







Limbile
Pagini
Legal

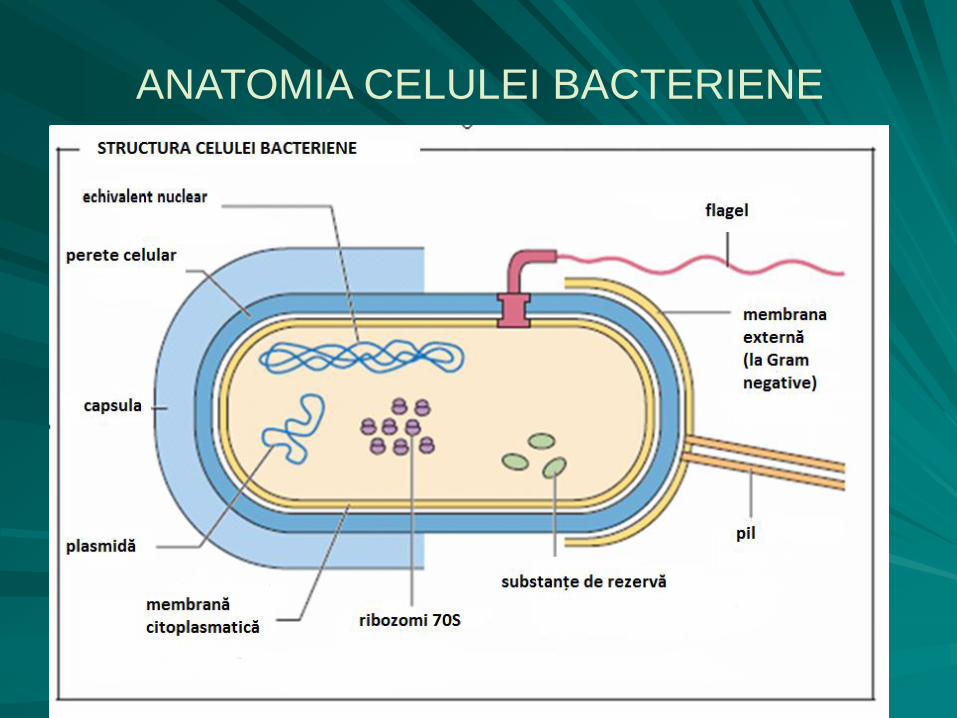
ANATOMIA CELULEI BACTERIENE

ANATOMIA CELULEI BACTERIENE
constituenţii celulari, în funcţie de poziţionarea lor faţă de peretele celular, sunt:
extraparietali:
capsula bacteriană,
stratul mucos,
flagelii,
pilii,
fimbriile,
intraparietali:
o membrana citoplasmatică,
o mezosomii,
o citoplasma,
o nucleosomul,
o plasmidele,
o ribozomii,
o vacuolele,
o incluziile,
o endosporul.
organizarea internă a bacteriilor a fost mult timp controversată, existând opinia că celulele bacteriene ar fi lipsite de o structură internă sau ar avea una foarte rudimentară;

ANATOMIA CELULEI BACTERIENE
Structurile celulei bacteriene pot fi:
– esenţiale pentru viaţa celulei, prezente invariabil la toate
celulele bacteriene (membrana citoplasmatică,
citoplasma, nucleosomul, ribozomii);
– accesorii, pot lipsi la unele bacterii (flagelul, plasmidele,
endosporul) sau apar doar în unele perioade de viaţă ale
celulei (incluziile, vacuolele, endosporul).

PERETELE CELULAR BACTERIAN
structură de înveliş bine definită, în general rigidă, care delimitează celula
bacteriană;
20 – 40% din greutatea uscată a celulei bacteriene, cu o grosime de 15 –
35nm;
În funcţie de prezenţa şi particularităţile de structură ale peretelui celular, în
regnul Prokaryotae se disting 4 categorii de bacterii:
– Firmacutes (de la firmus (lat.) = tare, cutis = înveliş), cu bacterii cu perete celular gros, rigid, lipsit de o membrană externă, caracteristic bacteriilor Gram (+);
– Gracilicutes (gracilis (lat.) = subţire, fin), cu bacterii cu perete celular subţire, cu o membrană externă, caracteristic bacteriilor Gram (-);
– Mollicutes sau Tenericutes (mollis, tener (lat.) = moale) cu bacterii din grupul Mycoplasma, lipsite de perete celular, la care membrana citoplasmatică ce conţine steroli devine înveliş extern;
– Mendosicutes (mendosus = o structură cu defecte), care cuprinde organismele din grupul Archaea, cu un perete celular atipic, din care lipseşte mureina, având în compoziţia lor chimică pseudomureină, ce le permite să trăiască în condiţii extreme de mediu.

PERETELE CELULAR BACTERIAN
LA BACTERIILE GRAM POZITIVE
gros (15 – 30 nm), rigid, alcătuit din mureina (peptidoglican, glicopeptid, mucopeptid, glucozaminopeptid), sensibilă la lizozim:
– Componenta glicanică - lanţuri paralele polizaharidice;
– Componenta peptidică este un tetrapeptid, cu o compoziţie în aminoacizi variabilă în funcţie de specie, care conţine L-alanină, D-alanină, D-glutamină, L-lizină;
la exteriorul stratului de mureină sunt prezenți acizii teichoici (teichos (gr.) = zid), polizaharide specifice, sub forma unor molecule polimerice lungi, flexibile, cu aspect tubular.
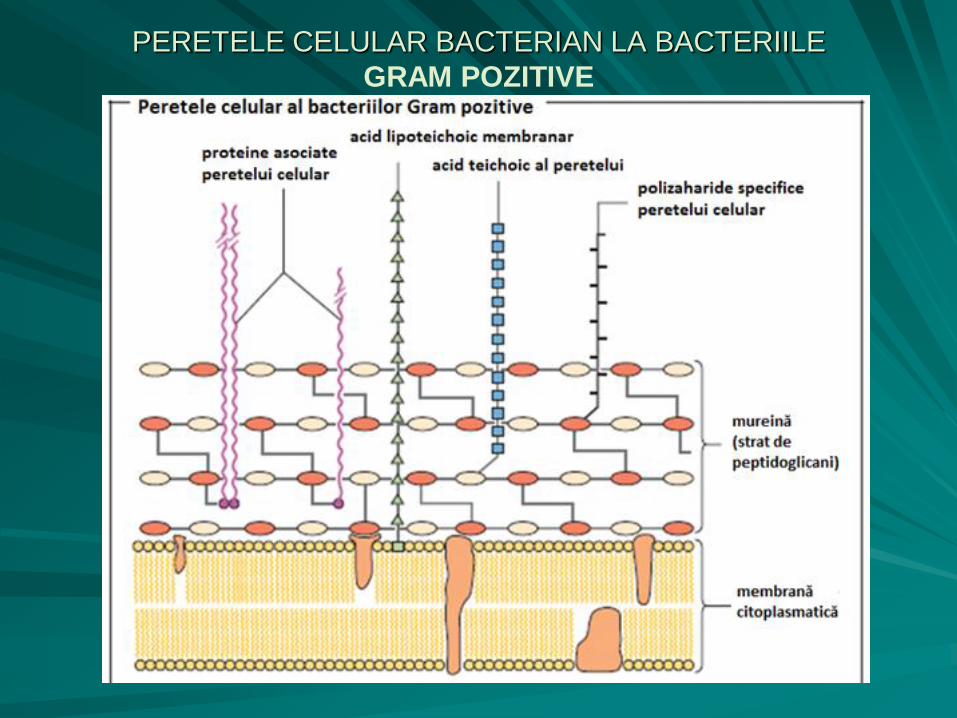
PERETELE CELULAR BACTERIAN LA BACTERIILE
GRAM POZITIVE

