
mg2seria2.files.wordpress.com · Web viewFigura 1 – Ciclul vieții și mărcile epigenetice în...
23
6. EPIGENETICA Suntem în era geneticii. Proiecte științifice uriașe sunt întreprinse pentru a afla care sunt secvențele de ADN care codifică toate caracteristicile noastre. Proiectul genomului uman ne- a dezvăluit întreaga secvență a genomului uman. În momentul în care a început proiectul genomului uman, ne așteptam ca majoritatea întrebărilor noastre să primească răspuns, că vom putea găsi explicația mutațiilor sau variațiilor bolilor doar prin cunoașterea secvenței. Acum știm că a fost o supraestimare, că secvența nu ne spune totul despre funcția ADN-ului și că există încă multe legături lipsă înainte de a înțelege toți factorii care determină cine suntem. În mod clar, există ceva mai mult decât secvența noastră de ADN care determină funcția genelor și a caracterelor pe care le afișăm în fenotipul nostru. Ceilalți factori care joacă un rol în reglarea genică alaturi de secventa ADN-ului sunt obiectul de studiu al acestei noi ramuri interesante a științei - epigenetica. De zeci de ani, una dintre cele mai interesante intrebări care i-a preocupat pe biologi a fost: Cum este posibil ca aceleași informații genetice în fiecare celulă să conducă la atât de multe tipuri celulare, prezentând atât de multe fenotipuri diferite odată ce celulele se specializează? Acum știm că acest lucru este obținut prin reglarea genetică foarte specifică: fiecare tip de celulă este definit de genele care sunt exprimate în acea celulă (prin transcriere și translație), ceea ce înseamnă că fiecare tip de celulă exprimă doar un subset restrâns de gene. Dacă o celulă umană conține aproximativ 25.000 de gene în genom, nu acel număr de gene este activ în oricare celulă specializată, ci mai degrabă ele exprimă doar acele gene care sunt importante pentru funcția lor. Deși acest lucru este adevărat în diferite celule ale corpului, știm, de asemenea, că procese similare pot fi observate și la oameni. Știm că există oameni identici din punct de vedere genetic (gemeni monozigoți) care apar atunci când un embrion se scindează în două părți devreme în cursul sarcinii. Cu toate acestea, gemenii identici pot avea fenotipuri diferite (de exemplu, gemenii sunt discordanți pentru înălțime în 30% din cazuri). Uneori, gemenii monozigoți poartă amândoi gene de risc pentru anumite boli, dar numai unul are fenotipul bolii. Acest lucru este acum atribuit faptului că, deși gemenii sunt identici din punct de vedere genetic, probabil ei sunt diferiți din punct de vedere epigenetic. Oamenii de știință au introdus termenul de epigenetică pentru a exprima faptul că, alături de ADN, există un alt nivel de informație ("epi" = "deasupra") ce determină care sunt genele care vor fi activate pentru fiecare tip de celulă. Aceasta se obține prin două mecanisme diferite: activitatea factorilor de transcripție specifici pentru fiecare linie celulară, și mărcile epigenetice specifice aplicate ADN-ului. Mărcile epigenetice sunt centrul acestui curs și subiectul principal al epigeneticii în general. Acestea reprezintă modificări moleculare în ADN sau proteine care reglează funcția ADN-ului și permit interpretarea secvenței de ADN: mărcile epigenetice permit cunoașterea începutului și sfârșitului genei, ele ajută la menținerea structurii cromozomilor, modifică gradul de transcripție a anumitor gene (unele gene sunt induse, în timp ce altele sunt reprimate). Controlul epigenetic este extrem de important și reglementat de-a lungul dezvoltării. Dacă controlul epigenetic nu funcționează corect, acest lucru poate duce la o gamă largă de anomalii la diferite nivele în dezvoltarea umană. 6.1. Definiție 1
Transcript of mg2seria2.files.wordpress.com · Web viewFigura 1 – Ciclul vieții și mărcile epigenetice în...

6. EPIGENETICA
Suntem în era geneticii. Proiecte științifice uriașe sunt întreprinse pentru a afla care sunt secvențele de ADN care codifică toate caracteristicile noastre. Proiectul genomului uman ne-a dezvăluit întreaga secvență a genomului uman. În momentul în care a început proiectul genomului uman, ne așteptam ca majoritatea întrebărilor noastre să primească răspuns, că vom putea găsi explicația mutațiilor sau variațiilor bolilor doar prin cunoașterea secvenței. Acum știm că a fost o supraestimare, că secvența nu ne spune totul despre funcția ADN-ului și că există încă multe legături lipsă înainte de a înțelege toți factorii care determină cine suntem. În mod clar, există ceva mai mult decât secvența noastră de ADN care determină funcția genelor și a caracterelor pe care le afișăm în fenotipul nostru. Ceilalți factori care joacă un rol în reglarea genică alaturi de secventa ADN-ului sunt obiectul de studiu al acestei noi ramuri interesante a științei - epigenetica.
De zeci de ani, una dintre cele mai interesante intrebări care i-a preocupat pe biologi a fost: Cum este posibil ca aceleași informații genetice în fiecare celulă să conducă la atât de multe tipuri celulare, prezentând atât de multe fenotipuri diferite odată ce celulele se specializează? Acum știm că acest lucru este obținut prin reglarea genetică foarte specifică: fiecare tip de celulă este definit de genele care sunt exprimate în acea celulă (prin transcriere și translație), ceea ce înseamnă că fiecare tip de celulă exprimă doar un subset restrâns de gene. Dacă o celulă umană conține aproximativ 25.000 de gene în genom, nu acel număr de gene este activ în oricare celulă specializată, ci mai degrabă ele exprimă doar acele gene care sunt importante pentru funcția lor. Deși acest lucru este adevărat în diferite celule ale corpului, știm, de asemenea, că procese similare pot fi observate și la oameni. Știm că există oameni identici din punct de vedere genetic (gemeni monozigoți) care apar atunci când un embrion se scindează în două părți devreme în cursul sarcinii. Cu toate acestea, gemenii identici pot avea fenotipuri diferite (de exemplu, gemenii sunt discordanți pentru înălțime în 30% din cazuri). Uneori, gemenii monozigoți poartă amândoi gene de risc pentru anumite boli, dar numai unul are fenotipul bolii. Acest lucru este acum atribuit faptului că, deși gemenii sunt identici din punct de vedere genetic, probabil ei sunt diferiți din punct de vedere epigenetic.
Oamenii de știință au introdus termenul de epigenetică pentru a exprima faptul că, alături de ADN, există un alt nivel de informație ("epi" = "deasupra") ce determină care sunt genele care vor fi activate pentru fiecare tip de celulă. Aceasta se obține prin două mecanisme diferite: activitatea factorilor de transcripție specifici pentru fiecare linie celulară, și mărcile epigenetice specifice aplicate ADN-ului. Mărcile epigenetice sunt centrul acestui curs și subiectul principal al epigeneticii în general. Acestea reprezintă modificări moleculare în ADN sau proteine care reglează funcția ADN-ului și permit interpretarea secvenței de ADN: mărcile epigenetice permit cunoașterea începutului și sfârșitului genei, ele ajută la menținerea structurii cromozomilor, modifică gradul de transcripție a anumitor gene (unele gene sunt induse, în timp ce altele sunt reprimate). Controlul epigenetic este extrem de important și reglementat de-a lungul dezvoltării. Dacă controlul epigenetic nu funcționează corect, acest lucru poate duce la o gamă largă de anomalii la diferite nivele în dezvoltarea umană.
6.1. Definiție
Epigenetica este definită ca știința care studiază schimbări heritabile în expresia genelor care nu se datorează modificărilor secvenței ADN. Modificările expresiei genice sunt obținute prin gradul de împachetare al ADN. Cu cât ADN este mai condensat, cu atât mai puțin active sunt genele din regiunea respectivă și viceversa. Diferitele grade de condensare a cromatinei sunt obținute prin modificări biochimice în ADN sau proteinele asociate, așa cum se va descrie în secțiunile următoare. Toate aceste modificări sunt denumite mărci epigenetice. Heritabilitatea mitotică se referă la procesul prin care statusul epigenetic al unei celule este conservat prin mitoză și este transferat la următoarea generație de celule fice. Acest lucru este important în menținerea identității celulare și a omogenității țesuturilor. Heritabilitatea este contracarată de perioadele în care se elimină semnele epigenetice. De exemplu, în cazul reprogramării epigenetice în dezvoltarea embrionară timpurie, mărcile vechi ale celulelor germinale sunt eliminate pentru a permite zigotului să obțină pluripotență. Odată ce celulele pluripotente încep să se diferențieze, vor începe să stabilească noi mărci epigenetice care vor determina identitatea și funcția noilor celule fice (Figura 1).
