
Honiges Ana
20
UNIVERSITATEA DIN ORADEA FACULTATEA DE ŞTIINłE ANA HÖNIGES Studii ecologice şi fiziologice la Orobanche sp. în ecosistemele naturale - Teză de doctorat - Conducător ştiinŃific: Prof. Univ. Dr. Aurel Ardelean ORADEA 2009
-
Upload
nguyenhuong -
Category
Documents
-
view
238 -
download
1
Transcript of Honiges Ana

UNIVERSITATEA DIN ORADEA
FACULTATEA DE ŞTIIN łE
ANA HÖNIGES
Studii ecologice şi fiziologice la Orobanche sp.
în ecosistemele naturale
- Teză de doctorat -
Conducător ştiin Ńific:
Prof. Univ. Dr. Aurel Ardelean
ORADEA
2009

CUPRINS
SUMMARY
8
PARTEA GENERAL Ă
1. INTRODUCERE 11 1.1. Genul Orobanche şi încadrarea lui sistematică 13 1.2. Morfologie şi anatomie 16 1.3. Ciclul de dezvoltare 24 1.4. Date comparative privind speciile de Orobanche din flora
spontană a României şi cele din Baden-Württemberg
28
PARTEA EXPERIMENTAL Ă
2. OBIECTIVE DE CERCETARE 32 3. MATERIAL ŞI METODE 33 3.1. ProvenienŃa materialului biologic pentru experimentele de
laborator 33
3.2. ProvenienŃa substanŃelor chimice 35 3.3. Determinarea speciilor de Orobanche 36 3.4. Cercetare ecologică comparativă a habitatelor studiate în
flora spontană din România şi în Baden-Württemberg 37
3.5. Experimente de laborator 38 3.5.1. Analiza solului 38 3.5.1.1. Umiditatea 38 3.5.1.2. Determinarea pH-ului 38 3.5.1.3. Determinarea conŃinutului de azot amoniacal 38 3.5.1.4. Determinarea conŃinutului de azot nitric 39 3.5.1.5. Determinarea conŃinutului în fosfor asimilabil 39 3.5.1.6. Determinarea carbonului organic 40 3.5.2. Investigarea seminŃelor de Orobanche prin microscopia
electronică de scanare (SEM) 41
3.5.2.1. Prepararea probelor 41 3.5.2.2. Efectuarea microfotografiilor SEM 42 3.5.3. Investigarea seminŃelor de Orobanche la microscopul optic
şi la microscopul electronic cu transmisie (TEM) 42
3.5.3.1. Prepararea probelor 43 3.5.3.2. Fixarea şi contrastarea probelor 44 3.5.3.3. Includerea probelor în răşină epoxidică 45 3.5.3.4. SecŃionarea probelor 46 3.5.3.5. Examinarea probelor la microscopul electronic cu transmisie 46

3.5.3.6. Examinarea secŃiunilor semifine la microscopul optic 47 3.5.4. Identificare ciupercilor endofite pe baza analizei secvenŃelor
genice 47
3.5.4.1. ReacŃia în lanŃ a polimerazei (Polymerase Chain Reaction, PCR)
47
3.5.4.2. Electroforeza în gel de agaroză 50 3.5.4.3. SecvenŃierea 51 3.5.5. Experimentele de germinaŃie 53 3.5.5.1. Cultivarea în seră a plantelor gazdă 53 3.5.5.2. ObŃinerea de exsudate radiculare de la plantele gazdă 54 3.5.5.3. Prepararea soluŃiei de GR 24 54 3.5.5.4. Pregătirea seminŃelor de Orobanche pentru experimente 55 3.5.5.5. Stimularea germinaŃiei 56 3.5.5.6. Adăugarea unor potenŃiale substanŃe active 56 3.5.5.7. Analiza microscopică a germinaŃiei 57 3.5.6. Analiza instrumentală a exsudatelor radiculare 57 3.5.6.1. Separarea exsudatului radicular în fracŃiunea hidrofilă şi
fracŃiunea hidrofobă 57
3.5.6.2. Derivatizarea fracŃiunii hidrofile (fracŃiunea I) pentru cromatografia de gaz cuplată cu spectrometria de masă (GC-MS)
58
3.5.6.3. Analiza prin metoda cromatografiei de gaz, cuplată cu spectrometria de masă (GC-MS)
58
3.5.6.4. Derivatizarea fracŃiunii hidrofobe (fracŃiunea II) pentru cromatografia de lichid de înaltă performanŃă (High Performance Liquid Chromatography, HPLC)
59
3.5.6.5. Analiza HPLC 59 3.5.7. Analiza instrumentală a extractelor radiculare 60 3.5.8. Analiza solului privind potenŃialii inhibitori de germina Ńie 60 3.5.8.1. ExtracŃia solului cu acetat de etil în Soxhlet 60 3.5.8.2. Derivatizarea substanŃei uscate 61 3.5.8.3. Analiza instrumentală prin GC-MS 61 3.5.9. Analiza exsudatelor radiculare privind strigolactonele 61 3.5.9.1. ExtracŃia cu acetatul de etil şi vaporizare până la uscare 61 3.5.9.2. Pregătirea probelor pentru cromatografia de lichid de înaltă
performanŃă cu spectrometria de masă în tandem HPLC-MS/MS 62
3.5.9.3. Desfăşurarea analizelor
62
4. REZULTATE 64 4.1. StaŃiuni cercetate 64 4.1.1. StaŃiuni în România 64 4.1.2. StaŃiuni în Baden-Wüttemberg 95 4.1.3. Concluzii parŃiale privind staŃiunile cercetate 119 4.2. Rezultatele analizelor de laborator 121 4.2.1. Analiza solului 121

4.2.2. Investigarea suprafeŃei tegumentului seminal la microscopul electronic de scanare (SEM)
122
4.2.3. Investigarea secŃiunilor semifine ale seminŃelor, la microscopul optic
129
4.2.4. Investigarea seminŃelor de Orobanche la microscopul electronic prin transmisie (TEM)
131
4.2.5. Analiza ADN pentru identificarea ascomicetelor endofite 133 4.2.6. Experimente de germinaŃie 134 4.2.6.1. Experimente de germinaŃie cu potenŃiale substanŃe active 136 4.2.7. Analiza instrumentală a exsudatelor radiculare pivind
inhibitorii de germina Ńie 140
4.2.7.1. Rezultatele analizei HPLC 140 4.2.7.2. Rezultatele analizelor GC-MS 153 4.2.8. Analiza solului privind inhibitorii poten Ńiali ai germinaŃiei 156 4.2.8.1. Spectrul de absorbŃie al extractului de sol 156 4.2.8.2. Rezultatele analizei GC-MS 157 4.2.9. Analiza exsudatelor radiculare pentru identificarea
strigolactonelor
158
5. DISCUłII PRIVIND REZULTATELE 162 5.1. Aspecte ecologice 162 5.1.1. Biotopurile din staŃiunile cercetate 162 5.1.2. Clima şi vremea 163 5.1.3. Interpretarea detaliată a datelor climatologice din patru staŃiuni 167 5.1.4. Interpretarea rezultatelor analizelor solului 195 5.2. Floarea şi biologia polenizării 196 5.3. Pierderi cauzate de insecte fitofage 198 5.4. Efecte alelopatice asupra speciilor de Orobanche 204 5.5. Strigolactonele şi specificitatea faŃă de planta gazdă 208 5.5.1. Strigolactonele şi micorizele 213 5.5.2. AcŃiunea formelor stereoizomerice ale strigolactonelor 213 5.5.3. Receptorii strigolactonelor 214 5.5.4. Ipoteza lui Wegmann privind specificitatea faŃă de plantele gazdă
la Orobanche 215
5.5.5 Analiza strigolactonelor din exsudatele radiculare 216 5.5.6. Cât de specifică este specificitatea faŃă de gazdă? 217 5.6. Aspecte fiziologice la speciile de Orobanche din flora spontană,
comparativ cu speciile de Orobanche dăunătoare agriculturii 220
6. CONCLUZII ŞI CONTRIBU łII 229 BIBLIOGRAFIE 233 IERBARE 254
Extras bibliografic al lucr ărilor ştiin Ńifice din teza de doctorat

