![[PPT]SINDROMUL KLINEFELTER - Seria 7 - Homeseria7.weebly.com/.../8/5/4085189/sindromul_klinefelter.ppt · Web viewDIFERENTIEREA SI MATURAREA SEXUALA: BAZE GENETICE, ENDOCRINOLOGIE,](https://static.fdocumente.com/doc/165x107/5b0a1e627f8b9a0c4b8b88c3/pptsindromul-klinefelter-seria-7-viewdiferentierea-si-maturarea-sexuala-baze.jpg)
EFECTUL UTILIZĂRII UNOR ANTIOXIDANŢI ÎN MEDIILE DE ... · primului globul polar (McGaughey,...
74
UNIVERSITATEA DE ŞTIINŢE AGRICOLE ŞI MEDICINĂ VETERINARĂ CLUJ-NAPOCA ŞCOALA DOCTORALĂ FACULTATEA DE ZOOTEHNIE ŞI BIOTEHNOLOGII Ing. Miclea Ileana Raluca EFECTUL UTILIZĂRII UNOR ANTIOXIDANŢI ÎN MEDIILE DE MATURARE A OVOCITELOR ŞI DE CULTURĂ A EMBRIONILOR REZUMAT AL TEZEI DE DOCTORAT Conducător ştiinţific Prof. dr. ing. Miclea Vasile Cluj-Napoca 2010
Transcript of EFECTUL UTILIZĂRII UNOR ANTIOXIDANŢI ÎN MEDIILE DE ... · primului globul polar (McGaughey,...

UNIVERSITATEA DE ŞTIINŢE AGRICOLE ŞI MEDICINĂ VETERINARĂ CLUJ-NAPOCA
ŞCOALA DOCTORALĂ FACULTATEA DE ZOOTEHNIE ŞI BIOTEHNOLOGII
Ing. Miclea Ileana Raluca EFECTUL UTILIZĂRII UNOR ANTIOXIDANŢI ÎN MEDIILE DE MATURARE A OVOCITELOR
ŞI DE CULTURĂ A EMBRIONILOR
REZUMAT AL TEZEI DE DOCTORAT Conducător ştiinţific
Prof. dr. ing. Miclea Vasile
Cluj-Napoca 2010

CUPRINS
CAPITOLUL 1. PRODUCEREA EMBRIONILOR SUINI .... 2
1.1. Foliculogeneza ............................................................... 2 1.2. Maturarea ovocitară ....................................................... 2 1.3. Dezvoltarea embrionului preimplantaţional................... 3
CAPITOLUL 2. BIOSINTEZA ŞI ROLUL RADICALILOR LIBERI...................................................................................... 3 CAPITOLUL 3. MECANISME DE APĂRARE ÎMPOTRIVA RADICALILOR LIBERI.......................................................... 4 SCOPUL ŞI OBIECTIVELE CERCETĂRII ........................... 5 CAPITOLUL 4. MATERIALE ŞI METODE.......................... 7
4.1. MATURAREA OVOCITARĂ...................................... 7 4.1.1. Medii utilizate ......................................................... 7 4.1.2. Recoltarea şi maturarea ovocitelor.......................... 8 4.1.3. Evaluarea maturării ovocitare în urma culturii cu antioxidanţi........................................................................ 8
4.2. FECUNDAŢIA IN VITRO ŞI CULTIVAREA EMBRIONILOR................................................................... 9
CAPITOLUL 5. REZULTATE ŞI DISCUŢII........................ 11 5.1. Determinarea influenţei α-tocoferolu-lui, acidului ascorbic şi luteinei asupra maturării ovocitelor .................. 11
5.1.1. Influenţa α-tocoferolului asupra expandării complexelor ovocită-cumulus ......................................... 11 5.1.2. Influenţa acidului ascorbic asupra expandării complexelor ovocită-cumulus ......................................... 13 5.1.3. Influenţa luteinei asupra expandării complexelor ovocită-cumulus .............................................................. 14 5.1.4. Evaluarea viabilităţii şi maturării nucleare a ovocitelor cultivate în mediu cu α-tocoferol................... 15
I

5.1.5. Evaluarea viabilităţii şi maturării nucleare a ovocitelor cultivate în mediu cu acid ascorbic................ 16 5.1.6. Evaluarea viabilităţii şi maturării nucleare a ovocitelor cultivate în mediu cu luteină .......................... 18 5.1.7. Determinarea conţinutului de glutation din ovocitele cultivate în medii de maturare suplimentate cu α-tocoferol....................................................................... 19 5.1.8. Determinarea conţinutului de glutation din ovocitele cultivate în medii de maturare suplimentate cu acid ascorbic ........................................................................... 20 5.1.9. Determinarea conţinutului de glutation din ovocitele cultivate în medii de maturare suplimentate cu luteină... 21
5.2. DETERMINAREA INFLUENŢEI Α-TOCOFEROLU- LUI, ACIDULUI ASCORBIC ŞI LUTEINEI ASUPRA DEZVOLTĂRII IN VITRO A EMBRIONILOR................ 22
5.2.1. Dezvoltarea embrionilor în mediul TALP suplimentat cu α-tocoferol, după 120 ore de la fecundaţie ........................................................................ 23 5.2.2. Dezvoltarea embrionilor în mediul TALP suplimentat cu luteină, după 120 ore de la fecundaţie .... 24 5.2.3. Dezvoltarea embrionilor în mediul NCSU-23 suplimentat cu α-tocoferol, după 48 ore de la fecundaţie ........................................................................ 25 5.2.4. Dezvoltarea embrionilor în mediul NCSU-23 suplimentat cu acid ascorbic, după 48 ore de la fecundaţie ...................................................................... 277 5.2.5. Dezvoltarea embrionilor în mediul NCSU-23 suplimentat cu luteină, după 48 ore de la fecundaţie .... 288 5.2.6. Dezvoltarea embrionilor în mediul NCSU-23 suplimentat cu α-tocoferol, după 120 ore de la fecundaţie ...................................................................... 299
II

5.2.7. Dezvoltarea embrionilor în mediul NCSU-23 suplimentat cu acid ascorbic, după 120 ore de la fecundaţie ...................................................................... 311 5.2.8. Dezvoltarea embrionilor în mediul NCSU-23 suplimentat cu luteină, după 120 ore de la fecundaţie .. 333 5.2.9. Comparaţie între NCSU-23 şi TALP ca medii de bază suplimentate cu antioxidanţi pentru dezvoltarea in vitro a embrionilor ........................................................ 344
CAPITOLUL 6. CONCLUZII.............................................. 355 BIBLIOGRAFIE..................................................................... 38
III

CAPITOLUL 1. PRODUCEREA EMBRIONILOR SUINI
1.1. FOLICULOGENEZA Foliculul este una din principalele componente ale ovarului cu rol în realizarea funcţiei de reproducţie. La formare foliculii se găsesc în stadiul primordial. În continuare, dezvoltarea lor se realizează printr-o serie de procese coordonate pe cale hormonală şi care implică creşterea ovocitei alături de proliferarea şi diferenţierea celulelor care o înconjoară. Aceste transformări asigură trecerea succesivă prin stadiile primar, secundar şi terţiar.
1.2. MATURAREA OVOCITARĂ În faza terţiară a dezvoltării foliculare ovocita suferă transformări în urma cărora dobândeşte competenţa de a participa la fecundaţie şi este considerată matură. Maturarea este definită ca fiind succesiunea de procese biologice prin care ovocita trece de la dezagregarea veziculei germinale şi până la încheirea celei de-a doua diviziuni meiotice şi eliminarea primului globul polar (McGaughey, 1983). Maturarea cuprinde două procese: unul desfăşurat în nucleu iar celălalt la nivelul citoplasmei. Ele se desfăşoară concomitent condiţionând dobândirea capacităţii ovocitei de a fi fecundată. În timp ce maturarea nucleară poate fi evaluată direct prin colorare fluorescentă aspectul citoplasmatic poate fi determinat numai indirect prin intermediul expandării celulelor cumulus-ooforus sau al determinării concentraţiei de glutation intracelular. Având în vedere conţinutului ridicat de lipide al ovocitelor la suine (160 µg) (McEvoy et al., 2000), prezenţa glutationului are o deosebită importanţă datorită rolului său de antioxidant.
2

1.3. DEZVOLTAREA EMBRIONULUI PREIMPLANTA-ŢIONAL Ovocita eliberată prin ovulaţie şi captată de oviduct este transportată la locul de întâlnire cu spermatozoidul unde va avea loc fecundaţia. Aceasta se desfăşoară după realizarea unor procese complexe la nivelul spermatozoizilor. Zigotul rezultat se va divide mitotic pentru a forma următoarele stadii embrionare, de 2 şi 4-8 celule. La început dezvoltării embrionul foloseşte piruvatul ca substrat metabolic. În stadiul de 4 celule are loc activarea genomului embrionar şi înlocuirea piruvatului cu glucoza. Protecţia sa împotriva stresului oxidativ este asigurată de către glutationul acumulat în timpul maturării în ovocită. Pe măsură ce au loc diviziunile numărul celulelor embrionare creşte şi între acestea se formează joncţiuni. Astfel, la 3-4 zile după ovulaţie (Ladoşi, 1999) embrionul apare ca o masă compactă înconjurată de zona pellucida şi poartă denumirea de morulă. În continuare vor avea loc procesele de compactare şi cavitaţie la finalul cărora rezultă blastocistul. Sub acţiunea enzimelor proteolitice şi a creşterii dimensiunilor embrionare zona pellucida se rupe iar blastocistul eclozează la şase zile după fecundaţie.
CAPITOLUL 2. BIOSINTEZA ŞI ROLUL RADICALILOR LIBERI
Potrivit lui Halliwell şi Gutteridge (2007), radicalii liberi sunt „orice specie de atomi capabilă de o existenţă independentă şi care conţine unul sau mai mulţi electroni neîmperecheaţi”. Cei mai importanţi radicali liberi sunt anionul
3

superoxid (O2·-), hidroxilul (OH·), peroxidul de hidrongen sau
apa oxigenată (H2O2) şi oxidul nitric (NO·). Cea mai mare parte a radicalilor liberi este produsă în mitocondrii unde 1%-2% din totalul electronilor care trec prin lanţul respirator celular se pierd ducând la formarea acestora (Boveris and Chance, 1973). Ei rezultă din activitatea unor enzime (citocrom P450, mono-oxigenazele, NADPH-oxidaza, xantin-oxidaza, ciclooxigenaza, lipooxigenaza) (Kovacic and Jacintho, 2001). In vitro, producerea radicalilor liberi este favorizată şi de lumina fluorescentă, concentraţia crescută a oxigenului şi prezenţa cationilor metalici precum Cu şi Fe. Radicalii liberi funcţionează ca molecule semnal (Hancock et al., 2001) dar în cantitate crescută şi în lipsa unor antioxidanţi adecvaţi produc stresul oxidativ. Acesta determină o serie de reacţii în lanţ care duc la depolarizarea membranei mitocondriale, eliberarea citocromului C, provoacă leziuni la nivelul acizilor nucleici şi oxidează acizii graşi polinesaturaţi ceea ce duce la moartea celulelor (Droge, 2002).
CAPITOLUL 3. MECANISME DE APĂRARE ÎMPOTRIVA RADICALILOR LIBERI
In vivo, ovocita şi embrionul sunt protejaţi împotriva radicalilor liberi de către antioxidanţi prezenţi în fluidul folicular şi oviductal. Antioxidanţii au fost definiţi de către Halliwell şi Gutteridge (2007) ca fiind “orice substanţă, care prezentă în concentraţii mici în comparaţie cu un substrat ce poate fi oxidat este capabilă să întârzie semnificativ sau să împiedice oxidarea acelui substrat”.
4

Ei pot fi de origine endogenă sau exogenă. În prima grupă se înscriu enzimele cu rol antioxidant (superoxid dismutaza, catalaza şi glutation peroxidaza) dar şi substanţe fără funcţie enzimatică precum glutationul, cisteamina şi hipotaurina. Din a doua categorie fac parte substanţe precum α-tocoferolul, acidul ascorbic sau carotenoidele (luteina). Aceştia sunt principalii antioxidanţi prezenţi în sistemele biologice, cu activitate în compartimentul celular liposolubil (α-tocoferolul, luteina) sau în cel hidrosolubil (acidul ascorbic). In vitro activitatea lor a fost testată în cazul a numeroase tipuri celulare, evidenţiindu-se un efect pozitiv mai mult sau mai puţin pronunţat.
SCOPUL ŞI OBIECTIVELE CERCETĂRII Pentru creşterea numărului şi calităţii embrionilor suini obţinuţi in vitro este necesară reducerea sau anularea efectelor negative ale stresului ce influenţează procesele de maturare a ovocitelor şi cultivare a embrionilor. Aceste deziderat se încadrează în contextul mai larg al optimizării protocoalelor de producere a embrionilor in vitro pentru conservarea biodiversităţii şi eficientizarea transferului de embrioni la suine. În acest scop, ne-am propus să studiem măsura în care pot fi îmbunătăţite mediile de maturare a ovocitelor şi cultivare a embrionilor prin adăugarea de antioxidanţi, şi anume α-tocoferol, acid ascorbic şi luteină. Ele sunt substanţe ce se găsesc în mod natural în sistemele biologice, protejându-le împotriva oxidării excesive şi contribuind la menţinerea homeostaziei.
5

În cazul ovocitelor s-a urmărit îmbunătăţirea maturării prin adăugare la mediile de cultură a câte unei substanţe antioxidante în cinci concentraţii specifice. Aprecierea maturării s-a realizat prin metode ce caracterizează aspectul citoplasmatic, cel nuclear dar şi relaţia cu celulele cumulus ooforus. Stabilirea gradului de expandare a celulelor cumulus ooforus s-a constituit ca şi criteriul morfologic. Ea a fost însoţită de colorarea fluorescentă pentru evaluarea viabilităţii, prezenţei leziunilor membranare şi observarea primului globul polar. În scopul aprecierii aspectului citoplasmatic al maturării s-a realizat cuantificarea conţinutului de glutation intraovocitar. Obiectivul următor, a fost stabilirea efectului prezenţei antioxidanţilor asupra dezvoltării morfologice şi fiziologice a embrionilor. În acest scop aceleaşi concentraţii de α-tocoferol, acid ascorbic sau luteină au fost adăugate la două medii de cultură. Evaluarea dezvoltării morfologice s-a făcut prin aprecierea stadiului de dezvoltare şi a calităţii embrionilor. Ea a fost realizată în două etape, adică la 48 şi 120 ore de la fecundaţie. Alegerea acestor momente de evaluare s-a datorat particularităţilor prezentate de embrionii suini cultivaţi in vitro, în special ritmul mai lent de dezvoltare. Astfel s-a putut inventaria atât stadiul de dezvoltare al embrionilor, dar şi evoluţia lor până la 120 ore în funcţie de tipul şi concentraţia substanţei antioxidante adăugate la mediu. La aceste obiective principale s-a adăugat compararea mediilor NCSU-23 şi TALP în vederea stabilirii celui mai potrivit pentru cultivarea embrionilor suini. Cercetările s-au desfăşurat în perioada 2006-2009 în cadrul laboratorului disciplinei Biotehnologia transferului de embrioni şi fecundaţie in vitro, al Facultăţii de Zootehnie şi Biotehnologii a USAMV Cluj-Napoca.
6