PERETELE CELULAR BACTERIAN
LA BACTERIILE GRAM NEGATIVE
subţire (8 – 12 nm), cu un strat fin de mureină sensibil la lizozim;
componenta peptidică a peretelui bacteriilor Gram negative conţine acid diaminopimelic (derivat al lizinei);
stratul de peptidoglicani este aderent la membrana citoplasmatică formând împreună cu aceasta membrana internă, la exterior stratul de peptidoglicani prezintă o membrană externă, cu o grosime de 6 – 20 nm, care nu este sensibilă la lizozim, ci la EDTA (etilen-diamino-tetraacetat) 30%;
între cele două membrane se delimitează spaţiul periplasmic
Membrana externă, cu o structură similară cu cea a membranei citoplasmatice, cuprinde:
– fosfolipide (35%) - dublu strat;
– proteine (15%) - inclavate în dublul strat fosfolipidic;
– lipopolizaharide (50%) (LPS) - prezintă trei porţiuni:
lipidul A, legat de stratul de fosfolipide;
componenta oligozaharidică (porţiunea centrală R);
componenta polizaharidică (antigenul somatic O), variabilă ca structură şi compoziţie chimică, determină specificitatea antigenică a celulei bacteriene.
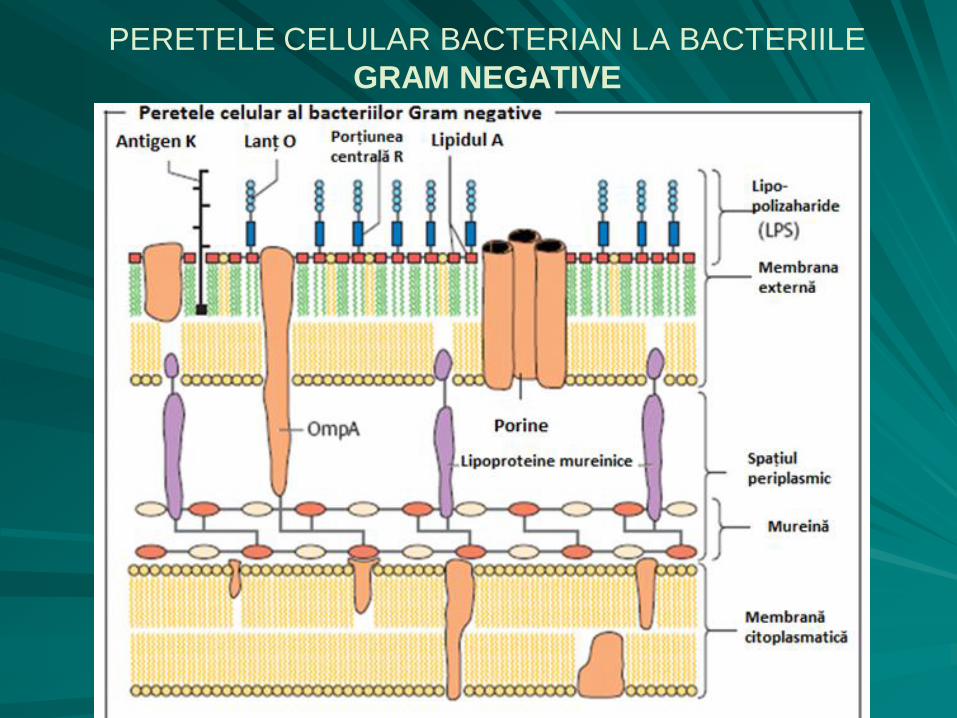
PERETELE CELULAR BACTERIAN LA BACTERIILE
GRAM NEGATIVE
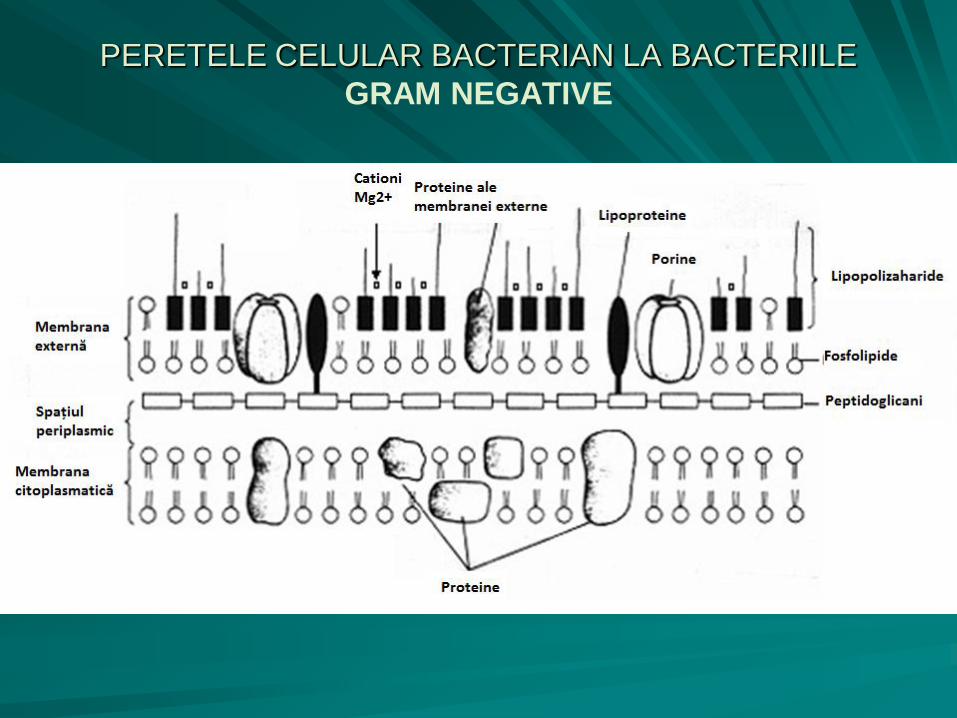
PERETELE CELULAR BACTERIAN LA BACTERIILE
GRAM NEGATIVE

FUNCŢIILE PERETELUI CELULAR BACTERIAN
menţine arhitectura structurală a celulei şi forma acesteia;
participă la procesele de creştere şi multiplicare bacteriană;
participă la procesul de sporogeneză;
funcţionează ca o barieră de permeabilitate între interiorul şi exteriorul celulei.
Acizii teichoici :
– asigură celulelor bacteriene Gram pozitive concentraţii adecvate de ioni metalici în micromediul de la suprafaţa celulei;
– inhibă fagocitoza şi măresc gradul de patogenitate al bacteriilor Gram pozitive patogene;
– acţionează ca receptori pentru bacteriofagi.
Membrana externă:
– scade capacitatea de fagocitoză a celulelor gazdă faţă de bacteriile Gram negative patogene;
– prin intermediul porinelor asigură trecerea unor substanţe din mediul extracelular în cel intracelular;
– antigenul somatic O conferă celulelor bacteriene specificitate antigenică;
– lipopolizaharidele au rol de endotoxină, răspunzătoare de efectele nocive asupra celulelor gazdă.

