
Virusologia Parazitologia Microbiologie Gheteu CURSURI Micro Primele 6 Final
46
VIRUSOLOGIE, MICROBIOLOGIE, PARAZITOLOGIE SUPORT DE CURS PENTRU SPECIALIZAREA MEDICINĂ DENTARĂ DIANA GHEŢEU Doctor în Biologie IAŞI 2011 UZ INTERN
-
Upload
ancuta-cpcp -
Category
Documents
-
view
64 -
download
6
Transcript of Virusologia Parazitologia Microbiologie Gheteu CURSURI Micro Primele 6 Final

VIRUSOLOGIE, MICROBIOLOGIE, PARAZITOLOGIE
SUPORT DE CURS PENTRU SPECIALIZAREA
MEDICINĂ DENTARĂ
DIANA GHEŢEU Doctor în Biologie
IAŞI 2011
UZ INTERN

Dr. biolog D. Gheţeu - Microbiologie
2
Cuprins I. Introducere în studiul organismelor patogene - metode clasice şi moderne de investigare.
Căi de transmitere ale organimelor patogene. Tipuri de infecţii II. Infecţia şi răspunsul imun
III. Bacteriile – morfologie, fiziologie, patogenitate. IV. Coci şi bacili patogeni V. V.1 Leptospirele. Rickettsiile.
V.2 Micologie - noţiuni generale. Dermatofiţii. VI. Candidoza şi actinomicoza
VII. Microflora normală. Mecanismele aderenţei, ocupării şi dezvoltării microorganismelor în cavitatea bucală
VIII. Microflora abcesului dentar. Microflora cariei dentare IX. Microbiologia plăcii dentare: formare şi interacţiuni microbiene. X. Noţiuni generale de parazitologie. Relaţia gazdă- parazit. Protozoonoze: agenţi etiologici,
transmitere şi combatere XI. Helmintoze: agenţi etiologici, transmitere şi combatere
XII. XII. 1 Noţiuni de entomologie medicală XII. 2Virusurile: structură, replicare.
XIII. Infecţiile virale cu virusurile: poliomielitei, herpetice, gripale, hepatice; HIV XIV. XIV. 1 Mecanisme de rezistenţă bacteriană la substanţe antibacteriene.
XIV.2 Vaccinoterapia în infecţiile virale Bibliografie 1. Baumgartner, J.C., Microbiological and molecular analysis of endodontic infections,
Endodontic Topics, 2004, 7, 35–51. 2. Buiuc, D., Neguţ, M., (sub redacţia), Tratat de microbiologie c1inică, Ed. Medicală,
1999. 3. Cajal, N., Tratat de virusologie, vol.l, Ed.Medicală, 1990. 4. Cemescu, C., Virusologie medicală, Ed.Medicală, 1998. 5. Duca, R., ş.a , Microbiologie medicală, Ed. Didactică, Bucureşti, 1979. 6. Georgescu, M., Microbiologie medicală şi imunologie fundamentală , I.M.F.,
Bucureşti, 1983. 7. Harley, J.P., Prescott, L.M., Laboratory Exercises in Microbiology, Fifth Edition,
Harley−Prescott: The McGraw−Hill Companies, 2002 8. Markell, Edward, David John, Wojciech Krotoski. Medical Parasitology. Philadelphia:
W.B Saunders, 1999. 9. Nitzulescu, I. Gherman, Parazitologie Medicală, Ed. Medicală, Bucureşti, 1986. 10. Pace, J.R. et al., Biofilms, Infection and Antimicrobial Therapy, Taylor & Francis
Group (Ed.) LLC, U.S., 2006. 11. Rădulescu, S., Meyer E., Diagnosticul bolilor parazitare. În Parazitologie medicală, Ed.
All, Bucureşti, 1992. 12. Shiba, C.R., et al., Atlas of medical parasitology, Kyokuseysia Co. Ltd., Japan, 2006 13. Titeica, M. (sub red), Practica laboratorului clinic – I.M.F., Bucureşti, 1981 14. Vojdani M., Zibaei MSc, Frequency of Bacteria and Fungi Isolated from Pumice in
Dental Laboratories, J Res Health Sci, 2006, Vol 6, No 1, pp. 33-38. 15. **Metode de laborator, vol. II, Ed. Medicală, Ministerul sănătăţii şi Academia de
Ştiinţe Medicale, Bucureşti, 1977 16. **Antibiograma difuzimetrică: tehnica de lucru şi citirea rezultatelor în „Imunologie” I.
Moraru, Ed. Medicală, Bucureşti, 1984

Dr. biolog D. Gheţeu - Microbiologie
3
Curs I
I. Introducere în studiul organismelor patogene - metode clasice şi moderne de investigare. Căi de transmitere ale organimelor patogene.
Tipuri de infecţii
MICROBIOLOGIA se ocupa cu studiul microorganismelor. MICROORGANISM / MICROB: bacterie, ciupercile microscopice, levurile, unele alge,
protozoare şi virusuri. MICROBIOLOGIA MEDICALĂ se ocupă cu studiul microorganismelor implicate în
patologia umană şi interacţiunile dintre om şi aceste microorganisme. Din punct de vedere biofizic, orice celulă vie este un complex autoorganizat de molecule
organice, care schimbă energie şi materie cu mediul inconjurator, capabil să crească şi are proprietatea de a se reproduce.
INFECŢIA desemnează procesul prin care agentul patogen pătrunde şi se multiplică în ţesuturile organismului-gazdă.
BOALA INFECŢIOASĂ defineşte prezenţa microorganismlelor şi modificările induse de acestea ca răspuns la agresiune, cu sau fără manifestări clinice.
Boala infecţioasă : este prezenţa microorganismlelor şi a modificărilor induse induse de acesta ca răspuns la agresiune, cu sau fără manifestări clinice.
Boala contagioasă: este acea afecţiune infecţioasă care se poate transmite pe diferite căi de la sursa de infecţie, indiferent de forma sa de manifestare (aparentă, subclinică, inaparentă, purtător) la un individ receptiv. Deci procesul infecţios nu este neapărat şi contagios. Exemple: tetanos, botulism, septicemiile – sunt boli infecţioase, însă netransmisibile.
VIRULENŢA = capacitatea unui agent patogen de a se înmulţi în ţesuturile afectate. Procesul infecţios este pătrunderea microorganismelor în organismul gazdă, urmată de
fixarea şi multiplicarea lor în ţesuturile acestuia. Postulatele lui Koch-Henle definesc încă într-o formă actuală agentul patogen infecţios:
poate fi pus în evidenţă mereu într-o boală infecţioasă; agentul patogen poate fi cultivat (izolat) şi în afara organismului bolnav; infectarea unui animal de laborator adecvat cu un agent microbian cultivat, trebuie să prezinte un tablou clinic tipic. Agentul microbian trebuie să poată fi din nou izolat din animalul infectat. Procesul infecţios cuantifică cel mai exact complexitatea fenomenuluiinfecţios.
Figura 1. Schema procesului infecţios.
Macroorganismul : apărare
Bariere naturale anatomice şi fiziologice
Mijloace de apărare specifice şi nespecifice
Microorganismul (antigen, enzime, toxice)
Se ataşează
Se multiplică
Modificări metabolice, morfologice, funcţionale şi clinice

Dr. biolog D. Gheţeu - Microbiologie
4
Infecţiile exogene sunt infecţiile unde agentul infecţios pătrunde în corp din exterior, penetrarea se realizează prin: piele, mucoase, oral, uretral, vaginal şi anal. Pot exista infectii exogene excepţionale şi anume:
- infecţii intrauterine, - transplacentare - infecţia intrapartum. În mod normal mucoasele şi pielea posedă mecanisme locale şi sistemice de apărare:
strat protector, mucus, efectori umorali sau celulari ai imunităţii. Dacă organismul este slăbit datorită unor tulburări ale sistemului imun sau unor leziuni focale, pătrunderea agenţilor infecţioşi este uşurată. Zonele frecvente de penetrare ai agentilor infecţioşi sunt: foliculii piloşi, glandele sudoripare şi sebacee, minuscule leziuni ale pielii.
Infecţiile endogene • Sunt generate de germenii care sunt deja existenţi în organism. • Acestea apar la persoanele care au un sistem imunitar slabit, sau la persoane în vârstă. • Atât germenii virulenti cât şi cei nevirulenti se pot multiplica în organismul uman, fără a determina apariţia bolii. • Atunci când sistemul imun şi/ sau mecanismele de apărare locală sunt deficitare, aceşti gemeni pot deveni infectioşi.
Infecţiile emergente • Sunt boli transmisibile, recent identificate şi clasificate taxonomic şi clinico-epidemiologic. • La sfârşitul secolului XX erau peste 30 de astfel de boli care pot să declanşeze epidemii periculoase şi anume:
- infecţia cu HIV, - febra Ebola, - sindromul pulmonar cu Hanta virus,
- febre virale hemoragice, - infecţia cu Campylobacter, Helicobacter, - encefalopatiile spongiforme transmisibile, - boala legionarilor, - boala Lyme ( Borrelia burgdorferi).
• Unele sunt boli nou apărute, altele existau de secole şi au fost recunoscute doar recent din cauza unor schimbări ecologice sau ambientale care au crescut riscul infecţiilor umane.
Infecţiile reemergente • Sunt boli infecţioase vechi ca: tuberculoza şi sifilisul, care au înregistrat o recrudescenţă semnificativă din cauza schimbărilor complexului de condiţii gazdă-agent, etiologie-mediu.
Infecţiile intraspitaliceşti sunt afectiuni survenite în interiorul unui spital, afectând bolnavii spitalizaţi. Ele se mai numesc şi infectii nosocomiale sau iatrogene sau infecţii încrucişate (cross infection). Aceste infecţii apar în timpul spitalizării, mai rar după spitalizare, în conditiile create de spitalizare şi de actul medical sau chirurgical
Evolutiv infectia de spital a parcurs trei etape: - prepasteuriană (prevalenţă crescută şi predominanta etiologică a germenilor înat
patogeni), - pasteuriană (datorită implementării asepsiei şi antisepsiei a scăzut semnificativ
prevalenţa), - modernă şi contemporană cu prevalentă în creştere şi implicarea predominantă a
unor bacterii condiţionat patogene. Caracteristicile tulpinilor care dau aceste infecţii sunt: rezistenta multiplă la antibiotice şi antiseptice, capacitatea de colonizare şi virulentă crescută.
Cele mai întâlnite bacterii care dau aceste infecţii şi care alcătuiesc fondul microbian de spital sunt:
- enterobacteriile ( E. coli, Klebsiella pneumoniae, Enterobacter spp.), - Pseudomonas aeruginosa, - Staphylococcus aureus, stafilococi coagulazo-negativi;

Dr. biolog D. Gheţeu - Microbiologie
5
- virusuri cu tropism respirator (gripal, paragripal, adenovirusuri, rinovirusuri, rujeolic, varicelos mai rar);
- enterovirusurile (polio, Coxsachie, ECHO); - fungi (Candida, Aspergillius); - protozoare Pneumocystis carinii, - Toxoplasma în cursul travaliului.
Riscul infecţiei intraspitaliceşti este mărit prin investigatiile medicale sau chirurgicale agresive (puncţii exploratorii sau endoscopia cavităţilor) sau prin actele terapeutice moderne (respiraţie asistată, intervenţii chirurgicale agresive, perfuzii, cateterisme venoase prelungite, medicaţie imunosupresivă).
Metode de investigare Componentele structurale ale microorganismelor joacă rol de antigen, provoacă răspuns
imun din partea organismului infectat, ceea ce rezultă în producere de anticorpi. Reacţia antigen – anticorp este specifică, antigenul poate fi legat numai de anticorpul ce s-a produs ca răspuns la stimulul antigenic respectiv. Aceste reacţii pot fi folosite pentru detectarea unui antigen necunoscut cu ajutorul unui anticorp cunoscut (metode de diagnostic directe) sau detectarea anticorpilor din serul bolnavilor cu antigene cunoscute (serodiagnostic, metode de diagnostic indirecte).
În cazul în care antigenul corespunde anticorpului, se formează complexul imun antigen-anticorp. În funcţie de cum se vizualizează/detectează formarea complexului imun există mai multe metode bazate pe reacţii antigen-anticorp:
reacţii de precipitare: complexele imune formează precipitat reacţii de aglutinare: complexele imune formează aglutinat RFC – se detectează consumarea complementului (acesta se leagă de complexele imune) reacţii în care elementul cunoscut este marcat – se detectează elementul marcat din complexul imun format
1. Reacţii de precipitare. Pot avea loc în medii lichide (metode folosite în bacteriologie) sau în medii solide. Antigenele: de natură coloidală (solubile), precipitinogene Anticorpii: precipitine Complexul imun: precipitat
2. Reacţii de aglutinare Antigenele: de natură corpusculară, aglutinogene Anticorpii: aglutinine Antigenele corespund cu anticorpii şi sunt prezente în cantităţi echivalente: se formează aglutinatul (reţea tridimensională de complexe imune), vizibil cu ochiul liber sub forma de grunji.
Reacţia de hemaglutinare, hemaglutino-inhibare (RHA-HAI) se foloseşte pentru detectarea virusurilor care prezintă pe suprafaţă hemaglutinine (HA). Exemplu metoda Hirst: detectarea anticorpilor antihemaglutinină din serul bolnavilor de gripă convalescenţi
Metode moderne – metode care folosesc un reactant imunologic marcat 1. Reacţia de imunofluorescenţă (RIF). Fluorocromii se leagă covalent de proteine
(imunoglobuline) = conjugat; emit fluorescenţă dacă se expun la raze UV. Preparatul se examinează la microscopul cu fluorescenţă. Puncte fluorescente pe fond întunecat confirmă rezultat pozitiv.
2. Metode imunoenzimatice (RIE, ELISA - enzyme-linked immunosorbent assay) Permit detectarea antigenelor libere şi a anticorpilor. Este posibilă determinare cantitativă. Reacţiile au loc în aparate automate, semiautomate.La reacţii participă: - un reactant imunologic cunoscut ataşat de un suport solid (plăci cu godeuri, lame, baghete - se pot folosi antigene, anticorpi, anticorpi monoclonali, proteina A - un reactant imunologic marcat enzimatic - peroxidaza

Dr. biolog D. Gheţeu - Microbiologie
6
- substrat specific (cromogenic). Se produce o modificare de culoare detectabilă spectrofotometric (determinare cantitativă) - substanţe pentru stoparea reacţiei (baze sau acizi puternici).
Identificarea microorganismelor prin detectarea acizilor nucleici Genomul conţine secvenţe specifice pentru fiecare microorganism, detectarea prezenţei
acestora într-un produs patologic sau în culturi celulare având valoare diagnostică. 1. Metode de hibridizare. Principiu: fragmente monocatenare de oligonucleotide
marcate (sonde), având secvenţă cunoscută (caracteristică unui virus) se vor atasa pe baza complementarităţii de secvenţe complementare situate pe acizii nucleici ale microorganismului de identificat.
Etape de lucru: - extragerea acizilor nucleici - denaturarea acizilor nucleici si fixarea acestora pe un suport - hibridizare: adăugarea sondelor (marcate enzimatic, cu fluorocrom sau cu substanŃe radioactive) si atasarea acestora de secvenţele omoloage - eliminarea sondelor nefixate - vizualizarea hibridizării. Vizualizarea se face în funcţie de tipul marcajului folosit.
Ex.1 Metoda Southern blot detectează secvenţe specifice de nucleotizi de ADN. Etape: - extragerea ADN-ului - denaturarea ADN-ului – se obŃin lanŃuri monocatenare - fragmentarea lanŃurilor cu enzime de restricŃie (acestea acŃionează în locusuri determinate, rezultând fragmente de lungime diferită, caracteristice) denaturare - separarea electroforetică a fragmentelor în gel de agaroză (separarea se face în funcŃie de mărimea fragmentelor) - transferarea pe membrană de nitroceluloză - adăugarea sondelor marcate cu substanŃe radioactive – atasarea de secvenŃele omoloage - se îndepărtează sondele în exces, nelegate - vizualizare prin autoradiografie
EX. 2. Northern blot – detectează ARN, este necesară revers-transcriptaza Ex. 3 Western blot – detectează proteine.
2. Reacţia în lanţ a polimerazei (PCR – polimerase chain reaction) Se detectează secvenţe specifice de acid nucleic după o prealabilă amplificare.
Etapele de lucru - detectarea ADN-ului - extragerea ADN-ului - amplificare: se realizează în aparatul PCR si constă din mai multe cicluri identice - denaturare (900C) - alinierea primerilor (500C) – 2 secvenŃe de oligonucleotide care se leagă prin complementaritate pe cele două catene la o anumită distanŃă între ele. - sinteza lanţurilor complementare (700C ) – elongaŃie; este necesară prezenŃa enzimei polimerază (Taq) si a nucleotidelor – se formează lanŃurile complementare astfel se dublează cantitatea de ADN - reluarea ciclului de n ori – se obŃin 2n copii ale secvenŃei de identificat - vizualizarea produsilor de amplificare - produsii de amplificare obŃinuŃi se separă electroforetic - colorare cu bromură de etidiu - vizualizare cu raze UV – prelucrare computerizată
Multiplex PCR – se amplifică mai multe gene odată Real time PCR RT-PCR - este necesară revers-transcriptaza

Dr. biolog D. Gheţeu - Microbiologie
7
Pentru studiul bacteriilor şi al fungilor, afinitatea tinctorială este un criteriu important de clasificare. Se folosesc în mod obişnuit două coloraţii principale şi anume: coloraţia albastru de metilen şi coloratia Gram, la care se poate adăuga (în funcţie de diagnostic) coloraţia Ziehl-Neelsen. Realizată de Christian Gram, bacteriolog danez, în 1884, este o coloraţie diferenţiată, foloseşte doi coloranţi de culori controstante (violet de genţiană, pentru colorarea iniţială şi fucsină bazică pentru recolorare), un mordant (soluţia Lugol) şi o soluţie de diferenţiere, decolorare (alcool-acetonă). Coloraţia Gram împarte toate bacteriile în Gram-pozitive şi Gram-negative în raport cu permeabilitatea diferită a peretelui dependentă de structura lor diferită.
Sursa de agenţi patogeni este reprezentată de un organism care are capacitatea să găzduiască, sa asigure supravieţuirea şi eventual multiplicarea şi întotdeauna diseminarea unui agent patogen, direct spre organismul receptiv sau indirect prin intermediul căilor de transmitere, prezentând sau nu semne clinice de boala.
Exemple de surse de agenti patogeni: - bolnavi cu forme tipice sau atipice de boală; - purtători de agenţi patogeni care pot să nu prezinte semne clinice de boală, dar pot fi:
- purtători preinfectioşi, viitorii bolnavi aflaţi în perioada de incubaţie a bolii; - purtători sănătoşi care au un anumit grad de imunitate şi o rezistentă generală nespecifică ridicată şi aceştia pot fi:
- temporari (câteva zile sau săptămâni), - cronici (luni, ani sau toată viaţa).
- purtători foşti bolnavi (după vindecarea clinică nu se sterilizează) - cunoscuţi (cei care au fost spitalizaţi) sau - necunoscuţi (cei cu forme atipice care au fost trataţi în ambulatoriu). - animale bolnave (inclusiv păsările) ele pot fi: - domestice, - peridomestice, - sălbatice.
Animalele pot transmite antropozoonozele (90 de maladii dintre care 20 interesează şi populaţia Europei). Bolile infecţioase la majoritatea animalele purtătoare de agenţi patogeni au aceleaşi caracteristici ca şi la oameni (stare febrilă, cahexie, modificarea constantelor sangvine etc.). Artropodele sunt vectori biologic activi care asigură în mod obligatoriu dezvoltarea cel puţin pentru un stadiu biologic al agentului patogen (ţântari, păduchele omului, căpuşele, puricii şoarecelui şi şobolanului etc.).
Modul de transmitere al organismelor patogene poate fi: • direct - se realizează între sursa de agent patogen şi organismul receptiv (inhalarea, contact fizic, sărutul, muşcătura, transferul transplacentar, transfuzia de sânge). • indirect - mai complex şi implică una sau mai multe căi de transmitere (apă, aer, sol, alimente, obiecte, mâini, insecte sau combinate).
Căi de transmitere ale organismelor patogene ► Aerul • Mecanismul de transmitere a agentului patogen prin această cale a fost descris de Pflügge in 1897. • În timpul expirării, aerul iese cu o presiune mare, produce picaturi septice care vor rămâne suspendate în aer pe o durată variabilă şi vor fi dispersate în raport cu dimensiunile lor (dispersia va fi intensă cand dimensiunile vor fi ≤ 10µ). • Picăturile septice sunt diseminate la o distantă de 1,5-2 m, sau dacă intervine tusea strănutul, vorbirea, cântarea, dispersia va fi la 5-10 m, datorită curentilor de aer. • După un timp picăturile septice pierd o parte din apa, devin mai uşoare, persistă mult in aer, sedimentează pe suprafeţe şi apoi se răspândesc în aer • Diseminând agenti patogeni aflaţi în interiorul masei de substanţe organice vor genera nucleosoli septici. • Ei se pot cupla cu particule minerale (praful din sol) şi generează praful septic, care se combină cu cele provenite din triturarea spontană a altor componente septice.

