 750 0,35 0,5 1,0 0,10045 0,20 1000 0,30 0,5 1,0 0,12086 0,25 1250 0,25 0,5 1,0 0,14917 0,30 1500 0,20 0,5 1,0 0,18008 0,35 1750 0,15 0,5 1,0 0,22009 0,40 2000 0,10 0,5 1,0 0,235110 0,45 2250 0,05 0,5 1,0 0,284311 0,50 2500 0,00 0,5 1,0 0,3285
Se adaugă 3 ml apă distilată şi se lasă la temperatura camerei pentru răcirea
eprubetelor, apoi se citeşte extincţia la spectrofotometru JASCO la 540 nm , faţă de apa
distilată. Se construieşte un grafic cu extincţia citită la 540 nm pe ordonată şi nmoli D-
xiloza pe abcisă, conform diagramei următoare :
Figura 5. Curba etalon de xiloză

Calculul rezultatelor
Din curba de etalonare se determină nmoli xiloza corespunzători extincţiei
măsurate.
Valorile citite pe grafic vor fi introduse in urmatoarea formula:
(nmoli xiloza probă – nmoli xiloza martor) x 1/30= U/ml/min
unde
1/30- factorul de transformare pentru un minut de reacţie enzimatică ;
O unitate de activitate xilanazică este definită ca fiind cantitatea de enzimă care în
condiţiile de reacţie indicate eliberează 1nmol xiloza/ml/minut la 40ºC.
4.5. Testarea produsului enzimatic în hrana animalelor de fermă
Testarea produsului enzimatic în hrana animalelor de fermă a implicat stabilirea
dozei optime, evaluarea efectelor tehnico-economice, întocmirea documentaţiei de
realizare a furajului, a schemei şi a procedurii de evaluare a conformităţii [24].
Figura 6. Tineret porcin din loturile experimentale

Produsul enzimatic realizat în urma cercetărilor abordate în cadrul proiectului, a
fost verificat în cadrul unui biotest pe tineret porcin, loturile experimentale şi observaţiile
fiind realizate în condiţiile de fermă la IBNA Baloteşti. Preparatul enzimatic s-a obţinut
din culturi de Aspergillus niger şi reprezintă un amestec de enzime cu activitate
amilolitică, proteolitică, celulozolitică şi xilanayică (tabelul 7). Doza de includere în
structura reţetei de nutreţ combinat a fost de 2 respectiv 5 kg / tona de nutreţ combinat.
Tabelul 7. Caracterizarea activităţii enzimatice a preparatului enzimatic
Specificaţie ActivitateAmilolitică 105,36UDNS/gProteolitică 1,01 UP/g
Celulozolitică 1,35 U/gXilanazica 3,95 U/g
În scopul cuantificării efectului administrării preparatului enzimatic în hrana
animalelor s-au format trei loturi omogene şi s-au elaborat noi soluţii nutriţionale.
Testul biologic s-a derulat pe un număr de 54 purcei din rasa Marele Alb conform
schemei experimentale din tabelul 8. Durata experimentului a fost de 18 zile, cu
înregistrarea zilnică a consumului de nutreţ combinat. Animalele au fost cântărite la
începutul şi sfârşitul experimentului.
Tabelul 8. Schema experimentală
SpecificaţieL O T
M E1 E2Număr de animale (cap) 18 18 18
Durata experimentului (zile) 18 18 18Variabila (cantitatea de preparat
enzimatic) Kg / tona de NC* - 2 5 *NC- nutreţ combinat
A fost utilizată o receptură de nutreţ combinat unică pentru cele trei loturi,
variabila constituind-o cantitatea de enzima inclusă în premixul vitamino-mineral într-o
pondere diferită la cele două loturi experimentale (E1-2 kg /to de NC; E2- 5 kg/ to de

NC) (tabelul 8). Ca ingrediente cerealiere s-a utilizat grâul (acesta fiind caracterizat
printr-un conţinut ridicat xilani şi arabinoxilani), ponderea de includere fiind de 30% şi
porumbul, în vederea unei mai bune echilibrări energetice a reţetei.
Tabelul 9. Structura recepturilor de nutreţ combinat şi indicii calitativi ai acestora
pentru purcei
Ingrediente % LOTM E1 E2
Porumb 28,31 28,21 27,81Grâu 30,00 30,00 30,00
Şrot soia 8,00 8,00 8,00Full fat soia 10,00 10,00 10,00Lapte praf 15,00 15,00 15,00Făină peşte 4,00 4,00 4,00
Lizină 0,49 0,49 0,49Metionină 0,20 0,20 0,20
Premix colină 0,10 0,10 0,10Fosfat monocalcic 1,70 1,70 1,70Carbonat de calciu 1,10 1,10 1,10
Sare 0,10 0,10 0,10Premix vitamino-mineral P1
Enzimă1,00
-1,000,20
1,000,50
Indici calitativiProteină brută (%) 21,62 21,62 21,62
Energie metabolizabilăKcal
Mj/kg NC332013,89
332013,89
332013,89
Lizină (%) 1,50 1,50 1,50Metionină + cistină (%) 0,90 0,90 0,90
Calciu (%) 1,10 1,10 1,10Fosfor (%) 0,90 0,90 0,90