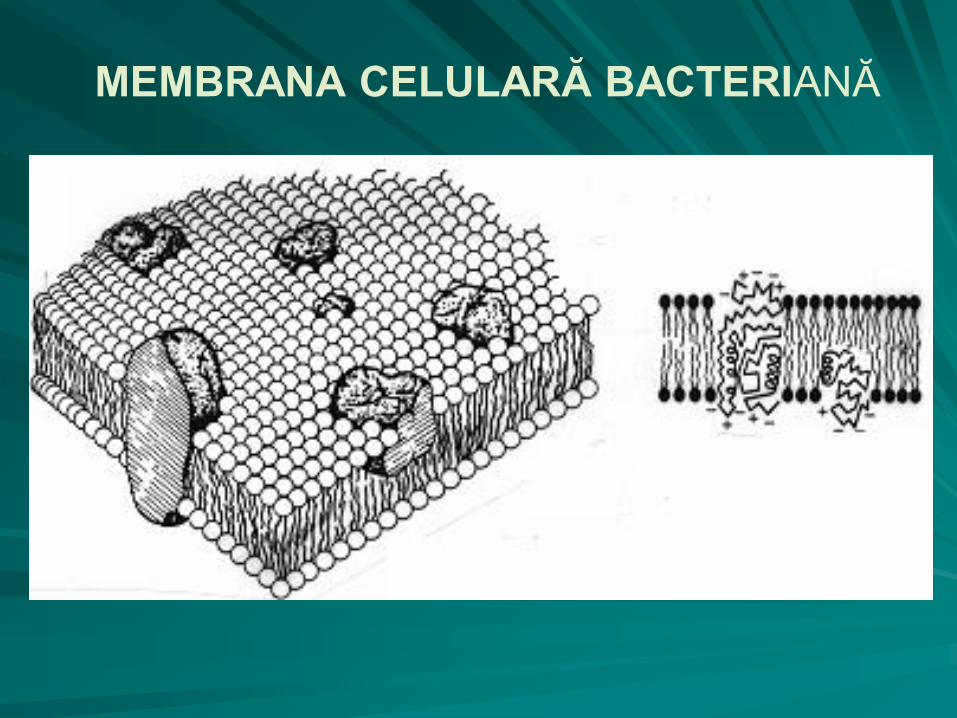
MEMBRANA CITOPLASMATICĂ
formaţiune structurală ce acoperă de jur împrejur citoplasma bacteriană, separând-o de suprafaţa internă a peretelui celular;
grosimea membranei este de 7 – 10 nm;
Singer şi Nicolson au propus în 1972 modelul mozaicului fluid pentru a descrie structura membranei plasmatice („un ocean de fosfolipide pe care plutesc iceberguri de proteine”).
MEMBRANA CELULARĂ BACTERIANĂ

MEMBRANA CITOPLASMATICĂ Structura membranei:
– Fosfolipidele – în dublu strat, cu grupările:
hidrofobe nepolare faţă în faţă, reprezentate de doi acizi graşi;
hidrofile polare, reprezentate de o grupare fosfat, orientate spre mediile apoase externe sau interne ale celulei;
– Proteinele - inclavate în dublul strat de fosfolipide; pot fi:
proteine integrate: transmembranare şi structurale;
proteine periferice (de suprafaţă), care sunt enzime active către peretele celular sau către citoplasmă;
– Glucidele - slab reprezentate în structura membranei plasmatice, sub formă de:
glicoproteine,
glicolipide.

ROLUL MEMBRANEI PLASMATICE
transportul substanţelor de la exteriorul celulei către interiorul acesteia şi în sens invers; este o barieră osmotică prin permeabilitatea pentru unele tipuri de molecule şi impermeabilitatea pentru altele.
echivalentul funcţional al mitocondriilor şi al cloroplastelor;
intervine în biosinteza peretelui celular;
intervine în mobilitatea orientată a bacteriei prin recepţionarea mesajelor chimice din mediu, determinate de substanţe repelente sau atractante;
intervine în sinteza şi eliminarea unor exoenzime şi a unor proteine toxice bacteriene (exotoxine).

CITOPLASMA BACTERIANĂ
sistem coloidal complex, corespunde stării fizice de gel;
formată din proteine, glucide, lipide, apă şi substanţe minerale;
în citoplasmă se găsesc: materialul nucleosomal, mezosomii, ribozomii, incluziile, vacuolele;
starea de gel permanent are ca rezultat o imobilitate a conţinutului celular în lipsa curenţilor citoplasmatici şi reprezintă o condiţie indispensabilă a nemiscibilităţii nucleosomului cu citoplasma, având în vedere lipsa membranelor intracelulare;
aspectul citoplasmei variază în funcţie de vârsta celulei bacteriene şi de condiţiile de cultivare:
– la celulele tinere (culturi de 24 – 48 h) şi în condiţii favorabile de mediu, citoplasma este densă, omogenă, fin granulară (datorită ribozomilor);
– la celulele îmbătrânite şi în condiţii nefavorabile de mediu, citoplasma îşi pierde treptat afinitatea pentru coloranţii bazici şi capătă un aspect granular, neomogen, cu multiple vacuole.