CAPITOLUL 4. MATERIALE ŞI METODE
4.1. MATURAREA OVOCITARĂ Pentru recoltarea şi maturarea ovocitelor s-au folosit protocoalele descrise de Grupen et al. (1995) şi Tao et al. (2004) cu modificări proprii.
4.1.1. Medii utilizate Ovarele au fost transportate într-o soluţie salină de 0,9% NaCl în apă bidistilată suplimentată cu penicilină (100 μg/ml) şi streptomicină (100 UI/ml). Pentru recoltare s-a folosit mediu M 199 suplimentat cu L-glutamină (3,4 g/l), bicarbonat de sodiu (2,2 g/l), HEPES (25 mM), penicilină (100 μg/ml) şi streptomicină (100 UI/ml) având pH-ul 7,4. Maturarea s-a realizat în mediu M 199 suplimentat cu L-glutamină (3,4 g/l), Chorulon (10 UI/ml), Folligon (10 UI/ml), ser fetal de viţel 10%, penicilină (100 μg/ml), streptomicină (100 UI/ml). Mediul de maturare astfel preparat a fost împărţit în tuburi tip eppendorf. În fiecare dintre acestea s-a pus un amestec format din α-tocoferol (dizolvat în etanol). α-tocoferolul a fost adăugat astfel încât să se obţină concentraţiile de 5, 10, 20, 40 şi 80 μM substanţă activă. În acelaşi mod, dar în tuburi diferite s-a procedat şi cu acidul ascorbic (dizolvat în apă ultrapură) obţinându-se concentraţii de 50, 150, 250, 500 şi 750 μM substanţă activă. Extractul de luteină (dizolvat în ser fetal bovin) la concentraţii de 2,5, 4, 5, 8 şi 10 μM substanţă activă a fost preparat prin amestec cu mediul de cultură. Mediul pentru denudarea şi spălarea ovocitelor a fost reprezentat de PBS suplimentat cu albumină serică bovină (BSA) (5 mg/ml). Mediul pentru evaluarea viabilităţii prin colorare fluorescentă are la bază tampon fosfat salin, PBS la
7

care s-au adăugat coloranţii 3’6’ diacetat de fluoresceină (FDA) (1 µg/ml), iodură de propidiu (PI) (50 µg/ml) şi Hoechst 33258 (20 µg/ml).
4.1.2. Recoltarea şi maturarea ovocitelor Ovarele recoltate de la femele abatorizate au fost transportate în laborator într-un interval de 2-3 ore în soluţie salină la 37oC. Recoltarea ovocitelor foliculare s-a efectuat prin tehnica puncţiei aspiratorii cu ajutorul unor seringi de 10 ml cu ace G 21. Ovocitele au fost colectat în plăci Petri, conţinând mediu de recoltare. Ovocitele care prezentau citoplasma cu granulaţii uniforme şi cumulus ooforus compact s-au spălat de 2 ori în mediu de recoltare şi transferate în picături de 30 µl mediu de maturare. La acesta au fost adăugate diferite concentraţii de α-tocoferol, acid ascorbic sau luteină. Ovocitele transferate în picături de mediu fără antioxidanţi au reprezentat lotul martor. Independent de lot, fiecare picătură a conţinut un număr de 5 ovocite. Maturarea in vitro s-a realizat într-o atmosferă controlată timp de 44-45 h, la temperatura de 37°C şi un amestec gazos de 5% CO2 în aer.
4.1.3. Evaluarea maturării ovocitare în urma culturii cu antioxidanţi La sfârşitul perioadei de maturare complexele ovocită-cumulus au fost evaluate la microscopul cu inversie. Indicatorul maturării ovocitelor este reprezentat de transformările apărute la nivelul complexelor ovocită cumulus ooforus (COC). Gradul de mucifiere şi expandare a acestor celule a determinat gruparea lor în 5 categorii de calitate notate de la 0 la 4 (Downs, 1989). Ovocitele din fiecare picătură s-au evaluat şi grupat numeric pe grade de expandare.
8

Colorarea fluorescentă s-a realizat prin incubarea ovocitelor timp de 15 min în soluţia de colorare după ce în prealabil ele fuseseră denudate şi spălate. Apoi s-a înregistrat numărul de celule care prezentau fluorescenţă verde sau roşie precum şi cele în al căror spaţiu perivitelin s-a observat primul globul polar colorat în albastru. În final s-a cuantificat cantitatea intraovocitară de glutation prin testului de reciclare al DTNB-GSSG reductazei (Anderson, 1985). În acest scop ovocitele au fost denudate şi congelate la -80°C în momentul analizei. Aceasta a presupus umplerea unui godeu al microplăcii utilizate cu 175 µl soluţie tampon (100 mM fosfat de potasiu, 1 mM EDTA) conţinând NADPH (0,33 mg/ml), 25 µl DTNB (6 mM), 45 µl apă pură şi 2,5 µl probă la care s-au adăugat în final 2,5 µl glutation reductază (250 U/ml). Urmărirea reacţiei enzimatice s-a realizat cu Synergy HT Multi-Mode Microplate Reader (Biotek) iar absorbanţa a fost măsurată la lungimea de undă de 412 nm, la fiecare 50 secunde timp de 10 minute. Concomitent s-au realizat o probă oarbă şi citiri pentru concentraţii cunoscute de GSH (250, 125, 62,5, 31,25, 15,5, 7,75 şi 4 pmoli) care au fost apoi utilizate pentru obţinerera unei curbe standard. În cazul fiecărui experiment s-a realizat comparaţia cu martorul pentru a determina diferenţele. Semnificaţia acestora a fost stabilită cu ajutorul analizei de varianţă şi interpretată prin testul diferenţelor limită (DL) şi testul Duncan (DS).
4.2. FECUNDAŢIA IN VITRO ŞI CULTIVAREA EMBRIONILOR Mediul pentru fecundaţie a fost reprezentat de TALP suplimentat cu penicilină (100 μg/ml) şi streptomicină (100 UI/ml). Embrionii au fost cultivaţi în TALP sau NCSU-23
9

suplimentat cu streptomicină (50 µg /ml), penicilină (75 µg/ml). În primele 48 ore după fecundaţie sursele de energie au fost reprezentate de piruvatul şi lactatul de sodiu la care s-a adăugat glucoza începând cu ziua a treia. Mediul TALP a fost suplimentat cu 5, 10, 20, 40 şi 80 μM α-tocoferol sau 2,5, 4, 5, 8 şi 10 μM luteină. La NCSU-23 sau adăugat aceleaşi concentraţii de α-tocoferolul sau luteină dar şi 50, 150, 250, 500 şi 750 μM acid ascorbic. După evaluarea gradului de expandare al celulelor cumulus, ovocitele au fost denudate şi incubate în mediu TALP alături de spermatozoizii capacitaţi. Pentru capacitare spermatozoizii s-au centrifugat de două ori în mediu Tyrode la 800 g, timp de câte 10 min. Determinarea concentraţiei spermatozoizilor s-a făcut cu ajutorul fotometrului SDM 5 (Minitub). După 16-18 ore de cultură, s-au îndepărtat spermatozoizii iar presupuşii zigoţi au fost cultivaţi în micropicături de mediu TALP sau NCSU-23. La 48 de ore după fecundaţie a fost evaluat stadiul de dezvoltare al embrionilor cuantificându-se numărul de entităţi care au atins stadiile de 2 celule, 4-8 celule şi morulă. În continuare dezvoltarea embrionară a fost monitorizată timp de 72 ore (3 zile de cultură) ajungându-se în final la un total de 120 ore (5 zile) de cultură post fertilizare. După această perioadă s-a determinat numărul de embrioni din fiecare picătură care au ajuns la 2 celule, 4-8 celule sau morulă. Fiecare lot a fost comparat cu martorul pentru a stabili diferenţele şi eventuala semnificaţie a acestora. Semnificaţia diferenţelor a fost determinată cu ajutorul analizei de varianţă şi interpretate prin testul diferenţelor limită (DL) şi prin testul Duncan (DS).
10

CAPITOLUL 5. REZULTATE ŞI DISCUŢII În cadrul experimentelor de maturare ovocitară s-au prelucrat un număr total de 1278 ovare aflate atât în faza foliculinică cât şi în cea luteică a ciclului sexual. Dintre acestea, 564 ovare au fost folosite pentru recoltarea ovocitelor cultivate în mediu suplimentat cu α-tocoferol, obţinându-se o rată de recoltare de 8,85 ovocite/ovar. Pentru recoltarea de ovocite care vor fi maturate în prezenţa acidului ascorbic s-au utilizat 395 ovare, cu o rată de 9,74 ovocite/ovar. În ceea ce priveşte ovocitele cultivate cu mediu ce conţinea luteină, recoltarea lor s-a realizat din 319 ovare, rezultând o rată de 10,86 ovocite/ovar.
5.1. DETERMINAREA INFLUENŢEI Α-TOCOFEROLU-LUI, ACIDULUI ASCORBIC ŞI LUTEINEI ASUPRA MATURĂRII OVOCITELOR
5.1.1. Influenţa α-tocoferolului asupra expandării complexelor ovocită-cumulus Complexele încadrate în categoriile 0 şi 1 nu prezentau expandarea celulelor cumulus ooforus sau aceasta era la un nivel minim în cazul straturilor exterioare. Procentele de COC corespunzătoare gradelor de expandare 0 şi 1 este semnificativ mai redus în cazul mediilor la care s-a adăugat α-tocoferol la concentraţiile de 10, 20, 40 şi 80 µM (tabelul 1). Complexele încadrate în gradul 2 prezintă caracteristic, expandarea şi mucifierea completă a unui segment din cumulus ooforus. El reprezintă situaţia de mijloc dintre ovocitele care sunt sau nu maturate.
11
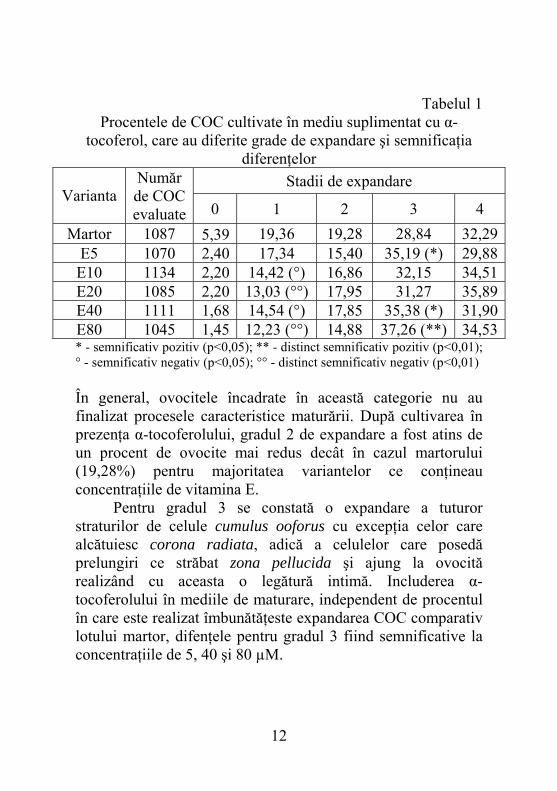
Tabelul 1 Procentele de COC cultivate în mediu suplimentat cu α-
tocoferol, care au diferite grade de expandare şi semnificaţia diferenţelor
Stadii de expandare Varianta
Număr de COC evaluate 0 1 2 3 4
Martor 1087 5,39 19,36 19,28 28,84 32,29 E5 1070 2,40 17,34 15,40 35,19 (*) 29,88 E10 1134 2,20 14,42 (°) 16,86 32,15 34,51 E20 1085 2,20 13,03 (°°) 17,95 31,27 35,89 E40 1111 1,68 14,54 (°) 17,85 35,38 (*) 31,90 E80 1045 1,45 12,23 (°°) 14,88 37,26 (**) 34,53 * - semnificativ pozitiv (p<0,05); ** - distinct semnificativ pozitiv (p<0,01); ° - semnificativ negativ (p<0,05); °° - distinct semnificativ negativ (p<0,01) În general, ovocitele încadrate în această categorie nu au finalizat procesele caracteristice maturării. După cultivarea în prezenţa α-tocoferolului, gradul 2 de expandare a fost atins de un procent de ovocite mai redus decât în cazul martorului (19,28%) pentru majoritatea variantelor ce conţineau concentraţiile de vitamina E. Pentru gradul 3 se constată o expandare a tuturor straturilor de celule cumulus ooforus cu excepţia celor care alcătuiesc corona radiata, adică a celulelor care posedă prelungiri ce străbat zona pellucida şi ajung la ovocită realizând cu aceasta o legătură intimă. Includerea α-tocoferolului în mediile de maturare, independent de procentul în care este realizat îmbunătăţeste expandarea COC comparativ lotului martor, difenţele pentru gradul 3 fiind semnificative la concentraţiile de 5, 40 şi 80 µM.
12
Gradul 4 ce reprezintă expandarea şi mucifierea tuturor straturilor de celule cumulus. El a fost atins de de majoritatea COC cultivate în cadrul tuturor variantelor experimentale fără ca diferenţele comparativ martorului să fie asigurate statistic.
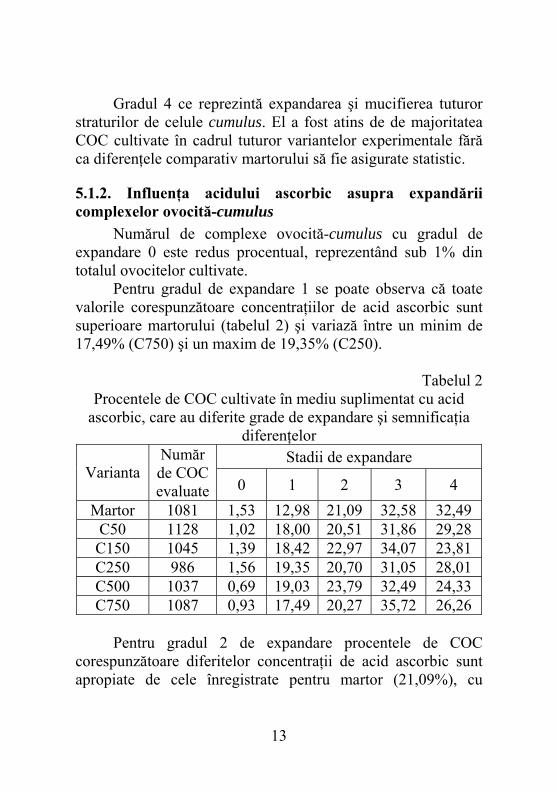
5.1.2. Influenţa acidului ascorbic asupra expandării complexelor ovocită-cumulus Numărul de complexe ovocită-cumulus cu gradul de expandare 0 este redus procentual, reprezentând sub 1% din totalul ovocitelor cultivate. Pentru gradul de expandare 1 se poate observa că toate valorile corespunzătoare concentraţiilor de acid ascorbic sunt superioare martorului (tabelul 2) şi variază între un minim de 17,49% (C750) şi un maxim de 19,35% (C250).
Tabelul 2 Procentele de COC cultivate în mediu suplimentat cu acid
ascorbic, care au diferite grade de expandare şi semnificaţia diferenţelor
Stadii de expandare Varianta
Număr de COC evaluate 0 1 2 3 4
Martor 1081 1,53 12,98 21,09 32,58 32,49 C50 1128 1,02 18,00 20,51 31,86 29,28 C150 1045 1,39 18,42 22,97 34,07 23,81 C250 986 1,56 19,35 20,70 31,05 28,01 C500 1037 0,69 19,03 23,79 32,49 24,33 C750 1087 0,93 17,49 20,27 35,72 26,26
Pentru gradul 2 de expandare procentele de COC corespunzătoare diferitelor concentraţii de acid ascorbic sunt apropiate de cele înregistrate pentru martor (21,09%), cu
13

