
Cod proiect: PN-II-RU-TE-2014-4-2093Tabel 1.1. Gradul de realizare a obiectivelor și...
43
1 Cod proiect: PN-II-RU-TE-2014-4-2093 ISTORIA EVOLUTIVĂ A UNUI GRUP DE SPECII DE COSAȘI (ISOPHYA CAMPTOXYPHA) EXTRAPOLATĂ UTILIZÂND METODE ACUSTICE, MOLECULARE ȘI DE MODELARE A NIȘEI ECOLOGICE Contract nr. 116 / 01.10.2015 Raport științific privind implementarea proiectului în perioada octombrie-decembrie 2015 și ianuarie-decembrie 2016 Etapa 1 (octombrie-decembrie 2015) Grupul de specii “Isophya camptoxypha”: evaluarea cunoștințelor despre distribuția, comunicarea acustică și genetica speciilor Etapa 2 (ianuarie-decembrie 2016) Studiul distribuției potențiale și a relațiilor filogenetice dintre speciile grupului “Isophya camptoxypha” Echipa proiectului Dr. Ionuț Ștefan Iorgu, director de proiect Dr. Elena Iulia Iorgu, cercetător postdoctoral Dr. Tiberiu Constantin Sahlean, cercetător postdoctoral Liviu Aurel Moscaliuc, student doctorand Daniela Mirela Dragoș, student doctorand Muzeul Național de Istorie Naturală "Grigore Antipa", București
Transcript of Cod proiect: PN-II-RU-TE-2014-4-2093Tabel 1.1. Gradul de realizare a obiectivelor și...

1
Cod proiect: PN-II-RU-TE-2014-4-2093
ISTORIA EVOLUTIVĂ A UNUI GRUP DE SPECII DE COSAȘI
(ISOPHYA CAMPTOXYPHA) EXTRAPOLATĂ UTILIZÂND METODE
ACUSTICE, MOLECULARE ȘI DE MODELARE A NIȘEI
ECOLOGICE
Contract nr. 116 / 01.10.2015
Raport științific
privind implementarea proiectului în perioada octombrie-decembrie 2015 și
ianuarie-decembrie 2016
Etapa 1 (octombrie-decembrie 2015)
Grupul de specii “Isophya camptoxypha”: evaluarea cunoștințelor despre
distribuția, comunicarea acustică și genetica speciilor
Etapa 2 (ianuarie-decembrie 2016)
Studiul distribuției potențiale și a relațiilor filogenetice dintre speciile
grupului “Isophya camptoxypha”
Echipa proiectului
Dr. Ionuț Ștefan Iorgu, director de proiect
Dr. Elena Iulia Iorgu, cercetător postdoctoral
Dr. Tiberiu Constantin Sahlean, cercetător postdoctoral
Liviu Aurel Moscaliuc, student doctorand
Daniela Mirela Dragoș, student doctorand
Muzeul Național de Istorie Naturală "Grigore Antipa", București

2
Etapa 1: Grupul de specii “Isophya camptoxypha”: evaluarea cunoștințelor
despre distribuția, comunicarea acustică și genetica speciilor
I. Rezumatul Etapei
Descrierea și scopul proiectului
Cuprinzând aproximativ 90 de specii, Isophya Brunner von Wattenwyl (1878) este
unul dintre cele mai numeroase genuri palearctice din ordinul Orthoptera. Aceste specii sunt
insecte brahiptere, cu morfologie aproape uniformă; contrar acesteia, structura ritmică a
semnalelor acustice ale masculilor prezintă diferențe clare interspecifice. Pentru a forma
cuplul, masculul și femela formează un duet acustic, componentă semnificativă a sistemului
de recunoaștere al partenerului la acești cosași. Grupul de specii Isophya camptoxypha se
definește ca fiind un complex alcătuit din șapte specii (Isophya camptoxypha, I. ciucasi, I.
nagyi, I. sicula, I. posthumoidalis, I. dochia, I. fatrensis), cu putere de dispersie geografică
limitată și restricționate la câteva regiuni montane din Carpați. Scopul acestui proiect este
investigarea relațiilor filogenetice și filogeografice între speciile grupului I. camptoxypha,
clarificând taxonomia și istoria lor evolutivă, sintetizate din studiul bioacusticii, biologiei
moleculare și modelarea nișei ecologice.
Etapa 1 presupune evaluarea cunoștințelor despre distribuția, comunicarea acustică și
genetica speciilor din grupul Isophya camptoxypha. În cadrul acestei etape, studiile noastre s-
au canalizat pe trei direcții:
- alcătuirea unor baze de date cu informații privind structura oscilografică a sunetului și
distribuția speciilor din grupul Isophya camptoxypha.
- optimizarea unui protocol de amplificare markeri ADN nucleari și mitocondriali,
utilizabil pentru taxonii analizați.
- diseminarea rezultatelor pe scară largă prin comunicarea sau prin publicarea națională
/ internațională a rezultatelor. Acest obiectiv s-a realizat prin participarea la o manifestare
științifică internațională.
Obiectivele și activitățile etapei 1 (2015)
1. Evaluarea comunicării acustice la speciile din grupul Isophya camptoxypha.
1.1. Recenzia exhaustivă a literaturii pentru a extrage elementele de acustică din
lucrările publicate și crearea unei baze de date cu informații.
1.2. Analiza discriminantă a sunetelor: comparație oscilografică intra- și interspecifică
a speciilor din grupul Isophya camptoxypha.
2. Studiul relațiilor filogenetice dintre speciile grupului Isophya camptoxypha, folosind
markeri moleculari.
2.1. Extracție de ADN genomic din indivizi aparținând speciilor din grupul Isophya
camptoxypha.
2.2. Optimizarea reacțiilor PCR a unor markeri moleculari (gene mitocondriale sau
nucleare), necesari în analiza filogenetică.

3
3. Evaluarea distribuției potențiale (anvelopei bioclimatice) actuale și trecute pentru
speciile din grupul Isophya camptoxypha.
3.1. Recenzia exhaustivă a literaturii pentru a extrage punctele de ocurență din
lucrările publicate și crearea unei baze de date cu informații.
3.2. Georeferențierea punctelor de ocurență care nu au informație spațială, folosind un
sistem GIS și hărți de referință (aeriene, hărți topografice actuale și anterioare).
4. Diseminarea rezultatelor.
4.1. Crearea site-ului web al proiectului.
4.2. Participare la o manifestare științifică.
II. Descrierea științifică și tehnică a etapei
Tabel 1.1. Gradul de realizare a obiectivelor și activităților
Obiectiv / Activitate Grad de
realizare
1. Evaluarea comunicării acustice la speciile din grupul Isophya camptoxypha. 100%
1.1. Recenzia exhaustivă a literaturii pentru a extrage elementele de acustică din lucrările
publicate și crearea unei baze de date cu informații. 100%
1.2. Analiza discriminantă a sunetelor: comparație oscilografică intra- și interspecifică a
speciilor din grupul Isophya camptoxypha. 100%
2. Studiul relațiilor filogenetice dintre speciile grupului Isophya camptoxypha, folosind
markeri moleculari. 100%
2.1. Extracție de ADN genomic din indivizi aparținând speciilor din grupul Isophya
camptoxypha. 100%
2.2. Optimizarea reacțiilor PCR a unor markeri moleculari (gene mitocondriale sau nucleare),
necesari în analiza filogenetică. 100%
3. Evaluarea distribuției potențiale (anvelopei bioclimatice) actuale și trecute pentru
speciile din grupul Isophya camptoxypha. 100%
3.1. Recenzia exhaustivă a literaturii pentru a extrage punctele de ocurență din lucrările
publicate și crearea unei baze de date cu informații. 100%
3.2. Georeferențierea punctelor de ocurență care nu au informație spațială, folosind un sistem
GIS și hărți de referință (aeriene, hărți topografice actuale și anterioare). 100%
4. Diseminarea rezultatelor. 100%
4.1. Crearea site-ului web al proiectului. 100%
4.2. Participare la o manifestare științifică. 100%
4
Obiectivul 1. Evaluarea comunicării acustice la speciile din grupul Isophya
camptoxypha.
Activitatea 1.1. Recenzia exhaustivă a literaturii pentru a extrage elementele de acustică
din lucrările publicate și crearea unei baze de date cu informații.
Uniformitatea morfologică a genului Isophya face aproape imposibilă identificarea
speciilor cu distribuție simpatrică și ridică probleme asupra validității taxonomice a unor
specii alopatrice. Contrar morfologiei, structura ritmică a semnalelor acustice ale masculilor
prezintă diferențe clare, chiar și atunci când sunt comparate specii cu un înalt grad de
similaritate morfologică.
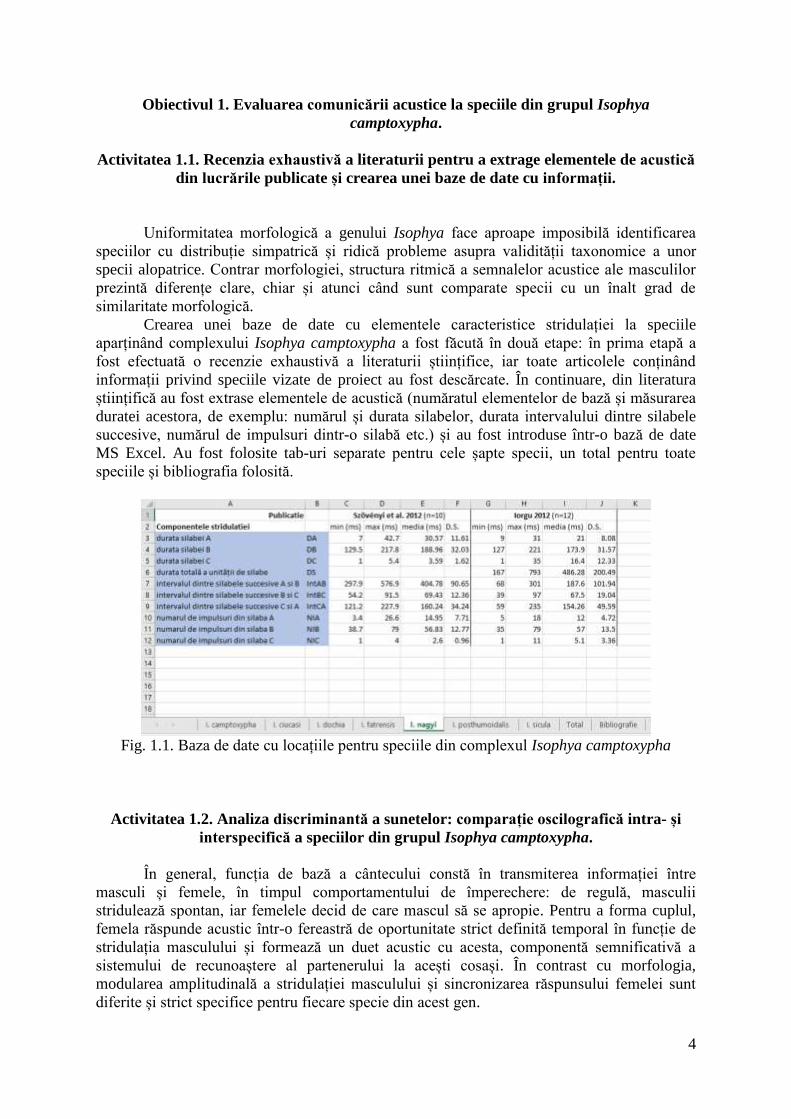
Crearea unei baze de date cu elementele caracteristice stridulației la speciile
aparținând complexului Isophya camptoxypha a fost făcută în două etape: în prima etapă a
fost efectuată o recenzie exhaustivă a literaturii științifice, iar toate articolele conținând
informații privind speciile vizate de proiect au fost descărcate. În continuare, din literatura
științifică au fost extrase elementele de acustică (număratul elementelor de bază și măsurarea
duratei acestora, de exemplu: numărul și durata silabelor, durata intervalului dintre silabele
succesive, numărul de impulsuri dintr-o silabă etc.) și au fost introduse într-o bază de date
MS Excel. Au fost folosite tab-uri separate pentru cele șapte specii, un total pentru toate
speciile și bibliografia folosită.
Fig. 1.1. Baza de date cu locațiile pentru speciile din complexul Isophya camptoxypha
Activitatea 1.2. Analiza discriminantă a sunetelor: comparație oscilografică intra- și
interspecifică a speciilor din grupul Isophya camptoxypha.
În general, funcția de bază a cântecului constă în transmiterea informației între
masculi și femele, în timpul comportamentului de împerechere: de regulă, masculii
stridulează spontan, iar femelele decid de care mascul să se apropie. Pentru a forma cuplul,
femela răspunde acustic într-o fereastră de oportunitate strict definită temporal în funcție de
stridulația masculului și formează un duet acustic cu acesta, componentă semnificativă a
sistemului de recunoaștere al partenerului la acești cosași. În contrast cu morfologia,
modularea amplitudinală a stridulației masculului și sincronizarea răspunsului femelei sunt
diferite și strict specifice pentru fiecare specie din acest gen.
5
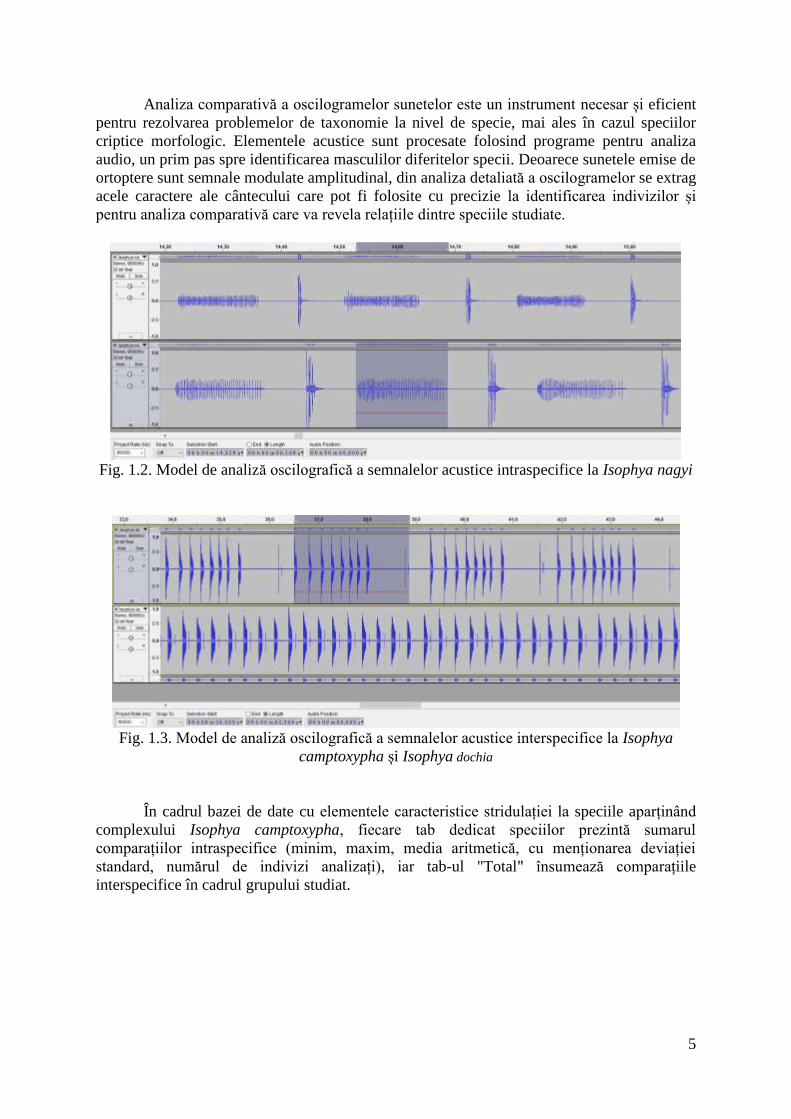
Analiza comparativă a oscilogramelor sunetelor este un instrument necesar și eficient
pentru rezolvarea problemelor de taxonomie la nivel de specie, mai ales în cazul speciilor
criptice morfologic. Elementele acustice sunt procesate folosind programe pentru analiza
audio, un prim pas spre identificarea masculilor diferitelor specii. Deoarece sunetele emise de
ortoptere sunt semnale modulate amplitudinal, din analiza detaliată a oscilogramelor se extrag
acele caractere ale cântecului care pot fi folosite cu precizie la identificarea indivizilor și
pentru analiza comparativă care va revela relațiile dintre speciile studiate.
Fig. 1.2. Model de analiză oscilografică a semnalelor acustice intraspecifice la Isophya nagyi
Fig. 1.3. Model de analiză oscilografică a semnalelor acustice interspecifice la Isophya
camptoxypha și Isophya dochia
În cadrul bazei de date cu elementele caracteristice stridulației la speciile aparținând
complexului Isophya camptoxypha, fiecare tab dedicat speciilor prezintă sumarul
comparațiilor intraspecifice (minim, maxim, media aritmetică, cu menționarea deviației
standard, numărul de indivizi analizați), iar tab-ul "Total" însumează comparațiile
interspecifice în cadrul grupului studiat.