NUCLEOSOMUL BACTERIAN Materialul genetic bacterian:
– Nucleosomul (nucleoid, material nuclear, echivalent nuclear, genofor), corespunde din punct de vedere funcţional cromozomului;
– plasmidele, care poartă gene accesorii.
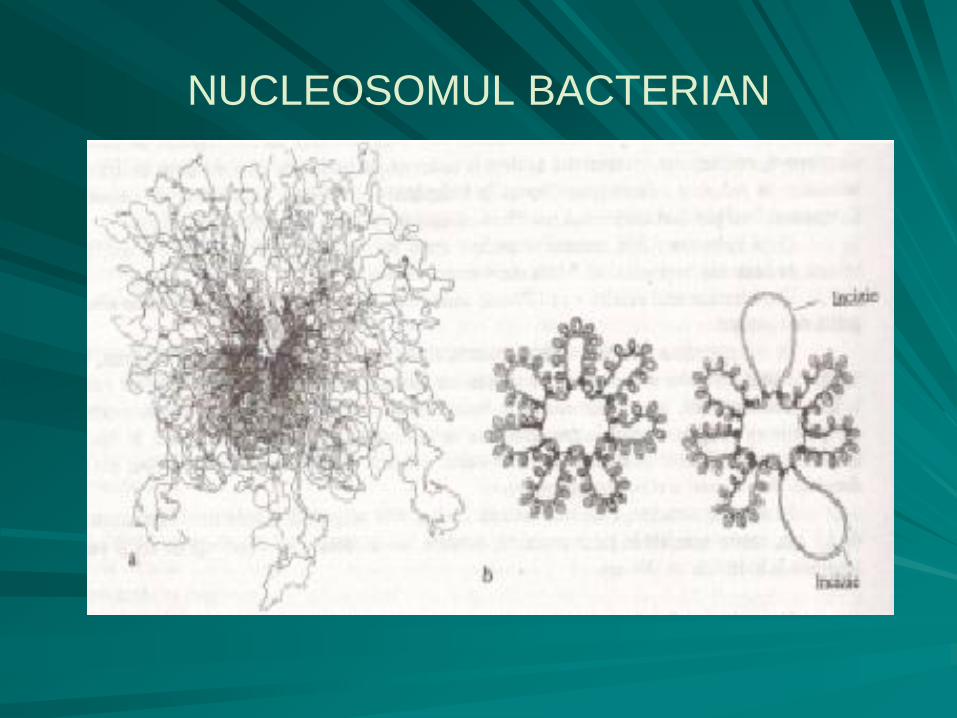
Nucleosomul bacterian
formă primitivă de organizare a nucleului, lipsită de membrană nucleară, inclavată direct în citoplasmă, într-o zonă numită nucleoplasmă, care este mai clară, în raport cu citoplasma propriu-zisă, care este mai densă;
extras prin metode speciale din celula bacteriană, sub forma unui corpuscul dens şi compact, nucleosomul s-a dovedit a fi alcătuit dintr-o singură moleculă de ADN, dublu catenară, circulară, covalent închisă, cu o lungime de aproximativ 1400µm şi diametrul de 2,5nm.
este cea mai mare moleculă biologică şi prin lungimea sa depăşeşte de 1000 de ori lungimea celulei bacteriene.
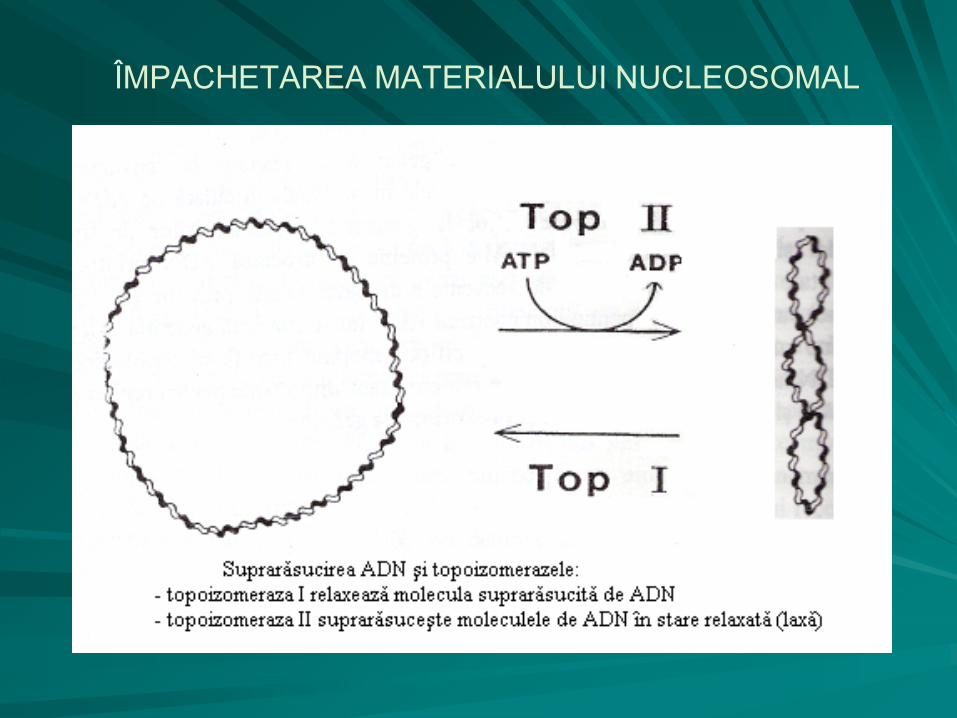
în structura dublu catenară a nucleoidului intervin modificări topologice, care constau într-un proces de împachetare prin suprarăsucire (supraspiralizare) după norme extrem de riguroase, ocupând astfel un volum de 1500 de ori mai mic.
NUCLEOSOMUL BACTERIAN
ÎMPACHETAREA MATERIALULUI NUCLEOSOMAL

FUNCŢIILE MATERIALULUI GENETIC
nucleosomul conţine informaţia genetică esenţială necesară existenţei celulelor bacteriene în mediul lor natural;
cuprinde aproximativ 3000 – 4000 gene, ce asigură: formarea structurilor celulare bacteriene,
metabolismul energetic şi biosintetic,
reglarea activităţilor celulare,
replicarea celulei bacteriene,
potenţialul de evoluţie a celulei.
pe medii neobişnuite, celulele bacteriene prezintă, pe lângă nucleosom, o informaţie genetică accesorie, reprezentată de plasmide: molecule de ADN dublu catenar, circular, covalent închis, numite şi minicromozomi, reprezentând 1 – 2% din mărimea nucleosomului bacterian, cu un număr mic de gene.