6.2. Mecanismele reglării epigenetice
În nucleu, ADN-ul este complexat cu proteine structurale (histone) pentru a forma nucleozomi. Majoritatea ADN-ului dublu-elicoidal este înfășurat în jurul unui miez octameric format din patru homodimeri histonici: H2A, H2B, H3 și H4. Două bucle de ADN sunt ținute în jurul octamerului histonei de către o altă proteină histonă denumită H1. Prin nivele multiple de împachetare, nucleozomii stabilesc conformații ale
1

cromatinei care pot fi relaxate (eucromatină) sau dense (heterocromatină). Acest lucru facilitează sau reprimă transcripția în celule în momente critice ale dezvoltării. De exemplu, regiunile telomerice și centromerice netranscrise sunt inhibate datorită mediului compact de heterocromatină, în timp ce genele extrem de active, de obicei localizate în eucromatină, sunt exprimate datorită unei cromatine mai deschise și accesibilității sporite pentru factorii de transcripție.
Anumite mărci epigenetice sunt asociate cu eucromatina și heterocromatina. Unele dintre cele mai importante mărci epigenetice vor fi menționate mai jos.
Figura 1 – Ciclul vieții și mărcile epigenetice în dezvoltarea celulelor somatice la om. Gameții fuzionează pentru a forma zigotul care va trece prin mitoze successive ce determină creșterea și diferențierea. Aceasta coincide cu o perioadă în care majoritatea mărcilor epigenetice care aparțin gameților specializați sunt eliminate, urmate de noi mărci epigenetice care se adaugă pe parcurs ce celulele somatice se specializează în funcție de propriul țesut și funcție. * Dezvoltarea celulelor germinale (prezentate aici în roșu) va conține un alt moment în care mărcile sunt îndepărtate și reintroduse (vezi secțiunile următoare privind amprentarea genomică).
-6.2.1. Metilarea ADN Probabil cel mai cunoscut proces epigenetic este metilarea ADN-ului. Metilarea ADN constă în
adăugarea unei grupări metil la cel de-al 5-lea atom de carbon al resturilor de citozină din ADN (citozina va fi transformată în 5-metil citozină, Figura 2).
Figura 2 – Metilarea citozinei. Resturile de citozină primesc o grupare metil la poziția celui de-al 5-lea atom de carbon prin activitatea metil transferazelor. Această reacție este reversibilă, sub efectul demetilazelor.
Transferul unei grupări metil la citozină se întâmplă în regiuni care conțin repetări ale bazelor citozină și guanină (repetări CG, cunoscute sub denumirea de regiuni CpG).
În 98% din genom, dinucleotidele CpG apar cu o frecvență redusă de 1/80 nucleotide. În restul de 2% din genom, acestea se găsesc în insule CpG, zone bogate în dinucleotide CpG cu lungimi variind de la 200
2

perechi de baze (bp) la mai multe kilobaze (kb) și cu o densitate CpG mai mare. Aproximativ 50-60% din gene conțin insule CpG în promotorii lor proximali (Figura 3). Astfel de insule CpG sunt aproape întotdeauna nemetilate în țesuturile normale. Siturile CpG din afara regiunilor promotor sunt aproape întotdeauna metilate (Figura 3).
Figura 3 – Metilarea ADN și localizarea acesteia în genom. Metilarea ADN are loc la nivelul dinucleotidelor CpG. Dinucleotidele CpG din cadrul insulelo CpG asociate promotorilor sunt de obicei nemetilate; la nivelul regiunilor intergenice, ele sunt de obicei metilate; elementele repetitive sunt de obicei metilate (cercurile goale reprezintă citozine nemetilate, cercurile întunecate pline simbolizează prezența metilării).
Metilarea ADN-ului este un semn epigenetic care este asociat cu heterocromatina. Siturile în care este prezentă metilarea ADN-ului pot interacționa cu factorii de transcripție sau pot recruta enzime care modifică histonele și care transformă în continuare această regiune într-o cromatină reprimată (condensată). Metilarea ADN la insulele CpG associate promotorilor cauzează, de obicei, represia genelor. Foarte important, lipsa generală de metilare în insulele CpG associate promotorilor poate fi perturbată în condiții patologice, în special în celulele canceroase, ducând la inhibiția aberantă a genelor.
Metilarea ADN la regiunile intragenice, intergenice sau fragmentelor repetitive determină represia acestor secvențe, prevenind astfel transpoziția, interferența transcripțională sau recombinarea ilegitimă care poate fi cauzată de aceste secvențe repetitive în genom. Prin aceasta, metilarea ADN-ului joaca un rol important in menținerea integrității genomului și limitarea ratelor de mutație.
Transferul grupării metil la citozină se produce simetric pe ambele catene ADN, astfel încât mărcile de metilare ale ADN-ului pot fi menținute prin diviziunea celulară (heritabilitate mitotică). Reacția este catalizată de ADN metiltransferaze (DNMT) care stabilesc și mențin metilarea ADN-ului după diviziunea celulară. De exemplu, DNMT1, principala metilază de întreținere a gradului de metilare, are o afinitate ridicată pentru ADN-ul hemimetilat (ADN dublu catenar care are citozine metilate pe o catenă, dar citozine nemetilate pe catena complementară). DNMT1 recunoaște aceste zone hemi-metilate și stabilește grupuri de metil la catena care conține citozine nemetilate.
Astfel, DNMT1 acționează pentru a susține modelele originale de metilare odată ce ADN-ul celulei s-a replicat și celula s-a divizat. Știm că prezența și menținerea metilării ADN este crucială pentru dezvoltarea normală. Șoarecii cărora li s-au eliminat genele DNMT (knock-out) mor in utero, dovedind că metilarea ADN-ului este esențială pentru viabilitate. Dimpotrivă, lipsa activității DNMT va duce la diluarea nivelelor de metilare din interiorul unei celule, contribuind astfel la îndepărtarea metilării ADN-ului. Acest proces poate fi pasiv (activitatea DNMT se oprește și nivelele de metilare a ADN vor fi reduse gradual cu fiecare replicare și diviziune celulară) sau poate fi activă, catalizată de demetilaze (enzime care elimină în mod activ grupele metil din resturile de 5-metil citozină). Aceasta implică faptul că metilarea ADN nu este un proces ireversibil, ci mai degrabă o marcă epigenetică dinamică ce poate fi modulată.
Un exemplu foarte bun de metilare a ADN-ului și de heritabilitate mitotică este inactivarea cromozomului X. Inactivarea X este un mecanism epigenetic de compensare a dozajului genic la mamifere, astfel încât bărbații și femeile au aceeași doză de gene pe cromozomul X. Inactivarea aleatorie X are loc la gastrulație în embrion. Există o șansă de 50-50% ca oricare dintre cei doi cromozomi X la mamiferele femele
3

să fie inactivat în orice celulă dată. După acest moment, această stare epigenetică este moștenită mitotic de către toate celulele fiice (Figura 4).
Figura 4 – Pisicile vărgate și inactivarea cromosomului X. Inactivarea aleatorie a cromosomului X și moștenirea sa mitotică sunt ușor observate în pisicile vărgate. Pisicile au o gena pe cromosomul X care codifică culoarea blănii (negru sau portocaliu). Pisicile femele care sunt heterozigote pentru această genă pot exprima ambele alele din cei doi cromozomi X. Cu toate acestea, în timpul dezvoltării embrionare, la gastrulație, unul dintre cei doi cromosomi X este inactivat aleatoriu în fiecare celulă disponibilă. Ulterior, celulele fiice vor menține acel cromosom într-o stare inactivă și vor transcrie gene de la celălalt cromosom. Ca rezultat, pisicile vor avea un aspect vărgat (calico cat), cu zone în portocaliu sau în negru, aleatoriu pe întreg corpul, conform cromozomului care a fost inactivat inițial în acea zonă. Datorită acestei explicații, blana vărgată poate fi văzută doar la femele, deoarece masculii moștenesc un singur cromosom X.