6
Obiectivul 2. Studiul relațiilor filogenetice dintre speciile grupului Isophya
camptoxypha, folosind markeri moleculari.
Activitatea 2.1. Extracție de ADN genomic din indivizi aparținând speciilor din grupul
Isophya camptoxypha.
Colectarea probelor şi extracţia ADN-ului genomic.
Au fost colectate probe de țesut de la 212 exemplare aparținând speciilor din grupul
Isophya camptoxypha, din 49 de situri (Tabel 2). De la fiecare exemplar a fost prelevat un
picior posterior (stâng) și apoi fixat în alcool 96%. ADN-ul genomic total a fost extras din
ţesut muscular provenit de la fiecare individ în parte, urmând etapele kiturilor-ului ISOLATE
II Genomic DNA Kit (Bioline, London, UK) sau NucleoSpin® Tissue MACHEREY-
NAGEL GmbH & Co.
Principiul extracției ADN presupune 4 etape:
1. Prima etapă a extracției de ADN genomic presupune liza celulară prin incubarea probei
într-o soluţie de proteinază K şi SDS.
2. A doua etapă este legarea ADN genomic de membrana de dioxid de siliciu din coloniţele
furnizate de kituri. Această etapă este facilitată de adăugarea de săruri şi etanol. Procesul de
legare la membrană este reversibil şi specific acizilor nucleici.
3. Spălări succesive ale ADN-ului. Contaminările sunt înlăturate prin spălări succesive în
două soluţii tampon diferite.
4. Eluția ADN. ADN genomic pur este eluat la final într-o soluţie uşor alcalină sau în apă
distilată.
Concentraţia şi puritatea ADN genomic extras au fost măsurate utilizând
spectrofotometrul NanoDrop ND-1000.
Integritatea ADN genomic a fost testată ulterior prin electroforeza a 5µl în gel de
agaroză 1,5 % cu 0,5μg/ml EtBr
Tabel 1.2. Exemplarele aparținând grupului Isophya camptoxypha de la care s-a extras ADN
genomic
Nr. Crt. Taxon Sit colectare Numărul de exemplare și codul
probelor
1. I. camptoxypha Polonia 3 exemplare – ICPXPOL 1, 2, 3
2. I. camptoxypha M-rea. Agapia (NT) 4 exemplare – AG 1, 2, 3, 4
3. I. camptoxypha M-rea. Vatra Moldoviței (SV) 4 exemplare – VMO 1, 2, 3, 4
4. I. camptoxypha M-rea. Humor (SV) 4 exemplare – MHU 1, 2, 3, 4
5. I. camptoxypha M-ții. Parâng 2 exemplare – PG 1, 2
6. I. camptoxypha Cheile Râmețului 4 exemplare – CRM 1, 2, 3, 4
7. I. camptoxypha Sadova (SV) 3 exemplare – SAD 1, 2, 3
8. I. camptoxypha Vânători Neamț (NT) 1 exemplar – VNE
9. I. camptoxypha M-ții. Nemira 4 exemplare – NEM1, 2, 3, 4
10. I. camptoxypha M-ții. Țibles 2 exemplare – TIB 1, 2
11. I. camptoxypha Tg. Neamț (NT) 4 exemplare – TN 1, 2, 3, 4
12. I. camptoxypha Mărișel (M-ții. Apuseni) 3 – MAR 1, 2, 3
13. I. camptoxypha Căpâlna, 2009 8 exemplare – A 108, A 210, A 216, A
217, A 218, A 219, A 221, A 222

7
14. I. camptoxypha Căpâlna, 2011 2 exemplare – CAP 1, 2
15. I. camptoxypha M-ții. Penteleu 8 exemplare – P 1, 2, 3, 4, 5, 6, 7, 8
16. I. camptoxypha M-ții. Pietrosul Bistritei 7 exemplare – PB 1, 2, 3, 4, 5, 6, 7
17. I. camptoxypha M-ții. Buila Vânturarița 5 exemplare – B 1, 2, 3, 4, 5
18. I. camptoxypha M-ții. Rarău 11 exemplare – R 1, 2, ,3, 4, 5, 6, 7, 8,
9, 10, 11
19. I. camptoxypha M-ții. Cozia 4 exemplare – COZ 4, 5, 6, 7
20. I. camptoxypha Ciuta (BZ) 1 exemplar – CIT
21. I. camptoxypha Pasul Mestecăniș (SV) 3 exemplare – ME 1, 2, 3
22. I. camptoxypha M-ții. Giumalău (SV) 2 exemplare – GIU 1, 2
23. I. camptoxypha M-ții. Piatra Craiului 4 exemplare – PIC 1, 2, 3, 4
24. I. camptoxypha Moeciu de Jos (BV) 2 exemplare – MJ 1, 2
25. I. camptoxypha M-ții. Bucegi 6 exemplare – BC 1, 2, 3, 4, 5, 6
26. I. camptoxypha M-ții. Postăvarul 4 exemplare – PTV 1, 2, 3
27. I. camptoxypha M-ții. Piatra Mare (BV) 3 exemplare – PMA 1, 2, 3
28. I. camptoxypha M-ții. Făgăraș, vf. Negoiu 3 exemplare – NEG 1, 2, 3
29. I. camptoxypha Sebeșul de Jos (SB) 3 exemplare – SBJ 1, 2, 3
30. I. camptoxypha M-ții. Cindrel 3 exemplare – CIN 1, 2, 3
31. I. camptoxypha M-ții. Ciucaș 13 exemplare – C1, 2, 3, 4, 5, 6, 7, 8,
9, 10, 11, 12, 13
32. I. camptoxypha M-ții. Călimani 3 exemplare – CAL 1, 2, 3
33. I. posthumoidalis Teceu Mic (MM) 2 exemplare – POST 1, 2
34. I. ciucasi M-ții. Vrancei, vf. Goru 3 exemplare – GOR 1, 2, 3
35. I. ciucasi M-ții. Ciucaș 4 – ICIU 1, 2, 3, 4
36. I. dochia M-ții. Gurghiu 3 exemplare – GRG 1, 2
37. I. dochia M-ții. Ceahlău 2 exemplare – DU 1, 2
38. I. dochia M-ții. Ceahlău 33 exemplare – Ch 1-33
39. I. dochia M-ții. Bistritei, vf. Budacu 4 exemplare – BUD 1, 2, 3
40. I. fatrensis Slovacia 2 exemplare – IFRASL 9,10
41. I. nagyi M-ții. Suhard 3 exemplare – SUH 1, 2, 3
42. I. nagyi Neagra Șarului (SV) 4 exemplare – NSA 1, 2, 3, 4
43. I. posthumoidalis Polonia 2 exemplare – IPOSTPL 4, 5
44. I. sicula Băile Harghita (HR) 4 exemplare – HRB 1, 2, 3, 4
45. I. camptoxypha Valea Berii, Muntele Roșu 2 exemplare – MRO 1, 2
46. I. beybienkoi Slovacia, Slovenski Karst 3 exemplare – IBBYSL 6, 7, 8
47. I. harzi M-ții. Cozia 4 exemplare – COZ 1, 2, 3, 4
48. I. pienensis Baia Mare 1 exemplar – IPI 4
49. I. pienensis Pasul Borsec (HR) 3 exemplare – IPI 1, 2, 3

8
Activitatea 2.2. Optimizarea reacțiilor PCR a unor markeri moleculari (gene
mitocondriale sau nucleare), necesari în analiza filogenetică.
Markerii mitocondriali sunt preferaţi în anumite studii de filogenie și filogeografie la
anumite specii criptice datorită mai multor factori:
- degradarea acestora se produce mai lent decât a ADN nuclear.
- ADN mitocondrial evolueaza mai rapid decât ADN nuclear.
- ADN mitocondrial se transmite la descendenţi fără a suferi procesul de
recombinare.
Noi am ales și am optimizat reactiile de amplificare PCR la doi markeri mitocondrali
și a unui singur marker nuclear:
1. Gena COI - gena care codifică pentru subunitatea I a citocrom C oxidazei. Gena COI
este un marker molecular frecvent utilizat în studiile de filogenie, fiind o regiune înalt
conservată a genomului mitocondrial, utilizată pentru diferenţierea speciilor înrudite.
S-a amplificat fragment de ~750 pb din gena COI cu ajutorul unor primeri universali.
2. Gena 16 s - gena pentru subunitatea 16 S a ARN ribosomal. Aceasta genă este de
asemenea utilizată frecvent în studii de filogenie. S-a amplificat un fragment de
aproximativ 500 bp cu ajutorul unor primeri universali.
3. Gena pentru ITS 1 – gena pentru spacerul de transcriere intern. Este o genă nucleară
situată între gena pentru subunitatea mică 5.8 S a ADN ribozomal și gena pentru
subunitatea mare 18 S a ADN ribozomal. Acest fragment se întâlnește repetat în
andem în genom și este de asemenea folosit în studii de filogenie și filogeografie. S-a
amplificat un fragment de aproximativ 350bp cu ajutorul primerilor. Pentru reacţiile
PCR a fost utilizat T Professional Thermocycler basic (Biometra).
Componentele mixului reacţiei de amplificare pentru cei trei markeri moleculari sunt
redate în tabelul 3.
Tabel 1.3. Componentele reacţiei PCR pentru amplificarea fragmentelor de interes
Reactiv Volumul în reacţie
ADN 5
Tag Buffer (ROVALAB) 10X 5
MgCl2 (ROVALAB) 50 mM 2.5
dNTP1 mM 10
LCO1490 1μM1 μM 5
HCO2198 1μM1 μM 5
Taq DNA Polvmerase5 U/μl 1,5 U
mQsteril 17,2
Volum final 50 μl
Amplificarea s-a realizat utilizând programele PCR cu parametrii menţionaţi în
tabelul 4, 5 și 6.
Tabel 1.4. Programul PCR pentru amplificarea fragmentului de interes din gena COI
Etapă Temperatură Durată Nr. Cicluri COI
Denaturare initial 95° 2 min 1×
Denaturare 94° 30 s
5× Aliniereprimeri 45° 1 min 30 s
Elongare 72° 1 min
Denaturare 94° 30 s 35×

9
Aliniereprimeri 52° 1 min 30 s
Elongare 72° 1 min
Elongarefinală 72° 5 min 1×
Tabel 1.5. Programul PCR pentru amplificarea fragmentului de interes din gena ITS 1
Etapă Temperatură Durată Nr. Cicluri ITS 1
Denaturare initial 95° 3 min 1×
Denaturare 94° 10 s
9× Aliniereprimeri 45° 30 s
Elongare 72° 50 s
Denaturare 94° 10 s
35× Aliniereprimeri 52° 30 s
Elongare 72° 1 min
Elongarefinală 72° 5 min 1×
Tabel 1.6. Programul PCR pentru amplificarea fragmentului de interes din gena 16S
Etapă Temperatură Durată Nr. Cicluri 16s
Denaturare initial 95° 3 min 1×
Denaturare 94° 30 s
35× Aliniereprimeri 52° 30 s
Elongare 72° 30 s
Elongarefinală 72° 5 min 1×
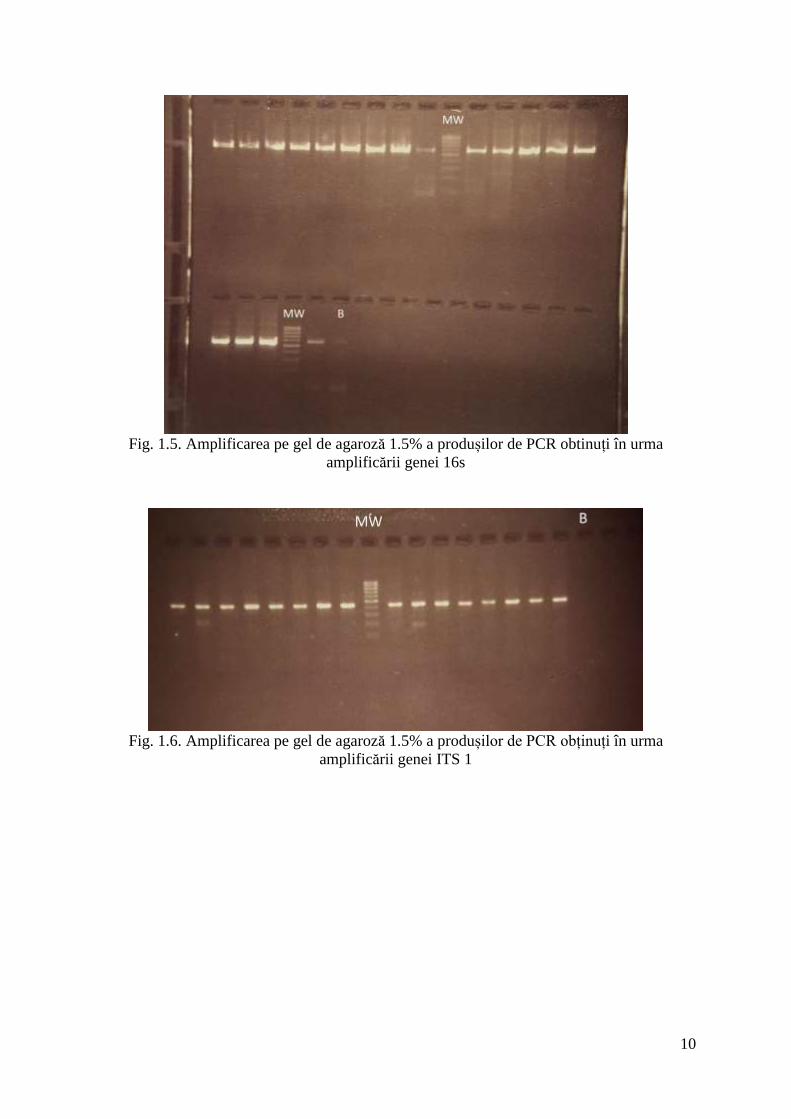
Rezultatele amplificării au fost verificate prin încărcarea şi migrarea produsului de
PCR în gel de agaroză 1.5% cu 0,5μg/ml EtBr (figurile 1.4-1.6).
Fig. 1.4. Amplificarea pe gel de agaroză 1.5% a produșilor de PCR obținuți în urma
amplificării genei COI
10
Fig. 1.5. Amplificarea pe gel de agaroză 1.5% a produșilor de PCR obtinuți în urma
amplificării genei 16s
Fig. 1.6. Amplificarea pe gel de agaroză 1.5% a produșilor de PCR obținuți în urma
amplificării genei ITS 1