Capitolul 5. Rezultate şi discuţii
5.1. Compoziţia mediilor pentru cultura de întreţinere
În urma testării celor doua medii menţionate în capitolul 4 s-a constatat că mediul
cu extract de malţ oferă cele mai bune condiţii pentru obţinerea culturilor de întreţinere
pentru toate tulpinile testate. Astfel, pe pe suprafata acestui mediu, la temperatura optimă
de dezvoltare (28ºC), după 7 zile, se formează un miceliu bogat, alb pufos şi un miceliu
aerian purtător de conidiofori coloraţi în negru intens. Miceliul este uniform, sporulat pe
toată suprafata. Tulpina de Trichoderma viride nu s-a dezvoltat pe mediul Czapek-Dox
simplu [25].
5.2 Analiza comparativă a biosintezei enzimelor hidrolitice cu tulpini Aspergillus niger şi Trichoderma viride pe diferite medii
Pentru a evalua nivelul de biosinteză a enzimelor componente ale complexului
studiat s-a utilizat, ca termen de comparaţie, produsul KEMZYME®VP DRY, un
complex multienzimatic destinat a fi utilizat în reţetele de hrană care conţin un nivel
ridicat de cereale şi proteine vegetale (şrot de soia). Acesta permite obţinerea unor
performanţe deosebite chiar în condiţiile excluderii din alimentaţie a surselor proteice de
origine animală. Conţine surse stabilizate de endo-1,3-beta-glucanaze (E.C 3.2.1.6),
celulaze (E.C 3.2.1.4), alfa-amilaze (E.C 3.2.1.1), bacilolizin (proteaze) (E.C 3.4.24.28),
şi endo-1,4-beta-xilanaze (E.C 3.2.1.8). Activitate secundară: lipaza [25]
Tabelul 10. Caracterizarea activităţii enzimatice a produsului KEMZYME®VP DRY
KEMZYME®VP DRYActivitatea
celulozolitică(C1-Cx) U/g
Activitate amilolitică
UDNS/g
Activitateproteolitică
U/g
2,50 620 1,12