Dr. biolog D. Gheţeu - Microbiologie
8
• Organismele receptive se vor contamina şi eventual infecta inhalând: picături, nucleosoli sau praf septic.
- Aerul încăperilor poate fi contaminat cu intensitate variabilă în funcţie de: destinatia acestora, densitatea surselor de agenti patogeni, şi calitatea decontaminării periodice.
- Prin această cale se pot transmite: difteria, gripa, scarlatina, tuberculoza etc. ► Solul • Este o altă cale de transmitere a infecţiei, 1 gram de sol poate conţine 2-5x109germeni. • Solul nu este un mediu prielnic dezvoltării agenţilor patogeni. • In afară de bacteriile care sporulează, ceilalţi agenti patogeni supravieţuiesc o perioadă limitată de timp zile sau luni). • In sol se pot întâlni: fungi, ouă şi larve de geohelminţi. Contaminarea şi eventual infectarea omului se poate face prin:
- cale digestivă, consumă apă sau vegetale crude nespălate, - cale respiratorie prin inhalarea pulberilor din solul contaminat, - prin tegumente cu integritate pierdută.
Prin această cale se pot transmite: tetanosul, poliomielita, salmoneloza, tuberculoza, parazitozele. ► Apa • 4/5 din bolile cunoscute în lume sunt legate de insalubritatea apei. Transmiterea agentilor patogeni se poate face: • direct - de la sursele de agenţi patogeni: dejecte (umane, animale), adăparea animalelor bolnave, scăldarea (oamenilor sau animalelor), depunerea cadavrelor animalelor; • indirect are loc prin: - aer, sol, alimente, recipiente • Transmiterea indirectă se produce în: inundaţii, averse de ploaie, cutremure, cataclisme. • În aceste momente apar defectiuni, avarii, când nu se face sau este insuficientă decontaminarea apei sau utilizarea neigienică a surselor de apă. Prin această cale se pot transmite: febrele tifo-paratifoidice. dizenteria, holera, hepatita
A, poliomielita etc. ► Alimentele • Atunci când nu sunt prelucrate igienic sau atunci când nu se respectă circuitele. Contaminarea lor poate fi: - primară (de la origine) sau - secundară (prin dejecte, animale bolnave sau purtătoare, aer, apă, sol, insecte sau mâini murdare). Prin alimente se pot transmite: trichineloza, tuberculoza, antraxul, bruceloza, salmoneloze etc. ► Obiectele • Pot constitui o altă cale de transmitere. Ele pot transmite: - direct (de la sursa de agenţi patogeni prin produsele eliminate: secreţii, spută, urină, fecale, sânge, puroi) sau - indirect (prin aer, sol, apă, mâini). Prin această cale se pot transmite: salmoneloze, tuberculoza, hepatita A, parazitoze.
► Transmiterea prin mâini murdare. Contaminarea poate fi - directă (de la agenjii patogeni) sau - indirectă (prin germeni vehiculaţi prin: aer, apă, sol, alimente, obiecte, insecte). Se pot transmite: febră tifoidă, dizenterie, hepatita A, parazitoze.
► Insectele pot fi surse de agenţi patogeni, gazde obligatorii cel puţin pentru un stadiu de dezvoltare (păduchii omului, căpuşele, ţânţarii, puricii şi alţi flebotomi). • Altele sunt vectori pasivi, artropodele: muştele, gândacii de bucătărie, furnicile, după ce contaminează asigura doar transmiterea pe distanţe şi perioade variabile de timp.

Dr. biolog D. Gheţeu - Microbiologie
9
Receptivitatea (gazdei) • Este starea organismului care nu-i conferă acesteia posibilitatea de a învinge o agresiune microbiană. • Gradul de receptivitate a unei populaţii depinde de - rezistenţa specifică (imunitatea) care este dependentă de prezenţa anticorpilor sau a unor celule cu capacitate antiinfecţioasă. - rezistentă nespecifică (generală) este dependentă de: integritatea barierelor cutanate şi a mucoaselor. • Imunitatea naturală care se realizează fără intervenţia omului şi este de:
- specie – omul şi anumite animale fac anumite boli specifice; - pasivă - urmare a transferului transplacentar de anticorpi, - activă - imunitatea care apare după trecerea prin boală.
• Imunitatea artificială Se realizează după interventia omului, prin programe prevenţionale. • Se utilizează produse biologice care oferă protectie specifică. Ea poate fi:
- activă - după administrarea vaccinurilor; - pasivă - după administrarea de seruri imune sau de imunoglobuline (în acest caz
organismul este doar consumator). • Rezistenţa specifică sau nespecifică poate fi sporită prin utilizarea de: anticorpi monoclonali cu acţiune idiotipică şi a sistemelor imunomodulatoare (imunostimulatoare ribozomale cu acţiune de apărare antiinfectioasă). • Gradul de receptivitate depinde şi de : vârstă, alimentatie, profesie.
Manifestarea infecţiilor în colectivităţile umane În funcţie de: numărul, dispersia temporală şi spaţială a unor îmbolnăviri se disting patru
forme: sporadică, endemică, epidemică şi pandemică. 1. În manifestarea sporadică, procesele morbide apar sub forma unui număr mic de îmbolnăviri, distribuite pe un teritoriu întins, la intervale mari de timp fără o legătură aparentă între ele. 2. În manifestarea endemică, morbiditatea este redusă, îmbolnăvirile nu au legătură aparentă între ele, dar există tendinta de concentrare spatială şi temporală care pot duce la izbucnirea unor epidemii. • Ele se pot întâlni în mod constant în anumite grupe de populatii, din unele zone geografice, sub forma de cazuri dispersate care periodic pot creşte ca frecventă (hepatita virală, dizenteria bacilara, guşa endemică). • Prin luarea unor măsuri eficiente, o morbiditate endemică se poate transforma într-o morbiditate sporadică sau poate fi eradicată pe teritoriul respectiv. • Sau când nu se iau măsuri se poate transforma în morbiditate endemo-epidemică • Sau când nu se iau măsuri se poate transforma în morbiditate endemo-epidemică 3. Manifestarea epidemică se caracterizează prin: - natura debutului (exploziv, lent), - extensivitate, - severitate (complicatii şi mortalitate), - durata de evolutie (scurtă, medie sau lungă), - modul de stingere (brusc, lent sau trenant) şi - perioada interepidemică (scurtă, medie sau lungă). Ex. epidemii hidrice, aerogene, alimentare etc. 4. Manifestarea pandemică este o acumulare de epidemii cu caracter extensiv, interesează arii mari de pe glob sau continente întregi şi în toate cazurile de imbolnăviri există legături evidente.
Forme de exprimare : 1. Infecţia inaparentă : asimptomatică clinic, însă implică prezenţa agentului patogen şi a moficărilor imunologice. Exemple: poliomielita, hepatita acută virală, rubeola, infecţia meningococică.

Dr. biolog D. Gheţeu - Microbiologie
10
2. Infecţia subclinică : tulburări funcţionale , modificări morfologice, biochimice. 3. Infecţia latentă : asimptomatică, determinată de starea de toleranţă a gazdei, faţă de agentul patogen. Exemple : infecţia herpetică, cu cu virus rubeolic, tuberculoză, bruceloză. 4. Infecţia persistentă : cu evoluţie lentă, este determinată de multiplicarea lentă, însăcontinuă a virusurilor, mai ales imunologic. Exemple : encefalita spongiformă, PESS ( panencefalita sclerozantă subacută). 5. Infecţia localizată: agentul patogen este la nivelul porţii de intrare, cu modificări locale şi facultativ generale (febră, frison), Exemple : foliculite, abcese, guturai. 6. Infecţia de focar: infecţie localizată, cu evoluţie cronică, cu modificări locale minime asociate unor manifestări generale clinice (febră, stare generală alterată, mialgii), sindrom inflamator paraclinic (VSH, fibrinogen). 7. Infecţia cronică: rar în bolile infecţioase (TBC, bruceloză), mai frecvent ca o posibilitate evolutivă (hepatita cronică postvirală).
Starea de purtător : - La persoanele sănătoase (purtătorii sănătoşi de hemolitic, meningococ, etc).streptococ - Poate finaliza o boală infecţioasă indiferent de forma clinică, realizând purtătorul convalescent de scurtă durată ( infecţia dizenterică, salmonelozică). - Purtătorul cronic ( bacilul dizenteric, VHB). Boala infecţiosă ciclică: este cea mai frecventă modalitate de manifestare a bolilor infecţiose, cu evoluţie etapizată : - Perioada de incubaţie : timpul de la momentul pătrunderii agentului patogen până la apariţia primului simptom clinic ; poate fi : o Scurtă( 30 – 60 minute în toxinfecţia stafilococică). o Medie (7 – 14 zile : rujeola, dizenteria, varicela, tusea convulsivă). o Lungă ( >14 zile : hepatita virală B , C, rabie). - Perioada de debut : invazie, prodromală. Exemplu : tetanostrismus, rujeolăKoplick ; sursa cea mai bogată de infecţie ; debutul poate fi : o Scurt (câteva ore) gripă , scarlatină, meningita bacteriană, tetanosul supraacut. o Câteva zile ( rujeolă, difteri, moninucleoză infecţioasă) - Perioada de stare : sindroame clinice particulare ( febril, digestiv, respirator, meningeal, icteric, ganglionar, hepatic, splenomegalic, eruptiv. - Perioada de convalescenţă :de scurtă sau lungă durată, consemnează atenuarea simptomatologiei clinice până la dispariţie ; poate fi abruptă ( in crisis) sau lent (in lisis). - Imunitatea post infecţioasă : o Puternică si de durată : scarlatină, rubeolă, rujeolă, infecţie urliană. o Slabă şi de scurtă durată cu posibile recidive : febră tifoidă, dizenterie bacteriană, erizipel. o Fără imunitate după boală : tetanos. - Ieşirea din infecţie se poate face cu : o Vindecare completă, cu restitutio ad integrum ( rujeolă, varicelă, scarlatină) şi după tratament complet şi corect. o Vindecare clinică însoţită de stare de stare de purtător temporar sau cronic. o Cu sechele : poliomielita, hepatite. o Cronicizare : hepatitele B , C , D, bruceloză , dizenterie bacteriană. - Evoluţia : o Nefavorabilă spre deces : diagnostic incorect, sau tardiv, consecinţă a unui tratament indecvat, forme severe. Letalitatea este 100% , neinfluenţată de tratament : rabia, PESS, SIDA. - Infecţia septicemică : septicemia o Focar primar la poarta de intrare, a prezenţei continue sau intermitente a agenţilor patogeni şi a toxinelor lor în sânge, precum şi a focarelor metastatice secundare. - Bacteriemia: focar primar, prezenţa continuă sau intermitentăa agenţilor patogeni şi a toxinelor lor în sânge. - Recidivele = reapariţia tuturor semnelor de boală după aparenta vindecare a unui episod. Exemplu : hepatite acute virale, meningite purulente, scarlatină.

Dr. biolog D. Gheţeu - Microbiologie
11
• Chiar şi flora normală a organismului poate deveni patogenă, mai ales dacă printr-u accident, sau o igienă deficitară, ajunge în zone nespecifice (ex. E. coli din intestin dacă ajunge în vezica urinară poate să genereze infectii urinare). • O altă modalitate de infecţie endogenă este atunci când un germen îşi modifică aspectul patogen prin căpătarea unor caractere sporite de patogenitate (virulenţă sau toxicitate).
Curs II. Infecţia şi răspunsul imun
Imunitatea reprezintă capacitatea organismului de a se apăra în faţa agresiunii agenţilor externi (termenul îşi are originea în limba latină, unde immunitas înseamnă "ferit de"). Pentru a putea îndeplini această proprietate, organismul trebuie să recunoască structurile proprii, considerate "self" şi să le deosebească de cele străine, denumite "nonself", reprezentate în principal de microorganisme, macromolecule, dar şi de celule tumorale sau ţesuturi transplantate.
Mecanismele de apărare ale organismului pot fi împărţite în două mari categorii: înnăscute şi dobândite, care interacţionează între ele.
Imunitatea înnăscută (nespecifică) este prezentă la toţi indivizii normali, fiind prima barieră în calea agresiunilor externe. Modul său de acţiune este nespecific, acelaşi indiferent de natura agentului străin. Mecanismele imunităţii înnăscute se comportă identic şi în cazul unei reexpuneri, nefiind dotate cu memorie imunologică.
În această categorie sunt incluse: barierele mecanice (reprezentate de tegumente şi mucoase - gastrointestinală, a tractului respirator şi a celui urogenital), produşii de secreţie (mucusul, secreţia gastrică acidă, enzimele digestive, lizozimul din lacrimi şi mucusul respirator, secreţia glandelor sebacee, secreţia acidă vaginală, ph-ul urinar, etc.), unele proteine din sânge (sistemul complementului, reactanţii de fază acută, citokinele) şi sistemul mononuclear fagocitic (care include neutrofilele şi monocitele din sânge şi macrofagele din ţesuturi). La acestea se adaugă alte mecanisme care contribuie la epurarea agenţilor exogeni, cum ar fi: clearence-ul muco-ciliar, tusea, strănutul, vărsăturile, peristaltice intestinală. Inflamaţia reprezintă, de asemenea, un mijloc important de apărare nespecifică, macrofagele şi limfocitele polimorfonucleare (PMN) fiind atrase la locul infecţiei de produşii eliberaţi de organismele infectante şi ţesuturile necrozante.
Când mijloacele de apărare nespecifice sunt depăşite, agentul pătrunde şi se dezvoltă în ţesuturile gazdei, producând intrarea în acţiune a unui sistem de apărare specific, mult mai eficace.
Imunitatea dobândită (specifică) se dezvoltă pe măsură ce organismul interacţionează cu diferiţi agenţi externi, interacţiune ce poartă numele de eveniment imunizant. La o nouă expunere la agentul respectiv, organismul va reacţiona cu mai multă promptitudine, celulele implicate în acest proces fiind dotate cu memorie imunologică.
Imunitatea dobândită poate fi naturală (câştigată în mod pasiv prin transfer transplacentar, sau în mod activ prin contactul cu antigenul) şi artificială (care la rândul ei poate fi obţinută pasiv prin seroterapie sau activ prin vaccinare).
Totalitatea organelor, celulelor şi moleculelor implicate în apariţia imunităţii dobândite alcătuiesc sistemul imun.
RĂSPUNSUL IMUN Răspunsul imun reprezintă totalitatea fenomenelor prin care organismul recunoaşte şi
neutralizează structurile agresoare. Contactul dintre Ag şi celula care-l recunoaşte este urmat de un proces de multiplicare şi
diferenţiere a acesteia, ce are drept rezultat sinteza de Ac specifici (răspuns imun umoral) sau

Dr. biolog D. Gheţeu - Microbiologie
12
apariţia limfocitelot T sensibilizate (răspuns imun celular). Efectele acestui prim contact poartă numele de răspuns imun primar.
În desfăşurarea răspunsului imun (indiferent că este umoral sau celular) există mai multe etape:
preluarea, prelucrarea şi prezentarea Ag recunoaşterea Ag activarea limfocitelor faza efectoare şi eliminarea Ag declinul răspunsului imun (restabilirea homeostaziei) menţinerea memoriei Ag RĂSPUNSUL IMUN UMORAL Esenţa răspunsului imun de tip umoral este secreţia de Ig (Ac) specifici ca urmare a
stimulării antigenice a liniei limfocitare B. Răspunsul imun care apare după o primă expunere a unui organism la Ag se numeşte
răspuns imun primar. Ac sunt iniţial de tip IgM, iar apoi nivelul acestora scade şi apar Ac din clasa IgG.
Răspunsul imun secundar se produce când acelaşi Ag se administrează după un interval de timp. Această proprietate este posibilă datorită faptului că în timpul desfăşurării procesului de diferenţiere, se desprinde un grup de celule care nu continuă maturarea şi multiplicarea spre celule efectoare, ci rămân într-o etapă timpurie de evoluţie (în faza G1 a ciclului mitotic). Rolul lor este acela de a păstra amintirea Ag, chiar după totala sa eliminare, de aceea poartă numele de celule cu memorie. O nouă stimulare antigenică produce proliferarea şi diferenţierea în plasmocite a unora dintre celulele B cu memorie. Ac specifici apar mai precoce, au o mai mare afinitate pt. Ag, nivelul lor seric creşte mai rapid, iar secreţia este mai prelungită. Aparţin clasei IgG. Acest răspuns imun presupune cooperarea intercelulară între limfocitele T şi B, fără ajutorul altor celule.
Majoritatea Ag naturale, în special proteinele, necesită intervenţia limfocitelor T pentru a determina un răspuns imun umoral, de aceea se numesc Ag timodependente. Există şi Ag timoindependente, Ag care nu necesită intervenţia limfocitelor Th. În acest tip de proces nu se selectează celule cu memorie, nu există răspuns imun secundar, secreţia este exclusiv de tip IgM, şi durează luni de zile. Acest efect prelungit se explică şi prin faptul că odată format complexul Ag-Ac este fagocitat şi digerat. Celulele fagocitare nu pot digera decât Ac, eliberând Ag odată cu moartea celulei. Ag se va reinsera într-un nou ciclu.
După sinteza Ig, acestea se cuplează cu Ag formând complexele imune. În cazul în care Ag este solubil, molecular, se formează complexe imune circulante care sunt epurate de
sistemul monocit/macrofag. În cazul în care Ag face parte dintr-o membrană celulară, Ac se fixeaxă pe celulă. Cuplarea ulterioară a
complementului duce la liza celulei sau la eliminarea celulei prin fagocitoză de către sistemul mononuclear-fagocitic.
RĂSPUNSUL IMUN CELULAR Răspunsul imun celular este implicat în: eliminarea celulelor infectate cu microorganisme cu habitat intracelular (viruşi,
micobacterii, etc.) apărarea antitumorală rejecţia grefelor. În funcţie de natura receptorului implicat în recunoaşterea Ag expus pe suprafaţa
celulei ţintă, se pot distinge 2 categorii mari de celule implicate în acest tip de răspuns imun: limfocitele Tc şi celulele citotoxice MHC-nespecifice (NK şi K). Ag care declanşează acest răspuns imun sunt timodependente.
După aproximativ 5 zile de la primul contact cu Ag apar în sânge celulele T cu memorie care sunt supuse unui proces de recirculaţie continuă. Ele însămânţează continuu ţesutul limfatic, conferindu-i memorie imună specifică. Un nou contact cu Ag induce proliferarea şi diferenţierea celulelor cu memorie spre celule efectoare, producând un răspuns imun în câteva ore.
REACŢIILE IMUNE

Dr. biolog D. Gheţeu - Microbiologie
13
În unele situaţii însă, răspunsul imun este inadecvat, exagerat, pierzându-şi rolul protectiv şi devenind nociv pentru organismul gazdă. Ansamblul fenomenolor rezultate din perturbarea proceselor imune, cu apariţia unui răspuns imun capabil să producă leziuni organismului gazdă, poartă denumirea de reacţie imună. Pentru realizarea reacţiei imune, organismul face apel la o serie de factori nespecifici celulari şi/sau serici. Acţiunea lor este complexă şi intricată, ei contribuind major la dezvoltarea inflamaţiei locale, urmarea reacţiei imune.
Reacţiile imune, prin caracterul lor sistemic, pot determina leziuni la nivelul tuturor aparatelor şi sistemelor organismului, iar când reacţia este foarte intensă poate produce chiar moartea individului.
În funcţie de mecanismul lor de producere, reacţiile imune au fost împărţite de Gell şi Coombs în patru tipuri principale:
- tipul I sau reacţia anafilactică - contactul Ag-Ac este urmat de eliberarea de substanţe biologic active ce acţionează predominent pe muşchuil neted şi vase
- tipul II sau reacţia citotoxică - Ac lezează direct celula alterându-i structura sau funcţia şi predispunând-o la liză sau fagocitoză
- tipul III sau reacţia prin complexe imune - depunerea tisulară a CI şi fixarea complementului determină atragerea PMN şi declanşarea procesului inflamator
- tipul IV sau hipersensibilitatea tardivă - apare prin exagerarea răspunsului imun celular.
Roitt descrie şi tipul V de reacţie imună, asemănător cu tipul II, dar în care Ac fixaţi pe celulă nu o distrug ci îi modifică numai funcţia.
ORGANELE LIMFATICE A. ORGANELE LIMFATICE PRIMARE sunt organele în care are loc diferenţierea şi
maturarea limfocitelor. Sunt considerate organe limfatice primare măduva osoasă şi ficatul, responsabile de diferenţierea limfocitelor B şi timusul, sediul de diferenţiere al limfocitelor T.
1. Măduva osoasă şi ficatul: Măduva osoasă conţine ţesutul hematopoetic şi reprezintă locul de origine al tuturor
celulelorsanguine la adult. Ea include şi precursorii celulelor limfatice, seria limfoidă reprezentând 5-15%.
Diferenţierea celulei stem în elemente limfoide se face în trepte succesive, fenomenul fiind independent de existenţa stimulării antigenice, măduva osoasă neavând de altfel căi aferente şi modalităţi de captare şi prelucrare a antigenului. Proliferarea şi maturarea precursorilor celulari din măduva osoasă se face sub influenţa unor citokine numite factori de stimulare ai coloniilor (colony-stimulating factors). Ei sunt sintetizaţi de celule medulare stromale şi de macrofagele din măduvă şi creează un mediu local favorabil hematopoezei. Măduva osoasă mai conţine şi un procent variabil (între 5 şi 15%) de plasmocite, formate la nivelul organelor limfatice periferice ca o consecinţă a stimulării limfocitelor B de către Ag.
Măduva osoasă reprezintă locul unde se desfăşoară post-natal diferenţierea limfocitelor numite B. La om, prenatal, acest proces se desfăşoară în ficat, în timp ce la păsări el are loc într-un organ limfo-epitelial numit "bursa lui Fabricius". De aici a provenit de altfel iniţial şi denumirea acestor limfocite -B- bursodependente. Ulterior, când s-a văzut că procesul se desfăşoară la om în măduvă "bone marrow", denumirea acestui tip de limfocite s-a consolidat. De menţionat că limfocite provenind din măduvă populează şi timusul (mai ales corticala) unde se pot diferenţia în limfocite T.
Ficatul reprezintă doar sediul primar, precoce de hematopoeză, fiind locul prenatal de diferenţiere şi maturare a limfocitelor B. La adult, doar în unele circumstanţe patologice (cum ar fi policitemia vera) ficatul îşi reia funcţia hematoformatoare.
2. Timusul: Timusul este un organ limfoepitelial care apare devreme în embriogeneză, creşte în
perioada intrauterină şi atinge o dezvoltare completă la naştere, pentru ca după pubertate să sufere un proces de involuţie, probabil indus hormonal prin corticoizi şi androgeni.