excepţia lui C150 (22,97%) şi C500 (23,79%). Acelaşi lucru se poate afirma şi despre variantele C150 (34,07%) şi C750 (35,72%) în cazul gradului 3 de expandare. Diferenţele existente nu sunt asigurate statistic. La analiza rezultatelor pentru ultimul grad de expandare se poate observa că valorile corespunzătoare concentraţiilor de acid ascorbic sunt inferioare celei înregistrate pentru martor. Rezultă că în ciuda capacităţii sale antioxidante acidul ascorbic nu a avut un efect benefic asupra expandării, probabil din cauza reducerii viabilităţii celulelor cumulus.
5.1.3. Influenţa luteinei asupra expandării complexelor ovocită-cumulus Pentru gradul 2 de expandare toate valorile caracteristice variantelor experimentale sunt mai reduse decât cele corespunzătoare martorului. Tendinţa descrescătoare nu este corelată cu nivelul antioxidantului. Valoarea înregistrată pentru varianta Lut 8 este semnificativ mai mică decât martorul. Referindu-ne la varianta Lut 8 observîm că, în mod constant are valori mici la gradele de expandare inferioare. Pentru gradul 3 de expandare procentele de COC au variat între 31,71% corespunzător mediului ce conţinea 5 µM luteină şi 36,00% pentru cel cu o concentraţie de 2,5 µM (tabelul 3). Varianta Lut 8 s-a situat în apropierea limitei inferioare cu un procent de COC de 31,75%. Acest fapt este corelat cu dinamica prezentată pentru gradul 2 de expandare la care s-a observat o diferenţă semnificativ mai mică pentru aceeaşi variantă, dar şi cu tendinţa observată în cazul celorlalte grade. Transportul luteinei în celulele cumulus este facilitat de către lipoproteinele prezente în serul fetal bovin.
14
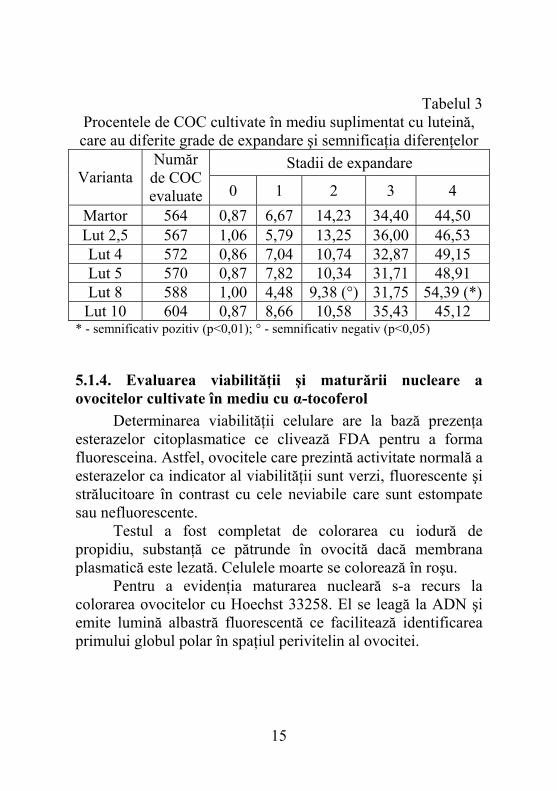
Tabelul 3 Procentele de COC cultivate în mediu suplimentat cu luteină, care au diferite grade de expandare şi semnificaţia diferenţelor
Stadii de expandare Varianta
Număr de COC evaluate 0 1 2 3 4
Martor 564 0,87 6,67 14,23 34,40 44,50 Lut 2,5 567 1,06 5,79 13,25 36,00 46,53 Lut 4 572 0,86 7,04 10,74 32,87 49,15 Lut 5 570 0,87 7,82 10,34 31,71 48,91 Lut 8 588 1,00 4,48 9,38 (°) 31,75 54,39 (*)
Lut 10 604 0,87 8,66 10,58 35,43 45,12 * - semnificativ pozitiv (p<0,01); ° - semnificativ negativ (p<0,05)
5.1.4. Evaluarea viabilităţii şi maturării nucleare a ovocitelor cultivate în mediu cu α-tocoferol Determinarea viabilităţii celulare are la bază prezenţa esterazelor citoplasmatice ce clivează FDA pentru a forma fluoresceina. Astfel, ovocitele care prezintă activitate normală a esterazelor ca indicator al viabilităţii sunt verzi, fluorescente şi strălucitoare în contrast cu cele neviabile care sunt estompate sau nefluorescente. Testul a fost completat de colorarea cu iodură de propidiu, substanţă ce pătrunde în ovocită dacă membrana plasmatică este lezată. Celulele moarte se colorează în roşu. Pentru a evidenţia maturarea nucleară s-a recurs la colorarea ovocitelor cu Hoechst 33258. El se leagă la ADN şi emite lumină albastră fluorescentă ce facilitează identificarea primului globul polar în spaţiul perivitelin al ovocitei.
15
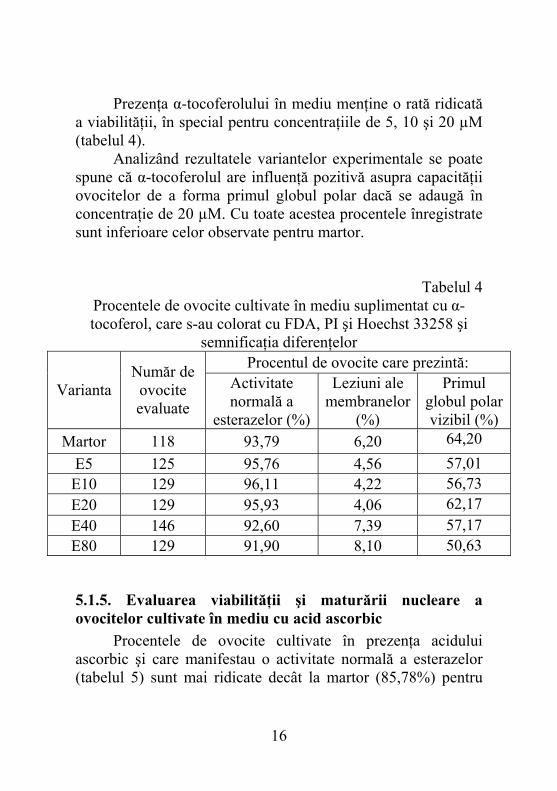
Prezenţa α-tocoferolului în mediu menţine o rată ridicată a viabilităţii, în special pentru concentraţiile de 5, 10 şi 20 µM (tabelul 4). Analizând rezultatele variantelor experimentale se poate spune că α-tocoferolul are influenţă pozitivă asupra capacităţii ovocitelor de a forma primul globul polar dacă se adaugă în concentraţie de 20 µM. Cu toate acestea procentele înregistrate sunt inferioare celor observate pentru martor.
Tabelul 4 Procentele de ovocite cultivate în mediu suplimentat cu α-tocoferol, care s-au colorat cu FDA, PI şi Hoechst 33258 şi
semnificaţia diferenţelor Procentul de ovocite care prezintă:
Varianta Număr de
ovocite evaluate
Activitate normală a
esterazelor (%)
Leziuni ale membranelor
(%)
Primul globul polar vizibil (%)
Martor 118 93,79 6,20 64,20 E5 125 95,76 4,56 57,01 E10 129 96,11 4,22 56,73 E20 129 95,93 4,06 62,17 E40 146 92,60 7,39 57,17 E80 129 91,90 8,10 50,63
5.1.5. Evaluarea viabilităţii şi maturării nucleare a ovocitelor cultivate în mediu cu acid ascorbic Procentele de ovocite cultivate în prezenţa acidului ascorbic şi care manifestau o activitate normală a esterazelor (tabelul 5) sunt mai ridicate decât la martor (85,78%) pentru
16
toate variantele experimentale, cu două excepţii, şi anume C250 (85,16%) şi C500 (82,13%). Prin compararea rezultatelor obţinute prin testele de colorare (FDA şi PI) în cazul vitaminei C rezultă că adăugarea acidului ascorbic, în anumite concentraţii la mediul de maturare ovocitară are acţiune favorabilă asupra păstrării viabilităţii ovocitelor.
Tabelul 5 Procentele de ovocite cultivate în mediu suplimentat cu acid ascorbic, care s-au colorat cu FDA, PI şi Hoechst 33258 şi
semnificaţia diferenţelor Procentul de ovocite care prezintă:
Varianta
Număr de
ovocite evaluate
Activitate normală a
esterazelor (%)
Leziuni ale membranelor
(%)
Primul globul polar vizibil (%)
Martor 164 85,78 14,22 42,72 C50 160 90,08 9,92 50,39 C150 139 89,30 10,70 53,18 C250 152 85,16 14,84 58,53 (**) C500 166 82,13 17,87 45,85 C750 160 88,59 11,41 56,22 (*) * - semnificativ pozitiv (p<0,05); ** - distinct semnificativ pozitiv (p<0,01); ° - semnificativ negativ (p<0,05) Pentru această substanţă antioxidantă procentele de ovocite cu primul globul polar variază de la un minim de 45,85% înregistrat pentru varianta C500 până la valoarea maximă de 58,53% în cazul variantei C250. Analiza statistică a relevat existenţa unor diferenţe semnificative faţă de martor pentru 750 µM şi distinct semnificative pentru concentraţia de 250 µM acid ascorbic.
17
5.1.6. Evaluarea viabilităţii şi maturării nucleare a ovocitelor cultivate în mediu cu luteină Rezultatele arată că prin adăugarea de 2,5 µM luteină (tabelul 6) la mediul de maturare al ovocitelor va rezulta un număr semnificativ mai mic de celule care prezintă leziuni ale oolemei decât în cazul în care se adaugă 8 µM din aceeaşi substanţă. Concomitent şi proporţional creşte numărul de ovocite viabile.
Tabelul 6 Procentele de ovocite cultivate în mediu suplimentat cu luteină, care s-au colorat cu FDA, PI şi Hoechst 33258 şi semnificaţia
diferenţelor Procentul de ovocite care prezintă:
Varianta
Număr de
ovocite evaluate
Activitate normală a
esterazelor (%)
Leziuni ale membranelor
(%)
Primul globul polar vizibil (%)
Martor 134 95,14 (ab) 4,86 (ab) 67,62 Lut 2,5 117 98,32 (a) 2,01 (b) 72,34 Lut 4 128 96,93 (ab) 3,40 (ab) 71,53 Lut 5 128 98,02 (ab) 2,31 (ab) 64,96 Lut 8 148 94,28 (b) 5,71 (a) 67,25 Lut 10 153 97,34 (ab) 2,66 (ab) 66,18 Diferenţa între oricare două valori, urmate de cel puţin o literă comună, este nesemnificativă. În cazul cultivării în mediu suplimentat cu luteină procentele de ovocite care prezentă primul globul polar sunt superioare situaţiei întâlnite la martor pentru concentraţiile de 2,5 şi 4 µM luteină. Situaţia coroborează observaţiile efectuate la evaluarea viabilităţii şi integrităţii membranelor, unde
18
varianta 2,5 µM este cea mai bine plasată între variantele experimentale dar nu depăşeşte lotul martor.
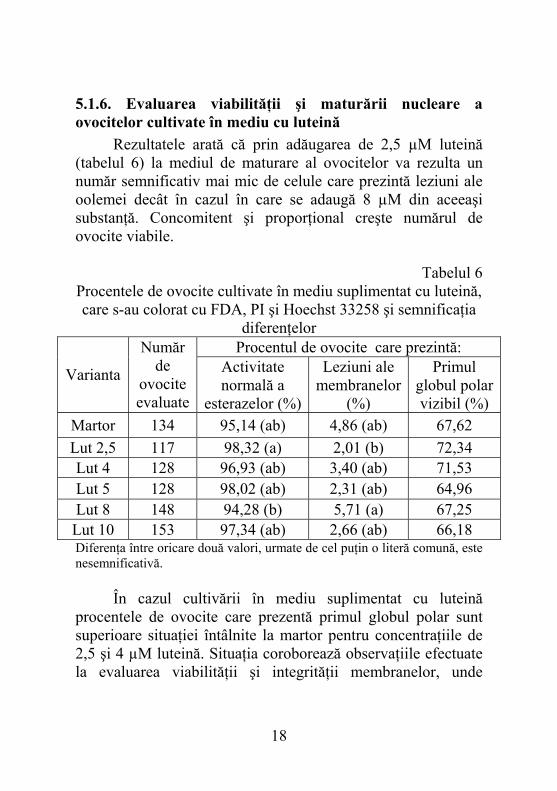
5.1.7. Determinarea conţinutului de glutation din ovocitele cultivate în medii de maturare suplimentate cu α-tocoferol Concentraţiile de glutation obţinute au oscilat între 12,94±0,77 pmoli/ovocit pentru varianta cu adaus de 80 µM α-tocoferol şi 14,09±0,78 pmoli/ovocit pentru cea suplimentată cu 5 µM. Diferenţele faţă de martor sunt semnificative pentru varianta E5 (figura 1).
pmoli/ovocit 13,19 14,09 13,53 13,50 13,27 12,94
M E5 E10 E20 E40 E80
*
Fig. 1. Concentraţia de glutation pentru ovocitele cultivate în
mediu cu α-tocoferol Dacă rezultatele sunt corelate cu ceea ce s-a obţinut în cazul celorlalte două metode de evaluare se ajunge la concluzia că α-tocoferolul în concentraţie de 5 µM favorizează metabolismul ovocitar ceea ce duce la creşterea viabilităţii şi
19

implicit acumularea unei cantităţi mai mari de glutation. De asemenea cantitatea mai mare de glutation demonstrează că α-tocoferolul asigură desfăşurarea normală a proceselor de maturate citoplasmatică. Concentraţiile rezultate se înscriu în limitele stabilite până în prezent pentru ovocitele maturate in vitro şi sugerează importanţa adăugării acestei substanţe la mediu pentru obţinerea unei maturări optime.
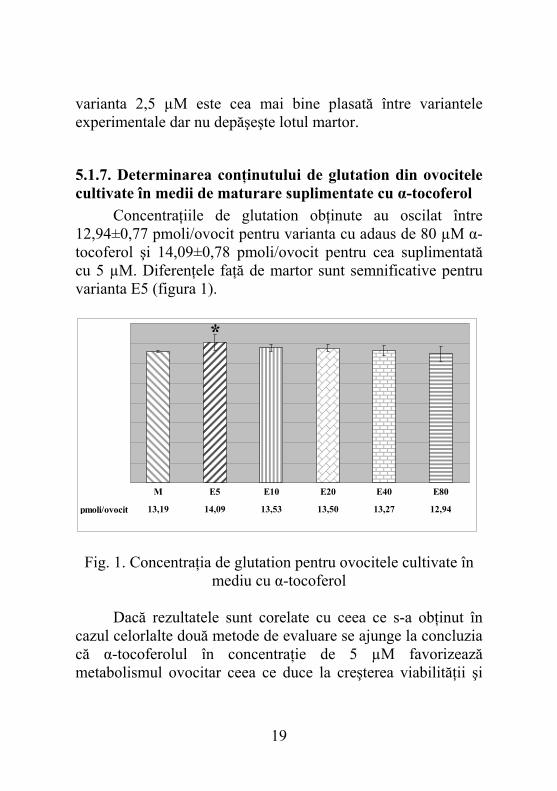
5.1.8. Determinarea conţinutului de glutation din ovocitele cultivate în medii de maturare suplimentate cu acid ascorbic Diferenţele observate au fost semnificativ mai mari comparativ lotului martor pentru concentraţia de 750 µM acid ascorbic şi semnificativ mai mici pentru 150 µM (figura 2). Având în vedere numărul semnificativ mai mare de ovocite la care s-a observat prezenţa primului globul polar, precum şi concentraţia crescută de glutation, putem considera că acidul ascorbic asigurat în concentraţie de 750 µM determină atingerea maturării nucleare şi citoplasmatice de către un număr mai mare de ovocite. Inventariind şi numărul crescut de celule cu activitate enzimatică normală care rezultă în urma cultivării cu acid ascorbic se poate afirma că deşi acesta are un efect nefast asupra expandării, nu pare a influenţa negativ viabilitatea ovocitelor, acumularea de glutation şi trecerea lor în metafaza II a diviziunii meiotice.
20
pmoli/ovocit 10,90 11,22 9,24 11,38 12,27 12,86
M C50 C150 C250 C500 C750
*
°
Fig. 2. Concentraţia de glutation pentru ovocitele cultivate în mediu cu acid ascorbic
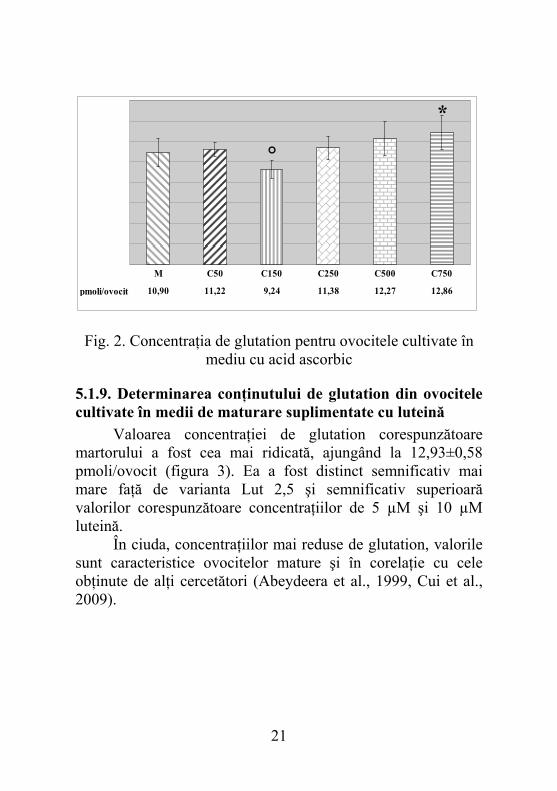
5.1.9. Determinarea conţinutului de glutation din ovocitele cultivate în medii de maturare suplimentate cu luteină Valoarea concentraţiei de glutation corespunzătoare martorului a fost cea mai ridicată, ajungând la 12,93±0,58 pmoli/ovocit (figura 3). Ea a fost distinct semnificativ mai mare faţă de varianta Lut 2,5 şi semnificativ superioară valorilor corespunzătoare concentraţiilor de 5 µM şi 10 µM luteină. În ciuda, concentraţiilor mai reduse de glutation, valorile sunt caracteristice ovocitelor mature şi în corelaţie cu cele obţinute de alţi cercetători (Abeydeera et al., 1999, Cui et al., 2009).
21