Din tabelul 11 se observă că tulpina de Aspergillus niger CBM-1 are activitate
proteolitică şi amilolitică, cele mai bune rezultate obtinându-se pe mediul de cultură M3.
De asemenea s-a constatat ca maximum de activitate amilolitică şi proteolitică s-a obţinut
la 48 ore : 12,3 UDNS/ml, 1,44 UP/ml, activitatea celulozolitică fiind considerată
satisfăcătoare, această tulpină a fost luată în considerare în experimentele ulterioare fără a
fi considerată cea mai bună.
Mediul de cultură final de fermentaţie s-a prelucrat integral prin absorbţie pe
carbonat de calciu tehnic, s-a uscat în curent de aer timp de 2 ore la 40ºC şi s-au obţinut
10g produs, care s-a caracterizat din punct de vedere al activităţii enzimatice.
Produsul enzimatic solid obţinut cu tulpina de Aspergillus niger CBM-1 prezintă
următoarele activităţi enzimatice:
Din tabelul 12, se observă că tulpina Aspergillus niger CMGB-401 aflată în
colecţia Facultăţii de Biologie, Universitatea Bucureşti este producătoare de enzime
amilolitice şi celulozolitice, rezultate bune oţinându-se la 48 de ore pe mediul de cultură
M4 (activitatea celulozolitică A.E.= 1,18 UC1-Cx/ml) şi la 72 de ore (activitatea amilolitică
A.E.= 40 UDNS/ml).
Din tabelul 13 se observă că tulpina Trichoderma viridae prezintă cea mai bună
activitate amilolitică (cuprinsă între 30-46 UDNS/ml), dar activitatea celulozolitică este de
aproximativ de 10 ori mai mică. Întrucât se urmăreşte obţinerea unui produs cu activitate
amilolitică şi celulozolitică s-a ajuns la concluzia ca cea mai potrivită tulpină pentru
scopurile noastre este tulpina de Aspergillus niger CMGB-401 pe mediul de cultură M4.
În tabelul 14 sunt redate valorile activităţii xilanazice pe probe obţinute prin
cultivarea timp de 72 de ore a tulpinilor de Aspergillus niger şi Trichoderma viridae pe
două variante de mediu: M1 (Aspergillus niger CBM-1 şi Trichoderma viridae CMGB-
401 uscate pe pat de carbonat de calciu) şi M4 (Aspergillus niger CMGB-401-cultura
finală lichidă).
Tabelul nr.11 Variaţia activităţii enzimatice hidrolitice pe parcursul biosintezei enzimelor cu tulpina de Aspergillus niger CBM-1
As
per
Me 24 ore 48 ore 72 ore
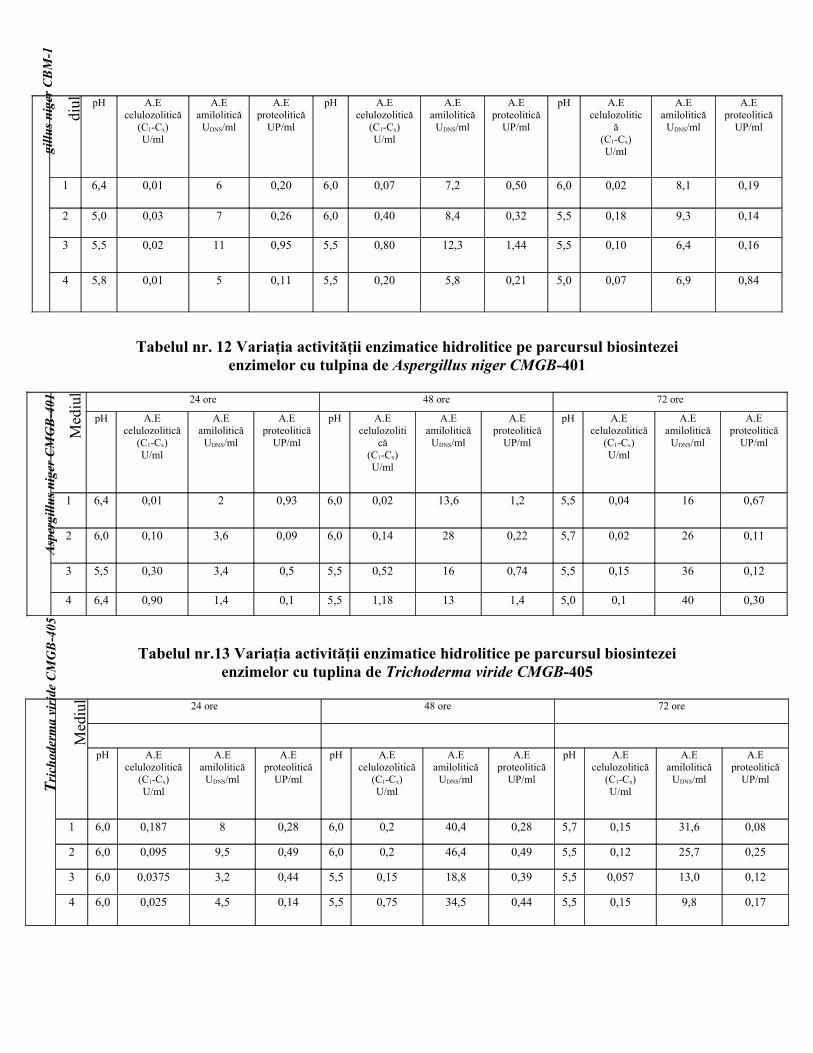
gillu
s n
iger
CB
M-1
diul pH A.E
celulozolitică(C1-Cx)U/ml
A.Eamilolitică
UDNS/ml
A.Eproteolitică
UP/ml
pH A.Ecelulozolitică
(C1-Cx)U/ml
A.Eamilolitică
UDNS/ml
A.Eproteolitică
UP/ml
pH A.Ecelulozolitic
ă(C1-Cx)U/ml
A.Eamilolitică
UDNS/ml
A.Eproteolitică
UP/ml
1 6,4 0,01 6 0,20 6,0 0,07 7,2 0,50 6,0 0,02 8,1 0,19
2 5,0 0,03 7 0,26 6,0 0,40 8,4 0,32 5,5 0,18 9,3 0,14
3 5,5 0,02 11 0,95 5,5 0,80 12,3 1,44 5,5 0,10 6,4 0,16
4 5,8 0,01 5 0,11 5,5 0,20 5,8 0,21 5,0 0,07 6,9 0,84
Tabelul nr. 12 Variaţia activităţii enzimatice hidrolitice pe parcursul biosintezeienzimelor cu tulpina de Aspergillus niger CMGB-401
Asp
ergi
llus
nige
r C
MG
B-4
01
Med
iul 24 ore 48 ore 72 ore
pH A.Ecelulozolitică
(C1-Cx)U/ml
A.Eamilolitică
UDNS/ml
A.Eproteolitică
UP/ml
pH A.Ecelulozoliti
că(C1-Cx)U/ml
A.Eamilolitică
UDNS/ml
A.Eproteolitică
UP/ml
pH A.Ecelulozolitică
(C1-Cx)U/ml
A.Eamilolitică
UDNS/ml
A.Eproteolitică
UP/ml
1 6,4 0,01 2 0,93 6,0 0,02 13,6 1,2 5,5 0,04 16 0,67
2 6,0 0,10 3,6 0,09 6,0 0,14 28 0,22 5,7 0,02 26 0,11
3 5,5 0,30 3,4 0,5 5,5 0,52 16 0,74 5,5 0,15 36 0,12
4 6,4 0,90 1,4 0,1 5,5 1,18 13 1,4 5,0 0,1 40 0,30
Tabelul nr.13 Variaţia activităţii enzimatice hidrolitice pe parcursul biosintezei enzimelor cu tuplina de Trichoderma viride CMGB-405
Tri
chod
erm
a vi
ride
CM
GB
-405
Med
iul 24 ore 48 ore 72 ore
pH A.Ecelulozolitică
(C1-Cx)U/ml
A.Eamilolitică
UDNS/ml
A.Eproteolitică
UP/ml
pH A.Ecelulozolitică
(C1-Cx)U/ml
A.Eamilolitică
UDNS/ml
A.Eproteolitică
UP/ml
pH A.Ecelulozolitică
(C1-Cx)U/ml
A.Eamilolitică
UDNS/ml
A.Eproteolitică
UP/ml
1 6,0 0,187 8 0,28 6,0 0,2 40,4 0,28 5,7 0,15 31,6 0,08
2 6,0 0,095 9,5 0,49 6,0 0,2 46,4 0,49 5,5 0,12 25,7 0,25
3 6,0 0,0375 3,2 0,44 5,5 0,15 18,8 0,39 5,5 0,057 13,0 0,12
4 6,0 0,025 4,5 0,14 5,5 0,75 34,5 0,44 5,5 0,15 9,8 0,17
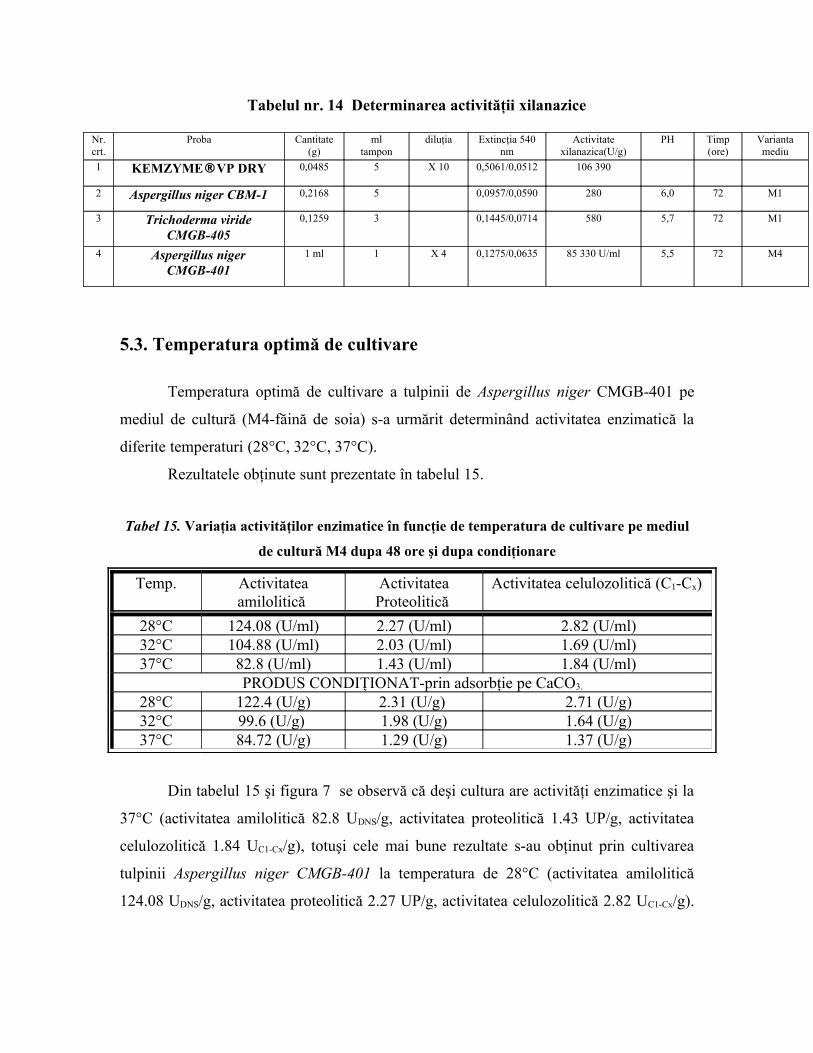
Tabelul nr. 14 Determinarea activităţii xilanazice
Nr.crt.
Proba Cantitate(g)
mltampon
diluţia Extincţia 540nm
Activitatexilanazica(U/g)
PH Timp(ore)
Variantamediu
1 KEMZYME®VP DRY 0,0485 5 X 10 0,5061/0,0512 106 390
2 Aspergillus niger CBM-1 0,2168 5 0,0957/0,0590 280 6,0 72 M1
3 Trichoderma virideCMGB-405
0,1259 3 0,1445/0,0714 580 5,7 72 M1
4 Aspergillus nigerCMGB-401
1 ml 1 X 4 0,1275/0,0635 85 330 U/ml 5,5 72 M4
5.3. Temperatura optimă de cultivare
Temperatura optimă de cultivare a tulpinii de Aspergillus niger CMGB-401 pe
mediul de cultură (M4-făină de soia) s-a urmărit determinând activitatea enzimatică la
diferite temperaturi (28°C, 32°C, 37°C).
Rezultatele obţinute sunt prezentate în tabelul 15.
Tabel 15. Variaţia activităţilor enzimatice în funcţie de temperatura de cultivare pe mediul
de cultură M4 dupa 48 ore şi dupa condiţionare
Temp. Activitateaamilolitică
ActivitateaProteolitică
Activitatea celulozolitică (C1-Cx)
28°C 124.08 (U/ml) 2.27 (U/ml) 2.82 (U/ml)32°C 104.88 (U/ml) 2.03 (U/ml) 1.69 (U/ml)37°C 82.8 (U/ml) 1.43 (U/ml) 1.84 (U/ml)
PRODUS CONDIŢIONAT-prin adsorbţie pe CaCO3.
28°C 122.4 (U/g) 2.31 (U/g) 2.71 (U/g)32°C 99.6 (U/g) 1.98 (U/g) 1.64 (U/g)37°C 84.72 (U/g) 1.29 (U/g) 1.37 (U/g)
Din tabelul 15 şi figura 7 se observă că deşi cultura are activităţi enzimatice şi la
37°C (activitatea amilolitică 82.8 UDNS/g, activitatea proteolitică 1.43 UP/g, activitatea
celulozolitică 1.84 UC1-Cx/g), totuşi cele mai bune rezultate s-au obţinut prin cultivarea
tulpinii Aspergillus niger CMGB-401 la temperatura de 28°C (activitatea amilolitică
124.08 UDNS/g, activitatea proteolitică 2.27 UP/g, activitatea celulozolitică 2.82 UC1-Cx/g).