11
3. Evaluarea distribuției potențiale (anvelopei bioclimatice) actuale și trecute pentru
speciile din grupul Isophya camptoxypha.
3.1. Recenzia exhaustivă a literaturii pentru a extrage punctele de ocurență din lucrările
publicate și crearea unei baze de date cu informații.
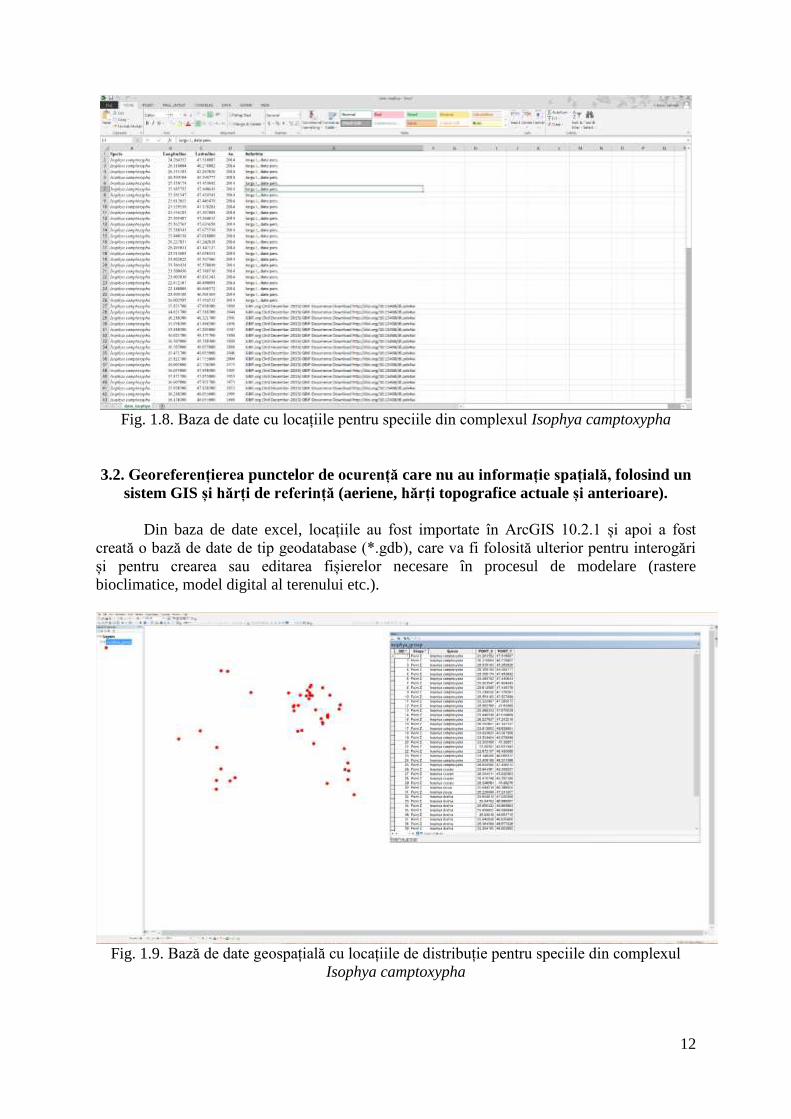
Pentru a construi baza de date cu puncte georeferențiate pentru speciile aparținând
complexului Isophya camptoxypha, în prima etapă a fost efectuată o recenzie exhaustivă a
literaturii științifice, iar toate articolele conținând informații privind speciile vizate au fost
descărcate și plasate într-un singur folder. În continuare, literatura științifică a fost organizată
folosind programul EndNote 7, în sensul că lucrările au fost introduse ca referințe într-o bază
de date EndNote (*.enl), iar articolele în format *.pdf au fost atașate acestei baze de date.
Fig. 1.7. Baza de date EndNote cu literatura științifică disponibilă pentru complexul Isophya
camptoxypha
În continuare, a fost creată o bază de date excel în care au fost introduse toate datele
de distribuție din literatura științifică, aceasta fiind apoi completată cu datele de distribuție
personale rezultate în urma deplasărilor în teren și cu datele disponibile on-line într-o locație
trasabilă (GBIF - http://www.gbif.org/). În cazul datelor din literatură pentru care lipseau
informațiile spațiale (latitudine și longitudine) au fost folosite ortofotoplanuri, harta militară
declasificată și informațiile autorului pentru a localiza spațial punctul de colectare. Dacă
autorul nu a oferit informații privind habitatul în locația de colectare, ci doar denumirea celei
mai apropiate localități, atunci punctul a fost amplasat într-un habitat favorabil speciei pentru
a fi util ulterior în procesul de modelare. În baza de date au fost incluse toate punctele din
literatura științifică, dar în procesul de modelare vor fi folosite doar punctele cu o rezoluție
mai mare de 1 km. Pentru locațiile prea generale (ex.: Munții Rarău, Munții Ciucaș)
georeferențierea nu a putut fi realizată, iar acele locații vor fi excluse din modelare.
12
Fig. 1.8. Baza de date cu locațiile pentru speciile din complexul Isophya camptoxypha
3.2. Georeferențierea punctelor de ocurență care nu au informație spațială, folosind un
sistem GIS și hărți de referință (aeriene, hărți topografice actuale și anterioare).
Din baza de date excel, locațiile au fost importate în ArcGIS 10.2.1 și apoi a fost
creată o bază de date de tip geodatabase (*.gdb), care va fi folosită ulterior pentru interogări
și pentru crearea sau editarea fișierelor necesare în procesul de modelare (rastere
bioclimatice, model digital al terenului etc.).
Fig. 1.9. Bază de date geospațială cu locațiile de distribuție pentru speciile din complexul
Isophya camptoxypha

13
4. Diseminarea rezultatelor.
Nr.
crt.
Rezultate
așteptate Rezultate livrate
1.
Crearea site-
ului web al
proiectului
Crearea site-ului web al proiectului, la adresa:
http://cercetare-antipa.ro/isophya/
2.
Participare la
o manifestare
științifică
Iorgu, I. Ş., Sahlean, T. C., Moscaliuc, L. A., Dragoș, M. & Iorgu, E. I.
- Mapping the general distribution of Isophya camptoxypha species
group (Insecta: Orthoptera) in the Romanian Carpathians and
application of IUCN Red List criteria for their future conservation.
International Zoological Congress of “Grigore Antipa” Museum,
București, 18-21 noiembrie 2015, p. 205-206
http://czga.ro/pozepagini/CZGA2015-
Book_of_abstracts(online_edition)_1.pdf

14
Etapa 2: Studiul distribuției potențiale și a relațiilor filogenetice dintre
speciile grupului “Isophya camptoxypha”
I. Rezumatul Etapei
În cadrul celei de-a doua etape, studiile noastre s-au canalizat pe următoarele direcții:
- activități de teren pentru a înregistra speciile de interes, pentru a colecta puncte de
distribuție pentru speciile țintă (și a investiga locațiile prezise de modele, dar unde specia nu a
fost găsită anterior) și pentru a colecta probe biologice. Stridulația insectelor colectate a fost
înregistrată în natură și în laborator pentru obținerea datelor privind sistemele de comunicare
acustică, înregistrarea masculilor care stridulează și a duetelor mascul-femelă.
- extracția ADN genomic de la probele colectate și amplificarea markerilor moleculari
necesari în analiza filogenetică și obținerea secvențelor ADN ale acestora. A fost realizată o
bază de date cu secvențe ADN provenind de la taxonii analizați.
- elaborarea unui model preliminar de distribuție potențială pentru speciile din
complexul “Isophya camptoxypha” folosind doi algoritmi de modelare: Maxent și Desktop
GARP. Modelele de distribuție potențială au fost recalibrate folosind date din teren, pentru a
crește acuratețea rezultatelor și transferarea modelelor pe climate din trecut. Au fost întocmite
hărți preliminare de distribuție a speciilor din grupul “Isophya camptoxypha”.
- diseminarea rezultatelor pe scară largă prin comunicarea sau prin publicarea
națională / internațională a rezultatelor. Acest obiectiv s-a realizat prin participarea la o
manifestare științifică internațională.
Obiectivele și activitățile etapei 2 (2016)
1. Evaluarea comunicării acustice la speciile din grupul “Isophya camptoxypha”.
1.1. Activități de teren pentru a înregistra speciile de interes și a colecta probe
biologice.
1.2. Înregistrarea cântecelor în natură/laborator pentru obținerea datelor privind
sistemele de comunicare acustică, înregistrarea masculilor care stridulează și a duetelor
mascul-femelă.
2. Studiul relațiilor filogenetice dintre speciile grupului “Isophya camptoxypha”, folosind
markeri moleculari.
2.1. Extracție de ADN genomic de la probele colectate.
2.2. Amplificarea markerilor moleculari necesari în analiza filogenetică și obținerea
secvențelor ADN ale acestora.
3. Evaluarea distribuției potențiale (anvelopei bioclimatice) actuale și trecute pentru
speciile din grupul “Isophya camptoxypha”.
3.1. Elaborarea unui model preliminar de distribuție potențială pentru speciile din
complexul “Isophya camptoxypha” folosind doi algoritmi de modelare: Maxent și Desktop
GARP.

15
3.2. Activități de teren pentru a colecta puncte de distribuție pentru speciile țintă și a
investiga locațiile prezise de modele, dar unde specia nu a fost găsită anterior și colectarea de
material biologic.
3.3. Recalibrarea modelelor de distribuție potențială folosind date din teren, pentru a
crește acuratețea rezultatelor și transferarea modelelor pe climate din trecut.
4. Rezultate așteptate:
4.1. Hărți preliminare de distribuție a speciilor din grupul “Isophya camptoxypha”.
4.2. Baza de date cu secvențe ADN provenind de la taxonii analizați.
4.3. Publicarea unui articol științific.
4.4. Participare la o manifestare științifică.
II. Descrierea științifică și tehnică a etapei
Tabel 2.1. Gradul de realizare a obiectivelor și activităților
Obiectiv / Activitate Grad de
realizare
1. Evaluarea comunicării acustice la speciile din grupul “Isophya camptoxypha”. 100%
1.1. Activități de teren pentru a înregistra speciile de interes și a colecta probe biologice. 100%
1.2. Înregistrarea cântecelor în natură/laborator pentru obținerea datelor privind sistemele de
comunicare acustică, înregistrarea masculilor care stridulează și a duetelor mascul-femelă. 100%
2. Studiul relațiilor filogenetice dintre speciile grupului “Isophya camptoxypha”, folosind
markeri moleculari. 100%
2.1. Extracție de ADN genomic de la probele colectate. 100%
2.2. Amplificarea markerilor moleculari necesari în analiza filogenetică și obținerea secvențelor
ADN ale acestora. 100%
3. Evaluarea distribuției potențiale (anvelopei bioclimatice) actuale și trecute pentru
speciile din grupul “Isophya camptoxypha”. 100%
3.1. Elaborarea unui model preliminar de distribuție potențială pentru speciile din complexul
“Isophya camptoxypha” folosind doi algoritmi de modelare: Maxent și Desktop GARP. 100%
3.2. Activități de teren pentru a colecta puncte de distribuție pentru speciile țintă și a investiga
locațiile prezise de modele, dar unde specia nu a fost găsită anterior și colectarea de
material biologic.
100%
3.3. Recalibrarea modelelor de distribuție potențială folosind date din teren, pentru a crește
acuratețea rezultatelor și transferarea modelelor pe climate din trecut. 100%
4. Diseminarea rezultatelor. 100%
4.1. Hărți preliminare de distribuție a speciilor din grupul “Isophya camptoxypha”. 100%
4.2. Baza de date cu secvențe ADN provenind de la taxonii analizați. 100%
4.3. Publicarea unui articol științific. 100%
4.4. Participare la o manifestare științifică. 100%