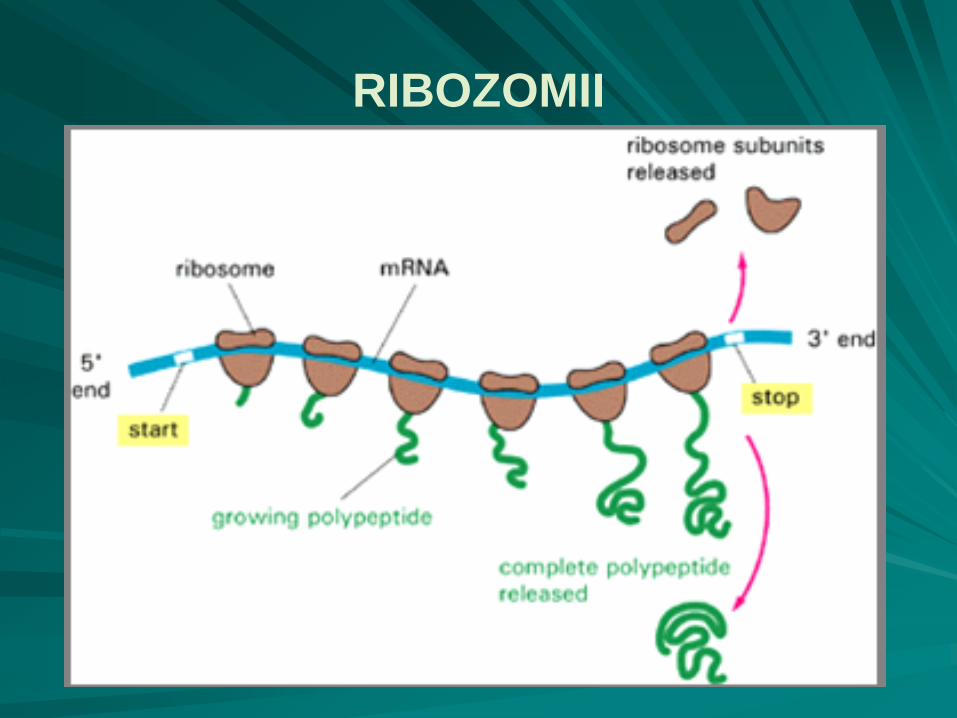
RIBOZOMII
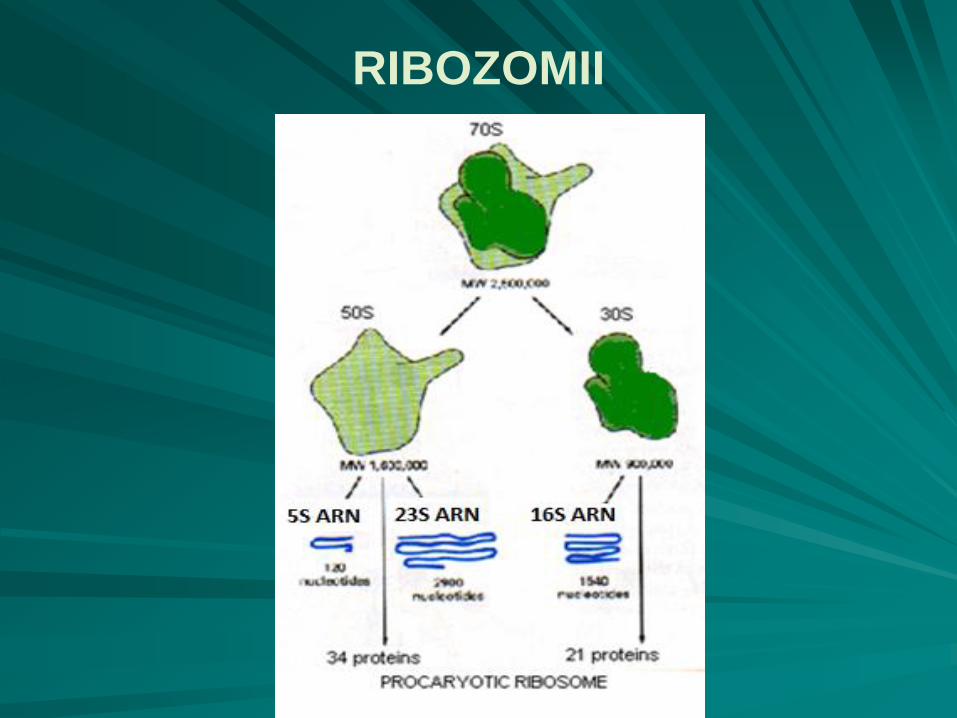
particule nucleoproteice intracitoplasmatice, de formă aproximativ sferică şi cu un diametru de 20nm, cu constanta de sedimentare 70S;
numărul ribozomilor în celula bacteriană este corelat cu activitatea ei fiziologică, mic în celulele în repaus şi mare în celulele fiziologic active, variază între 15000 – 100000/celulă;
sunt constituenţi esenţiali ai sistemului de traducere a informaţiei genetice, reprezentând adevărate fabrici de proteine, la nivelul lor desfăşurându-se ciclul ribozomal, ce determină iniţierea, creşterea şi terminalizarea lanţului polipeptidic;
ribozomii bacterieni 70S au două subunităţi:
o subunitate mare, 50S, cu formă de fotoliu
o subunitate mică, 30S, cu formă de halteră asimetrică, aşezată orizontal pe braţele şi spătarul fotoliului;
ribozomii 70S conţin 55 molecule de proteine şi 3 molecule de ARN ribosomal:
subunitatea mare - 34 molecule diferite de proteine (L1 - L34) şi două molecule de ARNr: 23S, 5S;
subunitatea mică - 21 molecule proteice (S1 - S21) şi o moleculă de ARNr, cu o constantă de sedimentare 16S.
RIBOZOMII
RIBOZOMII

INCLUZIILE BACTERIENE
formaţiuni structurale inerte, care apar în citoplasma bacteriilor aflate la sfârşitul perioadei de creştere activă, de regulă în condiţii de abundenţă a surselor nutritive şi în prezenţa unui dezechilibru între carbon şi azot;
reprezintă rezerve intracelulare pentru microorganisme, care dispar după trecerea celulelor respective în medii sărace în nutrienţi;