1. INTRODUCERE
Speciile genului Orobanche L. (verigel) aparŃin grupului de plante holoparazite, care reprezintă 1% din plantele superioare şi care duc o viaŃă total dependentă de planta gazdă. Ele şi-au pierdut capacitatea de fotosinteză, de aceea sustrag de la planta gazdă nu doar apă şi săruri minerale, ci şi substanŃe organice, pe seama cărora ele trebuie să îşi asigure dezvoltarea vegetativă şi reproducerea. Datorită rarităŃii şi vieŃii parazitare speciale, genul Orobanche a fost mereu considerat un grup botanic neobişnuit, studiat de un număr mic de botanişti. CunoştinŃele despre speciile relativ greu de diferenŃiat ale acestui gen au rămas fragmentare şi incomplete (Uhlich, Pusch & Barthel, 1995).
Genul Orobanche ar fi rămas doar un grup botanic deosebit dacă, din cei aproape 200 de taxoni, câŃiva nu s-ar fi dezvoltat ca buruieni rapace, dăunătoare agriculturii, capabile să distrugă recolte întregi. De aceea, în ultimele decenii, au luat o amploare deosebită cercetările şi încercarea de combatere a acestor plante parazite.
În România şi în regiunea Baden-Württemberg din Germania cresc 15 specii comune de Orobanche. Toate cele 15 specii comune se află pe Lista Roşie a plantelor în Baden-Württemberg (Breunig & Demuth, 1999); în Romania 3 specii se află pe Lista Roşie a plantelor: O. salviae, O lucorum, O. alsatica (Olteanu ş. a., 1994, Boşcaiu ş. a., 1994), iar alte 6 sunt considerate a avea diferite grade de ameninŃare, fără a fi inculse pe această listă: O. arenaria Borckhausen, O. purpurea Jaquin, probabil sporadică, O. lutea Baumgarten, sporadică, O. elatior Sutton, O. gracilis Smith, O. teucrii Hollandre, rară, O. reticulata Wallroth, O. alba Willdenow, rară (Drăgulescu, 2003; Pop, 1978).
În Flora României (Buia, 1961) sunt descrise 22 de specii ale genului Orobanche şi regiunile în care sunt răspândite. Ciocârlan (2000) descrie 21 specii de Orobanche în flora României. După Demuth (1996), în regiunea Baden-Württemberg (Sudul Germaniei) cresc tot 21 de specii de Orobanche, dintre care 20 în flora spontană.
Din cele 22 specii de Orobanche prezente în vegetaŃia din România, 16 sunt consemnate în conspectul speciilor din judeŃul Sibiu (Drăgulescu, 2003). Din acest motiv cercetările în teren au fost începute în această regiune, în special în RezervaŃia Naturală de Stepă Dealul Zakel (la Slimnic), Dealul Mocanu (la GuşteriŃa), Dealul Ritoi (la Slimnic).
În Baden-Württemberg, una din regiunile cele mai bogate în specii de Orobanche este Bergstraße, lângă Hemsbach, în care sunt prezente 17 specii (Röhner & Schwöbel, 2006). De aceea, staŃiunea Hemsbach (la Weinheim), Gemarkung Alteberg, a prezentat şi pentru noi un interes deosebit.
Au avut şi au încă loc numeroase iniŃiative de cercetare a grupului Orobanche, însă, speciile din flora spontană, dintre care unele sunt chiar ameninŃate cu dispariŃia, nu au stat în centrul acestora, cercetările fiind centrate exculsiv pe cele care parazitează speciile de plante cultivate.

2. OBIECTIVE DE CERCETARE Cercetările ecologice şi fiziologice ale speciilor rare de Orobanche în flora
spontană a României, în comparaŃie cu cele care cresc în regiunea Baden-Württemberg (sudul Germaniei) au ca scop aflarea cauzelor pentru care aceste specii devin tot mai rare, unele fiind chiar ameninŃate cu dispariŃia. Studiul condiŃiilor ecologice ale fiecărei staŃiuni urmăreşte verificarea următoarelor ipoteze:
• Schimbarea condiŃiilor climatice. • Producerea insuficientă de seminŃe periclitează menŃinerea speciei. • Polenizarea insuficientă, cauzată de dimensiunea foarte mică şi izolarea
populaŃiilor, având în vedere faptul că populaŃiile mici nu sunt interesante pentru insecte şi pentru că lipseşte schimbul de polen prin polenizarea încrucişată.
• Ovulele sunt consumate de insecte. • Ciupercile patogene pot distruge plantele. • Sensibilitatea redusă a seminŃelor, faŃă de stimulatorii de germinaŃie, scade
rata germinaŃiei. • SeminŃele de Orobanche sau seminŃele plantei gazdă conŃin inhibitori de
germinaŃie, care blochează declanşarea germinaŃiei, chiar dacă stimulatorii de germinaŃie sunt prezenŃi.
• Efectele alelopatice ale exsudatelor radiculare de la plantele însoŃitoare inhibă germinaŃia şi dezvoltarea la Orobanche.
Un rol deosebit îl are studiul climei şi vremii, pentru că seminŃele de Orobanche necesită condiŃii speciale, mai ales în timpul germinaŃiei (10 – 15 zile în sol umed şi cald pentru faza de condiŃionare, continuate cu 10 zile în sol umed şi temperatura de 20 °C, pentru germinaŃia propriu-zisă). Se urmăreşte, de asemenea, investigarea specificităŃii speciilor de Orobanche faŃă de gazdă.
Rezultatele aşteptate nu numai că vor lărgi cunoştinŃele despre speciile rare de Orobanche, ci, pe de o parte vor conduce la dezvoltarea unor metode pentru păstrarea speciilor rare, iar pe de altă parte, la identificarea unor căi noi de combatere a dăunătorilor în agricultură.
3. MATERIAL ŞI METODE
SeminŃele speciilor de Orobanche şi ale speciilor gazdă au fost procurate prin
intermediul Grădinii Botanice Universitare Macea, de la Grădina Botanică a UniversităŃii din Iaşi, de la alte instituŃii similare, ori prin recoltare directă din teren. InmulŃirea vegetativă necesară pentru unele specii gazdă s-a relizat la serele UniversităŃii Viena.
Pentru pregătire s-a efectuat studiul ierbarelor următoarelor instituŃii: Grădina Botanică a UniversităŃii “Babeş-Bolyai” din Cluj-Napoca, Muzeul “Brukenthal” Sibiu, Grădina Botanică Macea a UniversităŃii „Vasile Goldiş” din Arad şi colecŃia de preparate lichide a Muzeului Grădinii Botanice a UniversităŃii Bucureşti, unde speciile de Orobanche sunt conservate împreună cu gazdele lor în alcool.

Determinarea speciilor de Orobanche s-a efectuat pe baza caracterelor morfologice: forma de creştere, forma florii, dimensiunea şi alcătuirea ei, culoarea corolei (caracter nesigur, pentru că poate fi foarte variabil uneori), culoarea stigmatului, fidelitatea faŃă de planta gazdă (în măsura în care determinarea ei este sigură). Cea mai recentă monografie a speciilor de Orobanche din Europa (Uhlich ş. a., 1995) a fost, în acest sens, de mare ajutor.
Cercetare ecologică comparativă a habitatelor studiate în flora spontană din România şi Baden-Württemberg a avut în vedere datele din literatura de specialitate şi informaŃiile primite de la colegi botanişti, pe baza cărora au fost căutate staŃiuni ale speciilor de Orobanche în flora spontană. În Baden-Württemberg (sudul Germaniei) au fost utilizate pliantele Muzeului de ŞtiinŃe ale Naturii din Stuttgart, precum şi indicaŃile cercetătorilor care au realizat cartarea pentru flora din această zonă.
În fiecare staŃiune s-au desfăşurat următoarele activităŃi: - determinarea speciilor de Orobanche şi a stadiului lor de dezvoltare,
efectuându-se totodată şi fotografii; - în cazul în care fructele erau mature, s-au recoltat seminŃe pentru
experimente de laborator; - conspectarea plantelor însoŃitoare pe suprafaŃa de 1 mp în jurul speciei de
Orobanche luată în atenŃie; notarea specilor însoŃitoare este importantă nu doar în scopul determinării plantei gazdă, ci pentru a avea informaŃii despre posibilele reacŃii alelopatice, ca de exemplu inhibarea germinaŃiei seminŃelor de Orobanche prin exsudatul radicular al plantelor învecinate;
- descrierea topografiei staŃiunii investigate; - determinarea coordonatelor geografice cu Global Positioning System
(GPS), System Etrex Vista C; acest aparat are o marjă de siguranŃă de 0,1" (≅ 3 m); - recoltarea probelor de sol pentru determinarea de pH şi analiza conŃinutului
de nitrat, amoniu, fosfat, carbon organic; - în măsura în care a fost posibil, s-a încercat procurarea datelor referitoare la
climă şi la vreme pentru ultimii ani (în special privind temperatura şi precipitaŃiile). -încadrarea staŃiunilor cercetate în zonele climatice corespunzătoare pe baza
clasificării Köppen (Strahler& Strahler, 2005). În formule, prima literă denumeşte zona de climă, a doua tipul, iar următoarele, subtipul de climă. Diagrama climatică a fiecărei staŃiuni a fost întocmită pe baza datelor furnizate de cea mai apropiată staŃie meteorologică.
Experimente de laborator. Investigarea seminŃelor de Orobanche prin microscopia electronică de scanare (SEM) s-au efectuat pentru motivul că unii autori consideră morfologia suprafeŃei şi microstructura tegumentului seminal ca un criteriu taxonomic important. Datorită faptului că structura suprafeŃei seminŃelor din sol se păstrează neschimbată pentru mulŃi ani, analiza SEM ar putea să contribuie la determinarea seminŃelor de Orobanche din sol.
SeminŃelor de Orobanche au fost investigate prin intermediul microscopului optic şi al microscopului electronic cu transmisie (TEM) Pentru studiile de microscopie electronică, seminŃele de Orobanche sunt fixate cu glutaraldehidă în

