13. Ac Nucleici
47
ADN şi ARN MOLECULELE EREDITĂŢII http://www.dnai.org/timeline/index.html
-
Upload
dragos-ionut-criveanu -
Category
Documents
-
view
87 -
download
2
Transcript of 13. Ac Nucleici

ADN şi ARN
MOLECULELE EREDITĂŢII
http://www.dnai.org/timeline/index.html

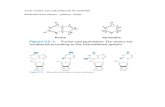
STRUCTURA PRIMARĂ (COVALENTĂ) A ADN
ADN este o macromoleculă cu structură polinucleotidică
un nucleotid este alcătuit din:
o oză
o bază azotată
una sau mai multe grupări fosfat

Pentozele din structura acizilor nucleici
DEOXIRIBOZA-în ADN (acidul deoxiribonucleic) RIBOZA-în ARN (acidul ribonucleic)

Structura bazelor azotate din ADN Heterocicluri cu N derivate de la:
purină BAZE PURINICE primidină BAZE PIRIMIDINICE

Tautomeria bazelor azotate (ceto enol şi amino imino)
in vivo, echilibrele sunt deplasate către formele ceto, respectiv amino
săgețile colorate indică atomii care pot participa la formarea legăturilor de hidrogen

Structura nucleozidelor și nucleotidelor NUCLEOZID: bază azotată legată -N-glicozidic la C1‘ al pentozei la formarea legăturii glicozidice participă:
N9 din bazele purinice N1 din bazele pirimidinice
NUCLEOTID: nucleozid esterificat la C5‘-OH cu una sau mai multe grupări fosfat

Nomenclatura bazelor, nucleozidelor şi nucleotidelor
din ADN
Baza Deoxiribonucleozid Deoxiribonucleotid (Deoxiribonucleozid-5-
monofosfat)
Adenina (A) Deoxiadenozina Deoxiadenilat (dAMP)
Guanina (G) Deoxiguanozina Deoxiguanilat (dGMP)
Timina (T) Deoxitimidina Deoxitimidilat (dTMP)
Citozina(C) Deoxicitidina Deoxicitidilat (dCMP)

Structura unui fragment de catenă ADN
nucleotidele sunt legate
prin punți 3‘-5’ fosfodiesterice într-o succesiune (secvență) liniară
o catenă polinucleotidică catenele polinucleotidice au polaritate:
la o extremitate se găsește gruparea 5’-P la cealaltă extremitate este gruparea 3‘-OH
secvența de nucleotide poate fi notată:
pApGpTpC....
prin convenție, notatrea începe cu
nucleotidul 5’

MODELUL ADN DUBLU CATENAR (ADNdc) (James Watson, Francis Crick, 1953)
A fost dedus pe baza imaginilor de difracție a razelor X prin fibre de ADN
STRUCTURA SECUNDARĂ A ADN

Caracteristicile modelului Watson-Crick
ADN celular este dublu catenar (bicatenar) alcătuit din două catene polinucleotidice cele două catene sunt antiparalele :
una are polaritate 5’-3‘iar cealaltă, 3‘-5’
răsucite helical în jurul unui ax comun (-helix de dreapta)
bazele purinice şi pirimidinice se găsesc în interiorul helixului
grupările fosfat şi unităţile deoxiriboză sunt la exterior
planurile bazelor sunt perpendiculare pe axa helixului
planurile ozelor formează unghiuri de ~90 cu planurile bazelor
diametrul helixului : 2 nm bazele adiacente:
se află la distanţă de 0,34 nm una de alta de-a lungul axei helixului, sunt rotite una faţă de cealaltă cu 36
structura helicală se repetă după fiecare 10 baze pe fiecare catenă, la intervale de 3,4 nm

cele două catene sunt asociate prin legături de hidrogen între perechile de baze
doar două tipuri de perechi de baze sunt permise de restricţiile sterice şi geometrice
perechi Watson-Crick: A.T şi G.C.
A formează două legături de hidrogen cu T
G formează trei legături de hidrogen cu C.
legăturile glicozidice ale unei perechi de baze nu sunt diametral opuse, B-ADN are o canelură mare (1,2 nm/ 0,85 nm) şi o canelură mică (0,6 nm/ 0,75 nm) fiecare este mărginită de atomi potenţial donori /acceptori de legături de hidrogen
în canelura mică se află O-2 al pirimidinei şi N-3 al purinei canelura mare este pe partea opusă