ENDOSPORUL BACTERIAN sporul bacterian este o formă primitivă de diferenţiere celulară, care constă în formarea în celula vegetativă a unui nou tip de celulă, cu ultrastructură, compoziţie chimică şi enzimatică diferită;
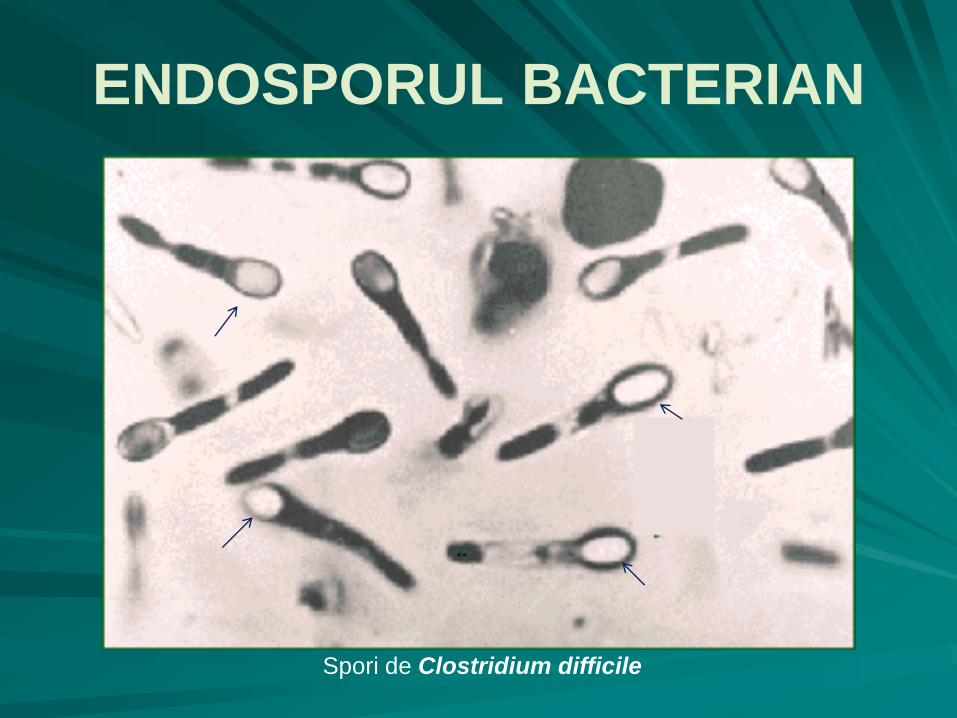
cea mai comună formă de spor – endosporul - sporul tipic, e frecvent întâlnit la bacteriile Gram pozitive cilindrice (de exemplu la genurile Bacillus, Clostridium) şi în mod excepţional la bacterii sferice (Sporosarcina);
are formă sferică sau ovalară, o refringenţă deosebită şi e foarte greu colorabil, altfel decât prin tehnici speciale energice;
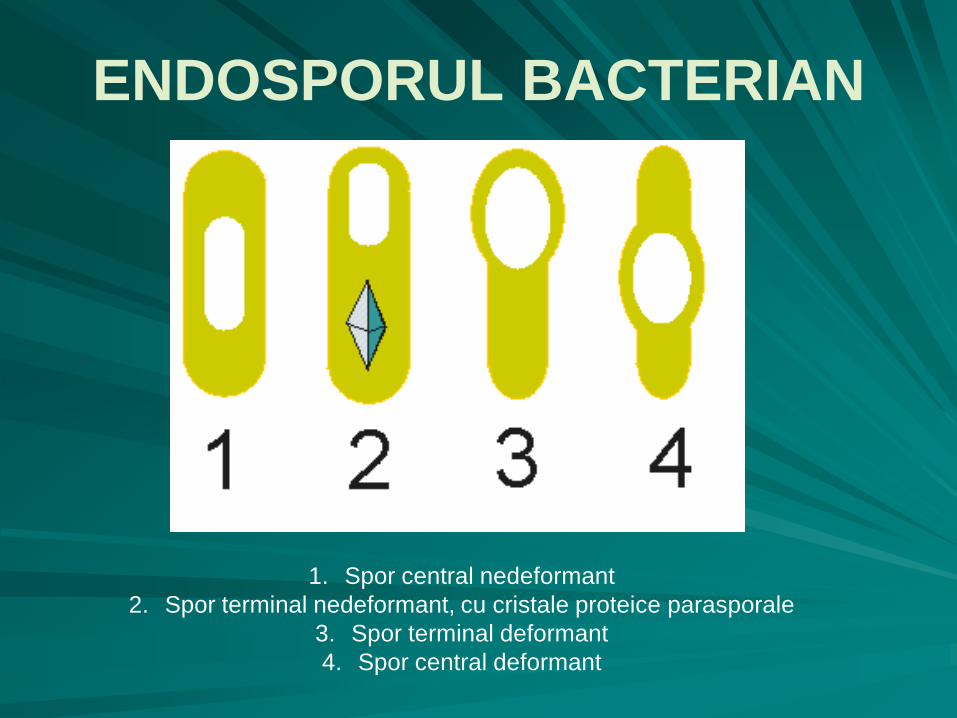
sporul poate fi:
terminal,
subterminal
ecuatorial (central)
nedeformant (terminal, subterminal, central)
deformant (terminal, ecuatorial).
ENDOSPORUL BACTERIAN
1. Spor central nedeformant
2. Spor terminal nedeformant, cu cristale proteice parasporale
3. Spor terminal deformant
4. Spor central deformant
ENDOSPORUL BACTERIAN
Spori de Clostridium difficile

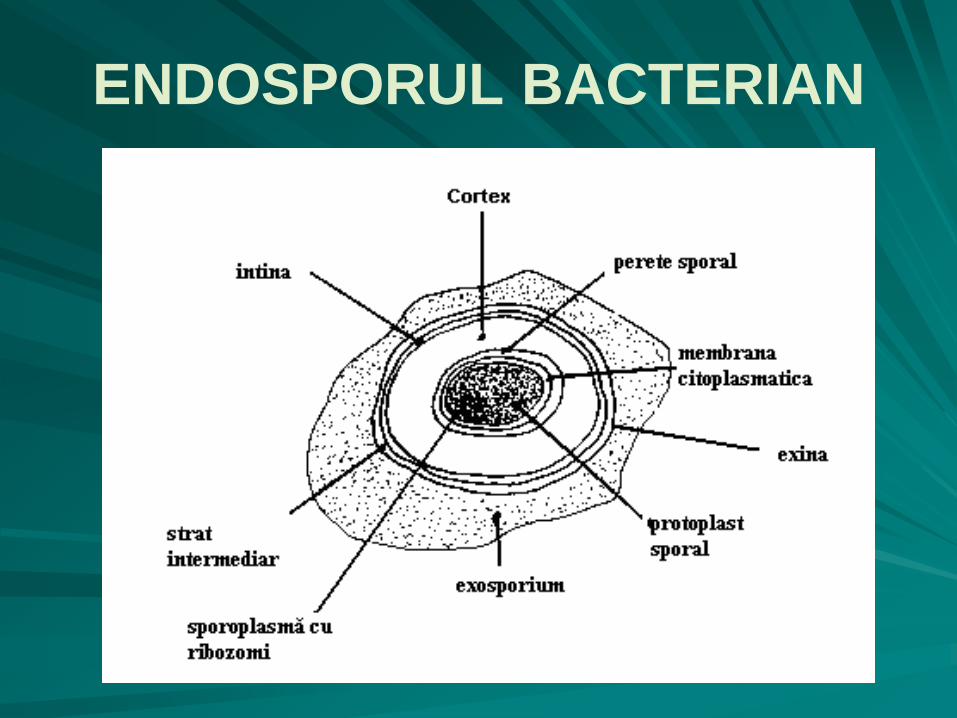
ENDOSPORUL BACTERIAN o porţiune centrală - protoplastul sporal, format din:
perete sporal,
citoplasmă granulară denumită sporoplasmă, în care se găsesc:
ribozomii
materialul nuclear – nucleoplasmă
o zonă transparentă - cortex, alcătuită din peptidoglican modificat;
un înveliş sporal lamelar cu 3 straturi suprapuse:
la interior este situată intina bistratificată;
un strat mijlociu;
exina multistratificată.
la unele bacterii există în jurul sporului un exosporium, uneori multistratificat, ancorat de spor prin filamente suspensoare;
unii spori au la una dintre extremităţi un smoc de apendici sporali, de aproximativ două – trei ori mai lungi decât sporul, cu aspect tubular şi cu striaţii orizontale sau în formă de pană.
ENDOSPORUL BACTERIAN

ENDOSPORUL BACTERIAN Compoziţia chimică a sporului
deosebirile dintre spor şi celula vegetativă nu sunt de ordin cantitativ, ci calitativ, referindu-se la starea apei: apa liberă din spor este în cantitate foarte redusă, între 3 – 10%, iar restul de 90 – 97% este în stare legată de constituenţii celulari, apa legată nu este implicată în procese metabolice; sporul este în stare de viaţă latentă (criptobioză), fără metabolism sau cu un metabolism extrem de redus; în celula vegetativă 70% din apă este în stare liberă, implicată în diverse procese metabolice;
particularităţile de structură şi compoziţie chimică a sporilor determină termorezistenţa acestora, deosebita rezistenţă la substanţele chimice (antiseptice, dezinfectanţi), rezistenţa la radiaţii.