16
Obiectivul 1. Evaluarea comunicării acustice la speciile din grupul Isophya camptoxypha
Activități de teren pentru a înregistra speciile de interes și a colecta probe biologice
Speciile din grupul studiat au o distribuție Central Europeană: partea de Est a Alpilor
și în bazinul Carpatic. În anul 2016 au fost întreprinse mai multe deplasări în teren în
următoarele zone din România: Lipova, Săvârșin (jud. Arad), Blaj (jud. Alba), Sovata (jud.
Mureș), Făgăraș, Agnita (jud. Sibiu), Munții Ciucaș (jud. Prahova), Gura Humorului, Cacica,
Putna, Munții Rarău, Munții Călimani, Vatra Dornei (jud. Suceava), Munții Maramureșului,
Munții Rodnei, Poienile de sub Munte (jud. Maramureș), Piatra Neamț, Poiana Largului,
Durău, Munții Ceahlău (jud. Neamț), pentru colectarea acestor insecte.
Înregistrările acustice ale diverselor specii studiate au fost efectuate în teren cu
ajutorul următorului echipament: reportofon digital Edirol R-09HR (frecvența de răspuns a
microfonului: 20–40000 Hz, fișiere .wav 96000 Hz, 24-bit) și un microfon extern Sennheiser
“shotgun” ME65 (Fig. 2.1). Amplitudinea perfectă pentru analiza oscilografică a cîntecului se
obține la intensitatea de -12 – -6 dB, respectiv prin poziționarea microfonului la aproximativ
30-50 cm de insectă. Perioada optimă a zilei pentru acest studiu este la apusul soarelui și în
primele ore ale nopții, când cosașii Isophya devin activi și încep să striduleze.
Fig. 2.1. Expediție în Carpați (stânga) pentru înregistrarea stridulațiilor (centru) speciilor din
complexul “Isophya camptoxypha” (dreapta)
Înregistrarea cântecelor în natură/laborator pentru obținerea datelor privind sistemele
de comunicare acustică, înregistrarea masculilor care stridulează și a duetelor mascul-
femelă
Introducere.
În general, funcția de bază a cântecului constă în transmiterea informației între
masculi și femele, în timpul comportamentului de împerechere: de regulă, masculii
stridulează spontan, iar femelele decid de care mascul să se apropie. Pentru a forma cuplul,
femela răspunde acustic într-o fereastră de oportunitate strict definită temporal în funcție de
stridulația masculului și formează un duet acustic cu acesta, componentă semnificativă a
sistemului de recunoaștere al partenerului la acești cosași. Modularea amplitudinală a

17
stridulației masculului și sincronizarea răspunsului femelei sunt diferite și strict specifice
pentru fiecare specie din acest gen.
Metode.
Înregistrările au fost efectuate cu un reportofon digital EDIROL R-09HR, sunetul
fiind captat la o frecvență de sampling de 96 kHz și rezoluție amplitudinală de 24-bit.
Temperatura ambientală a fost măsurată în timpul fiecărei înregistrări, deoarece densitatea
repetiției diferitelor elemente structurale ale stridulației sunt foarte dependente de temperatură
la animalele poikiloterme. Înregistrările audio și video s-au desfășurat în natură, dar și în
laborator (fig. 2.2).
Fig. 2.2. Frame-uri din înregistrări video ale mișcărilor aripilor în timpul stridulației la
speciile din complexul “Isophya camptoxypha”, în teren (stânga) și în laborator (dreapta)
Corelarea frame-urilor din înregistrările video cu semnalele acustice permit
vizualizarea comportamentului în timpul stridulației și mai ales a mișcărilor aripilor în timpul
producerii sunetelor. În laborator, insectele au fost înregistrate în terarii cu pereții din burete
(pentru prevenirea reverberațiilor), microfonul fiind așezat la o anumită distanța de aripile
insectei, în funcție de intensitatea sunetului emis de aceasta (fig. 2.3).
Fig. 2.3. Producerea sunetului prin mișcarea aripilor la o specie studiată: corelarea frame-
urilor dintr-o înregistrare video cu oscilograma sunetului (stânga) și poziționarea
microfonului față de aripile insectei în timpul înregistrărilor din laborator (dreapta)
Răspunsul femelei a fost obținut prin aducerea unei femele în terariul unui mascul
care stridulează și formarea spontană a unui duet acustic între cei doi parteneri. Înregistrarea
s-a realizat cu același echipament (Edirol R-09HR), dar folosind microfoanele stereo ale
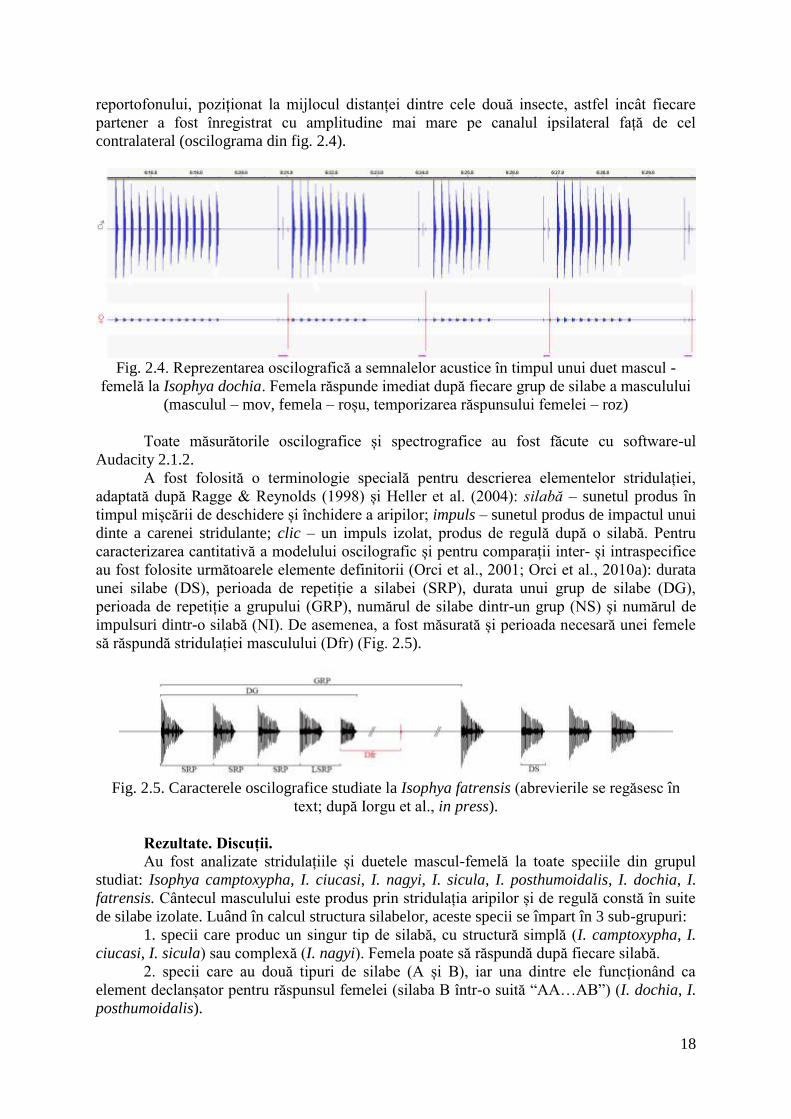
18
reportofonului, poziționat la mijlocul distanței dintre cele două insecte, astfel incât fiecare
partener a fost înregistrat cu amplitudine mai mare pe canalul ipsilateral față de cel
contralateral (oscilograma din fig. 2.4).
Fig. 2.4. Reprezentarea oscilografică a semnalelor acustice în timpul unui duet mascul -
femelă la Isophya dochia. Femela răspunde imediat după fiecare grup de silabe a masculului
(masculul – mov, femela – roșu, temporizarea răspunsului femelei – roz)
Toate măsurătorile oscilografice și spectrografice au fost făcute cu software-ul
Audacity 2.1.2.
A fost folosită o terminologie specială pentru descrierea elementelor stridulației,
adaptată după Ragge & Reynolds (1998) și Heller et al. (2004): silabă – sunetul produs în
timpul mișcării de deschidere și închidere a aripilor; impuls – sunetul produs de impactul unui
dinte a carenei stridulante; clic – un impuls izolat, produs de regulă după o silabă. Pentru
caracterizarea cantitativă a modelului oscilografic și pentru comparații inter- și intraspecifice
au fost folosite următoarele elemente definitorii (Orci et al., 2001; Orci et al., 2010a): durata
unei silabe (DS), perioada de repetiție a silabei (SRP), durata unui grup de silabe (DG),
perioada de repetiție a grupului (GRP), numărul de silabe dintr-un grup (NS) și numărul de
impulsuri dintr-o silabă (NI). De asemenea, a fost măsurată și perioada necesară unei femele
să răspundă stridulației masculului (Dfr) (Fig. 2.5).
Fig. 2.5. Caracterele oscilografice studiate la Isophya fatrensis (abrevierile se regăsesc în
text; după Iorgu et al., in press).
Rezultate. Discuții.
Au fost analizate stridulațiile și duetele mascul-femelă la toate speciile din grupul
studiat: Isophya camptoxypha, I. ciucasi, I. nagyi, I. sicula, I. posthumoidalis, I. dochia, I.
fatrensis. Cântecul masculului este produs prin stridulația aripilor și de regulă constă în suite
de silabe izolate. Luând în calcul structura silabelor, aceste specii se împart în 3 sub-grupuri:
1. specii care produc un singur tip de silabă, cu structură simplă (I. camptoxypha, I.
ciucasi, I. sicula) sau complexă (I. nagyi). Femela poate să răspundă după fiecare silabă.
2. specii care au două tipuri de silabe (A și B), iar una dintre ele funcționând ca
element declanșator pentru răspunsul femelei (silaba B într-o suită “AA…AB”) (I. dochia, I.
posthumoidalis).

19
3. specii care stridulează silabele în grupuri, fără un element special structural care să
declanșeze răspunsul femelei (I. fatrensis) (după Iorgu et al., in press).
La această ultimă specie, declanșarea răspunsului femelei în funcție de stimulii
acustici din stridulația masculului a făcut obiectul unui studiu recent (Iorgu et al., in press):
elementul funcțional din cântecul masculului este ultima silabă a unui grup, răspunsul femelei
fiind temporizat la un interval de 228–335 ms, unul dintre cele mai lungi de acest gen din
cadrul grupului [125–186 ms la I. harzi (Orci et al., 2010a), 40–70 ms la I. sicula (Orci et al.,
2010b), 13–34 ms la I. dochia (Iorgu, 2012), 70–130 ms la I. nagyi (Szövényi et al., 2012)].
Elementul declanșator se află în ultima silabă a unui grup, dar în acest caz există două
posibilități: începutul silabei sau finalul acesteia? Au fost examinate cele două cazuri, iar
efectul duratei ultimei silabe asupra femelei a fost negativ: timpul de reacție al femelelor nu
se modifică odată cu modificarea duratei ultimei silabe a masculului. Așadar, răspunsul
femelei ar depinde de începutul acestei silabe.
Rezultatul studiului arată unicitatea duetului dintre masculii și femelele acestei specii
și imposibilitatea formării unui cuplu acustic intraspecific. Pe viitor, studii asupra elementelor
spectrale ale sunetelor produse de speciile din acest complex vor completa cunoștințele
prezente despre datele oscilografice (după Iorgu et al., in press).

20
Obiectivul 2. Studiul relațiilor filogenetice dintre speciile grupului Isophya
camptoxypha, folosind markeri moleculari
Extracție de ADN genomic de la probele colectate
Colectarea probelor de la indivizi din genul Isophya
A fost constituită o colecție cu exemplare din genul Isophya din România și nu numai,
cu peste 600 de exemplare colectate. Toate exemplarele au fost fixate în alcool 96% și
etichetate. Etichetele conțin următoarele informații: Locul colectării, data colectării,
coordonatele GPS, colectorul.
Fig. 2.6. Exemple din colecția de indivizi de Isophya sp. Exemplarele sunt ținute individual în
recipiente în alcool etilic de 96%
Extracția ADN de la indivizi din grupul Isophya camptoxypha
Au fost colectate probe de țesut de la 307 exemplare aparținând speciilor din grupul
“Isophya camptoxypha”, din 78 de situri (Fig. 2.6, 2.7, 2.8). De la fiecare exemplar a fost
prelevat piciorul posterior stâng, fixat în alcool 96%. ADN-ul genomic total a fost extras din
ţesut muscular provenit de la fiecare individ în parte, urmând etapele kiturilor-ului ISOLATE
II Genomic DNA Kit (Bioline, London, UK).
Liza celulară începe prin incubarea probei într-o soluţie de proteinază K şi SDS.
Condiţiile necesare legării ADN la membrana de dioxid de siliciu din coloniţe sunt realizate
prin adăugarea de săruri şi etanol. Procesul de legare la membrană este reversibil şi specific
acizilor nucleici. Contaminările sunt înlăturate prin spălări succesive în două soluţii tampon
diferite. ADN genomic pur este eluat la final într-o soluţie uşor alcalină sau în apă distilată.
Eluția AND s-a realizat într-un volum final de 60-100 µl, în funcție de starea indivizilor.
ADN genomic extras a fost verificat atât din punct de vedere calitativ, cât şi cantitativ,
utilizând spectrofotometrul NanoDrop ND-1000. Concentrațiile DNA variază de la 6 ng/µl la
500 ng/µl.

21
Fig. 2.7. Exemple de cutii cu tuburi cu ADN genomic de la indivizi din grupul “Isophya
camptoxypha”. Tuburile cu ADN genomic sunt depozitate în congelatoare, la -20°C.
Datele privind concentrația ADN genomic și rapoartele A260/A280 și A260/A230 au
fost completate într-o bază de date cu probe ADN de la Isophya camptoxypha. De asemenea,
au fost completate în baza de date și următoarele aspecte: Id-ul probei, Specia probabilă,
Locul de colectare, Țara/județul, Data calendaristică, secvențele obținute de la markerii
moleculari amplificați, concentrația ADN genomic, Rapoartele de puritate ale ADN genomic
față de proteine și ARN. Această bază de date are atașate și fișiere fasta cu secvențe
amplificate de la fragmente din genele COI mitocondrial, 16s mitocondial, ITS1 și ITS2
nucleare.
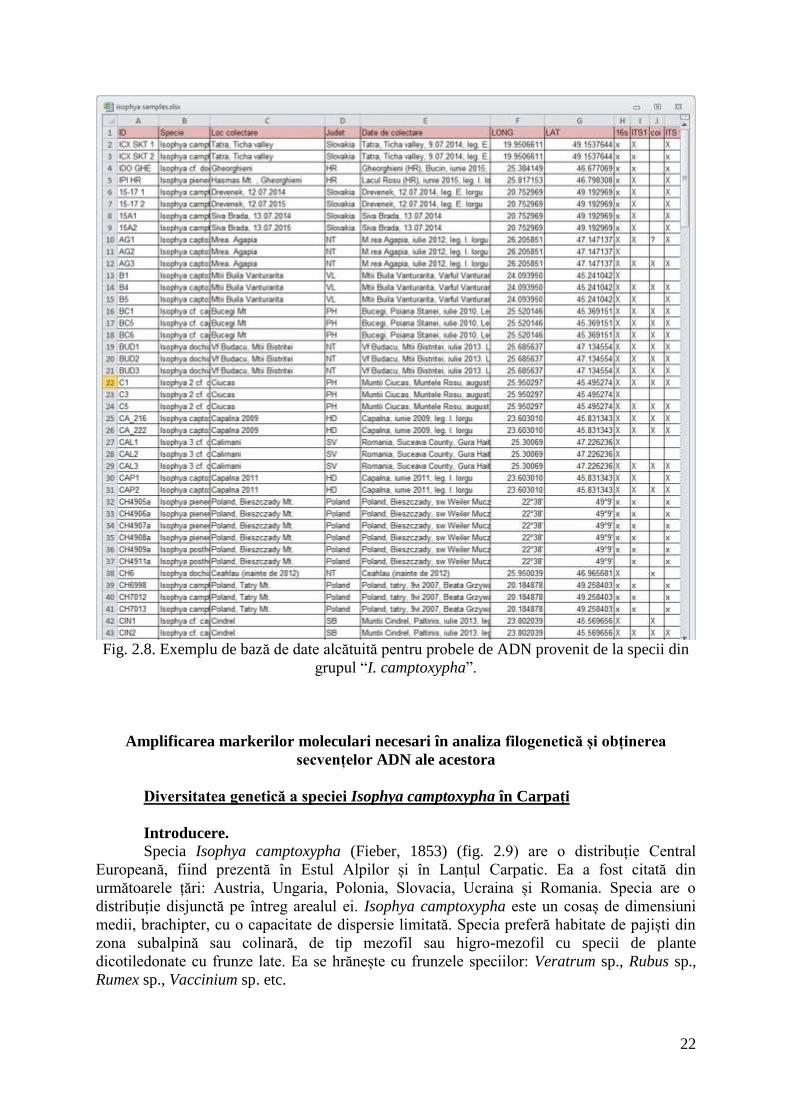
22
Fig. 2.8. Exemplu de bază de date alcătuită pentru probele de ADN provenit de la specii din
grupul “I. camptoxypha”.
Amplificarea markerilor moleculari necesari în analiza filogenetică și obținerea
secvențelor ADN ale acestora
Diversitatea genetică a speciei Isophya camptoxypha în Carpați
Introducere.
Specia Isophya camptoxypha (Fieber, 1853) (fig. 2.9) are o distribuție Central
Europeană, fiind prezentă în Estul Alpilor și în Lanțul Carpatic. Ea a fost citată din
următoarele țări: Austria, Ungaria, Polonia, Slovacia, Ucraina și Romania. Specia are o
distribuție disjunctă pe întreg arealul ei. Isophya camptoxypha este un cosaș de dimensiuni
medii, brachipter, cu o capacitate de dispersie limitată. Specia preferă habitate de pajiști din
zona subalpină sau colinară, de tip mezofil sau higro-mezofil cu specii de plante
dicotiledonate cu frunze late. Ea se hrănește cu frunzele speciilor: Veratrum sp., Rubus sp.,
Rumex sp., Vaccinium sp. etc.