pmoli/ovocit 12,93 10,67 11,82 11,60 11,93 11,63
M Lut 2,5 Lut 4 Lut 5 Lut 8 Lut 10
°°° °
Fig. 3. Concentraţia de glutation pentru ovocitele cultivate în mediu cu luteină
5.2. DETERMINAREA INFLUENŢEI Α-TOCOFEROLU- LUI, ACIDULUI ASCORBIC ŞI LUTEINEI ASUPRA DEZVOLTĂRII IN VITRO A EMBRIONILOR În cadrul experimentelor de fecundaţie in vitro şi cultură a embrionilor s-a folosit un număr de 5379 ovocite. Dezvoltarea in vitro a embrionilor prezintă unele particularităţi comparativ situaţiei in vivo. Acestea se referă la încetinirea ritmului evoluţiei şi dificultatea de a depăşi unele stadii considerate „puncte de control”. Cauzele acestor întârzieri sunt numeroase, ele referindu-se în principal la: condiţii de cultură suboptimale comparativ mediului oviductal, conţinutul scăzut de glutation, stresul oxidativ, întârzieri în activarea genomului embrionar, probleme de comunicare între celulele embrionare.
22
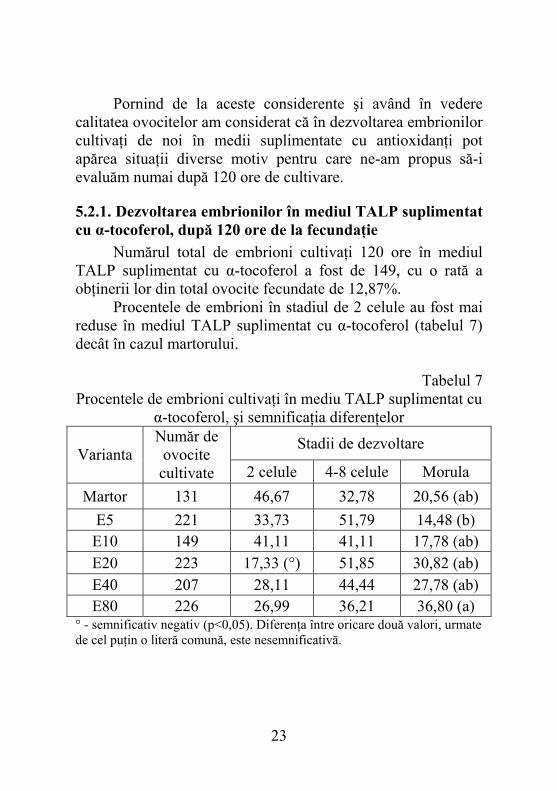
Pornind de la aceste considerente şi având în vedere calitatea ovocitelor am considerat că în dezvoltarea embrionilor cultivaţi de noi în medii suplimentate cu antioxidanţi pot apărea situaţii diverse motiv pentru care ne-am propus să-i evaluăm numai după 120 ore de cultivare.
5.2.1. Dezvoltarea embrionilor în mediul TALP suplimentat cu α-tocoferol, după 120 ore de la fecundaţie Numărul total de embrioni cultivaţi 120 ore în mediul TALP suplimentat cu α-tocoferol a fost de 149, cu o rată a obţinerii lor din total ovocite fecundate de 12,87%. Procentele de embrioni în stadiul de 2 celule au fost mai reduse în mediul TALP suplimentat cu α-tocoferol (tabelul 7) decât în cazul martorului.
Tabelul 7 Procentele de embrioni cultivaţi în mediu TALP suplimentat cu
α-tocoferol, şi semnificaţia diferenţelor
Stadii de dezvoltare Varianta
Număr de ovocite cultivate 2 celule 4-8 celule Morula
Martor 131 46,67 32,78 20,56 (ab) E5 221 33,73 51,79 14,48 (b) E10 149 41,11 41,11 17,78 (ab) E20 223 17,33 (°) 51,85 30,82 (ab) E40 207 28,11 44,44 27,78 (ab) E80 226 26,99 36,21 36,80 (a)
° - semnificativ negativ (p<0,05). Diferenţa între oricare două valori, urmate de cel puţin o literă comună, este nesemnificativă.
23
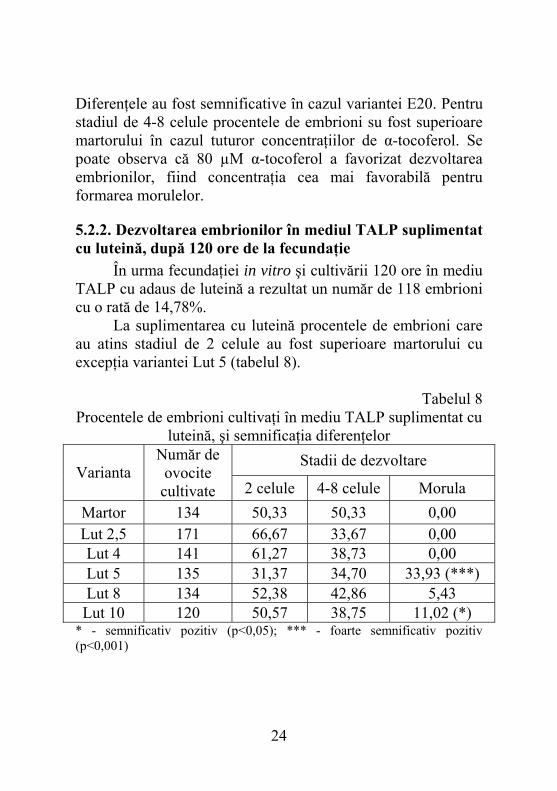
Diferenţele au fost semnificative în cazul variantei E20. Pentru stadiul de 4-8 celule procentele de embrioni su fost superioare martorului în cazul tuturor concentraţiilor de α-tocoferol. Se poate observa că 80 µM α-tocoferol a favorizat dezvoltarea embrionilor, fiind concentraţia cea mai favorabilă pentru formarea morulelor.
5.2.2. Dezvoltarea embrionilor în mediul TALP suplimentat cu luteină, după 120 ore de la fecundaţie În urma fecundaţiei in vitro şi cultivării 120 ore în mediu TALP cu adaus de luteină a rezultat un număr de 118 embrioni cu o rată de 14,78%. La suplimentarea cu luteină procentele de embrioni care au atins stadiul de 2 celule au fost superioare martorului cu excepţia variantei Lut 5 (tabelul 8).
Tabelul 8 Procentele de embrioni cultivaţi în mediu TALP suplimentat cu
luteină, şi semnificaţia diferenţelor Stadii de dezvoltare
Varianta Număr de
ovocite cultivate 2 celule 4-8 celule Morula
Martor 134 50,33 50,33 0,00 Lut 2,5 171 66,67 33,67 0,00 Lut 4 141 61,27 38,73 0,00 Lut 5 135 31,37 34,70 33,93 (***) Lut 8 134 52,38 42,86 5,43 Lut 10 120 50,57 38,75 11,02 (*)
* - semnificativ pozitiv (p<0,05); *** - foarte semnificativ pozitiv (p<0,001)
24

Spre deosebire de această situaţie, pentru stadiul de 4-8 celule procentul de embrioni dezvoltaţi în mediul TALP suplimentat cu luteină este mai redus decât cel corespunzător martorului. În cazul ultimului stadiu, concentraţiile de 5 µM şi 10 µM luteină au favorizat obţinerea unor procente de morule semnificativ mai mari faţă de cele observate la martor.
5.2.3. Dezvoltarea embrionilor în mediul NCSU-23 suplimentat cu α-tocoferol, după 48 ore de la fecundaţie Pornind de la diversitatea stadiilor de dezvoltare a embrionilor analizaţi după 120 ore de cultivare în mediul TALP am considerat necesar ca în următoarele experienţe să mai introducem o etapă de evaluare, la 48 ore după fecundaţie. Astfel s-a creat posibilitatea, nu numai să se inventarieze stadiul de dezvoltare al embrionilor, dar să şi urmărim cum au evoluat până la 120 ore în funcţie de tipul şi concentraţia de substanţe antioxidante suplimentate la mediul NCSU-23. Pentru stadiul de dezvoltare a embrionilor de 2 celule se observă că procentele corespunzătoare martorului (38,04%) sunt superioare celor caracteristice variantelor experimentale (tabelul 9). În cazul acestora valorile variază de la 18,26% pentru E40 până la 29,09% (E80). Diferenţele înregistrate în cazul variantelor ce conţineau 5 µM, 20 µM şi 40 µM s-au dovedit a fi semnificative dar inferioare lotului martor. În cazul stadiului de 4-8 celule sensul diferenţelor este opus faţă de stadiul anterior în cazul căruia procentele erau inferioare martorului. Această constatare ne conduce spre ideea că majoritatea embrionilor au ajuns într-un stadiu superior dacă s-au cultivat în mediu suplimentat cu α-tocoferol. Gradele de semnificaţie au variat, diferenţele fiind semnificative pentru
25
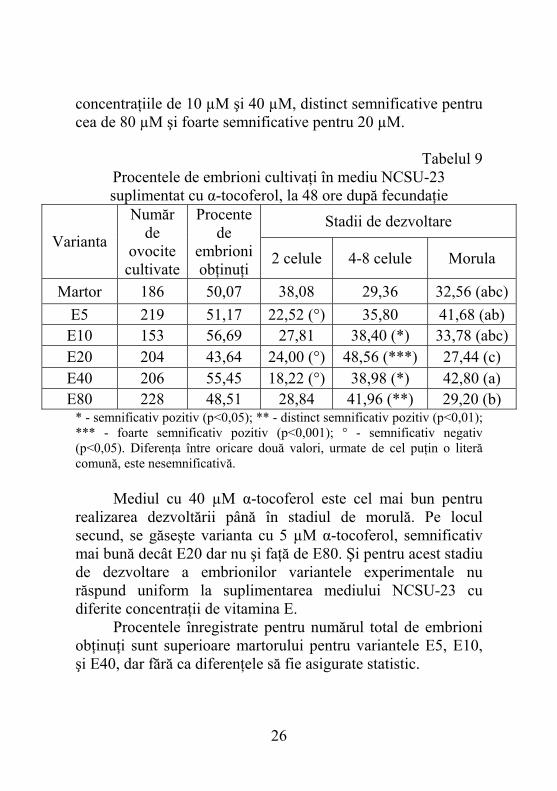
concentraţiile de 10 µM şi 40 µM, distinct semnificative pentru cea de 80 µM şi foarte semnificative pentru 20 µM.
Tabelul 9 Procentele de embrioni cultivaţi în mediu NCSU-23 suplimentat cu α-tocoferol, la 48 ore după fecundaţie
Stadii de dezvoltare Varianta
Număr de
ovocite cultivate
Procente de
embrioni obţinuţi
2 celule 4-8 celule Morula
Martor 186 50,07 38,08 29,36 32,56 (abc) E5 219 51,17 22,52 (°) 35,80 41,68 (ab) E10 153 56,69 27,81 38,40 (*) 33,78 (abc) E20 204 43,64 24,00 (°) 48,56 (***) 27,44 (c) E40 206 55,45 18,22 (°) 38,98 (*) 42,80 (a) E80 228 48,51 28,84 41,96 (**) 29,20 (b)
* - semnificativ pozitiv (p<0,05); ** - distinct semnificativ pozitiv (p<0,01); *** - foarte semnificativ pozitiv (p<0,001); ° - semnificativ negativ (p<0,05). Diferenţa între oricare două valori, urmate de cel puţin o literă comună, este nesemnificativă. Mediul cu 40 µM α-tocoferol este cel mai bun pentru realizarea dezvoltării până în stadiul de morulă. Pe locul secund, se găseşte varianta cu 5 µM α-tocoferol, semnificativ mai bună decât E20 dar nu şi faţă de E80. Şi pentru acest stadiu de dezvoltare a embrionilor variantele experimentale nu răspund uniform la suplimentarea mediului NCSU-23 cu diferite concentraţii de vitamina E. Procentele înregistrate pentru numărul total de embrioni obţinuţi sunt superioare martorului pentru variantele E5, E10, şi E40, dar fără ca diferenţele să fie asigurate statistic.
26
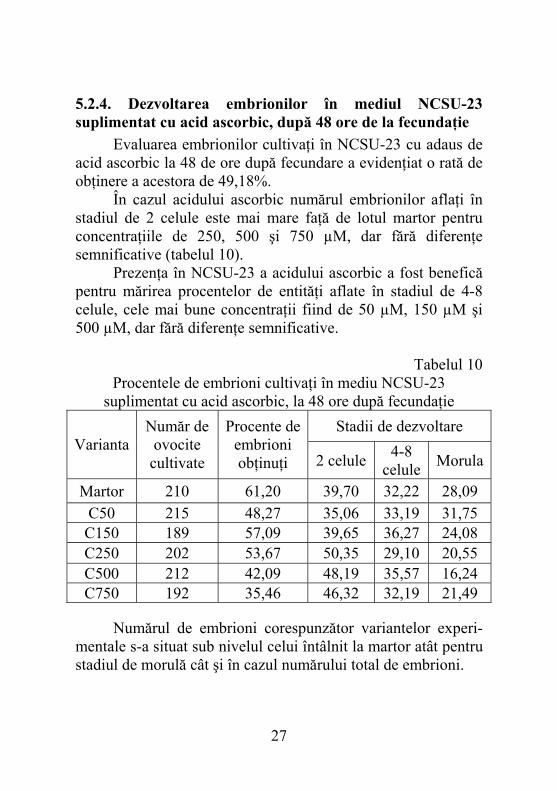
5.2.4. Dezvoltarea embrionilor în mediul NCSU-23 suplimentat cu acid ascorbic, după 48 ore de la fecundaţie
Evaluarea embrionilor cultivaţi în NCSU-23 cu adaus de acid ascorbic la 48 de ore după fecundare a evidenţiat o rată de obţinere a acestora de 49,18%. În cazul acidului ascorbic numărul embrionilor aflaţi în stadiul de 2 celule este mai mare faţă de lotul martor pentru concentraţiile de 250, 500 şi 750 µM, dar fără diferenţe semnificative (tabelul 10). Prezenţa în NCSU-23 a acidului ascorbic a fost benefică pentru mărirea procentelor de entităţi aflate în stadiul de 4-8 celule, cele mai bune concentraţii fiind de 50 µM, 150 µM şi 500 µM, dar fără diferenţe semnificative.
Tabelul 10 Procentele de embrioni cultivaţi în mediu NCSU-23
suplimentat cu acid ascorbic, la 48 ore după fecundaţie Stadii de dezvoltare
Varianta Număr de
ovocite cultivate
Procente de embrioni obţinuţi 2 celule 4-8
celule Morula
Martor 210 61,20 39,70 32,22 28,09 C50 215 48,27 35,06 33,19 31,75
C150 189 57,09 39,65 36,27 24,08 C250 202 53,67 50,35 29,10 20,55 C500 212 42,09 48,19 35,57 16,24 C750 192 35,46 46,32 32,19 21,49
Numărul de embrioni corespunzător variantelor experi- mentale s-a situat sub nivelul celui întâlnit la martor atât pentru stadiul de morulă cât şi în cazul numărului total de embrioni.
27
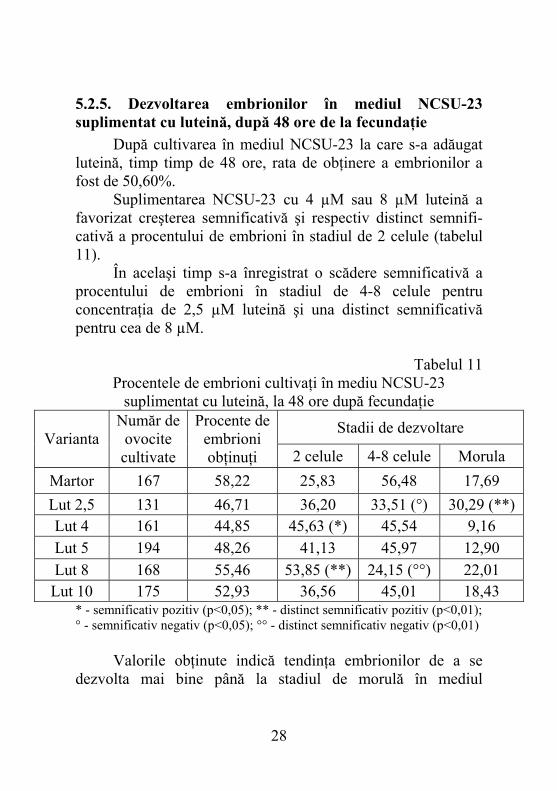
5.2.5. Dezvoltarea embrionilor în mediul NCSU-23 suplimentat cu luteină, după 48 ore de la fecundaţie
După cultivarea în mediul NCSU-23 la care s-a adăugat luteină, timp timp de 48 ore, rata de obţinere a embrionilor a fost de 50,60%. Suplimentarea NCSU-23 cu 4 µM sau 8 µM luteină a favorizat creşterea semnificativă şi respectiv distinct semnifi- cativă a procentului de embrioni în stadiul de 2 celule (tabelul 11). În acelaşi timp s-a înregistrat o scădere semnificativă a procentului de embrioni în stadiul de 4-8 celule pentru concentraţia de 2,5 µM luteină şi una distinct semnificativă pentru cea de 8 µM.
Tabelul 11 Procentele de embrioni cultivaţi în mediu NCSU-23
suplimentat cu luteină, la 48 ore după fecundaţie
Stadii de dezvoltare Varianta
Număr de ovocite cultivate
Procente de embrioni obţinuţi 2 celule 4-8 celule Morula
Martor 167 58,22 25,83 56,48 17,69 Lut 2,5 131 46,71 36,20 33,51 (°) 30,29 (**) Lut 4 161 44,85 45,63 (*) 45,54 9,16 Lut 5 194 48,26 41,13 45,97 12,90 Lut 8 168 55,46 53,85 (**) 24,15 (°°) 22,01 Lut 10 175 52,93 36,56 45,01 18,43
* - semnificativ pozitiv (p<0,05); ** - distinct semnificativ pozitiv (p<0,01); ° - semnificativ negativ (p<0,05); °° - distinct semnificativ negativ (p<0,01) Valorile obţinute indică tendinţa embrionilor de a se dezvolta mai bine până la stadiul de morulă în mediul
28