tampon cacodilat, postfixate cu tetraoxid de osmiu, sunt contrastate cu acetat de uranil, deshidratate cu acetonă, apoi incluse în răşină epoxidică sintetică. Preparatele sunt secŃionate la microtom şi observate la microscopul electronic de transmisie. Vizualizarea specimenelor se desfăşoară cu aparatul EM 109 de la Zeiss.
Pentru identificare ciupercilor endofite pe baza analizei secvenŃelor genice, ADN-ul ciupercilor se derulează cu ajutorul kitului DNeasy® Plant Mini Kit (Qiagen, Hilden). Prin intermediul reacŃiei în lanŃ a polimerazei PCR (Polymerase Chain Reaction), moleculele de ADN se înmulŃesc exponenŃial, se amplifică. La final ele sunt curăŃate şi separate prin electroforeză în gel. SecvenŃele de gene sunt apoi comparate cu datele din banca de gene.
Experimentele de germinaŃie s-au derulat în două serii independente: o serie simplă, de germinaŃie a seminŃelor de Orobanche în prezenŃa exsudatului radicular specific, respectiv a stimulentului sintetic GR 24, urmată de o serie mai complexă, în cadrul căreia s-a urmărit şi efectul altor substanŃe asupra acestui proces. Ca potenŃiale substanŃe active s-au introdus câte 1 mM de acid cinamic, acid ferulic, acid siringic, ca potenŃiali inhibitori de germinaŃie. Pe baza unor observaŃii, seria de germinaŃii a verificat şi acŃiunea următorilor compuşi:
• acidul benzoic este prezent în toate exsudatele radiculare analizate în această lucrare;
• metionina, după Vurro (2004), cauzează scurtarea radiculei şi prin aceasta, inhibă germinaŃia;
• acidul giberelinic, ca fitohormon, stimulează germinaŃia; el cauzează creşterea prin alungire a celulelor şi, probabil, poate să anuleze efectul de scurtare a radiculei, indus de metionină.
Analiza instrumentală a exsudatelor radiculare a necesitat separarea exsudatului radicular într-o fracŃiune hidrofilă şi o fracŃiune hidrofobă se efectuează cu coloana de Amberlite XAD 1180 (după WikEcology, după colecŃia de metode a Departamentului de Cercetarea Ecosistemelor, Universitatea Viena). Sunt prelucrate exsudate radiculare de la plantele gazdă: Artemisia campestris, Berberis vulgaris, Hedera helix, Carduus personata, Centaurea scabiosa, Cirsium vulgare, Galium verum, Genista lydia, Lycopersicon esculentum, Medicago sativa, Pelargonium zonale Rubus idaeus, Salvia aethiopis, Teucrium chamaedrys, Thymus pannonicus, Urtica dioica, precum şi de la plante însoŃitoare.
Pentru analiza GC-MS, concentraŃia probelor trebuie să fie de aproximativ 3 mg/ml. De aceea, este necesar ca sililarea să se efectueza în microflacoane. Extractele concentrate se dizolvă în 200 µl amestec de metanol-cloroform-apă, în proporŃie de 1:1:1 (v), transferate în microflacoane şi uscate din nou la SpeedVac. Urmează oximarea grupărilor aldehidice, de exemplu a glucidelor (Gullberg ş. a., 2004), timp de 17 h, cu 30 µl soluŃie 20 mg/l metoxiamină (CH3-O-NH2) în piridină, apoi timp de 1 h, trimetilsililarea cu 30 µl MSTFA, N-metil-N-(trimetilsilil)-trifluoro-acetamid (Fiehn ş. a., 2000).
Analizele GC-MS s-au efectuat cu sistemul Perkin Elmer Turbomass Autosystem XL cu cromatograf pentru gaze (vezi parametrii în tab. 3), dotat cu o