Din tabelul 16 se observă, de asemenea, că şi activitatea xilanazică este optimă la 22
-280C, iar la 370C activitatea scade de cca 4 ori.
Tabelul 16. Variaţia activităţii xilanazice în funcţie de temperatura de cultivare a mediului
de cultură (M4-făină de soia) dupa 48 de ore
TemperaturaCantitate probă (ml)
Sol. tampon(ml)
DiluţiaExtincţia (540 nm)
Activitate xilanazică (U/ml)
220C 2 2 X 5 0,4749/0,0941 1269,33280C 2 2 X 5 0,4749/0,0941 1269,33370C 2 2 X 5 0,1978/0,0941 345,66
Concluzia este că varianta optimă de cultivare a tulpinii Aspergillus niger CMGB-
401 este cea realizată la temperatura de 28°C.
124,08
2,272,82
104,88
2,03 1,69
82,8
1,431,84
0
20
40
60
80
100
120
140 Activitatea
enzimatică (U/ml)
28°C 32°C 37°CTemperatura
Activitateaamilolitică(UDNS/ml)
Activitatea proteolitică(U/g)
Activitateacelulozolitică(UDNS/g)
Figura 7. Efectul temperaturii de cultivare a tulpinii Aspergillus niger CMGB-401 pe
mediul de cultura (M4-faina de soia) asupra activitatii enzimatice, dupa 48 ore