suplimentat cu 2,5, 8 sau 10 µM faţă de cel conţinând 4 sau 5 µM luteină. Tendinţa este întărită de rezultatele analizei statistice care evaluează diferenţele înregistrate între martor şi varianta Lut 2,5 ca fiind distinct semnificative. Acest fapt sugerează o mai bună capacitate a mediului cu 2,5 µM luteină de a favoriza şi susţine dezvoltarea embrionilor suini până la stadiul de morulă. Procentele rezultate după analiza asupra numărului total de embrioni au fost mai reduse decât martorul, dar nesemnificative.
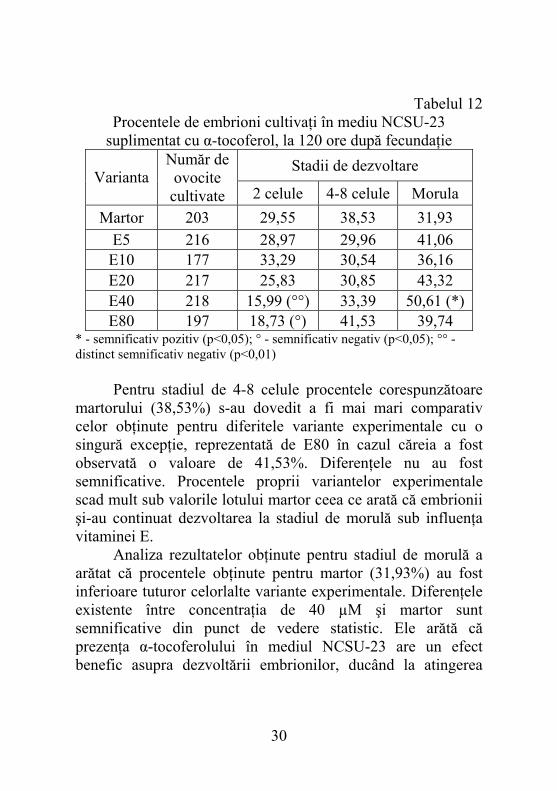
5.2.6. Dezvoltarea embrionilor în mediul NCSU-23 suplimentat cu α-tocoferol, după 120 ore de la fecundaţie Numărul total de embrioni cultivaţi până la 120 ore este cu puţin diferit (în plus sau în minus câţiva embrioni la unele loturi) comparativ evaluării la 48 ore. Diferenţele apar pentru că unii zigoţi au pornit mitozele mai târziu, iar alţi embrioni s-au eliminat atunci când a fost efectuat pasajul în mediu proaspăt la fiecare 48 ore. Prin compararea procentelor corespunzătoare martorului cu cele remarcate în cazul diferitelor concentraţii de α-tocoferol, se poate observa că mai există încă un număr important de embrioni rămaşi în stadiul de 2 celule. Ca un element pozitiv menţionăm situaţia variantelor E40 şi E80, la care concentraţiile mai mari de α-tocoferol au determinat reducerea procentelor de embrioni întârziaţi în stadiul de 2 celule, la valori de 15,99% şi respectiv 18,73%, diferenţele fiind semnificative. Aceste ultime procente sunt mai mici comparativ evaluării efectuate la 48 ore şi arată că o parte din embrioni au continuat să se dezvolte.
29
Tabelul 12 Procentele de embrioni cultivaţi în mediu NCSU-23
suplimentat cu α-tocoferol, la 120 ore după fecundaţie Stadii de dezvoltare
Varianta Număr de
ovocite cultivate 2 celule 4-8 celule Morula
Martor 203 29,55 38,53 31,93 E5 216 28,97 29,96 41,06 E10 177 33,29 30,54 36,16 E20 217 25,83 30,85 43,32 E40 218 15,99 (°°) 33,39 50,61 (*) E80 197 18,73 (°) 41,53 39,74
* - semnificativ pozitiv (p<0,05); ° - semnificativ negativ (p<0,05); °° - distinct semnificativ negativ (p<0,01) Pentru stadiul de 4-8 celule procentele corespunzătoare martorului (38,53%) s-au dovedit a fi mai mari comparativ celor obţinute pentru diferitele variante experimentale cu o singură excepţie, reprezentată de E80 în cazul căreia a fost observată o valoare de 41,53%. Diferenţele nu au fost semnificative. Procentele proprii variantelor experimentale scad mult sub valorile lotului martor ceea ce arată că embrionii şi-au continuat dezvoltarea la stadiul de morulă sub influenţa vitaminei E. Analiza rezultatelor obţinute pentru stadiul de morulă a arătat că procentele obţinute pentru martor (31,93%) au fost inferioare tuturor celorlalte variante experimentale. Diferenţele existente între concentraţia de 40 µM şi martor sunt semnificative din punct de vedere statistic. Ele arătă că prezenţa α-tocoferolului în mediul NCSU-23 are un efect benefic asupra dezvoltării embrionilor, ducând la atingerea
30

stadiului de morulă de către un număr semnificativ mai mare de entităţi. Situaţia prezentată aici este corelată cu ceea ce s-a observat pentru acelaşi grad de dezvoltare în intervalul de 48 ore, când E40 s-a dovedit a fi cea mai bună dintre variantele experimentale, chiar dacă diferenţele faţă de martor nu au fost semnificative. La 48 ore cele mai multe entităţi se aflau în stadiul de 4-8 celule. O comparaţie cu această situaţie ne arată că, vitamina E are o influenţă semnificativă şi pozitivă determinând creşterea numărului de embrioni aflaţi în stadiul de morulă după 120 ore de cultivare.
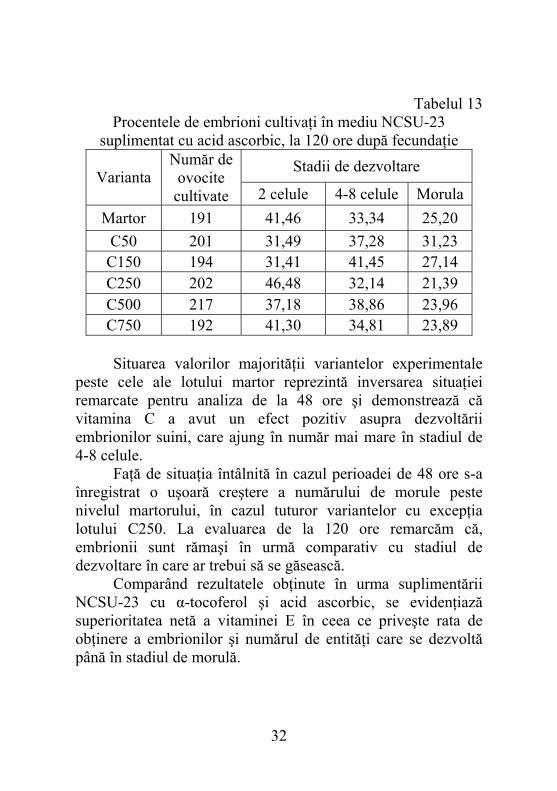
5.2.7. Dezvoltarea embrionilor în mediul NCSU-23 suplimentat cu acid ascorbic, după 120 ore de la fecundaţie Compararea procentelor de embrioni aflaţi în stadiul de 2 celule pentru diferite variante experimentale a arătat că martorul este superior tuturor concentraţiilor cu excepţia celei de 250 µM (46,48%) (tabelul 13). Comparativ evaluării efectuate la 48 ore, procentul de embrioni în stadiul de 2 celule este cu puţin mai mare în cazul lotului martor şi invariabil mai mic, dar la valori proprii pentru variantele experimentale. Motivarea situaţiei remarcate este întârzierea dezvoltării embrionare şi continuarea ei în cazul mediului cu diferite concentraţii de acid ascorbic. Pentru următorul stadiu de dezvoltare procentele corespunzătoare diferitelor variante, cu excepţia lui C250 (32,14%) s-au situat deasupra nivelului înregistrat pentru martor (33,34%).
31
Tabelul 13 Procentele de embrioni cultivaţi în mediu NCSU-23
suplimentat cu acid ascorbic, la 120 ore după fecundaţie Stadii de dezvoltare
Varianta Număr de
ovocite cultivate 2 celule 4-8 celule Morula
Martor 191 41,46 33,34 25,20 C50 201 31,49 37,28 31,23 C150 194 31,41 41,45 27,14 C250 202 46,48 32,14 21,39 C500 217 37,18 38,86 23,96 C750 192 41,30 34,81 23,89
Situarea valorilor majorităţii variantelor experimentale peste cele ale lotului martor reprezintă inversarea situaţiei remarcate pentru analiza de la 48 ore şi demonstrează că vitamina C a avut un efect pozitiv asupra dezvoltării embrionilor suini, care ajung în număr mai mare în stadiul de 4-8 celule. Faţă de situaţia întâlnită în cazul perioadei de 48 ore s-a înregistrat o uşoară creştere a numărului de morule peste nivelul martorului, în cazul tuturor variantelor cu excepţia lotului C250. La evaluarea de la 120 ore remarcăm că, embrionii sunt rămaşi în urmă comparativ cu stadiul de dezvoltare în care ar trebui să se găsească. Comparând rezultatele obţinute în urma suplimentării NCSU-23 cu α-tocoferol şi acid ascorbic, se evidenţiază superioritatea netă a vitaminei E în ceea ce priveşte rata de obţinere a embrionilor şi numărul de entităţi care se dezvoltă până în stadiul de morulă.
32
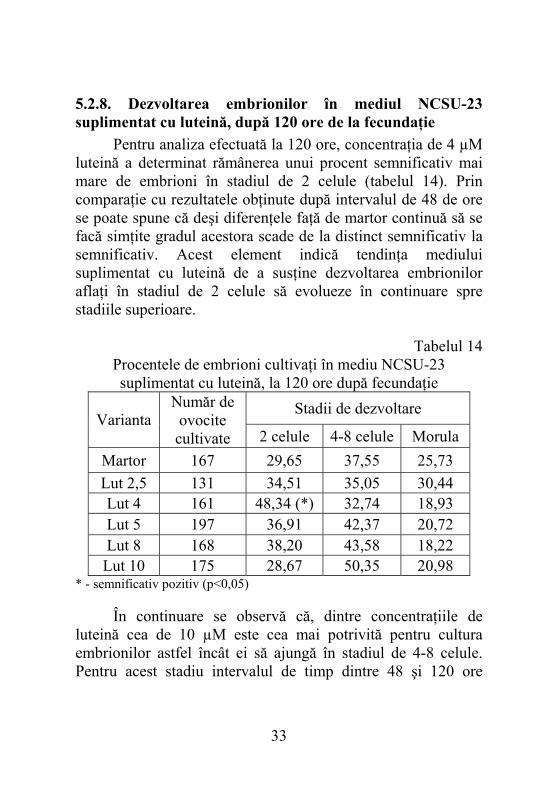
5.2.8. Dezvoltarea embrionilor în mediul NCSU-23 suplimentat cu luteină, după 120 ore de la fecundaţie Pentru analiza efectuată la 120 ore, concentraţia de 4 µM luteină a determinat rămânerea unui procent semnificativ mai mare de embrioni în stadiul de 2 celule (tabelul 14). Prin comparaţie cu rezultatele obţinute după intervalul de 48 de ore se poate spune că deşi diferenţele faţă de martor continuă să se facă simţite gradul acestora scade de la distinct semnificativ la semnificativ. Acest element indică tendinţa mediului suplimentat cu luteină de a susţine dezvoltarea embrionilor aflaţi în stadiul de 2 celule să evolueze în continuare spre stadiile superioare.
Tabelul 14 Procentele de embrioni cultivaţi în mediu NCSU-23 suplimentat cu luteină, la 120 ore după fecundaţie
Stadii de dezvoltare Varianta
Număr de ovocite cultivate 2 celule 4-8 celule Morula
Martor 167 29,65 37,55 25,73 Lut 2,5 131 34,51 35,05 30,44 Lut 4 161 48,34 (*) 32,74 18,93 Lut 5 197 36,91 42,37 20,72 Lut 8 168 38,20 43,58 18,22 Lut 10 175 28,67 50,35 20,98
* - semnificativ pozitiv (p<0,05) În continuare se observă că, dintre concentraţiile de luteină cea de 10 µM este cea mai potrivită pentru cultura embrionilor astfel încât ei să ajungă în stadiul de 4-8 celule. Pentru acest stadiu intervalul de timp dintre 48 şi 120 ore
33

egalizează diferenţele dintre martor şi variantele experimentale, ele devenind nesemnificative din punct de vedere statistic. Stadiul de morulă a fost atins de un procent de embrioni inferior martorului atunci când cultivarea s-a desfăşurat în mediu suplimentat cu 4 µM, 5 µM, 8 µM şi 10 µM luteină. În paralel s-a putut constata efectul pozitiv al concentraţiei de 2,5 µM. Privite în ansamblu datele referitoare la suplimentarea mediului de bază cu luteină demonstrează că la 120 ore se recuperează un număr redus de embrioni întârziaţi, la variantele experimentale. În cazul lotului martor embrionii întârziaţi care ajung la stadiul de morulă sunt mai numeroşi.
5.2.9. Comparaţie între NCSU-23 şi TALP ca medii de bază suplimentate cu antioxidanţi pentru dezvoltarea in vitro a embrionilor Mediul TALP are tendinţa de a susţine dezvoltarea embrionară numai până în stadiile sale timpurii (2 celule, 4-8 celule). Spre deosebire de acesta, mediul NCSU-23 favorizează creşterea numărului de embrioni aflaţi în stadiul de morulă. Analiza realizată pentru numărul total de embrioni oferă o privire generică asupra comportării celor două medii. În mediul NCSU-23 cu şi fără α-tocoferol sau luteină se obţin mult mai mulţi embrioni comparativ mediului TALP. Procentul total de embrioni obţinuţi şi dezvoltaţi este mai mare în cazul luteinei. Comparativ mediului TALP, NCSU-23 este superior atât la lotul martor cât şi la variantele experimentale.
34

CAPITOLUL 6. CONCLUZII 6.1. Concluzii privind determinarea influenţei α-tocoferolului, acidului ascorbic şi luteinei asupra maturării ovocitelor:
1. În urma coroborării rezultatelor obţinute prin aplicarea diferitelor metode de apreciere a maturării ovocitelor în medii suplimentate cu α-tocoferol, se remarcă creşterea procentului de ovocite viabile, care au concentraţii semnificativ mai mari de glutation şi grad de expandare ridicat a COC. Concentraţia optimă de α-tocoferol în mediile de maturare a ovocitelor suine este de 5 µM. Se evidenţiază de asemenea un procent mai mic de ovocite cu globul polar în spaţiul perivitelin, comparativ lotului martor. În consecinţă se poate afirma că prezenţa vitaminei E în mediul de maturare favorizează metabolismul ovocitar, ceea ce duce la creşterea viabilităţii şi a gradului de maturare citoplasmatică, dar are efect negativ asupra maturării nucleare.
2. Acidul ascorbic şi în special concentraţia de 750 µM a determinat obţinerea unui număr ridicat de ovocite viabile, mature nuclear şi citoplasmatic, dar la care expandarea celulelor cumulus ooforus este mai redusă.
3. Prezenţa în mediul de maturare a luteinei (concentraţia de 8 µM) a dus la creşterea semnificativă a procentelor de COC cu grad ridicat de expandare. Tendinţa s-a corelat cu numărul ridicat de ovocite viabile, mature nuclear, dar cu dificultăţi în realizarea maturării citoplasmatice.
4. Considerăm că diferenţele observate între cele trei substanţe sunt determinate de structurile chimice variate care duc la proprietăţi specifice. Astfel, α-tocoferolul şi luteina acţionează în compartimentul celular lipidic în timp ce acidul ascorbic este principalul antioxidant hidrosolubil. Aceste elemente confirmă ipoteza conform căreia carotenoidele sunt
35

parte a unei reţele de antioxidanţi intracelulari alături de vitaminele C şi E.
5. Pentru a evalua realizarea maturării sunt necesare tehnici multiple care să completeze metoda subiectivă dar rapidă, reprezentată de aprecierea gradului de expandare a celulelor cumulus ooforus. 6.2. Concluzii privind influenţa α-tocoferolului, acidului ascorbic şi luteinei asupra dezvoltării embrionilor în mediile TALP şi NCSU-23:
1. Adăugarea de α-tocoferol şi luteină la mediul TALP a dus la creşterea procentelor de embrioni ajunşi la 120 ore în stadiile de 4-8 celule şi morulă.
2. La 48 ore după fecundaţie prezenţa α-tocoferolului în mediul NCSU-23 are ca efect creşterea semnificativă a procentelor de embrioni dezvoltaţi până la stadiile de 4-8 celule (10, 20, 40, 80 µM) şi morulă (5, 40, 80 µM). Între 48-120 ore α-tocoferolul are efect pozitiv pentru că favorizează formarea morulelor.
3. Adăugarea acidului ascorbic la mediul NCSU-23, influenţează negativ dezvoltarea embrionară. După 48 ore de cultivare majoritatea embrionilor se găsesc în stadiile de 2 şi 4-8 celule. Evaluarea embrionilor la 120 ore arată că o parte a celor aflaţi în stadiul de două celule au evoluat la 4-8 celule fără a ajunge la morulă.
4. Evaluarea la 48 ore a arătat că suplimentarea mediului NCSU-23 cu luteină are efecte pozitive asupra dezvoltării embrionilor. Concentraţia de 2,5 µM luteină determină creşterea procentelor de embrioni ajunşi în stadiul de morulă, situaţie care se menţine şi la 120 ore.
5. Procentele corespunzătoare numărului total de embrioni cultivaţi în mediul NCSU-23 sunt net superioare faţă de TALP
36