coloană capilară pentru separare de 20 m x 0,18 mm, cu strat de separare de 0,18 µm.
Derivatizarea fracŃiunii hidrofobe (fracŃiunea II) pentru cromatografia de lichid de înaltă performanŃă (High Performance Liquid Chromatography, HPLC ) impune ca eluatele de coloană cu etanol să se evapore în Rotavapor până la uscare. În continuare se dizolvă în metanol (10 mg/ml) în care s-a adăugat o picătură de acid fosforic. Pentru dizolvare, sticluŃele se aşează pentru 15 min în baia de ultrasunete. 10 µl din conŃinut vor fi injectate cu ajutorul unui autosampler în sistemul Dionex HPLC.
Separarea probelor se desfăşoară în aparatul Dionex HPLC, dotat cu un schimbător pentru probe Famos, pompă HPLC P680 şi un detector UVD 340U-Photodioden-Array cu sistem de analiză a datelor. Cromatogramele se înregistrează la 229 nm şi absorbŃia se scanează la 220 – 590 nm. Separarea se derulează cu o coloană Synergi 4µ Max RP 80A (150 x 2 mm, Phenomenex).
Analiza exsudatelor radiculare privind strigolactonele s-a realizat prin cromatografia de lichid de înaltă performanŃă, cu spectroscopia de masă în tandem, HPLC-MS/MS . Exsudatul radicular se filtrează. Fiecare 100 ml se agită de 2 ori cu câte 400 ml acetat de etil în pâlnia de separare. Câte 400 ml din extractul în acetat de etil se spală cu câte 100 ml 0,2 M soluŃie apoasă de Na2HPO4 iar la final, se usucă cu MgSO4 anhidric. Etil acetatul se redistilează la rotavapor. Pentru a preveni impurificarea cu resturi de MgSO4, extractele se filtrează într-un balon de rotaŃie. Probele se concentrează până la uscare în recipiente mici de sticlă şi se sigilează. A fost utilizat BÜCHI ROTAVAPOR R-124 cu BÜCHI WATERBATH B-480. Parametrii aparatului: vacuum -0,95 Torr, temperatura de răcire 10°C, baia de apă 30°C.Probele se dizolvă în metanol 60 %, se filtrează şi se injectează într-o coloană fază-rezervă (C18) HPLC, care este conectată cu tandemul MS/MS. Strigolactonele se măsoară prin reacŃia multiplă de monitorizare (MRM).
4. REZULTATE
StaŃiuni cercetate. În această lucrare sunt cercetate 17 specii de Orobanche, în 26 de staŃiuni (13 în România şi 13 în Baden-Württemberg). De asemenea, pe baza datelor din literatură, au mai fost verificate 18 staŃiuni, în care însă, prezenŃa speciilor de Orobanche nu a putut fi confirmată.
StaŃiunile cercetate în România sunt răspândite în special în zone deluroase şi colinare (fig. 20). Localizarea lor a fost destul de dificilă, pentru că, în general, populaŃiile de Orobanche sunt mici, iar datele din literatură au caracter general.Ca şi staŃiunile cercetate în România, cele din Baden-Württemberg sunt situate în regiuni deluroase (fig.67). Ele au fost identificate mai uşor, dat fiind faptul că în Baden-Württemberg aproape toate speciile de Orobanche se află pe Lista Roşie, iar populaŃiile sunt cartografiate.Se constată atât scăderea numărului de indivizi în populaŃii cât şi a numărului de staŃiuni, chiar şi atunci când staŃiunile sunt rezervaŃii naturale, în pofida faptului că uşoara încălzire a vremii în ultimii ani ar trebui să favorizeze aceste specii.
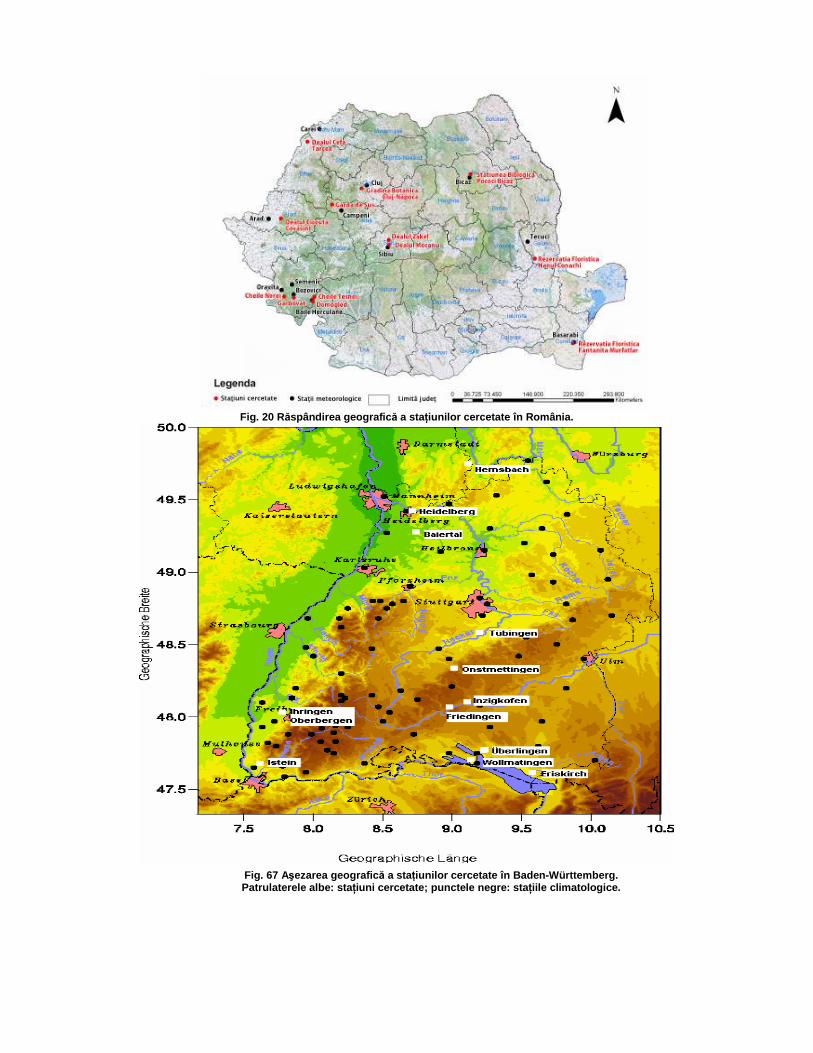
Fig. 20 Răspândirea geografic ă a sta Ńiunilor cercetate în România.
Fig. 67 Aşezarea geografic ă a sta Ńiunilor cercetate în Baden -Württemberg.
Patrulaterele albe: sta Ńiuni cercetate; punctele negre: sta Ńiile climatologice.
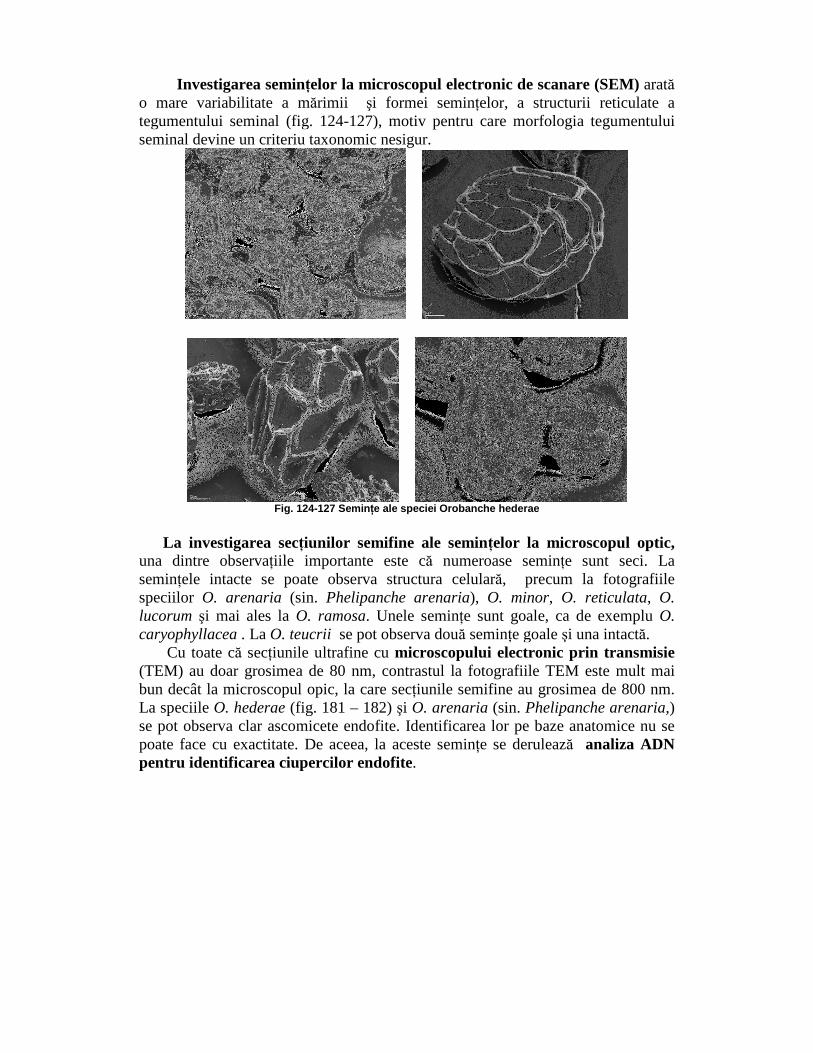
Investigarea seminŃelor la microscopul electronic de scanare (SEM) arată o mare variabilitate a mărimii şi formei seminŃelor, a structurii reticulate a tegumentului seminal (fig. 124-127), motiv pentru care morfologia tegumentului seminal devine un criteriu taxonomic nesigur.
Fig. 124-127 Semin Ńe ale speciei Orobanche hederae
La investigarea secŃiunilor semifine ale seminŃelor la microscopul optic,
una dintre observaŃiile importante este că numeroase seminŃe sunt seci. La seminŃele intacte se poate observa structura celulară, precum la fotografiile speciilor O. arenaria (sin. Phelipanche arenaria), O. minor, O. reticulata, O. lucorum şi mai ales la O. ramosa. Unele seminŃe sunt goale, ca de exemplu O. caryophyllacea . La O. teucrii se pot observa două seminŃe goale şi una intactă.
Cu toate că secŃiunile ultrafine cu microscopului electronic prin transmisie (TEM) au doar grosimea de 80 nm, contrastul la fotografiile TEM este mult mai bun decât la microscopul opic, la care secŃiunile semifine au grosimea de 800 nm. La speciile O. hederae (fig. 181 – 182) şi O. arenaria (sin. Phelipanche arenaria,) se pot observa clar ascomicete endofite. Identificarea lor pe baze anatomice nu se poate face cu exactitate. De aceea, la aceste seminŃe se derulează analiza ADN pentru identificarea ciupercilor endofite.
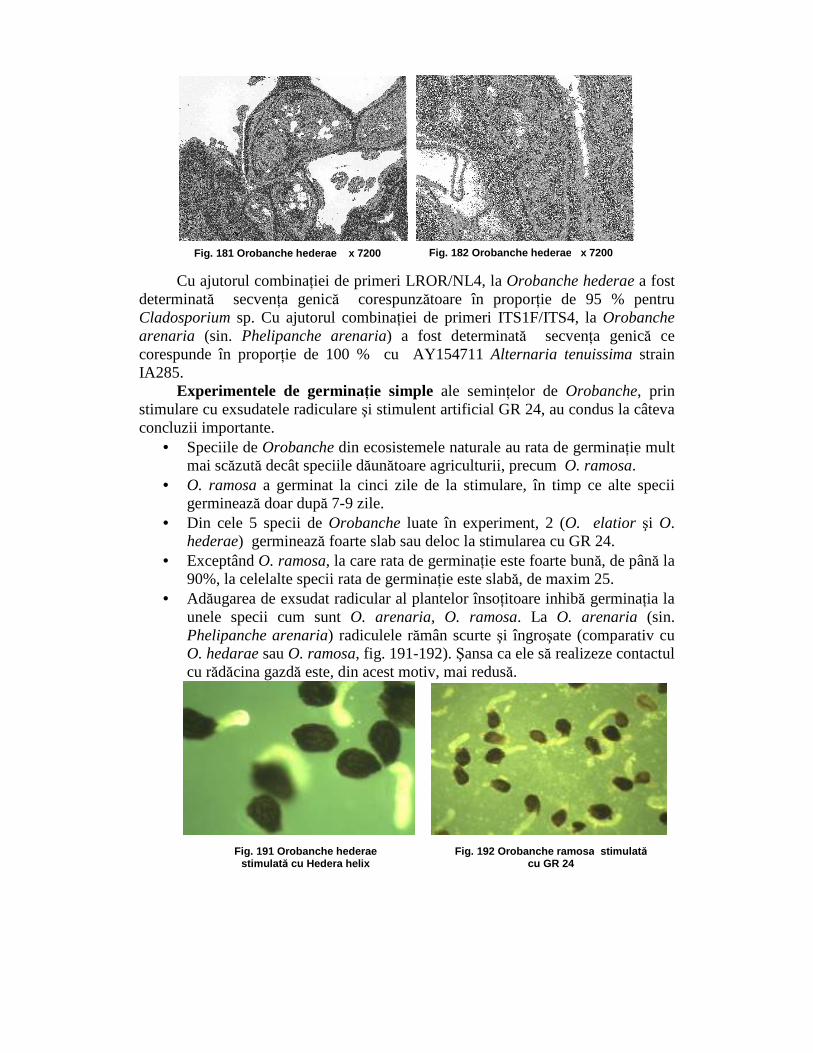
Cu ajutorul combinaŃiei de primeri LROR/NL4, la Orobanche hederae a fost determinată secvenŃa genică corespunzătoare în proporŃie de 95 % pentru Cladosporium sp. Cu ajutorul combinaŃiei de primeri ITS1F/ITS4, la Orobanche arenaria (sin. Phelipanche arenaria) a fost determinată secvenŃa genică ce corespunde în proporŃie de 100 % cu AY154711 Alternaria tenuissima strain IA285.
Experimentele de germinaŃie simple ale seminŃelor de Orobanche, prin stimulare cu exsudatele radiculare şi stimulent artificial GR 24, au condus la câteva concluzii importante.
• Speciile de Orobanche din ecosistemele naturale au rata de germinaŃie mult mai scăzută decât speciile dăunătoare agriculturii, precum O. ramosa.
• O. ramosa a germinat la cinci zile de la stimulare, în timp ce alte specii germinează doar după 7-9 zile.
• Din cele 5 specii de Orobanche luate în experiment, 2 (O. elatior şi O. hederae) germinează foarte slab sau deloc la stimularea cu GR 24.
• Exceptând O. ramosa, la care rata de germinaŃie este foarte bună, de până la 90%, la celelalte specii rata de germinaŃie este slabă, de maxim 25.
• Adăugarea de exsudat radicular al plantelor însoŃitoare inhibă germinaŃia la unele specii cum sunt O. arenaria, O. ramosa. La O. arenaria (sin. Phelipanche arenaria) radiculele rămân scurte şi îngroşate (comparativ cu O. hedarae sau O. ramosa, fig. 191-192). Şansa ca ele să realizeze contactul cu rădăcina gazdă este, din acest motiv, mai redusă.
Fig. 181 Orobanche hederae x 7200 Fig. 182 Orobanche hederae x 7200
Fig. 191 Orobanche hederae stimulat ă cu Hedera helix
Fig. 192 Orobanche ramosa stimulat ă cu GR 24
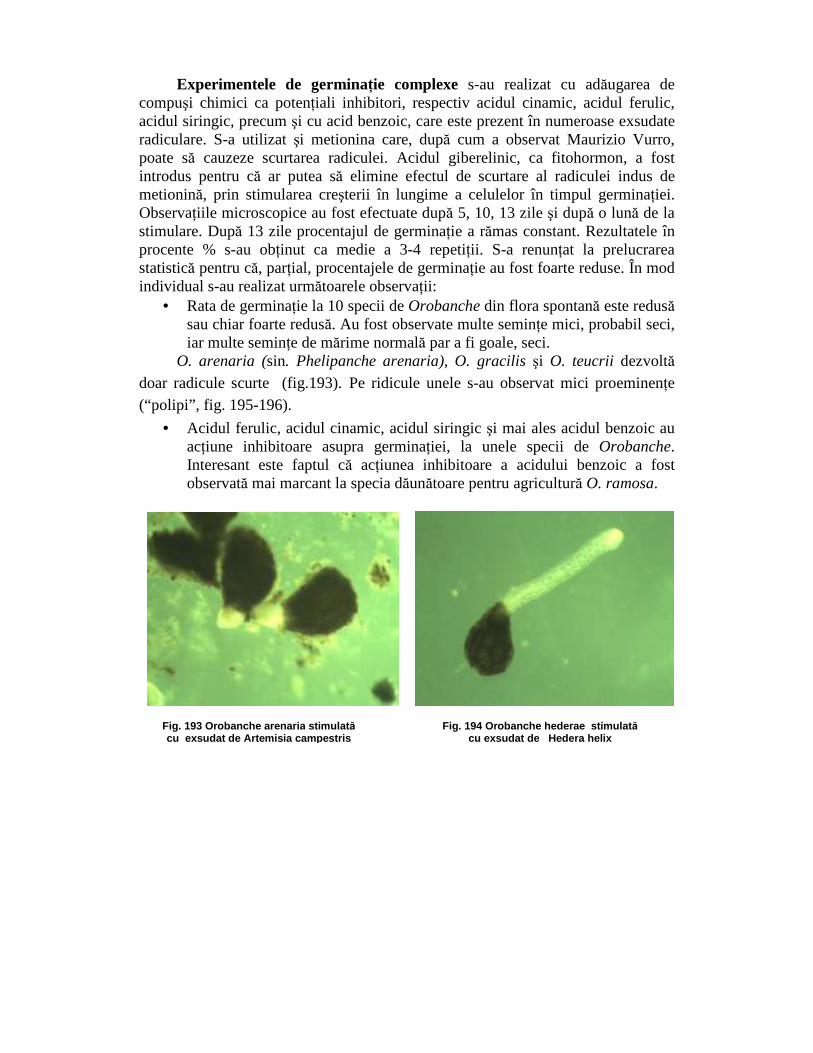
Experimentele de germinaŃie complexe s-au realizat cu adăugarea de compuşi chimici ca potenŃiali inhibitori, respectiv acidul cinamic, acidul ferulic, acidul siringic, precum şi cu acid benzoic, care este prezent în numeroase exsudate radiculare. S-a utilizat şi metionina care, după cum a observat Maurizio Vurro, poate să cauzeze scurtarea radiculei. Acidul giberelinic, ca fitohormon, a fost introdus pentru că ar putea să elimine efectul de scurtare al radiculei indus de metionină, prin stimularea creşterii în lungime a celulelor în timpul germinaŃiei. ObservaŃiile microscopice au fost efectuate după 5, 10, 13 zile şi după o lună de la stimulare. După 13 zile procentajul de germinaŃie a rămas constant. Rezultatele în procente % s-au obŃinut ca medie a 3-4 repetiŃii. S-a renunŃat la prelucrarea statistică pentru că, parŃial, procentajele de germinaŃie au fost foarte reduse. În mod individual s-au realizat următoarele observaŃii:
• Rata de germinaŃie la 10 specii de Orobanche din flora spontană este redusă sau chiar foarte redusă. Au fost observate multe seminŃe mici, probabil seci, iar multe seminŃe de mărime normală par a fi goale, seci.
O. arenaria (sin. Phelipanche arenaria), O. gracilis şi O. teucrii dezvoltă doar radicule scurte (fig.193). Pe ridicule unele s-au observat mici proeminenŃe (“polipi”, fig. 195-196).
• Acidul ferulic, acidul cinamic, acidul siringic şi mai ales acidul benzoic au acŃiune inhibitoare asupra germinaŃiei, la unele specii de Orobanche. Interesant este faptul că acŃiunea inhibitoare a acidului benzoic a fost observată mai marcant la specia dăunătoare pentru agricultură O. ramosa.
Fig. 193 Orobanche arenaria stimulat ă cu exsudat de Artemisia campestris
Fig. 194 Orobanche hederae stimul ată cu exsudat de Hedera helix