Dr. biolog D. Gheţeu - Microbiologie
14
Situat în mediastinul anterior, timusul este format din 2 lobi, acoperiţi de o capsulă fibroasă, fiecare lob este organizat în lobuli, separaţi între ei prin trabecule de ţesut conjunctiv. Lobulii la rândul lor, prezintă o zonă periferică (corticala) şi o zonă centrală (medulara).
În corticală se găsesc predominent celule limfocitare mari, imature care îşi au originea în măduva osoasă (în perioada adultă), caracterizate printr-o vie activitate de multiplicare. Pe măsură ce migrează spre medulară proliferează şi suferă un proces de maturaţie.
În medulară se găsesc predominent celule epiteliale reticulare ce contin în citoplasmă granulaţii bogate în hormoni timici, unele celule fiind aglomerate în grupuri constituind corpii Hassall; tot în medulară se găsesc limfocite mici, mature, asemănătoare celor din sângele periferic.
În esenţă, participare timusului la menţinerea homeostaziei imune priveşte două aspecte fundamentale:
a. realizează maturaţia precursorilor limfocitari T; b. asigură distribuţia limfocitelor T către anumite arii ale organelor limfatice periferice,
numite zone timodependente. Aceste zone sunt: zonele paracorticale ale ganglionilor limfatici şi zonele periarteriolare din pulpa albă a splinei. Acest lucru este realizat graţie unor receptori de tip "homing" pe care limfocitele T îi achiziţionează pe tot parcursul lor intratimic.
Formarea limfocitelor T mature este rezultatul unui şir complex de etape de proliferare, diferenţiere şi maturare, care implică exprimarea la suprafaţa celulelor a unor receptori esenţiali în recunoaşterea antigenelor (TCR, CD4, CD8), în activarea metabolică limfocitară (CD28, CD45) sau în adeziunea intercelulară ca (CD2, LFA1 etc.). Proliferarea este controlată de citokinele eliberate local, iar diferenţierea apare în urma contactului limfocitelor cu celulele stromei timusului, contact ce asigură generarea unor celule imunocompetente şi departajarea funcţională pe liniile helper, citotoxică, supresoare şi contrasupresoare.
Însă, după ce limfocitele T exprimă pe suprafaţa lor aceşti receptori, marea majoritate mor in situ prin apoptoză = moarte celulară programată. Acestea reprezintă aşa numitul contingent distrus, distrugerea lui împiedicând apariţia procesului de autoimunitate. Puţinele limfocite rămase viabile (contingentul posttimic), trec în circulaţie după un pasaj prin medulară. Distrucţia celulară prin apoptoză şi nu prin necroză nu lasă să pătrundă în mediul extracelular diversele debriuri celulare care ar putea declanşa un răspuns local inflamator, protejând astfel structura şi funcţia acestui organ. În plus, celulele apoptotice exprimă pe suprafaţa lor receptori care permit recunoaşterea şi ingestia lor de către macrofage şi epurarea lor rapidă.
B. ORGANELE LIMFATICE PERIFERICE reprezintă sediul unde se iniţiază şi se desfăşoară răspunsul imun. În această categorie sunt incluse atât organe capsulate ca splina şi ganglionii limfatici, cât şi ţesuturile limfoide difuze, asociate tubului digestiv (notat GALT - Gut Associated Lymphoid Tissue), căilor respiratorii (BALT - Branchial Associated Lymphoid Tissue) sau ţesutului cutanat (SALT - Skin Associated Lymphoid Tissue).
1. Splina este cel mai mare organ limfatic; ea este delimitată la exterior de o capsulă ce conţine fibre musculare netede, din care se desprind trabecule ce pătrund în parenchim şi compartimentează organul. În parenchimul splenic se disting două sectoare, diferite atât ca structură cât şi ca funcţie:
-pulpa roşie situată la periferia lobulilor, alcătuită predominent din macrofage cu rol în fagocitarea hematiilor senescente şi care aparţine sistemului mononuclear-fagocitic;
-pulpa albă situată în jurul arteriolei centrale care conţine ţesut limfoid numit PALS - Peiarteriolar Lymphoid Sheath); acesta la rândul ei are 2 zone, una strinc periarteriolară, ce conţine în special limfocite T şi formează (cum spuneam mai devreme) zona timodependentă a splinei şi alta la periferia PALS ce conţine aproape exclusiv limfocite B, organizate sub formă de foliculi - zona timoindependentă. Aceşti foliculi pot fi primari sau secundari, formaţi din celule B activate antigenic, caz în care poartă denumirea de centrii germinativi.
Între pulpa roşie şi cea albă, la periferia foliculilor limfatici, se găsesc sinusurile marginale, unde există atât limfocite T cât şi B, precum şi magrofage şi celule dendritice, absolut necesare (cum vom vedea în continuare) pentru cooperarea celulară din cadrul răspunsului imun.
Funcţia imunologică a splinei se exprimă în special în generarea unor răspunsuri imune faţă de Ag aflate în circulaţia sistemică.

Dr. biolog D. Gheţeu - Microbiologie
15
Rolul important pe care îl joacă splina în apărarea antiinfecţioasă este demonstrat de faptul că la bolnavii splenectomizaţi apar infecţii severe, mai ales cu germeni incapsulaţi (ca pneumococii şi meningococci).
2. Ganglionul limfatic constituie un adevărat filtru situat pe traiectul căilor limfatice. El este de asemenea delimitat de o capsulă fibroasă ce trimite spre interior septuri fibroase incomplete. Histologic, la nivelul ganglionului limfatic se disting trei zone:
- zona corticală - situată la periferie, alcătuită din limfocite B aranjate sub formă de foliculi primari sau secundari (centrii germinativi), cu o structură asemănătoare omonimilor lor din PALS. Foliculii constituie zona timoindependentă sau bursodependentă.
- zona paracorticală - situată sub cea corticală conţine predominant limfocite T (din acest motiv purtând denumirea de zonă timodependentă), precum şi un mare număr de macrofage.
- zona medulară - situată parahilar şi alcătuită din vase sanguine, sinusuri limfatice, limfocite B, T, plasmocite macrofage şi celule dendritice.
După stimularea antigenică, limfocitele T şi B migrează unele spre altele, pierzându-şi această repartiţie anatomică. Limfocitele T activate părăsesc ganglionul limfatic şi pătrund în circulaţia sistemică, iar limfocitele B activate migrează spre centrii germinativi unde sintetizează anticorpi.
O atenţie specială este acordată segmentului vascular denumit venulă postcapilară situată paracortical, ale cărei celule endoteliale sunt cilindrice şi înalte şi poartă denumirea de High Endothelial Venules - HEV. Această zonă cu celule cilindrice este locul de trecere al limfocitelor din circulaţia sanguină în ganglion. Ele au pe suprafaţa lor molecule de adeziune numai pentru limfocite, fiind neutre faţă de alte celule circulante.
3. Alte ţesuturi limfatice Zone de ţesut limfatic există în submucoasa unor porţiuni din tractul digestiv (GALT) sau
respirator (BALT), precum şi din tegument (SALT). În unele teritorii aceste colecţii sunt suficient de mari pentru a fi individualizate. Astfel, la nivelul tubului digestiv există în jurul faringelui inelul limfatic al lui Waldayer, compus din amigdalele faringiene, palatine, linguale, iar la nivelul ileonului se găsesc plăcile Peyer. Acestor structuri li se asociază apendicele. Colecţii similare de ţesut limfatic se găsesc şi la nivelul arborelui bronşic (mai ales la bifurcaţia bronhiilor mijloci sau la joncţiunea bronho-alveolară) şi în tegument.
Importanţa acestui sistem imun rezidă nu numai din faptul că el reprezintă un mijloc eficient de apărare la nivelul principalelor porţi de intrare ale organismului, controlând contactul cu o multitudine de Ag cu poartă de intrare digestivă, respiratorie şi cutanată, ci şi din faptul că ele sunt organe limfopoetice importante şi pot fi, la adult, surse de limfocite B sau T. CELULELE SISTEMULUI IMUN
Celulele sistemului imun sunt: limfocitele, celulele prezentatoare de Ag (CPA) şi celulele efectoare
A. LIMFOCITELE Limfocitele reprezintă componentul celular major al sistemului imun. Ele constituie o
categorie de celule ce grupează mai multe populaţii celulare similare morfologic, dar diferite din punct de vedere funcţional. Limfocitele provin din organele limfatice primare (timus, măduva osoasă şi, în perioada intrauterină, ficatul). O parte din aceste celule trec în circulaţia periferică şi migrează către organele limfatice periferice. Restul rămân pentru o perioadă în circulaţia generală ca limfocite circulante unde totalizează aproximativ 25% din elementele figurate albe. Între cele două sectoare există o recirculaţie permanetă, asigurând o supraveghere imună competentă şi continuă folosind un număr relativ mic de celule. Recirculaţia şi migraţia limfocitelor este reglată de moleculele de adeziune de pe suprafaţa lor, ce corespund unor alte molecule de adeziune de pe suprafaţa celulelor endoteliale sau matricei extracelulare, precum şi de chemokinele secretate de la nivel tisular.
Durata de viaţă a limfocitelor în sângele periferic variază între 1 şi 300 de zile, existând şi populaţii limfocitare care au o durată de viaţă lungă, de luni sau chiar ani, ele recirculând în permanenţă între cele două sectoare ca celule cu memorie.
Limfocitele sunt singurele celule din organism capabile să recunoască specific diferiţi determinanţi antigenici, fiind astfel responsabile de două dintre proprietăţile fundamentale ale răspunsului imun, specificitate şi memorie.

Dr. biolog D. Gheţeu - Microbiologie
16
Din punct de vedere morfologic, limfocitele naive, care nu au fost stimulate antigenic, au un diametru cuprins între 8 şi 10 m, un nucleu mare, cu heterocromatină densă şi citoplasmă puţină, ce conţine un număr redus de mitocondrii, lizozomi şi ribozomi, aflate în stare de repaus sau în faza G0 a ciclului celular. După contactul cu Ag, celulele intră în faza G1 a ciclului celular şi îşi schimbă caracteristicile morfologice (cresc în dimensiune, ajungând la un diametru de 10 -12 m, citoplasma e mai bine reprezentată şi conţine mai multe organite celulare).
Marea heterogenitate funcţională a limfocitelor este legată de expresia anumitor receptori de suprafaţă. Unii dintre aceşti receptori sunt exprimaţi numai în anumite stadii de diferenţiere sau activare, pe perioade scurte de timp, pe când alţii sunt permanenţi şi caracteristici diverselor linii celulare limfocitare. După mai multe denumiri utilizate pentru a desemna aceşti receptori, s-a stabilit utilizarea indicativului CD (de la cluster of differentation), în prezent cunoscându-se aproximativ 250 CD.
1. Limfocitele T La adultul sănătos celulele T reprezintă 60-80% din totalul limfocitelor, cea mai mare parte
având o durată de viaţă lungă, fiind celule intens recirculate. La microscopia optică diferenţierea dintre limfocitele T şi B este practic imposibilă. Prin prezenţa pe suprafaţa lor a unor receptori caracteristici, este posibilă însă identificarea lor.
Care sunt aceştia? 1. Receptorii limfocitului T, notaţi TCR (T cell receptor). Aceştia au proprietatea fundamentală de a
recunoaşte şi a lega Ag. Sunt transmembranari şi au o structură asemănătoare porţiunii Fab a moleculei de Ig, fiind alcătuiţi din 2 lanţuri diferite de polipeptide, legate între ele prin punţi disulfidice. Se cunosc 2 tipuri de TCR: TCR1, primul care apare în timpul vieţii intrauterine, compus din 2 lanţuri şi şi TCR2 prezent pe suprafaţa a 95% din limfocitele T, alcătuit din lanţuri polipeptidice notate şi . Ambele tipuri de receptori au un domeniu variabil, în porţiunea NH2-terminală, şi unele constante. Aceşti receptori sunt legaţi de membrana limfocitelor T prin intermediul unui alt grup de 5 lanţuri polipeptidice notate CD3 şi care au rolul de a transmite semnalul activator în interiorul celulei, formându-se astfel un complex TCR-CD3 care are proprietatea de a se lega atât de Ag cu care vine în contact celula, cât şi de moleculele Ag de histocompatibilitate.
Astfel, limfocitele T nu recunosc Ag native solubile (recunoscute de limfocitele B şi Ac) ci numai pe acelea cuplate cu MHC, fiind vorba de o recunoaştere asociativă.
2. CD2 - unul dintre primii markeri de suprafaţă, prin care celula fixează în jurul ei un număr de eritrocite de oaie cu formarea unor rozete (metoda este încă folosită pentru pentru identificarea şi separarea limfocitelor T). E prezent pe toate tipurile de limfocite T şi pe NK. Are rol în aderarea limfocitelor T la APC sau celulele ţintă (prin legarea de contraligandul său LFA3) şi rol accesor în activarea celulară.
3. CD4 glicoproteină de suprafaţă situată predominent pe suprafaţa celulelor Thelper (definitorie pentru această clasă de celule). CD4 reprezintă receptorul T pentru moleculele MHC de tip II. Stabilirea legăturii între limfocitul T CD4+ şi APC prin intermediul moleculei de MHC II contribuie esenţial la recunoaşterea Ag.
4. CD8 tot de natură glicoproteică, distribuit pe membrana limfocitelor T supresoare, contrasupresoare şi citotoxice. CD8 interacţionează cu moleculele MHC I.
5. CD5 se găsesc pe toate limfocitele T şi pe un subset de limfocite B. 6. CD28 prezent pe 95% dintre limfocitele T CD4+ şi 50% din cele CD8+ are rol în activarea celulară,
precum şi rol de moleculă de adeziune. 7. MHC I şi II 8. Receptori limfocitari T cu funcţie în adeziunea celulară Indiferent de particularităţi, funcţiile efectorii ale diverselor celule imune implică în mod obligatoriu
adeziunea intercelulară, proces căruia i se acordă în ultima vreme o atenţie tot mai mare. Adeziunea celulară este implicată atât în funcţiile efectorii imune cât şi în repartiţia specifică a diverselor tipuri de limfocite în anumite arii ale sistemului imun. Principalii receptori limfocitari implicaţi în aderarea intercelulară LFA (Ag asociat funcţiei limfocitare), VLA , CD2 şi CD4 pentru limfocitele Th şi CD8 pentru limfocilele Tc. Astfel, moleculele de adeziune au rol în:
-respingerea forţelor electrostatice de respingere dintre 2 celule -realizarea unui contact prelungit între celulele care urmeazăă să coopereze -favorizarea recunoaşterii Ag -rol accesor în activarea celulară. Clasificarea limfocitelor T se face după funcţia pe care o îndeplinesc, precum şi după tipul
markerilor de suprafaţă. Se deosebesc aslfel mai multe tipuri de celule T, cele mai importante fiind:
a. Limfocitele T ajutătoare sau helper - sunt CD4+ şi recunosc Ag specifice asociate cu Ag HLA II. - unele limfocite Th colaborează cu limfocitele B şi macrofagele în inducerea răspunsului imun umoral, iar
altele colaborează cu alte celule T la producerea limfocitelor Tc intervenind în răspunsul imun celular. - există 5 subtipuri, 2 fiind mai bine cunoscute: Th1 care intervin în răspunsul imun celular prin stimularea
dezvoltării limfocitelor Tc, creşterea activităţii macrofagice. Limfocitele Th2 care intervin în procesele atopice, coordonând secreţia de IgE, IgA şi IgG1, induc proliferarea eozinofilelor şi a mastocitelor. Ele îşi exercită acţiune prin sinteza unor proteine numite citokine. Astfel, diferenţierea lor nu se face pe criterii fenotipice ci în funcţie de

Dr. biolog D. Gheţeu - Microbiologie
17
setul de IL pe care îl elaborează. De ex. LTh1 elaborează: IL-2, IL-3, IFN-, limfotoxină, TNF- şi . LTh2 sintetizează: IL-4, IL-5, IL-6. Între cele 2 subtipuri există inter-relaţii stimulatoare şi inhibitoare. IFN- (secretat de Th1) deprimă maturarea Th2 cu consecinţe funcţionale. Th2 (prin IL-10) suprimă producţia de Th1 acţionând ca o celulă supresoare.
b. Limfocite T supresoare - sunt limfocite reglatoare care diminuă atât răspunsul imun umoral cât şi pe cel celular - majoritatea sunt celule CD8+ - formarea lor din din precursori este urmarea contactului cu Ag, fără implicarea MHC. - recunosc determinanţii Ag, elaborează factori supresori solubili cu funcţie de contrareceptori blocanţi - la rândul lor sunt supuse controlului inhibitor al limfocitelor T contrasupresoare. - prin stimulare Ts de către Ag proprii (self) este prevenit procesul de autoimunitate. c. Limfocitele T citotoxice - sunt responsabile de citotoxicitatea mediată celular; ea este implicată în apărarea antivirală, antitumorală şi
respingerea grefelor. - provoacă liza celulelor care poartă pe suprafaţa lor Ag - au determinanţi antigenici de tip CD8+ şi mai puţin CD4+ - pentru ca Ag să fie recunoscut de către limfocitele Tc, el trebuie să fie asociat cu Ag MHC I, similar celui al
limfocitelor Tc. Acest fenomen poartă numele de restricţie HLA. d. Limfocite T inductoare - sunt CD4+, şi recunosc Ag asociat cu MHC II - stimulează activitatea altor limfocite T (helper, suppresor, citotoxice) e. Limfocitele T cu memorie - sunt formate în timpul răspunsului imun primar, sunt CD4+ -au o viaţă lungă, de peste 40 ani, dar perioada lor activă este de numai 10-15 ani. Se poate astfel observa că limfocitele T joacă un rol esenţial în reglarea răspunsului imun, justificând
afirmaţia potrivit căreia "Limfocitul T este dirijorul orchestrei imunologice" 2. Limfocitele B Limfocitele B reprezintă 5-15% din totalul limfocitelor circulante; au o durată de viaţă
scurtă (câteva zile). Ele sunt precursorii celulelor care sintetizează Ac, plasmocitele. 3. Celule natural killer (NK) Celulele NK reprezintă un subset de limfocite care au capacitatea de a omorî celule
infectate şi celule care şi-au pierdut moleculele MHC I. De asemenea, celulele NK stimulează macrofagele să distrugă microbii fagocitaţi. Termenul de natural killer provine de la faptul că aceste celule îşi exercită funcţia citotoxică fără a necesita o activarea prealabilă. De asemenea, spre deosebire de limfocitele Tc, celulele NK nu îşi recunosc specific ţinta, similitudinea între Ag MHC de pe cele 2 tipuri de celule nefiind necesară. Activitate lor nu este deci supusă fenomenului de restricţiei HLA.
Celulele NK au pe suprafaţa lor şi receptori pentru porţiunea Fc a IgG (FcRIII-CD16). Acest lucru explică proprietatea lor fundamentală de citotoxicitate faţă de celule care au fixat pe suprafaţa lor Ac specifici şi de care se leagă prin intermediul acestor receptori. Acest fenomen poartă denumirea de citotoxicitate mediată de Ac, la realizarea lui nefiind necesară participarea complementului.
Activitatea celulelor NK este stimulată de citokine ca: IL-15, IL-12, IFN. B. CELULE PREZENTATOARE DE AG Celulele prezentatoare de Ag (CPA) sunt celule specializate în capturarea Ag,
prezentarea lui limfocitelor şi declanşarea unor semnale care favorizează activarea limfocitelor. Există 2 categorii de CPA: a. "profesionale": macrofagele, celulele dendritice şi LB b. "ocazionale": PMN (m.a. neutrofilele), celulele endoteliale. 1. Macrofagele Sistemul mononuclear fagocitic sau sistemul monocit/macrofag sau reticulo-endotelial
reuneşte celule (monocitul şi macrofagul) care au origine medulară comună, precum şi caractere histologice (sunt celule mononucleare) funcţionale (capacitate de fagocitoză) similare. Rolul lor fundamental este apărarea împotriva bacteriilor intracelulare şi a fungilor. Au origine în seria monocitară a sistemului hematopoetic; precursorii lor sunt monoblastul-promonocitul-monocitul,