în cazul ambelor substanţe antioxidante utilizate. Mediul TALP nu corespunde cerinţelor de dezvoltare a embrionilor suini. Adăugarea antioxidanţilor α-tocoferol sau luteină nu poate corecta acest neajuns.
37

BIBLIOGRAFIE SELECTIVĂ 1. ABEYDEERA, L.R., W.H. WANG, T.C. CANTLEY, R.S.
PRATHER, B.N. DAY, 1999, Glutathione content and embryo development after in vitro fertilisation of pig oocytes matured in the presence of a thiol compound and various concentrations of cysteine, Zygote, 7, 203–210.
2. ANDERSON, MARY, 1985. Determination of glutathione and glutathione disulfide in biological samples, Methods in Enzymology, 113, 548-555.
3. BOVERIS, A., AND CHANCE B., 1973, The mitochondrial generation of hydrogen peroxide. General properties and effect of hyperbaric oxygen, Biochemistry Journal, 134, 707-716.
4. BOYD, V., O.M. CHOLEWA, K.K. PAPAS, 2008, Limitations in the use of fluorescein diacetate/propidium iodide (FDA/PI) and cell permeable nucleic acid stains for viability measurements of isolated islets of Langerhans, Current Trends in Biotechnology and Pharmacy, 2(2), 28-304.
5. CHEW B.P., AND JEAN SOON PARK, 2004, Carotenoid action on the immune response, Journal of Nutrition, 134, 257S-261S.
6. CUI, M.S., Y.P. FAN, Y. WU, Z.D. HAO, S. LIU, X.J. CHEN, S.M. ZENG, 2009, Porcine cumulus cell influences ooplasmic mitochondria-lipid distributions, GSH-ATP contents and calcium release pattern after electro-activation, Theriogenology 71, 412-421.
7. DROGE, W., 2002, Free radicals in the physiological control of cell function, Physiology Reviews, 82, 47-95.
8. GRUPEN, C.G., H. NAGASHIMA, M.B. NOTTLE, 1995,
38

Cysteamine enhances in vitro development of porcine oocytes matured and fertilized in vitro, Biology of Reproduction, 53, 173-178.
9. GUERIN, P., S.E. MOUATASSIM, Y. MENEZO, 2001, Oxidative stress and protection against reactive oxygen species in the preimplantation embryo and its surroundings, Human Reproduction Update, 7(2), 175-189.
10. GUPTA, M.K., S.J. UHM, H.T. LEE, 2007, Cryopreservation of immature and in vitro matured porcine oocytes by solid surface vitrification, Theriogenology, 67, 238-248.
11. HALLIWELL, B., AND J.M.C. GUTTERIDGE, 2007, Free Radicals in Biology and Medicine, fourth edition, Clarendon Press, Oxford.
12. HANCOCK, J.T., R. DESIKAN, S.J. NEILL, 2001, Role of reactive oxygen species in cell signaling pathways, Biochemical Society Transactions, 29, 345-350.
13. HOMA, S., C. RACOW, R. MCGAUGHEY, 1986, Lipid analysis of immature pig oocytes, Journal of Reproduction and Fertility, 77, 425-434.
14. HOSSEIN, M.S., M.A. HASHEM, Y.W. JEONG, M.S. LEE, SUE KIM, J.H. KIM, O.J. KOO, S.M. PARK, E.G. LEE, S.W. PARK, S.K. KANG, B.C. LEE, W.S. HWANG, 2007, Temporal effect of α-tocoferol and L-ascorbic on in vitro fertilized porcine embryo development, Animal Reproduction Science, 100, 107-117.
15. KOVACIC, P., AND J.D. JACINTHO, 2001, Reproductive toxins: pervasive theme of oxidative stress and electron transfer, Current Medicinal Chemistry, 8(7), 863-892.
16. LADOŞI, I., 1999, Embriologie animală, Editura Victor Melenti, Cluj-Napoca.
39

17. MCEVOY, T., G. COULL, P. BROADBENT, J. HUTCHINSON, B. SPEAKE, 2000, Fatty acid composition of lipids in immature cattle pig and sheep oocytes with intact zona pellucida, Journal of Reproduction and Fertility, 118, 163–170.
18. MCGAUGHEY, R.W., 1983, Regulation of oocyte maturation, In: Finn, C.A., Oxford Reviews of Reproductive Biology, Oxford University Press, Oxford, 5, 106–130.
19. MENDOZA, C., E. RUIZ-REQUENA, E. ORTEGA, N. CREMADES, F. MARTINEZ, R. BERNABEU, E. GRECO, J. TESARIK, 2002, Follicular fluid markers of oocyte developmental potential, Human Reproduction, 17, 1017-1022.
20. OLSON, S.E. AND G.E. SEIDEL, 2000a, Reduced oxygen tension and EDTA improve bovine zygote development in a chemically defined medium, Journal of Animal Science, 78, 152-157.
21. TAO, Y., B. ZHOU, G. XIA, F. WANG, Z. WU, M. FU, 2004, Exposure to L-ascorbic acid or α-tocopherol facilitates the development of porcine denuded oocytes from metaphase I to metaphase II and prevents cumulus cells from fragmentation, Reproduction in Domestic Animals, 39, 52-57.
22. WARREN S., S. PATEL, C.M. KAPRON, 2000, The effect of vitamin E exposure on cadmium toxicity in mouse embryo cells in vitro, Toxicology, ,142, 119–126.
40

UNIVERSITY OF AGRICULTURAL SCIENCES AND VETERINARY MEDICINE
DOCTORAL SCHOOL FACULTY OF ANIMAL HUSBANDRY AND
BIOTECHNOLOGIES
Eng. Miclea Ileana Raluca
EFFECT OF ANTIOXIDANT SUPPLEMENTATION TO OOCYTE
MATURATION AND EMBRYO CULTURE MEDIA
SUMMARY OF THE PhD THESIS Scientific coordinator,
Prof. dr. ing. Miclea Vasile
Cluj-Napoca
2010

TABLE OF CONTENTS
CHAPTER 1. OOCYTE MATURATION AND EMBRYO PRODUCTION....................................................................... 42 CHAPTER 2. FREE RADICALS AND ANTIOXIDANTS.. 43 GOAL AND OBJECTIVES ................................................... 46 CHAPTER 3. MATERIALS AND METHODS .................... 47
3.1. OOCYTE MATURATION ......................................... 47 3.1.1. Media .................................................................... 47 3.1.2. Oocyte collection and maturation ......................... 47 3.1.3. Maturation assessment .......................................... 48
3.2. IN VITRO FERTILIZATION AND EMBRYO CULTURE .......................................................................... 49
CHAPTER 4. RESULTS AND DISSCUSION...................... 50 4.1. THE EFFECT OF α-tocoPHerol, ascorbic aciD AND lutein ON OOCYTE MATURATION................................ 51
4.1.1. The effect of α-tocopherol on cumulus expansion ........................................................................ 51 4.1.2. The effect of ascorbic acid on cumulus expansion ........................................................................ 52 4.1.3. The effect of lutein on cumulus expansion ........... 52 4.1.4. Fluorescent staining for the assessment of viability and nuclear maturation of oocytes cultured in α-tocopherol supplemented media...................................... 53 4.1.5. Fluorescent staining for the assessment of viability and nuclear maturation of oocytes cultured in ascorbic acid supplemented media ................................................ 53 4.1.6. Fluorescent staining for the assessment of viability and nuclear maturation of oocytes cultured in lutein supplemented media........................................................ 55 4.1.7. Glutathione content of oocytes cultured in α-tocopherol supplemented media...................................... 56
I I

4.1.8. Glutathione content of oocytes cultured in ascorbic acid supplemented media ................................................ 57 4.1.9. Glutathione content of oocytes cultured in lutein supplemented media........................................................ 58
4.2. THE EFFECT OF Α-TOCOPHEROL, ASCORBIC ACID AND LUTEIN ON IN VITRO EMBRYO DEVELOPMENT ............................................................... 59
4.2.1. Embryo development in α-tocopherol supplemented TALP medium, 120 hours after fertilization................... 60 4.2.2. Embryo development in lutein supplemented TALP medium, 120 hours after fertilization.............................. 61 4.2.3. Embryo development in medium NCSU-23 supplemented with α-tocopherol, 48 hours after fertilization...................................................................... 62 4.2.4. Embryo development in medium NCSU-23 supplemented with ascorbic acid, 48 hours after fertilization...................................................................... 63 4.2.5. Embryo development in medium NCSU-23 supplemented with lutein, 48 hours after fertilization .... 64 4.2.6. Embryo development in medium NCSU-23 supplemented with α-tocopherol, 120 hours after fertilization...................................................................... 65 4.2.7. Embryo development in medium NCSU-23 supplemented with ascorbic acid, 120 hours after fertilization...................................................................... 66 4.2.8. Embryo development in medium NCSU-23 supplemented with lutein, 120 hours after fertilization .. 67 4.2.9. Comparison between NCSU-23 and TALP as antioxidant supplemented media for in vitro embryo culture.............................................................................. 67
CHAPTER 5. CONCLUSIONS ............................................. 68
II II

CHAPTER 1. OOCYTE MATURATION AND EMBRYO PRODUCTION
Oocyte maturation is usually referred to as the set of processes occurring from the germinal vesicle stage to completion of the second meiotic division with the formation of the first polar body. It consists of two aspects, i.e. nuclear and cytoplasm maturation. Nuclear maturation refers to the resumption of meiosis and progression to the metaphase II (MII) stage, whereas cytoplasmic maturation encompasses other, poorly understood, maturational events. Among them are the migration of cortical granules to the vicinity of the membrane, a multiplication of mitochondria and ribosome number, both proportional with the increase in protein synthesis and cell metabolic activity in general. These processes are believed to progress in parallel to one another, and synchronization of nuclear and cytoplasmic maturation is essential for establishing optimal oocyte developmental potential. Hormonal supplements, such as FSH, eCG or hCG, are added to the in vitro maturation medium in order to mimic the in vivo situation and stimulate nuclear maturation. However, whereas nuclear maturation can be evaluated by nuclear staining methods such as the various Hoechst dyes that reveal the first polar body, cytoplasmic maturation can only be determined by indirect means such as cumulus oophorus expansion and measuring the content of reduced glutathione (GSH). However, only the first method leaves the oocyte intact and has the least effect on its viability and ability to sustain fertilization. Therefore it becomes the most important mean of assessing maturation.
42 42

Fluorescent staining has been used to assess enzyme activity and membrane integrity as a viability indicator for several somatic cells types such as Langerhans islets (Boyd et al., 2008) and following vitrification of pig oocytes (Gupta et al., 2007). Cytoplasmic maturation of pig oocytes can be improved by reducing oxidative stress caused by reactive oxygen species (Mendoza et al., 2002). The same can be said about the equally lipid rich swine embryos. Improved cytoplasmic and nuclear maturation leads to an increase in the number of oocytes that are able to undergo fertilization and hence to more embryos that can be used for embryo transfer. Although, the in vitro production of swine embryos has been greatly improved by the development of NCSU-23, NCSU-37 and their subsequent modifications it is still not as efficient as its in vivo counterpart. Hence, the need for further study both in the field of oocyte maturation as well as embryo culture.
CHAPTER 2. FREE RADICALS AND ANTIOXIDANTS The ambivalent relationship between living organisms and oxygen is a consequence of the fact that on one side it fuels aerobic life and on the other it is one of the main sources for the production of free radials. These are, as defined by Halliwell and Gutteridge (2007) “...any species capable of independent existence that contains one or more unpaired electrons”. Their formation is guided by various enzymes such as cytochrome P450 mono-oxygenases, NADPH-oxidase, xanthine-oxidase but can also result in the mitochondria, through the leakage of approximately 1%-2% of the electrons
43 43

from the electron transfer chain. In addition they result through the influence of exogenous agents such as radiation, heavy metals and pesticides In the cell, free radicals function as signal molecules by activating transcription factors and enzymatic reactions (Droge, 2002). In addition they play a significant role in steroidogenesis, follicle formation and progesterone release by the corpus luteum and together with antioxidant enzymes bring about follicle wall rupture, ovulation and luteal regression. They are also involved in regulating embryo development and implantation (Guerin et al., 2001). Although cells have various antioxidant systems that should scavenge endogenous free radicals, their endogenous overproduction and the exogenous sources lead to an imbalance in redox metabolism and therefore to oxidative stress. It causes chain reactions that result in mitochondrial depolarization, cytochrome c release, lipid peroxidation, transcription factor activation and DNA damage leading to apoptotic and non-apoptotic cell death. In the oocyte and embryo free radical levels are controlled by metabolic pathways mediated by enzymes such as glutathione. However, the oocyte contains a large quantity of lipid droplets. This is particularly evident in the oocytes of the domestic pig, where very high levels of lipid have been reported, 161 μg (McEvoy et al., 2000). Triacylglycerol was the major lipid component followed by cholesterol and phosphatidylcholine. Analysis of fatty acids esterified to the individual phospholipids and neutral lipids has shown that there are high levels of palmitic acid (16:0) and the monounsaturated fatty acid oleic acid (18:1). Triacylglycerol, free fatty acids and most of the phospholipids, particularly phosphatidylethanolamine, are considerably enriched in n-6
44 44

polyunsaturated fatty acids, specifically linoleic (18:2), arachidonic (20:4) and adrenic (22:4) acids (Homa et al., 1986). Protection of the fatty acid and lipid components of oocytes and embryos that render them susceptible to free radical or other oxidative injury may prevent the damage currently associated with culture. According to Halliwell and Gutteridge (2007) antioxidants are “any substance that delays, prevents or removes oxidative damage to a target”. The definition applies to enzymes as well as non enzymatic antioxidants. To defend themselves, living systems employ antioxidants that they either produce or take up from the environment. Among the latter are Cu, Se, Mg, Zn, ascorbic acid and α-tocopherol more broadly known as vitamins C and E. Tocopherols (vitamin E), the most important lipid soluble antioxidant in the cell can be found in significant amounts in the ovary and the follicular fluid. It protects polyunsaturated fatty acids in membranes against free radicals (Tao et al., 2004) and improves the development of bovine embryos (Olson and Seidel, 2000). Ascorbic acid is the most important antioxidant outside the cell (Warren et al., 2000). It functions as a reducing agent of oxygen and cytocromes c and a, but can also protect membranes against peroxidation. It can prevent apoptosis in cultured mouse follicles and improve swine oocyte maturation (Tao et al., 2004) and embryo development of swine embryos (Hossein et al., 2007). Lutein, part of the xanthophylls class of carotenoids is one of the most potent lipid soluble antioxidant that is readily incorporated in membrane bilayers of mitochondria and microsomes (Chew and Park, 2004).
45 45

GOAL AND OBJECTIVES In order to maximize the number and quality of in vitro produced embryos we need to reduce and eliminate stress factors from the culture environment. In turn, this will lead to improved protocols for cryopreservation and embryo transfer. Our goal has been to improve oocyte maturation and embryo culture media by adding antioxidants such as α-tocopherol, ascorbic acid and lutein. These compounds occur naturally in biological systems and function as protection against oxidative stress. Each antioxidant was added to the oocyte maturation medium separately, using 5 different concentrations. Cytoplasmic and nuclear maturation were assessed by glutathione quantification and fluorescent staining respectively. Cell viability was also established by fluorescent staining. In order to examine oocyte relation to the cumulus oophorus cells, their expansion was graded. Next, antioxidant influence on embryo morphology and physiology was evaluated through. Swine embryos were cultured in media supplemented with the same antioxidants at the same concentrations. Development to the 2 cell, 4-8 cells and morula stages was assessed at 48 and 120 hours. These moments were chosen taking into consideration the slower development rate of in vitro produced embryos. Two culture media, NCSU-23 and TALP were compared in order to establish the most suitable for swine embryo culture.
46 46