-6.2.2. Modificările histonice După ce genele histonelor sunt transcrise și translatate în proteine, moleculele histonelor pot primi
diferite grupări chimice adăugate ulterior. Numim aceste mărci modificări post-translaționale histonice. De cele mai multe ori aceste modificări au loc la cozile N-terminale ale proteinelor histonice, care sunt cozile ce ies din nucleozom, făcându-le accesibile enzimelor. Acetilarea și metilarea cozilor histonice sunt cel mai bine caracterizate (vezi mai jos). Alte grupuri chimice mai pot fi adăugate: ubiquitinilare, fosforilare, ADP-ribozilare, citrulinizare. Scopul cercetării în acest domeniu este de a studia cu ce sunt asociate toate aceste mărci, pentru a putea anticipa ce fel de rezultat ar putea avea loc pentru regiunea respectivă, în funcție de codul histonic din aceea zonă ADN (Figura 5).
Figura 5 – Modificările histonice. Modificări majore ale histonelor sunt asociate cu diferite grade de compactare a cromatinei. Acetilarea este asociată întotdeauna cu cromatina activă. Metilarea poate fi asociată atât cu zonele de eucromatină cât și cu heterochromatina.
4

Modificările posttranslaționale histonice se produc aproape exclusiv la nivelul histonelor H3 sau H4, nu în tipurile H1 sau H2. Diferitele modificări sunt corelate cu diferite funcții.
Acetilarea histonelor este corelată cu activarea genică (universal). Histonele sunt acetilate de histon-acetiltransferaze, iar grupările acetil sunt îndepărtate prin histon-deacetilaze. Modul în care acetilarea funcționează pentru a determina activarea genei este prin reducerea sarcinii pozitive a histonelor și diminuarea atracției dintre acestea și ADN (sarcină negativă). Acest lucru slăbește cromatina și atrage alte proteine care pot modifica în continuare poziția nucleozomilor pentru a scădea condensarea cromatinei (Figura 5).
Metilarea histonelor se corelează fie cu activitatea transcripțională, fie cu inactivitatea. Mono, di și tri-metilarea este posibilă. Grupările metil sunt adăugate prin histon-metil transferaze și sunt îndepărtate prin demetilaze. ex.: H3K4me = metilare (me) la al 4-lea rest de lizină (K4) al histonei 3 (H3) = aceasta este o marcă asociată cu eucromatina H3K9me3 = tri- metilare (me3) la al 9-lea rest de lizină (K9) al histonei 3 (H3) = aceasta este o marcă asociată cu heterocromatina Mecanismul de acțiune al metilării histonelor este acela de a acționa ca punct de amorsare pentru alte proteine care au funcția de a muta nucleozomii în jur și de a efectua remodelarea cromatinei pentru a influența gradul de condensare (Figura 5).
Este mai puțin cunoscut dacă modificările histonice sunt mitotic heritabile sau nu în comparație cu procesul descris anterior de metilare a ADN. Nu există încă un concept al modului în care o astfel de schimbare poate fi transmisă celulelor fiice, ceea ce face mai probabilă presupunerea că aceste modificări pot fi conservate numai pe parcursul ciclului de viață al celulei respective. Mai multe cercetări sunt în desfășurare pe această temă.
-6.2.3. ARN non-codantDupă cum sugerează și numele, ARN-urile non-codante sunt molecule de ARN care sunt codificate de genele din nucleu, dar nu
codifică proteine. ARN-urile non-codante sunt implicate mai degrabă în reglarea exprimării genelor prin mecanisme indirecte. Există multe clase de ARN-uri non-codante mici, medii sau lungi care sunt studiate în prezent. Este mult peste scopul acestui curs să descriem toate exemplele disponibile, dar mai degrabă ne vom concentra pe două clase de ARN non-codant și vom oferi câteva exemple.
Micro-ARN (miARN) – sunt molecule de ARN scurte care sunt implicate în inhibarea post-transcripțională a genelor. La mamifere, miARN-urile sunt transcrise în nucleu și sunt transportate în citoplasmă. În citoplasmă, miARN-urile pot lega moleculele de ARN mesager (ARNm) care codifică proteine. Dacă există o omologie de secvență între miARN și ARN mesager, atunci miARN se va lega la ARNm pe baza complementarității, blocând astfel translația ARN-ului mesager într-o proteină. Acesta este un mecanism de represiune translațională care reduce cantitatea de proteină codificată de către acea genă. Acest lucru nu modifică în sine compactarea cromatinei, ci mai degrabă este un mecanism de inhibiție a genei la un nivel post-transcripțional.
ARN non-codant lung (lncARN) – sunt molecule lungi de ARN care direcționează complexele epigenetice și ajută la stabilirea unor mărci epigenetice. Ele sunt transcrise în nucleu și vor rămâne în nucleu pentru a-și îndeplini funcția. ARN-urile non-codante lungi pot funcționa în mai multe moduri: ca și ghiduri care leagă, pe de-o parte, anumite regiuni ADN pe baza omologiei secvenței și, pe de altă parte, leagă proteinele sau enzimele care au roluri de remodelare a cromatinei, aducându-le la locul pe care îl recunosc; ele pot funcționa și ca schele, având capacitatea de a permite mai multor proteine sau complexe să acceseze un anumit sit al genomului, furnizând suportul care pune împreună mai multe proteine pentru a stabili alte mărci epigenetice. Un exemplu foarte bun pentru un ARN non-codant lung este Xist. Xist joacă un rol crucial în inactivarea cromosomului X. Am menționat anterior că inactivarea X este un mecanism epigenetic de compensare a dozajului genic care asigură că bărbații și femeile au aceeași doză de gene active de pe cromosomul X. Procesul necesită specificitate alelică, deoarece numai unul dintre cei doi cromosomi X ar trebui să fie inactivat, nu ambii. Primul eveniment în inactivarea X este transcrierea Xist din cromosomul care va deveni condensat. Pe de o parte, Xist se va lega de același cromozom din care a fost copiat. Pe de altă parte, Xist va lega de asemenea un complex proteic implicat în stabilirea unor modificări posttranslaționale histonice represive (în mod specific, H3K27me) (Figura 6).
Xist va acoperi treptat cromozomul X din care este transcris și va ghida acest complex de proteine pentru a răspândi semnele inhibitorii pe tot parcursul cromozomului X. Acesta este primul eveniment cheie implicat în inactivarea X. Metilarea ADN-ului, așa cum sa menționat în secțiunile anterioare, tinde să se întâmple mai târziu în procesul de inactivare X, odată ce s-au acumulat alte semne represive la locul de interes.
5

Figura 6 – ARN-ul non-codant lung Xist și mecanismul de acțiune. Xist leagă secvențele ADN pe baza complementarității și ghidează un complex enzimatic numit Polycomb Repressor 2 Complex (PRC2). PRC2 adaugă metilarea H3K27 la histonele adiacente care contribuie la heterochromatinizarea acelui cromozom X.
6.3. Epigenetica în medicină
După cum s-a discutat până în prezent, putem spune că reglarea epigenetică a expresiei genelor este un mecanism crucial pentru modularea activității genelor și producției de proteine în funcție de fenotipul fiecărei celule. De asemenea, ne oferă cunoștințe despre cum pot avea loc anumite procese cunoscute din biologie (cum ar fi inactivarea cromozomului X) și care sunt modificările chimice / moleculare care mediază aceste procese. Pe măsură ce cercetarea epigenetică se extinde, tot mai multe informații devin disponibile pentru a înțelege câteva mecanisme foarte importante pentru dezvoltarea normală. Cu toate acestea, disrupțiile controlului epigenetic au implicații și în ceea ce privește înțelegerea bolilor și, în cele din urmă, posibilitatea de a interveni terapeutic. În această secțiune ne vom concentra asupra modului în care reglarea epigenetică are un rol important în apariția bolilor. Ne vom concentra asupra bolilor genetice rare și vom descrie că uneori fenotipurile tulburărilor genetice pot fi cauzate de un dezechilibru epigenetic, nu de o eroare genetică. Mai mult, vom explica, de asemenea, modul în care epigenetica capătă din ce în ce mai multă atenție în patogeneza bolilor multifactoriale comune.