Dr. biolog D. Gheţeu - Microbiologie
18
Monocitele reprezintă 1-8% din leucocite, au un T1/2 de 8-72 ore, după care părăsesc torentul circulator şi migrează tisular, diferenţiindu-se în macrofage. Ele au un diametru de 10 – 15 m, un nucleu mare şi o citoplasmă bogată în lizozomi, vacuole şi filamente de citoschelet. Procesul de diferenţiere presupune: creşterea dimensiunilor celulare, amplificarea aparatului mitocondrial şi lizozomal, dezvoltarea proprietăţilor de fagocitoză şi sinteză, apariţia unor receptori pe suprafaţa celulei.
Macrofagele tisulare sunt heterogene, având morfologie, funcţii şi capacitătţi metabolice diferite, în funcţie de ţesutul căruia îi aparţin. Macrofagul este o celulă cu diametru de 10-25, cu nucleu mare, cu multe granule lizozomale şi vacuole în citoplasmă. Ele au căpătat denumiri specifice, în funcţie de locul de rezidenţă: Mf din ţesutul conjunctiv se numesc histiocite, cele din ficat - celule Kupffer, cele din ţesutul nervos - celule microgliale, cele din plămân - macrofage alveolare, cele din os - osteoclaste, cele din articulaţii - sinoviocite A, etc.
Figura 2. Liniile de diferenţiere celulară în cazul celulelor implicate în apărarea
oganismului Activarea macrofagelor nu este un proces specific, ca în cazul limfocitelor. El poate fi realizat sub influenţa
multor stimuli nespecifici: citokine (IFN- este cel mai puternic activator al macrofagului), contactul cu Ag, substanţe mitogene, etc. Durata de viaţă a macrofagului este de câteva luni.
Celulele sistemului monocit-macrofagic sunt implicate atât în apărarea nespecifică cât şi în cea specifică. În imunitatea nespecifică aceste celule fagocitează unele Ag şi sintetizează unele citokine care stimulează alte celule. În răspunsul imun, celule monocit-macrofagice au un rol deosebit de important şi complex, atât în etapa iniţială a răspunsului imun când au funcţia de CPA cât şi etapa finală, ca celule efectorii.
Ele au pe suprafaţa lor numeroşi receptori, dintre care cei mai importanţi sunt: receptori imuni: 1.- receptorii pentru fragmentul Fc al moleculele de Ig 2.- receptori pentru complement 3. receptori de adeziune celulara (LFA- antigenul asociat funcţiei leucocitare) receptori neimuni: (ei au rol în recunoaşterea celulelor senescente, eritrocitelor heterologe, bacteriilor,
fungilor, paraziţilor) 1.- pentru glicoproteine din structura peretelui bacterian, 2.- pentru proteine denaturate 3.- pentru hidrocarburi şi glicoproteine care se termină în fructoză şi manoză 4.- pentru fibronectină 5.- pt. unii hormoni, 6.- pt. Ag tumorale. De asemenea, macrofagul are o capacitate secretorie mare, această celulă putând elabora peste 50-100 de
produşi, dintre care menţionăm: enzime litice (elastaze, colagenaze, etc.), componente ale complementului (C1-

Dr. biolog D. Gheţeu - Microbiologie
19
C5, properdina), pirogeni endogeni, factori care influenţează diferenţierea celulară (de ex. colony-stimulating factor, factori de angiogeneză), factori citotoxici, citokine (IFN, IL, TNF-), unii factori de coagulare (V,VII, IX, X, protrombina, tromboplastina), radicali activi ai oxigenului.
2. Celulele dendritice Celulele dendritice reprezintă o populaţie leucocitară particulară, caracterizată morfologic
de prezenţa unor prelungiri citoplasmatice, iar funcţional prin capacitatea de a prezenta antigenul (sunt celule prezentatoare de antigen profesioniste) şi o extraordinară capacitate de a stimula limfocitele T native şi de a iniţia un răspuns imun.
Celulele dendritice au origine şi pot îndeplini rolul de CPA, datorită prezenţei pe suprafaţă a moleculelor MHC II şi a receptorilor pentru Ag. Ele au capacitatea de a capta Ag şi de a-l transporta la nivelul organelor limfatice periferice. După sediul lor pot fi împărţite astfel:
a. cele din organele nelimfatice: -celulele Langerhans care se găsesc în special în tegument (formează o reţea în stratul bazal şi în epiteliul
scuamos), dar şi la nivelul mucoasei bucale, faringe, col uterin. Pe suprafaţa lor au molecule HLA II, receptori pentru complement şi pentru Fc. Ele pot fixa astfel complexele imune. Sunt implicate mai ales în hipersensibilitatea cutanată de contact.
-celule interstiţiale situate în plămân, tractul digestiv, cord, ficat, rinichi b. cele din organele limfatice: -celulele interdigitale - se găsesc în zonele limfatice bogate în limfocite T, cu care cooperează (zonele
paracorticale ale ggl. limfatic, pulpa albă a splinei). -celulele foliculare - se găsesc în foliculii limfatici din corticala ganglionilor limfatici şi pulpa albă a splinei,
adică zone bogate în limfocite B. Multitudinea de arborizaţii le permite un contact intim cu limfocitele B, ele prezentând acestora Ag ca atare, nedegradat sau în complexe imune, într-un proces independent de MHC.
C. CELULELE EFECTOARE Limfocitele stimulate antigenic se transformă în celule efectoare, al căror rol esenţial este
acela de a elimina Ag. În această categorie sunt incluse: limfocitele T helper, limfocitele T citotoxice şi plasmocitele. Limfocitele T helper activate exprimă receptori de suprafaţă şi secretă numeroşi produşi care le permit să interacţioneze cu alte celule. Limfocitele T citotoxice activate dezvoltă granule care conţin proteine capabile să omoare celulele infectate viral sau celulele tumorale. Limfocitele B stimulate antigenic se transformă în celule capabile să sintetizeze imunoglobuline, numite plasmocite. Morfologia acestora este diferită: nuclei denşi, citoplasmă abundentă, cu reticul endoplamatic bine reprezentat (acesta fiind sediul de sinteză al imunoglobulinelor) şi complex Golgi perinuclear cu rolul de a converti imunoglubulinele în forma finală sub care sunt sintetizate.
IMUNOGLOBULINELE sau ANTICORPII Imunoglobulinele (Ig) reprezintă o familie de glicoproteine care au proprietatea de a
reacţiona în mod specific cu Ag a cărui recunoaştere de către sistemul imun a condus la sinteza lor, motiv pentru care li s-a conferit denumirea de Ac. Ele sunt sintetizate de limfocitele B fie asociate membranei celulare unde joacă rolul de receptori, fie sub formă serică. Ig reprezintă aprox. 20% din proteinele plasmatice şi migrază în cea mai mare parte în zona electroforetică gamma. În afară de ser, o cantitate variabilă de Ig se găsesc în lichidele extravasculare, secreţiile exocrine şi ţesutul limfatic, dar nu depăşesc bariera hemato-encefalică.
Structura generală a unei Ig În alcătuirea Ig, proporţia glucidică reprezintă doar 4-18%, restul fiind polipeptide. Fiecare
moleculă de Ig este alcătuită din cel puţin o unitate monomerică de bază. O asemenea unitate monomerică conţine 4 lanţuri polipeptidice, a căror coeziune este realizată prin punţi disulfidice. Aceste punţi sunt dispuse atât între lanţuri (intercatenare) cât şi în interiorul aceluiaşi lanţ polipeptidic (intracatenare) şi sunt esenţiale pentru configuraţia tridimensională a moleculei. Forma este cea a unui Y.
Cele 4 lanţuri sunt reprezentate de 2 lanţuri grele notate cu H (de la heavy=greu) cu o greutate moleculară de 55 – 75 kD şi 2 lanţuri uşoare notate L (de la light=uşor) a căror greutate moleculară este de aproximativ 24 kD. Aşezarea lor este simetrică. fiecare lanţ polipeptidic are o extremitate aminoterminală, denumită porţiune variabilă (V) şi un capăt carboxi-terminal denumit porţiune constatntă. Prin apropierea regiunilor variabile se realizează o structură tridimensională numită situs combinativ sau paratop, care interacţionează cu epitopul Ag specific. În afara acestora se mai descriu şi nişte regiuni numite hipervariabile sau CDR (complementarity Determining Region), caracterizate printr-o compoziţie foarte variabilă de amino-acizi, situate atât pe lanţutile H

Dr. biolog D. Gheţeu - Microbiologie
20
cât şi pe cele L. Numărul situsurilor combinative determină valenţa anticorpilor. de ex. el este 2 pentru IgG şi 10 pentru IgM.
Atât lanţurile H cât şi cele L nu sunt liniare, ci au o dispoziţie buclată, ca urmare a legăturilor disulfidice intracatenare. Regiunile buclate astfel create poartă denumirea de domenii. Lanţurile L au 2 domenii (unul în zona variabilă şi unul în cea constantă). Lanţurile H au un domeniu în zona variabilă şi 4-5 în cea constantă.
Porţiunea dintre primele 2 domenii ale regiunii constante ale lanţurilor H poartă denumirea de zona balamalei (hinge region) datorită frexibilităţii moleculei la acest nivel, ceea ce îi conferă un rol important în angajarrea legăturii dintre Ag şi Ac.
Tipurile de Ig După tipul de lanţ H, Ig se clasifică în 5 tipuri, cu structuri şi proprietăţi diferite, pe care,
foarte pe scurt, le vom trece în revistă în continuare: 1. IgG - reprezintă clasa dominantă din serul uman, adică 75% din totalul Ig - au cea mai mare durată de viaţă (3 săptămâni). - d.p.d.v. structural IgG este un monomer, alcătuit din 2 lanţuri H de tip şi 2 lanţuri uşoare care pot fi de tip
sau . Lanţurile H sunt împărţite în 4 subtipuri notate 1, 2 , 3 şi 4, care definesc cele 4 subtipuri de IgG: IgG1, IgG2, IgG3 şi IgG4.
- d.p.d.v. funcţional IgG au o importanţă deosebită, Ac răspunsului imun secundar aparţinând în cea mai mare parte acestei clase.
- au o distribuţie egală în sectorul intra- şi extravascular - IgG este singura Ig care poate traversa placenta, asigurând protecţia în primele luni după
naştere - IgG1 şi IgG3 se pot lega de componentul C1q al sistemului complementului, determinând
iniţierea activării acestuia pe calea clasică. Activarea complementului de către IgG impune participarea a minimum 2 molecule de Ac.
- IgG au de asemenea funcţii antitoxice şi de opsonizare 2. IgA - structural se aseamănă cu molecula de IgG, conţinând 2 lanţuri grele, dar de tip , şi 2
lanţuri L sau . - se cunosc 2 tipuri de IgA: IgA1 (majoritar) şi IgA2. - spre deosebire de celălalte tipuri de IgG, IgA sunt prezente atât în ser cât şi în diferite
secreţii. - din acest p.d.v se descriu: IgA serice cu o structură monomerică, (doar < de 10% fiind formate din
asamblarea a 2, 3 sau 4 monomeri), predominant de tip IgA1 şi IgA secretorie care este de tip IgA2. Acest tip se găseşte în secreţiile digestive, salivă, secreţiile lacrimale, bronşice, colostru, laptele matern. IgAs se prezintă în general sub forma unui dimer, reunit cu ajutorul unui lanţ polipeptidic J şi un nou polipeptid, numit componentă secretorie, ce se ataşează de lanţul H . Componenta secretorie este sintetizată de celulele epiteliale. Rolul său este acela de a conferi IgA rezistenţă faţă de acţiunea enzimelor proteolitice din secreţii. Limitând pătrunderea Ag în submucoase, IgA au rol în imunitatea locală.
3. IgM - este cea mai mare Ig, fiind alcătuită din 5 monomeri identici uniţi între ei prin lanţuri J şi
legături disulfidice la nivelul fragmentelor Fc. - lanţul H este de tip , iar cele L sunt fie fie - este principala Ig a răspunsului imun primar - are cea mai puternică acţiune de activare a căii clasice a complementului, o singură
moleculă de Ig M fiind suficeintă pentru a fixa C1q. -se poate sintetiza şi local, forma secretorie posedând ca şi IgA o componentă suplimentară. 4. IgD - reprezintă < de 1% din Ig serice - este un monomer, în care lanţul H este de tip - semnificaţia lor fiziologică nu este deplin elucidată, dar s-a dovedit că IgD, ca şi IgM au
rolul de receptor pnetu Ag la suprafaţa membranei limfocitului B. 4. IgE - se mai numesc şi reagine - au cel mai scrut timp de îmjumătăţire (2-3 zile) - sunt monomeri, lanţul H este de tip

Dr. biolog D. Gheţeu - Microbiologie
21
- se fixează prin fragmentul lor Fc de receptorii specifici de pe membrana bazofilelor circulante şi mastocitelor tisulare, putând declanşa astfel un răspuns de tip reaginic
- concentraţia lor plasmatică creşte în caz de infecţii parazitare, fungice, virale. SISTEMUL COMPLEMENTULUI Sistemul complementului reprezintă un complex de proteine serice ce conţine 25-30 de
componente (aflate în mod normal în stare inactivă) care îndeplinesc un rol important în apărarea organismului şi în procesul inflamator. El face parte din mijloacele de apărare nespecifică ale organismului. Activarea componentelor sale se face succesiv, "în cascadă". În urma activării rezultă o întreagă gamă compuşi biologic activi, care au capacitatea de a activa celule cu rol efector atât în imunitate cât şi în inflamaţie.
Principalele funcţii ale complementului sunt: Inflamatoare: Activare PMN, Mf Marginatia si diapedeza PMN Chemotaxie Degranularea mastocitelor Cresterea permeabilitatii vasculare Contractia mm. neted Eliminarea CI Funcţia citotoxică - în stadiile finale ale activării sale celulele ţintă sunt lizate. El este
implicat în liza multor celule cum ar fi: hematii, limfocite, bacterii, virusuri cu capside lipoproteice. Neutralizare virală Opsonizare - complementul activat se leagă de Ag sau complexe imune, favorizându-le
aderenţa şi fagocitoza. Privind aceste funcţii ne putem da seama că acest sistem participă atât în apărarea specifică
cât şi în cea nespecifică, rolul lui fundamental fiind acela de a corobora cele două tipuri de mecanisme, crescând eficienţa intrinsecă a fiecăruia dintre ele.
Ficatul este principalul sediu al sintezei proteinelor ce alcătuiesc acest sistem, deşi macrofagele tisulare şi fibroblaştii pot sintetiza şi ei unele componente.
Activarea componentelor complementului se face pe trei căi: calea clasică, calea alternă şi calea lectinelor.
Calea clasică Elementul iniţial este complexul imun Ag-Ac sau un Ag situat pe membrana celulară.
Activarea se realizează prin legarea componentului C1 de fragmentul Fc al moleculei de Ig. Calea clasică mai poate fi activată direct de unele virusuri şi endotoxine. C1 este un complex trimolecular ale cărui componente sunt notate C1q, C1r şi C1s, a căror activare se face în această ordine.
C1s activează C4, o moleculă de C1s putând cliva mai multe molecule de C4, ceea ce duce la amplificarea procesului. C4 este descompus în 2 componente: C4a şi C4b. C4a este o substanţă numită anafilatoxină care are următoarele proprietăţi: stimulează degranularea mastocitelor, eliberează aminelor vasoactive din mastocit şi bazofil, produce contracţia muşchiului neted, creşte permeabilitatea vasculară, eliberează amine vasoactive din granulocite.
C4b se leagă de următorul component ce întră în joc, C2. C2 este clivat sub acţiune C1s în 2 componente: C2a (o protează) şi C2b. C2b rămâne legat de C4b formând un complex enzimatic numit C3-convertaza, care determină clivarea componentului C3.
C3 este clivat astfel în C3a (şi ea o anafilatoxină) şi C3b, un component major al sistemului complementului. El se leagă de C4bC2b formând un complex trimolecular numit C5-convertaza. Tot C3b are capacitatea de a declanşa calea alternă de activare.
În plus, C3b şi C4b favorizează aderenţa imună şi fagocitoza, pentru că pe suprafaţa unor celule ca limfocitele, hematiile, celule fagocitare există receptori specifici pentru aceste componente ale complementului.
Calea alternă sau properdinică Această cale poate fi declanşată atât imunologic (IgA, IgG) cât şi neimunologic
(componente bacteriene, parazitare, unele enzime). În acest proces intervin mai mulţi factori, cu origini variate notaţi B, D, H, I.

Dr. biolog D. Gheţeu - Microbiologie
22
C3b protejat de Ag declanşant se leagă de factorul B, care, sub acţiunea factorului D este clivat în 2 componente: Ba (cu efect chemotactic pentru neutrofiele) şi Bb care rămâne legat de C3b, formând complexul C3bBb, care reprezintă pt. calea alternă C3-convertaza. El are deci capacitatea de a cliva C3, generând o cantitate sporită de C3b. Astfe, de o singură molecula de Bb se leagă mai multe molecule de C3b, formând un nou complex numit C5-convertaza căii alterne.
Ultimele secvenţe de activare sunt comune pentru ambele căi. C5-convertaza clivează C5 în C5a (anafilatoxină, eliberează SRS-A din mastocite şi factor chemotactic) şi C5b. C5b se leagă succesiv de C6, C7, C8 şi C9, formând complexul C5b6789 numit complexul de atac al membranei (MAC). El are asemănări structurale cu perforina (moleculă eliberată de celulele NK), având ca şi aceasta, proprietatea de a leza membranele celulare, inducând astfel liza celulei.
Activitatea biologică a complementului este controlată de mai multe elemente: viaţa efemeră a unor componente activate, proteine serice cu rol de a modula şi limita activarea unor componente.
ANTIGENELE Antigenul este o substanţă, cel mai adesea străină organismului, capabilă ca în
momentul pătrunderii sale în organism, să declanşeze un răspuns imun (umoral sau celular) şi să reacţioneze specific cu produşii acestui răspuns imun (limfocite T sensibilizate sau Ig).
Se impun însă câteva menţiunii faţă de această definiţie: imunogenul este o substanţă capabilă doar de a declanşa un răspuns imun există unele Ag care, în anumite condiţii legate de calea de pătrundere sau concentraţie,
pot induce inhibiţia, supresia răspunsului imun aşteptat, prin stimularea mecanismelor supresoare. Acestea se numesc Ag tolerogene.
nu toate Ag sunt exogene. Există şi Ag endogene, cum ar fi autoantigenele (structuri proprii devenite non-self) sau Ag tumorale.
Porţiunea din Ag care interacţionează cu Ac sau cu receptorii limfocitelor T poartă denumirea de determinant antigenic sau epitop. El este de fapt un subcomponent al macromoleculei antigenice. Acesta poate avea o structură liniară (recunoscuţi numai de limfocitele T) sau aminoacizii pot veni în contact fără ca pe lanţul polipeptidic să fie alăturaţi - epitop conformaţional- (recunoscuţi de limfocitele B). Epitopul poate fi situat la suprafaţa moleculei de Ag sau se poate găsi în interiorul acesteia, mai ales în structurile moleculare complexe, caz în care se poate exprima numai după degradarea parţială şi deplisarea restului moleculei de Ag. O moleculă de Ag poate avea mai mulţi determinanţi antigenici, fiecare dintre aceştia putând induce sinteza unui Ac, proprietate numită plurivalenţă.
Caracteristicile fundamentale ale Ag sunt imunogenicitatea, specificitatea şi adjuvanticitatea.
A. Imunogenicitatea Pentru ca o substanţă să fie imunogenă ea trebuie să îndeplinească următoarele condiţii: 1. să fie străină organismului - cu cât între organismul gazdă şi cel cu ale cărui
molecule înteracţionează sunt mai diferite filogenetic, cu atât acestea din urmă sunt mai imunogene. Există însăşi excepţii în care substanţe străine organismului nu sunt antigenice, iar altele proprii pot deveni antigenice în anumite condiţii (această situaţie este substratul bolilor autoimune).
2. să aibă o greutate moleculară mare - în general molecule < de 5-10 kdaltoni nu sunt imunogene. Şi în această privinţă există însă excepţii.
3. să aibă o structură chimică complexă - o moleculă complexă, cu un număr mare de determinanţi antigenici, este mai imunogenă. Cu excepţia lipidelor pure, toate macromoleculele sunt imunogene. Proteinele, mai ales cele bogate în aa aromatici, au cel mai puternic potenţial imunogen.
4. să fie digerabile - pt. a induce un răspuns imun, Ag trebuie să fie prelucrate de unele celule (m.a. macrofagele). cele care nu pot fi digerate de echipamentul enzimatic al acestora nu sunt imunogene.
5. să fie accesibilă spaţial - pentru a putea permite contactul cu paratopul 6. să aibă o remanenţă mare în organul gazdă - remanenţă legată de starea de
solubilitate. Cu cât o substanţă este mai insolubilă, cu atât este mai antigenică. B. Specificitatea