Structura ADN dublu catenar
Secvenţa bazelor azotate de-a lungul unei catene polinucleotidice poartă informaţia genetică

Forțele care stabilizează ADN dublu catenar
formarea legăturilor de hidrogen între perechile de baze: stabilitate termodinamică
conţinut informaţional, prin specificitatea perechilor de baze
Legături de hidrogen
sunt stabile când: atomii implicați se găsesc anumite distanțe unul de altul (0,28-0,30 nm pentru O şi N)
aranjamentul lor este liniar
criteriile geometrice le conferă specificitate
Forţele hidrofobe
bazele azotate sunt plate şi relativ insolubile în apă
tind să se aşeze una peste alta, perpendicular pe axa dublului helix (stivuirea bazelor)
helixul este stabilizat prin interacţiunile dintre norii electronici (-) ai bazelor stivuite în interiorul său

Variabilitatea conformației ADN dublu catenar
conformația descrisă de modelul Watson-Crick (ADN-B) corespunde majorității ADN celular
în anumite condiții, ADN dublu catenar
poate adopta și alte conformații: ADN-A și ADN-Z
poate forma structuri de tip triplex sau cvadruplex

CONFORMAŢIILE
ADN
DUBLU CATENAR

Caracteristica geometrică
Forma A Forma B Forma Z Sensul de răsucire al helixului
dreapta dreapta stânga
Diametrul helixului 2,3 nm 2 nm 1,8 nm
Unitatea repetitivă 1pb 1pb 2pb
Rotaţie/pb 33 36 -60/2
Nr. baze/tur de spiră 11 10,4(10,0-10,7) 12 (6 dinucleotide)
Înclinarea pb faţă de axa helixului
+19 -1,2 -9
Creşterea helixului/pb de-a lungul axei
0,26 nm 0,34 nm 0,37 nm
Pasul helixului 2,8nm 3,4 nm 4,5 nm
Unghiul mediu al răsucirii în elice
+18 +16 0
Conformaţia legăturii glicozidice
anti anti purine-sin;
pirimidine-anti
Proporţiile canelurii mari Extrem de îngustă şi foarte adâncă
Largă şi de adâncime intermediară
Aplatizată pe suprafaţa helixului
Proporţiile canelurii mici Foarte largă dar superficială
Îngustă şi de adâncime intermediară
Extrem de îngustă dar foarte adâncă

PROPRIETĂȚILE ADN
BICATENAR

Forma şi dimensiunile ADN bicatenar
(2x106 Da= 3000 pb= 1 m) Sursa pb.103
(kb)
Lungime
(m)
Virusuri
SV40 5,1 1,7
Fagul 48,6 17
Fagul T2 166 56
Bacterii
Mycoplasma 760 260
E. coli 4000 1360
Eucariote
Saccharomyces 13500 4600
Drosophila 165000 56000
Om 2900000 990000

Denaturarea şi renaturarea ADN
Separarea catenelor care formează dublul helix=denaturare (melting; topire)
in vivo, în timpul replicării (procesul de sinteză a ADN) in vitro, când o soluţie de ADN este adusă în condiţii de temperatură şi pH foarte diferite de condiţiile fiziologice

Curba denaturării termice a ADN
bazele azotate absorb radiații din domeniul UV (240 nm) reprezentând grafic creşterea absorbanţei în funcţie de temperatura la care este supusă soluţia ADN curbă de topire de formă sigmoidală absorbanţa creşte brusc într-un domeniu de temperatură relativ îngust cooperativitatea interacţiunilor care stabilizează dublul helix temperatura de topire (Tm-melting temperature) a ADN temperatura la care absorbanţa reprezintă 50% din valoarea maximă, corespunzătoare denaturării complete (separării complete a catenelor)

Temperatura de topire este caracteristică fiecărui ADN
Tm este determinată de: conţinutul de perechi G.C tăria ionică a soluţiei (a)-soluţii saline diluate; (b)-soluţii saline concentrate
oferă informații privind compoziția de baze azotate a ADN

Proprietăţile tolpologice ale ADN
Topologia ADN se referă la forma şi traiectoria spaţială a ADN
abordează interdisciplinar, cu instrumentele matematicii, probleme ale biologiei moleculare ca:
suprarăsucirea (supercoiling)
înnodarea (knotting)
catenarea (catenation)