-6.3.1. Boli rare – anomalii de amprentare genomicăAm menționat că diferențierea celulară și stabilirea de mărci epigenetice se întâmplă pe tot parcursul
dezvoltării. Acest lucru are sens, deoarece celulele se diferențiază din celule pluripotente embrionare în celule specializate. Cu toate acestea, ȋn unele momente în dezvoltare, aceste mărci epigenetice trebuiesc eliminate, așa cum este ilustrat pe scurt în Figura 1. În această secțiune descriem un proces care are lor în unul din aceste momente în care semnele epigenetice sunt îndepărtate și reintroduse: amprentarea genomică.
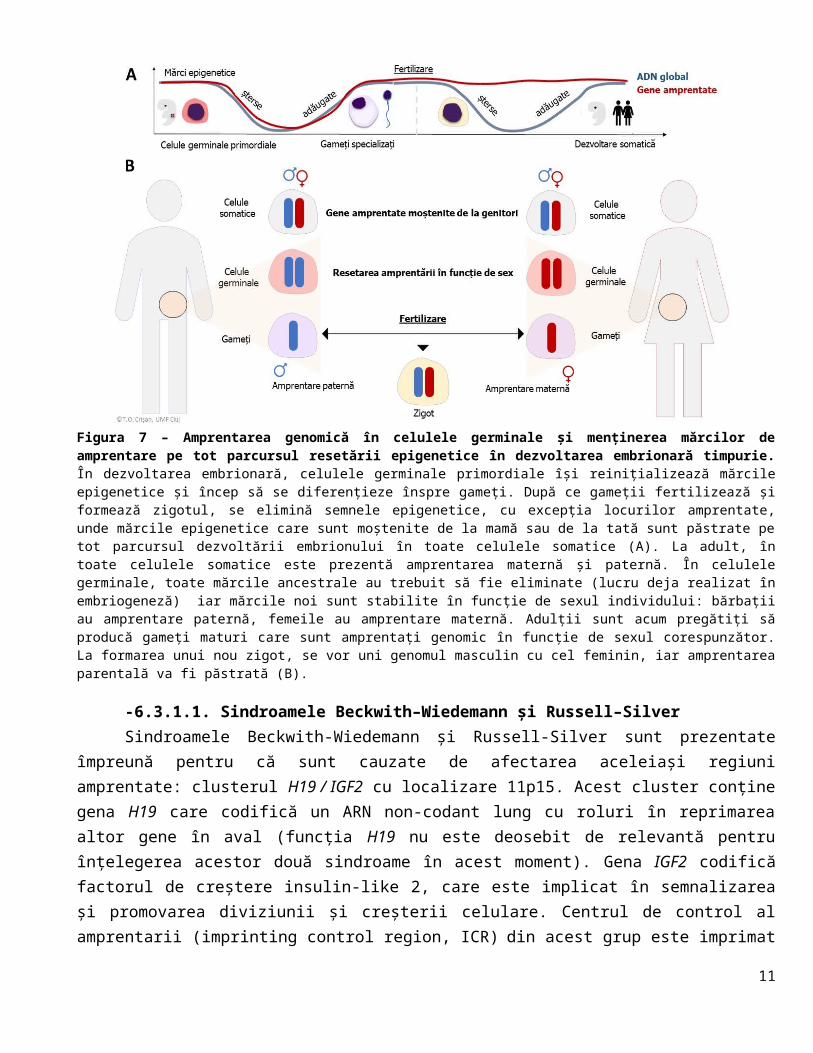
Amprentarea genomică este un fenomen epigenetic normal la om care se întâmplă când se formează gameții. Știm că gameții își încep formarea în timpul dezvoltării embrionare la locul gonadei primitive, urmând să se termine prin meioză pe durata vieții adulte. Reglarea epigenetică a genomului gameților începe cu eliminarea (aproape a) tuturor mărcilor epigenetice care au fost prezente în celulă și stabilirea de noi mărci specifice pentru diferențierea unui ovul sau a unui spermatozoid (Figura 7A). În acest moment, unele gene vor fi activate sau inhibate în mod diferit, în funcție de tipul de celulă care se va produce: ovul sau un spermatozoid. Aceste gene sunt ținta amprentării genomice și se numesc gene amprentate. Această activare sau inhibiție a unor gene precis alese va rezista la resetarea mărcilor epigenetice în embrionul nou format (Figura 7A). Aceasta înseamnă că, pentru unele zone ale genomului, embrionii vor moșteni alele care se vor comporta diferit dacă sunt moștenite de la mamă prin ovul, comparativ cu cazul în care sunt moștenite de la tată prin spermatozoid (Figura 7B).
Știm că amprentarea genomică este extrem de importantă pentru viabilitate, deoarece în condiiții experimentale, șoarecii care au două genomuri paterne sau două genomuri materne mor înainte de implantare. Prin urmare, genele amprentate vor fi gene la care va fi exprimată doar o singură alelă: deși avem două alele în genomul nostru pentru fiecare locus, în regiunile amprentate, una dintre cele două alele va fi activă și cealaltă va fi silențioasă din punct de vedere epigenetic într-un mod specific originii parentale. Cum se produce amprentarea genomica? De obicei, genele amprentate se găsesc în clustere în genom. Nu sunt aproape niciodată gene individuale activate sau inhibate prin amprentare, ci mai degrabă sunt grupuri de gene care sunt reglate împreună prin efectele unei singure regiuni numite centrul de control al amprentarii (imprinting control region, ICR) al acelui grup de gene. Metilarea / non-metilarea ICR determină efectele ulterioare care vor determina reglarea epigenetică a genelor găsite în acest cluster. Există multe regiuni amprentate în genomul uman. Toate acestea au mecanisme complexe de reglare
6

și este dincolo de scopul acestui curs să detalieze toate căile posibile în care ICR-urile pot determina amprentarea în diferite gene. Cu toate acestea, dorim să ne concentrăm pe exemplul a două regiuni și să descriem modul în care are loc reglarea în fiecare dintre acestea. Aceste exemple oferă o idee despre modul în care metilarea de la ICR poate funcționa în feluri diferite pentru a determina expresia unei singure alele pentru anumiți loci. Mai mult, explică cauza unor sindroame moștenite care nu pot fi pe deplin înțelese doar prin genetică. Pentru cele două exemple de mai jos, vom descrie mai întâi modul în care regiunea amprentată funcționează în situații normale. Ulterior, vom descrie modul în care aceasta poate fi bulversată pentru a provoca sindroame genetice.
Figura 7 – Amprentarea genomică în celulele germinale și menținerea mărcilor de amprentare pe tot parcursul resetării epigenetice în dezvoltarea embrionară timpurie. În dezvoltarea embrionară, celulele germinale primordiale își reinițializează mărcile epigenetice și încep să se diferențieze înspre gameți. După ce gameții fertilizează și formează zigotul, se elimină semnele epigenetice, cu excepția locurilor amprentate, unde mărcile epigenetice care sunt moștenite de la mamă sau de la tată sunt păstrate pe tot parcursul dezvoltării embrionului în toate celulele somatice (A). La adult, în toate celulele somatice este prezentă amprentarea maternă și paternă. În celulele germinale, toate mărcile ancestrale au trebuit să fie eliminate (lucru deja realizat în embriogeneză) iar mărcile noi sunt stabilite în funcție de sexul individului: bărbații au amprentare paternă, femeile au amprentare maternă. Adulții sunt acum pregătiți să producă gameți maturi care sunt amprentați genomic în funcție de sexul corespunzător. La formarea unui nou zigot, se vor uni genomul masculin cu cel feminin, iar amprentarea parentală va fi păstrată (B).
-6.3.1.1. Sindroamele Beckwith–Wiedemann și Russell–Silver Sindroamele Beckwith-Wiedemann și Russell-Silver sunt prezentate împreună pentru că sunt cauzate
de afectarea aceleiași regiuni amprentate: clusterul H19 / IGF2 cu localizare 11p15. Acest cluster conține gena H19 care codifică un ARN non-codant lung cu roluri în reprimarea altor gene în aval (funcția H19 nu este deosebit de relevantă pentru înțelegerea acestor două sindroame în acest moment). Gena IGF2 codifică factorul de creștere insulin-like 2, care este implicat în semnalizarea și promovarea diviziunii și creșterii celulare. Centrul de control al amprentarii (imprinting control region, ICR) din acest grup este imprimat patern (metilat pe cromozomul paternal) care se corelează cu transcripția IGF2 din cromozomul patern. În schimb, pe cromozomul matern, IGF2 nu va fi transcrisă și transcripția va avea loc la gena H19. Modul în care funcționează acest lucru este ilustrat în figura 8.