Dr. biolog D. Gheţeu - Microbiologie
23
Este calitatea de a induce un răspuns imun strict specific faţă de stimulul declanşator. C. Adjuvanticitatea Este capacitatea de alipire a moleculelor de Ag de celulele prelucrante. Ea este mai
mare la Ag prezente sub formă de particule decât la cele existente în soluţie. Această proprietate poate fi amplificată prin folosirea adjuvanţilor, care, în plus, protejează Ag de o degradare rapidă, prelungindu-i astfel remanenţa.
Clasificarea Ag Antigenele pot fi clasificate în funcţie de criterii structurale, chimice sau de relaţia
gazdă-Ag. A. Antigene complete şi incomplete Pentru ca un Ag să fie considerat complet el trebuie să îndeplineasă întocmai cerinţele
impuse de definiţie (să inducă un răspuns imun şi să reacţioneze cu produşii acestuia). Există însă molecule care datorită dimensiunilor mici, nu pot induce un răspuns imun
decât dacă se asociază cu un suport constituit din structuri proteice sau tisulare ce poartă denimirea de carrier, dar pot interacţiona cu produşii răspunsului imun. Asemenea molecule se numesc haptene sau Ag incomplete. Ele nu au imunogenicitate (nu pot declanşa singure un răspuns imun), dar au specificitate (reacţionează cu Ac specifici). Carrier-ul este responsabil de imunogenicitate, haptena este responsabilă de specificitate.
B. Clasificarea chimică Din acest p.d.v. Ag pot fi: 1. Proteine - cele mai antigenice structuri 2. Glucide - în general slab antigenice, moleculele ramificate (de ex. polizaharidele din
structura unor bacterii) pot fi însă puternic antigenice 3. Lipide - de asemenea slab antigenice în general. Unele însă, cum ar fi cardiolipinele,
pot fi antigenice şi au semnificaţie în unele boli. C. Relaţia gazdă-antigen Din acest p.d.v. se deosebesc mai multe situaţii: 1. Antigene heretogene, heterologe sau xenoantigene sunt Ag care se găsesc la specii
diferite. 2. Antigene heterogenetice sau heterofile sunt Ag prezente la specii diferite filogenetic,
dar care au structuri analoage (au în comun unul sau mai mulţi epitopi). Din această cauză pot reacţiona cu acelaşi Ac specific (corespunzător epitopului comun) dând reacţii încrucişate.
Ac heterofili pot avea o importanţă mare în practică. De exemplu: treponema pallidum are un epitop numit cardiolipină, care se găseşte şi în miocard. Detectarea Ac anticardiolipină stă la baza cunoscutei reacţii numită VDRL de diagnosticare a infecţiei cu Treponema pallidum. În unele boli autoimune, apare Ac anticardiolipină, ceea ce determină apariţia unor reacţii VDRL fals +.
3. Antigene alogeneice sau aloantigene sunt Ag caracteristice unei specii, dar care diferă între indivizii din interiorul speciei respective. Ele conferă specificitate de specie.
Din această categorie fac parte antigenele de histocompatibilitate (HLA), cu rol decisiv în răspunsul imun. Ele au căpătat o deosebită importanţă odată cu dezvoltarea transplantelor. De asemenea, s-a observat că unele grupe HLA prezintă o mare susceptibilitate la o anumită boală, de unde s-a născut ideea unei predispoziţii genetice.
Destinul Ag odată pătruns în organism este influenţat de mai mulţi factori: calea de pătrundere, solubilitatea sa, dimensiune, capacitatea organismului receptor de a-l elimina sau distruge. Primul contact este cel cu mecanismele imunităţii înăscute (bariere mecanice, produşi de secreţie, PMN, etc). Majoritatea substanţelor exogene sunt astfel eliminate.
când Ag ajunge direct în sânge, este transportat în splină, unde vine în contact cu limfocitele locale şi declanşează răspunsul imun.
când Ag pătrunde pe cale cutanată sau subcutanată, este captat de celulele Langerhans şi este transportat în ganglionul limfatic de drenaj. Aici se desfăşoară răspunsul imun, iar produşii acestuia (Ac sau limfocitele sensibilizate) ajung în sânge via canalul toracic,

Dr. biolog D. Gheţeu - Microbiologie
24
când Ag pătrunde pe calea mucoaselor respiratorie sau digestivă, răspunsul imun se desfăşoară în ţesului limfatic local. Ac acţionează local, iar limfocitele B şi T migrează spre alte locuri de rezidenţă.
În concluzie, Ag care reuşesc să depăşească barierele nespecifice, activează sistemul imun declanşând răspunsul imun. Curs III
Bacteriile – morfologie, fiziologie, patogenitate
Bacteriile sunt celule procariote, cu un singur cromozom, saprofite, strict sau condiţionat patogene. Dimensiuni: 0,1-300 µm. Se văd doar la microscop. Produc bacterioze (de exemplu pneumonia, tuberculoza, meningita etc.). Bacteriile au o capacitate mare de sinteză şi multiplicare. Se înmultesc asexuat, prin diviziune directă, în 10-12 minute în condiţii favorabile. Sunt sensibile la acţiunea antibioticelor şi chimioterapicelor.
Sporul bacterian este forma de rezistenţă care apare la unele specii în condiţii neprielnice de înmulţire.
Creşterea bacteriilor este procesul biologic prin care bacteriile îşi măresc volumul, datorită sintetizării de noi produşi şi acumulării apei.
Multiplicarea bacteriilor se realizează prin diviziune directă şi rar prin înmugurire. Diviziunea bacteriană are loc atunci când bacteriile se găsesc într-un mediu favorabil
(lichid sau solid): - prin diviziune directă sau binară - amitoza, prin sciziparitate, care se produce la majoritatea bacteriilor; ritmul de diviziune este foarte rapid de 10-20 min; excepţie face bacilul Koch care are un timp de 18-20 ore. - prin înmugurire sau ramificare (la un număr foarte redus de bacterii).
III.1. Morfologie Pentru descrierea şi identificarea bacteriilor există mai multe criterii: morfologia, afinitatea tinctorială, sporogeneza, mobilitatea etc.. Morfologic, bacteriile prezintă: Capsulă de natură polizaharidică complexă cu molecule neramificate lipsite de miez lipopolizaharidic, puternic hidratată (99% apă). Membrana externă - stratul exterior al peretelui celular, este triplu stratificată. Peretele bacterian -cu componenta principală mureina sau peptidoglicanul, polimeri micşti, din două tipuri de hexoze: N-acetilglucozamina (G) şi acidul N-acetilmuramic (M). Membrana citolplasmatică - delimitează citoplasma cu componentele sale structurate: o nucleoid- reprezentat de cromozomul bacterian, o ribozomi, etc. Flagel(i)- flexibil(i)- cu rol în mişcare. Pili (Fimbrii) – imobili. Protoplaştii sunt forme bacteriene lipsite de perete bacterian, create artificial, menţinute în medii osmotice capabile să protejeze de liză bacteriană. In medii hipotone aceste celule se sparg, iar la medii hipertone se ratatinează. Protoplaştii provin din bacterii Gram-pozitive. Sferoplaştii sunt forme bacteriene create artificial prin indepărtarea parţială a peretelui. Peretele este lipsit de mureină, însă conţine lipide, motiv pentru care au aspect sferic. Sferoplaştii provin bacterii Gram-negative. Grupe morfologice de bacterii: Sferică: cocii: stafilococi şi streptococi;
pneumococi (formă lanceolată); meningococi şi gonococi (reniformi).
Cilindrici: bacili: forma alungită, de bastonaş cu forme diferite:
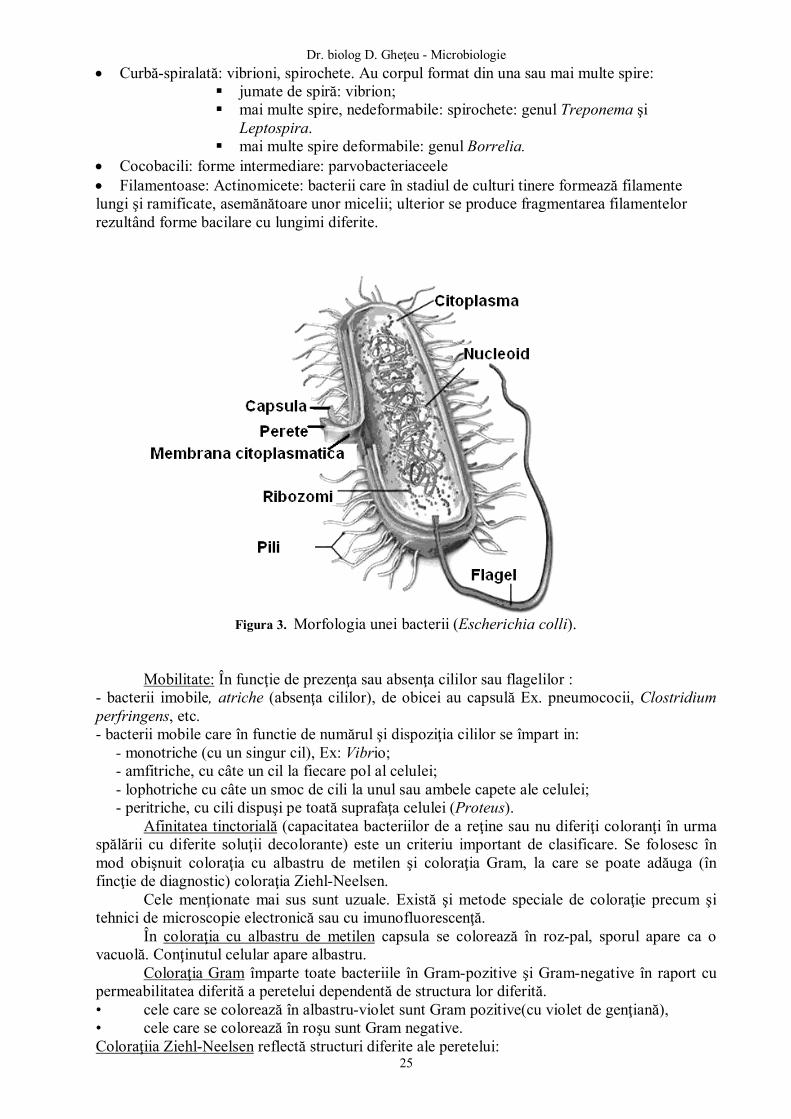
Dr. biolog D. Gheţeu - Microbiologie
25
Curbă-spiralată: vibrioni, spirochete. Au corpul format din una sau mai multe spire: jumate de spiră: vibrion; mai multe spire, nedeformabile: spirochete: genul Treponema şi
Leptospira. mai multe spire deformabile: genul Borrelia.
Cocobacili: forme intermediare: parvobacteriaceele Filamentoase: Actinomicete: bacterii care în stadiul de culturi tinere formează filamente lungi şi ramificate, asemănătoare unor micelii; ulterior se produce fragmentarea filamentelor rezultând forme bacilare cu lungimi diferite.
Figura 3. Morfologia unei bacterii (Escherichia colli).
Mobilitate: În funcţie de prezenţa sau absenţa cililor sau flagelilor : - bacterii imobile, atriche (absenţa cililor), de obicei au capsulă Ex. pneumococii, Clostridium perfringens, etc. - bacterii mobile care în functie de numărul şi dispoziţia cililor se împart in: - monotriche (cu un singur cil), Ex: Vibrio; - amfitriche, cu câte un cil la fiecare pol al celulei; - lophotriche cu câte un smoc de cili la unul sau ambele capete ale celulei; - peritriche, cu cili dispuşi pe toată suprafaţa celulei (Proteus).
Afinitatea tinctorială (capacitatea bacteriilor de a reţine sau nu diferiţi coloranţi în urma spălării cu diferite soluţii decolorante) este un criteriu important de clasificare. Se folosesc în mod obişnuit coloraţia cu albastru de metilen şi coloraţia Gram, la care se poate adăuga (în fincţie de diagnostic) coloraţia Ziehl-Neelsen.
Cele menţionate mai sus sunt uzuale. Există şi metode speciale de coloraţie precum şi tehnici de microscopie electronică sau cu imunofluorescenţă.
În coloraţia cu albastru de metilen capsula se colorează în roz-pal, sporul apare ca o vacuolă. Conţinutul celular apare albastru.
Coloraţia Gram împarte toate bacteriile în Gram-pozitive şi Gram-negative în raport cu permeabilitatea diferită a peretelui dependentă de structura lor diferită. • cele care se colorează în albastru-violet sunt Gram pozitive(cu violet de genţiană), • cele care se colorează în roşu sunt Gram negative. Coloraţiia Ziehl-Neelsen reflectă structuri diferite ale peretelui:

Dr. biolog D. Gheţeu - Microbiologie
26
• Acid-alcoolo-rezistente al căror perete foarte bogat în lipide devine permeabila) pentru colorant – fucsina - numai prin încălzire şi rezistă apoi la decolorare cu acid şi alcool, care se face la rece, ramânând colorate în roşu. • Acid-alcoolo-nerezistente care pierd culoarea roşie prin decolorare şi se recolorează cu al doilea colorant, albastrul de metilen, în albastru.
Staphylococcus aureus (ME) Leptospire (ME)
Vibrio (ME)
Bacilul Koch (MO)
Figura 4. Ilustrarea tipurilor morfologice de bacterii (ME-microscop optic, MO-microscop electronic).
Bacteriile Gram-pozitive au un perete gros (200 - 800Ả) alcătuit dintr-un strat rigid de peptidoglicani şi din acid teichoic. Cantitatea mare de peptidoglicani (70-80%) oferă acestor bacterii sensibilitate la lizozim şi penicilină. Lizozimul hidrolizează specific peptidoglicanii între N-acetil-glucozamină şi acidul N-acetilmuramic. Penicilina inhibă specific sinteza de peptidoglicani.
Bacteriile Gram-negative au un perete mai subtire de 100Ả cu un continut mult mai mic de peptidoglican (până la 20%) şi fără acid teichoic.
III.2 Fiziologia bacteriilor Caracteristicile metabolismului bacterian
- este un metabolism unicelular, se desfăşoară într-o singură celulă; - este un metabolism necompartimentat, caracteristic procariotelor (lipsa unor organite citoplasmatice). - se caracterizează printr-o mare flexibilitate metabolică, o capacitate crescută de adaptare în condiţii foarte variate de mediu; - este un metabolism de mare intensitate ceea ce asigură o multiplicare rapidă cu timp de generaţie de 20' (celulele de mamifere au timp de generaţie de cel puţin 24 ore).
Metabolismul bacterian este reprezentat de totalitatea proceselor biochimice intracelulare prin care substratul nutritiv este transformat în energie şi constituenţi celulari. Este de două tipuri:
- energetic - prin care se asigură respiraţia bacteriană şi este realizat prin reacţii catabolice; - de sinteză - prin care se asigură sinteza componentelor bacteriene şi se realizează prin
reacţii anabolice şi catabolice (reacţii biochimice de oxido-reducere, în cazul bacteriilor chimiosintetizante).
Respiraţia bacteriană este: în funcţie de natura acceptorului final de hidrogen- există tipuri de respiratii: o aerobă (oxibiotică) -acceptorul final este oxigenul atmosferic, 02, iar produsul rezultat este apa.

Dr. biolog D. Gheţeu - Microbiologie
27
o anaerobă (anoxibiotică) - acceptorul final este orice substanţă anorganică diferită de oxigen, ex. CO2 sau o substantă organică (fermentaţie). în funcţie de potenţialul redox, bacteriile pot trăi în medii cu cantitaţi variate de oxigen şi se împart în patru tipuri respiratorii: o bacterii strict aerobe - cresc numai în prezenţa unei cantităţi mari de oxigen; o bacterii strict anaerobe - cresc numai în absenta oxigenului, sunt lipsite de enzimele de mai sus ; o bacterii aerobe, facultativ anaerobe - au posibilitatea ambelor tipuri de respiratie, în funcţie de existenţa oxigenului. o bacterii anaerobe microaerofile - tolerează cantităţi mici de oxigen, probabil prin achiziţia unor enzime de tip catalazic. Metabolismul de sinteză
Este suma proceselor metabolice, ce concură la procurarea de materiale convertibile în energie şi componente celulare, necesare creşterii şi multiplicării bacteriene.
În functie de modul de procurare a acestor elemente, bacteriile sunt de trei tipuri : Autotrofe: utilizează pentru sinteze azotul şi carbonul din compuşi anorganici simpli: CO2, NH3, NO2, NO3. Heterotrofe: utilizează pentru sinteze azot şi carbon din substanţe organice diferite, produşi de alte organisme, putându-şi sintetiza singure metaboliţii esenţiali (majoritatea) Parotrofe: sunt bacteriile care nu-şi pot sintetiza singure metaboliţii esentiali. Ele se dezvoltă numai intracelular.Ex. Rickettsia prowazeki, agentul tifosului exantematic.
Unele bacterii sunt capabile să sintetizeze vitamine în cantităţi atât de mari încât servesc ca materie primă pentru extragerea acestor vitamine. Biotina este sintetizată de specii ca: Proteus vulgaris, Mycobacterium tuberculosis, Escherichia coli. Tiamina (vitamina Bl) este sintetizată de: E. coli, B. subtilis, S. typhi, Shigella, C. diphteriae. Piridoxina (vitamina B6) este sintetizată de unii lactobacili. Pentru om, flora microbiană intestinală constituie o importantă sursă de vitamine (vitamine din grupul B, vitamina K).
Mediile de cultură pentru bacterii Pentru identificarea unei specii bacteriene, examenul direct al produsului patologic recoltat de la bolnav este de cele mai multe ori insuficient. Precizarea cauzei care a produs boala necesită cultivarea agentului patogen pe un complex de substante nutritive adecvate care alcătuiesc mediile de cultură.
Aceste medii de cultură trebuie să îndeplineaseă anumite conditii: - să contină substanţe plastice şi energetice necesare cultivării microbului însămânţat, adică să asigure surse de azot, hidrati de carbon, săruri minerale, apă, vitamine şi factori de creştere necesare creşterii, dezvoltării şi reproducerii celulei bacteriene. - să satisfacă necesitătile de aerobioză sau anaerobioză ale bacteriei, ştiind că în timp ce bacteriile aerobe pot folosi oxigenul molecular, cele anaerobe nu se pot dezvolta în prezenta 02 liber. - să aibă o concentraţie de ioni de hidrogen (pH) optimă. - să fie sterile, pentru a permite izolarea în cultură pură a germenului respectiv.
Mediile de cultură sunt sub formă lichidă (bulion, apă peptonată, etc.) sau solidă (medii în care se încorporează agar).
Cultura bacteriană este rezultatul înmulţirii germenilor într-un mediu favorabil. Aspectul culturii depinde de: specie, de varsta culturii şi de mediul de cultură şi are rol în identificare. Bacteriile pot fi cultivate pe: medii artificiale (acelulare), culturi celulare, ou embrionat de găină, animale receptive (sisteme biologice sensibile -SBS).
Culturile bacteriene pe medii artificiale. În medii lichide - determină apariţia unei turbidităti (tulburări a mediului apos) după 18-20
ore în condiţii de crestere specifice bacteriei. În funcţie de aspectul turbidităţii culturile pot fi: Culturi "S" (smooth) - turbiditate omogenă, - germenii sunt tineri. - virulenţi, care disperseză rapid şi uniform.