Micrografii electronice ale unor molecule de ADN circular;
• 1-forma relaxată; 2-5-forme suprarăsucite
Suprarăsucirea ADN ADN circular purificat din bacterii şi eucariote
este de regulă suprarăsucit negativ (spre dreapta) suprarăsucirea negativă poate fi considerată o
formă de stocare a energiei libere utilizabilă în procese care necesită separarea catenelor ADN (replicarea, transcripţia)

Topoizomerazele
enzime care pot modifica gradul de suprarăsucire a ADN
bicatenar
catalizează scindarea temporară şi reformarea
legăturilor fosfodiesterice
există două tipuri de topoizomeraze care se deosebesc
prin mecanismul de acţiune şi prin funcţia celulară:
Topoizomeraze de tip I
Topoizomeraze de tip II

Aplicații medicale: inhibitorii topoizomerazelor Inhibitorii ADN girazei bacteriene
agenți antibacterieni
Ex. quinolonele (negram, urotrat) și aminocumarinele (novobiocina)
utilizați pentru tratarea infecțiilor urinare cauzate de colibacili
inhibă giraza bacterianăblochează replicarea cromozomului bacterian
acțiunea selectivă a acestor antibiotice se bazează pe diferența structurală dintre topoizomerazele bacteriene și cele umane
Ciprofloxacin
Novobiocina
Inhibitorii topoizomerazelor umane anticancerigeni blochează acțiunea reglatoare a topoizomerazelor
Ex. etopozidul (VP16) inhibă topoizomeraza II taxolul (extras din Taxus baccata) inhibă topoizomeraza I
Etopozid

STRUCTURA ARN Primară: secvenţa nucleotidelor
Secundară: formarea perechilor intracatenare de baze
Terţiară: conformaţia tridimensională

Structura primară a ARN
ARN are, ca şi ADN, structură polinucleotidică dar se deosebeşte de acesta sub trei aspecte:
scheletul oză-fosfat al ARN conţine riboza în loc de 2’-deoxiriboza
în ARN timina este înlocuită cu uracil
există în mod obişnuit în formă monocatenară
aceste modificări chimice minore au consecinţe structurale şi funcţionale majore.

Structura nucleozidelor și nucleotidelor ARN
NUCLEOZID: bază azotată legată b-N-glicozidic la C1‘ al ribozei La formarea legăturii glicozidice participă:
N9 din bazele purinice (G, C) N1 din bazele pirimidinice (U, C)
NUCLEOTID: nucleozid esterificat la C5 ‘-OH cu una sau mai multe grupări fosfat

Nomenclatura bazelor, nucleozidelor şi nucleotidelor
din ARN
Baza Ribonucleozid Ribonucleotid
(Ribonucleozid-5-monofosfat)
Adenina (A) Adenozina Adenilat (AMP)
Guanina (G) Guanozina Guanilat (GMP)
Uracil (U) Uridina Uridilat (UMP)
Citozina(C) Citidina Citidilat (CMP)
ARN este sintetizat totdeauna sub formă monocatenară, prin copierea informaţiei unei singure catene dintr-un duplex care serveşte ca matriţă.
deşi ARN poate forma duplexuri helicale lungi, asemănătoare structural cu ADN-A, acestea sunt neobişnuite în natură
datorită prezenței unor secvențe complementare de baze, în structura ARN există scurte regiuni dublu helicale formate prin împerecheri intracatenare ale bazelor azotate

De ce T în ADN și U în ARN? prin dezaminare oxidativă accidentală, C se transformă în U, putând genera o mutație
sistmele de reparare recunosc U (a cărui prezență este nefirească în ADN) și îl îndepărtează prin excizie, fiind apoi înlocuit cu C
în ADN, T este un semnal de normalitate, iar orice apariție a U este considerată o greșeală și tratată ca atare
dacă ADN ar fi avut U în stare normală, enzimele de reparare nu ar fi putut distinge U normal de U rezultat prin dezaminarea C
în ARN, această diferențiere nu se poate face; ca urmare, erorile rezultate din dezaminarea C nu sunt reparate
pentru ARN nu există un sistem de reparare a erorilor
ARN este o moleculă efemeră, în contrast cu ADN care trebuie conservat pe timp nelimitat

Elemente de structură secundară a ARN
Motive de structură secundară a ARN: (A)-agrafă, (B)-protuberanţă, (C)-buclă; (D) frunza de trifoi
Apar în regiunile cu secvențe complementare de baze azotate
(A) perechi convenţionale Watson Crick: A.U şi G.C (B) (B) perechea neconvenţională G.U
Perechi de baze din ARN