CHAPTER 3 MATERIALS AND METHODS
3.1. OOCYTE MATURATION Swine oocytes were collected and cultured according to the modified protocols of Grupen et al. (1995) and Tao et al. (2004).
3.1.1. Media Oocytes were collected in M 199 supplemented with L-glutamine (3.4 g/l), NaHCO3 (2.2 g/l), Hepes (25 mM), penicillin (100 μg /ml) and streptomycin (100 IU/ml). For oocyte maturation, M 199 was supplemented with L-glutamine (3.4 g/l), Chorulon (10 IU/ml), Folligon (10 IU/ml), foetal bovine serum 10%, penicillin (100 μg /ml) and streptomycin (100 IU/ml). For the first experiment α-tocopherol dissolved in 95% ethanol solution was added to the maturation medium in order to arrive at concentrations of 5, 10, 20, 40 and 80 µM. Lutein dissolved in foetal bovine serum was added to the maturation medium in order to arrive at concentrations of 2.5, 4, 5, 8, and 10 μM carotenoid and 10% foetal bovine serum. In the last experiment, ascorbic acid dissolved in ultrapure water was added to the maturation medium in order to arrive at concentrations of 50, 150, 250, 500 and 750 µM.
3.1.2. Oocyte collection and maturation Porcine ovaries were collected from pre-pubertal gilts and transported to the laboratory in a thermal container containing sterile saline solution (NaCl 0.9%) at 37°C supplemented with penicillin (100 μg /ml) and streptomycin (100 IU/ml). The contents of follicles of 2–6 mm in diameter
47 47

on the ovarian surface were aspired with a 10 ml syringe equipped with an 21-gauge needle and collected a Petri dishes containing harvest medium. Oocytes with a uniform ooplasm and compact cumulus cell mass were washed 2 times with harvest medium and then placed in 30 μl droplets of maturation medium containing the various lutein concentrations. All the droplets were covered in paraffin oil and incubated for 45 hours at 37°C in an atmosphere with 5% CO2.
3.1.3. Maturation assessment Cumulus oocyte complexes (COCs) were evaluated using an Olympus inverted phase contrast microscope, in order to assess cumulus oophorus expansion and/or the presence of the first polar body. Cumulus expansion was assessed by a subjective scoring method (Down, 1998). Each group was compared to the control in order to establish whether any differences existed between the degrees of cumulus expansion and if they were significant. After cumulus expansion assessment the COCs were transferred to PBS with 5 mg/ml bovine serum albumin (BSA) and mechanically denuded using a micropipette. Then they were transferred to PBS containing 1 µg/ml 3’, 6’ fluorescein diacetate (FDA), 50 µg/ml propidium iodide (PI), and 20 µg/ml Hoechst 33258, incubated for 15 minutes and viewed under ultraviolet illumination with an Olympus inverted microscope. Live oocytes appeared green (FDA positive), and their chromosomes were labelled with the Hoechst 33342 being blue under UV light. The cytoplasm of dead oocytes was FDA negative and they were stained with PI, thus appearing red. Glutathione concentration in oocytes was determined by the DTNB-GSSG reductase recycling assay (Anderson, 1985). Cumulus cells were removed and oocytes were stored in 5 µl of
48 48

1.25 M phosphoric acid at -80°C. Briefly, 175 µl of 0.33 mg/ml NADPH in assay buffer (100 mM sodium phosphate buffer containing 1 mM EDTA, pH 7.2), 25 µl of 6 mM 5,5'-dithiobis-(2-nitrobenzoic acid) in assay buffer, 45 µl of water and 2.5 µl sample were added into the microfuge tube and vortexed. In the end 2.5 µl of 250 IU/ml glutathione reductase was added to initiate the reaction. The formation of 5-thio-2-nitrobenzoic acid was followed continuously with a Synergy HT Multi-Mode Microplate Reader (Biotek) for 10 min with a reading recorded every 50s. Standards (250, 125, 62.5, 31.25, 15.5, 7.75 and 4 pmol) of glutathione and a sample blank lacking glutathione were also assayed. For all the experiments the differences between treatments were analyzed by the analysis of variance and interpreted using the Newman-Keuls method. For all comparisons, the values were considered statistically significant when p < 0.05.
3.2. IN VITRO FERTILIZATION AND EMBRYO CULTURE Spermatozoa were capacitated in Tyrode by centrifugation at 800 g for 10 minutes, 3 times and the SDM 5 photometer (Minitüb) was employed to establish concentration. After cumulus expansion assessment the oocytes were transferred to TALP and mechanically denuded using a micropipette. Afterwards they were placed in 30 µl droplets and the sperm cells were added. The concentration for fertilization was 1 x 106 spermatozoa/ml and it was computed according to the following formula: volume(µl) = 30 x 1 x 106 /the concentration of capacitated spermatozoa. After the sperm cells were added the droplets were covered with paraffin oil to
49 49

prevent media evaporation and incubated at 37°C and 5 % CO2 in air. After 16-18 hours the spermatozoa were removed mechanically using a micropipette and the presumed zygotes were cultured in NCSU-23 (Petters and Wells, 1993) droplets supplemented with α-tocopherol (5, 10, 20, 40 and 80 µM), ascorbic acid (50, 150, 250, 500 and 750 µM) or lutein (2.5, 4, 5, 8, and 10 µM) and covered with mineral oil. Embryo development was assessed at 48 and 120 hours after fertilization. In another experiment embryos were cultured for 120 hours in TALP supplemented with α-tocopherol or lutein. At 48 and 120 hours the number of embryos that had developed to the 2 cells, 4-8 cells and morula stages was counted and compared to the control, and the differences analyzed using the analysis of variance and interpreted using the Newman-Keuls method. For all comparisons, the values were considered statistically significant when p < 0.05.
CHAPTER 4. RESULTS AND DISSCUSION The oocyte maturation experiments employed 1278 ovaries. Oocytes cultured in α-tocopherol supplemented medium were collected form 564 ovaries at a rate of 8.85 oocytes/ovary. A number of 395 ovaries were used to collect oocytes that would be cultured in ascorbic acid supplemented medium at a rate of 9.74 oocytes/ovary. For the experiment involving lutein oocytes were collected from 319 ovaries at a rate of 10.86 oocytes/ovary.
50 50
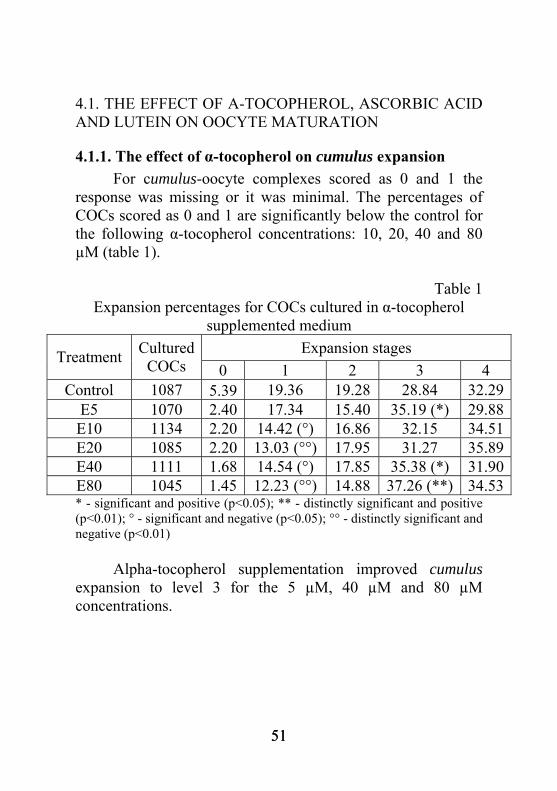
4.1. THE EFFECT OF Α-TOCOPHEROL, ASCORBIC ACID AND LUTEIN ON OOCYTE MATURATION
4.1.1. The effect of α-tocopherol on cumulus expansion For cumulus-oocyte complexes scored as 0 and 1 the response was missing or it was minimal. The percentages of COCs scored as 0 and 1 are significantly below the control for the following α-tocopherol concentrations: 10, 20, 40 and 80 µM (table 1).
Table 1 Expansion percentages for COCs cultured in α-tocopherol
supplemented medium Expansion stages Treatment Cultured
COCs 0 1 2 3 4 Control 1087 5.39 19.36 19.28 28.84 32.29
E5 1070 2.40 17.34 15.40 35.19 (*) 29.88 E10 1134 2.20 14.42 (°) 16.86 32.15 34.51 E20 1085 2.20 13.03 (°°) 17.95 31.27 35.89 E40 1111 1.68 14.54 (°) 17.85 35.38 (*) 31.90 E80 1045 1.45 12.23 (°°) 14.88 37.26 (**) 34.53 * - significant and positive (p<0.05); ** - distinctly significant and positive (p<0.01); ° - significant and negative (p<0.05); °° - distinctly significant and negative (p<0.01) Alpha-tocopherol supplementation improved cumulus expansion to level 3 for the 5 µM, 40 µM and 80 µM concentrations.
51 51
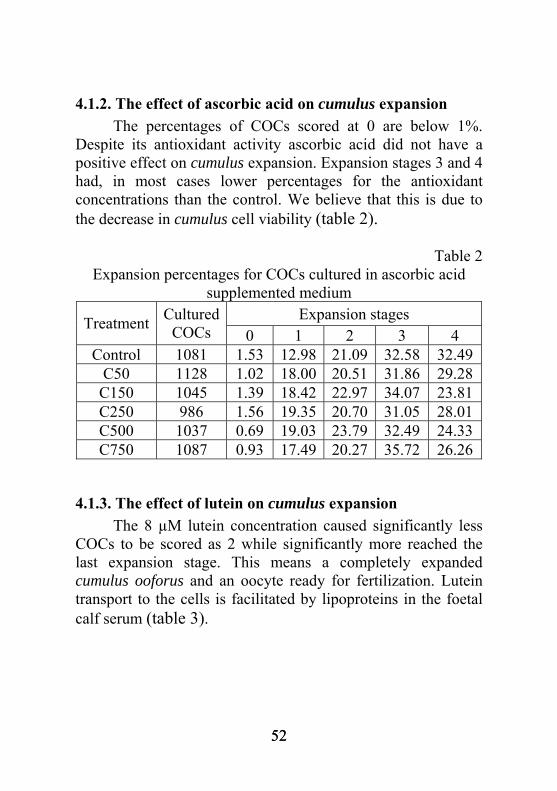
4.1.2. The effect of ascorbic acid on cumulus expansion The percentages of COCs scored at 0 are below 1%. Despite its antioxidant activity ascorbic acid did not have a positive effect on cumulus expansion. Expansion stages 3 and 4 had, in most cases lower percentages for the antioxidant concentrations than the control. We believe that this is due to the decrease in cumulus cell viability (table 2).
Table 2 Expansion percentages for COCs cultured in ascorbic acid
supplemented medium Expansion stages Treatment Cultured
COCs 0 1 2 3 4 Control 1081 1.53 12.98 21.09 32.58 32.49
C50 1128 1.02 18.00 20.51 31.86 29.28 C150 1045 1.39 18.42 22.97 34.07 23.81 C250 986 1.56 19.35 20.70 31.05 28.01 C500 1037 0.69 19.03 23.79 32.49 24.33 C750 1087 0.93 17.49 20.27 35.72 26.26
4.1.3. The effect of lutein on cumulus expansion The 8 µM lutein concentration caused significantly less COCs to be scored as 2 while significantly more reached the last expansion stage. This means a completely expanded cumulus ooforus and an oocyte ready for fertilization. Lutein transport to the cells is facilitated by lipoproteins in the foetal calf serum (table 3).
52 52
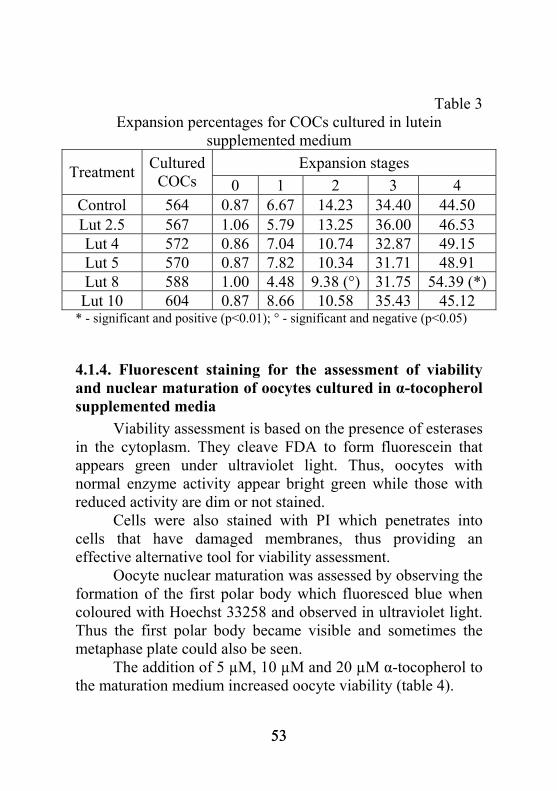
Table 3 Expansion percentages for COCs cultured in lutein
supplemented medium Expansion stages Treatment Cultured
COCs 0 1 2 3 4 Control 564 0.87 6.67 14.23 34.40 44.50 Lut 2.5 567 1.06 5.79 13.25 36.00 46.53 Lut 4 572 0.86 7.04 10.74 32.87 49.15 Lut 5 570 0.87 7.82 10.34 31.71 48.91 Lut 8 588 1.00 4.48 9.38 (°) 31.75 54.39 (*) Lut 10 604 0.87 8.66 10.58 35.43 45.12
* - significant and positive (p<0.01); ° - significant and negative (p<0.05)
4.1.4. Fluorescent staining for the assessment of viability and nuclear maturation of oocytes cultured in α-tocopherol supplemented media Viability assessment is based on the presence of esterases in the cytoplasm. They cleave FDA to form fluorescein that appears green under ultraviolet light. Thus, oocytes with normal enzyme activity appear bright green while those with reduced activity are dim or not stained. Cells were also stained with PI which penetrates into cells that have damaged membranes, thus providing an effective alternative tool for viability assessment. Oocyte nuclear maturation was assessed by observing the formation of the first polar body which fluoresced blue when coloured with Hoechst 33258 and observed in ultraviolet light. Thus the first polar body became visible and sometimes the metaphase plate could also be seen. The addition of 5 µM, 10 µM and 20 µM α-tocopherol to the maturation medium increased oocyte viability (table 4).
53 53
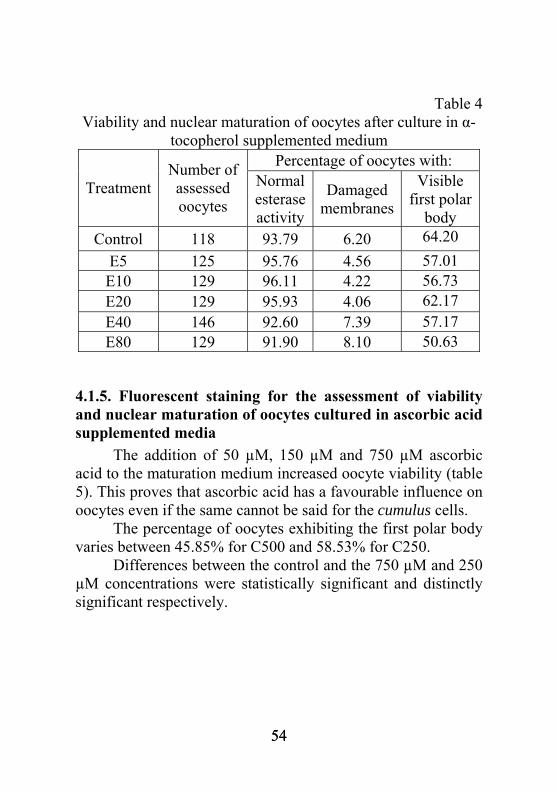
Table 4 Viability and nuclear maturation of oocytes after culture in α-
tocopherol supplemented medium Percentage of oocytes with:
Treatment Number of assessed oocytes
Normal esterase activity
Damaged membranes
Visible first polar
body Control 118 93.79 6.20 64.20
E5 125 95.76 4.56 57.01 E10 129 96.11 4.22 56.73 E20 129 95.93 4.06 62.17 E40 146 92.60 7.39 57.17 E80 129 91.90 8.10 50.63
4.1.5. Fluorescent staining for the assessment of viability and nuclear maturation of oocytes cultured in ascorbic acid supplemented media The addition of 50 µM, 150 µM and 750 µM ascorbic acid to the maturation medium increased oocyte viability (table 5). This proves that ascorbic acid has a favourable influence on oocytes even if the same cannot be said for the cumulus cells. The percentage of oocytes exhibiting the first polar body varies between 45.85% for C500 and 58.53% for C250. Differences between the control and the 750 µM and 250 µM concentrations were statistically significant and distinctly significant respectively.
54 54
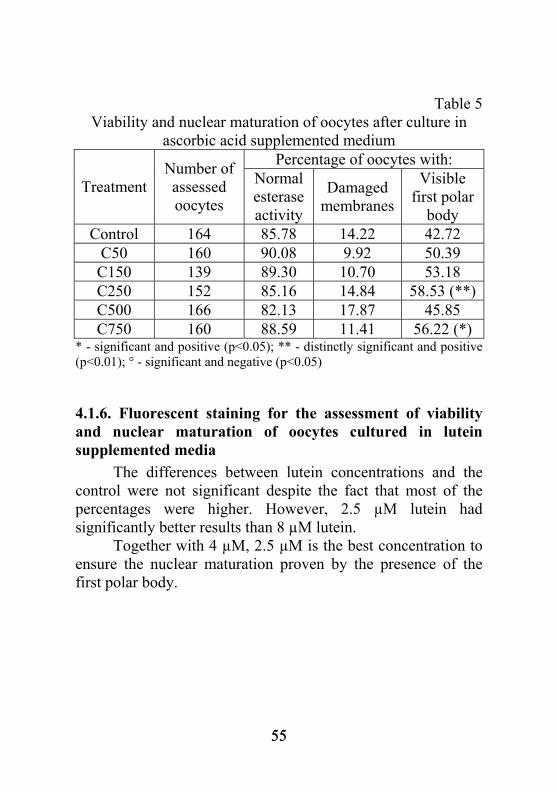
Table 5 Viability and nuclear maturation of oocytes after culture in
ascorbic acid supplemented medium Percentage of oocytes with:
Treatment Number of assessed oocytes
Normal esterase activity
Damaged membranes
Visible first polar
body Control 164 85.78 14.22 42.72
C50 160 90.08 9.92 50.39 C150 139 89.30 10.70 53.18 C250 152 85.16 14.84 58.53 (**)C500 166 82.13 17.87 45.85 C750 160 88.59 11.41 56.22 (*)
* - significant and positive (p<0.05); ** - distinctly significant and positive (p<0.01); ° - significant and negative (p<0.05)
4.1.6. Fluorescent staining for the assessment of viability and nuclear maturation of oocytes cultured in lutein supplemented media The differences between lutein concentrations and the control were not significant despite the fact that most of the percentages were higher. However, 2.5 µM lutein had significantly better results than 8 µM lutein. Together with 4 µM, 2.5 µM is the best concentration to ensure the nuclear maturation proven by the presence of the first polar body.
55 55
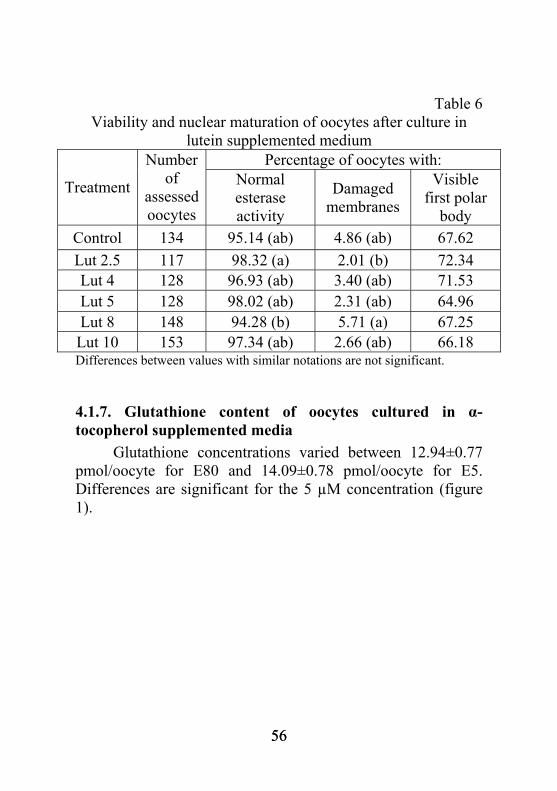
Table 6 Viability and nuclear maturation of oocytes after culture in
lutein supplemented medium Percentage of oocytes with:
Treatment
Number of
assessed oocytes
Normal esterase activity
Damaged membranes
Visible first polar
body Control 134 95.14 (ab) 4.86 (ab) 67.62 Lut 2.5 117 98.32 (a) 2.01 (b) 72.34 Lut 4 128 96.93 (ab) 3.40 (ab) 71.53 Lut 5 128 98.02 (ab) 2.31 (ab) 64.96 Lut 8 148 94.28 (b) 5.71 (a) 67.25 Lut 10 153 97.34 (ab) 2.66 (ab) 66.18 Differences between values with similar notations are not significant.
4.1.7. Glutathione content of oocytes cultured in α-tocopherol supplemented media Glutathione concentrations varied between 12.94±0.77 pmol/oocyte for E80 and 14.09±0.78 pmol/oocyte for E5. Differences are significant for the 5 µM concentration (figure 1).
56 56