Dr. biolog D. Gheţeu - Microbiologie
28
Culturi "R” (rough) - mediul este limpede. - germenii se găsesc sub formă de grunji (depuşi pe fundul vasului sau pe pereţi). - tulbură neomogen mediul. Sunt determinate fie de germeni avirulenţi, nepatogeni, sau de germeni patogeni îmbătrânite, care şi-au pierdut virulenta. Uneori creşterea în mediile lichide mai poate fi sub formă de: văl la suprafaţă, în cazul germenilor aerobi (bacilul tuberculos) sau depozit gros pe fundul vasului în cazul germenilor anaerobi.
Pe medii solide - produsul patologic se însămânţează prin diverse procedee de dispersie. Cultura bacteriană apare sub formă de colonie după 4 ore. Colonia bacteriană reprezintă totalitatea germenilor rezultaţi din multiplicarea unei singure celule bacteriene, ea este o cultură pură, sunt germeni identici genetic (monoclonali). Aspectul coloniilor Colonii de tip "S", Smooth = neted. Coloniile au suprafaţa netedă, lucioasă cu margini drepte, de consistenţă untoasă, cu un diametru de 1-2 mm, se pot detaşa uşor, sunt alcătuite din germeni virulenti. Colonii de tip "R", Rough = aspru. Coloniile au suprafată aspră, rugoasă, cu margini neregulnte, crenelate, nu se pot detaşa o parte din colonie cu ansa, ci doar colonia întreagă. Aceste colonii sunt formate din germeni cu structură antigenică afectată, batrâni, aviruleţii. În această formă nu se mai pot face identificări. Colonii de tip "M" care au suprafaţa netedă, mucoasă, sunt variante ale coloniilor "S" ale germenilor cu capsulă (Klebsiella). Ele au tendinţa de a se confrunta intre ele. Colonii de tip "G" sunt colonii pitice, ele rezultă în urma acţiunii antibioticelor asupra formelor "S" de germeni.
Populaţia care rezultă prin diviziunea unei bacterii creşte în progresie geometrică cu raţia 2. Timpul necesar pentru dublarea populatiei se numeşte timp de dedublare sau timp de generatie. Dinamica reală este caracterizată printr-o curbă cu patru faze.
1. Faza de lag - în primele 2 ore, numărul de indivizi este aproape constant (faza de latenţă), ea este o fază de acomodare a germenilor la noile condiţii de mediu.
2. Faza de multiplicare exponentială sau logaritmică Este o fază de creştere rapidiă a numărului bacteriilor. Bacteriile sunt virulente, antigenele sunt cele mai bine reprezentate. În această fază se recoltează germenii pentru prepararea vaccinurilor.
3. Faza staţionară. Prin consumarea substratului nutritiv, ritmul de diviziune scade, numărul germenilor care mor devine egal cu cel al germenilor care se divid, deci numărul total al bacteriilor rămâne constant. Această fază durează 2-3 zile. În această fază se fac identificările bacteriilor, coloniile sunt bine individualizate. Bacteriile din colecţiile purulente (abcese) se află în aceasta fază. 4. Faza de declin. Consumarea substratului nutritiv şi acumularea de metaboliţi toxici duce la moartea majoritătii bacteriilor. Celulele rămase vii sunt bătrâne, îşi pierd virulenlaţa şi patogenitatea. Faza durează 2-3 luni. În această fază apar sorii.
III. 3 Patogenitate Acţiunile agenţilor patogeni se manifestă prin: Adeziune. Participă adezinele bacteriene prin fimbrii; actiune obligatorie de penetrare în organism; flora bacteriana rezidentă se opune ca o "barieră" proliferării bacteriene exogene. Există receptori corespunzători pe mucoase. Invazia - prin endocitoză. Achiziţia factorilor nutritivi. Secreţia toxinelor
exotoxine - proteice: tetanos, difterie, botulism, scarlatina, gangrena, holera;

Dr. biolog D. Gheţeu - Microbiologie
29
endotoxine - lipopolizaharidice, constituente ale peretelui BGN - antigen O somatic, eliberate prin liza bacteriană, şoc endotoxinic;
Exemple de sinteză de enzime: coagulaza - S. aureus, streptokinaza - S. pyogenes, betalactamaze - S. aureus.
Însămânţarea Medii de cultură Antibiograma Figura 5. Medii şi tehnici utilizate în microbiologie.
Multiplicarea bacteriană: mecanism principal, bacterii cu multiplicare extracelulară; Acţiune antifagocitară prin: polizaharidul capsulei S. pneumoniae - (serotipuri), proteina M a peretelui S. pyogenes,- piogene - (septicemii, supuraţii, abcese); bacterii cu multiplicare intracelulară facultativă: în macrofage; micobacterii, brucele, produc boli lente, cu recăderi, hipersensibilizare de tip întârziat. bacterii cu multiplicare intracelulară obligatorie: Chlamydia; reacţii de hipersensibilizare (patogenia poststreptococică). Curs IV
Coci şi bacili patogeni patogeni Coci patogeni Stafilococii sunt coci gram-pozitivi, aerobi, imobili, nesporulaţi ce se dispun caracteristic
în „grămezi”, „ciorchini”, atât în produsele patologice cât şi în culturile pe medii solide. Genul Staphylococcus este împărţit în: Staphylococcus aureus, epidermidis şi saprophyticus. Din punct de vedere clinic este mai importantă clasificarea patogenă care crează două mari categorii: Stafilococii secretori de coagulază (foarte patogeni); Stafilococii coagulazo-negativi (mai puţin patogeni, S. epidermidis, S. saprofiticus).
Staphilococcus albus S. aureus
Figura 6. Colonii de Staphilococcus. Speciile de importantă medicală sunt: Staphylococcus aureus, Staphylococcus
epidermidis (care face parte din flora indigenă), Staphylococcus saprophyticus care poate da afecţiuni în sfera uro-genitală. Caractere morfologice. Sunt coci Gram pozitivi, cu de 0,8-1,5µm, în diplo sau grămezi neregulate (staphylos = ciorchine), necapsulaţi (cei mai multi), imobili, aerobi, facultativ anaerobi, saprofiţi sau patogeni. Posedă: catalază, coagulază, fosfatază, descompun glucoza prin fermentaţie.

Dr. biolog D. Gheţeu - Microbiologie
30
Caractere de cultură. Cresc pe medii simple şi în special pe mediile hiperclorurate (10-15% NaCI). Pe mediile solide formează colonii de tip "S" (smooth = neted). Elaborează pigmenţi endogeni:
- S. aureus elaborează pigment auriu - S. epidermidis pigment alb - S. saprophyticus pigment galben. Streptococii fac parte din familia Streptococcaceae, genul Streptococcus, unele specii
sunt patogene, altele fac parte din flora indigenă a orofaringelui, tractului intestinal, pielii. Cele mai importante specii pentru patologia umană sunt:
- Streptococcus pyogenes, - Streptacoccus agalactiae, - Enterococcus, - Streptococii viridans, - Streptococcus pneumoniae
Morfologie: Sunt coci dispuşi în lanţuri, Gram pozitivi, sunt saprofiţi, patogeni sau condiţionat
patogeni. Au dimensiuni de 0,5-1 µm, au în peretele bacterian o concentratie de mureină mare (80%), sunt imobili, nesporulaţi, uneori pot prezenta capsulă, ce le conferă o mare virulenţă.
Habitat: sunt ubiquitari (întâlniţi peste tot) şi în special pe tegumente şi mucoase. Caractere de hemoliză Dimensiunile coloniilor de streptococ sunt mult mai mici (cca. 0,5 mm) decât de
stafilococ (care au un diametru de 0,5-2 mm), sunt incolore şi dau hemoliză. În funcţie de elaborarea hemolizinei streptococii se împart în 4 categorii. Streptococii hemolitici (elaborează hemolizina β), dau hemoliză beta, care este completă, foarte întinsă, coloniile având marginile clare. Din această categorie fac parte serogrupurile: A (care este totdeauna patogen), C. G.
Rezistenţa - au rezistenţă scazută sunt distruşi de razele ultrviolete, dezinfectante şi antiseptice la concentraţii uzuale (sunt distrusi la 55oC timp de 20 min.); - streptococul din grupa A este sensibil la pencilina G, care se administrează parenteral/oral - streptococul din grupa B este rezistent la concentntii mari de NaCI 6.5%, la variatii mari de temperetura, la acţiunea bilei şi a sărurilor biliare şi la actiunea multor antibiotice.
Patogenitate Streptococii grup A pot da infecţii acute ale: tractului respirator (forma eruptivă-
scarlatina, forma neeruptivă: angina, faringita, etc); otita medie, sinuzita, meningita, pneumenie, bronhopneumonie, pleurezie, endocardita acută.
Localizarea cutanată a infecţiei streptococice este mai frecventă în zonele calde: intertrigo impetigo, zabăluţă (infecţia comisurii bucale), abcese. flegmoane.
Erizipelul este o infecţie streptococică cu localizare cutanată (atunci când există o soluţie de continuitate): la nivelul gambelor, feţei, cu caracter recidivant.
Virulenţa streptococilor piogeni este dată în special de proprietatea antifagocitară, care depinde de prezenţa componentelor celulare de suprafaţa: acidul hialuronic capsular şi proteina M.
Streptococcus pneumoniae (pneumococul) Este o bacterie comensală, care colonizează în mod normal tractul respirator superior, dar
şi tractul intestinal şi mucoasele uro-genitale. Procentajul de purtători oro-faringieni sănătoşi în populaţie este estimat la 30-70%. Bacteria este condiţionat patogenă, putând provoca îmbolnăviri în anumite condiţii. Este unul din principalii agenţi patogeni infecţioşi la vârsta copilăriei, precum şi la persoanele vârstnice şi/sau care au boli cronice sau trăiesc în condiţii speciale.
Morfologie - Sunt coci lanceolati dispuşi în diplo cu dimensiuni de 1/1,5µm, pneumococii virulenti au capsulă care înconjură ambele celule, dispuşi cu capetele rotunjite fată in fată. - Sunt imobili, nesporulaţi, Gram pozitivi, majoritatea sunt saprofiti, dar sunt şi tulpini patogene nu se găsesc în natură.

Dr. biolog D. Gheţeu - Microbiologie
31
Caractere de cultură: Cresc numai pe medii ce conţin sânge, cei încapsulati dau colonii de tip „S'. Sunt hemolitici, coloniile sunt mici, transparente şi înconjurate de o zonă de hemoliză a, ca la streptococii viridans.
Rezistenţă: Sunt sensibili în mediul extern, sunt distruşi de razele solare, de dezinfectante la concentraţii uzuale. Majoritatea tulpinilor sunt sensibile la penicilină.
Structură antigenică. Are antigene somatice şi solubile, cel mai important este Ag SSS (Substanta Solubilă Specifică), din constituţia capsulei, având o structură polizaharidică. Ag SSS împarte pneumococii în 83 de serotipuri (serotipul 3 dă cea mai mare capsulă).
Caractere de patogenitate. Pneumococii încapsulaţi sunt patogeni prin multiplicare şi invazivitate, nu au capacitate de toxigeneză. Determină pneumonia francă lobară (localizată doar la nivelul unui singur lob).
Figura 7. Streptococcus pneumoniae (pneumococul): imagine
la microscopul electronic (stânga) şi aspectul culturii (dreapta).
Pneumococul poate provoca următoarele tipuri de îmbolnăviri: - infecţii de vecinătate, prin răspândire în regiunile anatomice învecinate căilor respiratorii superioare: sinuzite, otită medie, otomastoidite. - infecţii invazive, prin diseminare sanguină (bacteriemie), cu constituirea de focare infecţioase la distanţă: septicemie şi meningită, care pot duce la deces, dar şi endocardită, pericardită, artrite septice etc. - pneumonie şi alte infecţii ale tractului respirator inferior, prin ambele mecanisme descrise anterior.
Neisseria meningitidis (meningococul) Germenii din acest gen sunt coci de formă reniformă, cu părtile adiacente concave, în
diplo, Gram negativi, uneori cu o tendintă de a rezista decolorarii. Toate speciile sunt aerobe, imobile, nu au flageli, unele tulpini posedă pili.Temperatura optimă de dezvoltare este între 35-37° C (sunt stenotermi - rezistă la variaţii mici de temperatură).
Neisseria meningitidis este reprezentată de coci ovalari, reniformi de dimensiuni de 0,6 - 1,0 µm dispuşi în diplo, Gram negativi. În produsul patologic (LCR) sunt intra- şi extracelulari.
Se gasesc numai la oameni (bolnavi sau sănătoşi). Sunt localizaţi pe tegumente, dar mai ales în cavitatea nazală şi bucală.
Caractere morfologice: germenii în diplo sunt înconjuraţi de o microcapsulă; în peretele lor predomină antigenul “O”, nu mureina, fiind Gram-negativi. Sunt germeni imobili, nesporulaţi.Unele tulpini prezintă pili în momentul izolarii, acestia pot să dispară dupa 1-2 subcultivări, nu par să aiba prea mare importantă în virulenţa germenului.
Meningococii sunt germeni patogeni, dar cu virulenţă scăzută, 60-70% din infectiile meningococice sunt inaparente. Cea mai frecventă localizare este cea nazală, foarte rar infecţia poate ajunge pe cale sanguină la meninge, unde va produce meningita (1 caz la 1000).
Dau rinite meningococice (cel mai frecvent), sinuzite, faringite, meningita cerebro spinală meningococică (MCSM). Patogenitatea se datorează în special proprietătilor sale antifagocitare date de către polizaharidul capsular şi de lipopolizaharidul care se află în peretele bacterian.
Meningita fulminantă (sindromul Waterhouse-Friederichsen): ▪ Este o formă supraacută a infecţiei sanguine cu meningococ. ▪ A fost considerat iniţial ca o formă fulgerătoare de septicemie meningococică prin prăbuşirea funcţiilor suprarenale datorită leziunilor hemoragice care se produc în această glandă. ▪ În această formă fulminantă, este caracteristic examenul LCR, este aproape clar, conţine un număr mic le PMN şi un numar mare de germeni.

Dr. biolog D. Gheţeu - Microbiologie
32
▪ Decesul pacientului poate surveni în 24-36 ore, dacă nu se face tratamentul corespunzător. Neisseria gonorrhoeae (gonococul) Neisseria-gonorrhoeae sau gonococul produce infecţia gonococică. Această bacterie a
fost descrisă de către Albert Neisser în 1879 în uretrite acute la bărbaţi şi în conjunctivita nou-născuţilor. N. gonorrhoeae este un parazit strict pentru specia umană. Se grefează pe celulele mucoasei genitale şi accidental pe alte mucoase: rectală, faringiană, conjunctivală. Conjunctivita care poate ajunge până la oftalmia gonococică la nou-născut este o infecţie exogenă Produsă în timpul naşterii atunci când mama are vaginită gonococică. La bărbat principala formă de boală este uretrita gonococică, la femei vaginita şi metrita.
Caractere morfo-tinctoriale. Sunt coci ovalari de 0,6-1,0 µm, în diplo, cu părtile concave adiacente, asemănători boabelor de cafea. Sunt Gram negativi, imobili, nesporulaţi, înconjurati de o capsulă comună. La suprafafaţa celulei au pili cu rol determinant în ataşarea bacteriei de celula gazdei, deci se găsesc la speciile virulente, lipsind la cele nevirulente sau la cele din colectiile vechi. Caractere de cultură. Gonococul nu se dezvoltă pe medii simple, este germenul cel mai greu de cultivat dintre toate Neisseriile.
Figura 8. Neisseria gonorrhoeae (gonococul) în examen direct la microscopul optic.
Bacilii patogeni Genul Escherichia face parte din familia Enterobacteriaceae1, sunt bacterii saprofite sau
condiţionat patogene. Cea mai importantă specie este Escherichia coli. În caz de boală aceasta poate fi pusă în evidentă în: sânge, urină, puroi, fecale, etc..
Sunt bacili Gram negativi, lungi de 2-3µm şi groşi de 0,5µm, ciliaţi, ceea ce face ca majoritatea să fie mobili; ei nu sunt sporulaţi.
Cei mai multi colibacili nu sunt patogeni. Virulenţa şi toxicitatea bacteriilor patogene sunt legate de prezenţa Ag K şi a unor toxine (enterotoxina, neurotoxina etc.) precum şi producerea hemolizinei şi factorului citotoxic necrotizant.
În anumite conditii E. coli poate să dea naştere la diferite afecţiuni locale sau generale: - afecţiuni genito-urinare (85 % patotipuri uropatogene datorată unor adezine, hemolizine, aerobactinei, care au receptori pe tractul urinar), - infectii intestinale.
Sunt 6 patotipuri diareigene: ETEC(enterotoxigen), EHEC (enterohemoragic),
1 Familia Enterobacteriaceae cuprinde 5 triburi, împărţite pe baza caracterelor biochimice. Tribul 1 - genul ESCHERICHIA - genul SHIGELLA Tribul 2 - genul SALMONELLA - genul ARIZONA - genul CITROBACTER Tribul 3 - genul KLEBSIELLA - genul SERRATIA - genul ENTEROBACTER Tribul 4 - genul PROTEUS
- genul PROVIDENCIA Tribul 5 - genul YERSINIA Genurile Shigella şi Salmonella sunt întotdeauna patogene. Celelalte genuri sunt saprofite, condiţionat patogene, cu excepţia speciei Yersinia pestis care este întotdeauna patogenă (produce ciuma sau pesta).

Dr. biolog D. Gheţeu - Microbiologie
33
EIEC (enteroinvaziv), EAggEC (enteroagregativ), EPEC(enteropatogen), DAEC (enteroaderent difuz).
Mai poate genera: endocardite, meningite, septicemii. Răspunsul imun este slab. Genul Salmonella. Este un grup al familiei Enterobacteriaceae, tribulul 2, format din bacili
Gram negativi, mobili, nesporulaţi, ce au o structură antigenică complexă şi patogenitate ridicată pentru om.
Clasificarea pe baza structurii antigenice a fost alcătuită o schemă de diagnostic în care genul Salmonella a fost împărtit în grupuri (cu antigene somatice comune) fiecare grupă este formată din specii care se diferentiază prin antigenele flagelare specifice. Germenii din acest gen se întâlnesc la omul bolnav sau purtătorii de germeni, la mamifere, reptile, în mediul extern: ape, sol, alimente.
Caractere morfologice şi tinctoriale. Sunt bacili cu lungime de 2-3 µm şi groşi de 0,6 µm, mobili, cu cili peritrichi bine dezvoltaţi, nu formează spori, majoritatea speciilor nu au capsule, sunt Gram negativi.
Caractere de cultură Sunt germeni aerobi, facultativi anaerobi. Cresc pe medii de cultură simple (bulion, apă peptonată, geloză nutritiva). Cresc sub forme „S" care tulbură uniform mediile lichide şi formează colonii netede,
rotunde, lucioase pe medii solide; sau sub formă „R", care sedimentează în mediile lichide şi dau naştere la colonii zbârcite, cu margini neregulate, pe medii solide.
Salmonella typhi este cea mai patogenă specie, îşi sintetizează totdeauna un flagel din proteine specifice, de aceea poate identificată. Kauffman şi White au stabilit că Salmonella cu Ag „O" = 9 sau 12 şi cu Ag"H" = d, este Salmonella typhi, iar cu Ag"H"= g sau m este Salmonella enteritidis. Acest fapt are mare importanţă practică deoarece cu ajutorul serurilor imune specifice se poate pune diagnosticul. Antigenul solubil este endotoxina, ea are un rol patogen mult mai important la salmonelele cu caracter invaziv, deoarece sunt puse în circulaţie, ducând la generalizare (febra tifoidă). Produsele patologice care se examinează sunt: sângele, măduva oaselor, materiile fecale, urina, bila, lichidul din petele lenticulare, în cazuri letale şi sângele din măduvă, fragmente din organe (ficat, splină, ganglioni mezenterici).
Caractere de patogenitate. Toate salmonelele sunt patogene, prin multiplicare şi endotoxinogeneză, iar cele ce prezintă şi o pseudocapsulă (Ag Vi") au şi caracter de invazivitate. La om salmonelozele sunt sub două forme principale: - Boli septicemice, cu evolutie gravă şi mortalitate ridicată, cum sunt febra tifoidă (produsă de bacilul tific) şi febrele paratifoide A. B şi C (produse de bacili parotidici A, B şi C). - Boli localizate, mai ales la nivelul tubului digestiv (toxiinfectiile alimentare şi enteritele), cu o durată mai scurtă şi de o gravitate mai redusă.
Imunitatea ▪ Foştii bolnavi de febră tifoidă şi parotifoidă dobândesc o imunitate destul de solidă, care poate dura toată viaţa. ▪ Imunitatea în toxiinfecţiile alimentare şi a enteritelor este slabă. ▪ Imunitatea antitifoparatifoidică se poate dobândi şi in mod artificial, prin vaccinare. ▪ Diagnosticul de laborator în infecţiile tifo-paratifoidice este bacteriologic şi imunologic
După boala sistemică apar anticorpi care protejează cel puţin 5-10 ani. Bacilul dizenteric (Shigella dizenteriae) Germenii genului Shigella produc dizenteria bacilară. Bacilii dizenterici sunt găsiţi în
materiile fecale ale bolnavilor de dizenterie şi ale purtătorilor. De asemenea se pot găsi în unele alimente contaminate, ape poluate.
Caractere morfo-tinctoriale. Sunt bacili Gram negativi, imobili şi nu sunt capsulaţi. Genul Shigella are 4 subgrupuri serologice: A, B, C, D. - Subgrupul A (Shigella dizenteriae) cuprinde 12 serotipuri. Acest grup este cel mai patogen. Shigella Shiga este cel mai patogen dintre toate tipurile, elaborează exotoxina neurotropă.