Structura terţiară a ARN
Formarea tripletului U.A.U
(A) Formarea unui pseudonod.
(B) Interacţiunile dintre bazele azotate care stabilizează structura

Funcțiile ARN
Cu excepţia anumitor virusuri, ARN nu are rol de material genetic şi nu este matriţă pentru propria sa replicare
ARN îndeplineşte însă roluri biologice numeroase şi diverse, prin care participă la exprimarea informaţiei genetice codificate în ADN
ARN mesager (ARNm): intermediar între gene şi ribozomi, dispozitive celulare de sinteză a proteinelor
ARN de transport (ARNt): adaptor între codonii din ARN mesager şi aminoacizi
ARN ribozomal (ARNr): rol structural dar şi catalitic, la formarea legăturilor peptidice
Pe lângă aceste trei clase majore, există multe alte tipuri de ARN care îndeplinesc funcţii celulare importante

Diversitatea funcţională a ARN
Tipuri Funcţii
ARN heterogen nuclear-ARNhn
Produs primar al transcripţiei, la eucariote;
este prelucrat la ARNm
ARN citoplasmatic de mici dimensiuni (small)-ARNsc
Prelucrarea ARNt; particulă de recunoaştere a semnalului-direcţionează secreţia proteinelor nou sintetizate prin reticulul endoplasmic
ARN nuclear de mici dimensiuni-ARNsn
În alcătuirea particulelor ribonucleoproteice de mici dimensiuni; participă la prelucrarea ARNm
ARN nucleolar de mici dimensiuni-ARNsno
Prelucrarea, maturarea şi metilarea ARNr, la eucariote
ARN telomerazic Matriţa telomerazei, enzima care adaugă repetări telomerice la capetele cromozomilor, la eucariote
micro ARN (ARNmi) reglează post-transcripţional expresia genelor prin procesul numit interferenţa ARN

ORGANIZAREA
MATERIALULUI GENETIC
ÎN CELULE

Genomurile eucariote
La organismele eucariote, genomul are două componente:
genomul nuclear
genomul organitelor celulare.
Genomul nuclear conţine cea mai mare parte a materialului genetic, este localizat în nucleu şi divizat în cromozomi.
Genomurile organitelor celulare au dimensiuni mai mici şi sunt localizate în mitocondrii la toate organismele eucariote;
Eucariotele fotosintetizante au şi genomuri cloroplastice.

Genomul nuclear constă dintr-o serie de molecule liniare de ADN, fiecare alcătuind un cromozom
la toate organismele eucariote există cel puţin doi cromozomi
moleculele de ADN sunt totdeauna liniare.
la acest nivel de organizare, aspectul variabil al genomurilor este numărul de cromozomi, care nu pare să fie corelat cu caracteristicile biologice ale organismului.
drojdiile au 16 cromozomi, de patru ori mai mult decât Drosophila.
nu există o corelaţie între mărimea genomului (cantitatea de ADN) şi numărul cromozomilor:
unele salamandre au genomul de 30 de ori mai mare decât genomul uman dar este format dintr-un număr de două ori mai mic de cromozomi.
caracterul neuniform al proceselor de evoluţie care au modelat arhitectura genomurilor la diferite organisme.
CARIOTIPUL UMAN

cea mai importantă funcţie a cromozomilor este de purtători ai genelor, unităţile funcţionale ale eredităţii.
gena este definită de obicei ca un segment de ADN care conţine instrucţiunile pentru sinteza unei anumite proteine sau a unui set de proteine înrudite
există şi gene al căror produs final nu este o proteină, ci o moleculă de ARN care poate îndeplini în celulă diverse funcţii structurale sau catalitice
cromozomii eucariotelor conţin, pe lângă gene, un mare exces de ADN care nu pare să poarte o informaţie vitală
ADN aparent nefolositor (junk DNA), a cărui secvenţă de nucleotide poate să nu fie importantă
funcţionează ca material de spaţiere între gene
implicat în:
evoluţia pe termen lung
exprimarea corectă a genelor
în contrast, genomurile procariote sunt extrem de compacte

Împachetarea ADN în cromozomi
cromozomii sunt mult mai scurţi decât moleculele de ADN pe care le conţin
la om, lungimea ADN din cromozomul 22, format din 48106 pb, este de 5 cm; în timpul mitozei, acest cromozom are doar 2m.
existenţa unui sistem organizat de compactare a ADN
compactarea trebuie să permită:
împachetarea ADN în spaţiul limitat al nucleului
desfăşurarea proceselor ca replicarea, repararea şi transcripţia (accesarea genomului)