Acest proces în care IGF2 este exprimată numai de pe cromosomul patern produce nivele normale de IGF2 și creșterea normală a copilului. Dacă, în condiții patologice, acest echilibru este întrerupt, iar copilul nu are cromozomul care se comportă în stilul matern, atunci va exista un exces de IGF2. Această
7

afecțiune are ca rezultat sindromul Beckwith-Wiedemann. Sindromul Beckwith-Wiedemann are caracteristica principală a creșterii accentuate (macrosomie), pacienții asociază trăsături craniofaciale, cum ar fi macroglosia, defectele peretelui abdominal, hemihidrotrofia, hipoglicemia, tumorile embrionare.
Figura 8 – Clusterul H19/IGF2 și sindroamele Beckwith–Wiedemann și Russell–Silver. În diagrama de mai sus este prezentat grupul H19 / IGF2, care conține următoarele componente (de la stânga la dreapta): gena IGF2, regiunea de control a amprentării (ICR), gena H19, regiuni de tip „enhancer” găsite în aval în secvența ADN. ICR este prezentat ca un oval care este fie gol, ceea ce reprezintă (ICR nemetilat) sau umplut în culoare neagră (ICR metilat). Enhancers sunt secvențe ADN care pot intra în contact cu alte regiuni ADN pe care le pot activa. Acest contact este posibil prin buclele ADN-ului: deoarece ADN-ul are o structură cuaternară, știm că zonele care ar fi foarte îndepărtate pot veni în contact strâns din cauza buclelor din cromatină. În situații normale, acest grup este amprentat patern = ICR este metilat pe cromozomul patern. Pe cromozomul matern, ICR este nemetilat. Când este nemetilat, ICR este legat de o proteină numită CTCF. CTCF este o proteină izolatoare, blochează accesul enhancers din aval la locusul IGF2, prin urmare IGF2 nu este exprimat. În consecință, enhancers vor lega gena H19. Spre deosebire pe cromozomul patern, CTCF nu poate lega ICR metilat, prin urmare enhancers vor putea stimula preferențial transcripția IGF2, în timp ce H19 nu va fi exprimat. Acest lucru asigură faptul că atât H19, cât și IGF2 sunt exprimate numai de pe un cromozom (fiecare). Dacă, în situații de boală, o persoană nu are partea care se comportă ca paternă sau lipsește partea care se comportă ca maternă, va exista o lipsă de echilibru în exprimarea genelor amprentate, respectiv, vor apărea Sindroamele Russell-Silver sau Beckwith-Wiedemann.
Dacă copilul nu are cromozomul care se comportă în stil patern, atunci va exista o producție insuficientă de IGF2. Această afecțiune are ca rezultat sindromul Russell-Silver. Sindromul Russell-Silver are caracteristica principală a restricției de creștere, determinând o statură semnificativ redusă la pacienții afectați, adesea cu o înălțime finală între 140 cm (feminin) și 151 cm (masculin). De asemenea, se asociază alte fenotipuri, cum ar fi dificultăți de hrănire, hipotonie musculară, hipoglicemie, asimetrie, rareori alte anomalii congenitale sunt prezente, întârzierea de vorbire poate fi asociată.
-6.3.1.2. Sindroamele Angelman și Prader–WilliSindroamele Prader-Willi și Angelman sunt un alt exemplu de sindroame care sunt prezentate într-un cuplu deoarece sunt cauzate de
afectarea aceleiași regiuni amprentate: clusterul Snrpn, cu localizare 15q11-13. Clusterul Snrpn conține multe gene diferite, dintre care se numără gena care codifică ARN-ul non-codant lung Snrpn. O altă genă importantă la acest grup este UBE3A, care codifică ubiquitin ligaza E3A, o enzimă importantă pentru procesele de degradare din interiorul celulelor. ICR din acest grup este metilat pe cromozomul matern - și acest lucru se corelează cu lipsa de transcripție în majoritatea genelor găsite în această regiune, cu excepția UBE3A. Pe cromozomul patern, ICR este nemetilat și cele mai multe gene sunt transcrise. O caracteristică a acestui cluster este faptul că, deși majoritatea genelor sunt active pe cromozomul patern, UBE3A este reprimată în mod specific în creier. Aceasta înseamnă că, în creier, singura sursă de UBE3A este cromozomul matern. Modul în care funcționează acest cluster este ilustrat în Figura 9.
8

Figura 9 – Clusterul Snrpn și Sindroamele Prader–Willi and Angelman Syndromes. În diagrama de mai sus este prezentat clusterul Snrpn, care conține următoarele componente (de la stânga la dreapta): reprezentarea schematică a diferitelor gene din acest grup, regiunea de control a amprentării (ICR), gena Snrpn, gena UBE3A. ICR este prezentat ca un oval care este fie gol, ceea ce reprezintă ICR nemetilat sau colorat în negru (ICR metilat). În situații normale, acest grup este amprentat matern, ceea ce înseamnă că ICR este metilat pe cromozomul matern. Pe cromozomul paternal, ICR este nemetilat. Lipsa de metilare pe cromozomul patern permite exprimarea ARN-ului non-codant lung Snrpn, care va lega ADN-ul descendent pe o secvenţă mai lungă. În creier, acest ARN ajunge până la locusul UBE3A și astfel inhibă expresia lui. Alte gene găsite în amonte de ICR pot fi, de asemenea, exprimate. Pe cromozomul matern, ICR este metilat, lncRNA Snrpn nu este transcris, prin urmare, nu există un efect inhibitor asupra genei UBE3A. Acest lucru permite ca UBE3A să fie transcris din cromozomul matern. Dacă, în situații de boală, o persoană nu are partea care se comportă ca paternă sau lipsește partea care se comportă ca maternă, va exista o lipsă de echilibru în exprimarea acestor gene amprentate respectiv, apar sindroamele Prader-Willi sau Angelman.
Situația în care UBE3A este exprimată numai în creier de la cromozomul matern produce niveluri normale de UBE3A și dezvoltarea neurologică normală a copilului. Dacă, în condiții patologice, acest echilibru este întrerupt, iar copilul nu are cromozomul care se comportă în stilul matern, atunci va exista o deficiență a UBE3A. Această afecțiune are ca rezultat sindromul Angelman. Sindromul Angelman are caracteristica principală a dizabilității intelectuale grave, acompaniind microcefalie. Pacienții asociază convulsii și manifestă o dispoziție fericită care este caracteristică acestui sindrom, cu momente inadecvate de râs pe fondul tulburărilor de dezvoltare neurologice. În schimb, expresia tuturor celorlalte gene din acest grup doar de pe cromozomul patern produce nivele normale ale acelor proteine corespunzătoare și dezvoltarea normală a copilului.
Dacă acest echilibru este întrerupt și copilul nu are cromozomul care se comportă în stilul patern, atunci va exista o deficiență în exprimarea genelor pentru acești loci. Această afecțiune va determina sindromul Prader-Willi, care are ca trăsătură principală dizabilitate intelectuală, însoțită de tulburări de comportament (comportament obsesiv-compulsiv), cu obiceiuri alimentare patologice până la obezitate morbidă, asociind hipogonadism. Pacienții în perioadă neonatală afișează un tonus muscular scăzut și eșec al creșterii.
-6.3.1.3. Cauzele și moștenirea anomaliilor de amprentareAm văzut în cele două secțiuni de mai sus că expresia necorespunzătoare a genelor amprentate este cee
ace cauzează anomaliile de amprentare. Dar cum putem explica faptul că aceste tulburări de expresie au loc? Menţionăm, mai jos, patru mecanisme majore care pot explica acest lucru.
a) Variația numărului de copii – deleție sau duplicație în regiunile amprentate. De exemplu, dacă există o deleție pe cromozomul 15q care implică clusterul de gene Snrpn, atunci fenotipul va varia în funcție de ce cromosom a suferit deleția - de ex. Deleția pe cromosomul matern rezultă în sindromul Angelman.
b) Disomie uniparentală – eroare reproductivă care determină moștenirea de la același părinte a ambilor cromozomi într-o pereche de omologi. De exemplu, dacă există o disomie uniparentală paternă a cromozomului 15, atunci pacientul are 2 cromozomi care sunt amprentați patern și nu are varianta amprentată matern - aceasta va duce din nou la sindromul Angelman.