ENDOSPORUL BACTERIAN
Sporogeneza - procesul de formare a sporilor, care se desfăşoară în 6 – 7 stadii succesive şi durează aproximativ 8 ore;
Germinarea sporilor - procesul de conversie a sporului în celula vegetativă, are loc atunci când sporul întâlneşte condiţii favorabile de mediu;
Δ Endosporul este şi o formă de conservare viabilităţii celulelor şi deci a speciei, nefiind o formă de multiplicare.
Δ Are rol în diseminarea celulelor bacteriene în natură, deoarece pot fi răspândiţi la distanţe foarte mari şi au o longevitate deosebită.

CAPSULA BACTERIANĂ
structură accesorie, care acoperă de jur împrejur celula bacteriană;
material macromolecular, gelatinos, vâscos, inegal dezvoltat de la o specie la alta;
forme de capsulă:
Microcapsula - peliculă fină, cu o grosime mai mică de 0,2µm, care acoperă de jur împrejur peretele celulei bacteriene;
Macrocapsula - structură organizată, aderentă de celulă, cu o grosime mai mare de 0,2µm, demonstrabilă prin metode citologice;
Stratul mucos - masă amorfă neorganizată, cu o grosime mult mai mare de 0,2µm;
Zoogleea (zoon = animal; glee = substanţă vâscoasă) constă din acelaşi material ca şi stratul mucos, dar cuprinde mai multe celule bacteriene, formând o asociaţie bacteriană mucilaginoasă.

FLAGELUL BACTERIAN
structuri extracelulare, organite de motilitate, cu o structură rudimentară în raport cu flagelul eucariotelor;
lungimea flagelului este variabilă, de la 4 - 5µm până la 15µm, ajungând uneori până la 70µm, flagelii fiind mult mai lungi decât celula bacteriană;
prezenţa lor poate fi dedusă pe baza mobilităţii accentuate a bacteriilor, care se deplasează în câmpul microscopic la întâmplare, în diferite direcţii;
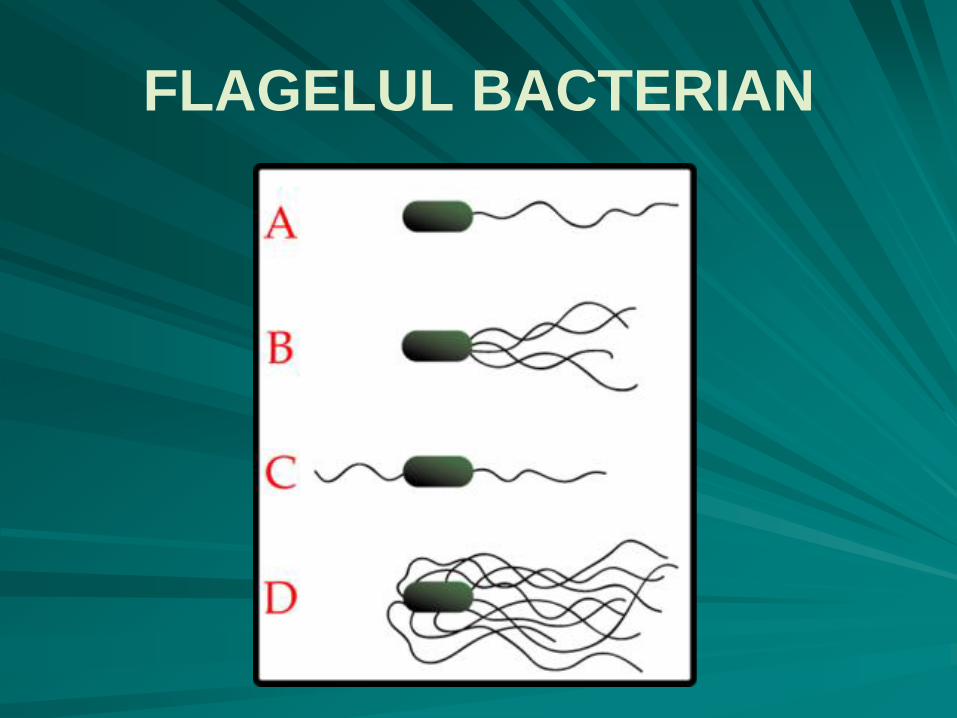
în funcţie de prezenţa sau absenţa flagelilor, numărul şi gruparea lor, au fost descrise mai multe tipuri de bacterii:
bacterii atriche (fără flagel)
bacterii monotriche (cu un singur flagel) - A
bacterii amfitriche (cu doi flageli, situaţi de obicei apical) - C
bacterii lofotriche (cu un smoc de flageli la unul din poli) - B
bacterii peritriche (cu flageli de jur împrejurul celulei) - D
FLAGELUL BACTERIAN