Nucleozomul
un nucleozom constă dintr-un segment un segment de ADN bicatenar de 146-147 pb care se înfăşoară de 1,65 ori spre stânga în jurul unui miez proteic, numit octamer histonic.
octamerul este alcătuit din câte două molecule de histone H2A, H2B, H3 şi H4, numite şi histone miez (core histone).
diametrul octamerului este de aproximativ 10 nm.
o singură moleculă de histonă H1, numită şi histonă de legătură (linker histone), se leagă la fiecare nucleozom, venind în contact atât cu miezul histonic cât şi cu ADN.
Micrografie electronică a fibrilei de cromatină- structura mărgele pe aţă

Micrografie electronică a fibrei de cromatină
Două modele ale fibrei de cromatină de 30 nm. (A) solenoid (B) zigzag
Fibra de cromatină de 30 nm este forma în care se găseşte cromatina în interfază, perioada dintre două diviziuni celulare se organizează sub formă de bucle pe un schelet proteic, într-o structură cu diametru de 30 nm. compactarea avansată conduce la heterocromatină, inactivă transcripţional

Genomurile procariote
majoritatea au dimensiuni mai mici de 5106 pb există și unele mult mai mari
ex. B. megaterium are un genom uriaş de 3107 pb
imaginea clasică:
un duplex ADN circular format prin legarea covalentă a capetelor 3’ şi 5’ ale fiecărei catene localizat în nucleoid
la E. coli:
cromozomul circular are circumferinţa de 1,6 mm bacteria are doar 1,02,0 m ADN-ul adoptă în celulă o formă compactă (este suprarăsucit) cu ajutorul unor proteine care se leagă la ADN structura rezultată nu se aseamănă cromozomului eucariot

Organizarea materialului genetic la procariote
modelul actual al cromozomului bacterian (E. coli ) ADN este ataşat la un miez proteic din care radiază 40-50 de bucle suprarăsucite
fiecare buclă este formată din circa 100 kb ADN
HU
cea mai abundentă dintre proteinele implicate în compactarea ADN bacterian este diferită structural de histone îndeplineşte o funcţie similară formează un tetramer în jurul căruia se înfăşoară circa 60 pb ADN

Genomurile virale Virusurile suntentităţi infecţioase acelulare, submicroscopice (20-300 nm) alcătuite din:
material genetic (o cantitate mică :106 Da -108 Da) o capsulă proteică
nu se pot reproduce independent replicarea lor se realizează prin infectarea unei celule gazdă
Genomurile virale prezintă o mare diversitate a structurală:
ADN bacteriofagii au genomurile formate din ADN bicatenar
ARN virusurile plantelor au de obicei genomuri din ARN monocatenar
ADN și ARN genomul citomegalovirusului: un miez ADN şi câteva segmente ARN
la unele specii virale există nucleotide anormale, ca hidroximetilcitozina genomurile virale ADN au stabilitate mai mare decât genomurile ARN
a. virus bacterian:bacteriofagul T4 b. virus animal (adenovirus) c. virus vegetal (mozaicul tutunului)

Virusurile sunt ansambluri supramoleculare
acţionează ca paraziţi intracelulari
irusurile sunt alcătuite din elemente genetice (ADN sau ARN) închise într-o capsulă proteică
Sunt entităţi infecţioase, nu organisme vii
Se reproduc numai în interiorul celulelor pe care le parazitează
Prezintă specificitate aproape absolută pentru celulele gazdă, în care se multiplică
Există virusuri specifice pentru toate tipurile de celule
A. Alcătuirea unui bacteriofag
B. Mecanismul infecţiei virale

Tipuri de genomuri virale

EXPRIMAREA GENOMULUI Imaginea clasică:
informaţia genetică este copiată prin procesul de replicare semiconservativă
fiecare catenă a ADN bicatenar este matriţă pentru sinteza unei catene complementare.
citirea şi utilizarea informaţiei genetice (exprimarea genomului) se realizează în două etape:
în procesul de transcripţie, segmente de ADN direcţionează sinteza unor molecule de ARN
în procesul de translaţie, moleculele ARN mesager direcţionează sinteza moleculelor proteice.
http://highered.mcgraw-hill.com/sites/0072437316/student_view0/chapter14/animations.html http://highered.mcgraw-hill.com/sites/0072437316/student_view0/chapter15/animations.html