5.3 Alegerea pH-ului optim de cultivare
Experimentele s-au realizat în pahare Erlenmayer cu capacitatea de 250 ml şi 500
ml care conţin 100 ml mediu cu făină de soia respectiv, 250 ml mediu cu tărâţe de grâu.
Culturile au fost agitate cu ajutorul unui agitator rotativ la 220 rpm timp de 48 de ore.
pH-ul mediului de cultură – sarja 2 (M4-mediu de cultura cu făină de soia) a variat astfel:
Varianta 1: pH mediului de cultură=5.5;
Varianta 2: pH mediului de cultură =6.5;
Varianta 3: pH mediului de cultură =7.0
Tabelul 17. Variaţia activităţilor enzimatice în funcţie de pH-ul mediului de cultură
(M4) după 48 ore de fermentaţie şi după condţionare
pH Activitatea amilolitică Activitatea proteolitică Activitatea celulozolitică(C1-Cx)
5.5 141.6 (U/ml) 1.95 (U/ml) 2.91 (U/ml)6.5 108 (U/ml) 1.29 (U/ml) 2.15 (U/ml)7.5 76.8 (U/ml) 1.15 (U/ml) 1.66 (U/ml)
PRODUS CONDIŢIONAT-prin adsorbţie pe CaCO3.
5.5 105.36 (U/g) 1.08 (U/g) 1.35 (U/g)6.5 96 (U/g) 1.01 (U/g) 1.28 (U/g)7.5 77.4 (U/g) 0.7 (U/g) 1.31 (U/g)
După 48 ore, s-a constatat că pH-ul mediului de cultură în cazul primei variante a
rămas constant 5-5,5, pe când pentru celelalte variante (2 şi 3) pH-ul mediilor de cultură a
scazut de la 6.5-7.5 la 5-5.5. Ulterior, mediile de cultură au fost condiţionate integral,

înainte de sporulare, prin adsorbţie pe CaCO3. .
141.6
1.952.91
108
1.292.15
76.8
1.151.66
0
20
40
60
80
100
120
140
160Activitatea
enzimatică (U/ml)
5.5 6.5 7.5pH
Activitateaamilolitică(UDNS/mlActivitatea proteolitică(U/g)
Activitateacelulozolitică(UDNS/g)
Figura 8. Variatia activitatii enzimatice a complexului enzimatic obtinut cu tulpina
Aspergillus niger CMGB-401 in functie de pH-ul mediului de cultura (M4 faina de soia),
dupa 48 ore
Din datele prezentate în tabelul 17 şi figura 8 s-a poate observa că tulpina de
Aspergillus niger CMGB-401 prezintă cele mai bune activităţi enzimatice la pH-ul
mediului de cultură de 5.5. În urma procesului de condiţionare prin adsorbţie pe CaCO3 s-
a observat o scădere a activităţilor enzimatice aşa cum rezultă din figura 9.