Cu toate că seminŃele de Orobanche au fost sterilizate cu mare atenŃie, deseori s-a observat că din micropilul unor seminŃe cresc hife de ciupercă, din acelaşi punct din care radicula iese din sămânŃă. Ca urmarea acestor observaŃii, s-au efectuat ulterior fotografii ale secŃiunilor fine la microscopul electronic de transmisie (TEM). Astfel s-a constatat prezenŃa unor ciuperci endofite, pentru care s-a derulat, de asemenea, identificarea prin metode de genetică moleculară.
Rezultatele analizei HPLC. Dacă spectrele identificate corespund cel puŃin în proporŃie de 95 % cu cele din banca de date, se tipăreşte o propunere de identificare. Identificarea este sigură doar dacă factorul de retenŃie la separarea HPLC corespunde spectrului de absorbŃie. Acidul ferulic şi acidul cumarinic au fost cromatografiaŃi în comparaŃie cu substanŃa pură.
Acidul benzoic a fost identificat cu siguranŃă la Artemisia campestris, Berberis vulgaris, Carduus personata, Centaurea scabiosa, Cirsium vulgare, Galium verum, Hedera helix, Pelargonium zonale, Teucrium chamaedrys, Thymus pannonicus şi la plantele însoŃitoare.
Prin analizele GC-MS a fost separată fracŃiunea hidrofilă a exsudatelor radiculare. Comuşii chimici prezenŃi au fost identificaŃi pe baza spectrelor de masă. Se constată prezenŃa glucidelor şi a poliolilor, precum şi a acizilor organici, ce pot constitui o sursă de hrană pentru microorganisme în sol.
Pentru analiza exsudatelor radiculare pentru identificarea strigolactonelor preparatele au fost prelucrate în Departamentul de Chimie Ecologică şi Cercetarea Ecosistemelor a UniversităŃii Viena, iar analizele au fost desfăşurate la Centrul de Cercetări al UniversităŃii Utsunomiya din Japonia (în colaborare cu prof. Koichi Yoneyama).
Pe abscisă este reprezentat timpul de retenŃie al HPLC în minute. Pe ordonată sunt marcate semnalele pentru fiecare din cele 7 intervale de masă căutate în cadrul reacŃiei multiple de monitorizare (Multiple Reaction Monitoring, MRM), caracteristică pentru compuşii din grupa strigolactonelor. Intervalele de masă sunt consemnate fiecare şi în partea dreaptă, sus. În lucrare sunt prezentate doar intervalele de masă corespunzătoare strigolactonelor prezente.
Pentru că orobanholul şi sorgomolul nu se separă pe coloana C18 (ODS) s-au realizat cromatograme de control cu o coloană fenil. Separarea a arătat că în toate preparatele este prezent orobanholul, dar sorgomolul este absent.
Fig. 195 Orobanche gracilis stimulat ă cu exsudat de Dorycnium herbaceum
Fig. 196 Orobanche caryophyllacea stimulat ă cu exsudat de Galium verum
„Polipi“
„Polipi“