9

c) Mutație moleculară la ICR sau în genele esențiale ale clusterului. Ambii cromozomi sunt în mod normal moșteniți de la părinții corespunzători, dar:
a. o mutație la ICR blochează procesul normal de metilare, iar regiunea nu poate fi amprentată correct;
b. o mutație într-o genă crucială imită fenotipul unei erori de amprentare (exemplu: mutație genetică în gena UBE3A într-o regiune altfel corect amprentată - gena va fi exprimată în mod normal, dar mutația în genă va duce la o proteină anormală cu efecte patologice - și acest lucru ar putea duce la sindromul Angelman.)
d) Eroare epigenetică. Ambii cromozomi sunt în mod normal moșteniți de la fiecare părinte, nu există mutații în secvența ADN, dar a apărut o eroare epigenetică ce a dus la imposibilitatea de a seta mărcile epigenetice corecte la acel locus. Acest fenomen se numește epimutație. Epimutațiile la locii amprentați sunt rare, iar cauzele acestor erori epigenetice sunt încă în curs de cercetare.
Știind că tulburările de amprentare sunt strâns dependente de sexul părintelui, deci vor exista și implicații asupra modului de transmitere a acestor boli. Cele mai multe boli genetice ereditare sunt transmise în conformitate cu legile mendeliane (v. capitolul anterior). Anomaliile de amprentare sunt excepții de la legile mendeliene de transmitere și urmează o transmitere dependentă de originea parentală.
În cazul clusterului Snrpn, de exemplu, sindromul Prader-Willi este moștenit de la tată, în timp ce sindromul Angelman este moștenit de la mamă. O femeie purtătoare nu ar putea avea niciodată un copil cu sindromul Prader-Willi. În schimb, un bărbat purtător, nu ar putea avea niciodată un copil cu sindromul Angelman (vezi explicația din Figura 10).
Figura 10. Transmitere dependentă de originea parentală în anomaliile de amprentare. Pedigree-ul de mai sus este un exemplu teoretic al unei tulburări de amprentare care urmează transmitere maternă. Acest lucru ar putea fi un exemplu de sindrom Beckwith-Wiedemann cauzat de mutații în clusterul H19 / IGF2, ceea ce a dus la afectarea amprentării materne la acest locus. Observați tatăl purtător din prima generație – acesta are un cromozom cu mutație, dar nu este afectat. El are un risc de 50% să transmită cromozomul cu mutația la urmașii săi - dar, din moment ce este bărbat, el trebuie să stabilească amprentare de tip patern în toți gameții, prin urmare mutația nu va avea niciun efect aici. În consecință, copiii care moștenesc această mutație de la tatăl lor vor fi purtători sănătoși. Aceeași situație se întâmplă și cu fiul său purtător din a doua generație. Fiica purtătoare a acestuia, totuși, trebuie să producă gameți care conțin cromozomi amprentați matern. Deoarece mutația nu permite efectuarea acestei amprentări în mod corect, acei copii care au moștenit cromozomul mutant vor fi afectați. În cea de a treia generație, bărbatul afectat nu va avea copii afectați, în timp ce fiica afectată are din nou un risc de 50% de a avea copii afectați. În multe cazuri, tulburările de amprentare provoacă un fenotip în care pacienții nu pot procrea, în astfel de cazuri un astfel de pedigree nu ar fi văzut în practică.
-6.3.2. Epigenetica și factorii de mediuÎn această secțiune vom analiza felul în care mediul înconjurător poate influența reprogramarea
epigenetică, conducând astfel la consecințe pentru sănătate. În primul rând, vom menționa posibile disrupții ale controlului epigenetic datorită intervenției medicale - de exemplu în tehnicile de reproducere asistată. Apoi vom descrie influențele de mediu care ar putea afecta reglarea genică - aceasta va fi mai importantă pentru înțelegerea influenței mediului asupra tulburărilor comune. În cele din urmă, vom avea o subsecțiune privind epigenetica cancerului, deoarece dezechilibrul epigenetic este unul dintre semnele distinctive ale fenotipului tumoral.
10
-6.3.2.1. Reproducerea asistatăTehnicile de reproducere asistată (TRA), în special injecția intra-citoplasmatică a spermatozoizilor
(ICSI) se asociază cu o incidență mai mare a tulburărilor de amprentare. Știm că aceste tulburări de imprimare ar putea fi rezultatul erorilor epigenetice (așa cum s-a discutat mai sus), prin urmare, aceasta a ridicat problema că TRA poate crește riscul de anomalii epigenetice - în regiuni amprentate, dar și în general în genom.
Știm că există două momente în dezvoltare în care reprogramarea epigenetică este crucială: în dezvoltarea celulelor germinale (mărcile epigenetice somatice sunt eliminate și se instalează semne epigenetice specifice celulelor
germinale) și în dezvoltarea embrionară timpurie (mărcile epigenetice ale gameților sunt îndepărtate pentru a permite pluripotență și dezvoltarea
ulterioară). Ambele sunt momente în care are loc manipularea în TRA: producția de gameți este stimulată pentru a obține mai multe ovule simultan de la mamă înainte de a fi supusă TRA, iar dezvoltarea embrionară timpurie se produce in vitro până la punctul de implantare în uterul mamei. Nu avem încă nicio dovadă precisă care demonstrează inducerea de anomalii epigenetice de către TRA însă e nevoie de mai multe studii experimentale pe această temă.
Trebuie de asemenea precizat că, având în vedere riscul foarte scăzut de anomalii epigenetice, chiar dacă TRA crește riscul de mai multe ori, tot va exista un număr extrem de mic de copii născuți cu aceste anomalii. Deci, cei mai multi copii născuţi prin TRA vor fi sănătoși în ciuda faptului că aceste metode ar putea crește riscul de anomalii epigenetice. Deci, problema consecințelor epigenetice în TRA este încă deschisă, iar studiile continuă să încerce să afle ce tulburări pot să apară și care este frecvența acestora în ART comparativ cu reproducerea fiziologică.
-6.3.2.2. Boli comune – boli multifactoriale.Odată ce ajungem să înțelegem la ce se referă epigenetica, putem de asemenea să ne imaginăm că m ărcile epigenetice trebuie să apară
nu numai în perioadele de reprogramare care au o importanță crucială pentru dezvoltarea normală, dar se întâmplă probabil în genom pe tot parcursul vieții, datorită influențelor mediului. Pe măsură ce corpul se adaptează la mediul înconjurător, este posibil să avem unele gene reglate pentru a adapta procesele noastre interne la cerințele sau stimulii externi. Acest lucru se știe că se întâmplă mai degrabă independent de secvența de ADN. Știm de asemenea că indivizii identici genetic (gemeni monozigoți) sunt foarte asemănători la naștere pentru nivelele de metilare a ADN-ului din întregul genom, dar modelul de metilare a ADN-ului genomic e semnificativ diferit dacă e măsurat în timpul vieții adulte.Apoi, este tentant să speculăm că mecanismele epigenetice are putea fi mediatori ai expunerii corpului la diferiți factori de mediu, care joacă un rol în susceptibilitatea noastră la boli.