141.6
105.36108
96
76.877.4
0
20
40
60
80
100
120
140
160
Activitate enzimatică
5.5 6.5 7.5pH
după 48 de ore
după condiţionare
Figura 9. Variaţia activitătii amilolitice după 48 de ore şi după condiţionare pe carbonat de
calciu
Tabelul 18. Variaţia activităţii xilanazice în funcţie de pH-ul mediului de cultură după 48
ore şi după condiţionare
PH Probă (ml sau g) Soluţie tampon
(ml)
diluţia Extincţia 540 nm Activitate xilanazică
(U/ml)
5.5 2 2 X5 0,5040/0,0941 1366,336.5 2 2 X5 0,5963/0,0941 16747.5 2 2 X5 0,5757/0,0941 1595,36
PRODUS CONDIŢIONAT-prin adsorbţie pe CaCO3.
5.5 0,0759 2 0,5181/0,0696 3,95 U/mg6.5 0,0742 2 0,3635/0,0624 2,67 U/mg
În ceea ce priveşte activitatea xilanazică a culturii de Aspergillus niger după 48 de
ore, funcţie de pH, se observă că aceasta este optimă la un pH 6,5-7, iar pentru produsul
condiţionat activitatea xilanazică este mai ridicată la pH = 5,5, dar scăderea nu este
dramatică la valoarea de pH = 6,5, ceea ce permite a se considera pH-ul 5,5 ca optim,
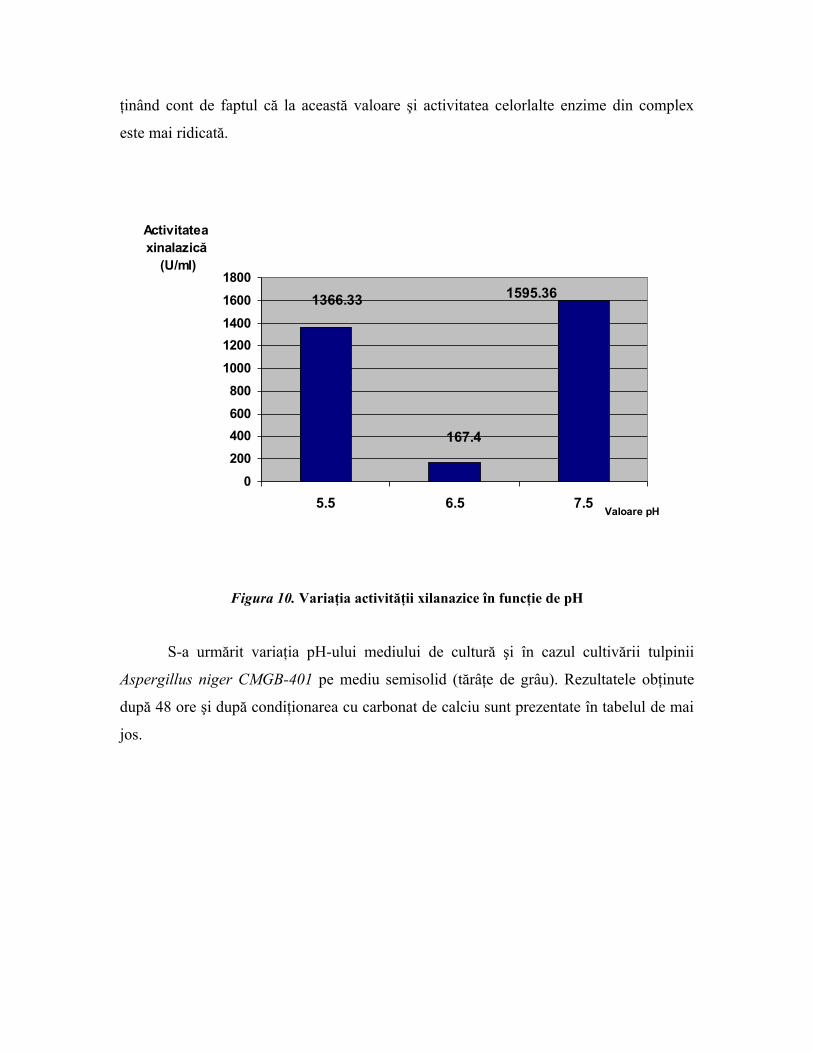
ţinând cont de faptul că la această valoare şi activitatea celorlalte enzime din complex
este mai ridicată.
1595.36
167.4
1366.33
0
200
400
600
800
1000
1200
1400
1600
1800
5.5 6.5 7.5Valoare pH
Activitatea xinalazică
(U/ml)
Figura 10. Variaţia activităţii xilanazice în funcţie de pH
S-a urmărit variaţia pH-ului mediului de cultură şi în cazul cultivării tulpinii
Aspergillus niger CMGB-401 pe mediu semisolid (tărâţe de grâu). Rezultatele obţinute
după 48 ore şi după condiţionarea cu carbonat de calciu sunt prezentate în tabelul de mai
jos.