Fig. 230 Strigolactonele din exsudatul radicular de Carduus personata, planta gazd ă pentru Orobanche reticulata
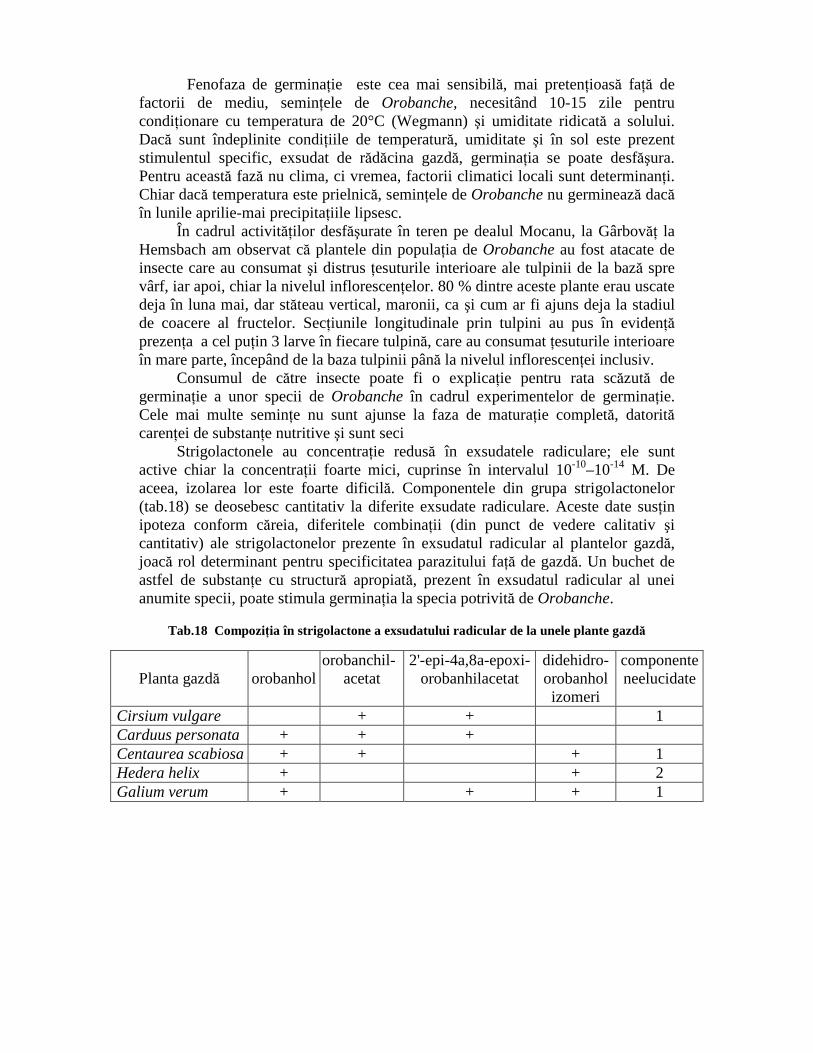
Amestecul de strigolactone prezente în diferite exsudate radiculare se deosebeşte cantitativ. Componentele necunoscute sunt strigolactone, a căror greutate moleculară nu este elucidată în detaliu până la această dată. Aceasta este valabil şi pentru isomeri ai didehidro-orobanholului.
5. DISCUłII PRIVIND REZULTATELE
Cu excepŃia speciei Orobanche hederae, care deseori preferă, la fel ca şi
gazda sa, locuri umbroase şi creşte mai ales sub arbori sau în tufişuri, speciile de Orobanche din flora spontană preferă staŃiuni însorite şi calde, deseori cu expoziŃie sudică, dar unele sunt întâlnite şi în pajişti pe suprafeŃe plane, chiar printre ierburi înalte (O. minor, O. coerulescens).
Speciile de Orobanche germinează la întuneric. Pentru hrănirea lor nu depind de lumină, ele ne având capacitatea de a face fotosinteză. Nu se cunoaşte exact, dacă lumina este necesară în dezvoltarea lor. În orice caz, la specia O. minor genele fitocromului sunt exprimate (Okazawa ş. a., 2004; Mathews, 2005).
În ecosistemele cercetate, nu s-au remarcat daune importante provocate speciilor gazdă, de către parazitul Orobanche în flora spontană.
După sistemul de clasificare Köppen, toate staŃiunile cercetate în Baden-Württemberg au aceleaşi caracteristici: climat temperat cald şi umed cu veri călduroase sau, ca formulă climatică: Cfb. În România, datorită altitudinii şi expoziŃiei foarte diferite a reliefului, climatele staŃiunilor studiate au nuanŃe clar diferite. Există, 6 staŃiuni Df (climă de pădure, rece cu zăpadă, dar cu veri calde) şi 5 Cf (climă temperată ploioasă) ca şi cele din Baden-Württemberg.
Speciile de Orobanche germinează, înfloresc şi fructifică în perioade diferite. Aceasta s-a constatat în toate staŃiunile în care cresc mai multe specii ale acestui gen. Fiecare este dependentă pentru dezvoltarea ei, de anumită „fereastră ecologică“ îngustă. VariaŃia factorilor climatologici de la un an la altul, este cauza pentru care speciile de Orobanche nu apar anual în anumită staŃiune, iar în staŃiunile cu mai multe speciile de Orobanche, compoziŃia floristică a acestora diferă de la an la an.
2’-epi -4a,8a-epoxi -orobanhilacetat
orobanhilacetat
orobanhol
Fenofaza de germinaŃie este cea mai sensibilă, mai pretenŃioasă faŃă de factorii de mediu, seminŃele de Orobanche, necesitând 10-15 zile pentru condiŃionare cu temperatura de 20°C (Wegmann) şi umiditate ridicată a solului. Dacă sunt îndeplinite condiŃiile de temperatură, umiditate şi în sol este prezent stimulentul specific, exsudat de rădăcina gazdă, germinaŃia se poate desfăşura. Pentru această fază nu clima, ci vremea, factorii climatici locali sunt determinanŃi. Chiar dacă temperatura este prielnică, seminŃele de Orobanche nu germinează dacă în lunile aprilie-mai precipitaŃiile lipsesc.
În cadrul activităŃilor desfăşurate în teren pe dealul Mocanu, la GârbovăŃ la Hemsbach am observat că plantele din populaŃia de Orobanche au fost atacate de insecte care au consumat şi distrus Ńesuturile interioare ale tulpinii de la bază spre vârf, iar apoi, chiar la nivelul inflorescenŃelor. 80 % dintre aceste plante erau uscate deja în luna mai, dar stăteau vertical, maronii, ca şi cum ar fi ajuns deja la stadiul de coacere al fructelor. SecŃiunile longitudinale prin tulpini au pus în evidenŃă prezenŃa a cel puŃin 3 larve în fiecare tulpină, care au consumat Ńesuturile interioare în mare parte, începând de la baza tulpinii până la nivelul inflorescenŃei inclusiv.
Consumul de către insecte poate fi o explicaŃie pentru rata scăzută de germinaŃie a unor specii de Orobanche în cadrul experimentelor de germinaŃie. Cele mai multe seminŃe nu sunt ajunse la faza de maturaŃie completă, datorită carenŃei de substanŃe nutritive şi sunt seci
Strigolactonele au concentraŃie redusă în exsudatele radiculare; ele sunt active chiar la concentraŃii foarte mici, cuprinse în intervalul 10-10–10-14 M. De aceea, izolarea lor este foarte dificilă. Componentele din grupa strigolactonelor (tab.18) se deosebesc cantitativ la diferite exsudate radiculare. Aceste date susŃin ipoteza conform căreia, diferitele combinaŃii (din punct de vedere calitativ şi cantitativ) ale strigolactonelor prezente în exsudatul radicular al plantelor gazdă, joacă rol determinant pentru specificitatea parazitului faŃă de gazdă. Un buchet de astfel de substanŃe cu structură apropiată, prezent în exsudatul radicular al unei anumite specii, poate stimula germinaŃia la specia potrivită de Orobanche.
Tab.18 CompoziŃia în strigolactone a exsudatului radicular de la unele plante gazdă
Planta gazdă
orobanhol
orobanchil- acetat
2'-epi-4a,8a-epoxi- orobanhilacetat
didehidro- orobanhol izomeri
componente neelucidate
Cirsium vulgare + + 1 Carduus personata + + + Centaurea scabiosa + + + 1 Hedera helix + + 2 Galium verum + + + 1