Dr. biolog D. Gheţeu - Microbiologie
34
- Subgrupul B (Shigella flexneri) cuprinde 6 tipuri, din care primele 4 au şi subtipuri. - Subgrupul C (Shigella baydii) cuprinde 15 tipuri serologice. - Subgrupul D (Shigella sonnei) cuprinde două faze: faza I (sau S) şi faza II (sau R)
Caractere de patogenitate. Se evidenţiază prin multiplicare şi toxigeneză, actioneaza la nivelul sigmoidului provocând ulceraţii superficiale. În dizenteria dată de Shigella apar trei simptome ce constituie sindromul dizenteric : scaune sangvinolente şi mucoase, colici abdominale, tenesme (senzaţie de defecare fără scaun).
În general boala evoluează cronic, uneori pe fondul unei rezistenţe scăzute, dau recidive. La copii este cea mai gravă dizenterie.Răspunsul imun este foarte slab.
Vibrionul holeric (Vibrio cholerae) Holera este o boală infecţioasă acută din grupul infecţiilor de carantină şi intestinale,
provocată de mai mulţi vibrioni (asiatic, El-tor, non O139), care pătrund în organismul uman prin tractul digestiv, caracterizându-se clinic prin simptome de deshidratare şi crampe sau convulsii musculare, manifestându-se prin diaree şi vome repetate, ce duc la dereglări circulatorii şi a metabolismului hidrosalinic. Primele 6 pandemii de holeră au fost provocate de vibrionul asiatic, iar acea de astăzi – de vibrionul El-tor. Ultimul este cu mult mai rezistent în mediul ambiant. El se păstrează destul de îndelungat la temperaturi joase (în experimente – până la 127 zile), în bazinele acvatice – câteva luni; în lapte şi produse lactate – 14-30 zile; pe suprafaţa legumelor şi a fructelor – 5-7 zile; în intestinul muştelor – 4-5 zile; în organismele hidrobionţilor (peşti, moluşte, broaşte, raci, crabi, stridii etc.) – un timp mai îndelungat.
De notat, că vibrionul El-tor după structura antigenică se subdivizează în serotipurile: Ogava, Inaba şi Higoşima. În epidemia din 1995 în ţara noastră preponderent (97%) s-a înregistrat serotipul Ogava.
Sursa de infecţie o constituie omul bolnav şi purtătorii de vibrioni. Ultimii în holera El-tor se întâlnesc mai frecvent decât în cea asiatică şi sunt periculoşi, deoarece pacientul nu are semne clinice.
Holera, ca şi celelalte infecţii intestinale, se transmite prin intermediul apei, produselor alimentare, mâinilor murdare, obiectelor de uz casnic contaminate cu excreţiile bolnavilor holerici şi prin intermediul muştelor. Un pericol evident în privinţa molipsirii de holeră El-tor îl prezintă contaminarea cu hidrobionţii (animalele acvatice sus menţionate) şi chiar consumarea în hrană a unor din ei.
Genul Corynebacterim include mai multe specii de bacili Gram pozitivi (poate fixa colorantul neregulat şi pot avea incluziuni de polimetafosfat), uşor incurbaţi, maciucaţi, aşezati în V. L mici grămezi neregulate, sau în palisade, nesporulaţi, necapsulari, imobili, aerobi, facultativ anaerobi, catalazo pozitivi. Dintre Corynebacteriile patogene pentru om, grupul Corynebacterium diphtheriae reuneşte trei specii: C. diphtheriae cu 4 biotipuri (gravis, mitis, intermedius şi belfanti ), C. ulcerans, C. pseudotuberculosis. C. diphtheriae are capacitatea de a elabora exotoxină în urma conversiei genetice.
Bacilul difteric - Corynebacterium diphtheriae este agentul cauzal al difteriei. Difteria este o boală infecţioasă gravă, care în forma ei caracteristică se manifestă printr-o leziune la poarta de intrare, cel mai frecvent la nivelul amigdalelor, unde produce un exudat fibrinos, aderent: "falsa membrană" de culoare gri-maronie (diphthera = membrană).
Ea se formează local sau se extinde acoperind faringele, obstructionand arborele traheo-bronşic producand crup laringian care poate provoca asfixie mecanică. La adăpostul falsei membrane, bacilii îşi continuă multiplicarea şi sinteza de toxină.
Tulpinile lizogene (infectate cu bacteriofag tox+) sintetizează toxina eliberată extracelular şi difuzată în organism pe cale sanguină şi limfatică produce leziuni importante la nivel cardiac (miocardită), nervos (demielinizare), renal (necroză tubulară), suprarenalism, muscular şi hepatic. Diagnosticul de laborator este bacteriologic.Este un diagnostic de mare urgentă.

Dr. biolog D. Gheţeu - Microbiologie
35
Bacilul Koch2 (Mycobacterium tuberculosis), bacilul tuberculos, manifestă un parazitism strict, el este prezent în leziunile omului sau animalelor bolnave de tuberculoză, poate fi găsit în praful din încăperile în care a fost răspândită sputa baciliferă sau pe obiectele contaminate de bolnav.
Sunt bacili subţiri cu lungimea de 3-10 µm şi lătime de 0,1-0,2 µm, uneori ramificati, drepţi sau uşor curbati sub formn literelor: X, Y, Z, cu granulaţii puse în evidentă prin coloraţii speciale, imobili, nesporulati, necapsulati.
Rezistentă. Bacilii rezistă luni sau ani în praf, sunt sensibili la acţiunea radiaţiilor solare şi UV, sunt sensibili la acţiunea antibioticelor: streptomicină, HIN, acidul para-amino salicilic, etambutol, rifampicină (sinerdol), dar pot câştiga uşor rezistentă.
Diagnosticul de laborator este bacteriologic şi se bazează prin punerea în evidenţă a bacilului Koch în produsele patologice, în acest scop se folosesc 3 metode: examenul direct, cultivarea şi inocularea la cobai.
În tuberculoza pleuro-pulmonară se recoltează: spută, aspirat bronşic sau cavitar, lichid de spalatura bronşică, piese de exereză chirurgicală, lichid de spălătură gastrică, fecale, piese necroptice, lichid pleural.
În tuberculoza uro-genitala: urină, sange menstrual, biopsie de endometru, piese de exereză chirurgicală.
În tuberculoza ganglionară: material din biopsie sau punctie. În tuberculoza ostio-articulară: lichid articular, ,puroi din abcese reci, fragmente osoase. In meningita tuberculoasă: lichid cefalorahidian. Cel mai folositor produs este sputa, pentru că tuberculoza pulmonară este forma cea mai
frecventă. Bacilul lepros (Mycobacterium leprae) este agentul etiologic al leprei umane. Lepra este o boală cu o evoluţie cronică, semnele de boală variază mult de la un pacient la altul; la congresul internaţional de combatere a leprei din anul 1953 s-a făcut categorisirea formelor de boală: 1. lepra nedefinită (prima fază cutanată eritematoasă) 2. lepra tuberculoasă (căderea părului, apar inflamaţii nodulare pe traiectul nervilor, descompuneri de ţesuturi) 3. lepra bordeline (formă instabilă, fiind definită de starea imună, care trece în forma gravă) 4. forma de lepră gravă (forma contagioasă, apar pete roşiatice, brune, „cap de leu”, inflamaţii pe traiectul sistemul limfatic şi nervos, descompunerea de ţesuturi cu tumefieri şi ulceraţii. Aceste procese se pot extinde în muşchi, oase tendoane şi organe, moartea fiind produsă de infecţiile secundare bacteriene.
Bacilul carbunos3 (Bacilus anthracis) este o bacterie gram pozitivă, care provoacă antraxul. Denumirea vine de la o leziune des întâlnită antraxul cutanat (pielea afectată este de culoare neagră, cu aspect de cărbune).
Cele mai multe cazuri de imbolnavire la om sunt reprezentate de infectiile cutanate si cele respiratorii. Afectarea intestinală apare foarte rar, iar meningita se manifestă doar în cazurile grave cu bacteriemie severă. Antraxul cutanat are o incubaţie in medie de 5 zile. Leziunile sunt localizate de obicei in
2 Genul MYCOBACTERIUM Germenii din acest gen apartin familiei Mycobacteriaceae. Această familie cuprinde mai multe specii, fiind aproape toate saprofite cu excepţia speciilor: Mycobacterium tuberculosis şi Mycobocterium leprae.
In anul 1882 Robert Koch descoperă bacilul tuberculozei; tot în acelasi an Paul Ehrlich descoperă acido-rezistenţa bacilului tuberculozei. In anul 1884 Robert Koch obţine cultura baciluli tuberculozei pe ser coagulat.
In 1891 tot R. Koch semnalează fenomenul Koch şi tuberculina. A. Calmette şi C. Guërin in perioada 1908-1921 au cultivat prin treceri la 21 zile, în bulion cartof glicerinat şi biliat o tulpină de Mycobocterium bovis virulentă izolată din lapte. In urma acestor treceri s-a obţinut o tulpină fără patogenitate, dar capabilă să determine rezistenta faţă de infecţia tuberculoasă, caracterele morfologice,tinctoriale, nutriţionale, de cultură au rămas cele iniţiale. Această tulpină (tulpina BCG - bacilul Calmette şi Guerin) a fost utilizată ca vaccin împotriva tuberculozei începând din 1921; în 1927 se organizează laboratoare de producţie a vaccinului BCG în 26 de tări , inclusiv în România. 3 Anthrakis (gr)= cărbune

Dr. biolog D. Gheţeu - Microbiologie
36
zonele expuse ale pielii: tegumentul feţei şi al membrelor superioare. La poarta de intrare apare o macula mica, rosie care ulterior va evolua catre forma de pustula si ulceratie, cu necroza si edem. Iniţial apare prurit (mancarime) iar catre final leziunea nu doare. De obicei pacienţii nu prezinta febră. Vindecarea spontană se produce in peste 90% din cazuri, dar exista si forme severe de boală, care netratate se vor finaliza prin deces. Contactul in antecedente cu animale sau produse de animale potential contaminate, orienteaza diagnosticul catre infectia cu bacilul antrax. Antraxul prin inhalaţie (boala lânarilor). Dupa cateva zile, simptomele se acutizeaza: febra crescuta, dispnee si hipotensiune arteriala. La examenul radiologic pulmonar, se observa largirea simetrica a mediastinului (mediastinita hemoragica). Antraxul gastrointestinal. Se constată o simptomatologie variabila: febra, greturi, varsaturi, dureri abdominale, scaune diareice cu sange sau chiar ascita (lichid in cavitatea peritoneala). Diareea severa determina deshidratare masiva si hemoconcentratie. Daca bacteria se localizeaza la nivelul faringelui va determina: febră, disfagie (înghiţire dificilă a alimentelor), durere în gât şi adenopatie regională.
Bacilul tetanic (Clostridium4 tetani) Este un bacil Gram pozitiv anaerob, cu capete nerotunjite de 3-4/0.4 µm, sporul terminal nu prinde coloranţi obişnuiţi diametrul transversal depăşeşte de 2-4 ori pe cel al corpului bacterian, are un contur refringent, o formă rotundă, mai rar ovalar. Bacilul tetanic este foarte mobil, prezentănd fenomenul de "căţărare' pe supafaţa gelozei, în condiţii de anaerobioză, asemănător cu cel al proteusului în condiţii aerobe.
Patogenitatea este determinată de o exotoxină pe care orice tulpină de Cl. tetani după 48-72 ore, în mediul lichid, o produce şi este suficientă pentru a provoca o intoxicaţie tetanică mortală la animalul de laborator. Acţiunea biologică a toxinei tetanice se exercită prin fixarea exclusivă şi ireversibilă pe sistemul nervos mai ales central (SNC).
Bacilul botulinic (Clostridium botulinum5). Este un bacil lung şi gros 4-6/0,8-1,2 µm, Gram pozitiv, mobil datorită cililor peritrichi, cu spor subterminal, deseori cultura poate pune în libertate mulţi spori.
Este lipsită de virulenţă, incapabil de multiplicare în organism, produce în afara organismului cea puternică toxină cunoscută, a cărei ingestie provoacă boala.
Este răspândit în sol şi se găseşte în intestinul omului şi animalelor. Sunt şapte serotipuri tipuri de Cl. botulinum (A, B, C, D, E, F, G) la noi este mai
răspândit tipul B, foarte rar tipul A. Gangrena gazoasă (Miozita clostridiană). Infecţie difuză necrozantă deosebit de gravă a
ţesutului muscular, cu extensie rapidă favorizată de prezenţa ţesuturilor devitalizate. Clostridiile gangrenei gazoase cele mai frecvente specii sunt: Cl. perfringens, Cl. histolyticum, Cl. oedematiens, Cl. septicum, Cl. sporogenes. Pot provoca la om tabloul clinic al gangrenei gazoase:
- mai frecvent la muşchii membrelor, - infecţia uterină postabortum, - infecţia post-operatorie, post-colecistectomie, - abces cerebral, infecţie sinuzală, poate afecta orice ţesut sau organ.
4 Genul Clostridium cuprinde numeroase specii, majoritatea sunt saprofite, dar un număr redus sunt
patogene. Sunt bacili Gram pozitivi, mobili datorită unor cili peritrichi (în afară de CI. perfringens) cu dimensiuni mari, strict anaerobi, formează spori ovoizi sau sferici care au dimensiuni mai mari decăt grosimea bacilului, deformăndu-l, sunt răspândiţi în mediul exterior mai ales în sol (anaerobi telurici). Patogenitatea este dată de invazivitate dar şi de toxigeneză. Anaerobii patogeni se împart în două grupe:
- anaerobi producători de toxine histotrope, clostridiiie gangrenei gazoase: Cl. perfringens, Cl. histolyticum, Cl. oedematiens, Cl. septicum, CI. sporogenes, etc.;
- anaerobi producători de toxine neurotrope: Cl. botulinum şi Cl. tetani. 5 Botulus = cărnat în limba latină. Clostridium botulinum a fost izolat în 1896 de Van Ermengem în cursul unei intoxicaţii cu cârnaţi.

Dr. biolog D. Gheţeu - Microbiologie
37
Curs 5
V.1 Leptospirele (Genul Brucella). Rickettsiile (Treponema pallidum)
Leptospirozele sunt boli cu potential septic, produse de genul Leptospira, caracterizate prin: - febră - stare septică - afectare hepatică şi renală ( icter + oligoanurie) Etiopatogenie: - Leptospira cu 280 de variante, gen Spirocetaceae (Treponema pallidum) - au forma spiralată - coloratie argentică; se vizualizează la microscopul cu fond întunecat - se găsesc în apele stătătoare: bălţi, lacuri , iazuri, mine inundate; în dejecţiile
de şobolan - frecvent şi la cabaline → extrageri manuale de placentă → veterinari. Contaminare: - pielea cu excoriaţii la oamenii desculţi, în special în orezării; - leptospirele au extremirăţile sub forma de cârlig la un capăt → capilare sanguine→ ajung
în circulaţia generală → diverse organe → ochi, rinichi, ficat, meninge, splină, plamâni. Dupa fixare la nivelul organelor, leptospirele se multiplică intens determinând leziuni
morfologice şi funcţionale, determinând şi o serie de sindroame, astfel: - la nivel pulmonar determina pneumonite - la nivelul ficatului apar leziuni datorita corpului microbian; - la nivelul rinichiului sunt afectate : interstitiul si mb. Bazala - la nivelui meningelui: plexurile coroide meningee - la nivelul ochiului: umoarea apoasa - la nivelui pielii: sdr eruptiv ( congestie tegumentara) - Starea septica induce → sdr. de CID → sdr. hemoragic. - Leptospirele dau cele mai multe cazuri de IRA; netratate duc direct la dializă - Sdr. de insuficienta hepato-renală, cu: şoc toxic, acidoză metabolică, transpiraţii reci,
hemoragii Tipuri de leptospiroze întâlnite: - leptospiroza cu L. pomona ( sursa de infectie porci şi sobolani) - leptospiroza cu L. Grippo- Typhosa (şoareci de camp, vite, cai) - leptospiroza cu L. canicola (câinele) - leptospiroza cu L. hyos (porc, soareci, vidră). Genul Brucella cuprinde şase specii dar trei sunt de interes uman producând
antropozoonoze: • Brucella melitensis (capre, oi), • Brucella abortus (vaci), • Brucella suis (porci). Sunt cocobacili (bacili foarte mici) cu lungimea de 1 µm şi grosimea de 0,3µm; sunt
încapsulaţi, imobili, nesporulaţi, aerobi, Gram negativi. Se găsesc în organismul infectat, animalele domestice care elimină bacteriile prin produsele
de avort, secreţii, lapte etc., urmate de contaminarea masiva a mediului extern (sol, apă, vegetaţie) şi de infectare a altor animale, omul fiind o gazdă accidentală, apărând ca verigă finală în lantul de transmisie a infecţiei.
Brucella se multiplică intracelular, de aceea este rezistentă la antibiotice. Calea de intrare este digestivă, prin alimentele contaminate, sau prin contaminarea tegumentelor (însă nu pe cele intacte). Animalul gestant bolnav prezintă avort spontan.
Bruceloza se mai numeşte febră ondulantă. După intrarea în organism cocobacilul se multiplică intracelular în ganglionii limfatici şi apoi pătrunde în circulatia sanguină.

Dr. biolog D. Gheţeu - Microbiologie
38
Boala are o evoluţie cronică, apar leziuni de tip proliferativ numite granuloame. Gronuloamele se observă la nivel cutanat, în ficat, splină,. sistem nervos, boala fiind o septicemie cu evoluţie lentă.
Rickettsiile sunt microorganisme care se situează la graniţa dintre bacterii şi virusuri. Se asemără cu bacteriile prin: dimensiuni, morfologie, organizare celulară internă, compoziţie chimică, multiplicare prin divziune directă. Se aseamănă cu virusurile deoorece nu cresc pe medii acelulare, ele se multiplică numai in ţesuturi vii, ca virusurile.
Familia cuprinde trei genuri: Rickettsia, Coxiella şi Ehrlichia. Aceste microorganisme sunt transmise de artropode (păduche, purice, căpuşă) multiplicându-se în corpul acestora. Specia tip al acestei familii este Rickettsia prowazeckii, care este specifică omului, celelalte specii au ca sursă animalele.
Figura 9. Căi de infecţie şi transmitere a brucelozei.
Rickettsia prowazeckii este agentul etiologic al tifosulul exantematic epidemic. Boala
apare în urma înţepăturii de păduche, păduchele face infecţia şi elimină bacteriile prin dejecţii pe pielea noii gazde, eliminând şi o substanţă pruriginoasă care determină scărpinarea. Prin leziunile care apar, rickettsiile pătrund in organism şi se multiplică în endoteliul vaselor mici.
Este caracteristică lezarea endoteliului vascular cu formarea de infiltrate perivasculare (nodulii Fraenkel) cu limfocite, monocite, histocite şi plasmocite, mai ales cutanat, sistemul nervos şi mai rar in miocard şi corticala renală.
După 7-14 zile de la înţepătură apare febră de 40-410 C care durează o săptămână. După a 5 a zi de febră apare erupţia, pete roşii. Această boală este o boală infectocontagioasă foarte gravă cu declarare obligatorie şi face parte din grupul bolilor pestilenţiale.
Treponema pallidum este agentul etiologic al sifilisului, cu transmitere în primul rând sexuală. Este un parazit exclusiv al omului şi nu se întâlneşte în mediul exterior, având o rezistenţă foarte scăzută.
Morfologie • Se examinează la microscopul cu fond întunecat. • Are o lungime de 10-15 µm, o grosime de 0,2-0,3 µm, formă helicoidală, spirele sunt în numar de 10-15 strânse şi regulate cu capetele efilate. • Între perete şi membrană are fibrile, care îi conferă mobilitate, prezintă mişcări de rotaţie şi flexie. • Se colorează Giemsa slab. • Se colorează bine prin impregnare argentică, coloralia Fontana-Tribondeau.
Patogenitate • Treponema pallidum este patogenă pentru om prin multiplicare intracelulară şi invazivitate. • Boala se numeşte SIFILIS sau LUES.

Dr. biolog D. Gheţeu - Microbiologie
39
• Boala se transmite prin contact sexual îi mult mai rar nesexual (cel mult 57, din cazuri, in urmatoarek situaţii: doică sănătoasă alăptând copil cu sifilis congenital sau doică bolnavă alăptând copil sănătos, transfuzie, medic consultând sau tratând fără masuri de protecţie bolnavi de sifilis). • Sifilisul este o boală cronică care în lipsa tratamentului (sau un tratament inadecvat) evoluează in 3 perioade distincte:
1. Sifilisul primar apare după inocularea in timpul actului sexual • Bacteriile pătrund în organism prin leziuni fine ale pielii şi mucoaselor datorită mobilitălii lor, treponemele ajung pe calea vaselor limfatice in ganglionii limfatici regionali şi apare bacteriemia. • La locul de inoculare apare după 3 săptămâni (între 10-60 zile şancrul "dur", care apare după multiplicarea treponemelor la locul de inoculare, atunci cand ating o concentraţie locală de 107 germeni/ml. • De obicei şancrul este unic şi este totdeauna însoţit de adenopatie regională, care este dură nesupurativă. • Fenomenele locale caracteristice leziunii primare se “vindecă” spontan după patru-şase săptămâni, fără a lăsa şi alte semne clinice de boală. • Anticorpii devin evidenţiabili în a doua jumătate a perioadei primare.
2. Sifilisul secundar la 2-3 luni de la contactul infectant (45-50 zile de la producerea şancrului) apar leziuni maculo-papuloase, neulcerative la nivelul tegumentelor şi mucoaselor. • Apar papule umede şi palide mai ales în regiunea ano-genitală, axilară, gură. • Se produc şi alte manifestări ca: meningita sifilitică, carioretinită, nefrită, periostitii. • În această fază se găsesc treponeme in tot organismul, bolnavul este foarte contagios. • După câteva luni (mai rar 1-3 ani) leziunile secundare se vindecă spontan, urmând o îndelungată fază de sifilis latent care poate dura ani sau zeci de ani.
- 25% din bolnavi se vindecă definitiv - în această fază 25% dintre bolnavi rămân latenţi toată viaţa. - 50% dintre bolnavi progresează către sifilisul terţiar. 3. Sifilisul terţiar este răspunsul imun celular la prezenţa Tr. pallidum în organism şi a
produşilor lor metabolici. Apare după 5-40 ani de la contactul infectant, intervalul până la debutul fazei terţiare se numeşte perioada de latenţă aparentă, fără simptome, dar cu un titru mare de anticorpi. se caracterizează prin: - leziunile granulomatoase (gome), tegumentare şi osoase, - modificări degenerative în sistemul nervos central (paralizie generală progresivă), - leziuni cardiovasculare (aortită, insuficienţă valvulară aortică), hepatice. Tratament • Se administrează penicilină. • Eficacitatea tratamentului se verifică prin diagnosticul serologic. • Dacă tratamentul este eficace, dispar anticorpii din serul de cercetat.
a b c
Figura 10. Agentul etiologic Treponema pallidum (a) şi manifestarea sifilisului la nivel tegumentar (b, c) (http://www.lib.uiowa.edu/hardin/MD/cdc/syphilis33.html).
V.2 Micologie- noţiuni generale. Dermatofiţii
Fungii sau ciupercile constituie astăzi un regn aparte. Aparatul vegetativ este alcătuit din

Dr. biolog D. Gheţeu - Microbiologie
40
filamente numite hife a căror totalitate alcătuieşte miceliul. Reproducerea la ciuperci se realizează prin spori.
După felul nutriţiei fungii pot fi paraziţi şi fungi saprofiţi. Aproape toti fungii sunt aerobi. La fungi peretele celular conţine chitina.
Înmulţirea poate realiza vegetativ prin simpla fragmentare a miceliului sau prin intermediul unor formaţiuni specializate numite spori. Sporii fungilor pot fi sexuati şi asexuati. Sporii sexuati: iau naştere direct sau indirect printr-un proces de încrucişare între 2 celule de
sex opus. Se întalnesc la fungii perfecţi.
Figura 11. Ciclul vital la fungi.
Ascospori ramân în celula în care s-au format şi poarta numele de ASCĂ. Acest tip de spori se intâlneste la clasa Ascomycetes.
Bazidiospori iau naştere pe o formatiune specializată numita bazidie, caracteristici fungilor din clasa Bazidiomycetes.
Oospori se formează din fuziunea a 2 celule de sex opus de pe hife vecine. Aparatul hifal se numeşte oogonie.
Zigospori. Sporii asexuaţi: se formează prin simpla derivare din celule vegetative prin fragmentarea unei
hife specializate sau a hifei modificate caracteristic pentru a forma corpi reproducatori. Zoospori se foremază în celule speciale denumite zoosporangi din care se elimină prin ruperea
peretelui sporangial. Sporangiospori apar în sporange care se dezvoltă la capatul terminal al unei hife denumite
sporangiofor. Sporii se eliberează prin ruperea peretelui. Conidiospori sunt aşezaţi la capatul unei hife fertile numita conidiofor în lanţuri din care se
desprind usor. Sunt caracteristici fungilor din clasa Ascomycetes. Clamidiosporii iau naştere prin dezvoltarea unui perete gros, rezistent, îin jurul unei celule din
interiorul unei hife; se intalnesc la toti fungii. Artrosporii se formeaza la fungii care au miceliu septat prin fragmentarea unei hife miceliene;
nu sunt rezistenti la uscaciune. Sporii fungilor sunt în general coloraţi şi dau coloniilor mature o culoare caracteristică;
se desprin usor de pe miceliu. Sporii adaptaţi la mediu acvatic sunt mobili cu ajutorul unor flageli. Au viaţă latentă şi o rezistenţă mare la uscăciune, presiune osmotică, temperatură şi radiaţiiUV.
Dermatofiţii produc infecţii care afecteaza zonele superficiale ale corpului: epiderma şi fanere (păr, unghii). Principalii agenţi etiologici implicaţi in producerea dermatomicozelor aparţin genurilor Epidermophyton, Microsporum şi Tricophyton.
TRICOFIŢIA este caracterizată de leziuni supurative ale pielii şi firelor de păr, cu caracter inflamator, cauzate de Tricophyton. Transmisă de la animale, se numeşte şi sicosis parazitar sau kerion celsi. Frecventă la copii, tricofiţia capului apare sub formă de placardă rotundă, bine delimitată, eritematoasă sau eritemato-veziculară, de 2-4 cm pe măsura extinderii.