Datorită interacțiunii gene-mediu, mărcile epigenetice pot fi modificate în celule sau țesuturi pentru a direcționa adaptarea mecanismelor biologice. Mai jos, vom menționa câteva exemple de descoperiri în care expunerea la diverși stimuli de mediu determină, pe de o parte, schimbarea mărcilor epigenetice în genomul celulelor și, pe de altă parte, prin această reprogramare epigenetică joacă un rol în patogenia unor boli.-a)Astmul. Astmul este o reacție de hipersensibilitate de tip I în care maturarea celulelor T helper (Th) este înclinată spre un răspuns Th2. Celulele Th2 vor produce mai multe IgE care sunt implicate în patogeneza astmului. Cercetatorii au fost în măsură să demonstreze că această progresie accentuată înspre Th2 este mediată de factori epigenetici, cum ar fi metilarea ADN-ului si modificările histonice (H3K9ac, H3K4me2) la nivelul unor gene care codifică citokine cu rol în dezvoltarea celulelor T.-b) Diabetul și bolile metabolice. Un număr mare de dovezi științifice obținute în studii pe oameni și șoareci arată că hiperglicemia poate provoca modificări epigenetice în celulele vasculare la promotorii unor factori de transcripție proinflamatori, și că acest lucru ar putea juca un rol în inflamația cronică. Metilarea ADN la nivelul altor gene a fost asociată cu infarctul miocardic la femei. Diferite modificări histonice au fost asociate cu expunerea la hiperglicemie. Acest ansamblu de studii a condus la termenul de "memorie glicemică" sau "memorie metabolică", care este colectiv considerat un exemplu important în care stimulii metabolici modifică reglarea genică în celule, cu consecințe relevante asupra sănătății. Modificări similare s-au observant și în bolile metabolice, cum ar fi bolile cardiovasculare, obezitatea, iar lista se extinde progresiv.-c) Bolile neurologice. Boala Alzheimer, schizofrenia și tulburările de spectru autist arată o varietate de anomalii epigenetice. Faptul că procesele epigenetice pot apărea în ciuda diviziunii celulare au creat optimism în domeniul tulburărilor neurologice, deoarece aceste procese ar putea fi țintite prin viitoare terapii în celulele care nu se divid, cum sunt neuronii.-d) Biologia vaccinurilor și bolile infecțioase. Studiile recente arată că vaccinurile pot avea efecte heteroloage (influențează răspunsul imun împotriva altor agenți patogeni decât cei pentru care sunt administrați). Cel mai evident exemplu la momentul actual este vaccinarea BCG (Bacillus Calmette-Guerin) pentru prevenirea tuberculozei. Sa observat că nou-născuții vaccinați cu BCG au o mortalitate semnificativ mai mică (ne-datorată tuberculozei) în comparație cu copiii care nu au primit vaccinul. Studiile in vitro demonstrează că BCG are efecte epigenetice (H3K4me3 și H3K27ac la nivelul unor gene implicate în răspunsul imun), care stimulează răspunsul antiinfecțios înspre mai mulți agenți patogeni decât M. tuberculosis, oferind o protecție generală crescută.
11

-6.3.2.3. Epigenetica în cancerÎn cancer, cea mai timpurie aberație epigenetică găsită a fost o lipsă globală de metilare a genomului.
Hipometilarea la nivelul regiunilor intergenice și elementelor repetitive este asociată cu o instabilitate genomică marcată și o recombinare ilegitimă care provoacă mutații. Această hipometilare a ADN-ului genomic este o caracteristică găsită în toate cancerele studiate vreodată și este chiar mai consistentă decât mutațiile genetice!
Cu toate acestea, unele tipuri de cancer (cum ar fi cancerul colorectal) sunt asociate cu o hipermetilare a anumitor promotori genici și inhibarea acelor gene. Asocierea fumatului cu risc crescut de cancer colorectal s-a dovedit a fi prezentă numai în grupul de pacienți care a prezentat această hipermetilare.
Virusul hepatitei B (HBV) este cunoscut a fi implicat în patogeneza cancerului hepatic. Integrarea ADN-ului VHB în genomul gazdei a fost asociată cu hipermetilarea în hepatocite.
Rolul folaților și al donatorilor de grupări metil în limitarea cancerului sau influențarea riscului de cancer a fost studiat extensiv și a adus rezultate modeste sau negative. Totuși, s-a constatat o corelație între aportul de folat în perioade peri-concepționale și în timpul sarcinii cu un risc mai mic de afecțiuni maligne la copii. Aceste studii necesită replicare și înțelegere mai aprofundată.
Cunoașterea acestor variații ale nivelelor de metilare în cancer s-a dovedit a fi o resursă excelentă pentru dezvoltarea de terapii noi. Tratamentele bazate pe utilizarea enzimelor metiltransferaze sau demetilaze ar putea, sperăm, să aducă noi strategii de limitare a cancerului. În prezent se efectuează multe studii clinice și multe alte ținte epigenetice trebuie încă să fie transpuse în terapie în acest domeniu.
6.4. Epigenetica în cercetare – transmiterea transgenerațională
Una dintre cele mai fascinante întrebări din domeniul epigeneticii la om este: există o transmitere epigenetică transgenerațională mediate prin gameți? Dacă mediul poate schimba reglarea și funcția genomului nostru, va fi posibil ca această schimbare să fie transmisă copiilor noștri? Sau se va pierde odată cu moartea organismului care a suferit aceste schimbări?
Am văzut cum se modifică statusul epigenetic în diferite momente ale dezvoltării: avem mai multe momente în care semnele epigenetice sunt în mare parte eliminate și re-adăugate în funcție de tipul celular. Ar fi, oare, posibil ca schimbările obținute pe parcursul vieții să găsească modalități de păstrare, în ciuda acestor perioade de îndepărtare / remodelare a statusul nostru epigenetic? (Pentru claritate, trebuie să precizăm că această întrebare exclude mecanismele de amprentare discutate mai sus, care sunt strict controlate, și se referă mai degrabă la noi modificări ale genomului care au avut loc la un individ datorită expunerii sale specifice la mediul înconjurător.)
Mai jos, vom evidenția câteva dintre numeroasele constatări care susțin transmiterea epigenetică transgenerațională la șoareci și la oameni.-a. Un studiu a investigat transmiterea de la o generație la alta a unei expuneri traumatice: șoarecii au fost învățați să se teamă de mirosul acetofenonei prin învățarea condiționată (inițial au fost expuși unor stimuli dureroși aplicați imediat după mirosul acetofenonei, ceea ce a dus la teama doar de acetofenonă, în absența durerii, în timpul expunerilor ulterioare). Mai târziu, șoarecii condiționați au fost lăsați să se reproducă, iar puii din următoarele două generații au manifestat frică de acetofenonă, moștenită de la părinții lor. Acest lucru s-a dovedit a fi produs prin hipometilarea CpG la nivelul genei care codifică un receptor olfactiv (Olfr151). Această hipometilare s-a dovedit a fi prezentă în gameții indivizilor expuși, și acest lucru a fost considerat mecanismul transmiterii transgeneraționale în acest exemplu (Figura 11).-b. Unul dintre cele mai frecvent întâlnite exemple la șoareci este cel al genei Agouti. Gena Agouti este implicată în mai multe efecte metabolice, dar și în culoarea blănii la șoareci: comută producția de pigment de la o culoare maro (așa cum se vede la tipul sălbatic) la galben. Dacă există hipometilare în amonte de promotorul genei - gena va fi exprimată, iar culoarea blănii va fi galbenă. Dacă se produce metilarea, culoarea blănii va fi maro. Nu există diferențe genetice între șoarecii de culoare maro și cei de culoare galben, va fi doar o variație epigenetică responsabilă pentru acest lucru. Aceste nivele diferite de metilare se întâmplă stocastic (aleatoriu) în mare măsură dependente de dieta mamei: nutriție bogată în donatori ai grupului de metil => inhibarea expresiei Agouti, culoarea brună a blănii puilor; nutriție săracă la donatorii de grupuri de metil => Expresia Agouti, culoarea galbenă a blănii puilor.-c. Unul dintre cele mai răspândite exemple și studii la om este un studiu epidemiologic care a luat naștere în perioada "foametei olandeze". Aceasta a fost o perioadă de aproape un an între 1944-1945, când aprovizionarea cu alimente a fost redusă în Țările de Jos datorită izolării din timpul celui de-al doilea război
12

mondial și a unei ierni foarte reci, care a blocat accesul în regiune. A fost păstrată o bună evidență a populației din vremea respectivă și au fost posibile studii de urmărire în timp pentru a investiga dacă au avut loc consecințe ulterioare perioadei de foamete. S-a observat că expunerea la foamete în perioada peri-concepțională a fost asociată cu un risc mai mare de tulburări metabolice (obezitate, diabet, boli cardiovasculare) la vârsta de adult în cazul descendenților care au fost concepuți în timpul foametei. Acest lucru nu a fost observat la alți copii ai acelorași părinți dacă momentul concepției a fost în afara perioadei de foamete.-d. Un studiu epidemiologic similar a avut loc în rândul populației din China afectată de foametea severă dintre anii 1958-1961, cauzată atât de probleme politice cât și meteorologice. S-a observant o asociere între foametea/malnutriția prenatală și schizofrenia la adult. Această asociație a fost confirmată și în studiul iarnii olandeze.-e. În cele din urmă, un studiu epidemiologic a putut investiga situația inversă în comparație cu cele două exemple de mai sus: este cunoscut faptul că o zonă din Suedia (parohia Ӧverkalix) are o evidență bună a aprovizionării cu alimente. Aici, în anumite perioade de timp a fost înregistrată creșterea disponibilității alimentelor, în timp ce în alte perioade au fost a fost înregistrat un deficit de alimente. S-a constatat o asociere conform căreia aprovizionarea cu alimente în copilărie la bărbați a modificat metabolismul la nepoții lor. Acesta a fost unul dintre primele exemple în care un stimul de mediu este asociat cu un efect transgenerațional la om.