Caracteristici fiziologice ale speciilor de Orobanche din flora spontană, comparativ cu speciile dăunătoare agriculturii :
• DependenŃa strânsă de condiŃiile climatice. Biotopurile naturale (inclusiv rezervaŃiile naturale) nu sunt aprovizionate cu apă în mod artificial;
• SeminŃe puŃine, pentru că inflorescenŃele sunt mai mici şi au flori mai puŃine decât la speciile dăunătoare agriculturii;
• PuŃine seminŃe mature datorită atacului unor insecte. Rata de germinaŃie este o dovadă a numărului mic de seminŃe intacte şi mature.
• Radicule (tuburi germinative) scurte, cu posibilitate foarte redusă de a atinge rădăcina gazdă;
• Specificitate mai redusă faŃă de gazdă, comparativ cu speciile dăunătoare; • PrezenŃa inhibitorilor de germinaŃie în rizosferă (acidul benzoic); • Prin exsudatul radicular al plantelor însoŃitoare poate interveni declanşarea
germinaŃiei suicide; în acest sens lucrarea prezentă aduce unele argumente. Prin germinaŃia suicidă potenŃialul de seminŃe din sol se reduce, iar germinaŃia în masă, la momentul în care condiŃiile de mediu sunt optime (în anii favorabili), nu mai poate fi realizată
6. CONCLUZII ŞI CONTRIBU łII
Cercetările au vizat populaŃiile genului Orobanche ce se dezvoltă în
ecosisteme naturale din România şi Baden-Württemberg (sudul Germaniei), zone asemănătoare din punct de vedere al condiŃiilor fizico-climatice şi a biodiversităŃii acestui gen.
Se poate constata un regres natural, datorită condiŃiilor staŃionale, atât în privinŃa numărului de staŃiuni, cât şi a numărului de exemplare din populaŃiile de Orobanche ale florei spontane. La 70 % din populaŃiile observate, numărul de exemplare este sub 30.
Cercetarea în România a fost focalizată pe 13 din cele 22 specii de Orobanche care cresc aici: O. alba, O. arenaria, O. caryophyllacea, O. coerulescens, O. elatior, O. gracilis, O. lucorum, O. lutea, O. minor, O. purpurea, O. reticulata, O. salviae şi O. teucrii. Dintre acestea, 5 specii se află pe Lista Roşie a plantelor : O. caesia, O. salviae, O lucorum, O. aegyptiaca, O. alsatica. Pentru 6 specii, considerate a avea diferite grade de periclitare, ar fi necesară inculderea lor pe Lista Roşie a Plantelor din România, şi anume: O. arenaria Borckhausen, O. purpurea Jaquin, O. lutea Baumgarten, O. elatior Sutton, O. gracilis Smith, O. teucrii Hollandre, O. reticulata Wallroth, O. alba Willdenow. Ar fi necesară, de asemenea, cartografierea populaŃiilor de Orobanche după modelul de lucru din Baden-Württemberg.
În Baden-Württemberg, au fost luate în studiu 11, din cele 21 de specii ale genului Orobanche răspândite aici: O. alsatica, O. arenaria, O. caryophyllacea, O. elatior, O. hederae, O. lutea, O. mayeri, O. minor, O. picridis, O. purpurea şi O. teucrii. Din 21 specii de Orobanche prezente în regiunea Baden-Württemberg, 19 se află pe Lista Roşie a plantelor din această zonă. În RezervaŃia Naturală Zeller