23
Fig. 2.9. Mascul și femelă de Isophya camptoxypha (Foto: I.Ș. Iorgu)
Din punct de vedere morfologic, specia aparține unui supra-grup numit “Isophya
pyrenaea”, care este format din 16 specii de cosași (Warchałowska–Śliwa et al., 2008).
Datorită similarităților morfologice foarte mari și a lipsei unor piese genitale sclerotizate la
masculi, cel mai bun criteriu de discriminare pentru acest gen de cosași este stridulația.
Pentru o bună diferențiere a speciilor se analizează elementele structurale ale cântecului
masculilor, care sunt specific fiecărei specii, precum și duetele formate între masculi și
femele (Heller et al., 2004).
Scopul studiului nostru a fost să analizăm diversitatea genetică a speciei cu ajutorul
secvențelor a două fragmente de gene mitocondriale (16s și COI) și de a testa ipoteza
corelării grupurilor filogenetice descoperite la acestă specie cu distribuția lor geografică.
Materiale și metode.
Prelevarea probelor
Am colectat 83 de exemplare adulte de I. camptoxypha din 37 de populații de pe
întreg arealul acestei specii. Colectarea s-a realizat manual sau prin cosire cu fileul
entomologic. Fiecare individ a fost prins viu, înregistrat acustic pentru a se determina clar
dacă aparține acestei specii și apoi a fost fixat în alcool etilic 96% pentru analize genetice
ulterioare. Ca outgroup s-au folosit 3 exemplare din grupul “Isophya modesta” (după
Grzywacz-Gibala et al., 2010): 2 exemplare de Isophya modesta și un exemplar de Isophya
longicaudata (fig. 2.10).

24
Fig. 2.10. Harta distribuției populațiilor de I. camptoxypha luate în analiză (roșu). Albastru –
specii din outgroup (Isophya modesta)
Extracția DNA și amplificarea markerilor genetici
ADN genomic a fost extras din țesut muscular provenit din piciorul posterior de la
fiecare individ în parte, cu ajutorul uni kit de izolare ADN de la Bioline Genomic II DNA kit,
conform instrucțiunilor producătorului.
S-au amplificat două fragmente genice din ADN mitocondrial, cu ajutorul unor
primeri specifici universali:
1. un fragment de 662 perechi de baze, provenind de la subunitatea I a genei Citocrom
C oxidaza mitocondrială (coi) - cu ajutorul primerilor: (LCO1490 – LCO2198) (Folmer,
1994);
2. un fragment de 494 perechi de baze, din gena ARN ribozomal, subunitatea 16s
(16s) – cu ajutorul primerilor (16sSa – 16sSb) (Simon et al., 1994).
Reacţia PCR pentru genotipare a fost realizată într-un volum total de 50 μl cu
următoarele componente: 100-200 ng ADN matriță, 10 μl buffer de reactie 5X Green
GoTaq® Reaction Buffer Promega, 1.5-2.5 mM MgCl2, fiecare dNTP la o concentraţie de 0.1
mM, 0.1 μM din fiecare primer (unul dintre ei marcat cu o “coadă” M13), 1.5 U de ADN
polimerază GoTaq® DNA Polymerase, și apă distilată sterilă până la volumul final.
Amplificarea PCR s-a realizat într-un thermal cycler de tip BioRad PTC 200 PCR
engine după următoarele protocoale:
COI - o etapă de denaturare iniţială la 95°C pentru 2 min, urmată de 5 cicluri cu
denaturare la 95°C pentru 30 s, alinierea primerilor la 45°C pentru 1 min 30 s şi elongare la
72°C pentru 1 min, urmate de 30 cicluri cu denaturare la 95°C pentru 30 s, alinierea la 52 °C
pentru 1 min 30 s şi elongare la 72°C pentru 1 min, urmate de o etapă finală de elongare la
72°C pentru 5 min.

25
16s - o etapă de denaturare iniţială la 95°C pentru 3 min urmată de 35 cicluri cu
denaturare la 95°C pentru 30 s, alinierea primerilor la 48°C pentru 45 s şi etapa de elongare la
72°C pentru 45 s, urmate de o etapă finală de elongare la 72°C pentru 5 min.
După reacția de amplificare, produșii de PCR sau fost încarcați, migrați și vizualizați
pe un gel de agaroză de concentrație 1.5 %, colorat cu bromură de etidiu. Fragmentele de
interes au fost excizate din gel și purificate utilizând un kit specific FavorPrep™ GEL/PCR
Purification Kit de la Favorgen Corp., conform instrucțiunilor fabricantului.
Secvențierea produșilor de PCR s-a realizat unidirecțional apelând la serviciile
Macrogen.com.
Analize moleculare
Secvențele obținute au fost curățate și aliniate cu ajutorul programelor Codon Code
Aligner (codonCode Corp.) și Mega 7 v. 7.0.14 (Tamura, Stecher & Kumar, 2016).
Modelul de substituție a nucleotidelor cu cea mai mare probabilitate a fost calculat cu
ajutorul programului jModelTest 2.1.5 (Darriba et al., 2012; Guindon & Gascuel, 2003),
ordonate după algoritmul Akaike Information Criterion (AIC).
Pe baza secvențelor concatenate de 16s și COI și acestor analize preliminarii au fost
reconstruiți arbori filogenetici după următoarele metode: Maximum Likehood (ML),
Maximum Parsimony (MP), Neighbour Joining (NJ) și Bayesian Inference (BI). Programul
Mega 7 v7.0.14 a fost utilizat pentru reconstrucția arborilor cu ajutorul metodelor Maximum
Likehood (1000 de iterații de tip Bootstrap, deleția completă a gapurilor), Maximum
Parsimony (1000 de iterații de tip Bootstrap, deleția completă a gapurilor), Neighbour Joining
(1000 de iterații de tip Bootstrap, deleția completă a gapurilor). Reconstrucția filogenetică
prin metoda Bayesian Inference a fost realizată prin programul MrBayes v3.2.2 (Ronquist &
Huelsenbeck 2003; Ronquist et al., 2005) cu 1.000.000 de generații simulate și cu o rată de
extracție de 100 de generații).
Diversitatea genetică, numărul de haplotipuri și divergențele genetice au fost calculate
folosind programul DnaSP v5.10.1 (Librado & Rozas, 2009). Ca parametri de variație
genetică au fost calculate numărul de situri polimorfice/segregative (S), numărul de
haplotipuri (H), diversitatea haplotipică și diversitatea nucleotidelor (π), pentru totalul
secvențelor, pentru fiecare clad în parte și pentru fiecare subclad în parte.
Estimarea timpului de divergență a cladelor filogenetice a fost realizată cu ajutorul
programului Beast v1.8.1 (Drummond et al., 2012), folosind un model de ceas molecular
strict și rate de substituție cunoscute pentru cei doi markeri (0.0177 substituții/sit/Mya pentru
COI și 0.0054 substituții/sit/Mya pentru 16s) determinate după alte specii de Isophya
(Chobanov et al., 2016 și Papadopoulou et al., 2010). Mărimea populației în timp a fost
estimată după un model de speciație de tip Yule, lungimea lanțului Markov-Chain-Monte-
Carlo (MCMC) a fost setat la 10×106 cu un burn in de 106, în timp ce ceilați parametrii au
rămas în ”default”.
De asemenea, am folosit algoritmul Median-Joining implementat în programul
NETWORK Software v4.6.1.0 (http://www.fluxus-engineering.com) pentru a ilustra pattern-
urile filogenetice și geografice în diversitatea haplotipurilor.
Rezultate.
Caracteristicile secvențelor, diversitatea haplotipică și distanțele genetice între
principalele clade
În cei 86 de indivizi (incluzând outgroup-ul), secvențele concatenate au însumat 1156
perechi de baze. În aceste secvențe au fost detectate 52 de haplotipuri de I. camptoxypha și 3
haplotipuri în outgroup (2 de I. modesta și 1 de I. longicaudata). În total, 186 de situri s-au