Tabelul 19. Variaţia activităţilor enzimatice în funcţie de pH-ul mediului semisolid (cu
tărâţe de grâu) după 48 ore şi după condiţionare
PH Activitatea amilolitică
Activitatea proteolitică
Activitatea celulozolitică
(C1-Cx)
Activitate xilanazică
(U/mg)
5.5 75.12 (U/ml) 1.98 (U/ml) 1.45 (U/ml)6.5 66.96 (U/ml) 1.55(U/ml) 1.05 (U/ml)
7.5 52.32 (U/ml) 1.29 (U/ml) 1.27 (U/ml)PRODUS CONDIŢIONAT-prin adsorbţie pe CaCO3.
5.5 76.32 (U/g) 1.22 (U/g) 0.98 (U/g) 8,74
6.5 58.88 (U/g) 1.4 (U/g) 0.81 (U/g)7.5 50.88 (U/g) 1.48 (U/g) 1.22 (U/g)
Examinand comparativ rezultatele prezentate în tabelele 17 -19 care redau variaţia
activităţilor enzimatice în funcţie de pH pe mediul M4 (stabilit în prima fază ca fiind
mediul optim de dezvoltare) şi un mediu semisolid cu tărâţe de grâu ca sursă de carbon
(activitatea amilolitică 75.12 UDNS/g, activitatea proteolitică 1.98 UP/g, activitatea
celulozolitică 1.45 UC1-Cx/g), s-a constatat că biosinteza enzimelor hidrolitice (amilaza,
proteaza, celulaza) ca şi a xilanazei este favorizată în mediul lichid (M4-făină de soia) la
pH=5.5. Mediul lichid este mai avantajos din punct de vedere economic ceea ce îl
recomandă pentru folosirea în condiţii de micropilot şi pilot.
5.4. Testarea complexului enzimatic ca aditiv furajer
Greutatea medie iniţială a animalelor supuse testării a fost de 8 kg. După 18 zile
de la începutul experimentului s-a efectuat cântărirea individuală finală iar rezultatele
obţinute au fost prelucrate statistic (tabelul 18, figura 8). Performanţele au fost
îmbunătăţite în cazul loturilor la care a fost inclusă enzima în structura reţetei de nutreţ
combinat dar nu s-au semnalat diferenţe semnificative în ceea ce priveşte greutatea
corporală şi sporul mediu zilnic (P>0.05). O posibilă explicaţie a îmbunătăţirii
performanţelor la purcei prin adăugarea de enzimă ar fi efectul asupra unor factori ca:
timpul de tranzit intestinal al hranei, producţiile endogene, schimbările în polizaharurile
fără amidon, degradarea sau schimbarea fibrei[26].

Tabelul 20. Performanţe bioproductive
SpecificaţieL O T
M E1 E2Greutate corporala iniţiala (kg) 8,75 8,71 8,96Greutate corporala finală (kg)* 12,75 a ± 1,94 12,83 a ± 1,90 13,23 a ± 2,86
Spor mediu zilnic (kg) 0,222a ± 0,09 0,229a ± 0,08 0,237a ± 0,12Consum mediu zilnic de NC (kg) 0,439 0,446 0,450
Consum specific -kg NC /Kg spor- % faţă de martor
1,98100,00
1,9598,48
1,9095,96
*aceeaşi literă diferenţe nesemnificative între loturi (P>0.05)
8.75 12
.75
8.71 12
.83
8.96 13
.23
0
2
4
6
8
10
12
14
G. initiala G. finala
M E1 E2
Figura 10. Dinamica greutăţii corporale
În ceea ce priveşte sporul mediu zilnic (tabelul 20, figura 11), se constată de
asemenea, o uşoară îmbunătăţire în cazul loturilor experimentale (0,237 kg la lotul E1,
respectiv, 0,229 kg la lotul E2, faţă de 0,222 kg la lotul martor), fără ca diferenţele să fie
semnificative. Se apreciază că enzima ameliorează efectul antinutritiv şi eficientizează
producţia [26].

0.22
2
0.22
9
0.23
7
0.21
0.22
0.23
0.24
smz
M
E1
E2
Figura 11. Sporul mediu zilnic
Consumul mediu zilnic de nutreţ combinat a fost asemănător la cele trei loturi
(tabelul 20, figura 12). S-a constatat o reducere a consumului specific în cazul loturilor
cu adaos de preparat enzimatic (1,95 kg NC/kg spor la lotul E1 respectiv, 1,90 kg NC/kg
spor în cazul lotului E2 comparativ cu 1,98 kg NC/kg spor la lotul M).
0
0.3
0.6
0.9
1.2
1.5
1.8
2.1
M
E1
E2
M 0.439 1.98
E1 0.446 1.95
E2 0.450 1.90
c.m.z NC Consum specific
Figura 12 . Consumul mediu zilnic (kg) şi consumul specific (kg NC/Kg spor)

CONCLUZII
În cadrul cercetărilor experimentale prezentate în proiectul de faţă s-au testat
două tulpini de Aspergillus niger şi o tulpină Trichoderma viridae din colecţia Centrului
de Biotehnologii Microbiene şi respectiv a Facultăţii de Biologie, Universitatea
Bucureşti, în ceea ce priveşte capacitatea acestora de a produce un complex de enzime
hidrolitice (proteaze, amilaze, celulaze, xilanaze) utilizabil ca aditiv furajer.
Cele trei tulpini de fungi au fost cultivate în sistem submers, pe 4 medii de
cultură preluate din literatura de specialitate; activitatea polienzimatica a mediilor de
fermentaţie şi a produselor finite (obţinute prin filtrarea mediului de fermentaţie,
concentrare şi adsorbţie pe CaCO3 tehnic) s-a evaluat comparativ cu preparatul
KEMZYME®VP DRY (Belgia).
Rezultatele obţinute au evidenţiat capacitatea bioproductivă superioară a tulpinii
Aspergillus niger CMGB-401 în ceea ce priveşte complexul enzimatic studiat.
În scopul determinării condiţiilor optime de cultivare a tulpinii de Aspergillus
niger CMGB-401 pentru biosinteza enzimelor hidrolitice (amilaze, proteaze, celulaze,
xilanaze), au fost cercetaţi o serie de parametrii care au o importanţă semnificativă pentru
acest tip de bioprocese şi anume: compoziţia mediul de cultură, temperatura, pH-ul
mediului de cultură şi gradul de agitare şi aerare al culturii.
În ceea ce priveşte compoziţia mediului de cultură, dintre cele cinci medii testate
(patru medii lichide şi un mediu semisolid pe bază de tărâţe de grâu), cele mai bune
rezultate s-au obţinut cu mediul lichid M 4, având următoarea compoziţie (g%):
Făină de porumb-2%;
Făină de soia-3%;
(NH4)2HPO4-0,5%;
MgSO4x7H2O-0,05%;
CaCl2-0,1%
În cazul diferitelor variante de pH şi temperatură folosite, s-a observat că tulpina
de Aspergillus niger CMGB-401 conduce cele mai bune activităţi enzimatice, în urma