pmoli/ovocit 13,19 14,09 13,53 13,50 13,27 12,94
M E5 E10 E20 E40 E80
*
Fig. 1. Glutathione concentration for oocytes cultured in α-
tocopherol supplemented media If the results are analyzed together with those from previous test it becomes apparent that 5 µM α-tocopherol has a beneficial effect on oocyte metabolism leading to increased viability and a higher glutathione concentration. The concentrations are within the limits for mature oocytes.
4.1.8. Glutathione content of oocytes cultured in ascorbic acid supplemented media The differences were significantly greater than the control for C750 and significantly lower for C150 (figure 2). The 750 µM ascorbic acid concentration leads to a better cytoplasmic and nuclear maturation. Although it has a detrimental effect on cumulus expansion, media supplementation with ascorbic acid had a beneficial influence
57 57
of glutathione concentration and oocyte ability to reach the second metaphase.
pmoli/ovocit 10,90 11,22 9,24 11,38 12,27 12,86
M C50 C150 C250 C500 C750
*
°
Fig. 2. Glutathione concentration for oocytes cultured in
ascorbic acid supplemented media
4.1.9. Glutathione content of oocytes cultured in lutein supplemented media Control oocytes had the highest glutathione content, 12.93±0.58 pmol/ooocyte (figure 3). It is significantly higher than Lut 2.5, Lut 5 and Lut 10. Despite this situation, values for all the lutein concentrations are characteristic for mature oocytes as established by Abeydeera et al (1999) and Cui et al (2009).
58 58
pmoli/ovocit 12,93 10,67 11,82 11,60 11,93 11,63
M Lut 2,5 Lut 4 Lut 5 Lut 8 Lut 10
°°° °
Fig. 3. Glutathione concentration for oocytes cultured in lutein supplemented media
4.2. THE EFFECT OF Α-TOCOPHEROL, ASCORBIC ACID AND LUTEIN ON IN VITRO EMBRYO DEVELOPMENT A number of 5379 oocytes were used for in vitro fertilization and embryo culture experiments. In vitro embryo development is slower in vitro than in vivo and sometimes it becomes impossible to surpass certain checkpoints. The main motives have to do with a low glutathione concentration, oxidative stress, delays in genome activation and unsuitable culture conditions. That is why we decided to assess embryo development at 120 hours.
59 59
4.2.1. Embryo development in α-tocopherol supplemented TALP medium, 120 hours after fertilization A number of 149 embryos were cultured in α-tocopherol supplemented TALP medium. Embryos resulted at a rate of 12.87%. The percentages of embryos at the 2 cell stage were lower in α-tocopherol supplemented medium than for the control (table 7).
Table 7 Percentages of embryos at different cleavage stages after
culture in TALP medium supplemented with α-tocopherol Percentages of embryos at each
development stage Treatment Number of oocytes 2 cells 4-8 cells Morula
Control 131 46.67 32.78 20.56 (ab) E5 221 33.73 51.79 14.48 (b) E10 149 41.11 41.11 17.78 (ab) E20 223 17.33 (°) 51.85 30.82 (ab) E40 207 28.11 44.44 27.78 (ab) E80 226 26.99 36.21 36.80 (a)
° - significant and negative (p<0.05). Differences between values with similar notations are not significant Among the 5 concentrations 80 µM α-tocopherol was the most suitable for embryo development to the morula stage, although it was not significantly better than the control.
60 60
4.2.2. Embryo development in lutein supplemented TALP medium, 120 hours after fertilization After fertilization a number of 118 embryos developed, at a rate of 14.78%. The percentages of embryos in the 2 cell stage were greater for the lutein concentrations than the control, with the exception of Lut 5 (table 8).
Table 8 Percentages of embryos at different cleavage stages after
culture in TALP medium supplemented with lutein Percentages of embryos at each
development stage Treatment Number of oocytes 2 cells 4-8 cells Morula
Control 134 50.33 50.33 0.00 Lut 2.5 171 66.67 33.67 0.00 Lut 4 141 61.27 38.73 0.00 Lut 5 135 31.37 34.70 33.93 (***) Lut 8 134 52.38 42.86 5.43 Lut 10 120 50.57 38.75 11.02 (*)
* - significant and positive (p<0.05); *** - very significant and positive (p<0.001) The percentages of embryos in the 4-8 cell stage for the lutein concentrations are bellows those for the control. The first two lutein concentrations and the control did not support embryo development to the morula stage. However, the differences for Lut 5 and Lut 10 were significant and very significant, respectively.
61 61
4.2.3. Embryo development in medium NCSU-23 supplemented with α-tocopherol, 48 hours after fertilization Given the diversity of development stages at 120 hours, we deemed necessary an additional evaluation at 48 hours. Thus, it became possible to observe embryo development dynamics. For the 2 cell stage, differences between the control and lutein concentrations are significantly lower for three of the five concentrations, namely 5 µM, 20 µM and 40 µM (table 9).
Table 9 Percentages of embryos at different cleavage stages, at 48
hours after fertilization and culture in α-tocopherol supplemented NCSU-23
Percentages of embryos at each development stage Treatment Number
of oocytes
Cultured embryos
(%) 2 cells 4-8 cells Morula Control 186 50.07 38.08 29.36 32.56 (abc)
E5 219 51.17 22.52 (°) 35.80 41.68 (ab) E10 153 56.69 27.81 38.40 (*) 33.78 (abc) E20 204 43.64 24.00 (°) 48.56 (***) 27.44 (c) E40 206 55.45 18.22 (°) 38.98 (*) 42.80 (a) E80 228 48.51 28.84 41.96 (**) 29.20 (b)
* - significant and positive (p<0.05); ** - distinctly significant and positive (p<0.01); *** - very significant and positive (p<0.001); ° - significant and negative (p<0.05); Differences between values with similar notations are not significant. For the next stage percentages are greater when embryos were cultured in α-tocopherol supplemented media than the
62 62
control. The differences are significantly higher for 10 µM and 40 µM, distinctly significant for 80 µM and very significant for 20 µM α-tocopherol. Among the 5 concentrations 40 µM and 5 µM α-tocopherol were the most suitable for embryo development to the morula stage, although they were not significantly better than the control.
4.2.4. Embryo development in medium NCSU-23 supplemented with ascorbic acid, 48 hours after fertilization
Embryos developed from fertilized oocytes at a rate of 49.18%. The embryo yield (table 10) for most ascorbic acid concentrations was higher than the control when the 2 cell and 4-8 cell stages were concerned.
Table 10 Percentages of embryos at different cleavage stages, at 48
hours after fertilization and culture in ascorbic acid supplemented NCSU-23
Percentages of embryos at each development stage Treatment
Number of
oocytes
Cultured embryos
(%) 2 cells 4-8 cells Morula Control 210 61.20 39.70 32.22 28.09
C50 215 48.27 35.06 33.19 31.75 C150 189 57.09 39.65 36.27 24.08 C250 202 53.67 50.35 29.10 20.55 C500 212 42.09 48.19 35.57 16.24 C750 192 35.46 46.32 32.19 21.49
63 63
Both the percentages of embryos that had reached the morula stage and total number of embryos were lower than the control.
4.2.5. Embryo development in medium NCSU-23 supplemented with lutein, 48 hours after fertilization
Embryos developed from fertilized oocytes at a rate of 49.18%. The percentages of embryos in the 2 cell stage were significantly higher than the control for Lut 4 and Lut 8 (table 11).
Table 11 Percentages of embryos at different cleavage stages, at 48 hours after fertilization and culture in lutein supplemented
NCSU-23 Percentages of embryos at each
development stage Treatment Number
of oocytes
Cultured embryos
(%) 2 cells 4-8 cells Morula Control 167 58.22 25.83 56.48 17.69 Lut 2.5 131 46.71 36.20 33.51 (°) 30.29 (**) Lut 4 161 44.85 45.63 (*) 45.54 9.16 Lut 5 194 48.26 41.13 45.97 12.90 Lut 8 168 55.46 53.85 (**) 24.15 (°°) 22.01 Lut 10 175 52.93 36.56 45.01 18.43
* - significant and positive (p<0.05); ** - distinctly significant and positive (p<0.01); ° - significant and negative (p<0.05); °° - distinctly significant and negative (p<0.01)
64 64
After the results were analyzed it also became apparent that the differences between control and Lut 2.5 were significant and negative while those between control and Lut 8 were distinctly significant and negative. This indicates a better embryo development in medium supplemented with 2.5, 8 or 10 µM lutein. This is strengthened by the significant differences between control and Lut 2.5 when the morula stage is concerned.
4.2.6. Embryo development in medium NCSU-23 supplemented with α-tocopherol, 120 hours after fertilization Despite the fact that the percentages for E40 and E 80 are significantly lower than the control there still are numerous embryos in the 2 cell stage (table 12).
Table 12 Percentages of embryos at different cleavage stages, at 48
hours after fertilization and culture in α-tocopherol supplemented NCSU-23
Percentages of embryos at each development stage Treatment Number of
oocytes 2 cells 4-8 cells Morula Control 203 29.55 38.53 31.93
E5 216 28.97 29.96 41.06 E10 177 33.29 30.54 36.16 E20 217 25.83 30.85 43.32 E40 218 15.99 (°°) 33.39 50.61 (*)E80 197 18.73 (°) 41.53 39.74
* - significant and positive (p<0.05); ° - significant and negative (p<0.05); °° - distinctly significant and negative (p<0.01)
65 65
The percentage of embryos to have reached the morula stage is greater for the 40 µM concentration than the control. These results show that α-tocopherol has a beneficial influence on embryo development
4.2.7. Embryo development in medium NCSU-23 supplemented with ascorbic acid, 120 hours after fertilization The percentages of embryos for the antioxidant supplemented media are higher than the control for the 2 cell and 4-8 cell stages but not the morula (table 13). Most embryos should be in later development stage such as morula.
Table 13 Percentages of embryos at different cleavage stages, at 48
hours after fertilization and culture in ascorbic acid supplemented NCSU-23
Percentages of embryos at each development stage Treatment
Number of
oocytes 2 cells 4-8 cells Morula Control 191 41.46 33.34 25.20
C50 201 31.49 37.28 31.23 C150 194 31.41 41.45 27.14 C250 202 46.48 32.14 21.39 C500 217 37.18 38.86 23.96 C750 192 41.30 34.81 23.89
66 66
4.2.8. Embryo development in medium NCSU-23 supplemented with lutein, 120 hours after fertilization Differences between the control and Lut 4 were significant and positive for the 2 cell stage. Among the 5 lutein concentrations 2.5 µM proved to be the best and ensured the development to the morula stage of a higher percentage of embryos.
Table 14 Procentele de embrioni cultivaţi în mediu NCSU-23 suplimentat cu luteină, la 120 ore după fecundaţie
Percentages of embryos at each development stage Treatment Number of
oocytes 2 cells 4-8 cells Morula Control 167 29.65 37.55 25.73 Lut 2.5 131 34.51 35.05 30.44 Lut 4 161 48.34 (*) 32.74 18.93 Lut 5 197 36.91 42.37 20.72 Lut 8 168 38.20 43.58 18.22 Lut 10 175 28.67 50.35 20.98
* - significant and positive (p<0.05)
4.2.9. Comparison between NCSU-23 and TALP as antioxidant supplemented media for in vitro embryo culture Embryos cultured in TALP medium develop to lower stages (2 cells, 4-8 cells) while NCSU-23 increases the percentage that reached the morula stage. More embryos develop in NCSU than TALP irrespective of the antioxidant supplementation.
67 67

CHAPTER 5. CONCLUSIONS 5.1. The effect of α-tocopherol, ascorbic acid and lutein on oocyte maturation:
6. Oocytes maturated in α-tocopherol supplemnted media (5 µM) have a higher glutathione content, better cumulus expansion and are more likely to be viable. Supplementation of the maturation medium with α-tocopherol leads to an increase in viability and cytoplasm maturation at the expense of nuclear maturation.
7. Ascorbic acid supplementation (750 µM) results in increased cytoplasm and nuclear maturation, with negative effects on cumulus expansion.
8. Lutein supplementation leads to an increase in cumulus expansion and percentage of viable oocytes while cytoplasmic maturation suffered ill effects.
9. Differences between antioxidants are the effect of their chemical structure and properties. While α-tocopherol and lutein act in the lipid soluble compartment of the cell, ascorbic acid is the foremost water soluble antioxidant. This confirms the hypothesis that together with α-tocopherol and ascorbic acid, carotenoids are part of an antioxidant network that functions in the cell.
10. Thorough maturation assessment requires various techniques to complete the subjective but fast scoring of the cumulu oophorus. 5.2. The effect of α-tocopherol, ascorbic acid and lutein on embryo development in TALP and NCSU-23 media:
6. TALP supplementation with α-tocopherol and lead to an increase in the number of embryos that at 120 hours reached the 4-8 cell and morula stages.
68 68

7. At 48 hours after fertilization it becomes apparent that α-tocopherol supplemented NCSU-23 increases embryo development to the 4-8 cell (10, 20, 40, 80 µM) and morula (5, 40, 80 µM) stages. At 120 hours even more embryos have reached the morula stage.
8. Ascorbic acid addition to NCSU-23 has a detrimental effect on embryo development. Both at 48 and 120 hours most embryos were at the 2 cell and 4-8 cell stages.
9. Lutein (2.5 µM) supplementation has a beneficial effect on embryo development to the morula stage. This is apparent at 48 hours as well as 120 hours.
10. More embryos develop in NCSU-23 than TALP. The addition of α-tocopherol and lutein cannot compensate for the inadequacies that become apparent for TALP.
69 69


