26
dovedit variabile, iar dintre acestea, 153 au fost informative parsimonial, fără să fie detectată
vreo diferență semnificativă între cei doi loci.
În cadrul secvențelor din specia I. camptoxypha (83 de indivizi) au fost descoperite 52
de haplotipuri. Secvențele conțin 1028 situri invariabile și 128 situri polimorfice dintre care
114 informative parsimonial. S-a observat o diversitate haplotipică foarte ridicată (0.986)
(vezi tabelul 2).
Tabelul 2.2. Parametrii de variație genetică în cadrul speciei I. camptoxypha. Abrevieri:
mărimea probei (N), nr. de situri polimorfice (S), numărul de haplotipuri (H), diversitatea
haplotipica (h) și diversitatea nucleotidica (π)
Grup N S H h π
Total 83 128 52 0.986 0.03122
Subclad I Carpathians 41 74 26 0.974 0.01277
Clad II Carpathians 37 49 23 0.964 0.00757
Subclad Eastern Alps 5 3 3 0.7 0.00104
Media distanței genetice între grupuri este prezentată în tabelul 2.3:
CLAD II CLAD I CLAD III OUTGROUP
CLAD_II Carpathians
0.006 0.006 0.009
SUBCLAD_I Carpathians 0.053
0.005 0.010
SUBCLAD_Eastern Alps 0.037 0.036
0.010
OUTGROUP 0.098 0.106 0.099
Analizele filogenetice
Conform algoritmului BIC, modelul evolutiv de substituție a nucleotidelor cu cea mai
mare probabilitate s-a dovedit a fi Hasegawa-Kishino-Yano (HKY, nst=2) cu o distribuție de
tip Gamma a ratei de variație pe situri și și o proporție de situri invariabile (HKY+G+I).
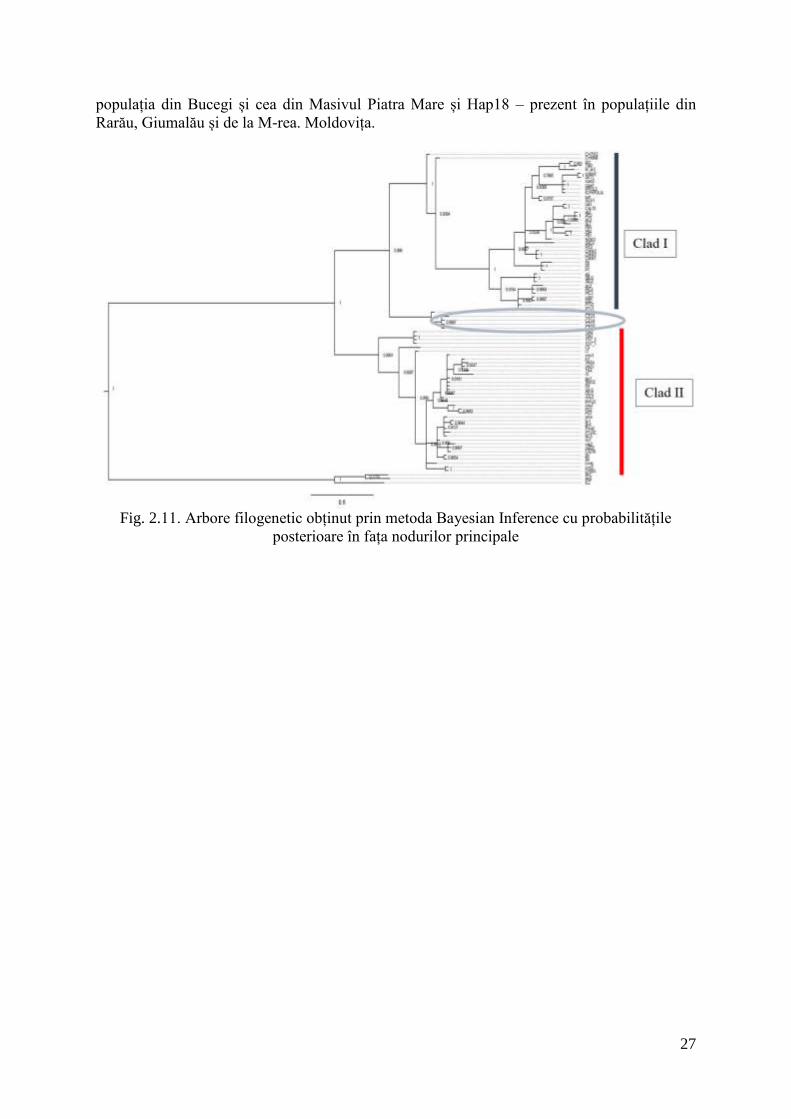
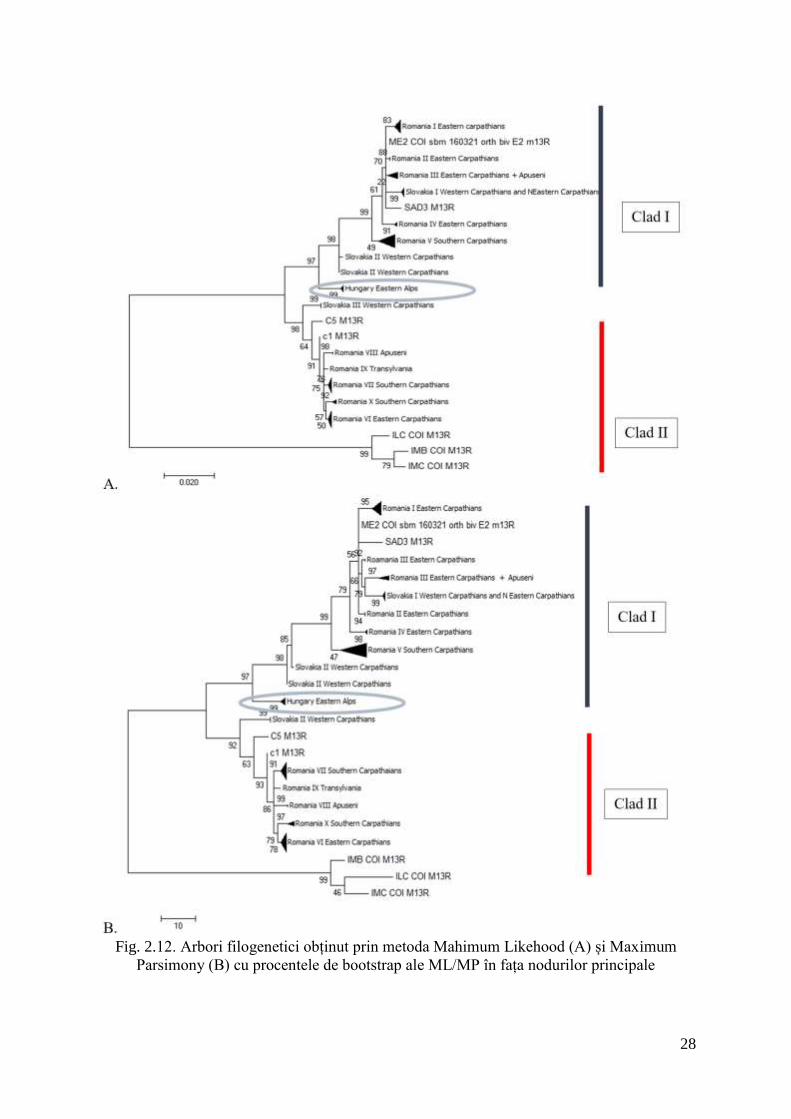
Analizele filogenetice de tip ML, MP, NJ și BI au generat arbori foarte asemănători și
înalt congruenți din punct de vedere al ordinii cladelor și poziția nodurilor majore. Fiecare
metodă a generat câte un arbore care prezenta 2 clade majore pentru specia I. camptoxypha.
Nu s-a dovedit o corelație între cele două clade filogenetice și distribuția geografică, fiecare
clad ocupând același areal în Carpați, având astfel o distribuție simpatrică. De asemenea, se
distinge prezența unui clad separat, format din probe provenind din Alpii de est, Munții
Kozseg din Ungaria (fig. 2.11, 2.12).
Cu toate acestea, privind distribuția în Carpați a celor două clade (fig. 2.13) putem
vedea că deși acestea sunt simpatrice, haplotipurille din Cladul I din Carpați nu ocupă
aceleași masive muntoase ca haplotipurile din Cladul II. Există o singură excepție: în Masivul
Postăvarul au fost descoperite haplotipuri unice aparținând la cele două claduri.
De asemenea, din cele 52 de haplotipuri identificate, 50 s-au dovedit a fi private
pentru masivele muntoase analizate (populațiile analizate), doar două haplotipuri găsindu-se
în 2, respectiv 3 populații foarte apropiate. Aceste două haplotipuri sunt: Hap4 – prezent în
27
populația din Bucegi și cea din Masivul Piatra Mare și Hap18 – prezent în populațiile din
Rarău, Giumalău și de la M-rea. Moldovița.
Fig. 2.11. Arbore filogenetic obținut prin metoda Bayesian Inference cu probabilitățile
posterioare în fața nodurilor principale
28
Fig. 2.12. Arbori filogenetici obținut prin metoda Mahimum Likehood (A) și Maximum
Parsimony (B) cu procentele de bootstrap ale ML/MP în fața nodurilor principale
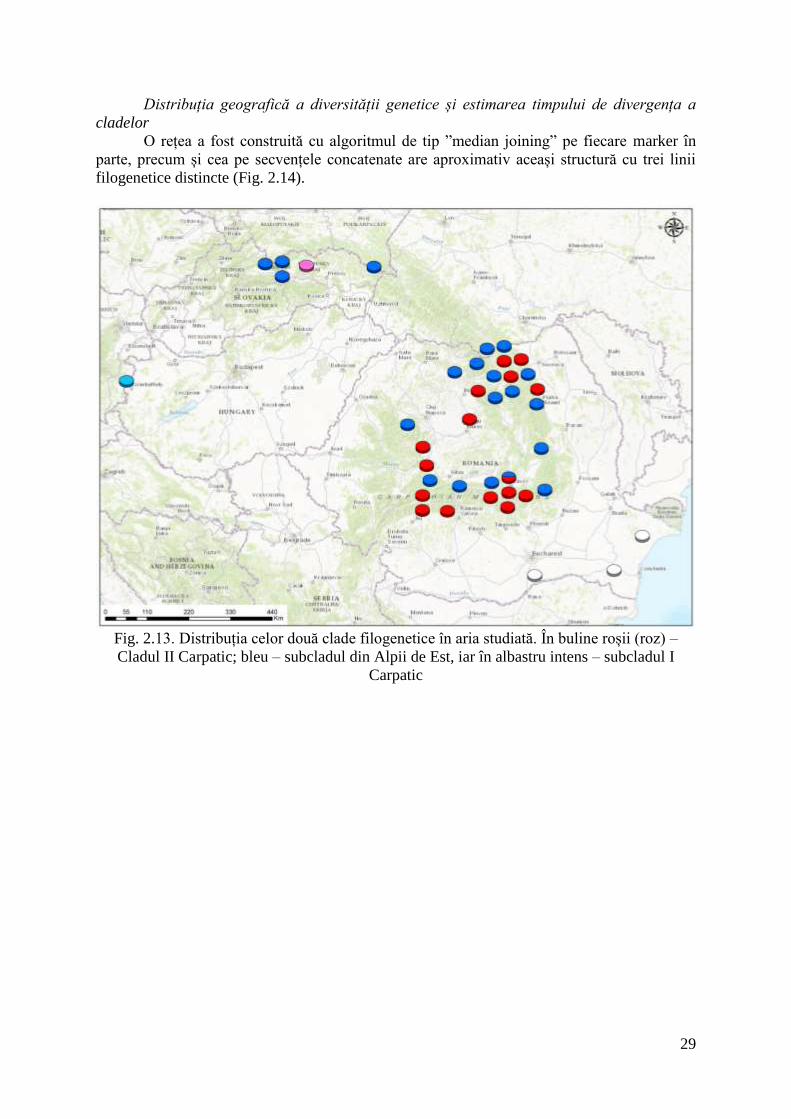
29
Distribuția geografică a diversității genetice și estimarea timpului de divergența a
cladelor
O rețea a fost construită cu algoritmul de tip ”median joining” pe fiecare marker în
parte, precum și cea pe secvențele concatenate are aproximativ aceași structură cu trei linii
filogenetice distincte (Fig. 2.14).
Fig. 2.13. Distribuția celor două clade filogenetice în aria studiată. În buline roșii (roz) –
Cladul II Carpatic; bleu – subcladul din Alpii de Est, iar în albastru intens – subcladul I
Carpatic

30
Fig. 2.14. Rețeaua de haplotipuri pe secvențe ale genei COI mitocondriale, realizată prin
algoritmul Median-Joining. Cercurile galbene reprezintă haplotipurile, mărimea cercurilor
arată numărul de secvențe prezent în fiecare haplotip.
În urma analizei secvențelor în programul BEAST v1.8.1, prin metoda de inferență
colescentă de tip Bayesian Markov Chain Monte Carlo s-a aproximat momentul de
divergență a cladelor principale la 1.96 milioane de ani. Acestă vechime a cladelor ne indică
faptul că cele două clade majore din cadrul I. camptoxypha s-au format cu mult înainte de
ultima perioadă glaciară. De asemenea, subcladul identificat in Munții Koszeg din Ungaria s-
a desprins de subcladul Carpatic aproximativ acum 1.3 milioane de ani. În cadrul Carpaților,
Cladul II și Subcladul I au avut linii filogenetice divergente care au aproximativ aceeași
vârstă (1 milion de ani).
De asemenea, separarea dintre grupurile I. camptoxypha și I. modesta s-a realizat
înainte de separarea speciilor I. modesta și I. longicauda (4.42 milioane de ani).
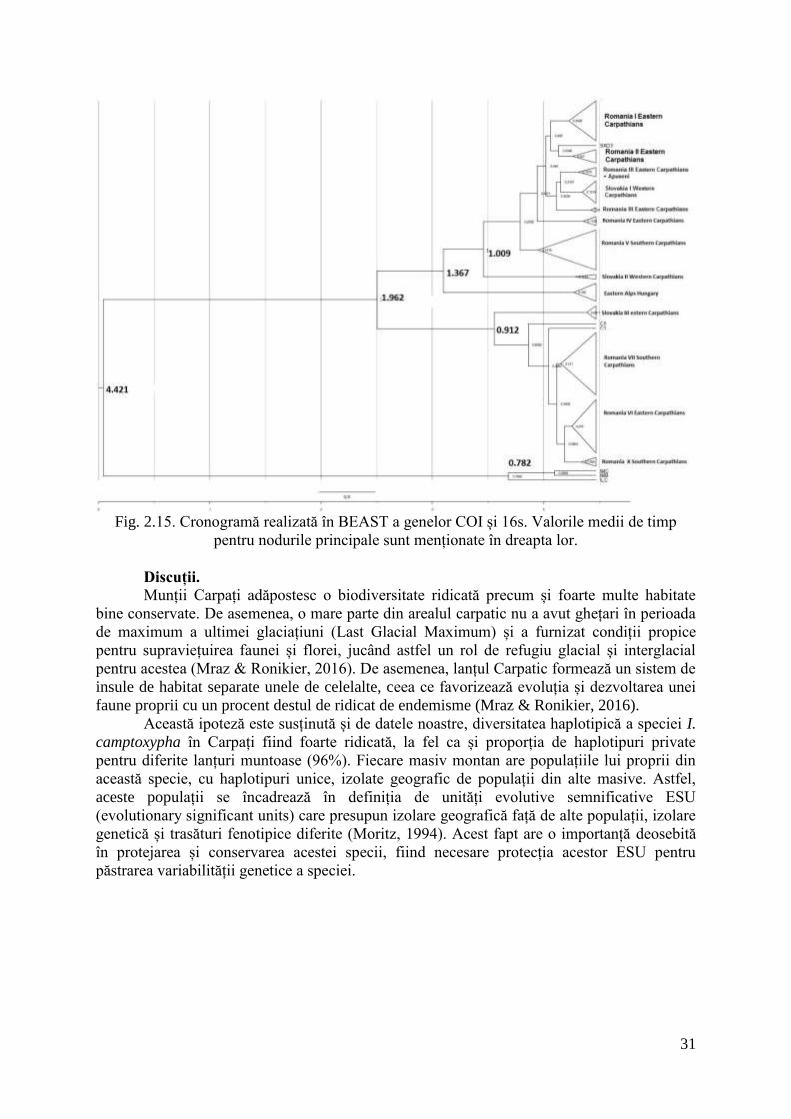
31
Fig. 2.15. Cronogramă realizată în BEAST a genelor COI și 16s. Valorile medii de timp
pentru nodurile principale sunt menționate în dreapta lor.
Discuții.
Munții Carpați adăpostesc o biodiversitate ridicată precum și foarte multe habitate
bine conservate. De asemenea, o mare parte din arealul carpatic nu a avut ghețari în perioada
de maximum a ultimei glaciațiuni (Last Glacial Maximum) și a furnizat condiții propice
pentru supraviețuirea faunei și florei, jucând astfel un rol de refugiu glacial și interglacial
pentru acestea (Mraz & Ronikier, 2016). De asemenea, lanțul Carpatic formează un sistem de
insule de habitat separate unele de celelalte, ceea ce favorizează evoluția și dezvoltarea unei
faune proprii cu un procent destul de ridicat de endemisme (Mraz & Ronikier, 2016).
Această ipoteză este susținută și de datele noastre, diversitatea haplotipică a speciei I.
camptoxypha în Carpați fiind foarte ridicată, la fel ca și proporția de haplotipuri private
pentru diferite lanțuri muntoase (96%). Fiecare masiv montan are populațiile lui proprii din
această specie, cu haplotipuri unice, izolate geografic de populații din alte masive. Astfel,
aceste populații se încadrează în definiția de unități evolutive semnificative ESU
(evolutionary significant units) care presupun izolare geografică față de alte populații, izolare
genetică și trasături fenotipice diferite (Moritz, 1994). Acest fapt are o importanță deosebită
în protejarea și conservarea acestei specii, fiind necesare protecția acestor ESU pentru
păstrarea variabilității genetice a speciei.

32
Fig. 2.16. Lanțul Carpatic și principalele bariere fizico-geografice ce au influențat de-a lungul
timpului distribuția diversității genetice la diverse specii (după Mraz & Ronikier, 2016)
Speciile cu dispersie redusă tind să prezinte pattern-uri de diferențiere foarte
pronunțate datorită lipsei de geneflow (Kuo & Avise, 2005). Mai multe studii asupra unor
specii cu capacitate de dispersie redusă au descoperit divergențe genetice profunde de-a
lungul populațiilor din Carpați, precum și persistența “in situ” a liniilor evolutive pe o
perioadă foarte mare de timp. Acestea au fost corelate cu diferite bariere fizico-geografice
(Babick et al., 2005, Mraz et al., 2006, Theissinger et al., 2013, Homburg et al., 2013 etc.).
În cazul speciei I. camptoxypha, am identificat două linii filogenetice diferite, cu o
divergență genetică foarte mare, dar care nu s-au corelat cu nicio barieră geografică din lanțul
carpatic.
Concluzii.
- Specia I. camptoxypha este o specie cu abilitate de dispersie redusă, cu o diferențiere
genetică puternică și prezintă numeroase populații izolate, cu haplotipuri private în diferite
masive montane care pot fi considerate ca potențiale ESU.
- Datele noastre sugerează că cele două clade principale prezente în Carpați au fost izolate
unul de celălalt pentru o perioadă lungă de timp. Se presupune că în timpul perioadelor dintre
glaciațiuni s-au petrecut evenimente de colonizare multiple, care au avut loc din mai multe
refugii.
- Pentru un tablou mai complet al istoriei evolutive a acestei specii este necesară realizarea
mai multor analize, care presupun un efort de sampling mai mare, precum și analiza unor
markeri genetici nucleari independenți.