Horn, din Onstmettingen, a fost observată specia endemică, ameninŃată cu dispariŃia, Orobanche mayeri K. Bertsch & F. Bertsch 1948 (O. alsatica var. mayeri Suessenguth & Ronniger 1942), prezentă exclusiv în această staŃiune, reprezentată prin doar 2 exemplare. Regresul populaŃiei este dramatic, cu toate că specia este ocrotită în acestă rezervaŃie naturală.
StaŃiunile cercetate sunt definte foarte exact cu ajutorul coordonatelor geografice determinate prin sistemului GPS. Aceste date nu mai apar, până în present, în literatura de specialitate.
Schimbările climatice joacă un rol secundar şi, în principiu, ar trebui să favorizeze dezvoltarea speciilor de Orobanche. Din datele climatologice colectate şi prelucrate pentru lunile de primăvară din ultimii 4 ani, în 4 staŃiuni, reiese o tendinŃă clară de creştere a volumului precipitaŃiilor şi a temperaturii în lunile martie-mai. Privită în ansamblu, repartiŃia precipitaŃiilor pe luni şi zile este foarte neregulată. În lunile uscate de primăvară nu sunt asigurate valorile optime de condiŃionare şi de germinaŃie, ceea ce face ca populaŃiile de Orobanche să nu apară în fiecare an.
Speciile rare de Orobanche, cum sunt O. salviae, O lucorum, O. alsatica, O. mayeri, O. bartlingii, O. aegyptiaca, O. caesia, O. alba, O. arenaria, O. gracilis au dezavantaje biologice, comparativ cu puŃinele specii dăunătoare agriculturii (O. cumana, O. cernua, O. crenata, O. ramosa, O. minor). Ele au seminŃe mai puŃine, inflorescenŃele lor fiind mult mai mici. Prin atacul insectelor fitofage (observate în zonele Sibiu, GârbovăŃ şi Hemsbach), plantele au pierdut o parte însemnată din seminŃe, pe două căi: pe de o parte prin consumul lor de către insecte, iar pe de altă parte, seminŃele intacte rămân seci, planta ne mai putând să asigure apa şi substanŃele nutritive necesare maturării lor (datorită distrugerii cilindrului central al tulpinii, în urma atacului). Aceasta s-a confirmat prin experimentele de germinaŃie în condiŃii standard de laborator şi prin investigaŃii elecronomicroscopice. Unele specii de Orobanche din flora spontană (O. arenaria, O. gracilis şi O. teucrii) formează doar radicule scurte, cu care au şanse reduse de a stabili contactul cu rădăcina plantei gazdă, pentru formarea haustorului. GerminaŃia lor, cu astfel de radicule scurte, este una de tip suicid.
Cât de profunde sunt implicaŃiile genetice ale schimbului deficitar de gene între aceste populaŃii izolate geografic nu s-a putut stabili. Datorită dezavantajelor biologice, nu se întrevede pericolul ca speciile de Orobanche din flora spontană să devină buruieni dăunătoare agriculturii. Trecerea spre speciile dăunătoare ar putea să fie realizată doar prin mutaŃii adaptative (O. foetida) sau prin hibridizare (O. lavandulacea).
S-au efectuat numeroase experimente de germinaŃie cu exsudatele radiculare ale plantei gazdă corespunzătoare şi sub influenŃa suplimentară a unor inhibitori potenŃiali de germinaŃie. În experimente paralele a fost utilizat GR 24, ca stimulent artificial de germinaŃie. Astfel, s-a constatat că unele specii de Orobanche nu sunt stimulate de GR 24 (O. elatior, O. lutea, O. hederae). Acesta este un aspect important, pentru că în literatură GR 24 este considerat stimulent standard pentru acest tip de germinaŃie. La toate experimentele derulate, specia O.

ramosa, ca specie dăunătoare agriculturii, a fost introdusă pentru studiu comparativ.
Pentru cercetarea efectelor alelopatice ale florei însoŃitoare, s-au efectuat investigaŃii cu ajutorul cromatografiei de lichid de înaltă performanŃă (HPLC) şi prin intermediul cromatografiei gazoase, în tandem cu spectrometria de masă (GC-MS). La jumătate din exsudatele plantelor gazdă analizate şi chiar la cele de la flora însoŃitoare, acidul benzoic a fost prezent în cantităŃi considerabile. Identitatea lui s-a verificat atât prin timpul de retenŃie la separarea HPLC cât şi prin spectrul de absorbŃie. PrezenŃa substanŃelor inhibitoare ale germinaŃiei din familia acidului cinamic nu a putut fi confirmată. Experimentele standardizate de germinaŃie atestă variaŃia gradului de sensibilitate faŃă de substanŃele inhibitoare ale germiaŃiei, în funcŃie de specie .
Chiar mai importantă decât inhibarea germinaŃiei, este germinaŃia suicidă, provocată de flora însoŃitoare. Aproape toate rădăcinile plantelor însoŃitoare exudează strigolactone, care stimulează germinaŃia seminŃelor de Orobanche, dar aceste plante sunt rezistente şi nu pot fi parazitate. GerminaŃia suicidă poate reduce drastic rezerva de seminŃe din sol şi, în consecinŃă, se reduce potenŃialul de seminŃe ce ar putea germina în anii următori, când biotopul va oferii condiŃii optime.
Pe baza observaŃiilor efectuate pe parcursul experimentelor de germinaŃie, conform cărora, prin micropilul unor seminŃe ies hife de ciuperci, s-au derulat investigaŃii de microscopie electronică. După ce au fost remarcate ascomicete endofite, a urmat identificare lor prin analize de genetică moleculară. Au fost determinate Alternaria tenuissima strain IA 285 şi Cladosporium sp. Rolul acestor ciuperci microscopice pentru germinaŃia seminŃelor, ar putea constitui obiectul unor cercetări viitoare.
Strigolactonele au fost mai întâi izolate din exsudatul radicular al unor plante gazdă pentru speciile de Orobanche din flora spontană, prin intermediul cromatografiei de lichid de înaltă performanŃă, cu spectrometria de masă în tandem (HPLC-MS/MS). S-a constatat prezenŃa unor strigolactone cunoscute la specii gazdă noi, care nu au mai fost cercetate, şi s-au stabilit indicii asupra unor strigolactone noi, a căror structură încă nu este complet elucidată. InvestigaŃiile au arătat că exsudatele radiculare conŃin un amestec specific de compuşi din grupa strigolactonelor. Aspectul este foarte important, pentru că susŃine ipoteza conform căreia, specificitatea faŃă de gazdă la Orobanche, este determinată de compoziŃia în strigolactone a exsudatului radicular.
Investigarea suprafeŃei seminŃelor cu ajutorul microscopului electronic de scanare (SEM), pentru a verifica valoarea taxonomică a infrastructurii tegumentului seminal, a confirmat faptul că aceasta nu poate constitui un criteriu taxonomic esenŃial. Deosebirea dintre cele două secŃii, Orobanche şi Trionychon (Phelipanche), este clară, chiar fără a Ńine seama de aspectul tegumentului seminal. Structura tegumentului seminŃelor se conservă în sol pentru o perioadă îndelungată. De aceea, metoda a fost propusă de diferiŃi autori, pentru a identifica seminŃele din sol şi pe acestă bază, pentru a lua măsuri agronomice de prevenŃie (asolamente specifice şi rotaŃia culturilor).

Bibliografie selectivă
1. Beck-Mannagetta G. (1930) Orobanchaceae. In: Engler A. Das Pflanzenreich. Regni vegetabilis conspectus IV, Leipzig, Wilhelm Engelmann Verlag, 1–347.
2. Cristea V., Gafta D. & Pedrotti F. (2004) Fitosociologie. Editura Presa Universitară Clujeană: 5-120.
3. Demuth S., Kleinsteuber A., Lange D., Philippi G., Siegmund S., Voggesberger M. &, Wörz A. (1996) Die Farn- und Blütenpflanzen Baden-Württembergs, Band 5, Verlag Eugen Ulmer, Stuttgart: 361-399.
4. Drăgulescu C. & Bude M.C. (1999) Dinamica vegetaŃiei din RezervaŃia “Dealul Zakel” (Jud. Sibiu). Acta oecologica, vol. 6, nr. 1-2, 1999: 45-52.
5. Jones M. (1989) Taxonomic and ecological studies on the genus Orobanche in the British Isles. PhD Thesis University of Liverpool.
6. Kuijt J. (1969) The Biology of Parasitic Flowering Plants. Berkeley, CA: University of California Press.
7. Parker C. & Riches C.R. (1993) Parasitic Weeds of the World: Biology and Control. Wallingford, Oxon, UK: CAB International: 332.
8. Schneeweiß G.M. (2004) Phylogeny and Evolution of holoparasitic Orobanche (Orobanchaceae). Dissertation Universität Wien.
9. Teryokhin E.S. (1997) Weed Broomrapes: systematics – ontogenesis – biology – evolution. Aufstieg-Verlag, Augsburg, Germany.
10. Toma C., Andronache A. & Gostin I. (2007) Researches Regarding the Histo-Anatomy and Floral Morphogenesis in some Orobanche L. Species. In: Analele ştiinŃifice ale UniversităŃii ”Al. I. Cuza” Iaşi: 11-25.
11. Uhlich H., Pusch J. & Barthel K.-J. (1995) Die Sommerwurzarten Europas. Die neue Brehm-Bücherei Bd. 618. Westarp Wissenschaften, Magdeburg.
12. Yoneyama K., Yoneyama K., Takeuchi Y., Sekimoto H. (2007) Phosphorus deficiency in red clover promotes exudation of orobanchol, the signal for mycorrhizal symbionts and germination stimulant for root parasites. Planta 225: 1031-1038.


