Subiectul transmiterii transgeneraționale a modificărilor epigenetice este foarte controversat, deoarece studiile se bazează în cea mai mare parte pe asocieri și nu pe dovedirea acestor mecanisme. Studiile la om nu pot fi efectuate cu exactitate și nu pot fi studii corect controlate din cauza problemelor etice și tehnice. Există mai multe modele animale care fac acest concept interesant, dar, totuși, avem nevoie de dovadă la oameni pentru a ști cu adevărat dacă și cum o persoană poate transmite moștenirea sa epigenetică la generația următoare.
Figura 11. Reprezentarea schematică a unuia dintre experimentele lui Dias și Ressler care prezintă moștenirea transgenerațională prin gameți a comportamentului indus de miros la șoareci. Șoarecii au fost condiționați să se teamă de mirosul de acetofenonă și apoi au fost lăsați să se reproducă. Puii din prima generație (F1) și din a doua generație (F2) au arătat un comportament alterat față de acetofenonă, sugerând moștenirea unei trăsături dobândite de la strămoșii lor (mijloc). Pentru a distinge dacă această moștenire a fost biologică (transmisă prin gameți) sau indusă social de către părinte, s-a realizat schimbarea mediului de viață (cross-fostering): puii nou-născuți au fost luați de la părinți și au fost lăsați să fie crescuți de șoareci naivi. Modificările comportamentale au fost prezente și de această dată, argumentând în favoarea transmiterii prin gameți (panoul inferior). Concluzia că transmiterea este biologică a fost întărită prin experimente de fertilizare in vitro (FIV): gameții șoarecilor condiționați au fost utilizați pentru FIV într-o instituție diferită, folosind șoareci naivi străini. Efectele comportamentale au fost în continuare observate la puii astfel obținuți (panoul superior).
6.5. Concluzii
Domeniul epigeneticii este un subiect care se extinde rapid, care este foarte relevant pentru înțelegerea multor mecanisme biologice. Încă nu s-au găsit răspunsuri la numeroase întrebări de cercetare privind cauzele, mecanismele și rezultatele controlului epigenetic. Cu toate acestea, epigenetica este un domeniu care ne oferă un alt nivel de informație, alături de genetică, pentru a explica variațiile dintre noi. De
13

asemenea, ne permite să studiem modul în care expresia genică poate fi controlată prin mecanisme moleculare care stabilesc mărci epigenetice. Faptul că modificările epigenetice sunt inductibile de factori externi înseamnă de asemenea că, în condițiile propice, regalarea epigenetică ar putea fi reversibilă. Aceasta oferă ținte de tratament promițătoare pentru bolile care implică modificări epigenetice.
-Mesaje de ținut minte Epigenetica este știința care studiază reglarea genică => efecte genetice care nu sunt cauzate de
variațiile în secvența ADN, ci de gradul expresiei genice. Deja avem cunoștințe cu privire la unele procese epigenetice cunoscute și acceptate (diferențierea
celulară, inactivarea cromosomului X). Multe din descoperirile făcute au elucidat unele mecanisme de control epigenetic (metilare ADN,
modificări posttranslaționale histonice, ARN non-codant). Totuși, multe întrebări rămân neelucidate, iar cercetarea continua în domeniu.
Importanța epigeneticii în medicină este imensă: -1) boli rare de amprentare sunt explicate de diferențe în reglarea epigenetică la anumite regiuni cromosomiale, în funcție de originea parental;-2) boli comune pot să fie mediate și, eventual tratate, prin modificările epigenetice instituite în genom, prin interacțiunea dintre genetică-epigenetică-factorii de mediu de pe parcursul vieții; -3) alte studii sunt în derulare pentru a descoperi dacă modificările epigenetice dobândite pe parcursul vieții pot fi transmise de la o generație la alta.
Selected references and further reading:-1. Epigenetics Ethical, Legal and Social Aspects, Reinhard Heil, Stefanie B. Seitz, Harald König, Jürgen Robienski Editors, Springer Fachmedien Wiesbaden GmbH 2017, DOI 10.1007/978-3-658-14460-9-2. The use of mouse models to study epigenetics, Blewitt Marnie, Whitelaw E. Cold Spring Harb Perspect Biol. 2013, 5(11): a017939. doi: 10.1101/cshperspect.a017939.-3. Genomic imprinting: recognition and marking of imprinted loci , Lara K. Abramowitz and Marisa S. Bartolomei Curr Opin Genet Dev. 2012, 22(2): 72–78. doi: 10.1016/j.gde.2011.12.001-4. The evolution of genomic imprinting: theories, predictions and empirical tests , M M Patten, L Ross, J P Curley, D C Queller, R Bonduriansky, and J B Wolf, Heredity 2014 Aug; 113(2): 119–128. doi: 10.1038/hdy.2014.29-5. Fat dads must not be blamed for their children’s health problems , Moore and Stanier BMC Medicine 2013http://www.biomedcentral.com/1741-7015/11/30 (Please also see related research article here http://www.biomedcentral.com/1741-7015/11/29) -6. Environmental epigenetics: prospects for studying epigenetic mediation of exposure–response relationships , Victoria K. Cortessis, Duncan C. Thomas, A. Joan Levine, Carrie V. Breton, Thomas M. Mack, Kimberly D. Siegmund, Robert W. Haile, Peter W. Laird, Human Genetics (2012) 131:1565–1589, DOI 10.1007/s00439-012-1189-8-7. Trained immunity: A smart way to enhance innate immune defence. Jos van der Meer, Leo A.B. Joosten, Niels Riksen, Mihai G.Netea. Mol Immunol. 2015, 68(1):40-4. doi: 10.1016/j.molimm.2015.06.019.-8. Targeting histone methyltransferases and demethylases in clinical trials for cancer therapy , Ludovica Morera, Michael Lübbert and Manfred Jung, Clinical Epigenetics (2016) 8:57 DOI 10.1186/s13148-016-0223-4-9. Killing Me Softly: The Fetal Origins Hypothesis, Douglas Almond and Janet Currie J Econ Perspect. 2011, 25(3): 153–172. doi:10.1257/jep.25.3.153.-10. Transgenerational epigenetic inheritance: More questions than answers, Lucia Daxinger and Emma Whitelaw, Genome Research 2010 http://www.genome.org/cgi/doi/10.1101/gr.106138.110. -11. The sins of the father, Virginia Hughes, Nature. 2014, 507: 22-24.-12. Parental olfactory experience influences behavior and neural structure in subsequent generations, Brian G Dias and Kerry J Ressler, Nature Neuroscience 2014, 17: 89–96 doi:10.1038/nn.3594-13. Cardiovascular and diabetes mortality determined by nutrition during parents’ and grandparents’ slow growth period, G Kaati, LO Bygren and S Edvinsson, European Journal of Human Genetics 2002, 10: 682 – 688.
Useful web resources:-1. Epigenetic Control of Gene Expression, MOOC held by Marnie Blewitt, https://www.coursera.org/learn/epigenetics -2. Learn Genetics – Genomic Imprinting http://learn.genetics.utah.edu/content/epigenetics/imprinting/ -3. GeneEd - Genetics, Education, Discovery https://geneed.nlm.nih.gov/topic_subtopic.php?tid=35-4. What is epigenetics https://www.whatisepigenetics.com/what-is-epigenetics/ -5. Centre for Research into Genetic Imprinting official website (University of Southampton, UK, funded by the NIHR): https://www.southampton.ac.uk/geneticimprinting
14