33
3. Evaluarea distribuției potențiale (anvelopei bioclimatice) actuale și trecute pentru
speciile din grupul “Isophya camptoxypha”
Activități de teren pentru a colecta puncte de distribuție pentru speciile țintă și a
investiga locațiile prezise de modele, dar unde specia nu a fost găsită anterior și
colectarea de material biologic.
Punctele de ocurență au fost colectate în teren folosind o unitate GPS Garmin Oregon
450t, în format GPX și folosind proiecția WGS84. Pentru colectarea datelor s-au efectuat
deplasări în zonele unde distribuția speciilor nu era cunoscută, dar modelul preliminar
construit pe baza datelor anterioare indica o favorabilitate ridicată pentru specii. Datele
colectate au fost stocate într-o bază de date geospațială compatibilă cu principalele programe
GIS (QuantumGIS, DivaGIS, ArcGIS).
Fig. 2.17. Harta de distribuție a unei specii din complexul studiat: Isophya dochia

34
Elaborarea unui model preliminar de distribuție potențială pentru speciile din
complexul “Isophya camptoxypha” folosind doi algoritmi de modelare: Maxent și
Desktop GARP. Recalibrarea modelelor de distribuție potențială folosind date din
teren, pentru a crește acuratețea rezultatelor și transferarea modelelor pe climate din
trecut.
Materiale și metode.
Puncte de distribuție
Pentru crearea modelelor de distribuție au fost folosite atât datele existente în
literatură cât și datele colectate de membrii echipei în timpul activităților de teren. Baza de
date a fost rarefiată la o rezoluție de ~1 km folosind funcția “Trimm duplicate occurrences”
din ENMTools 1.4.4 (Warren et al., 2010), pentru a elimina punctele duplicate (i.e. prezența a
mai mult de 1 punct în fiecare celulă a rasterului).
Gruparea datelor de distribuție
Deoarece complexul “Isophya camptoxypha” cuprinde numeroase specii cu un areal
de distribuție limitat, adesea reprezentat doar de câte un singur masiv muntos izolat, datele de
prezență au fost grupate pentru modelare, rezultând o bază de date cu 86 de puncte de
distribuție după rarefierea spațială. Gruparea este susținută și de datele morfologice care nu
prezintă diferențe între specii, ceea ce indică o evoluție recentă a grupului.
Variabile climatice
Datele climatice utilizate pentru modelare au fost reprezentate de setul de 19 variabile
bioclimatice disponibile gratuit pe site-ul Worldclim (http://www.worldclim.org/, Hijmans et
al., 2005) și prezentate în continuare (Tabel 4). Pentru proiecțiile pe paleoclimate a fost
folosit același set de 19 variabile bioclimatice, acestea fiind calculate din simulări cu Modele
Globale de Climat (GCM) și disponibile pe site-ul Worldclim. Perioadele temporale de
referință au fost reprezentate de ultima perioadă interglaciară (LIG) (~120.000 – 140.000
î.e.n.), ultimul maxim glaciar (LGM) (~22.000 î.e.n.) și Holocenul Mijlociu (MH) (~6.000
î.e.n.). Datele pentru ultima perioadă interglaciară sunt furnizate de Otto-Bliesner et al.
(2008), iar datele pentru ultimul maxim glaciar și holocenul mijlociu sunt obținute din
Modelul Comunitar de Climat (CCSM4). Datele pentru ultima perioadă interglaciară și
pentru holocenul mijlociu au o rezoluție de ~1 km, iar datele pentru ultimul maxim glaciar au
o rezoluție de 2.5 minute de arc.
Tabel 2.4. Setul de 19 variabile bioclimatice utilizate pentru modelare, descărcate de pe site-
ul Worldclim
Cod Denumire
Bio1 Temperatura medie anuală
Bio2 Variația diurnă (media lunară(temp max – temp min)
Bio3 Izotermalitate (bio2/bio7)(*100)
Bio4 Sezonalitatea temperaturii (SD*100)
Bio5 Temperatura maximă a celei mai calde luni
Bio6 Temperatura minimă a celei mai reci luni
Bio7 Variația anuală a temperaturii (bio5-bio6)
Bio8 Temperatura medie a celui mai ploios sfert
Bio9 Temperatura medie a celui mai uscat sfert
Bio10 Temperatura medie a celui mai cald sfert
Bio11 Temperatura medie a celui mai rece sfert

35
Bio12 Precipitații anuale
Bio13 Precipitații în cea mai ploioasă lună
Bio14 Precipitații în cea mai uscată lună
Bio15 Sezonalitatea precipitațiilor (coeficient de variație)
Bio16 Precipitații în cel mai ploios sfert
Bio17 Precipitații în cel mai uscat sfert
Bio18 Precipitații în cel mai cald sfert
Bio19 Precipitații în cel mai rece sfert
Algoritmi de modelare
Inițial s-a dorit folosirea a doi algoritmi de modelare (Maxent și GARP) pentru a
obține modele de ansamblu, dar algoritmul GARP, implementat în programul Desktop GARP
nu a putut fi utilizat ca urmare a instabilității extensiei Dataset Manager care are rol de a crea
o bază de date cu variabilele utilizate în modelare și care trebuie să furnizeze informații
despre variabile către Desktop GARP.
În consecință am folosit programul Maxent 3.3.3k (Phillips et al., 2006) care
implementează algoritmul de entropie maximă – o versiune a modelelor liniare generalizate
(Renner & Warton, 2013). Avantajul programului constă în capabilitatea de a genera automat
pseudoabsențe folosind datele de background, producerea unor modele robuste chiar și
folosind un set redus de date, acceptarea unei game largi de date în format ASCII sau SWD
și disponibilitatea gratuită.
Modelele au fost antrenate pe o zonă care reprezintă arealul actual cunoscut al speciei
și apoi au fost transferate pe o scală spațială mai largă sau pe paleoclimate. Zona de antrenare
a fost determinată folosind funcția “Minimum bounding geometry” din ArcGIS 10.2.1 și
argumentul “Convex hull” pentru a genera o anvelopa care cuprinde toate punctele
cunoscute, la care a fost adăugat apoi un buffer generic de 10 km.
Setul de date de distribuție a fost împărțit în două seturi, 75% din puncte fiind folosite
pentru antrenare și 25% pentru testarea modelului generat. Pentru împărțirea setului de date a
fost folosită extensia SDMToolbox v1.1c (Brown, 2014) pentru ArcGIS disponibilă gratuit
(http://sdmtoolbox.org/).
Modelul a fost rulat folosind “Auto features”, 104 puncte de background și a fost
activată funcția de jackknife pentru a determina contribuția variabilelor în producerea
modelului. Algoritmul a fost lăsat să ruleze până la convergență sau pentru 1000 de iterații.
Ulterior variabilele cu o contribuție mai mică de 5% în generarea modelului au fost eliminate
și modelul a fost generat din nou și inspectat rapid pentru acuratețe folosind AUC și opinia
expertului.
Evaluarea modelelor
Modelele au fost evaluate în fază inițială pe bază de AUC și opinia expertului, iar
modelele finale au fost evaluate statistic folosind AUC parțial implementat în Partial ROC
(Narayani, 2008) și inspecția vizuală de către expert. În cazul AUC parțial s-a folosit raportul
dintre AUC la un nivel de confidență de 95% și AUC generat randomic (50%), precum și
testul z pentru a verifica semnificația statistică a modelelor obținute comparativ cu modele
generate randomic.
Rezultate.
Acuratețea modelelor
Modelele generate au avut un raport AUC0.05/AUC0.5 cuprins între 1.033 și 1.0342
(Fig. 2.18): prezent 1.033251±0.01294; LIG 1.033373±0.01281; LGM 1.034091±0.013057;
MH 1.034245±0.013442. Toate modelele au dovedit predicții semnificativ mai bune față de
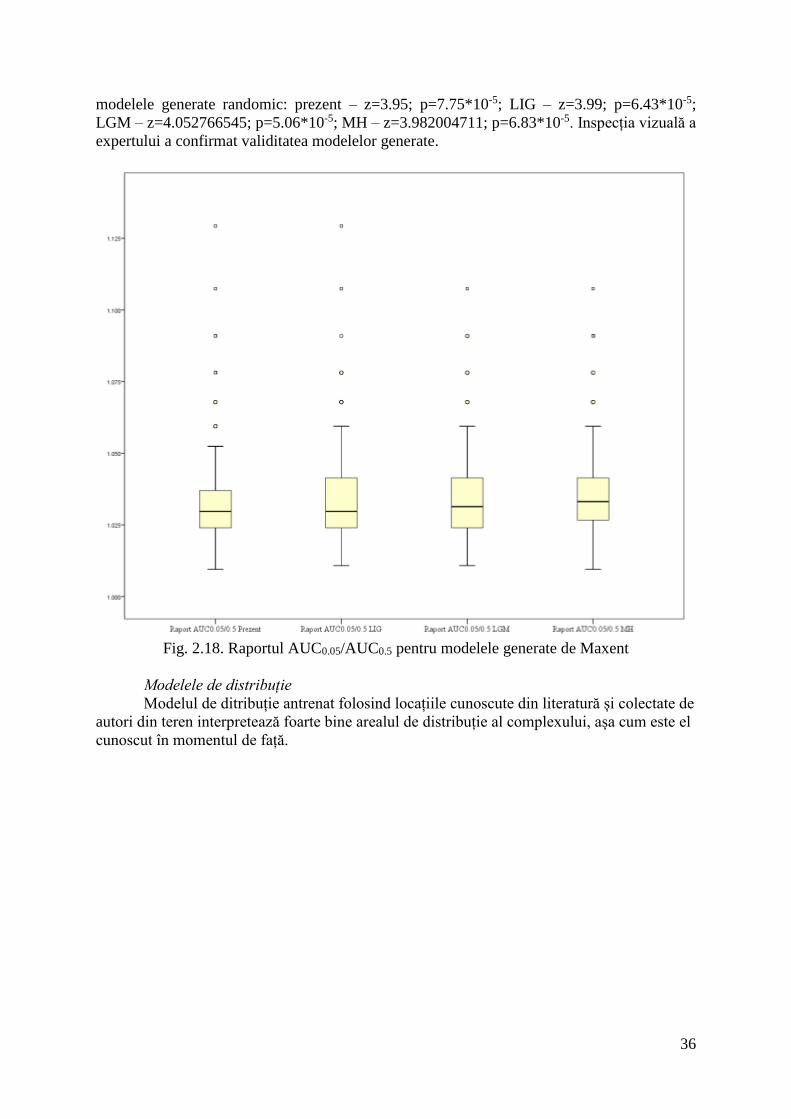
36
modelele generate randomic: prezent – z=3.95; p=7.75*10-5; LIG – z=3.99; p=6.43*10-5;
LGM – z=4.052766545; p=5.06*10-5; MH – z=3.982004711; p=6.83*10-5. Inspecția vizuală a
expertului a confirmat validitatea modelelor generate.
Fig. 2.18. Raportul AUC0.05/AUC0.5 pentru modelele generate de Maxent
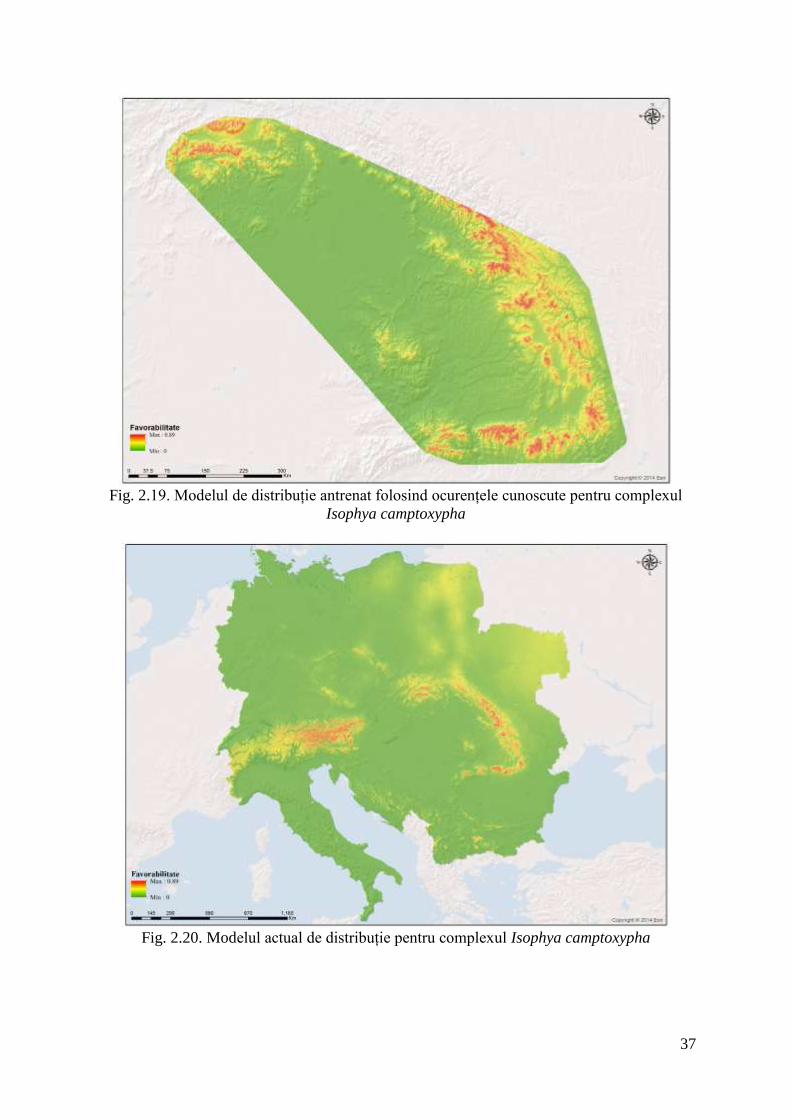
Modelele de distribuție
Modelul de ditribuție antrenat folosind locațiile cunoscute din literatură și colectate de
autori din teren interpretează foarte bine arealul de distribuție al complexului, așa cum este el
cunoscut în momentul de față.
37
Fig. 2.19. Modelul de distribuție antrenat folosind ocurențele cunoscute pentru complexul
Isophya camptoxypha
Fig. 2.20. Modelul actual de distribuție pentru complexul Isophya camptoxypha
38
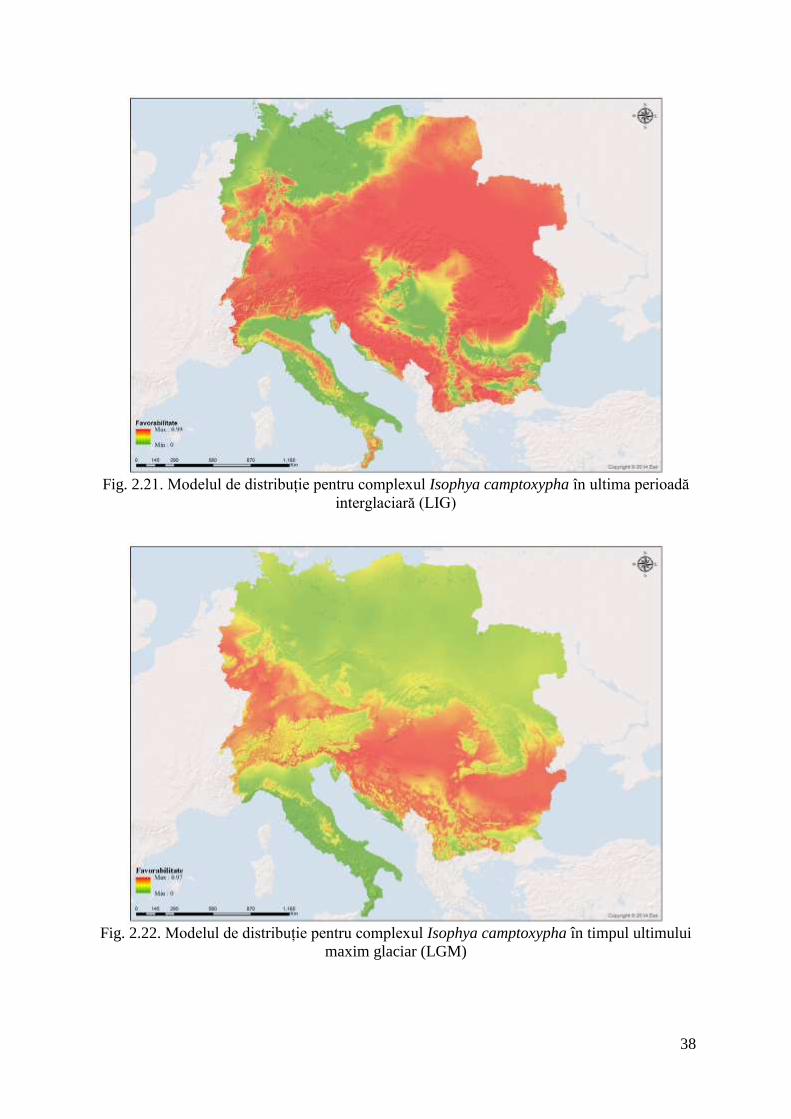
Fig. 2.21. Modelul de distribuție pentru complexul Isophya camptoxypha în ultima perioadă
interglaciară (LIG)
Fig. 2.22. Modelul de distribuție pentru complexul Isophya camptoxypha în timpul ultimului
maxim glaciar (LGM)