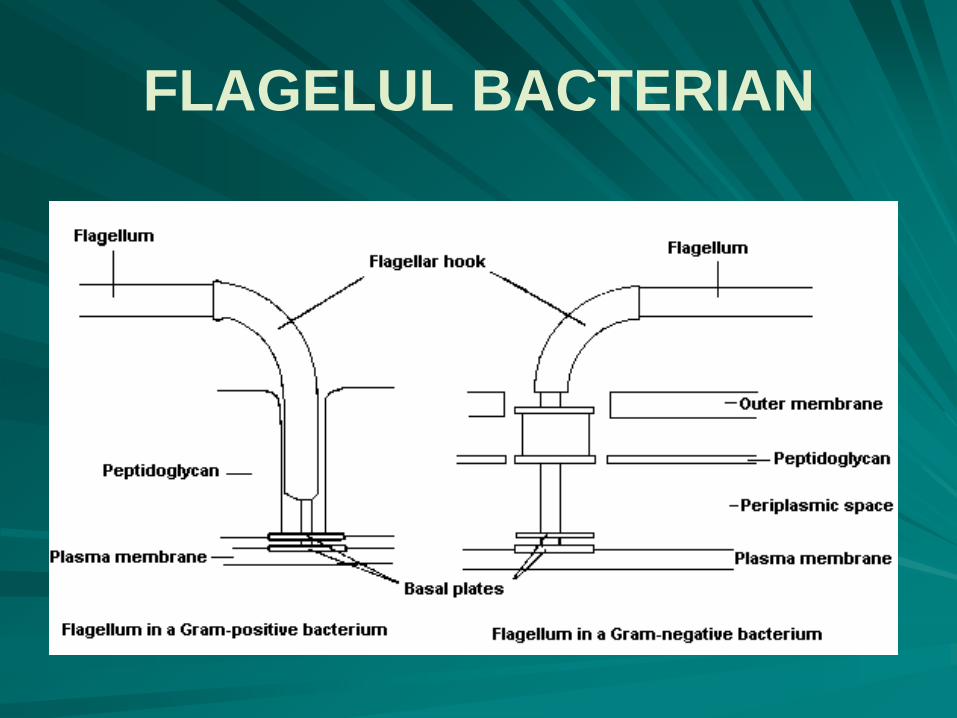
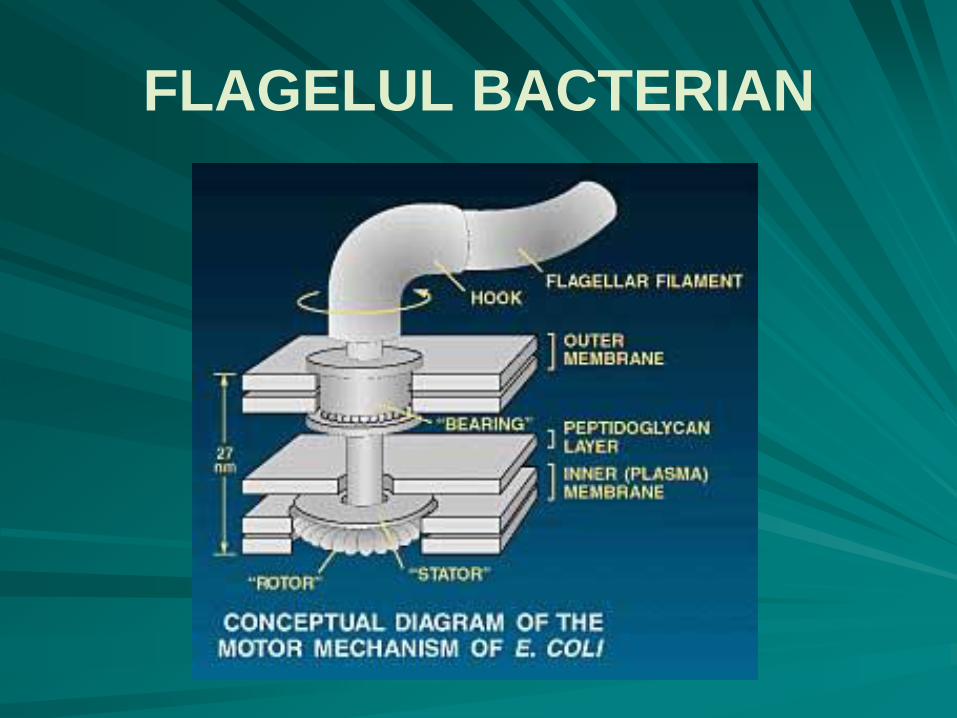
FLAGELUL BACTERIAN - ULTRASTRUCTURA
o porţiune bazală rotatorie, cu caracter de motor rotativ :
la bacteriile Gram pozitive porţiunea bazală e simplă, formată din două discuri unite între ele printr-un ax central, precum butonii unei manşete;
la bacteriile Gram negative porţiunea bazală este formată din mai multe discuri unite printr-un ax central:
discul M, la nivelul membranei plasmatice, disc mobil,
discul S, în regiunea supramembranară;
discul P, legat de lanţul de peptidoglicani;
discul L, la nivelul lipopolizaharidelor membranei externe a peretelui celular.
o porţiune în formă de cârlig, are caracter de articulaţie flexibilă universală, putându-se roti împreună cu filamentul cu 360˚;
extracelular flagelul propriu-zis, cu un diametru constant pe toată lungimea sa, de 20nm, cu mişcări de bici, nu ondulatorii.
FLAGELUL BACTERIAN
FLAGELUL BACTERIAN

FLAGELUL BACTERIAN
prezenţa flagelilor asigură deplasarea activă a bacteriilor în medii lichide;
în medii neutre din punct de vedere chimic bacteriile se deplasează la întâmplare: o secundă în linie dreaptă, apoi o rostogolire în 0,1 secunde cu schimbarea direcţiei de deplasare, urmată de o nouă deplasare în linie dreaptă timp de o secundă;
viteza de deplasare a bacteriilor este mai mare decât a macroorganismelor, raportată la dimensiunile proprii: bacteriile flagelate se deplasează cu 20-80µm pe secundă, ceea ce reprezintă o viteză de 40 lungimi/secundă.

FLAGELUL BACTERIAN
mişcarea orientată preferenţial a bacteriilor = taxie, în funcţie de:
– substanţele chimice din mediu (chimiotaxie):
substanţele chimiotactic pozitive = atractante
substanţele chimiotactic negative = repelente
– lumină (fototaxie),
– concentraţia de oxigen (aerotaxie),
– temperatură (termotaxie).
PILII
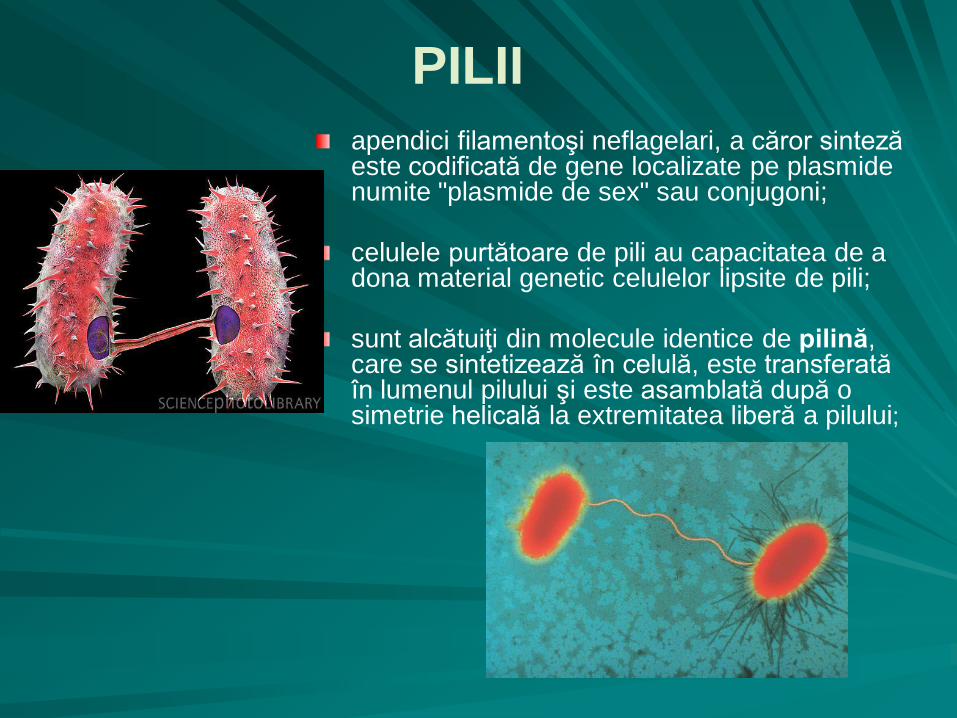
apendici filamentoşi neflagelari, a căror sinteză este codificată de gene localizate pe plasmide numite "plasmide de sex" sau conjugoni;
celulele purtătoare de pili au capacitatea de a dona material genetic celulelor lipsite de pili;
sunt alcătuiţi din molecule identice de pilină, care se sintetizează în celulă, este transferată în lumenul pilului şi este asamblată după o simetrie helicală la extremitatea liberă a pilului;

FIMBRIILE
structuri de tipul apendicelor filamentoase drepte şi rigide, mai scurte şi mai subţiri decât flagelii, sintetizate sub controlul unor gene situate pe cromozomul bacterian;
sunt alcătuite din molecule de fimbrilină aşezate după o simetrie helicală, determinând o structură tubulară;
sunt considerate structuri adaptative, cu rolul de a mări suprafaţa de contact a bacteriei cu mediul înconjurător, fiind utilizate pentru absorbţia nutrienţilor;
Top Related