cultivării la pH = 5,5 şi la temperatura de 280C (activitatea amilolitică 141,6 UDNS/g,
activitatea proteolitică 1,95 UP/g, activitatea celulozolitică 2,27 UC1-Cx/g, activitatea
xilanazică 1366,33 U/ml).
Condiţionarea produsului polienzimatic s-a realizat prin concentrarea mediului
integral din finalul fermentaţiei, tratarea cu CaCO3 a concentratului şi uscarea sub vid a
amestecului rezultat.
Testarea ca aditiv furajer a produsului polienzimatic condiţionat a evidenţiat
faptul că utilizarea acestuia nu a influenţat semnificativ greutatea corporală şi sporul
mediu zilnic a purceilor în perioada crizei de înţărcare, posibil din cauza faptului că
nutreţul combinat a fost optimizat în principii nutritivi specifici acestei categorii de
porcine;
Consumul specific la loturile de purcei la care s-a utilizat preparat enzimatic este
cu 1,52% - 4,04% mai mic faţă de martor, in funcţie de cantitatea de produs introdusă în
raţia zilnică.
Rezultatele obţinute sugerează eficienţa utilizării preparatelor enzimatice în
recepturi de nutreţ combinat deficitare în principii nutritivi.

BIBLIOGRAFIE
1. David Cowan, International Journal for Food Chemistry, pag. 32-38, 1992.
2. Sun Y., Cheng J., Biores. Technol. 83: 1-11, 2002.
3. Rosgaard I., Pederson S., Cherry J.R., Harris P., Meyer AS., Biotechnol Prog.,
22(2):493-498, 2006.
4. Puls J., Schuseil J., Hemicellulose and Hemicellulases, MP Coughlan and GP
Hazlewood (Eds), Portland Press, 1993.
5. Betts W.B., Dart R.K., Ball A.S., Pedlar S.L., Biodegradation: Natural and Synthetic
6. A.L. Demain, N.A. Solomon, Biology of Industrial Microorganisms,1985.
7. Bielecki Stainslaw, Rita Pyc, Institute of Technical Biochemistry, Technical
University of Eoodz, Poland, pag. 153-157.
8. Z. Park, S. Kang, J. Lee, S. Hong, S. Kim, Applied Microbiology and Biotechnology,
vol. 58,nr. 6, pag. 761-766, 2002.
9. P. V. Gawande and M.Y. Kamat, Journal of Applied Microbiology, vol.87, issue 4,
pag. 511, 1999.
10. Stefana Jurcoane, Biotehnologii*Fundamente*Bioreactoare*Enzime, Editura Tehnica
Bucuresti, pag. 308-318,2000
11. Liming Xia, Peilin Cen, Process Biochemistry 34, 909-912, 1999.
12. Kamm B., Kamm M., Schmidt M., Sarke I., Kelinpeter E., Chemosphere, 62, 97-105,
2006

13. Levine J.S, Biomass burning and global change, vol. 1, 1996,. Levine J.S (ed), The
MIT Press, Cambridge, Massachusetts, USA, pp 35.
14. Eustace A. Iyayi, sp., African Journal of Biotechnology vol.3, 186-188,2004.
15. Dierick, N.A. and J.A. Decuypere, Supplementary enzymes to improve
utilization of pig diets. Proceedingsa 45th Annual Meeting of EAAP, Edinburgh,
1994.
16. Luiza Jecu, Industrial Crops and Products, 11,pag.1-5, 2000.
17. Costa-Fereira M., Matos de Sousa J., Procedeengs of Bioenergy-I: From concept to
Commercial Processes, Tomar, Portugalia, 5-10.03.2006.
18. Malherbe S., Cloete T.E., Environ. Sci. Biotechnol. 1:105-114, 2003
19. Kanuf M., Monirussaman M., International Sugar Journal, 106: 147-150, 2004
20. Miller G. L., Anal. Chem., 31, 426-428,1959.
21. Petterson and Porath, Eur. J. Biochem, 1974.
22. Iordachescu D., Dumitru I.F., Biochimie practica,151, 1980.
23. Baylei M. J., Biely P., J. Biotechnol., 23, 257-270,1959.
24. Vasile Anca, Ciurescu Georgeta, Moldovan I., Editura Cartea Universitara, vol.
8, 111-116, 2005.
25. Varga E., Rczey K., Zacchi G.,. Appl. Biochem. Biotechnol., 113:509-523, 2004
26. D.I.Kitchen, Enzyme applications in corn/soia diets fed pigs, in Biotechology in
the Feed
Industry, Proceedings of Alltech’s 13th Annual Symposium Nottingham Univ. Press.
T.P. Lyons& K.A. Jacques (Ed), 1977.


