39
Fig. 2.23. Modelul de distribuție pentru complexul Isophya camptoxypha în timpul
Holocenului Mijlociu
Modelul general actual de distribuție al speciei (fig. 2.20) evidențiază foarte bine
spațiul geografic ocupat de acest complex și chiar suprinde zone noi dinspre Alpi unde
existența complexului este cunoscută dar punctele nu au fost folosite pentru antrenare cu
scopul de a testa performanța modelului.
Transferând modelele pe paleoclimate se observă că nișa ecologică a speciei acoperea
o mare parte din zona de studiu în perioada interglaciară (LIG), cuprinzând atât zone de
altitudine mai joasă, cât și zonele muntoase. Odată cu răcirea climei și instalarea perioadei
glaciare dimensiunea nișei se reduce iar complexul Isophya camptoxypha s-a refugiază către
zonele cu altitudini mai joase și climat mai blând. În Holocenul Mijlociu, când climatul era
mai blând decât astăzi speciile complexului au început să se retragă din nou către altitudini
mai mari, dar se observă că nișa climatică era mult mai mare față de cea actuală.

40
Bibliografie
Babik W, Branicki W, Crnobrnja-Isailovic J, Cogalniceanu D, Sas I, Olgun K, Poyarkov NA,
Garcia-Paris M, Arntzen JW (2005) Phylogeography of two European newt species:
discordance between mtDNA and morphology. Molecular Ecology 14: 2475-2491.
Brown JL (2014) SDMtoolbox: a python-based GIS toolkit for landscape genetic,
biogeographic, and species distribution model analyses. Methods in Ecology and Evolution
DOI: 10.1111/2041-210X.12200.
Chobanov DP, Kaya S, Grzywacz B, Warchałowska‐Śliwa E, Çıplak B (2016). The
Anatolio‐Balkan phylogeographic fault: a snapshot from the genus Isophya (Orthoptera,
Tettigoniidae). Zoologica Scripta (online first).
Darriba D, Taboada GL, Doallo R, Posada, D (2012) jModelTest 2: more models, new
heuristics and parallel computing. Nature Methods 9: 772.
Drummond AJ, Suchard MA, Xie D, Rambaut A (2012) Bayesian phylogenetics with
BEAUti and the BEAST 1.7. Molecular Biology and Evolution 29: 1969-1973.
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of
mitochondrial cytochrome c oxidase subunit I from diverse metazoan
invertebrates. Molecular marine biology and biotechnology 3(5): 294-299.
Grzywacz-Gibała B, Chobanov DP, Warchałowska-Sliwa E (2010) Preliminary phylogenetic
analysis of the genus Isophya (Orthoptera: Phaneropteridae) based on molecular data.
Zootaxa 2621: 27-44.
Guindon S, Gascuel O (2003) A simple, fast and accurate method to estimate large
phylogenies by maximum-likelihood. Systematic Biology 52: 696-704.
Heller K-G, Orci KM, Grein G, Ingrisch S (2004) The Isophya species of Central and
Western Europe (Orthoptera: Tettigonioidea: Phaneropteridae). Tijdschrift voor Entomologie
147: 237-258.
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A (2005) Very high resolution
interpolated climate surfaces for global land areas. International Journal of Climatology 25:
1965-1978.
Homburg K, Drees C, Gossner MM, Rakosy L, Vrezec A, Assmann T (2013) Multiple
glacial refugia of the low-dispersal ground beetle Carabus irregularis: molecular data
support predictions of species distribution models. PLoS ONE 8: e61185.
Iorgu IŞ (2012) Acoustic analysis reveals a new cryptic bush-cricket in the Carpathian
Mountains (Orthoptera, Phaneropteridae). ZooKeys 254: 1-22.
Iorgu IȘ, Kristin A, Szovenyi G, Kanuch P, Jarcuska B, Sahlean TC, Iorgu EI, Orci KM (in
press) Distinctive male-female acoustic duetting support the specific status of Isophya
fatrensis, a West-Carpathian endemic bush-cricket (Insecta: Orthoptera: Tettigoniidae:
Phaneropterinae). Bioacoustics. The International Journal of Animal Sound and its

41
Recording.
Librado P, Rozas J (2009) DnaSP v5: a software for comprehensive analysis of DNA
polymorphism data. Bioinformatics 25: 1451-1452.
Kuo CH, Avise JC (2005) Phylogeographic breaks in low-dispersal species: the emergence of
concordance across gene trees. Genetica 124(2-3): 179-186.
Moritz C (1994) Defining ’Evolutionarily Significant Units’ for conservation. Trends in
Ecology & Evolution 9: 373-375.
Mraz P, Gaudeul M, Rioux D, Gielly L, Choler P, Taberlet P, Intrabiodiv. Consortium (2007)
Genetic structure of Hypochaeris uniflora (Asteraceae) suggests vicariance in the Carpathians
and rapid post-glacial colonization of the Alps from an eastern Alpine refugium. Journal of
Biogeography 34: 2100-2114.
Mráz P, Ronikier M (2016) Biogeography of the Carpathians: evolutionary and spatial facets
of biodiversity. Biological Journal of the Linnean Society 119(3): 528-559.
Narayani B (2008) Tool for Partial-ROC (Biodiversity Institute, Lawrence, KS), ver 1.0.
Orci KM, Szövényi G, Nagy B (2001) A description of the song of Isophya beybienkoi
Mařan 1958 (Orthoptera: Tettigoniidae). Biologia (Bratislava) 56(5):489–495.
Orci KM, Szövényi G, Nagy B (2010a) A characterization of the pair forming acoustic
signals of Isophya harzi (Orthoptera, Tettigonioidea, Phaneropteridae). Acta Zoologica Acad.
Sci. Hung. 56(1): 43-53.
Orci KM, Szövényi G, Nagy B (2010b) Isophya sicula sp. n. (Orthoptera: Tettigonioidea), a
new, morphologically cryptic bush-cricket species from the Eastern Carpathians (Romania)
recognized from its peculiar male calling song. Zootaxa 2627: 57-68.
Otto-Bliesner BL, Marshall SJ, Overpeck JT, Miller GH, Hu A (2006) Simulating Arctic
climate warmth and icefield retreat in the last interglaciation. Science 311(5768): 1751-1753.
Papadopoulou A, Anastasiou I, Vogler AP (2010) Revisiting the insect molecular clock: the
Mid-Aegean Trench calibration. Molecular Biology and Evolution 27: 1659-1672.
Phillips S, Anderson R, Schapire R (2006) Maximum entropy modeling of species
geographic distributions. Ecological Modelling 190: 231-259.
Ragge DR, Reynolds WJ. 1998. The songs of the grasshoppers and crickets of Western
Europe. Colchester: Harley Books, 591 pp.
Renner IW, Warton DI (2013) Equivalence of MAXENT and Poisson Point Process Models
for Species Distribution Modeling in Ecology. Biometrics 69: 274-281.
Ronquist F, Huelsenbeck JP (2003) MrBayes version 3.0: Bayesian phylogenetic inference
under mixed models. Bioinformatics 19: 1572-1574.

42
Ronquist F, Huelsenbeck JP, van der Mark P (2005) MrBayes. Ver. 3.1. [Computer software
and manual. Draft 5/26/2005]. Available via http://mrbayes.csit.fsu.edu/manual.php.
Simon C, Frati F, Beckenbach A, Crespi B, Liu H et al. (1994) Evolution, Weighting, and
phylogenetic utility of mitochondrial gene-sequences and a compilation of conserved
polymerase chain-reaction primers. Annals of the Entomological Society of America 87: 651-
701.
Szövényi G, Puskás G, Orci KM (2012) Isophya nagyi, a new phaneropterid bush-–cricket
(Orthoptera: Tettigonioidea) from the Eastern Carpathians (Căliman Mountains, North
Romania). Zootaxa 3521: 67-79.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: Molecular
Evolutionary Genetics Analysis version 6. Molecular Biology and Evolution 30: 2725-2729.
Theissinger K, Balint M, Feldheim KA, Haase P, Johannesen J, Laube I, Pauls SU (2013)
Glacial survival and postglacial recolonization of an arctic-alpine freshwater insect
(Arcynopteryx dichroa, Plecoptera, Perlodidae) in Europe. Journal of Biogeography 40: 236-
248.
Warchałowska-Śliwa E, Chobanov DP, Grzywacz B, Maryańska-Nadachowska A (2008)
Taxonomy of the genus Isophya (Orthoptera, Phaneropteridae, Barbitistinae): comparison of
karyological and morphological data. Folia biologica (Kraków) 56(3-411): 227-241.
Warren DL, Glor RE, Turelli M (2010) ENMTools: a toolbox for comparative studies of
environmental niche models. Ecography 33: 607-611.
4. Diseminarea rezultatelor
Nr.
crt.
Rezultate
așteptate Rezultate livrate
1.
Hărți
preliminare
de distribuție
a speciilor
din grupul
Isophya
camptoxypha
Au fost realizate hărți de distribuție pentru fiecare dintre speciile
complexului “Isophya camptoxypha”, folosind software-ul ArcGIS
10.2.1. De asemenea, datele au fost introduse într-o bază de date
geospațială folosită pentru modelare.
2.
Baza de date
cu secvențe
ADN
provenind de
la taxonii
analizați
A fost generată o bază de date cu secvențe ADN provenind de la
taxonii analizați (Id-ul probei, Specia probabilă, Locul de colectare,
Țara/județul, Data calendaristică, secvențele obținute de la markerii
moleculari amplificați, concentrația ADN genomic, Rapoartele de
puritate ale ADN genomic față de proteine și ARN). Această bază de
date are atașate și fișiere fasta cu secvențe amplificate de la fragmente
din genele COI mitocondrial, 16s mitocondial, ITS1 și ITS2 nucleare.

43
3.
Publicarea
unui articol
științific
Iorgu I.Ș., Kristin A., Szovenyi G., Kanuch P., Jarcuska B., Sahlean
T.C., Iorgu E.I., Orci K.M. - Distinctive male-female acoustic
duetting support the specific status of Isophya fatrensis, a West-
Carpathian endemic bush-cricket (Insecta: Orthoptera: Tettigoniidae:
Phaneropterinae). Bioacoustics. The International Journal of Animal
Sound and its Recording [IF 1.369] (accepted on 11.11.2016)
http://www.tandfonline.com/toc/tbio20/current
4.
Participare la
o manifestare
științifică
Iorgu I.Ș., Kristin A., Szovenyi G., Kanuch P., Jarcuska B., Sahlean
T.C., Iorgu E.I., Orci K.M. - Acoustic behaviour of Isophya fatrensis,
the last phaneropterine species with unknown acoustic signal pattern
from Central Europe. First European Congress on Orthoptera
Conservation, Trier, Germany, 18-20 March 2016, p. 21.
https://www.uni-
trier.de/fileadmin/fb6/prof/BIO/Claudes_Datenbank_sortiert/08Heusc
hrecken_Tagung/Programm/Abstractband_Heuschreckentagung_Trier
_Homepage_11.03.16.pdf
Iorgu I.Ș. - The hunt for peculiar Plump Bush-crickets in the largest
European volcanic caldera (Insecta: Orthoptera: Tettigoniidae).
International Zoological Congress of Grigore Antipa Museum,
Bucharest, Romania, 16-19 November 2016, p. 50.
http://czga.ro/pozepagini/CZGA2016_Book_of_abstracts_on_line_edi
tion_.pdf
Iorgu E.I., Iorgu I.Ș., Vadana M.I., Krapal A.M., Popa A.F., Popa
O.P., Popa L.O. - Preliminary data on the genetic diversity of Isophya
camptoxypha in the Carpathians. International Zoological Congress of
Grigore Antipa Museum, Bucharest, Romania, 16-19 November 2016,
p. 80.
http://czga.ro/pozepagini/CZGA2016_Book_of_abstracts_on_line_edi
tion_.pdf
Data, Director de proiect,
05 Decembrie 2016 Dr. Ionuț Ștefan Iorgu


