Dr. biolog D. Gheţeu - Microbiologie
41
Apar: febră, dureri musculare, articulare, inflamarea ganglionilor limfatici. Pe pielea fără păr se localizează pe dosul mânii sau degetelor cu aspectul foliculitelor supurate.
MICROSPORIA data de speciile genului Microsporium, poate fi transmisă de la animale şi determină apariţia de plăci mici, rotunde de 2-5 cm cu scvame asemănătoare tărâţei. Pe pielea glabră spectul este de herpes circinat înconjurat de reacţie inflamtorie.
FAVUSUL este o dermatomicoză a pielii capului şi glabre caracterizată de leziuni tipuce sub formă de godeuri galbene cu centrul deprimat în care se găsesc adevărate colonii. în jurul perilor parazitaţi. Cauzată de Achorion sp., adesea transmisă de la animale (A. quinckeanum- la şoareci).
În toate cazurile, măsurile profilactice şi de combatere presupun igiena strictă, localizarea şi îndepărtarea sursei de infecţie şi tratamentul cu antimicotice.
Fungii dermatofiţi se individualizează prin cel puţin 2 caractere cu impact direct asupra identificării lor în laboratorul clinic:
- folosesc keratina ca element nutritiv (sursa de azot), ceea ce explica tropismul lor pentru epidermă, păr şi unghii; sunt de obicei incapabile să penetreze ţesutul subcutanat;
- prezintă caractere morfologice particulare: hife în forma de candelabru, de coarne de cerb, uneori spiralate şi macroconidii caracteristice, complet diferite de ale altor fungi.
Genul Tricophyton este capabil să invadeze părul, epiderma şi unghiile, în timp ce genul Microsporum doar părul şi epiderma iar genul Epidermophyton afecteaza epiderma si unghiile. Principalele specii de dermatofiţi izolate din probe clinice sunt în ordinea frecvenţei: Tricophyton rubrum, Tricophyton mentagrophytes, Epidermophyton flocossum, Tricophyton tonsurans, Microsporum canis şi Tricophyton verrucosum.
Micozele cutanate sunt probabil cele mai frecvente infecţii fungice la om şi sunt de obicei denumite cu termenul latin tinea ("vierme inelar"). Macroscopic, leziunea are un aspect inelar, marginea sa constituind zona de infecţie activă, progresivă; vindecarea se produce în zona centrală a inelului. Pentru a indica zona corpului afectată se folosesc termeni latini: tinea corporis (corp), tinea cruris (zona inghinala), tinea capitis (scalp, păr), tinea barbae (barba), tinea unghium (unghii), etc.
Deoarece tratamentul dermatomicozelor este îndelungat, costisitor şi cu potenţial toxic, este necesar ca mai intâi să se stabilească cu certitudine diagnosticul pe baza examenului micologic.
Microscopia directă examinează scuamele sau firele de păr în preparat umed montat între lamă şi lamelă cu solutie KOH.
Invazia părului este imparţită în 3 tipuri. Locul formarii artroconidiilor (corpi formatori de spori) clasifică speciile cauzatoare ale infecţiei astfel:
-speciile ectotrix: conidiile se formează la exteriorul invelişului părului. Cuticula este distrusă iar zonele afectate arată fluorescenţă verde-galbenă la lampa Wood. Este cauzată de Microsporum canis, M. distortum, M. ferrugineum, M. audouinii.
-speciile endotrix: conidiile se formează în învelişul părului, fiecare fiind plină de spori şi hife; cuticula nu este afectată iar părul nu prezintă fluorescenţă la lampa Wood; este cauzată de speciile de Trichophyton.
Microsporum canis M. audouini Tricophyton rubrum
Figura 12. Aspect microscopic al unor agenţi etilogici pentru tinea.
Dr. biolog D. Gheţeu - Microbiologie
42
Figura 13: Dermatofitoze- aspect microscopic.
Figura 14: Epidermophyton floccosum: aspectul microscopic al ciupercii şi aspectul macroscopic al
infecţiei Factorii de risc pentru tinea cuprind:
-condiţii de umezeală, băile comunale, -status imunocompromis, sindrom Cushing -atopie, predispoziţie genetică, activităţi care presupun contactul pielii.
a b c d
Figura 15. Aspecte structurale la fungi: a, b- conidii cu conidiofori; c- miceliu septat, d- Aspergillus fumigatus.
Aspergillus flavus Aspergillus niger Aspergillus ochraceous Aspergilom
Figura 14. Aspecte structurale diferite specii de fungi şi infecţie Genul Aspergillus este unul din cele mai bogate şi mai raspândite genuri din natură.
Unele specii sunt dăunătoare omului, animalelor şi plantelor, altele sunt folositoare fiind utilizate la producerea unor enzime, acizi organici, pigmenţi, antibiotice, diverse băuturi din orez, brânzeturi şi altele. Aspergillus prezintă hife ramificate, incolore sau colorate în nuanţe deschise şi numai la cateva specii hifele sunt colorate în brun sau de alta culoare. Conidioforii sunt lungi cu perete neted de culoare brun inchis. Sterigmele sunt uniseriate si sunt dispuse paralel cu axul conidioforului. Conidiile sunt sferice dispuse în lanţuri.
Aspergillus fumigatus este una din cele mai comune specii de Aspergillus care cauzeaza boli la oamenii cu un sistem imunitar slăbit. Se poate găsi aproape oriunde în mediul înconjurator în special în materiile organice în descompunere. Are un genom stabil, fără cicluri sexuale cunoscute; se reproduce prin formarea sporilor care sunt eliberaţi în mediul înconjurător. Se izolează pe medii uzuale, la temperatura de 25°C şi spre deosebire de alte specii de Aspergillus el este termofil (se dezolta bine la 48°C ).
Este capabil să se dezvolte la temperatura corpului uman; sporii sunt inhalaţi de aproape

Dr. biolog D. Gheţeu - Microbiologie
43
toata populaţia şi în mod normal eliminaţi rapid de catre sistemul imunitar. Este agentul cauzal al diverselor aspergiloze intalnite la om cu localizari diverse: pulmonare, nazale, oculare, cerebrale, osoase, cardiovasculare si viscerale. În aspergiloza pulmonară simptomele pot include: febra, slăbiciune generală, dureri de piept, pierderi de greutate, sânge în urina, linii roşii subţiri sau vase de sânge sparte sub unghii.
Alternaria alternata este un mucegai de interior (se dezvolta în spaţiile închise), destul de comun. Intr-un studiu efectuat a fost găsit în 87% din casele examinate. Este considerat unul dintre cele mai importanţi alergeni; Alternaria alternata a fost implicat în cazurile de alergie la persoanele care au fost expuse la lemn, ori rumeguş de lemn, de asemenea cauzează pete maronii pe plante. CURS VI
Candidoza şi actinomicoza
Candidoza bucală este determinată cel mai frecvent de Candida albicans, dar pot fi implicate şi alte genuri precum Candida tropicalis, Candida krusei, Candida parapsilosis sau Candida glabrata. Inmulţirea acestei ciuperci este controlată de o serie de bacterii prezente la nivelul mucoasei bucale. Apariţia unei noi bacterii la nivelul gurii ce rupe echilibrul dintre microorganismele deja prezente aici, va permite ciupercii să se inmuţeasca în mod necontrolat. Candidoza bucală este mai frecventă la nou-nascuţi, sugari şi batrâni.
Unele medicamente permit ciupercilor să se înmulţeasca necontrolat: - antibioticele, în special cele cu spectru larg (de exemplu, tetraciclina); - contraceptive orale (pilulele); - medicamente ce scad capacitatea de apărare a organismului precum corticosteroizii.
Modul de transmitere a ciupercii ce determina candidoza bucală diferă în funcţie de vârstă.
a b c
Figura 16. Candida albicans: agent etiologic (a-MO, b-ME) şi aspect al infecţiei în candidoza bucală (c).
Candidoza bucală, în forma sa uşoară, poate ramâne asimptomatică. In formele medii sau severe apar simptomele.
La sugari - pete (depozite) de culoare albă localizate la nivelul gurii şi pe limba cu aspect brânzos. Candidoza bucală poate fi confundata cu depozitele de lapte ce raman dupa alaptare. Aceste pete alb-branzoase sunt aderente la mucoasa si nu pot fi indepartate cu usurinta. Daca sunt frecate, pot sangera - limba si gura devin dureroase si pot apare dificultati la inghitire; - scăderea apetitului: sugarul poate refuza masa, mama confundând uşor acest simptom cu starea de saţietate (lipsa foamei) a copilului. In aceste conditii, sugarul poate deveni agitat; - eritemul fesier: apare atunci cand infecţia micotică se extinde si la nivel intestinal, ciuperca fiind prezenta în scaunul sugarului; - mameloane dureroase şi eritematoase (roşii) la mamă: candidoza bucală a sugarului poate trece neobservată până în momentul în care mameloanele mamei devin roşii şi dureroase, în special în timpul sau după alăptare.
La adulţi - senzaţie de arsură la nivelul gurii şi a gâtului (de obicei acestea sunt simptome incipiente); - pete albe aderente de mucoasa gurii şi a limbii. Ţesutul din jurul acestor depozite albicioase

Dr. biolog D. Gheţeu - Microbiologie
44
este roşu şi dureros. Daca este frecat (în timpul periajului dinţilor, de exemplu) poate sângera; - apariţia unui gust neplăcut sau a dificultăţilor de a gusta alimentele.
Dacă infecţia micotică nu este tratată sau nu se remite spontan există riscul ca aceasta să afecteze şi alte organe: la nivelul gatului (esofagului), a vaginului sau a pielii.
Pentru a preveni apariţia candidozei bucale ar trebui urmate recomandarile de mai jos: - menţinerea unei igiene orale riguroase, aceasta implicând perierea dinţilor, curăţarea acestora cu
aţă dentară şi folosirea unei ape de gură antiseptice zilnic. Daca pacientul a prezentat în antecedente o infecţie micotica a gurii, se recomanda schimbarea periuţei de dinti;
- spălarea mâinilor; - dacă se foloseste un antibiotic lichid se recomandă clătirea gurii cu apă imediat după
administrarea acestuia; - tratarea afecţiunilor medicale ce favorizeaza apariţia candidozei, precum diabetul zaharat,
neoplasmele sau HIV; - folosirea unui spacer pentru a administra corticosteroizii inhalatori, se recomanda clătirea gurii
cu apă după fiecare administrare; - la pacienţii cu SIDA se recomandă clatirea zilnică a gurii cu o soluţie antiseptică.
Figura 17. Mecanismul infecţiei în aspergiloza pulmonară: Aerosoli cu conidii de Aspergillus fumigatus inhalaţi ajung în alveole unde, in mod normal sunt eleiminate de macrofagele alveolare şi PMN. Imunitatea scăzută perimte germinarea conidiei şi invazia. A fost semnalată de asemenea invazia directă a epiteliului ciliat.
Figura 18. Mecanismul infecţiei în candidoză: Candida albicans traversează microvilii direct sau prin germinaţie şi ajunge, prin intermediul circulaţiei la organe unde se localizează de obicei în cortex unde formează pseudohife. Normal sunt atacate şi eliminate de mononucleare şi PMN. Factorii virulenţei (adezinele, fenotipuri variabile, proteinele antioxidante sau enzimele invasive) favorizează invazia în organism.
Candidoza genitală este dată tot de Candida albicans. La femei:
- vulvo-vaginita - inflamaţie a vulvei şi a vaginului (vezi anatomia feminina) - aparitia unei scurgeri (în cantitate mare) alb-cremoasă, urât mirositoare. Ca la majoritatea infecţtiilor apare şi o mancărime jenantă, senzaţii de durere sub forma de arsură fără o localizare precisă. De obicei se asociază şi cu infecţie la nivelul aparatului urinar determinând dureri la micţiune (urinare).
- netratată - se poate extinde, de la o simplă vulvo-vaginită, până la nivelul întregului perineu (zona dintre picioare), la nivelul stinghiilor, coapselor, zona anusului. In timp mucoasa vaginală sufera un proces de atrofie determinând dureri continui.
La bărbaţi - balanita sau balanopostita - inflamatia glandului şi a şantului dintre gland şi prepuţ - se manifestă
mai sever cu aparitia unor bubiţe cu puroi care se pot sparge sau se usca; în unele cazuri apare şi o pseudomembrana alb-gălbuie. Apar senzaţii intense de mâncărime local, dureri sub forma de arsură.
- netratată - se extinde ajungand pe scrot şi zona stinghiilor. La ambele sexe se poate manifesta doar prin atingere urinară - la nivelul uretrei -
scurgere abundentă / redusă de puroi, cu urini tulburi, dureri la micţiune, micţiuni dese, prurit. Este definit şi termenul de candidoza conjugală când cei doi parteneri sexuali au o

Dr. biolog D. Gheţeu - Microbiologie
45
candidoză genito-urinară care se manifestă (uneori femeia poate purta fără nici o manifestare o infecţie cu Candida).
Tratament - dupa un examen la un medic specialist care a indicat şi obţinut rezultatele unui examen microscopic ce pune în evidenţă ciuperca, tratamentul se poate începe cu o gama foarte larga de antimicotice: Amfotericina B, Nistatin, Natamicina, Terbinafina, Itraconazol, Fluconazol, Butoconazol, Clotrimazol, Ketoconazol, Flucitozina, Ciclopiroxolamina, etc.
Tratamentul se efectuează concomitent la ambii parteneri sexuali. Tratamentul cu antimicotice trebuie susţinut şi de o foarte bună igienă locală prin spălături zilnice (chiar de mai multe ori pe zi) cu săpunuri care sunt neutre sau bazice, spălături în care s-a dizolvat bicarbonat de sodiu (chiar din cel alimentar - 2 - 3 linguri la un lighenaş de apă), soluţii antiseptice pentru spălături vaginale din comerţ.
Actinomicoza este o boală parazitară cutanată (dermatomicoză) sau viscerală (visceromicoză), provocată de microorganisme (Actinomyces – ex. A. israelii, Actinobacillus), comună omului şi animalelor (în special a bovinelor, dar care apare şi la cai, porci, câini şi pisici). Se manifestă pe piele mai ales în regiunea capului, în jurul cavităţii bucale şi a gâtului sau, mai rar, la nivelul organelor (plămâni, creier, inimă, intestin, amigdale, mucoase bucale, glande salivare, etc) prin apariţia unor nodozităţi subcutanate. Cel mai frecvent, infecţia se localizează la nivelul maxilarului inferior. Nodozităţile sunt dure la început, apoi se înmoaie şi excretă puroi cu granule galbene (puroiul conţine filamentele şi sporii microorganismului parazitar). Limba se umflă şi se rigidizează (“limbă de lemn”).
Figura 19. Infecţia cu actinomicete (Actinomyces naeslundii- stânga).
Actinomicetele, agenţii etiologi ai actinomicozei se gasesc în mod normal în flora
bacteriană a cavităţii bucale şi mult mai puţin în cea a tractului gastrointestinal sau genital feminin. Aceste microorganisme nu sunt virulente şi nu pot determina o boala clinic manifestă dacă imunitatea gazdei este în parametri normali. Insă, dacă în mucoasa bucală apar mici leziuni iar ţesutul îşi pierde integritatea şi există şi fragmente tisulare devitalizate, bacteria devine capabilă să invadeze structurile profunde şi să disemineze în organism, producând afecţiunea cunoscută sub numele de actinomicoză.
Deşi actinomicetele sunt izolate frecvent din abcese, boala este considerată a fi plurimicrobiană, deoarece în unele cazuri pot fi identificate până la 10 tipuri diferite de bacterii din produsele microbiologice recoltate. Acestea sunt considerate bacterii însotiţoare, care nu pot determina singure boala dar care sporesc considerabil patogenitatea actinomicetelor, participand la stimularea secretiei de toxine si de enzime care pot inhiba raspunsul imun al gazdei. In plus, se pare ca ele cresc si capacitatea de invazie a actinomicetelor in sine. Unele actinomicete, considerate azi a fi bacterii gram-pozitive, care colonizează cavitatea bucală (Actinomyces naeslundii, Actinomyces radicidentis), produc afecţiuni stomatologice şi halenă (miros urât al gurii). Infecţiile sunt mai frecvente la persoanele care vin în contact cu animalele infestate. Este discutată şi foarte probabilă posibilitatea infestării prin resturile celulozice agricole (paie). În ceea ce priveşte speciile de microfloră, înmulţirea lor masivă are loc mai ales atunci când imunitatea este scăzută.
Actinomicoza pulmonară este o infecţie bacteriană a plamânului, ce poate avea caracter subacut sau cronic. Afecţiunea se extinde constant în ţesutul pulmonar, determinand o stare permanentă de inflamaţie granulomatoasă, precum şi formarea a numeroase abcese pulmonare. In unele cazuri, actinomicoza pulmonară poate fi insoţita si de alte localizări ale infectiei, cum ar fi regiunea cervicofacială, abdominală sau chiar pelvină (în special la femei). O dată ce infecţia incepe sa se producă, iar leziunea iniţială a permis actinomicetelor să patrundă

Dr. biolog D. Gheţeu - Microbiologie
46
profund in tesuturi, sistemul imun se activeaza şi organismul încearcă să lupte împotriva bacteriilor. Reacţia inflamatorie are drept rezultat apariţia supuraţiilor, granuloamelor şi în final a fibrozei. Infecţia se răspândeşte prin contiguitate, nerespectând planurile anatomice şi invadând din aproape în aproape ţesuturile pe care le întalneşte.
Cele mai frecvente simptome pe care le acuză pacienţii cu actinomicoză pulmonară sunt: - Durere toracică la inspir profund; - Tuse cu expectoraţie sangvinolentă sau franc hemoragică; - Febră, frisoane; - Alterarea starii generale;
- Letargie, astenie, oboseală; - Anorexie; - Transpiraţii nocturne; - Dispnee; - Scădere ponderală neintenţională.
Dacă actinomicoza pulmonară are şi alte complicaţii infecţioase (actinomicoza diseminată), pacienţii pot prezenta: modificări ale tranzitului intestinal, disconfort abdominal difuz, greţuri, vărsături (în diseminările abdominale); disconfort abdominal, scurgeri sau sângerari vaginale anormale (în cazul actinomicozei pelvine ).
Actinomicoza este o afecţiune ce poate sa apară oriunde, fiind mai frecventă în comunităţile cu un statut socioeconomic mai precar sau în cazul pacienţilor cu o igiena deficitară. Datorită tratamentului antibiotic, prognosticul bolii este favorabil, cu condiţia ca acesta să fie instituit la timp.


















